
Bu içeriği görüntülemek için JoVE aboneliği gereklidir. Oturum açın veya ücretsiz deneme sürümünü başlatın.
Method Article
Oda ve Plaka Tabanlı Respirometreler Kullanarak Hücrelerdeki ve Dokulardaki Biyoenerjetik Değerleri Değerlendirmek için Yüksek Çözünürlüklü Respirometri
Bu Makalede
Özet
Yüksek çözünürlüklü respirometreler kullanılarak oksidatif fosforilasyonun değerlendirilmesi, mitokondri ve hücresel enerji metabolizmasının fonksiyonel analizinin ayrılmaz bir parçası haline gelmiştir. Burada, oda ve mikroplaka tabanlı yüksek çözünürlüklü respirometreler kullanılarak hücresel enerji metabolizmasının analizi için protokoller sunuyoruz ve her cihazın temel faydalarını tartışıyoruz.
Özet
Yüksek çözünürlüklü respirometri (HRR), bireysel hücresel enerji durumlarının analizi ve çeşitlendirilmiş substrat-unkuplör-inhibitör titrasyonu (SUIT) protokolleri kullanılarak solunum komplekslerinin değerlendirilmesi için oksidatif fosforilasyonun gerçek zamanlı olarak izlenmesine olanak tanır. Burada, iki yüksek çözünürlüklü respirometri cihazının kullanımı gösterilmiştir ve kültürlenmiş hücrelerin, iskelet ve kalp kası liflerinin ve beyin ve karaciğer gibi yumuşak dokuların analizi için geçerli temel bir protokol koleksiyonu sunulmaktadır. Kültürlenmiş hücreler ve dokular için protokoller, her ikisi de standart solunum protokollerini kapsayan oda bazlı bir respirometre ve mikroplaka tabanlı bir respirometre için kültürlenmiş hücreler için sağlanmıştır. Karşılaştırmalı amaçlar için, çoklu solunum sistemi eksikliğine neden olan mitokondriyal translasyonda eksik olan CRISPR mühendisliği HEK293 hücreleri, solunumdaki hücresel kusurları göstermek için her iki cihazla birlikte kullanılır. Her iki respirometre de, incelenen araştırma sorusuna ve modeline bağlı olarak ilgili teknik yararları ve uygunlukları ile hücresel solunumun kapsamlı bir şekilde ölçülmesine izin verir.
Giriş
Mitokondri, enerjinin temel hükmünü yerine getirir ve nükleotitlerin, lipitlerin ve amino asitlerin anabolizması, demir-kükürt kümesi biyogenezi gibi temel hücresel biyoenerjetik ve metabolik süreçlere katkıda bulunan bölümlere ayrılmış bir organeldir ve kontrollü hücre ölümü gibi sinyalleşmeye dahil edilir 1,2,3 . Oksidatif fosforilasyon yoluyla mitokondriyal biyoenerjetik maddeler, hücre içindeki hemen hemen tüm hücresel süreçlere katkıda bulunur ve sonuç olarak, birincil veya ikincil kökenli mitokondriyal işlev bozuklukları, geniş bir hastalık koşulları spektrumu ile ilişkilidir 4,5. Mitokondriyal disfonksiyon sadece yapı veya mitokondriyal yoğunluktaki değişiklikleri değil, aynı zamanda solunum sisteminin kalitesinde ve düzenlenmesinde de değişiklikler içerir6. Bu kalitatif eleman substrat kontrolünü, kuplaj özelliklerini, post-translasyonel modifikasyonları, krista dinamiklerini ve solunum süperkomplekslerini kapsar 7,8. Bu nedenle, hücrenin enerji metabolizmasını değerlendirmek için deneysel ve tanısal yaklaşımlar için mitokondriyal biyoenerjetik ilaçların doğru analizi sağlık ve hastalıkta önemlidir.
Mitokondriyal oksidatif fosforilasyon (OXPHOS), adenozin trifosfat (ATP)9 yoluyla hücresel enerji üretimi için solunum sistemi veya elektron transfer sistemi (ETS) içindeki bir dizi reaksiyondur. I ve II kompleksleri boyunca elektron akışından karmaşık IV'e kadar enerjiyi kullanmak için çok enzimatik adım, iç mitokondriyal membran boyunca bir elektrokimyasal proton gradyanı üretir ve daha sonra adenozin difosfatın (ADP) karmaşık V (F1FO ATP sentaz) yoluyla ATP'ye fosforilasyonu için kullanılır (Şekil 1A).
İlk olarak, trikarboksilik döngü (TCA), glikoliz ve piruvat oksidasyonu sırasında iki elektron taşıyıcıları üretilir: nikotinamid adenin dinükleotid (NADH) ve dihidroflavin adenin dinükleotid (FADH2). NADH, kompleks I'de (NADH dehidrojenaz) oksitlenir, bu sırada iki elektron koenzim Q'ya aktarılır (kinon kinol'e indirgenir), protonlar membranlar arası boşluğa (IMS) pompalanır. İkincisi, kompleks II (Süksinat dehidrojenaz) FADH2'yi oksitler ve protonları pompalamadan elektronları koenzim Q'ya besler. Üçüncüsü, kompleks III'te (Sitokrom c oksidoredüktaz), koenzim Q'dan gelen elektronlar sitokrom c'ye aktarılırken, protonlar IMS'ye pompalanır. Dördüncüsü, sitokrom c, elektronları protonları pompalamak için son kompleks olan ve oksijenin protonları asimile etmek için bir elektron alıcısı olarak işlev gördüğü ve sonuçta su oluşturan kompleks IV'e (Sitokrom c oksidaz) aktarır. Mitokondrinin tükettiği ve bir oksigrafla ölçülebilen bu oksijendir. Son olarak, karmaşık I, karmaşık III ve karmaşık IV'ten üretilen protonlar, karmaşık V'yi döndürmek için kullanılır, böylece ATP9 üretilir.
Önemli olarak, elektron transferi sadece doğrusal bir şekilde gerçekleşmez, aksi takdirde elektron taşıma zinciri olarak gösterilir. Bunun yerine, elektronlar çoklu solunum yolları aracılığıyla koenzim Q havuzuna aktarılabilir ve yakınsak elektron akışını kolaylaştırabilir. Örneğin NADH-substratları ve süksinat, sırasıyla karmaşık I ve karmaşık II yoluyla girebilir. Yağ asidi oksidasyonundan elde edilen elektronlar, elektron transfer flavoprotein kompleksi aracılığıyla bağışlanabilir. Gerçekten de, OXPHOS'un kapsamlı bir analizi, uygun yakıt substratları ile bütünsel bir yaklaşım gerektirir (Şekil 1A).

Şekil 1: Mitokondriyal oksidatif fosforilasyon ve spesifik substrat ve inhibitör protokolleri. (A) Mitokondri ve elektron transfer sisteminin şeması (CI-CIV) ve mitokondriyal F1F0 ATP sentaz (CV). Tüm yapılar PDB'dendir. Rakamlar sadece bu çalışmada açıklanan substratları ve inhibitörleri göstermektedir). (B) Bir mHRR cihazında standart protokol kullanarak bozulmamış HEK293 hücrelerinde oksijen akışının örnek izi. (C) Bir cHRR cihazında standart protokol kullanarak bozulmamış HEK293 hücrelerinde oksijen akışının örnek izi. (D) İlgili SUIT protokolüne sahip sağlıklı bir donörden geçirgenleştirilmiş insan fibroblastlarındaki oksijen akışının örnek izi. Kısaltmalar: 1 = Bozulmamış hücrelerin rutin solunumu; 2 = Devlet 2; 3 = Durum 3(I); 4 = cytC ile Durum 3(I); 5 = Durum 3 (I+II); 6 = Sızıntı (OM); 7 = ETS kapasitesi; 8 = S (ÇÜRÜKLÜK); 9 = ROX; 10 = TMPD; 11 = Az. ROT = Rotenon, = Antimisin, ATP = Adenozin trifosfat, Az = Azid, OM = Oligomisin, FCCP = Karbonil siyanür p-trifloro-metoksifenil-hidrazon; Asc = Askorbat, TMPD = N,N,N′,N′-tetrametil-p-fenilendiamin, Succ = Süksinat, M = Malat, P = Piruvat, ADP = Adenozin difosfat, NAD = Nikotinamid adenin dinükleotid, IMS = Membranlararası boşluk, FAD = Flavin adenin dinükleotid. Bu şeklin daha büyük bir versiyonunu görüntülemek için lütfen buraya tıklayın.
{kind=link}
HRR kullanılarak mitokondriyal OXPHOS kapasitesinin analizi, sadece primer mitokondriyal defektler10,11 için değil, kanser ve yaşlanma12 gibi biyolojinin diğer tüm alanlarına uzanan tanısal değeri olan enstrümantal bir biyokimyasal yöntem haline gelmiştir. HRR, bireysel veya kombine mitokondriyal solunum kompleksi eksikliğini doğrudan yansıtan ve dolaylı olarak hücresel işlev bozukluğu ve değişmiş enerji metabolizması9 ile ilişkili mitokondriyal OXPHOS kapasitesinin analizi ile hücresel solunumun belirlenmesine izin verir. Metodolojik olarak, solunum ölçümleri hücreler, doku veya izole mitokondri11,13,14 kullanılarak gerçekleştirilir, dondurulmuş materyal sadece kısmen uygun15,16 ile gerçekleştirilir. Dondurulmuş dokunun, süperkompleks stabilitesi korunmuş sağlam bir ETS'ye sahip olduğu gösterilmiştir15. Böylece, geleneksel TCA ara ürünlerinin aksine, ilgili substratlar doğrudan ETS'ye beslenir. Bununla birlikte, ETS ve ATP sentezi arasındaki bağlantı, membran bütünlüğü donma hasarı (buz kristali oluşumu) nedeniyle tehlikeye girdiğinden kaybolur.
