
Method Article
Generierung von Monozyten-abgeleiteten dendritischen Zellen mit unterschiedlichen sialylierten Phänotypen
In diesem Artikel
Zusammenfassung
Ein einzigartiges, umfassendes Protokoll zur Generierung von de-sialylierten humanen monozyten-abgeleiteten dendritischen Zellen (mo-DCs) aus isolierten mononukleären Zellen des peripheren Blutes (PBMCs) unter Verwendung einer Sialidase-Behandlung wird vorgestellt. Des Weiteren werden Methoden zur Beurteilung der phänotypischen und funktionellen Charakterisierung von mo-DCs beschrieben und evaluiert, wie die Behandlung mit Sialidase das Reifungsniveau von mo-DCs verbessert.
Zusammenfassung
Sialinsäuren sind negativ geladene Monosaccharide, die typischerweise an den Enden von Glykanen auf der Zelloberfläche zu finden sind. Aufgrund ihrer Hydrophilie und biophysikalischen Eigenschaften sind sie an zahlreichen biologischen Prozessen beteiligt, wie z.B. der Modulation der Immunantwort, der Erkennung von Selbst- und Nicht-Selbstantigenen, Kohlenhydrat-Protein-Wechselwirkungen usw. Der zelluläre Gehalt an Sialinsäure wird durch Sialidase reguliert, die die Entfernung von Sialinsäureresten katalysiert. Mehrere Studien haben gezeigt, dass Sialo-Glykane entscheidend für die Überwachung der Immunüberwachung sind, indem sie mit cis- und trans-inhibitorischen Siglec-Rezeptoren auf Immunzellen interagieren. In ähnlicher Weise werden Glyko-Immun-Checkpoints bei Krebs zu entscheidenden Zielen für die Entwicklung von Immuntherapien. Darüber hinaus werden dendritische Zellen (DCs) aufgrund ihrer einzigartigen Rolle als professionelle Antigen-präsentierende Zellen (APC) und ihrer Fähigkeit, adaptive Immunantworten auszulösen und immunologisches Gedächtnis zu erzeugen, als wichtige Komponente in Immuntherapien, insbesondere in der Krebsforschung, angesehen. Nichtsdestotrotz hängt die Funktion von DCs von ihrer vollen Reifung ab. Unreife DCs haben eine gegenläufige Funktion zu reifen DCs und einen hohen Sialinsäuregehalt, was ihren Reifegrad weiter dämpft. Dadurch wird die Fähigkeit unreifer DCs, T-Zellen zu aktivieren, herunterreguliert, was zu einer beeinträchtigten Immunantwort führt. Folglich induziert die Entfernung von Sialinsäure von der Zelloberfläche menschlicher DCs deren Reifung und erhöht so die Expression von MHC-Molekülen und die Antigenpräsentation. Darüber hinaus kann es die Expression von co-stimulatorischen Molekülen und IL-12 wiederherstellen, was dazu führt, dass DCs eine höhere Fähigkeit haben, T-Zellen in Richtung eines Th1-Phänotyps zu polarisieren und zytotoxische T-Zellen spezifisch zu aktivieren, um Tumorzellen abzutöten. Daher hat sich Sialinsäure als Schlüsselmodulator von DCs herauskristallisiert und wird als neuartiges Ziel verwendet, um ihre therapeutische Verwendung voranzutreiben. Diese Studie bietet einen einzigartigen Ansatz zur Behandlung von in vitro Monozyten-abgeleiteten DCs mit Sialidase, der darauf abzielt, DC-Populationen mit unterschiedlichen Sialinsäure-Phänotypen auf der Zelloberfläche und maßgeschneiderten Reifungs- und Stimulationsprofilen zu generieren.
Einleitung
Sialinsäure-tragende Glykane (Sialoglykane) haben aufgrund ihrer immunmodulatorischen Rolle großes Interesse geweckt. Das Monosaccharid Sialinsäure, das beim Menschen am häufigsten in Form von N-Acetylneuraminsäure vorkommt, stellt grundlegende Liganden für Lektine dar, die eine anerkannte Rolle in der Immunologie spielen, wie z. B. Selektine und Siglecs. Diese Lektine erkennen Sialoglykane entweder auf derselben Zelle (cis) oder auf verschiedenen Zellen (trans) und spielen eine wichtige Rolle bei Wirt-Pathogen-Interaktionen und verschiedenen physiologischen und pathologischen zellulären Aktivitäten 1,2,3. Da Sialinsäure die terminalen Positionen von Glykokonjugaten auf der Zelloberfläche einnimmt, kann sie außerdem die darunter liegenden Strukturen verbergen und so den Zell-zu-Zell-Kontakt durch unspezifische abstoßende Effekte oder durch Behinderung der Detektion durch andere Lektine hemmen4. Die Aktivität einer Vielzahl von Sialyltransferasen (die Sialinsäuren übertragen) und von Sialidasen (die Sialinsäurebindungen spalten) innerhalb der Zelle bestimmt die Menge an Sialinsäure, die an der Oberfläche vorhanden ist. Darüber hinaus können lösliche Sialyltransferasen und Sialidasen, die vom Wirt oder von Krankheitserregern exprimiert werden, die Menge an Sialinsäure auf der Zelloberfläche extrinsisch modifizieren 5,6.
