
このコンテンツを視聴するには、JoVE 購読が必要です。 サインイン又は無料トライアルを申し込む。
Method Article
ヒト単球由来マクロファージにおける炎症性カスパーゼ誘導近接の可視化
要約
このプロトコルは、ヒト血液サンプルから単球由来マクロファージ(MDM)を取得するワークフロー、細胞の生存率と挙動を損なうことなく炎症性カスパーゼ二分子蛍光相補(BiFC)レポーターをヒトMDMに効率的に導入する簡単な方法、および生細胞における炎症性カスパーゼ活性化を測定するためのイメージングベースのアプローチを記述しています。
要約
炎症性カスパーゼは、カスパーゼ−1、−4、−5、−11、および−12を含み、開始カスパーゼのサブグループに属する。カスパーゼ-1は、炎症シグナル伝達の正しい調節を確実にするために必要であり、インフラマソームへの動員に続く近接誘導二量体化によって活性化される。カスパーゼ-1は単球系細胞系譜に豊富であり、炎症誘発性サイトカインインターロイキン(IL)-1βおよびIL-18の成熟を活性分泌分子に誘導する。他の炎症性カスパーゼでは、カスパーゼ−4および−5(ならびにそれらのマウスホモログカスパーゼ−11)は、ピロトーシスを誘導することによってIL−1β放出を促進する。カスパーゼ二分子蛍光相補(BiFC)は、カスパーゼ活性化の読み出しとして炎症性カスパーゼ誘導近接を測定するために使用されるツールです。インフラマソームに結合する領域を含むカスパーゼ-1、-4、または-5プロドメインは、黄色蛍光タンパク質Venus(Venus-N [VN]またはVenus-C [VC])の非蛍光断片に融合され、カスパーゼが誘導近接したときに蛍光金星複合体を改質する。このプロトコルは、ヌクレオフェクションを使用してこれらのレポーターを初代ヒト単球由来マクロファージ(MDM)に導入する方法、炎症性カスパーゼ活性化を誘導するために細胞を治療する方法、および蛍光および共焦点顕微鏡を用いてカスパーゼ活性化を測定する方法を記述している。このアプローチの利点は、生細胞における炎症性カスパーゼ活性化複合体の成分、要件、および局在を同定するために使用することができることである。しかし、細胞の生存率と挙動を損なわないように、慎重な制御を考慮する必要があります。この技術は、インフラマソームレベルでの動的カスパーゼ相互作用の分析、ならびにヒト血液サンプル由来の生きたMDMおよび単球における炎症シグナル伝達カスケードの尋問のための強力なツールである。
概要
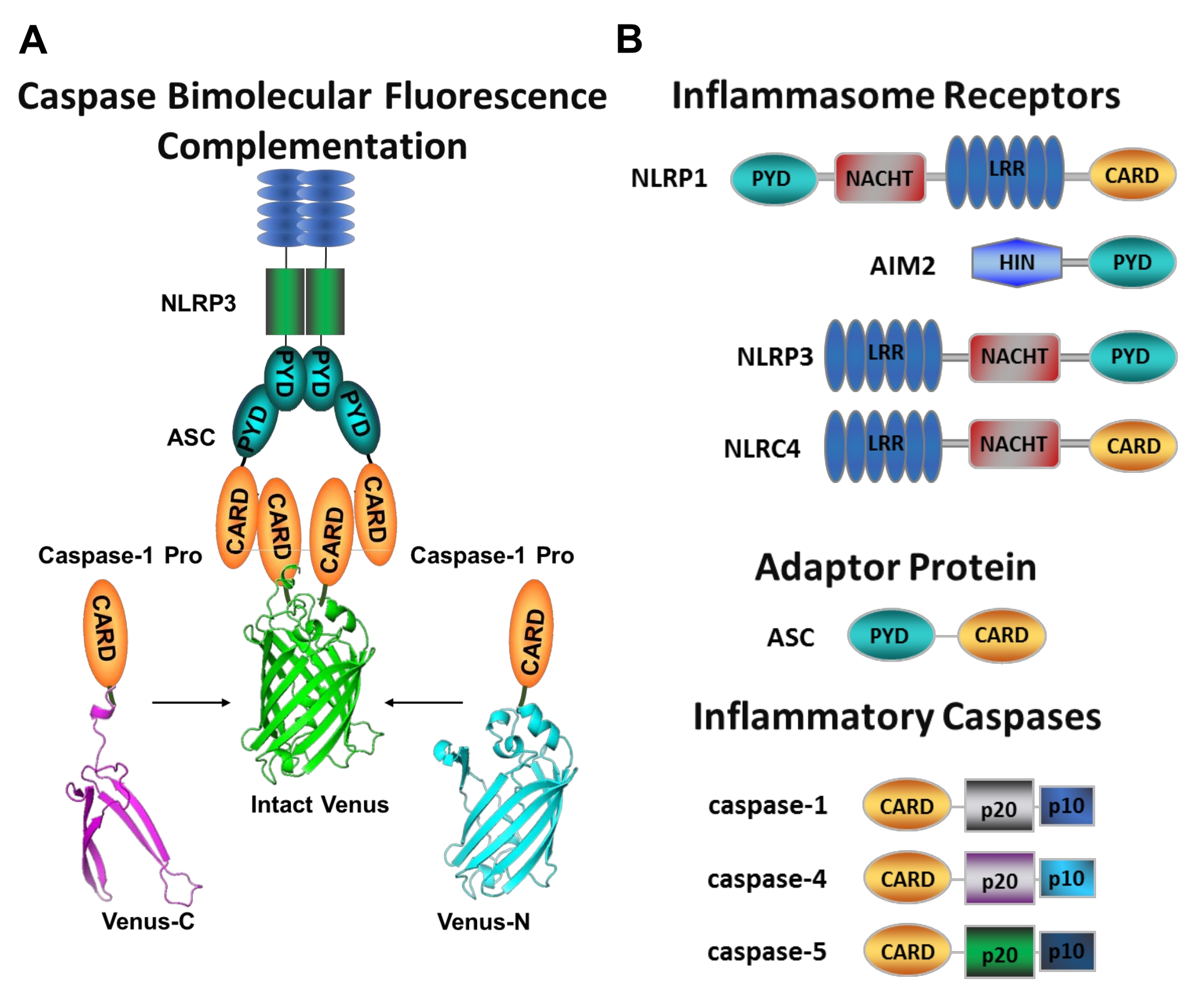
カスパーゼは、システインアスパラギン酸プロテアーゼのファミリーであり、イニシエーターカスパーゼおよび死刑執行人カスパーゼにグループ化することができる。死刑執行人カスパーゼは、カスパーゼ-3、-6および-7を含む。それらは二量体として細胞内に自然に見出され、アポトーシス1を実行するために開始カスパーゼによって切断される。開始カスパーゼには、ヒトカスパーゼ−1、−2、−4、−5、−8、−9、−10および−12が含まれる。それらは、近接誘導二量体化によって活性化され、自己タンパク質分解切断によって安定化される不活性ザイモゲン(プロカスパーゼ)として見出される2,3。炎症性カスパーゼは、開始カスパーゼ2のサブセットであり、ヒトにおけるカスパーゼ−1、−4、−5、および−12、ならびにマウス4、5におけるカスパーゼ−1、−11、および−12を包含する。アポトーシスの役割ではなく、炎症において中心的な役割を果たします。それらは、病原性侵入者8,9に応答して放出される最初のサイトカインであるプロインターロイキン(IL)−1βおよびプロIL−18 6,7のタンパク質分解的プロセシングおよび分泌を媒介する。カスパーゼ-1は、その活性化プラットフォームへの募集時に活性化される。インフラマソームと呼ばれる大きな分子量のタンパク質複合体(図1A)10。カスパーゼ−4、−5、および−11の二量体化は、非正準インフラマソーム経路11、12を介してこれらのプラットフォームとは独立して起こる。
正準インフラマソームは、インフラマソームセンサータンパク質、アダプタータンパク質ASC(CARDを含むアポトーシス関連スペック様タンパク質)、およびエフェクタータンパク質カスパーゼ-110からなる細胞質ゾル多量体タンパク質複合体である。最もよく研究された正準インフラマソームは、ピリンドメイン(NLRP)、NLRP1およびNLRP3を含むNOD様受容体ファミリー、CARD(NLRC)、NLRC4を含むNLRファミリー、および黒色腫2に存在しない(AIM2)である。それらはそれぞれ、ピリンドメイン、CARD、またはその両方のドメインを含む。CARD ドメインは、CARD を含むカスパーゼとそのアップストリーム アクティベーター間の相互作用を仲介します。したがって、N末端ピリンドメイン(PYD)とC末端CARDモチーフ13,14とからなる足場分子ASCは、NLRP110、NLRP3 15、およびAIM216インフラマソームへのカスパーゼ-1のリクルートに必要である。