Solunum deneyleri normalde geçirgenleşmemiş veya geçirgenleştirilmiş hücrelerde veya dokularda endotermler için 37 ° C'lik fizyolojik bir sıcaklıkta gerçekleşir. Birincisi sitozolik metabolik bağlamı göz önünde bulundururken, ikincisi, spesifik substratların (ve inhibitörlerin) eklenmesiyle bireysel OXPHOS komplekslerinin ve ATPaz'ın enerjik katkısını sağlar. Substratların ve inhibitörlerin dizilimi ve varyasyonu, OXPHOS fonksiyonunun çeşitli bilimsel sorularını ele almak için çeşitli SUIT protokolleri17 ve tahliller18'in geliştirilmesine yol açmıştır (12 altında gözden geçirilmiştir). Hücresel solunumun temel protokolü dört farklı durumu değerlendirir: i) rutin solunum - herhangi bir substrat veya inhibitör eklemeden ilgili solunum ortamındaki solunum, ancak endojen substratları tüketir. Bu durum, örneğin değiştirilmiş metabolit profillerinin neden olduğu genel OXPHOS veya ikincil kaynaklı solunum kusurlarını ortaya çıkarabilir. Daha sonra, ATPaz inhibitörü oligomisinin eklenmesi, iç mitokondriyal membranın protonlara geçirgenliğini ortaya çıkarır, ii) sızıntı solunumu olarak tanımlanır. Eşinsiz karbonil siyanür p-trifloro-metoksifenil-hidrazon (FCCP) gibi bir protonoforun müteakip titrasyonlanması, iii) ayrışmamış solunum olarak tanımlanan açık transmembranlı proton devre modunda ETS kapasitesinin maksimum olduğu durumun belirlenmesini sağlar. Önemli olarak, mitokondriyal membranlara aşırı mekanik hasar yoluyla deneysel müdahalelerle de ayrılmamış bir durum ortaya çıkabilir. Tersine, bağlanmamış durum, fizyolojik olarak kontrol edilen içsel bir mekanizma ile solunum ayrışmasını ifade eder. Son olarak, kompleks III inhibitörü antimisin ve kompleks I inhibitörü rotenonun eklenmesiyle ETS'nin tamamen inhibe edilmesi, mitokondriyal olmayan oksijen tüketen süreçlerden kalan oksijen tüketimini (ROX) belirler (Şekil 1A-C).
Mitokondriyal biyoenerjetik beş ayrı solunum durumundan oluşur19,20. Durum 1 solunum, endojen olarak mevcut olanlar dışında, herhangi bir ek substrat veya ADP içermez. ADP'nin eklenmesinden sonra, ancak yine de, hiçbir substrat yok, durum 2 solunumu elde edilir. Elektron transferine ve ATP sentezine izin veren substratlar eklendiğinde, durum 3 solunumuna ulaşılır. Bu durumda, OXPHOS kapasitesi ADP, inorganik fosfat, oksijen, NADH ve süksinat bağlantılı substratların doygunluk konsantrasyonlarında tanımlanabilir. Durum 4 solunumu veya LEAK solunumu, yeterli substratlara sahipken ADP'siz veya kimyasal olarak inhibe edilmiş ATP sentezleri olmayan bir durum olarak tanımlanabilir. Son olarak, kapalı bir oda ortamında tüm oksijen tükendiğinde (anoksik), durum 5 solunumu gözlenir.
Deneysel modele ve araştırma sorusuna bağlı olarak farklı uygulanabilirliğe sahip kapalı odacıklı bir sistemde zaman içinde oksijendeki azalmanın işlevi olarak ölçülen oksijen tüketiminin analizi yoluyla OXPHOS'un mevcut gerçek zamanlı değerlendirmesine hakim olan iki cihazla hücresel enerji durumlarını14 ile değerlendirmek için çeşitli yöntemler mevcuttur: Oroboros 2k yüksek çözünürlüklü respirometre ve Seahorse XF hücre dışı akı analizörü. Her iki cihaz da oksijen tüketim oranlarını, saniyede oksijen pikomollerinde (pmol) (O2) bir azalma olarak, oda veya mikroplaka kuyusu içinde mutlak bir değer olarak kaydeder. Kütle başına spesifik oksijen tüketimi, ilgili oksijen tüketiminin hücre sayısı (milyonlarca), doku ağırlığı (mg) veya protein miktarı başına belirli bir tampon tarifinde normalleştirilmesiyle elde edilir.
O2k (Oroboros Instruments), polarografik oksijen sensörü (oda tabanlı yüksek çözünürlüklü respirometre olarak kısaltılır: cHRR) ile donatılmış kapalı iki odacıklı bir sistemdir. Her deney odası, manyetik karıştırıcılar tarafından homojen tutulan 2 mL sıvı tutar. Polarografik oksijen sensörü, oksijeni ölçmek için amperometrik bir yaklaşım kullanır: altın bir katot, bir gümüş / gümüş klorür anot ve bir KCI çözeltisi arasında, üzerine bir voltajın (0.8 V) uygulandığı bir elektrokimyasal hücre oluşturur. Tahlil ortamından gelen oksijen, 25 μm florlu etilen propilen membrandan (O2-geçirgen) yayılır ve katotta indirgenerek hidrojen peroksit üretir. Anotta, gümüş hidrojen peroksit tarafından oksitlenir ve bir elektrik akımı üretir. Bu elektrik akımı (amper) doğrusal olarak kısmi oksijen basıncı ile ilişkilidir. Oksijen konsantrasyonunu hesaplamak için kısmi oksijen basıncı ve tahlil ortamının oksijen çözünürlük faktörü kullanılır. Oksijen kısmi basıncı deneysel sıcaklığa bağlı olduğundan ve polarografik ölçümler sıcaklığa duyarlı olduğundan, sıcaklıktaki dalgalanmalar bir Peltier ısıtma bloğu tarafından hassas (±0.002 ° C) düzenlemeye ihtiyaç duyar. Sıcaklık 4 °C ile 47 °C aralığında kontrol edilebilir.
Seahorse XF hücre dışı akı analizörü (Agilent), üç floresan elektrodunun her bir kuyucukta zaman içinde oksijen tüketimini ölçtüğü 24 veya 96 delikli mikroplaka formatına sahip plaka tabanlı bir sistemdir (mikroplaka tabanlı yüksek çözünürlüklü respirometre olarak kısaltılır: mHRR). Tahlil sırasında otomatik enjeksiyon için tahlil kartuşundaki maksimum dört port mevcuttur. Bir tahlil, her biri üç fazlı birden fazla döngü içerir: 1) karıştırma, 2) bekleme ve 3) ölçüm. Ölçüm aşamasında, sensör probları mikroplakaya indirilir ve yayılan ışığı ölçmek için 7-10 μL hacim içeren geçici olarak kapalı bir oda oluşturulur. Bu ışık, fosforesans söndürmeye dayalı olarakO2'yi algılayan sensör problarının ucundaki polimer gömülü floroforlar tarafından yayılır. Floresan sinyalinin yoğunluğuO2 ile orantılıdır ve sensörün ve tahlil ortamının sıcaklığından etkilenir. Bu nedenle, doğru oksijen tahmini, herhangi bir numune olmadan arka plan kuyusu ile göreceli bir yaklaşım gerektirir. Oksijen konsantrasyonunun geri kazanılması, karıştırma aşamasında, sensör geçici odanın üzerindeki hacmi karıştırmak için yukarı ve aşağı hareket ettiğinde gerçekleşir. Her döngü bir oksijen tüketim oranını hesaplar. Sıcaklık 16 °C ile 42 °C aralığında kontrol edilebilir.
HRR, primer ve mitokondri ile ilişkili hastalıklarda ve genel hücresel metabolizmada hücresel biyoenerjetik değerleri değerlendirmek için altın standarttır. Bu çalışmada, hücre ve dokularda OXPHOS fonksiyonunu değerlendirmek için HRR için temel protokoller sağlanmıştır.

