
Method Article
シアル化表現型の異なる単球由来樹状細胞の作製
要約
シアリダーゼ処理を使用して、単離された末梢血単核細胞(PBMC)から脱シアル化ヒト単球由来樹状細胞(mo-DC)を生成するための独自の包括的なプロトコルが提示されます。さらに、mo-DCの表現型および機能特性を評価し、シアリダーゼ処理がmo-DCの成熟レベルをどのように改善するかを評価する方法が記載されている。
要約
シアル酸は負に帯電した単糖類で、通常、細胞表面の糖鎖の末端に見られます。それらの親水性と生物物理学的特性のために、それらは免疫応答の調節、自己および非自己抗原の認識、炭水化物とタンパク質の相互作用など、多くの生物学的プロセスに関与しています。シアル酸の細胞内含有量はシアリダーゼによって調節され、シアリダーゼはシアル酸残基の除去を触媒します。いくつかの研究により、シアロ糖鎖は、免疫細胞上の シス および トランス 阻害性シグレック受容体に関与することにより、免疫監視のモニタリングに重要であることが示されています。同様に、がんにおける糖鎖免疫チェックポイントは、免疫療法を開発するための重要な標的になりつつあります。さらに、樹状細胞(DC)は、専門抗原提示細胞(APC)としての独自の役割と、適応免疫応答を誘発し、免疫学的記憶を生成する能力により、免疫療法、特にがん研究における重要な要素として想定されています。それにもかかわらず、DCの機能は、その完全な成熟に依存しています。未熟なDCは、成熟DCとは逆の機能を持ち、シアル酸含有量が高いため、成熟レベルがさらに低下します。これにより、未熟なDCがT細胞を活性化する能力がダウンレギュレートされ、免疫応答が損なわれます。その結果、ヒトDCの細胞表面からシアル酸を除去すると、成熟が誘導され、MHC分子の発現と抗原提示が増加します。さらに、共刺激分子とIL-12の発現を回復させることができ、その結果、DCはT細胞をTh1表現型に分極させ、細胞傷害性T細胞を特異的に活性化して腫瘍細胞を殺す能力を高めます。そのため、シアル酸はDCの重要なモジュレーターとして浮上し、その治療用途を前進させるための新しい標的として使用されています。この研究は、 in vitro 単球由来の DC をシアリダーゼで処理するための独自のアプローチを提供し、異なる細胞表面シアル酸表現型とテーラード成熟および共刺激プロファイルを持つ DC 集団を生成することを目的としています。
概要
シアル酸担持糖鎖(シアログリカン)は、免疫調節の役割から大きな関心を集めています。N-アセチルノイラミン酸の形でヒトに最も多く存在する単糖類のシアル酸は、セレクチンやシグレックなど、免疫学において役割が認められているレクチンの基本的なリガンドです。これらのレクチンは、同じ細胞(cis)または異なる細胞(trans)のシアログリカンを認識し、宿主と病原体の相互作用やさまざまな生理学的および病理学的細胞活性に重要な役割を果たします1,2,3。さらに、シアル酸は細胞表面の複合糖質の末端位置を占めるため、その下にある構造を隠蔽し、非特異的な反発作用や他のレクチンによる検出を妨害することで細胞間の接触を阻害することができる4。細胞内のさまざまなシアリルトランスフェラーゼ(シアル酸を転移する)とシアリダーゼ(シアル酸結合を切断する)の活性によって、表面に存在するシアル酸の量が決まります。さらに、宿主または病原体によって発現される可溶性シアリルトランスフェラーゼおよびシアリダーゼは、細胞表面上のシアル酸の量を外因的に変化させることができる5,6。
異常なシアル化は、いくつかの病理学的状態の特徴です。自己免疫疾患では、シアル酸が自己抗原を識別し、炎症反応を調節するのに役立つため、低シアル化は無制限の免疫活性化と臓器損傷に寄与する可能性があります7。逆に、高シアリル化は、シアリル-Tn、シアリル-ルイス抗原、ポリシアル酸、ガングリオシドなどのシアログリカンの過剰発現をもたらし、これはいくつかの癌の特徴を構成します8,9。また、高シアリル化は、N-アセチルグルコサミントランスフェラーゼ(GNT-V)などの特異的酵素の発現増加にも依存し、がんの増殖や転移に関与する高シアル化トリ結合型および/またはテトラ結合型N結合型糖鎖を生成する10。また、シアル酸の含有量はタンパク質の安定性と機能を調節し、関連する発がん性プレーヤーの役割の鍵となります11。したがって、シアル化の増加は、腫瘍の発生、転移、薬剤耐性、および免疫回避を促進する可能性があります。さらに、シアログリカンのアップレギュレーションにより、腫瘍は免疫細胞上の抑制性シグレック受容体と相互作用し、免疫監視を回避することができます。そのため、シアロ糖鎖は現在、糖鎖免疫チェックポイントであり、魅力的な治療標的であると考えられています。例えば、免疫細胞受容体Siglec(シアル酸結合免疫グロブリン様LECtin)が免疫抑制の役割を果たすため、Siglec免疫軸の阻害剤はすでに初期の臨床試験に行われている12。
酵素は、研究や治療戦略のためのツールとして糖鎖プロファイルを調節するために使用されてきました13,14。シアリルルイスXなどのシアル化糖鎖は細胞遊走と癌転移に重要であるため、シアリダーゼは癌細胞の悪性腫瘍を変化させるために用いられてきた15。同時に、シアル酸の切断を妨げるシアリダーゼ阻害剤は、シアル酸依存性ウイルス感染症を治療するための診療所に到達しています16。近年、シアル酸調節は、シグレック免疫軸におけるリガンドとしてのシアル酸の重要な役割により、さらに関心を集めており、免疫応答からのがんの逃避を減らすための新しい手段を提供しています。この関心は、2022年のノーベル賞受賞者であるベルトッツィ氏と彼女のチームが、多様なシアロ糖鎖を選択的に切断し、抗がん免疫応答を改善するいくつかの戦略に貢献したことでさらに強化されました17。したがって、シアリダーゼベースの戦略は、糖鎖免疫チェックポイント療法の有望なモダリティを表しています。免疫系の細胞の糖鎖型は、細胞の種類とその活性化状態に依存します。T細胞に関しては、糖鎖はT細胞の発生と胸腺細胞の選択、T細胞の活性、分化、増殖の病態生理学的ステップにおいて重要な役割を果たします18。例えば、糖タンパク質上のポリラクトサミンは、Bリンパ球とTリンパ球の基礎レベルとマクロファージの活性化に影響を与えます19。マクロファージでは、異なる糖鎖発現パターンが、腫瘍微小環境(TME)へのマクロファージの動員に重要な役割を果たします20。したがって、免疫細胞によるO結合型およびN型糖鎖の発現は、がんや自己免疫疾患の治療アプローチにおける潜在的な糖鎖バイオマーカーとして使用できる可能性があります。
樹状細胞(DC)は、抗がん免疫などの免疫応答を誘発する独自の能力を持つ特異的抗原提示細胞である21。DCは、抗原をT細胞に提示するために抗原提示MHC分子のアップレギュレーションを受け(シグナル1)、T細胞を活性化するための共刺激分子(シグナル2)、およびIL-12などの炎症誘発性サイトカインを上方制御して、1型ヘルパーT細胞の増殖(シグナル3)を誘発する必要があります22。その結果、免疫プロファイルは厳密に制御され、チェックポイントは健康な細胞が攻撃されるのを防ぐために不可欠です。DCは腫瘍細胞に対するさまざまな免疫応答を刺激できるため、細胞ベースのワクチンとして使用され、かなりの数の臨床研究がそれらの潜在的な利点を実証しています23,24。FDAが2010年に最初のDCベースのワクチンを承認した後25,26、他の多くのDCベースのワクチンが開発されています。DCベースのワクチンは、主にex vivoで製造され、腫瘍に対する免疫反応を誘発するために患者に投与されます。しかし、現在、不十分または短期間の成熟は、DCの臨床効果を制限する要因の1つであり、高価なサイトカインカクテルを使用する必要があります。適切な成熟がなければ、DCは臨床状況でT細胞を活性化できません。代わりに、DCは免疫チェックポイントを発現し、細胞傷害性T細胞が腫瘍細胞に対して作用するのを防ぐ寛容性免疫応答を引き起こします。
ヒトDCは表面がシアル化しており、このシアル化は成熟時および全体的な免疫応答中に減少します27。DCの成熟は、これらのシアル酸をシアリダーゼで除去することによって誘導できます。脱シアリル化は、NF-κB転写因子の核への転座により、IL-12を含むさまざまなサイトカインを大幅にアップレギュレートします6,28。さらに、脱シアリル化は、MHC-Iおよび抗腫瘍免疫応答による抗原の交差提示を改善します29,30。したがって、DCシアリル化において主要な役割を有するシアリルトランスフェラーゼST3Gal.lおよびST6Gal.lのノックアウトは、マウスDCにおいてより成熟した表現型を生成する31。
シアリダーゼ治療は、抗原提示の増加、共刺激分子の発現の増加、サイトカイン産生の増加など、DC成熟のあらゆる側面を刺激する方法を提供し、上記の欠点に対処し、DCが効果的な反応を引き出すことを可能にします。この記事では、細菌のシアリダーゼを使用して生存可能な脱シアル化ヒトDCを取得する手順を紹介します。脱シアル化DCは成熟プロファイルの改善を示し、 in vitroで抗腫瘍免疫応答を増強する細胞モデルとして使用できます。DCは血中単球から得られ、サイトカインインターロイキン-4(IL-4)および顆粒球マクロファージコロニー刺激因子(GM-CSF)の存在下で in vitro で分化されます。この研究では、細胞表面のシアル酸を分析するレクチンベースの方法と、DC成熟レベルの免疫表現型分析方法についても説明します。ここで説明する手順は、他の種類の細胞を脱シアリル化するために使用でき、重要な糖免疫チェックポイントであり、免疫調節に関連するシアロ糖鎖の役割を調査するためのアプローチを提供します。