各インフラマソームは、明確な炎症促進性刺激を認識する独自のセンサータンパク質にちなんで名付けられました(図1B)。この経路の活性化因子は、正準刺激と呼ばれる。インフラマソームは、微生物成分および組織ストレスのセンサとして機能し、炎症性カスパーゼ17の活性化を介して堅牢な炎症反応を誘発するように組み立てられる。インフラマソームアセンブリは、カスパーゼ−1活性化を開始し、その主要基質であるプロ−IL−1βおよびプロIL−18の成熟および分泌を媒介する。このプロセスは、2段階のメカニズムを介して行われます。第1に、プライミング刺激は、NF−κB経路の活性化を介して、ある種のインフラマソームタンパク質およびプロIL−1βの発現をアップレギュレートする。第二に、細胞内(正準)刺激は、プロカスパーゼ-1 6,7のインフラマソーム集合および動員を誘導する。
カスパーゼ−4およびカスパーゼ−5は、マウスカスパーゼ−11 11のヒトオルソログである。それらは、グラム陰性菌18、19、20の外膜に見られる分子である細胞内リポ多糖(LPS)および赤血球溶血21の産物である細胞外ヘムによって、インフラマソーム非依存的に活性化される。LPSがこれらのタンパク質のCARDモチーフに直接結合し、それらのオリゴマー化を誘導することが提案されている20。カスパーゼ-4またはカスパーゼ-5の活性化は、孔形成タンパク質ガスデルミンD(GSDMD)の切断を介してピロトーシスと呼ばれる炎症型の細胞死を誘導することによってIL-1β放出を促進する18,19。さらに、カスパーゼ−4およびGSDMD媒介性パイロプトーシス死から生じるカリウムイオンの流出は、NLRP3インフラマソームの活性化およびその後のカスパーゼ−1の活性化を誘導する22,23。したがって、カスパーゼ−4、−5、および−11は、特定の刺激に応答してパイロプトーシスおよびカスパーゼ−1活性化を誘導することができるLPSの細胞内センサと考えられる11、24。

図1:炎症性カスパーゼおよびカスパーゼ-二分子蛍光相補(BiFC)アッセイ。(A)カスパーゼ-BiFC系を示す図では、金星の各非蛍光断片(Venus-CまたはVenus-N)に連結された2つのカスパーゼ-1プロドメイン(C1-pro)がNLRP3活性化プラットフォームにリクルートされ、金星にリフォールドと蛍光が強制されます。この複合体は、顕微鏡下で緑色の斑点として現れ、開始カスパーゼ活性化の最初のステップである炎症性カスパーゼ誘発近接の読み出しとして機能する。(b)インフラマソーム成分および炎症性カスパーゼのドメイン組織を示す模式図。この図の拡大版を表示するには、ここをクリックしてください。
{kind=link}
特定の開始剤カスパーゼ活性化を測定することは困難であり、イメージングアプローチによってこれを行うために利用可能な方法は多くない。カスパーゼ二分子蛍光相補(BiFC)は、生細胞における炎症性カスパーゼ活性化を直接可視化するために使用できます(図1A)25。この技術は、最近、ヒト単球由来マクロファージ(MDM)21における使用に適応されている。カスパーゼBiFCは、炎症性カスパーゼ活性化における第1ステップを測定し、二量体化を促進するために近接を誘導する。CARD含有カスパーゼプロドメインをコードするプラスミドの発現は、光線上黄色蛍光タンパク質Venusの非蛍光断片に融合(Venus−C[VC])およびVenus−N[VN])が使用される。2つのカスパーゼプロドメインが活性化プラットフォームにリクルートされるか、または誘導された近接を受けると、金星の2つの半分が近接し、リフォールドおよび蛍光を発することを余儀なくされる( 図1A、B参照)。これは、特定の炎症性カスパーゼ活性化のリアルタイム読み出しを提供する。
ヒトMDMは、危険シグナルや病原体産物を同定するインフラマソーム遺伝子やパターン認識受容体を豊富に発現しています。これは、炎症性カスパーゼ経路の尋問に理想的な細胞型を提供する。さらに、それらは末梢血から、さらには患者サンプルから誘導して、特定の疾患状態における炎症性カスパーゼ活性化を評価することができる。このプロトコルでは、ヌクレオフェクションを使用してBiFCカスパーゼレポーターをMDMに導入する方法、エレクトロポレーションベースのトランスフェクション法、炎症性カスパーゼ活性化を誘導するために細胞を治療する方法、および顕微鏡アプローチを使用して活性カスパーゼ複合体を視覚化する方法について説明しています。さらに、この方法論は、これらの複合体の分子組成、細胞内局在化、動態、およびこれらの高度に秩序付けられた構造のサイズを決定するために適合させることができる25、26、27。
プロトコル
このプロトコルは、ベイラー医科大学のヒト研究倫理委員会のヒトサンプルの操作に関するガイドラインに従っています。血液サンプルは、ヒトサンプルの機関安全ガイドラインに従って取り扱われます。血液サンプルは地域の血液バンクで得られ、そこでクエン酸リン酸デキストロース(CPD)溶液で収集されます。しかしながら、ヘパリンナトリウム、リチウムヘパリン、またはEDTAのような他の抗凝固剤を用いて収集された血液も、このプロトコル28,29に使用することができる。
1. ヒト単球の単離とマクロファージへの分化
- 地域の血液バンクで非同定健常者から抗凝固血液を取得し、以下に示すように末梢血単核球(PBMC)を単離する。