Die aberrante Sialylierung ist ein Merkmal verschiedener pathologischer Zustände. Bei Autoimmunerkrankungen kann die Hyposialylierung zu einer ungebremsten Immunaktivierung und Organschädigung beitragen, da Sialinsäure hilft, Selbstantigene zu unterscheiden und Entzündungsreaktionen zu regulieren7. Umgekehrt führt die Hypersialylierung zu einer Überexpression von Sialoglykanen wie Sialyl-Tn, Sialyl-Lewis-Antigenen, Polysialinsäure und Gangliosiden, die ein Kennzeichen einiger Krebsarten darstellen 8,9. Die Hypersialylierung hängt auch von einer erhöhten Expression spezifischer Enzyme wie der N-Acetylglucosaminyltransferase (GNT-V) ab, die hypersialylierte Tri- und/oder Tetra-Antennen-N-verknüpfte Glykane erzeugt, die mit Krebswachstum und Metastasierung in Verbindung gebracht werden10. Der Sialinsäuregehalt reguliert auch die Stabilität und Funktion von Proteinen, die für die Rolle relevanter onkogener Akteure von entscheidender Bedeutung sind11. Daher kann eine erhöhte Sialylierung die Tumorentwicklung, Metastasierung, Arzneimittelresistenz und Immunevasion begünstigen. Darüber hinaus ermöglicht die Hochregulierung von Sialoglykanen Tumoren, mit hemmenden Siglec-Rezeptoren auf Immunzellen zu interagieren und eine Immunüberwachung zu vermeiden. Aus diesem Grund gelten Sialoglykane heute als Glyko-Immun-Checkpoints und attraktive therapeutische Ziele. So befinden sich Inhibitoren der Siglec-Immunachse bereits in frühen klinischen Studien, da der Immunzellrezeptor Siglec (Sialsäure-bindendes Immunglobulin-like LECtin) eine immunhemmende Rolle spielt12.
Enzyme wurden zur Modulation des Glykanprofils als Werkzeuge für Studien oder therapeutische Strategien verwendet13,14. Sialidase wurde eingesetzt, um die Malignität von Krebszellen zu verändern, da sialylierte Glykane wie Sialyl Lewis X für die Zellmigration und die Krebsmetastasierung entscheidend sind15. Gleichzeitig haben Sialidase-Inhibitoren, die die Sialinsäurespaltung hemmen, Kliniken zur Behandlung von Sialinsäure-abhängigen Virusinfektionen erreicht16. In jüngster Zeit hat die Sialinsäure-Modulation aufgrund der entscheidenden Rolle von Sialinsäuren als Liganden in der Siglec-Immunachse weiter an Interesse gewonnen, was bedeutet, dass sie neuartige Mittel zur Verringerung der Krebsflucht aus Immunantworten bieten. Dieses Interesse wurde durch den Beitrag der Nobelpreisträgerin von 2022 Bertozzi und ihres Teams zu mehreren Strategien weiter gestärkt, die selektiv verschiedene Sialoglykane spalten und die Immunantwort gegen Krebs verbessern17. Daher stellen Sialidase-basierte Strategien eine vielversprechende Modalität für die Glyko-Immun-Checkpoint-Therapie dar. Der Glykophenotyp von Zellen des Immunsystems ist abhängig von der Art der Zelle und ihrem Aktivierungsstatus. In Bezug auf T-Zellen spielen Glykane eine Schlüsselrolle bei den pathophysiologischen Schritten der T-Zell-Entwicklung und der Thymozytenselektion, der T-Zell-Aktivität, -Differenzierung und -Proliferation18. Zum Beispiel beeinflusst Polylactosamin auf Glykoproteinen die basalen Spiegel von B-Lymphozyten und T-Lymphozyten und die Makrophagenaktivierung19. In Makrophagen spielen unterschiedliche Glykanexpressionsmuster eine wichtige Rolle bei der Rekrutierung von Makrophagen in die Tumormikroumgebung (TME)20. Daher könnte die Expression von O- und N-chromosomalen Glykanen durch Immunzellen als potenzielle Glykobiomarker für therapeutische Ansätze in der Behandlung von Krebs und Autoimmunerkrankungen genutzt werden.
Dendritische Zellen (DCs) sind spezifische Antigen-präsentierende Zellen mit einer einzigartigen Fähigkeit, Immunantworten auszulösen, wie z. B. die Anti-Krebs-Immunität21. DCs müssen eine Hochregulierung ihrer Antigen-präsentierenden MHC-Moleküle durchlaufen, um Antigene für T-Zellen zu präsentieren (Signal 1), kostimulatorische Moleküle zur Aktivierung von T-Zellen (Signal 2) und pro-inflammatorische Zytokine wie IL-12, um die Proliferation von Typ-1-Helfer-T-Zellen auszulösen (Signal 3)22. Das daraus resultierende Immunprofil ist streng reguliert, und Checkpoints sind unerlässlich, um zu verhindern, dass gesunde Zellen angegriffen werden. Da DCs verschiedene Immunantworten gegen Tumorzellen stimulieren können, werden sie als zellbasierte Impfstoffe eingesetzt, und eine beträchtliche Anzahl klinischer Studien hat ihren potenziellen Nutzen nachgewiesen23,24. Nachdem die FDA im Jahr 2010 den ersten DC-basierten Impfstoff zugelassenhat 25,26, wurden viele weitere DC-basierte Impfstoffe entwickelt. DC-basierte Impfstoffe werden hauptsächlich ex vivo hergestellt und Patienten verabreicht, um Immunantworten gegen Tumore hervorzurufen. Eine unzureichende oder kurze Reifung ist jedoch derzeit einer der Faktoren, die die klinische Wirksamkeit von DCs einschränken und dazu führen, dass teure Zytokincocktails verwendet werden müssen. Ohne ausreichende Reifung können DCs T-Zellen unter klinischen Bedingungen nicht aktivieren. Stattdessen exprimieren die DCs Immun-Checkpoints und lösen eine tolerogene Immunantwort aus, die zytotoxische T-Zellen daran hindert, gegen Tumorzellen zu wirken.