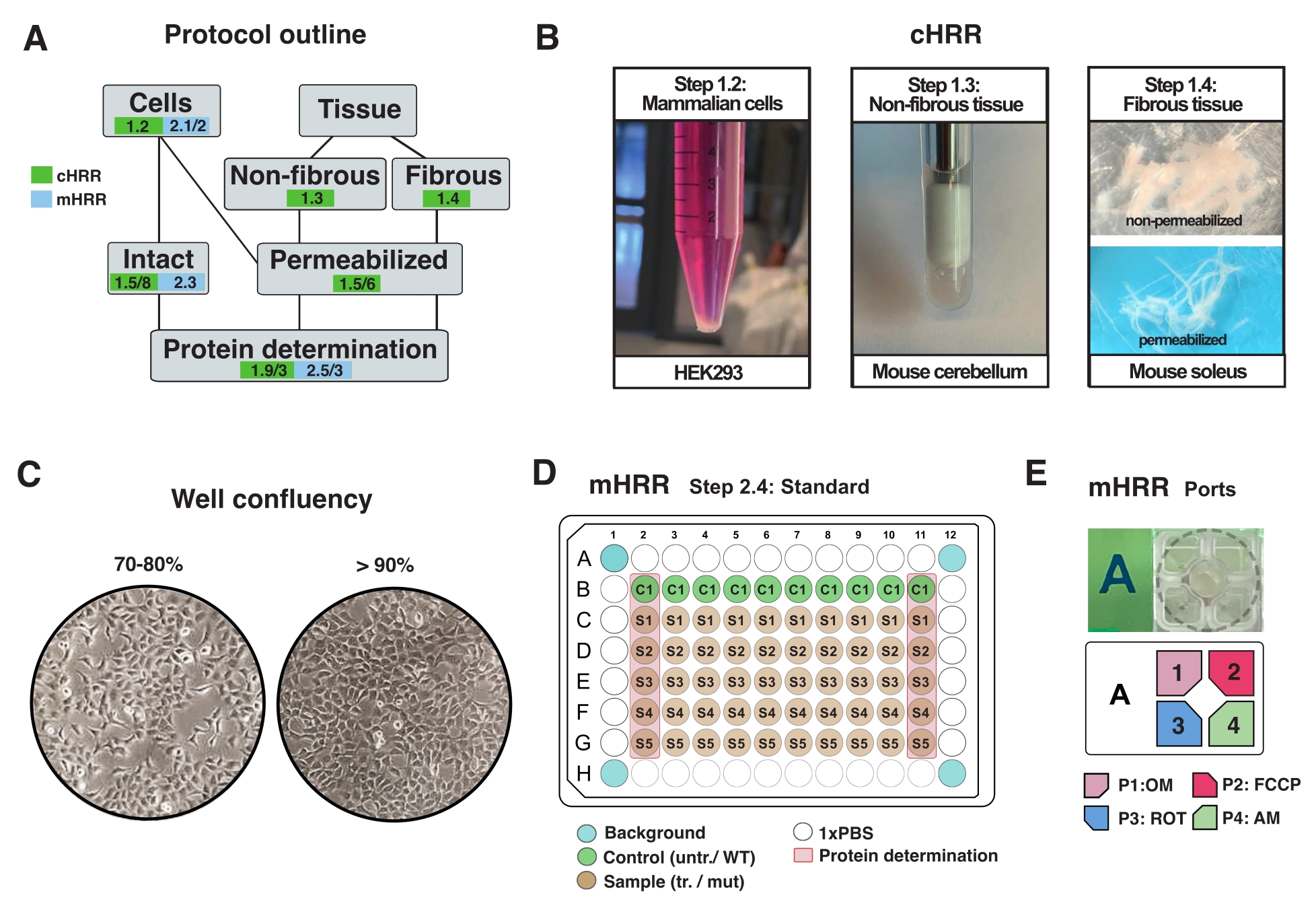
Şekil 2: cHRR için hücre ve doku preparatları için iş akışı ve mHRR respirometrisi için hücre hazırlığı . (A) Sağlanan protokollerin ana hatları. (B) Memeli hücreleri (adım 1.2): HEK293 pelet 3 x 106 hücreye eşittir (sol panel). Fibröz olmayan doku (adım 1.3): 2 mL Teflon çömlekçide (orta panel) murin beyincik lizatının hazırlanması. cHRR respirometrisi için saponun neden olduğu iskelet kası geçirgenliği (adım 1.4) sağ panel). (C) Standart mikroplaka tohumlama düzeni (adım 2.4) ve mHRR respirometrisi için ökaryotik hücrelerin (HEK293) analizi için akıcılık kontrolü. (D, E) mHRR respirometrisi için enjeksiyon port yükleme şeması (adım 2.4). Bu şeklin daha büyük bir versiyonunu görüntülemek için lütfen buraya tıklayın.
{kind=link}
Protokol
Tüm hayvan deneyleri, Ulusal Hayvan Deneyleri İnceleme Kurulu ve Güney Finlandiya Bölgesel Devlet İdari Ajansı'na uygun olarak gerçekleştirilir. Bu çalışmada erkek C57BL/6JOlaHsd fareler (4-6 aylık) kullanıldı. İnsan hücre hatlarının kullanımı için onay, Helsinki Üniversitesi kurumsal etik komitesinden alınmıştır.
1. Yüksek çözünürlüklü respirometri: Oda tabanlı respirometre (cHRR)
NOT: Protokolün bu bölümündeki deneyler Oroboros O2k-Core: Oxygraph-2k (Malzeme Tablosu) kullanılarak gerçekleştirilmiştir.
- Oksijen sensörlerinin kalibrasyonu
- 2.1 mL mitokondriyal solunum ortamında (MiR05, Tablo 1, çözünürlük faktörü: 0.92) 37 ° C'de respirometreleri >45 dakika boyunca önceden çalıştırın ve tarif edildiği gibi oksijen kalibrasyonu yapın21. Temel varyasyon ± 4 pmol/s içindeyse devam edin.
NOT: Arka plan sinyalindeki büyük dalgalanmalar, sensör zarının gerekli bakımını veya önceki deneylerden odada kalan inhibitör izlerini gösterebilir. Bir dizi deneyden önce enstrümantal bir arka plan oksijen akısı düzeltmesi önerilir25. - Sensör membranının zaman içindeki performansını izlemek için oksijen kalibrasyon değerlerini kaydedin.
NOT: Bu, sensör işlevini, sinyal-gürültü kararlılığını ve sensör membran bakımının ne zaman gerekli olduğunu ortaya çıkarır. Ortam basıncına bağlı olarak, MiR05'te 180-200 μmol arasında oksijen çözünür. - Solunum ortamına herhangi bir numune eklemeden önce odadaki tüm sıvıyı çıkarın.
NOT: Solunum odalarının hacmini düzenli olarak tam olarak 2 mL olacak şekilde değerlendirin.
- 2.1 mL mitokondriyal solunum ortamında (MiR05, Tablo 1, çözünürlük faktörü: 0.92) 37 ° C'de respirometreleri >45 dakika boyunca önceden çalıştırın ve tarif edildiği gibi oksijen kalibrasyonu yapın21. Temel varyasyon ± 4 pmol/s içindeyse devam edin.
- Yüksek çözünürlüklü respirometri için hücrelerin hazırlanması
- Kültür HEK293 hücreleri, Dulbecco'nun Modifiye Kartal ortamındaki (DMEM) 10cm2 çapındaki tabaklarda,% 10 ısı-inaktive fetal sığır serumu (FBS), GlutaMax, Esansiyel olmayan amino asitler ve Na-Piruvat22 ve idrar 23 ile desteklenmiş yüksek glikozlu, 37 ° C'de bir inkübatörde% 5 CO2'de OXPHOS kusurlu metabolizmayı desteklemek için.
NOT: Her türlü ökaryotik hücre kültürlenebilir. Çoğu hücre tipi için, 10cm2'lik bir kabın kültürlenmesi yeterli hücrelere yol açar (genellikle >3 x 106 hücre). Hücresel metabolizma ve solunum üzerindeki etkilerden kaçınmak için mikoplazma enfeksiyonunu rutin olarak kontrol edin. - %90'ı aşmadan hücreleri büyütün (Şekil 2C).
NOT: %>90 akıcılığa sahip hücreler solunum üzerinde büyümeye bağlı inhibitör etkiler gösterebilir (senkronize veya post-mitotik değilse). - Hücreleri 1x PBS ile yıkayın, 1 mL ılık% 0.25 tripsin ile ayırın, ılık DMEM (5 mL / 10 cm2 plaka) ekleyerek tripsini devre dışı bırakın ve hücreleri bir hemositometre ile sayın.
- 5 dakika boyunca 300 x g'de 2,5 x 10 6 hücreye eşit hücre çözeltisini nazikçe santrifüj edin, süpernatantı tamamen çıkarın ve 2,5 mL sıcak MiR05 (1 x 10 6 hücre / mL) içinde yeniden askıya alın (Şekil 2A).
- Süspansiyon hücreleri için, 2.5 x 106 hücreye eşit çözeltiyi sayın ve çıkarın, pelet ve adım 1.2.4'te belirtildiği gibi devam edin.
- Geçirgenlik optimizasyonu (adım 1.6), geçirgenleştirilmiş hücre veya doku (adım 1.5) veya bozulmamış hücreler (adım 1.7) için SUIT protokolünü çalıştırın
NOT: Tutarlı sonuçlar için, hücre konsantrasyonunun sabit tutulması önerilir (örneğin, 1 x 106 hücre/mL). Solunum, respirometre24'teki hücre yoğunluğundan bağımsız olmasına rağmen, hücre sayıları sabit tutulursa, substratlar ve inhibitörler deneyler boyunca karşılaştırılabilir konsantrasyondadır.