メモ: 組織培養層流フードですべての手順を実行します。滅菌チューブのみを使用し、手袋を着用してください。廃棄時にすべての血液関連製品に10%の漂白剤を加えてください。滅菌PBS(1x)またはDPBS(Caなし)2+ および Mg2+) は、同じ意味で使用できます。- 希釈バッファーを調製する:1x滅菌PBSに2%FBSおよび0.5mM EDTAを補充する。
- 培養液を調製する:RPMI-1640培地にFBS(10%(v/v))、グルタマックス(2mM)、ペニシリン/ストレプトマイシン(50I.U./50μg/mL)を補充する
- 製造元のプロトコルに従って、ランニングバッファー(材料表)を予冷します。
- 全血を2倍量の希釈バッファーで希釈する。血清学的ピペットを用いて、15mLの抗凝固血液を30mLの希釈緩衝液を含む50mLチューブに移す。反転で優しく混ぜる。
- 10 mLの全血または30 mLの希釈血液ごとに、15 mLの密度勾配培地を50 mLの空のチューブに加える。
- ステップ1.1.5からの密度勾配培地を、25mLの血清学的ピペットを用いて、希釈血液30mLでゆっくりと着実に層状にする。ピペットの先端をチューブとチューブの壁に傾けた角度に保ちます。
- チューブをスイングバケット遠心分離機に慎重に移します。2 つのフェーズを乱さないでください。チューブを室温(RT)で400 x g で25分間遠心分離し、加減速を最小値に設定します。
- 10mLピペットを使用して上部(透明)プラズマ層を慎重に除去し、漂白剤(10%)の入った容器に廃棄します。
- 末梢血単核球(PBMC、 図2)の相間(白色)層を10mLピペットで集め、新鮮な50mLチューブに移す。同じドナーの異なるチューブからの白色層を30mLまでの50mLチューブに組み合わせる。
- 各チューブをステップ1.1.1の希釈緩衝液で全量50mLにし、300 x g および4°Cで10分間遠心分離する。上清を10mLピペットで取り出し、漂白剤の入った容器に捨てる(10%)。
- p1000マイクロピペットを用いてステップ1.1.3から予め冷却されたランニングバッファーの1mLに各細胞ペレットを再懸濁する。同じドナーからの細胞懸濁液を新しい15mLチューブに混ぜ合わせる。各チューブの容量を予め冷却されたランニングバッファーで15mLにし、反転させてよく混合する。
- ステップ1.1.11から細胞懸濁液の20μLアリコートを取り、1x滅菌PBSを使用して1:100希釈液を調製する。血球計数器を用いて細胞数を決定する。
- ステップ1.1.11からの細胞懸濁液を300 x g および4°Cで10分間遠心分離し、10 mLピペットで上清を除去した。必要に応じて、p200マイクロピペットを使用して上清を完全に除去します。
- 単離したPBMCを、1 x107 細胞ごとに80 μLの予冷MACSランニングバッファーに再懸濁し、最大800 μLのバッファーを追加します。
- 抗ヒトCD14マイクロビーズを各1 x107 細胞あたり20 μL、または血液サンプルあたり最大100 μL(希釈されていない血液約100 mL)まで加える。反転させてよく混合し、チューブ回転子の上に4°Cで20分間連続混合する。
- チューブ回転子からサンプルを取り出し、各チューブに予冷ランニングバッファーを10mL加え、300 x g (加速度=5、減速=5)で4°Cで10分間遠心分離した。
- 上清を10 mLピペットで除去し、500 μLの予冷ランニングバッファー(2 x 108/mL)に最大1 x108細胞を再懸濁します。
- CD14陽性細胞の単離は、製造元の指示に従って手動または自動システム(材料表)を用いて磁気細胞選別を行う。
- CD14陽性選択後のステップ1.1.18から細胞懸濁液の20μLアリコートを採取し、1x滅菌PBSを用いて1:100希釈液を調製する。血球計数器で細胞を数えて細胞数を決定します。
- CD14陽性細胞を300 x g およびRTで10分間遠心分離する。10mLピペットまたは真空システムを用いて上清を除去する。
- ステップ1.1.20の細胞ペレットを、ステップ1.1.2から予め加温した培養培地に再懸濁し、最終細胞密度が1 x107 cells/mLになるまで再懸濁する。
- 単離したCD14陽性単球を5 x106 細胞の細胞密度で播種する。
- 10 cmの組織培養皿に、50 ng/mLの顆粒球 - マクロファージコロニー刺激因子(GM-CSF)を添加したステップ1.1.2の培養液10mLを加える。
- ステップ1.1.21の細胞懸濁液0.5 mLを培養液に滴下し、プレートを静かに旋回させる。細胞を加湿組織培養インキュベーター(37°C、5%CO2)中で一晩インキュベートする。
- 翌日、真空システムを用いて培地を吸引し、一晩付着しなかった細胞を除去した。GM-CSFを添加した新鮮な培養液(50ng/mL)を10mL加え、加湿組織培養インキュベーター(37°C、5%CO2)で細胞を7日間インキュベートして、完全な分化を可能にする(GM-CSFにおける分化の様々な段階におけるCD14+単球の出現については 図3A を参照のこと)。2〜3日ごとに培養液を交換し、毎回新鮮なGM-CSF(50ng/mL)を補充する。

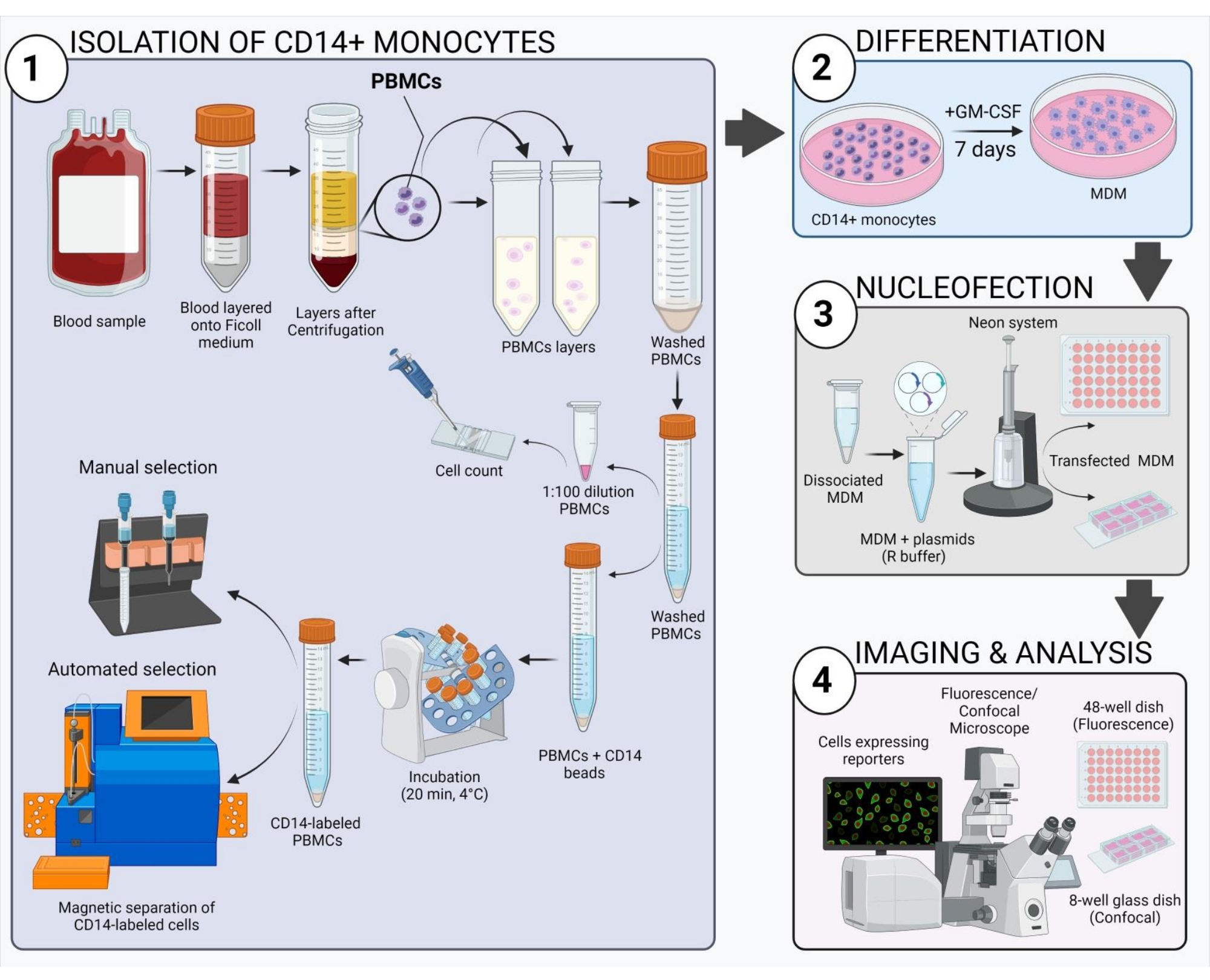
図 2: 実験ワークフローの概略概要。 この図の拡大版を表示するには、ここをクリックしてください。
{kind=link}