Menschliche DCs haben stark sialylierte Oberflächen, und diese Sialylierung nimmt mit der Reifung und während einer allgemeinen Immunantwort ab27. Die Reifung von DCs kann durch Eliminierung dieser Sialinsäuren mit Sialidase induziert werden. Die Desialylierung reguliert verschiedene Zytokine, einschließlich IL-12, aufgrund der Translokation des NF-kB-Transkriptionsfaktors in den Zellkernstark hoch 6,28. Darüber hinaus verbessert die Desialylierung die Antigen-Kreuzpräsentation durch MHC-I und die Anti-Tumor-Immunantwort29,30. Dementsprechend erzeugt der Knockout der Sialyltransferasen ST3Gal.l und ST6Gal.l, die eine wichtige Rolle bei der DC-Sialylierung spielen, einen reiferen Phänotyp in murinen DCs31.
Die Sialidase-Behandlung bietet eine Methode zur Stimulierung aller Aspekte der DC-Reifung, einschließlich einer erhöhten Antigenpräsentation, einer erhöhten Expression von kostimulierenden Molekülen und einer erhöhten Zytokinproduktion, um die oben genannten Mängel zu beheben und DCs in die Lage zu versetzen, wirksame Reaktionen hervorzurufen. In diesem Artikel wird ein Verfahren vorgestellt, um lebensfähige desialylierte humane DCs durch die Verwendung einer bakteriellen Sialidase zu erhalten. Desialylierte DCs zeigen ein verbessertes Reifungsprofil und können als Zellmodelle verwendet werden, um die Anti-Tumor-Immunantwort in vitro zu verstärken. Die DCs werden aus Blutmonozyten gewonnen, die dann in vitro in Gegenwart des Zytokins Interleukin-4 (IL-4) und des Granulozyten-Makrophagen-Kolonie-stimulierenden Faktors (GM-CSF) differenziert werden. In dieser Arbeit werden auch lektinbasierte Methoden zur Analyse von Sialinsäure an der Zelloberfläche und Methoden zur Immunphänotypisierung des DC-Reifungsgrades beschrieben. Das hier beschriebene Verfahren kann zur Desialylierung anderer Zelltypen verwendet werden und bietet somit einen Ansatz, um die Rolle von Sialoglykanen zu untersuchen, die wichtige Glyko-Immun-Checkpoints sind und für die Immunmodulation relevant sind.
Protokoll
Die Zellen wurden aus den Buffy-Mänteln gesunder anonymer Blutspender isoliert, bei denen es sich um Freiwillige handelte, die von der nationalen Blutbank Instituto Português do Sangue e da Transplantação (IPST) zur Verfügung gestellt wurden, nachdem die schriftliche und informierte Zustimmung der Spender eingeholt worden war (IMP.74.52.4). Die Verwendung von Blut wurde von der Ethikkommission (IPST 30072015) gemäß der Richtlinie 2004/23/EG über Qualitäts- und Sicherheitsstandards für die Spende, Beschaffung, Testung, Verarbeitung, Konservierung, Lagerung und Verteilung von menschlichen Geweben und Zellen genehmigt (portugiesisches Gesetz 22/2007 vom 29. Juni). Die IPST-Biobank sammelt und lagert Blut in einem speziellen Plastiksammelbeutel, der Citratphosphatdextrose (CPD), eine konservierende und gerinnungshemmende Lösung, enthält, um die Integrität des Blutes bis zur Verarbeitung zu erhalten. Um zu beurteilen, ob das biologische Material für eine Manipulation geeignet ist, wird eine serologische Kontrolle auf Treponema pallidum, Hepatitis-B-Virus (HBV), Hepatitis-C-Virus (HCV) und Humanes Immundefizienz-Virus (HIV) durchgeführt, die alle negativ sein müssen. Für die vorliegende Studie wurde der Buffy Coat von IPST zu Untersuchungszwecken zur Verfügung gestellt, zusammen mit Informationen über das Entnahmedatum, die serologischen Ergebnisse, die Blutgruppe und das Alter des Spenders32 Jahre. Der Buffy Coat kann maximal 1 Tag bei Raumtemperatur bleiben.
1. Gewinnung von aus Monozyten gewonnenen dendritischen Zellen
HINWEIS: Es ist wichtig zu erwähnen, dass bei der Manipulation von menschlichem peripherem Blut spezifische universelle Sicherheitsvorkehrungen und eine angemessene Materialentsorgung in Betracht gezogen werden sollten. Bevor Sie beginnen, vergewissern Sie sich, dass alle erforderlichen Reagenzien und Materialien vorbereitet und einsatzbereit sind.