- Kültür HEK293 hücreleri, Dulbecco'nun Modifiye Kartal ortamındaki (DMEM) 10cm2 çapındaki tabaklarda,% 10 ısı-inaktive fetal sığır serumu (FBS), GlutaMax, Esansiyel olmayan amino asitler ve Na-Piruvat22 ve idrar 23 ile desteklenmiş yüksek glikozlu, 37 ° C'de bir inkübatörde% 5 CO2'de OXPHOS kusurlu metabolizmayı desteklemek için.
- Yüksek çözünürlüklü respirometri için fibröz olmayan dokunun (örneğin beyin, karaciğer) hazırlanması
- Homojen bir doku parçasını, 30-40 mg ağırlığında tüketin veya tüm organı kullanın (bu durumda fare beyinciği).
NOT: Doku hemen kullanılmazsa, çoğu doku için 2 saate kadar koruma sağlayan 2 mL buz gibi soğuk MiR05'te saklayın. Bireysel doku depolama sürelerinin zaman serilerine göre değerlendirilmesi gerekir. - Dokuyu bir Whatman filtre kağıdı ile kurutun (dikkat: yumuşak doku maddesi yapışma eğilimindedir).
- 30-40 mg'lık doku parçasını buz soğutmalı 2 mL politetrafloroetilen çömlekçi Elvehjem homojenizatöre yerleştirin.
- Doku-tampon oranını korumak için 20 mg / mL elde etmek için uygun miktarda MiR05 ekleyin. Uygun mekanik geçirgenlik için yetersiz veya aşırı sıvıyı önlemek için toplam miktarı 1,5 mL ve >2 mL'< tutun.
- Havaneyi yerleştirin, havaneyi dikkatlice geri çekerek dokuyu yavaşça lize edin ve aşırı doku hasarına neden olan bir vakum oluşumunu önleyin.
- Lize edilene kadar toplamda 7 vuruş (bir yukarı ve aşağı vuruş olarak tanımlanan 1x) gerçekleştirin (büyük döküntü içermeyen bulanık bir sıvı olarak görünür) (Şekil 2B).
NOT: Uygun lizis için inme sayısı, sitokrom C yanıtı yoluyla dış mitokondriyal membran bütünlüğü değerlendirilerek her doku için test edilmelidir (adım 1.5.11). Lize edilmesi zor bağ dokusu veya damar parçaları kalabilir. - Lize edilmiş dokuyu 15 mL'lik bir santrifüj tüpüne boşaltın.
- Çömlekçinin içini lize adımında kullanılan eşit miktarda MiR05 ile yıkayın (örneğin, 1.5 mL) ve şimdi 10 mg / mL doku lizatında 3-4 mL MiR05 içeren 15 mL tüpe ekleyin.
- 37 °C'ye kadar ısıtmak için oda başına 2 mL düz MiR05 ekleyin.
- Soğuktan 37 ° C'ye kadar olan stresi en aza indirmek için oda başına her lizattan 500 μL (5 mg'a eşit) pipetleme yapmadan önce tüpü eşit dağılım için yavaşça döndürün.
- Odayı kapatmadan önce oda içeriğinin 37 °C'ye ısınması için >3 dakika bekleyin. Tıpanın üstündeki fazla sıvıyı çıkarın (kapandıktan sonra oda başına miktar: 4 mg).
- Standart geçirgenlik için SUIT protokolünü çalıştırın (adım 1.5).
- Homojen bir doku parçasını, 30-40 mg ağırlığında tüketin veya tüm organı kullanın (bu durumda fare beyinciği).
- Yüksek çözünürlüklü respirometri için fibröz dokunun (iskelet kası, kalp kası) hazırlanması
- Sert dokuyu çıkarın, bir diseksiyon mikroskobu altında 2 mL buz gibi soğuk BIOPS'ta (Tablo 2) keskin forseps kullanarak bağ dokusunu ve yağını kaslardan çıkarın.
- Lif demetlerini (~ 4 mg) uzunlamasına eksen boyunca keskin forsepslerle ayırın. Lifleri, ağ benzeri bir yapı elde etmek için yeterince gevşetin (Şekil 2B).
NOT: Uygun mekanik lif ayrımı ve geçirgenliği, kırmızı pigment miyoglobininin kaybı ve yarı saydamlığın artması ile gösterilir. - Lif demetini saponin içinde (BIOPS'ta 50 μg / mL, taze hazırlanmış) 4 ° C'de 20 dakika boyunca yıkayın ve geçirgenleştirin (lifler yarı saydam hale gelir, tam geçirgenleşmeyi gösterir, Şekil 2B).
- Lifleri MiR05'te iki kez 4 ° C'de yıkama başına 5 dakika boyunca yıkayın.
- Filtre kağıdı ile kurutun ve 2,1 mL MiR05 ile doldurulmuş hazneye eklemeden önce tartın.
- Tapaları tamamen kapatmadan tanıtın, ardından 20 mL'lik bir şırınga kullanarak odaları2 mL saf O2 ile oksijenlendirin ve tıpaları dönen bir hareketle bükerek odaları kapatın. Oksijen difüzyon sınırlamasını önlemek için deney sırasındaO2 konsantrasyonunu 300-500 μM arasında tutun.
- Hücrelerde veya dokularda rutin solunumu değerlendirmek için protokol
- 1.5.2-1.5.3 numaralı adımlarda belirtildiği gibi odaya numune ekleyin.
- 2.3 mL sıcak MiR05 hücre süspansiyonu ekleyin (standart giriş: adım 1.2'de olduğu gibi 1 x 106 hücre / mL veya adım 1.3'te olduğu gibi 2 mg doku / mL)
- İskelet ve kalp kası (adım 1.4): 1.4.4-1.4.6 adımlarını göz önünde bulundurarak önceden ısıtılmış 2.3 mL sıcak MiR05'e ~ 4 mg saponin-geçirgenleştirilmiş lif ekleyin
- Odaları 37 °C'de ve 700 rpm karıştırma hızında çalıştırın. Tıpayı dönen bir hareketle döndürerek ortamın gazdan arındırılmasına ve haznelerin kapanmasına izin vermek için >3 dakika bekleyin. Peltier blok stabilizasyonu, ayarlanan sıcaklığa ulaşıldığını gösterir.
- (İSTEĞE BAĞLI) Kalan kabarcıkların tıpanın kılcal damarından kaçmasına izin vermek için karıştırıcı hızını 300 rpm olarak değiştirin.
- Tıpanın üstündeki fazla sıvıyı aspire edin. Rutin/durum 1 solunumu kaydetmek için herhangi bir numune türüyle kararlı bir oksijen akısı sinyali elde edilene kadar 10 dakika bekleyin, Şekil 1B).
- Geçirgenleştirilmiş hücrelerde ve dokuda solunum ölçümleri için, adım 1.6 ile devam edin. Adım 1.8 ile sağlam hücreler için.
- Geçirgenleştirilmiş hücrelerde veya dokularda OXPHOS analizi için protokol
- Hücreleri geçirgenleştirmek için 5 μg / mL'lik son konsantrasyon için lize (geçirgenleştirilmiş) doku örneği kullanın veya hücreleri geçirgenleştirmek için 1 μL sayısallaştırma (dimetil sülfoksit (DMSO) içinde 8.1 mM sayısalonin stoğu) ekleyerek hücreleri geçirgenleştirin. Akı düşecek ve >5 dakikada stabilize olmalıdır.
DİKKAT: Digitonin, solunum yolları, cilt ile temas halinde veya yutulduğunda akut toksiktir.
NOT: Tüm kimyasalların enjeksiyonu hassas cam şırıngalarla yapılır. Şırıngaları çapraz kontaminasyonu önlemek için sadece belirtilen kimyasallar için kullanın ve kullanımdan sonra su ve EtOH ile iyice yıkayın. Tıkanmış şırıngalar, sıcak ddH2O'da ultrasonikasyon veya herhangi bir kimyasal tıkanıklığı yerinden çıkarmak için bir temizleme teli gerektirebilir. Odalara hava girmesini önlemek için her zaman ilgili stok çözeltisinin bir fazlasını şırıngaya geri çekin. Her enjeksiyondan sonra hava girişi için odaların içini kontrol edin. Akı platolarına kadar her adımı kaydedin. - Hızlı bir şekilde art arda ekleyin: 1 mM'lik bir nihai konsantrasyon için 5 μL 0.4 M malat (M), 5 mM'lik bir son konsantrasyon için 5 μL, 2.0 M piruvat (P; taze olarak hazırlanmış), 5 mM'lik bir son konsantrasyon için 4 μL 2.5 M glutamat (G).
- Önceki akı platolarından sonra, 1.25 mM'lik bir son konsantrasyon için 0.5 M adenozin difosfat (ADP, -80 ° C'de depolanan alikotlar) 5 μL (kas dokusu için 10 μL) ekleyin.
NOT: Kas gibi dokuların doygunluğa ulaşması için farklı bir konsantrasyona ihtiyacı olabilir. - 10 μM'lik son konsantrasyon için 5 μL 4 mM sitokrom C (cytC) ekleyin.
NOT: Hücrelerin geçirgenlik kalitesini değerlendirmesi için isteğe bağlıdır. - 10 mM'lik son konsantrasyon için 16 μL 1,25 M süksinat (S) ekleyin. (İSTEĞE BAĞLI) ADP konsantrasyonunun doygunluğunu kontrol etmek için 2 mM'lik bir son konsantrasyon için 3 μL 0,5 M ADP ekleyin.