エレクトロポレーション成分の調製
注: このプロトコルは、10 μL ネオンチップ用に設計されています (材料表)。各トランスフェクションには、1 ~ 2 x105 個のセルを使用します。トランスフェクトした細胞を 48 ウェルプレートまたは 8 ウェルチャンバーディッシュ (1 ウェルあたり 10 μL トランスフェクトされた細胞) に播種することをお勧めします。1x滅菌DPBS(Ca2+ およびMg2+なし)をPBSの代わりに使用することができる。
- 7日目に、RPMI-1640培地にFBS(10%(v/v))およびグルタマックス(2mM)を補充することにより、抗生物質非含有培地を調製した。
- 無血清RPMI-1640培地、トリプシン-EDTA(0.25%)溶液、1x滅菌PBS(Ca2+ およびMg2+なし)、および37°Cの水浴中にステップ1.1.2からの完全な培養培地を入れた。
- ガラス底ディッシュ(共焦点顕微鏡用)を使用する場合は、ポリD-リジン臭化水素酸塩でディッシュをコーティングする。
- 8 ウェルチャンバーのディッシュに 200 μL のポリ D-リジン臭化水素酸塩 (1x 滅菌 PBS 中 0.1 mg/mL) をコーティングし、RT で 5 分間インキュベートします。
- ポリD-リジン溶液を吸引し、1x滅菌PBSでガラスを1回洗浄する。PBS を吸引し、ステップ 2.4 に進みます。
- 48ウェルプレートまたは8ウェルチャンバーディッシュのウェルあたり200μLの抗生物質非含有培地を添加し、トランスフェクト細胞をプレートする準備ができるまで加湿組織培養インキュベーター(37°C、5%CO2)でプレインキュベートする。
3. エレクトロポレーション用細胞の作製
注:7日間の分化期間の終わりに10cmディッシュからのMDMの収量は、約1.5 x106 細胞である。1x滅菌DPBS(Ca2+ およびMg2+なし)をPBSの代わりに使用することができる。このプロトコルは、ほとんどのマクロファージが細胞生存率および完全性の維持とともにプレートから剥離されるように最適化された。MDMは、細胞培養プレートから剥離することが困難である。したがって、細胞を解離させるためにステップ3.2および3.3を2回実行する必要があるかもしれない。トリプシン-EDTA(0.25%)による各インキュベーション時間が5分を超えないようにしてください。
- 完全に分化したマクロファージから培地を10cmディッシュ上で吸引し、細胞単層を温かい無血清RPMI-1640培地で洗浄する。メディアを完全に取り外してください。
- 10cmディッシュあたり2 mLの温かいトリプシン-EDTA(0.25%)溶液を加えて細胞を回収し、加湿組織培養インキュベーター(37°C、5%CO2)で5分間インキュベートした。
- p1000マイクロピペットを用いてトリプシン-EDTA(0.25%)溶液をディッシュ領域全体に上下に穏やかにピペッティングすることにより、細胞剥離を完了します。細胞懸濁液を、ステップ1.1.2からの5mLの温かい完全培養培地を含む15mL円錐管に移す。
- 皿を明視野顕微鏡に持って行き、様々な視野で細胞の剥離を確認します。まだかなりの量のセルが付着している場合は、手順 3.2 ~ 3.3 を繰り返します。
- 細胞懸濁液を250 x g でRTで5分間遠心分離する。
- 培地を吸引し、37°Cに予め加温した1x滅菌PBSの10mLに細胞を再懸濁する。 20 μLのアリコートを採取し、血球計数器を用いて細胞数を決定した。
- 目的のトランスフェクションあたり 1 ~ 2 x 105 個の細胞を取り、15 mL チューブに入れます。予め加温した1x滅菌PBSで15mLの最終容量に持参する。RTで5分間、250 x g で遠心分離します。
- PBSを吸引し、250 x gで1分間もう一度遠心分離 します。p200マイクロピペットを用いて細胞ペレットから残留PBSを除去する。
4. ヒト単球由来マクロファージへのカスパーゼBiFC成分のヌクレオフェクション
メモ: プロトコルのこのセクションは、Neon トランスフェクションシステム(材料表)を使用して実行されます。このプロトコルは、10 μL のネオンチップを使用して 1 ~ 2 x105 個の細胞をトランスフェクトする手順を概説しています (材料表)。100 μL のネオンチップを使用する場合は、それに応じてスケールアップします。細胞を再懸濁バッファー R に 15 分以上さらすと、細胞の生存率とトランスフェクション効率が低下する可能性があるため、避けてください。
- レポータープラスミド(すなわち、mCherryまたはdsRedmitoを100ng/μL)をヌクレアーゼフリー水または0.5x TEバッファーで希釈して、トランスフェクトされた細胞を視覚化する。
- カスパーゼ BiFC プラスミドをヌクレアーゼフリー水または 0.5x TE バッファーで適切な濃度に希釈し、プラスミドの総容量がトランスフェクションの総容量 10 μL (つまり、300 ng/μL の C1 Pro-VC および 300 ng/μL の C1 Pro-VN) を超えないようにします。