- Isolierung von mononukleären Zellen des peripheren Blutes
- Greife auf den menschlichen Buffy-Mantel zu.
HINWEIS: Buffy Coat ist ein Nebenprodukt, das aus Blut gewonnen wird, das durch Leukapherese32 gewonnen wird und durch Zentrifugation mit weißen Blutkörperchen angereichert wird. Alle Schritte wurden in einer Biosicherheitswerkbank (BSC) mit vertikaler Durchflusskammer durchgeführt. - Öffnen Sie die Verpackung des Buffy Coats, indem Sie das versiegelte Auslassröhrchen mit einem Skalpell durchschneiden, und füllen Sie den Inhalt in ein 50-ml-Röhrchen. Übertragen Sie 7 ml Buffy-Coat-Probe in ein steriles konisches 15-ml-Röhrchen und fügen Sie 6 ml phosphatgepufferte Kochsalzlösung (PBS) hinzu, um eine Vorwäsche durchzuführen. Dieser erste Waschschritt ist notwendig, um die Probe von der beträchtlichen Menge an roten Blutkörperchen (RBC) und Plasma zu reinigen, so dass die Probe für die Gradiententrennung mit einem Dichtegradientenmedium optimiert ist (siehe Materialtabelle).
- Das Röhrchen wird bei Raumtemperatur 10 min bei 1.100 x g in einer Zentrifuge mit Schwenkrotor und bei gelöster Bremse zentrifugiert (siehe Materialtabelle).
- Nach der Zentrifugation wird die Leukozytensuspension (der weiße Ring zwischen Plasma und Erythrozyten) mit einer Pasteurpipette aufgefangen und in ein neues steriles konisches 15-ml-Röhrchen überführt.
- Füllen Sie die Leukozytensuspension bis zu 10 ml mit PBS, um den nächsten Trennschritt zu unterstützen, und mischen Sie durch vorsichtiges Auf- und Abpipettieren.
- Bereiten Sie die Lösung des Dichtegradientenmediums (Dichte: 1,077 g/ml) vor: Geben Sie 3 ml Dichtegradientenmedium in ein neues steriles konisches 15-ml-Röhrchen und lassen Sie es auf Raumtemperatur erwärmen.
- 5 ml der verdünnten Leukozytensuspension (aus Schritt 1.1.5) werden in das konische Röhrchen mit dem Dichtegradientenmedium (5:3) gegeben, um eine Dichtegradiententrennung durchzuführen. Geben Sie die Probe langsam, Tropfen für Tropfen, unter Verwendung der Röhrchenwände hinzu, um eine Störung des Dichtegradientenmediums zu vermeiden.
- Gradientenabscheidung: Die Dichtegradientenmittelsuspension wird bei Raumtemperatur 30 min bei 1.100 x g in einer Zentrifuge mit Schwenkrotor und bei gelöster Bremse zentrifugiert.
- Nach dem Zentrifugieren die konischen Röhrchen vorsichtig aus der Zentrifuge entfernen. Nach diesem Schritt sind eine Reihe von gut definierten Schichten sichtbar, darunter die folgenden, beginnend von unten: eine rote Schicht (Erythrozyten und Granulozyten), ein Dichtegradientenmedium, eine dünne blasse Schicht aus mononukleären Zellen des peripheren Blutes (PBMC) und Plasma.
- Sammeln Sie die dünne Schicht PBMCs mit einer Pasteurpipette und vermeiden Sie es, das Dichtegradientenmedium darunter oder zu viel Plasma darüber aufzunehmen. Geben Sie die PBMC-Probe in ein neues konisches 50-ml-Röhrchen, füllen Sie es bis zu 25 ml mit PBS und mischen Sie die Probe durch vorsichtiges Auf- und Abpipettieren.
- Zentrifugieren Sie die Proben bei Raumtemperatur für 10 Minuten bei 600 x g (normale Bremse), um Restzellen und Ablagerungen abzuwaschen, und verwerfen Sie den Überstand, indem Sie das Röhrchen vorsichtig umdrehen.
HINWEIS: Wenn es zu viel Kontamination der roten Blutkörperchen gibt, was beobachtet werden kann, wenn das Zellpellet oder der Buffy Coat nicht vollständig abgetrennt ist oder rötlich erscheint, wird empfohlen, die verbleibenden Erythrozyten zu lysieren. In diesem Fall werden 5 ml Erythrozyten-Lysepuffer (siehe Materialtabelle) zugegeben, gründlich gemischt und 5 Minuten lang inkubiert. Füllen Sie bis zu 40 ml mit PBS, zentrifugieren Sie die Proben 10 Minuten lang bei Raumtemperatur bei 900 x g (normale Bremse) und verwerfen Sie den Überstand, indem Sie das Röhrchen vorsichtig umdrehen. - Füllen Sie die Probe auf 10 ml mit PBS auf und nehmen Sie ein Aliquot, um die Zellen zu zählen. Um Blutplättchen zu entfernen, zentrifugieren Sie bei Raumtemperatur 5 Minuten lang bei 400 x g (normale Bremse) und entsorgen Sie den Überstand, indem Sie das Röhrchen vorsichtig umdrehen.