- Hücreler ve lifli olmayan dokular için, 1 μg / mL'lik son konsantrasyon için 2 μL 1 mg / mL oligomisin (OM) ekleyin.
DİKKAT: Kullanılan tüm ETS inhibitörleri oldukça toksiktir.
NOT: Oligomisin, ETS kapasitesini baskılayabildiği ve kas dokusu için ihmal edildiği için optimum konsantrasyon için titrasyon gerektirebilir. Kas dokusu test edildiğinde veO2 300 μM'nin altındaysa burada oksijenlenin. - FCCP'yi 2 mM'lik bir stoktan titre edin, solunum ve solunumda maksimum olarak ayrılmayana kadar (teorik: bağlanmamış) 0,2 μL adımlarla 0,6 μL ekleyin.
- 0.5 μM'lik son konsantrasyon için 1 μL'lik 1 mM rotenon (ROT) ekleyin. 1 μg / mL'lik bir son konsantrasyon için 2 μL 1 mg / mL antimisin () stoğu ekleyin.
- Pistonu büküm hareketiyle yavaşça kaldırarak tüm odalarda benzer bir oksijen seviyesine (~ 150 μM) ulaşmak için odaları yeniden oksijenlendirin.
- 2 mM'lik bir son konsantrasyon için 5 μL 0.8 M askorbat ekleyin ve hemen ardından kompleks IV aktivitesini değerlendirmek için 0.5 mM'lik bir son konsantrasyon için 5 μL 0.2 M N, N, N′, N′-tetrametil-p-fenilendiamin (TMPD) ekleyin (isteğe bağlı).
- TMPD ile tepeO2 akısına ulaşıldığında hemen 10 mM'lik son konsantrasyon için 5 μL 4 M azid ekleyin. Karmaşık IV baz seviyesi hesaplaması için TMPD'nin otomatik oksidasyonunu test etmek üzere >5 dakika boyunca çalışmaya devam edin.
- Hücre sayımını çalıştırma öncesi onaylamak için hücreleri yeniden sayın ve adım 1.9 ile devam edin.
NOT: Digitonin-geçirgenleştirmenin (yalnızca hücreler için), maksimum akıya ulaşmak ve mitokondriyal membran bütünlüğünü etkilememesi için deneme deneylerinde titre edilmesi gerekir (bkz. adım 1.7). Sitokrom c ilavesinden sonra solunum hızında %>10 artış olan geçirgen numuneler (özellikle kas dokusu), dış mitokondriyal membran hasarı nedeniyle ileri analizlerden çıkarılmalıdır. EtOH çözünmüş kimyasalların eklenmesinden sonra akıda kısa süreli bir düşüş beklenir.
- Hücreleri geçirgenleştirmek için 5 μg / mL'lik son konsantrasyon için lize (geçirgenleştirilmiş) doku örneği kullanın veya hücreleri geçirgenleştirmek için 1 μL sayısallaştırma (dimetil sülfoksit (DMSO) içinde 8.1 mM sayısalonin stoğu) ekleyerek hücreleri geçirgenleştirin. Akı düşecek ve >5 dakikada stabilize olmalıdır.
- Hücreler için optimal geçirgenlik koşullarını belirlemek için protokol
- Adım 1.2 ve 1.5.2'de açıklandığı gibi hücreler ekleyin.
- 10 mg / mL digitonin stoğundan 10 μL alın ve 5 mg / mL'ye seyreltmek için 10 μL DMSO ekleyin.
- 1 μL rotenon ekleyin (1 mM stok). 10 μL süksinat (2 mM stok) ve 5 μL ADP (0,5 M stok) ekleyin.
- Solunum daha fazla artmayana ve maksimal olana kadar tekrar tekrar 1 μL digitonin (adım başına 2.5 mg) titre edin.
NOT: Solunumdaki bir azalma, aşırı miktarda digitonin konsantrasyonunu gösterir.
- Sağlam hücrelerde OXPHOS analizi için protokol
- Rutin solunumdan sonra (adım 1.6.1-1.6.6), 10 nM'lik son konsantrasyon için 2 μL 0.01 mM oligomisin ekleyin.
- FCCP'yi 2 mM stoktan titratın, solunumda daha fazla artış olmayıncaya kadar sonraki 0,2 μL adımlarla 0,6 μL ekleyin ve solunum maksimum olarak ayrılamaz (teorik: bağlanmamış)
- 0.5 μM'lik son konsantrasyon için 1 μL 1 mM rotenon ekleyin. 1 μg / mL'lik bir son konsantrasyon için 2 μL 1 mg / mL antimisin stoğu ekleyin.
- Pistonu büküm hareketiyle yavaşça kaldırarak odayı aynı oksijen seviyesine (~ 150 μM) yeniden oksijenlendirin.
- 2 mM'lik son konsantrasyon için 5 μL 0.8 M askorbat ekleyin. Kompleks IV aktivitesini değerlendirmek için derhal 0,5 mM'lik bir son konsantrasyon için 5 μL 0,2 M TMPD ekleyin.
NOT: TMPD otomatik oksidasyona eğilimli olduğundan daha büyük bir deney setinden önce yeni bir parti hazırlayın. Etkinlik, -20 °C'de depolandığında zamanla azalabilir. - TMPD ile tepeO2 akısına ulaşıldığında hemen 10 mM'lik son konsantrasyon için 5 μL 4 M azid ekleyin. Karmaşık IV baz seviyesi hesaplaması için TMPD'nin otomatik oksidasyonunu test etmek üzere >5 dakika boyunca çalışmaya devam edin.
- Çalıştırma öncesi hücre sayısını onaylamak için hücreleri yeniden sayın ve adım 1.9 ile devam edin.
- Çalıştırma sonrası örnek koleksiyonu
- Her odadan (karıştırıcılar açıkken) tam olarak 1 mL MiR05 süspansiyonunu 1,5 mL'lik bir tüp 1,5 mL'lik bir tüpe toplayın.
- Geçirgen hücreler için 1000 x g'da veya doku lizatı için 20.000 x g'da santrifüj. Süpernatantı çıkarın ve daha fazla işlem için peleti -80 °C'de dondurun (bölüm 3).
- SUIT protokollerinin analizi
- Bir substrat veya inhibitör ekledikten sonra her platodaki oksijen akışını (pmol/s, girişe normalleştirilmiş) analiz edin (Şekil 1C ve Şekil 3A). Değerleri bir e-tabloya dışa aktarın.
- Artık oksijen tüketimi (ROX, Şekil 1C ve Şekil 3C) değerini her deneysel çalışmanın tüm değerlerinden çıkarın. Kompleks IV solunum elde etmek için TMPD'den azid kalıntı solunumu çıkarın.
- Hücre (Şekil 3A, B) veya doku girişi (Şekil 5A,B) için normalleştirilmiş mutlak değerleri çizin. Akı kontrol oranlarını hesaplayın (adım 1.11) veya bunları protein girişine normalleştirin (Şekil 3C).
- Akı kontrol oranı hesaplaması
- Akı kontrol oranlarını (FCR)9,26 kullanarak solunum fonksiyonu indeksi ve bağlantı kontrolü elde edin.
NOT: Bu, mitokondriyal miktardan bağımsız olarak içsel mitokondriyal kalitenin değerlendirilmesine izin verir. Ek olarak, akı kontrol oranları (FCR), reaktif kalite kontrolüne izin veren aynı hücre hatları içinde karşılaştırılabilir (ilgili FCR'ler, Şekil 1B-D ve Şekil 3C'de belirtilen numaralı referans değerleri aracılığıyla elde edilir). - Denklem 1'i kullanarak OXPHOS'un LEAK'e bağlanması için solunum kontrol oranını hesaplayın.
Denklem 1: FCRADP = 5/6 = Durum 3 / Durum 4 - Denklem 2'yi kullanarak NADH'ye bağımlı solunumu değerlendirmek için FCR'yi hesaplayın
Denklem 2: FCR durum 3 (I) = 3/5 = Durum 3 (I) / Durum3 ( I+II) - Denklem 3'ü kullanarak süksinat bağımlı solunumu değerlendirmek için FCR'yi hesaplayın.
Denklem 3: FCRdurumu 3 (II) = 8/7 = Sçürüme / ETSkapasitesi - Denklem 4'ü kullanarak birleştirilmiş ve ayrıştırılmamış olarak değerlendirmek için FCR'yi hesaplayın.
Denklem 4: FCReşli/bağlanmamış = 5/7 = Durum 3 (I+II) / ETSkapasitesi - Mitokondriyal dış membran bütünlüğünü test etmek için Denklem 5'i kullanın.
Denklem 5: % mitokondriyal dış zar hasarı = 3/4 = Durum 3 (I) / Durum 3 (I) cyt c ile
- Akı kontrol oranlarını (FCR)9,26 kullanarak solunum fonksiyonu indeksi ve bağlantı kontrolü elde edin.
2. Yüksek çözünürlüklü respirometri: Mikroplaka bazlı respirometre (mHRR)
NOT: Protokolün bu bölümündeki deneyler, Seahorse XFe96 Hücre Dışı Akı Analizörü (Malzeme Tablosu) kullanılarak gerçekleştirilmiştir.