- 目的のトランスフェクションあたり 1.5 mL の滅菌マイクロチューブを調製し、適切な量のレポータープラスミド (すなわち、50 ng または 0.5 μL) およびカスパーゼ BiFC プラスミド (すなわち、300 ng または 1 μL の C1 Pro-VC および 300 ng または 1 μL の C1 Pro-VN フラグメント) を追加します。マイクロチューブは常にフードに入れてください。
- ピペットステーション、装置、チップ、エレクトロポレーションチューブ、およびピペットを滅菌層流フード内に置く。
メモ:ピペットステーション、デバイス、チップ、エレクトロポレーションチューブ、およびピペットは、ネオントランスフェクションシステムに含まれています。 - 製造元の指示に従って、ピペットステーションの高電圧コネクタとセンサーコネクタをデバイスの背面ポートに接続します。ピペットステーションを装置の近くに保管してください。
- 電源コードを背面のACインレットに接続し、デバイスをコンセントに接続します。電源スイッチを押して、デバイスの電源を入れます。
- デバイスの電源を入れて適切に接続したときに表示される起動画面にトランスフェクションパラメータを入力します。電圧を押し、1000と入力し、Doneを押して電圧を1000 Vに設定します。幅を押し、40と入力し、Doneを押してパルス持続時間を40ミリ秒に設定します。 最後に、# パルスを押して「2」と入力し、「完了」を押して電気パルス数を 2 に設定します。
- エレクトロポレーションチューブ(キットに付属)の1つを取り、RTで3 mLの電解バッファーE(チップ10 μL用、キットに付属)を充填します。エレクトロポレーションチューブをピペットステーションのピペットホルダーに挿入します。チューブ側面の電極が内側を向いていること、およびチューブを挿入するときにカチッという音が聞こえることを確認します。
- ステップ3.8から細胞ペレットを取り、1〜2 x105 個の細胞ごとに10μLの予備加温再懸濁R緩衝液(キットに付属)を加える。p20マイクロピペットで優しく混ぜる。ステップ4.3でセットアップした各チューブに10 μLの細胞懸濁液を加え、p20マイクロピペットで穏やかに混合します。
- ピペットを取り、 プッシュ ボタンを押して2番目のストップまでチップを挿入します。クランプがピストンの先端の取り付けステムを完全に持ち上げ、ピペットの上部に隙間が見られないことを確認します。
- サンプルを吸引するには、ピペットの プッシュ ボタンを押して最初のストップまで押し、細胞/プラスミドDNA混合物を含む最初のチューブに浸します。混合物をゆっくりとピペットチップに吸引します。
メモ:気泡はエレクトロポレーション中にアーク放電を引き起こす可能性があり、デバイスによって検出された場合は電気パルスの送達を妨げる可能性があるため、避けてください。気泡が観察された場合は、内容物をチューブに放し、もう一度吸引してみてください。 - サンプルを含むピペットをピペットホルダーに慎重に挿入します。ピペットがカチッと音を立て、正しく配置されていることを確認します。
- タッチスクリーンの [スタート ]を押し、電気パルスが届くまで待ちます。画面上のメッセージは、完了を示します。
- ステーションからピペットをゆっくりと取り外し、プッシュボタンをゆっくりと押して最初のストップまで押して、ステップ2.4から予め加温した抗生物質を含まない培地で対応するウェルにトランスフェクトされた細胞懸濁液を直ちに加えます。
注:このチップは、同じプラスミドに対して最大3回再利用できます。それ以外の場合は、 プッシュ ボタンを2番目のストップに押してバイオハザード廃棄物容器に捨てます。 - 細胞/プラスミドDNA混合物を含む各チューブについて、ステップ4.10~4.14を繰り返します。
- 形質移入細胞でプレートを静かに揺らし、加湿組織培養インキュベーター(37°C、5%CO2)中で1〜3時間インキュベートする。
- ステップ1.1.2から予め加温した培養培地(完全培地)を各ウェルに200μL加える。このディッシュを加湿組織培養インキュベーター(37°C、5%CO2)に再度置く。遺伝子発現に少なくとも24時間かかります。
- 翌日、落射蛍光顕微鏡を用いて細胞生存率およびトランスフェクション効率を検査する。
- 製造元の指示に従って落射蛍光顕微鏡と蛍光光源ボックスの電源を入れ、培養皿を顕微鏡ステージに置きます。
- 10x または 20x の対物レンズと 568 nm (RFP) フィルターを選択します。
- 細胞の生存率を推定するには、 透過光LED(TL) ボタンを押して、選択したフィールド内のすべての細胞を視覚化します。顕微鏡接眼レンズを覗き込みながら、細胞が観察されるまでフォーカスノブを回し、選択したフィールドに細胞が付着していないか確認します。
注: 完全に付着した細胞は生細胞を表し、浮遊細胞は非生存細胞を表します。ウェルのコンフルエンシーが高い場合、非結合細胞の存在は、細胞数の過大評価の結果であり、低い生存率の結果ではない可能性がある。しかしながら、浮遊細胞の高含量を伴う低いコンフルエンシーは、エレクトロポレーション中のアーク放電、プラスミド毒性、または再懸濁R緩衝液への過剰曝露から生じる可能性のある低い生存率を意味する。後者の動作を示す井戸は使用しないでください。 - トランスフェクション効率を推定するには、上記のように透過光下で選択されたフィールドの細胞に焦点を合わせます。選択したフィールドのセルの合計数をカウントします。 透過光 LED (TL) をオフにして、 反射光 LED ボタン (RL) を押してオンにします。