HINWEIS: Falls eine beträchtliche Anzahl von Blutplättchen vorhanden ist, zentrifugieren Sie zweimal zweimal bei Raumtemperatur bei 200 x g (normale Bremse). Die Identifizierung der Blutplättchen erfolgt durch Visualisierung der Probe während der Zählung der Zellen.
- Greife auf den menschlichen Buffy-Mantel zu.
- Monozytenisolierung durch immunmagnetische Trennung
- Bereiten Sie den Mikroperlenpuffer vor, indem Sie PBS mit 0,5 % Rinderserumalbumin (BSA) und 2 mM Ethylendiamintetraessigsäure (EDTA) ergänzen. Sterilisieren Sie die Lösung durch Filtration (0,2 μm) und bewahren Sie den Puffer gekühlt auf (2-8 °C).
- Führen Sie die Isolierung von Monozyten CD14+ durch magnetisch aktivierte Zellsortierung (MACS) durch.
- Berechnen Sie nach der Zellzählung mit einem automatischen Zellzähler (Schritt 1.4.1) das entsprechende Volumen des Mikrokügelchenpuffers und der immunmagnetischen CD14-Kügelchen (siehe Materialtabelle). Stellen Sie sicher, dass diese Lösungen auf Eis gelegt werden. 80 μl Mikrokügelchenpuffer pro 1 x 10 7 Zellen und 20 μl CD14-Kügelchen pro 1 x 107 Zellen hinzufügen.
- Resuspendieren Sie das Zellpellet in den zuvor festgelegten Volumina und inkubieren Sie es 15 Minuten lang bei 4 °C (2-8 °C).
HINWEIS: Falls eine Überprüfung der Monozytenspiegel in den PBMC-Proben erforderlich ist, führen Sie eine durchflusszytometrische Analyse mit Färbeantikörpern (z. B. CD14 [monoklonales TÜK4]) durch. Befolgen Sie Schritt 3.2, um Details zur durchflusszytometrischen Analyse zu erhalten. - Geben Sie 1-2 ml Mikroperlenpuffer pro 1 x 107 Zellen hinzu, zentrifugieren Sie 10 Minuten lang bei Raumtemperatur bei 600 x g (normale Bremse), um ungebundene Kügelchen zu entfernen, und verwerfen Sie den Überstand, indem Sie das Röhrchen vorsichtig umdrehen.
- Bereiten Sie die LS-Spalte vor. Die LS-Säulen enthalten ferromagnetische Kugeln, die, wenn sie auf einem Magneten platziert werden, eine positive, sanfte Retention von magnetisch markierten Zellen33 ermöglichen. Unmittelbar vor dem Gebrauch eine LS-Säule (siehe Materialtabelle) auf den Magneten stellen, mit 3 ml Mikroperlenpuffer spülen, ohne vollständig auszutrocknen, und sofort mit dem nächsten Schritt fortfahren.
HINWEIS: Lassen Sie die Säule während des Eingriffs niemals austrocknen, um die Ausbeute nicht zu beeinträchtigen. - Resuspendieren Sie das Zellpellet in 500 μl Mikroperlenpuffer pro 1 x 108 Zellen. Wenn die Zellzahl größer als 4 x 108 ist, verwenden Sie ein 40-μm-Zellsieb, um eine Zellaggregation zu verhindern.
- Geben Sie die Zellsuspension in den Säuleneinlass, platzieren Sie ein konisches 15-ml-Röhrchen unter dem Säulenauslass, um die negative Zellfraktion aufzufangen, und waschen Sie die Säule dreimal mit 3 ml Mikroperlenpuffer. Die negative Fraktion umfasst die Zellen, die nicht mit den CD14-Kügelchen gesammelt wurden (d.h. die CD14− -Zellen).
- Nach dem letzten Waschen nehmen Sie die Säule vom Magneten, legen Sie sie auf ein steriles konisches 15-ml-Röhrchen, pipettieren Sie 5 ml Mikroperlenpuffer in den Säuleneinlass, führen Sie sofort den Spritzenkolben in den Säuleneinlass ein und drücken Sie, um die Zielzellen zu dosieren.
- Sammeln Sie die magnetisch markierten Zellen (CD14+ -Zellen) und nehmen Sie ein Aliquot, um die Zellen zu zählen, wie in Schritt 1.4.1 beschrieben.
- Beide Zellfraktionen, CD14−- und CD14+-Zellen, werden bei Raumtemperatur 10 min bei 600 x g (normale Bremse) zentrifugiert. Verwerfen Sie den Überstand, bewahren Sie die CD14+-Fraktion für die nächsten Schritte auf und bewahren Sie die CD14-Fraktion für zukünftige Assays, wie z. B. Co-Kultur-Assays, auf. Bei Bedarf können die Zellen aus der CD14−Fraktion in RPMI-1640 20% FBS und 10% DMSO bei −80 °C kryokonserviert werden.
- Differenzierung von Monozyten in dendritische Zellen
- Bereiten Sie das vollständige RPMI-1640-Medium vor, indem Sie das RPMI-1640-Basismedium (mit 11,1 mM Glukose) mit 10 % fötalem Kälberserum (FBS), 1 % 2 mM L-Glutamin, 1 % nicht-essentiellen Aminosäuren (NEAA), 1 % Natriumpyruvat und 1 % 100 μg/ml Penicillin/Streptomycin ergänzen (siehe Materialtabelle).