プロトコル
細胞は、国立血液銀行であるInstituto Português do Sangue e da Transplantação(IPST)によって提供されたボランティアである健康な匿名の献血者のバフィーコートから、書面によるインフォームドナーの同意が得られた後に単離されました(IMP.74.52.4)。血液の使用は、ヒト組織および細胞の提供、調達、検査、加工、保存、保管、および配布の品質と安全性の基準に関する指令2004/23/EC(ポルトガル法22/2007、6月29日)に従って、倫理委員会(IPST 30072015)によって承認されました。IPSTバイオバンクは、保存・抗凝固液であるクエン酸リン酸デキストロース(CPD)が入った特定のプラスチック採取バッグに血液を採取・保管し、処理まで血液の完全性を維持します。生体物質が操作に適しているかどうかを評価するために、 梅毒トレポネーマ、B型肝炎ウイルス(HBV)、C型肝炎ウイルス(HCV)、およびヒト免疫不全ウイルス(HIV)の血清学的管理が行われ、これらはすべて陰性でなければなりません。本研究では、バフィーコートは、ドナー32の採取日、血清学的結果、血液型、年齢に関する情報とともに、調査目的でIPSTから提供されました。バフィーコートは、室温で最大1日間保存できます。
1. 単球由来樹状細胞の入手
注:ヒトの末梢血を操作する場合は、特定の普遍的な安全対策と適切な物質廃棄を検討する必要があることに言及することが重要です。開始する前に、必要なすべての試薬と材料が準備され、使用する準備ができていることを確認してください。
- 末梢血単核細胞の単離
- 人間のバフィーコートにアクセスします。
注:バフィーコートは、白血球アフェレーシス32によって収集された血液に由来する副産物であり、遠心分離によって白血球が豊富になります。すべてのステップは、垂直フローチャンバーのバイオセーフティキャビネット(BSC)で行いました。 - 密封されたアウトレットチューブをメスで切断してバフィーコートのパッケージを開き、内容物を50mLのチューブに移します。滅菌済み 15 mL コニカルチューブあたり 7 mL のバフィーコートサンプルを移し、6 mL のリン酸緩衝生理食塩水(PBS)を加えて予備洗浄を行います。この最初の洗浄ステップは、大量の赤血球(RBC)および血漿からサンプルを洗浄し、密度グラジエント培地によるグラジエント分離用にサンプルを最適化するために必要です( 材料表を参照)。
- チューブを室温で1,100 x g で10分間、スイングローター付き遠心分離機でブレーキをオフにして遠心分離します( 材料表を参照)。
- 遠心分離後、白血球懸濁液(血漿と赤血球の間の白いリング)をパスツールピペットで回収し、新しい滅菌済み15 mLコニカルチューブに移します。
- 白血球懸濁液を最大10 mLのPBSで満たして次の分離ステップに役立て、ピペッティングで静かに上下させて混合します。
- 密度グラジエント培地(密度:1.077 g/mL)溶液を調製します:3 mLの密度グラジエント培地を新しい滅菌済み15 mLコニカルチューブに入れ、室温まで温めます。
- 希釈した白血球懸濁液(ステップ1.1.5から)5 mLを密度勾配培地(5:3)を含むコニカルチューブに添加し、密度勾配分離を行います。密度勾配培地を乱さないようにチューブ壁を使用して、サンプルをゆっくりと一滴ずつ加えます。
- グラジエント分離:密度グラジエント培地懸濁液を室温で1,100 x g で30分間、スイングローター付き遠心分離機でブレーキをオフにして遠心分離します。
- 遠心分離後、遠心分離機から円錐形のチューブを慎重に取り外します。このステップの後、下から順に、赤色層(赤血球および顆粒球)、密度勾配培地、末梢血単核細胞(PBMC)の薄い淡い層、および血漿を含む、明確に定義された層の範囲が表示されます。
- パスツールピペットを使用してPBMCの薄層を回収し、密度勾配培地を下に、またはプラズマを上に取りすぎないようにします。PBMC サンプルを新しい 50 mL コニカルチューブに入れ、PBS を 25 mL まで充填し、ピペッティングでゆっくりと上下させてサンプルを混合します。
- 試料を室温で600 x g (ノーマルブレーキ)で10分間遠心分離し、残留細胞や破片を洗い流し、チューブを慎重に反転させて上清を廃棄します。
注:赤血球の汚染が多すぎる場合(細胞ペレットまたはバフィーコートが完全に分離されていないか、赤みがかっているように見える場合に観察される)、残りの赤血球を溶解することをお勧めします。この場合、5 mLのRBC溶解バッファー( 材料表を参照)を加え、十分に混合し、5分間インキュベートします。PBSを最大40 mLまで充填し、サンプルを室温で900 x g (ノーマルブレーキ)で10分間遠心分離し、チューブを慎重に反転させて上清を廃棄します。 - サンプルをPBSで10 mLまで満たし、アリコートを取って細胞をカウントします。血小板を除去するには、室温で400 x g (通常のブレーキ)で5分間遠心分離し、チューブを慎重に反転させて上清を廃棄します。
注:血小板の数が多い場合は、室温で200 x g (通常のブレーキ)で10分間遠心分離機を2回かけます。血小板は、細胞を数えながらサンプルを視覚化することによって識別されます。
- 人間のバフィーコートにアクセスします。
- 免疫磁気分離による単球単離
- PBS に 0.5% ウシ血清アルブミン(BSA)および 2 mM エチレンジアミン四酢酸(EDTA)を添加して、マイクロビーズバッファーを調製します。濾過(0.2μm)により溶液を滅菌し、緩衝液を冷蔵(2〜8°C)します。
- 磁気活性化セルソーティング(MACS)による単球CD14+ 単離を実施します。
- 自動セルカウンターを使用して細胞をカウントした後(ステップ1.4.1)、必要なマイクロビーズバッファーとCD14免疫磁気ビーズ( 材料表を参照)の適切な量を計算します。これらの溶液が氷上に保たれていることを確認してください。1 x 107 細胞あたり80 μLのマイクロビーズバッファーと、1 x 107 細胞あたり20 μLのCD14ビーズを加えます。
- 細胞ペレットを以前に決定した容量で再懸濁し、4°C(2〜8°C)で15分間インキュベートします。
注:PBMCサンプル中の単球レベルの検証が必要な場合は、染色抗体(CD14 [モノクローナルTÜK4]など)を使用してフローサイトメトリー解析を実施してください。ステップ3.2に従って、フローサイトメトリー解析の詳細を確認してください。 - 1 x 107 細胞あたり1〜2 mLのマイクロビーズバッファーを加え、室温で600 x g (ノーマルブレーキ)で10分間遠心分離して結合していないビーズを除去し、チューブを慎重に反転させて上清を廃棄します。
- LSカラムを準備します。LSカラムには強磁性球が含まれており、磁石上に置くと、磁気的に標識された細胞をポジティブで穏やかに保持することができます33。使用直前にLSカラム( 材料表参照)を磁石上に置き、完全に乾燥させないまま3 mLのマイクロビーズバッファーで洗い流し、直ちに次のステップに進んでください。
注:収率の低下を防ぐため、手順中はカラムを乾燥させないでください。 - 細胞ペレットを1 x 108 細胞あたり500 μLのマイクロビーズバッファーに再懸濁します。細胞数が 4 x 108 より大きい場合は、40 μm のセルストレーナーを使用して細胞の凝集を防ぎます。
- 細胞懸濁液をカラムインレットに添加し、15 mL のコニカルチューブをカラムアウトレットの下に置き、陰性細胞分画を回収し、3 mL のマイクロビーズバッファーでカラムを 3 回洗浄します。陰性画分は、CD14ビーズで収集されなかった細胞(すなわち、CD14− 細胞)を含む。
- 最終洗浄後、カラムを磁石から取り外し、滅菌済みの 15 mL コニカルチューブに置き、5 mL のマイクロビーズバッファーをカラムインレットにピペットで移し、すぐにシリンジプランジャーをカラムインレットに挿入し、押して目的細胞を分注します。
- 磁気的に標識された細胞(CD14+ 細胞)を収集し、ステップ1.4.1の説明に従って、アリコートを取って細胞をカウントします。
- CD14− 細胞とCD14+ 細胞の両方の細胞分画を、室温で600 x g で10分間遠心分離します(通常のブレーキ)。上清を廃棄し、次のステップのためにCD14+ 画分を保持し、必要に応じて共培養アッセイなどの将来のアッセイのためにCD14− 画分を保存します。必要に応じて、CD14− 画分由来の細胞をRPMI-1640 20% FBSおよび10% DMSOで−80°Cで凍結保存することができる。
- 樹状細胞への単球分化
- RPMI-1640ベース培地(11.1 mMグルコースを含む)に10%ウシ胎児血清(FBS)、2 mMのL-グルタミン1%、非必須アミノ酸(NEAA)1%、ピルビン酸ナトリウム1%、および100 μg/mLペニシリン/ストレプトマイシン1%を添加して、完全なRPMI-1640培地を調製します( 材料表を参照)。
- 単球をmo-DCに分化させ、5~6日かけて行います。
- 得られたCD14+ 細胞の数に必要な培地の体積を計算し、以下の実験セットアップに従って細胞をプレーティングします。