- レポーター遺伝子蛍光(赤血球)に細かく着目し、赤色蛍光細胞の総数をカウントした。これらのステップ (4.18.4-4.18.5) を繰り返して、ウェルごとに少なくとも 2 つのフィールドを増やします。
5. トランスフェクトMDMおよびカスパーゼBiFCデータ取得の治療
注:落射蛍光または共焦点顕微鏡を使用して細胞を画像化する予定の場合、カスパーゼ依存性細胞死(主にアポトーシス)を予防するために、選択した刺激による治療の前にqVD-OPh(20μM)による1時間の治療が推奨される。これは、細胞がアポトーシスによって浮き上がるのを防ぐためにイメージングに使用され、焦点面から移動するときに細胞をイメージングすることを非常に困難にします。活性化プラットフォームおよび関連するカスパーゼBiFCへのカスパーゼ動員は、カスパーゼの触媒活性に依存しないことに留意し、その結果、カスパーゼ阻害はこのステップに影響を及ぼさないであろう。
- トランスフェクションの約24時間後に選択した刺激で治療し、各薬剤に必要な期間インキュベートする。
- 工程1.1.2の培養液にヘペス(20 mM, pH 7.2-7.5)および2-メルカプトエタノール(55 μM)を補充してイメージング培地を調製する。
- 予め加温した画像化媒体に所望の濃度の刺激を加え、穏やかに混合する。
- p1000マイクロピペットで細胞から培地を慎重に取り出し、ステップ5.1.2の刺激溶液500 μLをウェルの側面に加えます。
- 未処理のコントロールウェルを稼働させるには、刺激を与えずにイメージング培地を加える。
- 細胞を加湿組織培養インキュベーター(37°C、5%CO2)中で、各処置について指示される限りインキュベートする。
- 落射蛍光または共焦点顕微鏡を使用して細胞を視覚化します。
- 製造元の指示に従って、顕微鏡と蛍光灯の光源をオンにします。
- 10倍または20倍の対物レンズを選択し、培養皿を顕微鏡ステージに置きます。
- 顕微鏡接眼レンズを使用して、568nmフィルター下の細胞を見つけ、dsRedmito/mCherryレポーターを発現する細胞(赤血球)に焦点を合わせます。
- 視野内のすべての赤いセルを数え、その数を記録します。
- 同じ視野で、488または512フィルタ(GFPまたはYFP)に変更し、緑色(金星陽性またはBiFC陽性)である赤色細胞の数を数え、その数を記録します。
- 少なくとも3つの個々の視野から少なくとも100個のdsRedmito/mCherry陽性細胞を数える。
- 視野あたりの金星陽性形質移入細胞の割合を計算し、得られた割合を各処理(ウェル)で平均して標準偏差を求めます。
- 落射蛍光または共焦点顕微鏡を用いて細胞を画像化する
注:20倍の対物レンズまたはそれ以上の倍率を使用して共焦点画像を取得するには、顕微鏡にロングパス対物レンズが装備されていない限り、細胞をガラス皿にメッキする必要があります。- ステップ 5.2.1 から 5.2.3 に従ってください。40x、60x、または63xのオイル対物レンズで共焦点顕微鏡を使用する場合は、対物レンズにオイルを一滴置きます。
- カメラによって取得されたコンピュータ画面上の細胞のライブ画像を視覚化します。蛍光画像には蛍光発光光源を使用するか、共焦点画像には光源をレーザーに切り替えます。
- ジョイスティックコントロールとフォーカスホイールを使用して、セルのフォーカスと位置を微調整します。
- 512 nm または 488 nm (YFP または GFP) および 568 nm (RFP) レーザーのレーザー出力と露光時間の割合を設定して、画像内の信号が良好に見え、飽和に達しないようにします。
- ライブキャプチャをオンにして、結果の画像を確認します。両方のチャンネルの表示ヒストグラムで、各蛍光に明確なピークが見られることを確認します。
- 必要に応じてレーザー出力と露光時間を調整します。これらの値をできるだけ低く保ちながら、両方の蛍光シグナル(RFPとGFP/YFP)を検出できるようにします。
- 細胞のライブ画像を視覚化しながら、プレートの各ウェルについてmCherry/dsRedmitoレポーターを発現する1つ以上の細胞を含むフィールドの複数の代表画像を撮影し、データを保存します。
結果
選択されたCD14+単球をGM-CSFと共に7日間インキュベートした後、細胞形態は分化期間の過程で変化し(図3A)、球状浮遊細胞からスピン状および完全に付着する(3日目および4日目)まで、 そして最後に、完全に分化したときに細胞をより広げる(7日目)。次いで、完全に分化した細胞をプレートから剥離し、カスパーゼBiFC対(VCおよびVN)をレポータープラス...
ディスカッション
このプロトコルは、ヒト血液サンプルから単離された単球からマクロファージを得るワークフローと、細胞の生存率および挙動を損なうことなく炎症性カスパーゼBiFCレポーターをヒトMDMに効率的に導入する方法を記述している。
このプロトコルは、BiFC技術35 を利用して、カスパーゼリクルートメントドメイン(CARD)における炎症性カスパーゼを、分割蛍...