- Führen Sie eine Monozytendifferenzierung in Mo-DCs durch, die über ~5-6 Tage erfolgt.
- Berechnen Sie das Volumen des Mediums, das für die Anzahl der erhaltenen CD14+ -Zellen erforderlich ist, und bestücken Sie die Zellen gemäß dem folgenden Versuchsaufbau.
ANMERKUNG: In diesem Protokoll wurden die Zellen bei einer Konzentration von 1,3 x 106 Zellen/ml plattiert, um den Zelltod und Messfehler zu berücksichtigen, und das Medium wurde durch Zugabe von 1.000 U/ml GM-CSF und 750 U/ml IL-4 (siehe Materialtabelle) in ein vollständiges Medium und gründliches Mischen hergestellt. - Geben Sie das entsprechende Mediumvolumen zu den CD14+ -Zellen und resuspendieren Sie sie, indem Sie mit einer Pasteurpipette auf und ab pipettieren. Die Zellsuspension wird in 24-Well-Platten (pro Well: 1,3 x 106 Zellen/ml) aufgetragen und in einem Kulturinkubator bei 37 °C mit 5 % CO2 inkubiert.
- Wechseln Sie das Nährmedium und ergänzen Sie es alle 2-3 Tage mit frischen Zytokinen (in der Regel einmal pro Differenzierungsprozess). Entfernen Sie dazu vorsichtig die Hälfte des Nährmediums, ohne die Zellen zu stören. Die gleiche Menge frisches Medium mit der entsprechenden Konzentration an Zytokinen wird zugegeben, wie zuvor in der Anmerkung zu Schritt 1.3.2.1 beschrieben, und für die verbleibende Differenzierungszeit inkubiert.
HINWEIS: DCs sind, wenn sie sich von Monozyten unterscheiden, lose adhärente Zellen. Vollständig differenzierte unreife mo-DCs sind spindelförmige, frei schwebende und locker adhärente Zellen. Die Zellen können auch Rosetten bilden, insbesondere wenn sie reifsind 34. - Um die Zellen nach der Differenzierung zu sammeln, verwenden Sie eine Mikropipette, um die gesamte Zellsuspension in ein steriles konisches Röhrchen zu überführen, und waschen Sie die Vertiefungen der Platte zweimal mit PBS, indem Sie vorsichtig auf den Boden klopfen (Abbildung 1A).
HINWEIS: Vermeiden Sie es, die stark anhaftenden Zellen zu sammeln, da es sich wahrscheinlich um Makrophagen handelt. Um eine unsachgemäße Zellreifung oder -aktivierung zu vermeiden, stellen Sie sicher, dass die Zellen mit äußerster Vorsicht behandelt werden. - Zentrifugieren Sie die Zellen bei Raumtemperatur für 10 Minuten bei 180 x g (normale Bremse), um Rückstände oder abgestorbene Zellen zu entfernen, und resuspendieren Sie sie in das geeignete Medium/Puffer für den Versuchsaufbau.
- Berechnen Sie das Volumen des Mediums, das für die Anzahl der erhaltenen CD14+ -Zellen erforderlich ist, und bestücken Sie die Zellen gemäß dem folgenden Versuchsaufbau.
- Führen Sie die Reifung von Mo-DCs durch.
- Für den Fall, dass die Reifung von mo-DCs erforderlich ist, verwenden Sie eine Well-Platte oder einen Kolben unter Berücksichtigung des zuvor verwendeten Beispiels der Zellkonzentration (1,3 x 10 6 Zellen/ml) und verabreichen Sie einen Zytokincocktail, indem Sie das Medium mit einem Zytokincocktail ergänzen, der IL-1β (10 ng/ml),IL-6 (1.000 U/ml), Prostaglandin E2 (PGE2; 1 μg/ml) umfasst. und Tumornekrosefaktor-α (TNF-α; 10 ng/ml) (siehe Materialtabelle). Die Zellen werden bei 37 °C mit 5 %CO2 für 24 h oder 48 h inkubiert.
- Zellzählung und Lebensfähigkeit
- Führen Sie eine Zellzählung und eine Trypanblaufärbung durch.
- Um die Anzahl der Zellen und die Lebensfähigkeit einer Zellsuspension zu bestimmen, wird ein Aliquot von 10 μl aus der Zellsuspension entnommen und mit 10 μl Trypanblau (1:1 Verdünnung) gemischt.
- Nehmen Sie 10 μl der vorherigen Mischung und verwenden Sie den automatischen Zellzähler, um die Anzahl der Zellen gemäß den Anweisungen des Herstellers zu zählen.
HINWEIS: Wenn die Konzentration der Zellen zu hoch ist, verdünnen Sie das Aliquot und berücksichtigen Sie nach der Zellzählung den Verdünnungsfaktor in den Berechnungen. - Passen Sie die Zellzahl und das Medium/den Puffer für den Versuchsaufbau an.
- Bestimmung der Zelllebensfähigkeit und Apoptose30.
ANMERKUNG: In dieser Arbeit wurde nach der Sialidase-Behandlung (Abschnitt 2) der Viabilitätstest durchgeführt.- Die mo-DCs werden mit 5 μg/ml 7-Aminoactinomycin D (7-AAD) und Annexin V gefärbt und die Apoptose gemäß den Anweisungen des Herstellers bestimmt (siehe Materialtabelle).