注:このプロトコルでは、細胞死および測定誤差を考慮するために、細胞を1.3 x 106 細胞/mLの濃度でプレーティングし、1,000 U/mLのGM-CSFおよび750 U/mLのIL-4( 材料表を参照)を完全な培地に添加し、完全に混合することによって培地を調製しました。 - CD14+ 細胞に適切な量の培地を添加し、パスツールピペットでピペッティングして上下に再懸濁します。細胞懸濁液を24ウェルプレート(ウェルあたり:1.3 x 106 細胞/mL)に播種し、37°Cの培養インキュベーターで5% CO2でインキュベートします。
- 培地を交換し、2〜3日ごとに(通常は分化プロセスごとに1回)新鮮なサイトカインを添加します。これを行うには、細胞を乱すことなく培地の半分を慎重に除去します。ステップ1.3.2.1の注で前述したように、適切な濃度のサイトカインを含む同量の新鮮な培地を添加し、残りの分化期間インキュベートします。
注:DCは、単球と分化する場合、緩く接着する細胞です。完全に分化した未熟mo-DCは、紡錘体型で、自由に浮遊し、緩やかに接着する細胞です。細胞はまた、特に成熟するとロゼットを形成することがあります34。 - 分化後の細胞を採取するには、マイクロピペットを使用して細胞懸濁液全体を滅菌コニカルチューブに移し、プレートのウェルをPBSで2回洗浄し、底部を軽くたたきます(図1A)。
注:密着性の高い細胞はおそらくマクロファージであるため、収集しないでください。細胞の不適切な成熟や活性化を避けるために、細胞の取り扱いには細心の注意を払ってください。 - 細胞を室温で180 x g (通常のブレーキ)で10分間遠心分離し、残留物や死細胞を除去し、実験セットアップに適した培地/バッファーに再懸濁します。
- 得られたCD14+ 細胞の数に必要な培地の体積を計算し、以下の実験セットアップに従って細胞をプレーティングします。
- mo-DCの熟成を行います。
- mo-DCの成熟が必要な場合は、以前に使用した細胞濃度の例(1.3 x 10 6 cells/mL)を考慮してウェルプレートまたはフラスコを使用し、IL-1β(10 ng/mL)、IL-6 (1,000 U/mL)、プロスタグランジンE2(PGE2;1 μg/mL)からなるサイトカインカクテルを培地に添加してサイトカインカクテルを投与します。 および腫瘍壊死因子-α(TNF-α; 10 ng/mL)( 材料表を参照)。細胞を37°Cで5%CO2 で24時間または48時間インキュベートします。
- 細胞のカウントと生存率
- 細胞カウントとトリパンブルー染色を行います。
- 細胞数と細胞懸濁液の生存率を測定するには、細胞懸濁液から10 μLのアリコートを採取し、10 μLのトリパンブルー(1:1希釈)と混合します。
- 前のミックスを10 μL取り、自動セルカウンターを使用して、メーカーの指示に従って細胞数をカウントします。
注:細胞の濃度が高すぎる場合は、アリコートを希釈し、細胞計数後に、計算で希釈係数を考慮してください。 - 実験セットアップの細胞数と培地/バッファーを調整します。
- 細胞生存率とアポトーシスを決定する30。
注:この研究では、シアリダーゼ処理(セクション2)に続いて、生存率アッセイを実施しました。- mo-DC を 5 μg/mL の 7-アミノアクチノマイシン D(7-AAD)およびアネキシン V で染色し、メーカーの指示に従ってアポトーシスを決定します( 材料表を参照)。
- フローサイトメトリー29,30を使用して結果を解析します。
- 細胞カウントとトリパンブルー染色を行います。
2. シアリダーゼによる細胞の処理
注:mo-DCへの分化後、6日目に、細胞はシアリダーゼ処理アッセイの準備が整います。
- 目的の実験セットアップを考慮して、1.3 x 10 6 cells/well の 24 ウェルプレートの 10 ウェルから ~10 x 106 mo-DC を回収し、新しい滅菌 15 mL コニカルチューブに移します。
注:ある程度のセル損失を想定します。通常、この段階では、mo-DCとその前駆体は増殖せず、mo-DCへの分化中に20%の生存率の損失を経験するため、検出される濃度は1.3 x 106 細胞/mLです。 - 室温で300 x g (通常のブレーキ)で5〜7分間遠心分離し、上清を捨てて死細胞と破片を除去します。
- 10 mLのRPMI-1640培地(11.1 mMグルコースを含む)を加え、室温で300 x g (通常のブレーキ)で4分間遠心分離し、上清を捨て、2 mLのRPMI-1640を加え、完全に混合します。
- RPMI-1640 の細胞 1 mL を新しい滅菌マイクロチューブ #1 および #2 に入れます。各マイクロチューブには、約 5 x 106 個の細胞が含まれます。
- マイクロチューブ #1 に、 クロストリジウム・パーフリンゲンス由来のシアリダーゼ 500 mU を添加します( 材料表を参照)。マイクロチューブ #2 に、ネガティブコントロールである模擬処理シアリダーゼを添加し、観察された効果がシアル酸の除去に直接関連し、アーチファクトによるものではないかどうかを確認します。模擬処理されたシアリダーゼは、酵素を100°Cで20分間沸騰させることによって得られる熱不活化シアリダーゼです。
- 37°Cで60分間インキュベートします。
- インキュベーション後、細胞を同じ番号(#1および#2)の新しい滅菌15 mLコニカルチューブに入れます。約4 mLの完全なRPMI-1640培地(10%FBSを含む)を両方のチューブに追加して、酵素反応を停止します。
- 室温で300× g (ノーマルブレーキ)で4分間遠心分離し、上澄み液を廃棄する。
- 各チューブに5 mLの完全RPMI-1640培地を加え、ウェルあたり1 mLの細胞をプレーティングします。
3. シアル酸プロファイルの決定
- レクチン染色
- 細胞を回収し、室温で300 x g (通常のブレーキ)で5分間洗浄します。
- 細胞をRPMI-1640 + 10% FBSに再懸濁し、細胞(100,000/100μL)をマイクロチューブに分配します。
- RPMI-1640 の 10% FBS で、各レクチン ( Sambucus nigra (SNA) レクチン、ピーナッツ凝集素 (PNA) レクチン、 Maackia amurensis (MAA) レクチン ( 材料表参照) の濃度 0.01 mg/mL を使用して、フローサイトメトリー用の染色を行います。4°Cで30分間インキュベートします。
- 10% FBSまたは10% BSAを含むPBS1 mLで細胞を洗浄し、室温で300 x g (ノーマルブレーキ)で4分間遠心分離します。
- ビオチン化レクチンで染色した細胞に、0.0005 mg/mLのストレプトアビジン-PE( 材料表参照)を添加し、暗所で室温で15分間インキュベートします。細胞を1 mLのPBSで洗浄し、室温で300 x g (通常のブレーキ)で4分間遠心分離します。
- 上清を廃棄し、各チューブに300 μLの2%パラホルムアルデヒド(PFA 2%)を加えます。チューブを光から保護し、必要に応じて、データが取得されるまで4°Cで保管してください。
- サンプル調製から1週間以内にフローサイトメーターを使用してデータを取得します29,30。
- フローサイトメトリー
- 細胞を1 mLのPBSに再懸濁し、フローサイトメーターでサンプルを取り込み、すぐにデータを取得します。
- データ取得が遅れた場合は、300 μL の 2% PFA に再懸濁し、1 週間以内にデータを取得してください。
- 共焦点レーザー走査型顕微鏡
- 直径12 mmのポリリジンコーティングガラスカバーガラスに細胞を播種し、室温で5分間インキュベートします。
- カバーガラスを室温で100 x g (通常のブレーキ)で1分間遠心分離し、細胞の接着を促進します。
- 室温で4%PFAで30分間固定してから、PBS中の1%BSAで洗浄します。
- FITC標識SNAレクチン(0.01 mg/mL)を使用して、細胞表面のα2,6結合型シアル酸を染色します( 材料表を参照)。
- 共焦点顕微鏡で画像を取得します( 材料表を参照)。
- Zスタック処理後、代表的な共焦点断面画像を選択します。
- 補正全細胞蛍光(CTCF)を用いて染色強度を分析的に定量します。
注:CTCF = 積分密度 − (選択した細胞の面積 × バックグラウンド測定値の平均蛍光)29。
4. mo-DCの成熟プロファイリング
- 抗体染色とフローサイトメトリー
- 目的の細胞の新しいサンプルを採取して、抗体染色を行います。細胞を室温で300 x g (通常のブレーキ)で5分間洗浄し、細胞をマイクロチューブ(チューブあたり100,000細胞)に分配します。
- 目的の抗体(ab)、MHI-I、MHC-II、CD80、およびCD86を使用してフローサイトメトリーの染色を行います( 材料表を参照)。
- 蛍光標識abを暗所で室温で15分間インキュベートします。
- 細胞を1 mLのPBSで洗浄し、室温で300 x g(通常のブレーキ)で5分間遠心分離します。
注:標識のないabを使用する場合は、蛍光標識二次abを添加し、製造元の指示に従って暗所で15分間インキュベートします。細胞を1 mLのPBSで洗浄し、室温で300 x g (通常のブレーキ)で5分間遠心分離します。 - すべてのマイクロチューブに最大100 μLのPBSを添加し、細胞を300 μLの2%パラホルムアルデヒド(PFA 2%)に再懸濁し、データ取得までチューブを4°Cの暗所に保ちます。