- Hücre kültürü
- Her türlü hücreyi kültüre alın. Hücre bağlanmasını kolaylaştırmak için aderanlar (örneğin, kollajen, laminin) kullanılabilir. Burada, HEK293 hücreleri daha önce olduğu gibi kültürlenir (adım 1.3).
- Deneyden bir gün önce, deney gününde (% 80-% 100) ideal akıcılık elde etmek için hücreleri ayırın ve belirlenmiş bir mHRR 96 delikli mikro plakaya aktarın (Şekil 2C).
NOT: mHRR için mikroplaka hücre yoğunlukları kritiktir. Hücre hatlarının bireysel büyüme özellikleri veya büyümeyi etkileyen tedaviler, deney gününde karşılaştırılabilir bir akıcılık sağlamak için hesaba katılmalıdır.
- Yüksek çözünürlüklü respirometri için hücrelerin hazırlanması
- Tohumlamadan önce hücreleri yeterince hasat edin ve yeniden askıya alın
NOT: Replikasyonlar için hücrelerin aynı seyreltmeden tohumlanması önerilir. - Hücreleri, bireysel hücre hatlarının büyüme oranlarına veya tedavi altındaki büyüme özelliklerine göre tohumlayın.
NOT: Standart bir 96 delikli mikro plaka üzerinde optimizasyon yapın ve hücre yoğunluğunu 96 kuyucuklu tahlil spesifik bir mikroplakaya tahmin edin. Bu kurulumda, 96 kuyucuğun kuyusu başına 7 x 104 HEK293 WT hücresi tohumlandı. 96 delikli plakanın ilk ve son sütunları protein tayini için kullanılır (Şekil 2C). Dört köşe kuyusu herhangi bir hücre içermemelidir ve deneysel arka plan düzeltmesi için kullanılır. İdeal olarak, kenar etkisini en aza indirmek için kenarlara yakın kuyular boştur (örneğin; hücreler sıcaklık etkilerinin neden olduğu değişmiş büyüme hızlarını gösterir) (Şekil 2C, D).
- Tohumlamadan önce hücreleri yeterince hasat edin ve yeniden askıya alın
- Sensör plakalarının hazırlanması, inhibitörlerin yüklenmesi
- Tahlil gününde, 0.4 mL 1 M Glikoz, 0.4 mL 200 mM Glutamin ve 0.4 mL 100 mM Na-Piruvat ile 38.8 mL orta takviye edin.
NOT: mHRR solunum, pH 7.4'te özel tamponsuz DMEM ortamı gerektirir. Genel olarak, 96 delikli bir mikroplaka ile yapılan bir deney için 40 mL yeterli olmalıdır. - Solunum testi ortamını 37 ° C'ye ısıtın ve hücre kültürü ortamını, kuyucuk başına 80 μL ile iki kez yıkayarak solunum testi ortamı ile değiştirin.
- Plakayı hücrelerle birlikte tahlilden önce 60 dakika boyunca CO2 içermeyen 37 ° C'lik bir inkübatöre yerleştirin.
NOT: Bu adım, CO2 solunum sonuçlarını etkileyebileceğinden ve ortamdaki serum tahlil sırasında kabarcıklar üretebileceğinden, plakanın gazını gidermek için gereklidir. - Ön ısınma inhibitörü OM, FCCP, ROT ve için 37 ° C'ye kadar aliquots ve sensör plakasını inkübatörden çıkarın.
- OM, FCCP, ROT ve'yi 3 mL tahlil ortamında sırasıyla 1.5 μM, 1.125 μM ve 1 μM'lik son kuyucuk konsantrasyonuna kadar seyreltin. Şekil 2E'de gösterildiği gibi ayrı bağlantı noktalarına doldurun.
NOT: Sensör kartuşunu doldurmak için çok kanallı pipet kullanılması önerilir. Bileşikleri enjekte etmek için basınçlı hava kullanıldığından, bir port bir bileşikle doldurulduğunda tüm portlar eşit miktarda sıvı hacimle doldurulmalıdır. ROT ve tek bir bağlantı noktasında birleştirilebilir. İnhibitörler EtOH veya DMSO'da çözülebilir. - Enjeksiyon portlarını inceleyin ve her port için eşit bir yükleme hacmini doğrulayın.
NOT: Tüm portların altında enjeksiyon için bir delik bulunur. Sensör plakasını hareket ettirirken dikkatli olunmalıdır. Hava kabarcıkları bir iğne kullanılarak çıkarılabilir.
- Tahlil gününde, 0.4 mL 1 M Glikoz, 0.4 mL 200 mM Glutamin ve 0.4 mL 100 mM Na-Piruvat ile 38.8 mL orta takviye edin.
- Sağlam hücrelerde oksijen değerlendirmesi için protokol
- Tahlilden önceki gün, 2.4.2-2.4.7 adımlarını uygulayın.
- Kalibrant çözeltisinin 20 mL'lik bir kısmını 50 mL'lik konik bir tüpe boşaltın.
- Hücre Dışı Akı Testi Kiti'ni açın ve içindekileri çıkarın.
- Sensör kartuşunu ters çevrilmiş olarak yardımcı plakanın yanına yerleştirin. Pipet, yardımcı plakanın her bir kuyucuğuna 200 μL kalibrant çözeltisi yerleştirin.
- Tüm sensörlerin suya batırılmasına dikkat ederek sensör kartuşunu yardımcı plakaya takın.
- Plakayı gece boyunca CO 2 olmadan veya en az12 saat boyunca 37 ° C'lik bir inkübatöre yerleştirin. İnkübatörün içindeki nemin, kalibrantın buharlaşmasını önlemek için yeterli olduğunu doğrulayın.
- Ertesi gün kullanıma hazır olması için mikroplaka tabanlı sistemi ve bilgisayarı açın (makinenin bir tahlil yapmadan önce 37 ° C'ye dengelenmesi için en az 3 saat gerekir).
NOT: Sinyal kararlılığı için, ölçüm noktalarını solunum durumu başına 3 ölçüm döngüsü yerine 6'ya yükseltin. Her döngü 3 dakikalık karıştırma ve 3 dakikalık ölçümlemeden oluşur. - XF testi gününde, 2.4.9-2.4.20 adımlarını uygulayın.
- Hücre kültürü plakasının akıcılığını, hücrelerin morfolojisini ve arka plan kuyularının boş olduğunu doğrulayın.
- Hücreleri, 2.4.11-2.4.12 adımlarında belirtildiği gibi hazırlanan solunum ortamıyla yıkayın.
- Kültür ortamının 20 μL hariç tamamını her bir kuyucuktan çıkarın. Kültür ortamı gece boyunca buharlaşma nedeniyle 80 μL ise (yaklaşık 5 μL) 55 μL'yi çıkarın.
- Hücreleri 90 μL tahlil ortamı ile iki kez yıkayın. Son olarak, 100 μL tahlil ortamı ekleyin. Son hacim 120 μL olmalıdır.
NOT: Her deneysel duruma aynı yıkama prosedürünün uygulandığından emin olmak için bu adımda çok kanallı bir pipet önerilir (plaka kurulumuna bağlıdır). Aspire ederken, plakayı 45° açıyla eğin ve pipet uçlarını aspirasyon ve sıvı enjeksiyonu için kuyucukların köşesine yerleştirin. Bazı hücreler hücre kültürü plakasının dibinden kolayca ayrılabileceğinden, yıkama sırasında dikkatli olmak zorunludur. - Plakayı, tahlilden önce 60 dakika boyunca CO2 içermeyen 37 ° C'lik bir inkübatöre yerleştirin.
- Hidratlanmış sensör kartuş plakasını CO2 içermeyen inkübatörden alın.
- Eski kalibrant çözeltisini atın ve 37 ° C'ye kadar önceden ısıtılmış taze kalibrant çözeltisi ile değiştirin.
- İnhibitörleri ve tahlil ortamını hazırlayın (toplam 12 mL tahlil ortamı için inhibitör başına 3 mL) ve inhibitörün portlara yüklenmesi için bir pipet rezervuarı kullanın.
- Yazılımı açın ve önceden tasarlanmış veya yeni bir şablon çalıştırın. Plaka haritasını doldurun, titrasyonları ve ölçüm döngülerini ayarlayın ve ardından optik sensörlerin kalibrasyonunu başlatmak için Başlat'a basın.
- Kapağı yüklü kartuştan çıkarın ve otomatik olarak makineden dışarı kayan yuvaya yerleştirin ve plakanın sağ alt köşesindeki işaretlerin yuvanın sağ alt köşesindeki üçgenle aynı hizada olduğunu doğrulayın.
- Yaklaşık 20 dakika süren otomatik kalibrasyonu gerçekleştirmek için Devam'a tıklayın.
- Kalibrasyondan sonra, kalibrant içeren yardımcı plakayı çıkarın.
- Kapağı hücreleri içeren mikro plakadan çıkarın ve makine tarafından istendiğinde plakayı yuvaya yerleştirin. Çalıştırmayı başlatmak için Devam'a tıklayın.
- Çalıştırma sonrası örnek koleksiyonu
- Plakayı makineden çıkarın, hücreleri rahatsız etmeden kalan tahlil ortamını dikkatlice çıkarın ve daha fazla işlem için tüm plakayı -80 ° C'de dondurun (bölüm 3).