- Analysieren Sie die Ergebnisse mit Hilfe der Durchflusszytometrie29,30.
- Führen Sie eine Zellzählung und eine Trypanblaufärbung durch.
2. Behandlung der Zellen mit Sialidase
HINWEIS: Nach der Differenzierung in mo-DCs sind die Zellen am sechsten Tag bereit für den Sialidase-Behandlungstest.
- Unter Berücksichtigung des gewünschten Versuchsaufbaus werden ~10 x 10 6 mo-DCs aus 10 Wells der 24-Well-Platten mit 1,3 x 106 Zellen/Well gesammelt und in ein neues steriles konisches 15-ml-Röhrchen überführt.
HINWEIS: Gehen Sie von einem Zellverlust aus. Typischerweise beträgt die gefundene Konzentration in diesem Stadium 1,3 x 106 Zellen/ml, da die mo-DCs und ihre Vorläufer nicht proliferieren und während der Differenzierung in mo-DCs einen Lebensfähigkeitsverlust von 20 % erleiden. - Bei Raumtemperatur 5-7 Minuten bei 300 x g (normale Bremse) zentrifugieren und den Überstand entsorgen, um abgestorbene Zellen und Ablagerungen zu entfernen.
- Fügen Sie 10 ml RPMI-1640-Medium (mit 11,1 mM Glukose) hinzu, zentrifugieren Sie es 4 Minuten lang bei Raumtemperatur bei 300 x g (normale Bremse), verwerfen Sie den Überstand, fügen Sie 2 ml RPMI-1640 hinzu und mischen Sie es gründlich.
- Geben Sie 1 ml Zellen in RPMI-1640 in neue sterile Mikroröhrchen, #1 und #2; Jedes Mikroröhrchen enthält ca. 5 x 106 Zellen.
- In das Mikroröhrchen #1 geben Sie 500 mU Sialidase von Clostridium perfringens (siehe Materialtabelle). Fügen Sie dem Mikroröhrchen #2 mit Mock behandelte Sialidase hinzu, bei der es sich um eine Negativkontrolle handelt, um zu bestätigen, ob die beobachteten Effekte in direktem Zusammenhang mit der Sialinsäureentfernung stehen und nicht auf Artefakte zurückzuführen sind. Bei der nachgeahmten Sialidase handelt es sich um hitzeinaktivierte Sialidase, die durch 20-minütiges Kochen des Enzyms bei 100 °C gewonnen wird.
- 60 min bei 37 °C inkubieren.
- Nach der Inkubation werden die Zellen in neue sterile konische 15-ml-Röhrchen mit der gleichen Nummerierung #1 und #2 gegeben. Geben Sie etwa 4 ml des vollständigen RPMI-1640-Mediums (mit 10 % FBS) in beide Röhrchen, um die enzymatische Reaktion zu stoppen.
- Bei Raumtemperatur 4 min bei 300 x g (normale Bremse) zentrifugieren und den Überstand entsorgen.
- Geben Sie 5 ml des vollständigen RPMI-1640-Mediums in jedes Röhrchen und geben Sie 1 ml Zellen pro Well.
3. Bestimmung des Sialinsäureprofils
- Lektin-Färbung
- Sammeln und waschen Sie die Zellen bei Raumtemperatur für 5 Minuten bei 300 x g (normale Bremse).
- Resuspendieren Sie die Zellen in RPMI-1640 + 10% FBS und verteilen Sie die Zellen (100.000/100 μl) in den Mikroröhrchen.
- Färbung für die Durchflusszytometrie in RPMI-1640 mit 10 % FBS unter Verwendung einer Konzentration von 0,01 mg/ml für jedes Lektin: Sambucus nigra (SNA) Lektin, Erdnussagglutinin (PNA) und Maackia amurensis (MAA) Lektin (siehe Materialtabelle). 30 min bei 4 °C inkubieren.
- Waschen Sie die Zellen mit 1 ml PBS mit 10 % FBS oder 10 % BSA und zentrifugieren Sie sie 4 Minuten lang bei Raumtemperatur bei 300 x g (normale Bremse).
- Zu den mit den biotinylierten Lektinen gefärbten Zellen werden 0,0005 mg/ml Streptavidin-PE (siehe Materialtabelle) zugegeben und 15 Minuten lang bei Raumtemperatur im Dunkeln inkubiert. Waschen Sie die Zellen mit 1 ml PBS und zentrifugieren Sie sie 4 Minuten lang bei Raumtemperatur bei 300 x g (normale Bremse).
- Verwerfen Sie den Überstand und geben Sie in jedes Röhrchen 300 μl 2 % Paraformaldehyd (PFA 2 %). Schützen Sie die Röhren vor Licht und lagern Sie sie bei Bedarf bis zur Datenerfassung bei 4 °C.
- Erfassen Sie die Daten mit einem Durchflusszytometer innerhalb von 1 Woche nach der Probenvorbereitung29,30.
- Durchflusszytometrie
- Resuspendieren Sie die Zellen in 1 ml PBS und entnehmen Sie die Probe mit einem Durchflusszytometer zur sofortigen Datenerfassung.