- フローサイトメーターを使用してデータを取得します。
注:染色および固定後、サンプルはフローサイトメトリーですぐに、または1週間以内に取得できます。この場合、チューブは暗所で4°Cで保管してください。
5. 酵素結合免疫吸着法(ELISA)
注:この研究では、メーカーの指示に従ってELISAアッセイを使用してIFN-γ産生を測定しました( 材料表を参照)。
- プレートをコーティングバッファーでコーティングする場合は、捕捉抗体を希釈し(1:100、捕捉抗体をPBSで)、このワーキング溶液100 μLを各ウェルに移し、室温で一晩インキュベートします。
- 捕捉抗体を完全に廃棄します。
- ブロッキングバッファー(例:PBS + 2% BSA + 0.05% Tween20)を添加し、室温で1時間インキュベートしてからブロッキングバッファーを除去します。
- 標準試料とサンプルをそれぞれの混合液と希釈液とともに添加し、室温で2時間インキュベートします。洗浄バッファーで5回洗浄します。
- ビオチン化検出器抗体を添加し、室温で2時間インキュベートした後、5回洗浄します。
- poly-HRP-streptavidin-HSを添加し、室温で30分間インキュベートした後、洗浄バッファーで5回洗浄します。
- TMB基質( 材料表を参照)を添加し、使用する試験システムを考慮して室温で最大60分間インキュベートします。洗浄バッファーで5回洗浄します。
- 450 nmのマイクロプレートリーダーでサンプルを読み取ります。
結果
単球単離とmo-DCへの単球分化
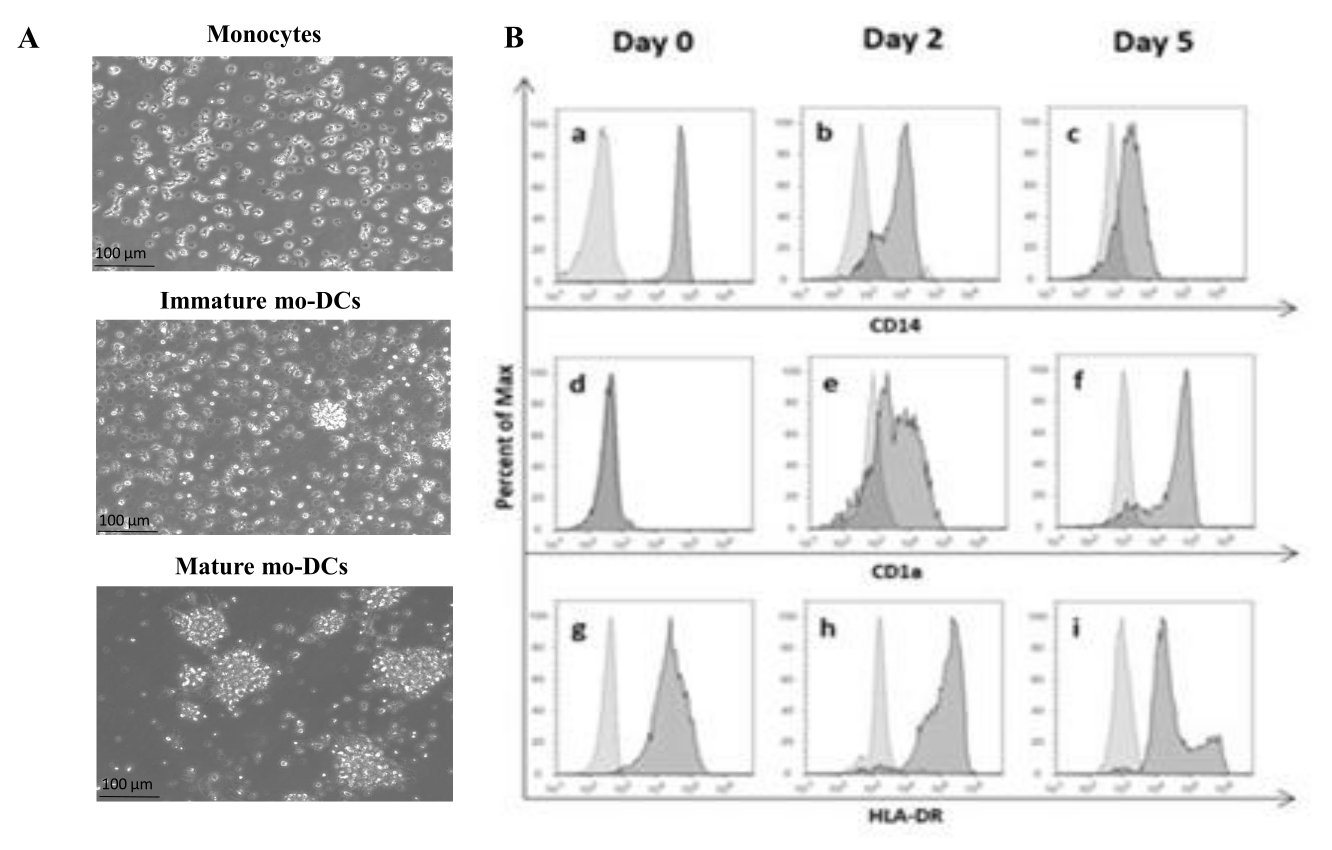
プロトコルに従い、ヒトPBMCを密度勾配培地による密度勾配分離を用いてバフィーコートから単離し、十分に洗浄しました。トリパンブルーを使用して、ステップ1.4.1で前述したように、単離日に生細胞のカウントを行いました。続いて、CD14+ 単球単離をポジティブセレクションにより実施した。これを達成するために、PBMCをCD14抗原を認識する抗体を含む磁気ビーズとインキュベートしました。選択されたCD14+ 単球を、GM-CSFおよびIL-4を添加した培地で5〜6日間培養し27 、未熟なmo-DCに分化させた(図1A)。mo-DCの成熟は、IL-6、IL-1β、TNF-α、およびPGE235 などのサイトカインのカクテルを適用することによって得ることができる(図1A)。
分化過程において、IL-4およびGMCSF刺激の結果として、細胞表現型が変化することが期待されます。データによると、mo-DCは、主に単球によって発現される表面マーカーCD14の発現を失い(図1B)、ヒトDCによって発現されるマーカーであるCD1aの有意な発現を獲得する36,37。また、mo-DCは、ヒトDCおよび他の抗原提示細胞によって発現される抗原提示分子であるMHC-II(HLA-DR)の発現も向上する38(図1B)。
細胞表面のシアル酸含有量
mo-DCをシアリダーゼで処理すると、mo-DCの表面のシアル酸含有量が減少するが、これは炭水化物に結合できるタンパク質であるレクチンで染色することで確認できる39。細胞表面からα2,3とα2,6が結合したシアル酸を除去するため、Mo-DCは、T抗原-Galβ1-3GalNAcα1-Ser/Thrを認識するPNAと、α2,3-とα2,6-シアル酸に結合するMAAとSNAレクチンで染色しました。シアリダーゼ処理の有効性は、フローサイトメトリーおよび共焦点顕微鏡法によって評価されました(図2)。 図2Aに示すように、シアリダーゼ処理はMAAおよびSNA結合を有意に減少させ、PNA染色を増加させました。シアリダーゼ処理後のSNA染色の減少は、細胞表面でのSNA染色が有意に減少したことを示す共焦点顕微鏡分析によってさらに確認されました(図2B)。
シアリダーゼ処理mo-DCの機能評価
シアリダーゼ処理がmo-DC機能にどのように影響するかを評価するために、シアリダーゼ処理後のmo-DCの成熟レベルを評価しました。 図3Aに示すように、シアリダーゼ処理は、抗原提示分子MHC IおよびMHC IIの発現、ならびにCD80およびCD86共刺激分子の発現の有意な増加をもたらす。モダコがT細胞応答を誘導する能力に対するシアル酸除去の効果を評価するために、腫瘍細胞ライセートをロードしたシアリダーゼ処理モダクションを使用して自家T細胞をプライミングしました(図3)。次に、得られたT細胞のプロファイルを、Th1サイトカインIFN-γを分泌する能力に基づいて特徴付けました。 図3Bに示すように、完全にシアル化mo-DCでプライミングされたT細胞と比較すると、シアリダーゼ処理されたmo-DCでプライミングされたT細胞は、有意に高いレベルのIFN-γを分泌しました。これらの結果は、シアリダーゼ処理されたmo-DCが自家T細胞をプライミングする能力を向上させたことを示唆しています。
細胞生存率
シアリダーゼ処理後、処理が細胞に対して細胞傷害性がないことを確認するために生存率アッセイを実施しました。処理後、mo-DCを7-AADおよびアネキシンVで染色し、非生細胞およびアポトーシス細胞を検出し、フローサイトメトリーで解析しました(図4)。データは、未処理細胞(図4、左パネル)とシアリダーゼ処理細胞(図4、右パネル)の間で細胞生存率に有意差がないことを示しています。

図1:単離された単球のmo-DCへの分化。 (A)CD14+単球をバフィーコートから単離し、37°Cで1.3 x 106細胞/mLの濃度で培養しました。 単球は、750 U/mL の IL-4 および 1,000 U/mL の GM-CSF を添加した培地で分化しました。0日目にヒトバフィーコートから単離した単球の形態の顕微鏡解析(トップ画像)。未熟なmo-DC;細胞は、IL-4およびGM-CSFを用いて5日間分化しました(中央画像)。IL-6、IL-1β、TNF-α、およびPGE2サイトカインを24時間使用することにより、成熟したmo-DCが得られました(下図)。スケールバー:100 μm。 (B)細胞は、分化期間を通じて0日目、2日目、および5日目にフローサイトメトリーを使用して分析されました。以下の抗体を細胞表面マーカーの特性評価に用いた:(a-c)CD14;(d-f)CD1a、および(g-i)HLA−DR(MHCクラスII)。この図は、少なくとも3つの独立したアッセイの代表的なヒストグラムを示しています。パネル(B)は、Videira et al.40, patent WO2017002045A1から改変したものである。この図の拡大版をご覧になるには、ここをクリックしてください。
{kind=link}

図2:細胞表面からα2,6およびα2,3結合シアル酸を除去するためのヒトmo-DCのシアリダーゼ処理。 (A)レクチン染色を用いたフローサイトメトリーによるmo-DCの解析によるシアリダーゼ処理の有効性試験。ヒトmo-DCをシアリダーゼ(灰色のバー)で処理するか、未処理(白いバー)で処理し、SNAレクチン([2,6]-シアル酸を認識)、MAAレクチン([2,3]-シアル酸を認識)、およびPNAレクチン(T抗原-Galβ1-3GalNAcα1-Ser/Thrを認識)で染色しました。これらの値は、少なくとも 3 つの独立したアッセイの平均蛍光強度(MFI)を表します。統計的有意性は、未処理のDCとシアリダーゼ処理されたDCの差を参照して、両側対応のあるt検定(*P < 0.05または***P < 0.0001)を使用して決定されました。 シアリダーゼ処理は、α(2,3)結合シアル酸の除去に起因する、ヒトmo-DCにおけるMAA結合を減少させ、PNA染色を増加させました。シアリダーゼ処理後のα(2,6)結合シアル酸の除去は、SNA染色の減少によって検出されました。(B)シアリダーゼで処理し、観察用のカバーガラス上に調製したmo-DCの共焦点顕微鏡。さまざまな細胞からさまざまなzスタック画像を収集し、平均染色強度を含むように処理しました。スケールバー:20 μm。パネル(A)は、Silva et al.30;パネル(B)は、Silva et al.29から改変したものである。この図の拡大版をご覧になるには、ここをクリックしてください。
{kind=link}

図3:成熟マーカーの高発現を誘導するmo-DCのシアリダーゼ処理。 (A)シアリダーゼで処理したmo-DCは、完全シアル化mo-DCよりも高い成熟表現型を示しました。フローサイトメトリーを使用して、いくつかの成熟マーカーの発現を評価しました。mo-DCをシアリダーゼで37°Cで1時間処理しました。グラフ値は、少なくとも 3 つの独立したアッセイの平均蛍光強度 (MFI) (平均 ± SEM) を表しています。統計学的に有意な差は、未処理条件とシアリダーゼ処理条件の差を参照して、t検定(*P < 0.05、**P < 0.01)を使用して計算されました。(B)全腫瘍抗原をロードした脱シアル化ヒトmo-DCは、特異的なT細胞応答を誘導した。mo-DCをシアリダーゼで37°Cで1時間処理するか、未処理のままにした後、全腫瘍細胞抗原の供給源としてMCF-7ライセート(TL)をロードしました。mo-DCと自家T細胞との共培養は、IL-2(10 U/mL)の存在下で4〜8日間実施した。脱シアル化mo-DCでプライミングされたT細胞は、Th1サイトカインであるIFN-γの分泌が有意に高いことを示しました。mo-DCによるT細胞刺激後、共培養上清に分泌されたサイトカインをELISAで測定した(n = 7)。グラフ値は、濃度(pg/mL)(SEM±平均)を表しています。統計的に有意な差は、t検定(*P < 0.05)を用いて計算した。この図は Silva et al.30 から修正されています。この図の拡大版をご覧になるには、ここをクリックしてください。