開示事項
著者らは、競合する金銭的利益はないと宣言している。
謝辞
この技術の開発に貢献したLBHの過去と現在の研究室のメンバーに感謝します。このラボは、NIH/NIDDK T32DK060445 (BEB)、NIH/NIDDK F32DK121479 (BEB)、NIH/NIGMS R01GM121389 (LBH) によってサポートされています。図2は、Biorenderソフトウェアを使用して描画されました。
資料
| Name | Company | Catalog Number | Comments |
| 48 well tissue culture2:34 plates | Genesee Scientific | 25-108 | |
| 10 cm Tissue Culture Dishes | VWR | 25382-166 | |
| 2 Mercaptoethanol 1000x | Thermo Fisher Scientific | 21985023 | |
| 8 well chambered coverglass with 1.5 HP coverglass | Cellvis | c8-1.5H-N | |
| AutoMACS columns | Miltenyi (Biotec) | 130-021-101 | For automated separation using AutoMACS Pro Separator only |
| AutoMACS Pro Separator | Miltenyi (Biotec) | 130-092-545 | For automated separation using AutoMACS Pro Separator only |
| AutoMACS Pro Washing Solution | Miltenyi (Biotec) | 130-092-987 | For automated separation using AutoMACS Pro Separator only |
| AutoMACS Rinsing Solution | Miltenyi (Biotec) | 130-091-222 | For automated separation using AutoMACS Pro Separator only |
| AutoMacs running buffer | Miltenyi (Biotec) | 130-091-221 | For manual or automated separation using QuadroMACS or AutoMACS pro Separator |
| Axio Observer Z1 motorized inverted microscope equipped with a CSU-X1A 5000 spinning disk unit | Zeiss | Any confocal microscope equipped with a laser module fitted with laser lines of 568 nm (RFP) and 488 or 512 nm (GFP or YFP) wavelengths can be used | |
| AxioObserver A1, Research Grade Inverted Microscope | Zeiss | Any epifluorescence microscope with fluorescence filters capable of exciting 568 nm (RFP) and 488 or 512 nm (GFP or YFP) wavelengths can be used | |
| CD14+ MICROBEADS | Miltenyi (Biotec) | 130-050-201 | For manual or automated separation using QuadroMACS or AutoMACS pro Separator |
| DPBS without calcium chloride and magnesium chloride | Sigma | D8537-6x500ML | |
| DsRed mito plasmid | Clontech | 632421 | Similar plasmids that can be used as fluorescent reporters can be found on Addgene |
| Fetal Bovine Serum | Thermo Fisher Scientific | 10437028 | |
| Ficoll-Paque PLUS 6 x 100 mL | Sigma | GE17-1440-02 | |
| GlutaMAX Supplement (100x) | Thermo Fisher Scientific | 35050079 | |
| GM-CSF | Thermo Fisher Scientific | PHC2011 | |
| Hemin BioXtra, from Porcine, ≥96.0% (HPLC) | Sigma | 51280-1G | |
| HEPES | Thermo Fisher Scientific | 15630106 | |
| Inflammatory caspase BiFC plasmids | Available by request from LBH lab | ||
| LPS-EB Ultrapure | Invivogen | TLRL-3PELPS | |
| LS Columns | Miltenyi (Biotec) | 130-042-401 | For manual separation using QuadroMACS Separator only |
| MACS 15 mL Tube Rack | Miltenyi (Biotec) | 130-091-052 | For manual separation using QuadroMACS Separator only |
| MACS MultiStand | Miltenyi (Biotec) | 130-042-303 | For manual separation using QuadroMACS Separator only |
| mCherry plasmid | Yungpeng Wang Lab | Similar plasmids that can be used as fluorescent reporters can be found on Addgene | |
| Neon Transfection System | Thermo Fisher Scientific | MPK5000 | Includes Neon electroporation device, pipette and pipette station |
| Neon Transfection System 10 µL Kit | Thermo Fisher Scientific | MPK1096 | Includes resuspension buffer R, resuspension buffer T, electrolytic buffer E, 96 x 10 µL Neon tips and Neon electroporation tubes |
| Nigericin sodium salt, ready made solution | Sigma | SML1779-1ML | |
| Penicillin-Streptomycin (10,000 U/mL) | Thermo Fisher Scientific | 15140122 | |
| Poly-D-Lysine Hydrobromide | Sigma | P7280-5mg | |
| QuadroMACS Separator | Miltenyi (Biotec) | 130-090-976 | For manual separation using QuadroMACS Separator only |
| qVD-OPh | Fisher (ApexBio) | 50-101-3172 | |
| RPMI 1640 Medium | Thermo Fisher Scientific | 11875119 | |
| Trypsin-EDTA (0.25%), phenol red | Thermo Fisher Scientific | 25200072 | |
| UltraPure 0.5 M EDTA, pH 8.0 | Thermo Fisher Scientific | 15575020 | |
| Zeiss Zen 2.6 (blue edition) software | Zeiss | Any software used to operate the confocal microscope of choice |
参考文献
- Boatright, K. M., et al. A unified model for apical caspase activation. Molecular Cell. 11 (2), 529-541 (2003).
- Pop, C., Salvesen, G. S. Human caspases: activation, specificity, and regulation. Journal of Biological Chemistry. 284 (33), 21777-21781 (2009).
- Boice, A., Bouchier-Hayes, L. Targeting apoptotic caspases in cancer. Biochimica et Biophysica Acta (BBA) - Molecular Cell Research. 1867 (6), 118688 (2020).
- Bolívar, B. E., Vogel, T. P., Bouchier-Hayes, L. Inflammatory caspase regulation: maintaining balance between inflammation and cell death in health and disease. The FEBS Journal. 286 (14), 2628-2644 (2019).
- Martinon, F., Tschopp, J. Inflammatory caspases: linking an intracellular innate immune system to autoinflammatory diseases. Cell. 117 (5), 561-574 (2004).
- Lamkanfi, M., Vishva, M. D. Mechanisms and functions of inflammasomes. Cell. 157 (5), 1013-1022 (2014).
- Viganò, E., et al. Human caspase-4 and caspase-5 regulate the one-step noncanonical inflammasome activation in monocytes. Nature Communications. 6, 8761 (2015).
- Cerretti, D. P., et al. Molecular cloning of the interleukin-1 beta converting enzyme. Science. 256 (5053), 97 (1992).
- van de Veerdonk, F. L., Netea, M. G., Dinarello, C. A., Joosten, L. A. Inflammasome activation and IL-1β and IL-18 processing during infection. Trends in Immunology. 32 (3), 110-116 (2011).