3. Bisinkoninik asit testi (BCA testi) kullanılarak protein tayini
- Protein ekstraksiyonu için kullanılan ve BCA ile uyumlu tamponda seyreltilmiş sığır serum albümini (BSA) hazırlayın: 2 mg / mL, 1.5 mg / mL, 1 mg / mL, 0.5 mg / mL, 0.5 mg / mL, 0.25 mg / mL ve 0 mg / mL.
- mHRR için kuyucuk başına 20 μL veya cHRR için 1,5 mL'lik bir tüp içinde bulunan pelet başına 100 μL ile uygun bir lizis tamponunda (örneğin, RIPA) yeniden süspansiyon yaparak proteinleri ekstrakte edin.
- mHRR plakasını veya protein lizatları içeren 1,5 mL tüpü buz üzerinde 30 dakika boyunca inkübe edin.
- Protein lizatını içeren 1,5 mL'lik tüpü 20 dakika boyunca 20.000 x g'de 4 ° C'de santrifüj edin ve elde edilen süpernatantı yeni bir temiz 1,5 mL tüpe aktarın.
- Kopyalarda numune başına 10 μL kullanın ve bir mikrotitre plakasında standartlar kullanın. 200 μL BCA çalışma reaktifi ekleyin ve >15 dakika kuluçkaya yatırın.
- Plakayı 562 nm dalga boyunda standart bir spektrofotometrede okuyun ve BSA standart eğrisini kullanarak protein konsantrasyonlarını hesaplayın.
- Solunum sonuçlarını protein konsantrasyonuna normalleştirin.
NOT: Protein miktarına normalleştirme, hücre tohumlama yoğunluklarını veya ıslak ağırlık girişini doğrulamaya izin verir. Ekstrakte edilen proteinler, örneğin ETS'nin alt birimlerine karşı sonraki immünoblotlama için uygundur, ancak doğal numuneyi tam olarak temsil etmez (örneğin, fosforilasyon bölgelerinin kaybı).
Sonuçlar
Burada, ökaryotik hücrelerdeki, fibröz olmayan dokudaki (örneğin, beyincik) ve fibröz dokudaki (örneğin, iskelet kası) mitokondriyal biyoenerjetik maddeleri belirlemek için protokoller sunuyoruz. Ökaryotik hücreler için, mitokondriyal translasyonla ilişkili iki farklı proteinin CRISPR mühendisliği nakavtına sahip HEK293, çoklu (CRISPRKO1) ve şiddetli / tam OXPHOS eksikliği (CRISPRKO2) ile sonuçlanan cHRR (Şekil 3A-C) veya mHRR (Şekil 3A-D...
Tartışmalar
Geleneksel olarak, mitokondriyal biyoenerjetik Clark tipi oksijen elektrotları ile çalışılmıştır. Bununla birlikte, çözünürlük ve verim eksikliği, teknolojik ilerleme için garanti edildi. Bugüne kadar, O2k (cHRR olarak adlandırılır) ve Seahorse XF96 Akı Analizörü (mHRR olarak adlandırılır) hücresel biyoenerjetik alanında yaygın olarak benimsenmiştir. Burada, cHRR veya mHRR kullanarak mitokondriyal solunumun değerlendirilmesi yoluyla hücresel enerji metabolizmasının analizi için anlaşıl...
Açıklamalar
Açıklanacak çıkar çatışması yok.
Teşekkürler
Bu çalışma, Finlandiya Akademisi (CBJ), Magnus Ehrnroot Vakfı (CBJ) ve Entegre Yaşam Bilimleri Enstitüsü (RA) Doktora bursu ile desteklenmiştir.
Malzemeler
| Name | Company | Catalog Number | Comments |
| 2 mL Potter-Elvehjem Glass/PTFE Tissue Grinder/Homogenizer | Omni International | 07-358029 | |
| 95% O2, 5% CO2 medical gas mixture | Potter for tissue grinding | ||
| ADP | Sigma | A 4386 | |
| Antimycin A | Sigma | A 8674 | Chemical |
| Ascorbate | Merck | PHR1279-1G | Chemical, dissolve in ethanol |
| BSA (fatty accid free) | Sigma | A 6003 | Chemical |
| CaCO3 | Sigma | C 4830 | Chemical |
| Cytochrome c | Sigma | C 7752 | Chemical |
| Digitonin | Sigma | D 5628 | Chemical |
| Dithiothreitol | Sigma | D 0632 | Chemical, dissolve in DMSO |
| D-Sucrose | Roth | 4621.1 | Chemical |
| Dulbecco’s modified Eagle’s medium (High glucose) | Fisher Scientific | 41965-039 | Chemical |
| Dulbecco’s modified Eagle’s medium (No Glucose) | Fisher Scientific | A14430-01 | |
| EGTA | Sigma | E 4378 | |
| Etomoxir | Sigma | E1905 | Chemical |
| Falcon 15 ml Conical Centrifuge Tubes | Fisher Scientific | AM12500 | Chemical |
| Falcon 50 ml Conical Centrifuge Tubes | Fisher Scientific | AM12501 | |
| FCCP | Sigma | C 2920 | |
| Glucose | Sigma | G7021 | Chemical, dissolve in ethanol |
| Glutamate | Sigma | G 1626 | Chemical |
| GlutaMax (100x) (200 nM L-alanyl-L-glutamine dipeptide) | Fisher Scientific | 35050061 | Chemical |
| HEK293 cells | ATTC | CRL-1573 | |
| Hemocytometer | Fisher Scientific | 0267151B | Instrument for cell counting |
| Hepes | Sigma | H 7523 | Chemical |
| Imidazole | Fluka | 56750 | Chemical |
| KCl | Merck | 1.04936 | Chemical |
| L-carnitine | Sigma | C0283 | Chemical |
| Malate | Sigma | M 1000 | Chemical |
| MES hydrate | Sigma | M8250 | Chemical |
| MgCl2 | Sigma | M 9272 | Chemical |
| Na2ATP | Sigma | A 2383 | Chemical |
| Na2Phosphocreatine | Sigma | P 7936 | Chemical |
| Na-pyruvate (100 mM) (100x) | Fisher Scientific | 11360070 | |
| NEAA (Non-essential amino acids) 100x | Fisher Scientific | 11140035 | |
| Normal FBS (10x) | Fisher Scientific | 10500064 | |
| O2k-Core: Oxygraph-2k | Oroboros Instruments | 10000-02 | High-resolution respirometry instrument |
| O2k-Titration Set | Oroboros Instruments | 20820-03 | Hamilton syringes for chemical injections |
| Oligomycin | Sigma | O 4876 | Chemical, dissolve in ethanol |
| Palmitoylcarnitine | Sigma | P 4509 | Chemical |
| Penicillin-Streptomycin | Fisher Scientific | 15140122 | |
| Pierce BCA Protein Assay Kit | Fisher Scientific | 23227 | |
| Pyruvate | Sigma | P 2256 | Chemical |
| RIPA-Buffer | Fisher Scientific | 89900 | Chemical |
| Rotenone | Sigma | R 8875 | Chemical, dissolve in ethanol |
| Saponin | Sigma | S7900 | Chemical |
Seahorse XF DMEM assay medium pack, pH 7.4 | Agilent, Santa Clara, CA | 103680-100 | |
| Seahorse XFe96 Extracellular Flux Analyzer | Agilent, Santa Clara, CA | High-throughput respirometry instrument | |
| Seahorse XFe96 FluxPak | Agilent, Santa Clara, CA | Includes assay plates, cartridges, loading guides for transferring compounds to the assay cartridge, and calibrant solution. | |
| Small scissors | Fisher Scientific | 08-951-20 | |
| Sodium azide | Sigma | S2002 | Chemical |
| Succinate | Sigma | S 2378 | Chemical |
| Taurine | Sigma | T 8691 | Chemical |
| TMPD | Sigma | T 3134 | Chemical |
| Trypan Blue solution | Merck | 72-57-1 | Chemical |
| Trypsin 0.25% EDTA | Fisher Scientific | 25200056 | |
| Two thin-edged forceps | Fisher Scientific | 12-000-122 | |
| Uridine stock (500x) | Sigma | U3750 | Chemical |
Referanslar
- McBride, H. M., Neuspiel, M., Wasiak, S. Mitochondria: More than just a powerhouse. Current Biology. 16 (14), 551-560 (2006).
- Mehta, M. M., Weinberg, S. E., Chandel, N. S. Mitochondrial control of immunity. Beyond ATP. Nature Reviews Immunology. 17 (10), 608-620 (2017).
- Spinelli, J. B., Haigis, M. C. The multifaceted contributions of mitochondria to cellular metabolism. Nature Cell Biology. 20 (7), 745-754 (2018).
- Gnaiger, E. Capacity of oxidative phosphorylation in human skeletal muscle. New perspectives of mitochondrial physiology. International Journal of Biochemistry and Cell Biology. 41 (10), 1837-1845 (2009).
- Gorman, G. S., et al. Mitochondrial diseases. Nature Reviews Disease Primers. 2, 1-23 (2016).