{kind=link}

図4:シアリダーゼ処理がヒトmo-DCの生存率に及ぼす影響の欠如。 未処理のmo-DC(左パネル)とシアリダーゼ処理したmo-DC(右パネル)をアネキシンVと7-AADで二重染色し、フローサイトメトリーで染色を解析しました。このデータは、未処理細胞とシアリダーゼ処理細胞の間で細胞生存率に有意差を示さなかったことから、mo-DCがシアリダーゼ処理に耐え、免疫学的機能を発揮する生存能力を維持できることを示唆しています。この図は Silva et al.30 から修正されています。 この図の拡大版をご覧になるには、ここをクリックしてください。
{kind=link}
ディスカッション
単球単離
この原稿では、ヒト単離単球CD14+ (図1A)からmo-DCを生成し、その後、これらの細胞の表面のシアル酸含有量を減らすためにシアリダーゼ処理を行うプロトコルについて説明します。
ヒトDCを得るには、末梢血や組織から直接入手する方法や、幹細胞や単球などの前駆体から分化させる方法など、さまざまな方法があります。末梢血から単離された単球から分化したDCを得ることは、他のDC源と比較して大量の単球を得ることが容易であるため、はるかに簡単である41。それでも、単離された単球の割合を高くするには、すべてのプロトコルステップに注意深く従う必要があります。例えば、密度勾配培地は細胞に対して有毒である可能性があり、細胞死を防ぐためには、密度勾配培地との長時間の細胞接触を避け、細胞を完全に洗浄する必要があります。細胞の生存率の損失を避けるために、細胞操作はできるだけ早く行う必要があります。PBMCから単球を単離するには、磁気活性化細胞選別(MACS)法を用いてポジティブセレクションを行い、単球を大量に作製するのに適した技術です。さらに、他の単球選択法と比較して、MACS単離単球由来のmo-DCは、抗腫瘍T細胞活性を刺激するより大きな能力を有する42。このプロトコルでは、単離後、単球をIL-4およびGM-CSFと5〜6日間インキュベートして、未熟なmo-DCに分化させました(図1)。その結果、単離された単球は形態学的(図1A)および表現型(図1B)で未熟なmo-DCに分化することが示されました。さらに、分化の過程で、mo-DCはCD14マーカーの発現を失い、T細胞への抗原提示に必要なCD1aおよびMHC-IIの発現を獲得しました(図1B)。
この単球の単離とmo-DCへの分化は、このプロトコルの制限です。単離プロセスは、細胞死を避けるために慎重かつ迅速に実行する必要がある繊細なステップであり、このステップは、新しい実験のためにmo-DCが必要になるたびに実行する必要があります。分化プロセスには5〜6日かかるため、この分析法をハイスループット分析に採用することは困難です。それにもかかわらず、単離法およびサイトカインを使用してmo-DCを分化することは、実験目的で in vitro で多数の機能的なmo-DCを生成するのに有用です。従ってこのプロトコルで生成されたmo-DCは、シアリダーゼ処理、フローサイトメトリー、ELISA、共焦点顕微鏡検査などを受けることができ、この方法の重要性と有用性を強調しています30。
未熟なmo-DCとシアリダーゼ治療
シアリダーゼはシアル化調節に不可欠であり、細胞表面の糖鎖からシアル酸を除去する役割を担っています。mo-DCでは、シアリダーゼによるシアル酸除去により、これらの細胞が成熟し、抗原交差提示とそれに続くT細胞の活性化と抗腫瘍活性が増加します30。
未熟なヒトmo-DCは、成熟したmo-DCと比較して、細胞表面α(2,6)およびα(2,3)結合シアル酸27の含有量が高い31,43。さらに、mo-DCをシアリダーゼで処理してシアル酸を除去すると、DCの成熟が改善されます28,30,31。この実験のために選択されたシアリダーゼは、細菌Clostridium perfringensからのものでした。しかし、肺炎球菌、コレラ菌、ネズミチフス菌44、ヒルのマクロブデラ・デコラ45、さらにはホモ・サピエンス46など、他の生物もシアリダーゼを産生し、これらの生物由来のシアリダーゼも実験的に使用されています。 しかし、各シアリダーゼは異なる基質特異性を有する。さらに、シアリダーゼ酵素の使用には限界があります。例えば、治療中のmo-DCの操作は、これらの細胞をさらに刺激することができる。さらに、シアリダーゼの量とインキュベーション時間は、使用する細胞の種類とシアル酸組成に基づいて最適化する必要があります。シアル酸の除去は、細胞が細胞表面のシアル酸含有量を回復させるため、永続的な効果ではなく、むしろ一時的な現象です。シアリダーゼ以外にも、シアリルトランスフェラーゼ阻害剤の使用、シアリルトランスフェラーゼ遺伝子の遺伝子ノックアウト、シアル酸模倣物によるシアル酸の代謝遮断など、細胞表面のシアル酸分子を減少させる方法があります47,48,49。それにもかかわらず、これらの方法は細胞に明確な影響を与える可能性があり、脱シアリル化に加えて、細胞の生存率を考慮する必要があります。シアリダーゼ酵素処理は、細胞の生存率を維持しながら、細胞表面のシアル酸を効果的かつ一過性に除去するための実用的な方法です。
この研究では、500 mU/5 x 106 cells/mLの濃度で未熟mo-DCにシアリダーゼを添加し、細胞を37°Cで60分間インキュベートしました。処理は、細胞の生存率を維持し、血清中に存在するシアル化分子間の相互作用を避けるために、血清なしでRPMI-1640を用いて行った30。シアリダーゼ処理は、RPMI以外の緩衝液、例えば50 mM酢酸ナトリウム、pH 5.1(C. perfingens sialidaseの場合)、PBS50,51,52を用いて行うことができます。それにもかかわらず、RPMI-1640は、手順中に一定の実験条件を維持し、成熟の誘発を回避し、シアリダーゼバッファーまたはPBS53,54,55,56によって引き起こされる可能性のあるストレスを軽減するため、DCの最も一般的な培地です。シアリダーゼとインキュベートした後、酵素反応が停止したことを保証するために、血清を添加した培地で細胞を完全に洗浄することが重要です。血清中のシアル化分子の存在は、シアリダーゼの基質として競合するため、迅速な反応停止が保証されます。
フローサイトメトリーおよび共焦点顕微鏡による表面マーカーの特性評価
シアル酸プロファイルの決定には、プロトコールセクション3で、レクチン染色、フローサイトメトリー、共焦点レーザー走査型顕微鏡を使用しました。細胞染色手順では、どちらの場合も、細胞の凝集と死を避けるためにレクチン濃度とインキュベーション条件を最適化しました。レクチンの非特異的結合を避けるために、FBSまたはBSAのいずれかを少なくとも2%含むバッファー中で4°Cでインキュベーションを行うことが重要です。このプロトコルでは、10% FBSを含むRPMI-1640を使用して、一定の実験条件を維持し、細胞ストレスを回避しました。共焦点顕微鏡では、形態の保存、自己消化の防止、抗原性の維持のために、染色前の細胞の固定が不可欠です。
フローサイトメトリーによるmo-DC表現型の解析では、シアリダーゼ処理されたmo-DCは、MMAおよびSNAレクチンと比較して、細胞表面に結合したPNAレクチンの量が有意に多く、シアリダーゼ処理後に減少することが示されました(図2A)。予想通り、PNAは非シアル化抗原を認識するため、MAAとSNAはそれぞれα2,3-およびα2,6-シアル酸に直接結合するため、PNA染色が増加した30。この染色は、このプロトコルを使用して細胞表面からシアル酸を効果的に除去することを確認しています。処理を検証し、細胞表面のシアル酸含有量を分析するために使用できる別の方法は、 図2Bに例示されるように、レクチン染色とそれに続く共焦点顕微鏡検査です。
前者の例に加えて、シアル酸含有量を評価および特性評価するための代替アプローチとして、ウェスタンブロッティングによるレクチンプロービングなどがあります。シアル酸の種類と結合に明確な選好性を持つレクチンのグループであるシグレックなどの代替シアル酸特異的レクチンも利用可能である57。いずれかの技術(フローサイトメトリー、顕微鏡、ウェスタンブロット)でレクチンを使用する以外に、抗体を使用してシアル酸含有量を特徴付けることも可能です。例えば、α2,8-シアル酸は、ポリシアル酸58に特異的なクローン735などの抗体によって評価することができます。さらに、シアリダーゼ処理後、細胞の表現型およびT細胞を活性化する能力を評価することにより、細胞の生物学的または治療的効率を機能的にテストすることができる40。実際、提供された例に示されるように、シアリダーゼ処理されたmo-DCは、より高い成熟表現型を示し、抗原提示分子および共刺激分子の発現が上昇しました。
さらに、シアリダーゼ処理されたmo-DCは、抗原をロードし、T細胞または他の細胞と共培養することができ、表現型、サイトカイン分泌プロファイル、またはその他の特徴に関して研究することができます。提供された例では、シアリダーゼで処理されたmo-DCに腫瘍抗原をロードし、T細胞を活性化するために使用できることがデータから示されています。実際、得られたT細胞はIFN-γ分泌の増加を示し、これはシアル酸不足がT細胞を活性化するmo-DCの能力を高める効果に関する以前の報告と一致しています27,28,29,30,31。