- Martinon, F., Burns, K., Tschopp, J. The inflammasome: a molecular platform triggering activation of inflammatory caspases and processing of proIL-beta. Molecular Cell. 10 (2), 417-426 (2002).
- Kayagaki, N., et al. Noncanonical inflammasome activation targets caspase-11. Nature. 479 (7371), 117-121 (2011).
- Kayagaki, N., et al. Noncanonical inflammasome activation by intracellular LPS independent of TLR4. Science. 341 (6151), 1246-1249 (2013).
- Masumoto, J., et al. ASC, a novel 22-kDa protein, aggregates during apoptosis of human promyelocytic leukemia HL-60 cells. The Journal of Biological Chemistry. 274 (48), 33835-33838 (1999).
- Bertin, J., DiStefano, P. S. The PYRIN domain: a novel motif found in apoptosis and inflammation proteins. Cell Death and Differentiation. 7 (12), 1273-1274 (2000).
- Agostini, L., et al. NALP3 forms an IL-1beta-processing inflammasome with increased activity in Muckle-Wells autoinflammatory disorder. Immunity. 20 (3), 319-325 (2004).
- Bürckstümmer, T., et al. An orthogonal proteomic-genomic screen identifies AIM2 as a cytoplasmic DNA sensor for the inflammasome. Nature Immunology. 10 (3), 266-272 (2009).
- Latz, E., Xiao, T. S., Stutz, A. Activation and regulation of the inflammasomes. Nature Reviews Immunology. 13 (6), 397-411 (2013).
- Kayagaki, N., et al. Caspase-11 cleaves gasdermin D for noncanonical inflammasome signalling. Nature. 526 (7575), 666-671 (2015).
- Shi, J., et al. Cleavage of GSDMD by inflammatory caspases determines pyroptotic cell death. Nature. 526 (7575), 660-665 (2015).
- Shi, J., et al. Inflammatory caspases are innate immune receptors for intracellular LPS. Nature. 514 (7521), 187-192 (2014).
- Bolívar, B. E., et al. Noncanonical Roles of Caspase-4 and Caspase-5 in Heme-Driven IL-1β Release and Cell Death. The Journal of Immunology. 206 (8), 1878-1889 (2021).
- Schmid-Burgk, J. L., et al. Caspase-4 mediates noncanonical activation of the NLRP3 inflammasome in human myeloid cells. European Journal of Immunology. 45 (10), 2911-2917 (2015).
- Baker, P. J., et al. NLRP3 inflammasome activation downstream of cytoplasmic LPS recognition by both caspase-4 and caspase-5. European Journal of Immunology. 45 (10), 2918-2926 (2015).
- Cullen, S. P., Kearney, C. J., Clancy, D. M., Martin, S. J. Diverse activators of the NLRP3 inflammasome promote IL-1β secretion by triggering necrosis. Cell Reports. 11 (10), 1535-1548 (2015).
- Sanders, M. G., et al. Single-cell imaging of inflammatory caspase dimerization reveals differential recruitment to inflammasomes. Cell Death & Disease. 6, 1813 (2015).
- Charendoff, C. I., Bouchier-Hayes, L. Lighting up the pathways to caspase activation using bimolecular fluorescence complementation. Journal of Visualized Experiments: JoVE. (133), e57316 (2018).
- Bouchier-Hayes, L., et al. Characterization of cytoplasmic caspase-2 activation by induced proximity. Molecular Cell. 35 (6), 830-840 (2009).
- Riedhammer, C., Halbritter, D., Weissert, R. Peripheral blood mononuclear cells: Isolation, Freezing, thawing, and culture. Methods in Molecular Biology. 1304, 53-61 (2016).
- Betsou, F., Gaignaux, A., Ammerlaan, W., Norris, P. J., Stone, M. Biospecimen science of blood for peripheral blood mononuclear cell (PBMC) functional applications. Current Pathobiology Reports. 7 (2), 17-27 (2019).
- Perregaux, D., et al. IL-1 beta maturation: evidence that mature cytokine formation can be induced specifically by nigericin. The Journal of Immunology. 149 (4), 1294-1303 (1992).
- Cheneval, D., et al. Increased mature interleukin-1β (IL-1β) secretion from THP-1 cells induced by nigericin is a result of activation of p45 IL-1β-converting enzyme processing. Journal of Biological Chemistry. 273 (28), 17846-17851 (1998).
- Fernandes-Alnemri, T., et al. The pyroptosome: a supramolecular assembly of ASC dimers mediating inflammatory cell death via caspase-1 activation. Cell Death and Differentiation. 14 (9), 1590-1604 (2007).
- Lu, A., et al. Unified polymerization mechanism for the assembly of ASC-dependent inflammasomes. Cell. 156 (6), 1193-1206 (2014).
- Muller-Eberhard, U., Javid, J., Liem, H. H., Hanstein, A., Hanna, M. Brief report: Plasma concentrations of hemopexin, haptoglobin and heme in patients with various hemolytic diseases. Blood. 32 (5), 811-815 (1968).
- Shyu, Y. J., Liu, H., Deng, X., Hu, C. D. Identification of new fluorescent protein fragments for bimolecular fluorescence complementation analysis under physiological conditions. Biotechniques. 40 (1), 61-66 (2006).
- Thornberry, N. A., et al. A novel heterodimeric cysteine protease is required for interleukin-1βprocessing in monocytes. Nature. 356 (6372), 768-774 (1992).
- Jensen, K., Anderson, J. A., Glass, E. J. Comparison of small interfering RNA (siRNA) delivery into bovine monocyte-derived macrophages by transfection and electroporation. Veterinary Immunology and Immunopathology. 158 (3-4), 224-232 (2014).
- Tada, Y., Sakamoto, M., Fujimura, T. Efficient gene introduction into rice by electroporation and analysis of transgenic plants: use of electroporation buffer lacking chloride ions. Theoretical and Applied Genetics. 80 (4), 475-480 (1990).
- Bhowmik, P., et al. Targeted mutagenesis in wheat microspores using CRISPR/Cas9. Scientific Reports. 8 (1), 6502 (2018).
- Sansom, D. M., Manzotti, C. N., Zheng, Y. What's the difference between CD80 and CD86. Trends in Immunology. 24 (6), 314-319 (2003).
- Kurokawa, M., Kornbluth, S. Caspases and kinases in a death grip. Cell. 138 (5), 838-854 (2009).
転載および許可
このJoVE論文のテキスト又は図を再利用するための許可を申請します
許可を申請さらに記事を探す
This article has been published
Video Coming Soon
Copyright © 2023 MyJoVE Corporation. All rights reserved