- Boushel, R., Gnaiger, E., Schjerling, P., Skovbro, M., Kraunsøe, R., Dela, F. Patients with type 2 diabetes have normal mitochondrial function in skeletal muscle. Diabetologia. 50 (4), 790-796 (2007).
- Cogliati, S., et al. Mitochondrial cristae shape determines respiratory chain supercomplexes assembly and respiratory efficiency. Cell. 155 (1), 160-171 (2013).
- Kühlbrandt, W. Structure and function of mitochondrial membrane protein complexes. BMC Biology. 13, 1-11 (2015).
- Gnaiger, E. Mitochondrial pathways and Respiratory control. An introduction to OXPHOS analysis. Bioenergetics communications. 5th ed. , (2020).
- Jackson, C. B., et al. Mutations in SDHD lead to autosomal recessive encephalomyopathy and isolated mitochondrial complex II deficiency. Journal of Medical Genetics. 51 (3), 170-175 (2014).
- Pesta, D., Gnaiger, E. High-resolution respirometry: OXPHOS protocols for human cells and permeabilized fibers from small biopsies of human muscle. Methods in Molecular Biology. 810, 25-58 (2012).
- Horan, M. P., Pichaud, N., Ballard, J. W. O. Review: Quantifying mitochondrial dysfunction in complex diseases of aging. Journals of Gerontology - Series A Biological Sciences and Medical Sciences. 67 (10), 1022-1035 (2012).
- Doerrier, C., Garcia-Souza, L. F., Krumschnabel, G., Wohlfarter, Y., Mészáros, A. T., Gnaiger, E. High-resolution fluorespirometry and oxphos protocols for human cells, permeabilized fibers from small biopsies of muscle, and isolated mitochondria. Methods in Molecular Biology. 1782, 31-70 (2018).
- Zhang, J., et al. Measuring energy metabolism in cultured cells, including human pluripotent stem cells and differentiated cells. Nature Protocols. 7 (6), 1068-1085 (2012).
- García-Roche, M., Casal, A., Carriquiry, M., Radi, R., Quijano, C., Cassina, A. Respiratory analysis of coupled mitochondria in cryopreserved liver biopsies. Redox Biology. 17, 207-212 (2018).
- Acin-Perez, R., et al. A novel approach to measure mitochondrial respiration in frozen biological samples. The EMBO Journal. 39 (13), 1-18 (2020).
- Cell metabolism assay kits. Seahorse assay kits and media Available from: https://www.agilent.com/en/product/cell-analysis/real-time-cell-metabolic-analysis/xf-assay-lits-reagents-cell-assay-media (2021)
- Chance, B., Williams, G. R. A method for the localization of sites for oxidative phosphorylation. Nature. 176 (4475), 250-254 (1955).
- Gnaiger, E., et al. Mitochondrial respiratory states and rates. MitoFit Preprint Arch. , (2019).
- Gnaiger, E. O2k-procedures: SOP O2k quality control 1: Polarographic oxygen sensors and accuracy of calibration Section Page. Oroboros. 03 (18), 1-21 (2020).
- Robinson, B. H., Petrova-Benedict, R., Buncic, J. R., Wallace, D. C. Nonviability of cells with oxidative defects in galactose medium: A screening test for affected patient fibroblasts. Biochemical Medicine and Metabolic Biology. 48 (2), 122-126 (1992).
- King, M. P., Attardi, G. Human cells lacking mtDNA: Repopulation with exogenous mitochondria by complementation. Science. 246 (4929), 500-503 (1989).
- Makrecka-Kuka, M., Krumschnabel, G., Gnaiger, E. High-resolution respirometry for simultaneous measurement of oxygen and hydrogen peroxide fluxes in permeabilized cells, tissue homogenate and isolated mitochondria. Biomolecules. 5 (3), 1319-1338 (2015).
- Fasching, M., Gnaiger, E. O2k quality control 2: Instrumental oxygen background correction and accuracy of oxygen flux. Mitochondrial Physiology Network. 14 (06), 1-14 (2016).
- Gnaiger, E., Lassnig, B., Kuznetsov, A., Rieger, G., Margreiter, R. Excess capacity of cytochrome c oxidase. Journal of Experimental Biology. 1139, 1129-1139 (1998).
- Gnaiger, E., et al. Mitochondria in the Cold. Life in the Cold. , 431-442 (2000).
- Fontana-Ayoub, M., Fasching, E., Gnaiger, Selected media and chemicals for respirometry with mitochondrial preparations. Mitochondrial Physiology Network. 02 (17), 1-9 (2014).
- Gerencser, A. A., et al. Quantitative microplate-based respirometry with correction for oxygen diffusion. Analytical Chemistry. 81 (16), 6868-6878 (2009).
- Krumschnabel, G., Eigentler, A., Fasching, M., Gnaiger, E. Use of safranin for the assessment of mitochondrial membrane potential by high-resolution respirometry and fluorometry. Methods in Enzymology. 542, 163-181 (2014).
- Nászai, A., Terhes, E., Kaszaki, J., Boros, M., Juhász, L. Ca(2+)N it be measured? Detection of extramitochondrial calcium movement with high-resolution fluorespirometry. Scientific Reports. 9 (1), 1-13 (2019).
- Pajak, B., et al. 2-Deoxy-d-Glucose and its analogs: From diagnostic to therapeutic agents. International Journal of Molecular Sciences. 21 (1), 234 (2019).
- Mercier-Letondal, P., Marton, C., Godet, Y., Galaine, J. Validation of a method evaluating T cell metabolic potential in compliance with ICH Q2 (R1). Journal of Translational Medicine. 19 (1), 1-15 (2021).
- Sauerbeck, A., et al. Analysis of regional brain mitochondrial bioenergetics and susceptibility to mitochondrial inhibition utilizing a microplate based system. Journal of Neuroscience Methods. 198 (1), 36-43 (2011).
- Jackman, M. R., Willis, W. T. Characteristics of mitochondria isolated from type I and type IIb skeletal muscle. American Journal of Physiology - Cell Physiology. 270 (2), 673-678 (1996).
- Ponsot, E., et al. Mitochondrial tissue specificity of substrates utilization in rat cardiac and skeletal muscles. Journal of Cellular Physiology. 203 (3), 479-486 (2005).
- Schönfeld, P., Reiser, G. Why does brain metabolism not favor burning of fatty acids to provide energy-Reflections on disadvantages of the use of free fatty acids as fuel for brain. Journal of Cerebral Blood Flow and Metabolism. 33 (10), 1493-1499 (2013).
- Calderon-Dominguez, M., Mir, J. F., Fucho, R., Weber, M., Serra, D., Herrero, L. Fatty acid metabolism and the basis of brown adipose tissue function. Adipocyte. 5 (2), 98-118 (2016).
- Divakaruni, A. S., Rogers, G. W., Murphy, A. N. Measuring mitochondrial function in permeabilized cells using the seahorse XF analyzer or a clark-type oxygen electrode. Current Protocols in Toxicology. 2014, 1-16 (2014).
- Iuso, A., Repp, B., Biagosch, C., Terrile, C., Prokisch, H. Assessing mitochondrial bioenergetics in isolated mitochondria from various mouse tissues using Seahorse XF96 analyzer. Methods in Molecular Biology. 1567, 217-230 (2017).
- Rogers, G. W., et al. High throughput microplate respiratory measurements using minimal quantities of isolated mitochondria. PLoS ONE. 6 (7), 21746 (2011).
- Jordá, A., Zaragozá, R., Portolés, M., Báguena-Cervellera, R., Renau-Piqueras, J. Long-term high-protein diet induces biochemical and ultrastructural changes in rat liver mitochondria. Archives of Biochemistry and Biophysics. 265 (2), 241-248 (1988).
- Jackson, C. B., Gallati, S., Schaller, A. QPCR-based mitochondrial DNA quantification: Influence of template DNA fragmentation on accuracy. Biochemical and Biophysical Research Communications. 423 (3), 441-447 (2012).
- Hirsch, H. M. Tissue autoxidation inhibitors: II. The presence of inhibitor in intact cells; Assay of liver and hepatoma effect on radio-oxidations. Cancer Research. 16 (11), 1076-1082 (1956).
- Picard, M., et al. Mitochondrial structure and function are disrupted by standard Isolation methods. PLoS ONE. 6 (3), 18317 (2011).
- Tanumihardja, E., Slaats, R. H., Van Der Meer, A. D., Passier, R., Olthuis, W., Van Den Berg, A. Measuring both pH and O2 with a single On-Chip sensor in cultures of human pluripotent stem cell-derived cardiomyocytes to track induced changes in cellular metabolism. ACS Sensors. 6 (1), 267-274 (2021).
- Harms, F., Stolker, R. J., Mik, E. Cutaneous respirometry as novel technique to monitor mitochondrial function: A feasibility study in healthy volunteers. PLoS ONE. 11 (7), 159544 (2016).
- Levitsky, Y., et al. Micro-respirometry of whole cells and isolated mitochondria. RSC Advances. 9 (57), 33257-33267 (2019).
Yeniden Basımlar ve İzinler
Bu JoVE makalesinin metnini veya resimlerini yeniden kullanma izni talebi
Izin talebiThis article has been published
Video Coming Soon
JoVE Hakkında
Telif Hakkı © 2020 MyJove Corporation. Tüm hakları saklıdır