結論として、このプロトコルは、シアリダーゼによる処理によってシアル酸含有量の操作のためのmo-DCを生成するための実行可能で実行可能で実用的な方法を示しています。このプロトコルは、さまざまな目的とアプリケーションに役立つ方法論を提示します。この方法は、免疫細胞の成熟と応答におけるシアル酸の役割を理解する上で重要な役割を果たすだけでなく、免疫調節ツールとしても使用できます。
開示事項
著者は、競合する金銭的利益やその他の利益相反がないことを宣言します。
謝辞
執筆者らは、欧州委員会のGLYCOTwinning GA 101079417およびEJPRD/0001/2020 EU 825575からの資金提供を認めています。FCT 2022.04607.PTDC、UIDP/04378/2020、UIDB/04378/2020 (UCIBIO)、LA/P/0140/2020 (i4HB) の助成金に基づく Fundação para a Ciência e Tecnologia (FCT) Portugal。FCT-NOVAです。また、Stemmattersは、SI I&のPrograma Operacional Regional do Norte(Norte 2020)を通じて、Fundo Europeu de Desenvolvimento Regional (FEDER)から資金提供を受けました。DT DCMatters プロジェクト (NORTE-01-0247-FEDER-047212)。FCT-NOVAとGLYCOVID NOVA SaudeのBiolabs施設に感謝します。
資料
| Name | Company | Catalog Number | Comments |
| 15 mL conical tube | AstiK’s | CTGP-E15-050 | Obtaining Monocyte-derived Dendritic Cells; Treatment of Cells with Sialidase |
| 24-well plate | Greiner Bio-one | 662 160 | Obtaining Monocyte-derived Dendritic Cells; Treatment of Cells with Sialidase |
| 50 mL conical tube | AstiK’s | CTGP-E50-050 | Obtaining Monocyte-derived Dendritic Cells |
| 7-Aminoactinomycin D (7-AAD) | BioLegend | 420404 | Obtaining Monocyte-derived Dendritic Cells |
| Annexin V | Immunotools | 31490013 | Obtaining Monocyte-derived Dendritic Cells |
| Attune Acoustic Focusing Flow Cytometer | Thermo Fisher Scientific | Determination of Sialic Acid Profile; Maturation Profiling of mo-DCs | |
| BSA | Sigma - Aldrich | A3294-100G | Obtaining Monocyte-derived Dendritic Cells; Determination of Sialic Acid Profile |
| CD14 (Monoclonal TÜK4) | Miltenyi Biotec | 130-080-701 | Obtaining Monocyte-derived Dendritic Cells |
| CD80 | Immunotools | 21270803 | Maturation Profiling of mo-DCs |
| CD86 | Immunotools | 21480863 | Maturation Profiling of mo-DCs |
| Cell counting slides and trypan blue | EVE | EVS-050 | Obtaining Monocyte-derived Dendritic Cells |
| Centrifuge | Eppendorf | 5430 R | Obtaining Monocyte-derived Dendritic Cells; Treatment of Cells with Sialidase; Determination of Sialic Acid Profile; Maturation Profiling of mo-DCs |
| Density gradient medium (Histopaque) | Sigma - Aldrich | 10771-100ML | Obtaining Monocyte-derived Dendritic Cells |
| EDTA | Gibco, ThermoFisher | 15400054 | Obtaining Monocyte-derived Dendritic Cells |
| Elisa kit (IFN-γ) | Immunotools | 31673539 | Maturation Profiling of mo-DCs |
| EVE automated cell count | NanoEntek | 10027-452 | Obtaining Monocyte-derived Dendritic Cells |
| Fetal bovine serum (FBS) | Gibco | 10500064 | Obtaining Monocyte-derived Dendritic Cells; Treatment of Cells with Sialidase; Determination of Sialic Acid Profile |
| Granulocyte-macrophage colony-stimulating factor (GM-CSF) | Miltenyi Biotec | 130-093-864 | Obtaining Monocyte-derived Dendritic Cells |
| Human CD14 microbeads (Immunomagnetic beads) | Miltenyi Biotec | 130-050-201 | Obtaining Monocyte-derived Dendritic Cells |
| Interleukin (IL)-1β | Sigma - Aldrich | I9401 | Obtaining Monocyte-derived Dendritic Cells |
| Interleukin (IL)-4 | Miltenyi Biotec | 130-093-919 | Obtaining Monocyte-derived Dendritic Cells |
| Interleukin (IL)-6 | Sigma - Aldrich | SRP3096 | Obtaining Monocyte-derived Dendritic Cells |
| L-glutamine | Gibco | A2916801 | Obtaining Monocyte-derived Dendritic Cells |
| LS column and plunger | Miltenyi Biotec | 130-042-401 | Obtaining Monocyte-derived Dendritic Cells |
| Maackia amurensis (MAA) lectin (MAA lectin - Biotinylated) | Vector labs | B-1265-1 | Determination of Sialic Acid Profile |
| MHC-I (HLA-ABC) | Immunotools | 21159033 | Maturation Profiling of mo-DCs |
| MHC-II (HLA-DR) | Immunostep | HLADRA-100T | Maturation Profiling of mo-DCs |
| Microtubes | AstiK’s | PCRP-E015-500 | Obtaining Monocyte-derived Dendritic Cells; Treatment of Cells with Sialidase; Determination of Sialic Acid Profile; Maturation Profiling of mo-DCs |
| Neuraminidase (Sialidase) | Roche | 11585886001 | Treatment of Cells with Sialidase |
| Non-essential amino acids (NEAA) | Gibco | 11140-050 | Obtaining Monocyte-derived Dendritic Cells |
| Paraformaldehyde (PFA 2%) | Polysciences Europe | 25085-1 | Determination of Sialic Acid Profile; Maturation Profiling of mo-DCs |
| Paraformaldehyde (PFA 4%) | Biotium | 22023 | Determination of Sialic Acid Profile |
| Pasteur pipettes | Labbox | PIPP-003-500 | Obtaining Monocyte-derived Dendritic Cells |
| Peanut (Arachis hypogaea) Agglutinin (PNA) lectin (PNA lectin - FITC) | Vector labs | FL-1071 | Determination of Sialic Acid Profile |
| Penicillin/streptomycin | Gibco | 15140163 | Obtaining Monocyte-derived Dendritic Cells |
| Phosphate Buffered Saline (PBS) | NZYTech | MB18201 | Obtaining Monocyte-derived Dendritic Cells; Treatment of Cells with Sialidase; Determination of Sialic Acid Profile; Maturation Profiling of mo-DCs |
| Prostaglandin E2 (PGE2) | Sigma - Aldrich | P0409 | Obtaining Monocyte-derived Dendritic Cells |
| RBC lysis buffer | BioLegend | 420302 | Obtaining Monocyte-derived Dendritic Cells |
| RPMI-1640 medium (containing 11.1 mM glucose) | Gibco | 31870074 | Obtaining Monocyte-derived Dendritic Cells; Treatment of Cells with Sialidase; Determination of Sialic Acid Profile |
| Sambucus nigra lectin (SNA lectin - Biotinylated) | Vector labs | B-1305-2 | Determination of Sialic Acid Profile |
| Sambucus nigra lectin (SNA lectin - FITC) | Vector labs | FL-1301-2 | Determination of Sialic Acid Profile |
| Sodium pyruvate | Thermofisher | 11360-070 | Obtaining Monocyte-derived Dendritic Cells |
| SpectroMax190 | Molecular Devices | Maturation Profiling of mo-DCs | |
| Streptavidin-PE | BioLegend | 405203 | Determination of Sialic Acid Profile; Maturation Profiling of mo-DCs |
| Tetramethylbenzidine (TMB) | Sigma - Aldrich | T0440 | Maturation Profiling of mo-DCs |
| Tumour necrosis factor-α (TNF-α) | Sigma - Aldrich | H8916 | Obtaining Monocyte-derived Dendritic Cells |
| Zeiss LSM710 confocal microscope | Zeiss | Determination of Sialic Acid Profile |
参考文献
- Varki, A., Gagneux, P. Multifarious roles of sialic acids in immunity. Annals of the New York Academy of Sciences. 1253, 16-36 (2012).
- Bochner, B. S., Zimmermann, N. Role of Siglecs and related glycan-binding proteins in immune responses and immunoregulation. The Journal of Allergy and Clinical Immunology. 135 (3), 598-608 (2015).
- Smith, B. A. H., Bertozzi, C. R. The clinical impact of glycobiology: Targeting selectins, Siglecs and mammalian glycans. Nature Reviews Drug Discovery. 20, 217-243 (2021).
- Schauer, R. Sialic acids as regulators of molecular and cellular interactions. Current Opinion in Structural Biology. 19 (5), 507-514 (2009).
- Manhardt, C. T., Punch, P. R., Dougher, C. W. L., Lau, J. T. Y. Extrinsic sialylation is dynamically regulated by systemic triggers in vivo. Journal of Biological Chemistry. 292 (33), 13514-13520 (2017).
- Cabral, M. G., et al. Human dendritic cells contain cell surface sialyltransferase activity. Immunology Letters. 131 (1), 89-96 (2010).
- Bordron, A., et al. Hyposialylation must be considered to develop future therapies in autoimmune diseases. International Journal of Molecular Sciences. 22 (7), 3402 (2021).
- Julien, S., Videira, P. A., Delannoy, P. Sialyl-Tn in cancer: (How) did we miss the target. Biomolecules. 2 (4), 435-466 (2012).
- Munkley, J. Aberrant sialylation in cancer: Therapeutic opportunities. Cancers. 14 (17), 4248 (2022).
- Dennis, J. W., Laferte, S., Waghorne, C., Breitman, M. L., Kerbel, R. S. S1-6 Branching of Asn-linked oligosaccharides is directly associated with metastasis. Science. 236 (4801), 582-585 (1987).
- Pinho, S. S., Reis, C. A. Glycosylation in cancer: Mechanisms and clinical implications. Nature Reviews Cancer. 15 (9), 540-555 (2015).
- Manni, M., Läubli, H. Targeting glyco-immune checkpoints for cancer therapy. Expert Opinion on Biological Therapy. 21 (8), 1063-1071 (2021).
- Sjögren, J., Lood, R., Nägeli, A. On enzymatic remodeling of IgG glycosylation; Unique tools with broad applications. Glycobiology. 30 (4), 254-267 (2020).
- Trastoy, B., et al. Sculpting therapeutic monoclonal antibody N-glycans using endoglycosidases. Current Opinion in Structural Biology. 72, 248-259 (2022).
- Pascoal, C., et al. Sialyl LewisX/A and cytokeratin crosstalk in triple negative breast cancer. Cancers. 15 (3), 731 (2023).
- von Itzstein, M., et al. Rational design of potent sialidase-based inhibitors of influenza virus replication. Nature. 363 (6428), 418-423 (1993).
- Gray, M. A., et al. Targeted glycan degradation potentiates the anticancer immune response in vivo. Nature Chemical Biology. 16 (12), 1376-1384 (2020).
- Fernandes, &. #. 1. 9. 4. ;., et al. Glycans as shapers of tumour microenvironment: A sweet driver of T-cell-mediated anti-tumour immune response. Immunology. 168 (2), 217-232 (2023).
- Togayachi, A., et al. Polylactosamine on glycoproteins influences basal levels of lymphocyte and macrophage activation. Proceedings of the National Academy of Sciences of the United States of America. 104 (40), 15829-15834 (2007).
- Park, D. D. Resident and elicited murine macrophages differ in expression of their glycomes and glycan-binding proteins. Cell Chemical Biology. 28 (4), 567-582 (2021).
- Steinman, R. M. Dendritic cells and immune-based therapies. Experimental Hematology. 24 (8), 859-862 (1996).
- Sabado, R. L., Bhardwaj, N. Directing dendritic cell immunotherapy towards successful cancer treatment. Immunotherapy. 2 (1), 37-56 (2010).
- Pardoll, D. M. The blockade of immune checkpoints in cancer immunotherapy. Nature Reviews Cancer. 12 (4), 252-264 (2012).
- Steinman, R. M., Banchereau, J. Taking dendritic cells into medicine. Nature. 449 (7161), 419-426 (2007).
- So-Rosillo, R., Small, E. J. Sipuleucel-T (APC8015) for prostate cancer. Expert Review of Anticancer Therapy. 6 (9), 1163-1167 (2006).
- Cheever, M. A., Higano, C. S. PROVENGE (Sipuleucel-T) in prostate cancer: The first FDA-approved therapeutic cancer vaccine. Clinical Cancer Research. 17 (11), 3520-3526 (2011).
- Videira, P. A., et al. Surface α2-3- and α2-6-sialylation of human monocytes and derived dendritic cells and its influence on endocytosis. Glycoconjugate Journal. 25 (3), 259-268 (2008).
- Cabral, M. G., et al. The phagocytic capacity and immunological potency of human dendritic cells is improved by α2,6-sialic acid deficiency. Immunology. 138 (3), 235-245 (2013).
- Silva, Z., et al. MHC class I stability is modulated by cell surface sialylation in human dendritic cells. Pharmaceutics. 12 (3), 249 (2020).
- Silva, M., et al. Sialic acid removal from dendritic cells improves antigen cross-presentation and boosts anti-tumor immune responses. Oncotarget. 7 (27), 41053-41066 (2016).
- Crespo, H. J., et al. Effect of sialic acid loss on dendritic cell maturation. Immunology. 128, 621-631 (2009).
- Council of Europe. Guide to the Preparation, Use and Quality Assurance of Blood Components. Council of Europe. , (2017).
- LS Columns. Miltenyi Biotec Available from: https://www.miltenyibiotec.com/US-en/products/Is-columns.html#130-042-401 (2012)
- Nair, S., Archer, G. E., Tedder, T. F. Isolation and generation of human dendritic cells. Current Protocols in Immunology. 07, (2012).
- Wu, X., Xu, F., Liu, J., Wang, G. Comparative study of dendritic cells matured by using IL-1β, IL-6, TNF-α and prostaglandins E2 for different time span. Experimental and Therapeutic Medicine. 14 (2), 1389-1394 (2017).
- Naeim, F., Nagesh Rao, P., Song, S., Phan, R. Chapter 2 - Principles of Immunophenotyping. Atlas of Hematopathology. , 29-56 (2018).
- Cernadas, M., Lu, J., Watts, G., Brenner, M. B. CD1a expression defines an interleukin-12 producing population of human dendritic cells. Clinical and Experimental Immunology. 155, 523-533 (2009).
- Santambrogio, L., Strominger, J. L. The ins and outs of MHC class II proteins in dendritic cells. Immunity. 25 (6), 857-859 (2006).
- Raposo, C. D., Canelas, A. B., Barros, M. T. Human lectins, their carbohydrate affinities and where to find them. Biomolecules. 11 (2), 188 (2021).
- Videira, P. A. Q., et al. Patent WO2017002045. A viable cell population, method for production and uses thereof. Portugal patent. , (2017).
- Bai, L., Feuerer, M., Beckhove, P., Umansky, V., Schirrmacher, V. Generation of dendritic cells from human bone marrow mononuclear cells: Advantages for clinical application in comparison to peripheral blood monocyte derived cells. International Journal of Oncology. 20 (2), 247-253 (2002).
- Marques, G. S., Silva, Z., Videira, P. A. Antitumor efficacy of human monocyte-derived dendritic cells: Comparing effects of two monocyte isolation methods. Biological Procedures Online. 20, 4 (2018).
- Bax, M., et al. Dendritic cell maturation results in pronounced changes in glycan expression affecting recognition by Siglecs and galectins. Journal of Immunology. 179 (12), 8216-8224 (2007).
- Chinoy, Z. S., Montembault, E., Moremen, K. W., Royou, A., Friscourt, F. Impacting bacterial sialidase activity by incorporating bioorthogonal chemical reporters onto mammalian cell-surface sialosides. ACS Chemical Biology. 16 (11), 2307-2314 (2021).
- Chou, M. -. Y., Li, S. -. C., Li, Y. -. T. Cloning and expression of sialidase L, a NeuAcα2→3Gal-specific sialidase from the leech, Macrobdella decora. Journal of Biological Chemistry. 271 (32), 19219-19224 (1996).
- Crespo, H. J., Lau, J. T. Y., Videira, P. A. Dendritic cells: A spot on sialic acid. Frontiers in Immunology. 4, 491 (2013).
- Büll, C. Metabolic sialic acid blockade lowers the activation threshold of moDCs for TLR stimulation. Immunology & Cell Biology. 95 (4), 408-415 (2017).
- Ohmi, Y., et al. Majority of alpha2,6-sialylated glycans in the adult mouse brain exist in O -glycans: SALSA-MS analysis for knockout mice of alpha2,6-sialyltransferase genes. Glycobiology. 31 (5), 557-570 (2021).
- Chung, C., et al. Integrated genome and protein editing swaps α-2,6 sialylation for α-2,3 sialic acid on recombinant antibodies from CHO. Biotechnology Journal. 12 (2), 1600502 (2017).
- Hyvärinen, S., Meri, S., Jokiranta, T. S. Disturbed sialic acid recognition on endothelial cells and platelets in complement attack causes atypical hemolytic uremic syndrome. Blood. 127 (22), 2701-2710 (2016).
- Powell, L. D., Whiteheart, S. W., Hart, G. W. Cell surface sialic acid influences tumor cell recognition in the mixed lymphocyte reaction. Journal of Immunology. 139, 262-270 (1987).
- Corfield, A. P., Higa, H., Paulson, J. C., Schauer, R. The specificity of viral and bacterial sialidases for α(2-3)- and α(2-6)-linked sialic acids in glycoproteins. Biochimica et Biophysica Acta - Protein Structure and Molecular Enzymology. 744 (2), 121-126 (1983).
- Tkachenko, N., Wojas, K., Tabarkiewicz, J., Rolinski, J. Generation of dendritic cells from human peripheral blood monocytes - Comparison of different culture media. Folia Histochemica et Cytobiologica. 43, 25-30 (2005).
- Kim, S. J., et al. Human CD141+ dendritic cells generated from adult peripheral blood monocytes. Cytotherapy. 21 (10), 1049-1063 (2019).
- Calmeiro, J., et al. In-depth analysis of the impact of different serum-free media on the production of clinical grade dendritic cells for cancer immunotherapy. Frontiers in Immunology. 11, 593363 (2021).
- Stamatos, N. M., et al. LPS-induced cytokine production in human dendritic cells is regulated by sialidase activity. Journal of Leukocyte Biology. 88 (6), 1227-1239 (2010).
- Lehmann, F., Tiralongo, E., Tiralongo, J. Sialic acid-specific lectins: Occurrence, specificity and function. Cellular and Molecular Life Sciences. 63 (12), 1331-1354 (2006).
- Frosch, M., Görgen, I., Boulnois, G. J., Timmis, K. N., Bitter-Suermann, D. NZB mouse system for production of monoclonal antibodies to weak bacterial antigens: Isolation of an IgG antibody to the polysaccharide capsules of Escherichia coli K1 and group B meningococci. Proceedings of the National Academy of Sciences of the United States of America. 82 (4), 1194-1198 (1985).
転載および許可
このJoVE論文のテキスト又は図を再利用するための許可を申請します
許可を申請さらに記事を探す
This article has been published
Video Coming Soon
Copyright © 2023 MyJoVE Corporation. All rights reserved