- Für eine verzögerte Datenerfassung resuspendieren Sie 300 μl 2 % PFA und erfassen die Daten innerhalb von 1 Woche.
- Konfokale Laser-Scanning-Mikroskopie
- Die Zellen werden auf polylysinbeschichtete Glasdeckgläser mit einem Durchmesser von 12 mm plattiert und 5 Minuten lang bei Raumtemperatur inkubiert.
- Zentrifugieren Sie die Deckgläser bei Raumtemperatur für 1 min bei 100 x g (normale Bremse), um die Zelladhäsion zu fördern.
- 30 Minuten bei Raumtemperatur mit 4 % PFA fixieren, vor dem Waschen mit 1 % BSA in PBS.
- Verwenden Sie FITC-konjugiertes SNA-Lektin (0,01 mg/ml), um α2,6-verknüpfte Sialinsäuren auf den Zelloberflächen zu färben (siehe Materialtabelle).
- Erfassen Sie Bilder mit einem konfokalen Mikroskop (siehe Materialtabelle).
- Wählen Sie nach der Z-Stapel-Verarbeitung repräsentative konfokale Querschnittsbilder aus.
- Analytische Quantifizierung der Färbeintensität mit der korrigierten Gesamtzellfluoreszenz (CTCF).
HINWEIS: CTCF = Integrierte Dichte − (Fläche der ausgewählten Zelle × Mittlere Fluoreszenz der Hintergrundmesswerte)29.
4. Reifeprofilierung von Mo-DCs
- Antikörperfärbung und Durchflusszytometrie
- Entnehmen Sie eine neue Probe der interessierenden Zellen, um eine Antikörperfärbung durchzuführen. Waschen Sie die Zellen 5 Minuten lang bei Raumtemperatur bei 300 x g (normale Bremse) und verteilen Sie die Zellen in den Mikroröhrchen (100.000 Zellen pro Röhrchen).
- Führen Sie die Färbung für die Durchflusszytometrie mit den gewünschten Antikörpern (ab), MHI-I, MHC-II, CD80 und CD86 durch (siehe Materialtabelle).
- Das fluoreszenzkonjugierte ab wird 15 min bei Raumtemperatur im Dunkeln inkubiert.
- Waschen Sie die Zellen mit 1 ml PBS und zentrifugieren Sie sie 5 Minuten lang bei Raumtemperatur bei 300 x g (normale Bremse).
HINWEIS: Wenn Sie unmarkiertes ab verwenden, fügen Sie fluoreszenzkonjugiertes sekundäres ab hinzu und inkubieren Sie es 15 Minuten lang im Dunkeln gemäß den Anweisungen des Herstellers. Waschen Sie die Zellen mit 1 ml PBS und zentrifugieren Sie sie 5 Minuten lang bei Raumtemperatur bei 300 x g (normale Bremse). - Geben Sie zu allen Mikroröhrchen bis zu 100 μl PBS, resuspendieren Sie die Zellen in 300 μl 2 % Paraformaldehyd (PFA 2 %) und halten Sie die Röhrchen bis zur Datenerfassung bei 4 °C im Dunkeln.
- Erfassen Sie die Daten mit einem Durchflusszytometer.
HINWEIS: Nach der Färbung und Fixierung können die Proben sofort oder innerhalb eines Zeitraums von 1 Woche durch Durchflusszytometrie entnommen werden. Lagern Sie die Röhrchen in diesem Fall bei 4 °C im Dunkeln.
5. Enzymgebundener Immunosorbent-Assay (ELISA)
HINWEIS: In dieser Arbeit wurde die IFN-γ-Produktion mit dem ELISA-Assay gemäß den Anweisungen des Herstellers gemessen (siehe Materialtabelle).
- Um die Platte mit einem Beschichtungspuffer zu beschichten, wird der Einfangantikörper verdünnt (1:100, Einfangantikörper in PBS), 100 μl dieser Arbeitslösung in jede Vertiefung überführt und über Nacht bei Raumtemperatur inkubiert.
- Verwerfen Sie den Fängerantikörper vollständig.
- Fügen Sie den Blockierungspuffer hinzu (z. B. PBS + 2 % BSA + 0,05 % Tween20) und inkubieren Sie 1 Stunde lang bei Raumtemperatur, bevor Sie den Blockierungspuffer entfernen.
- Den Standard und die Probe mit der jeweiligen Mischung und den Verdünnungen zugeben und 2 h bei Raumtemperatur inkubieren. Fünfmal mit Waschpuffer waschen.
- Fügen Sie den biotinylierten Detektorantikörper hinzu und inkubieren Sie ihn 2 Stunden lang bei Raumtemperatur, gefolgt von fünf Wäschen.
- Poly-HRP-Streptavidin-HS zugeben und 30 Minuten bei Raumtemperatur inkubieren, gefolgt von fünf Wäschen mit Waschpuffer.
- TMB-Substrat zugeben (siehe Materialtabelle) und bis zu 60 Minuten bei Raumtemperatur unter Berücksichtigung des verwendeten Testsystems inkubieren. Fünfmal mit Waschpuffer waschen.
- Lesen Sie die Proben auf einem Mikroplatten-Reader bei 450 nm ab.
Ergebnisse
Monozytenisolierung und Monozytendifferenzierung in mo-DCs
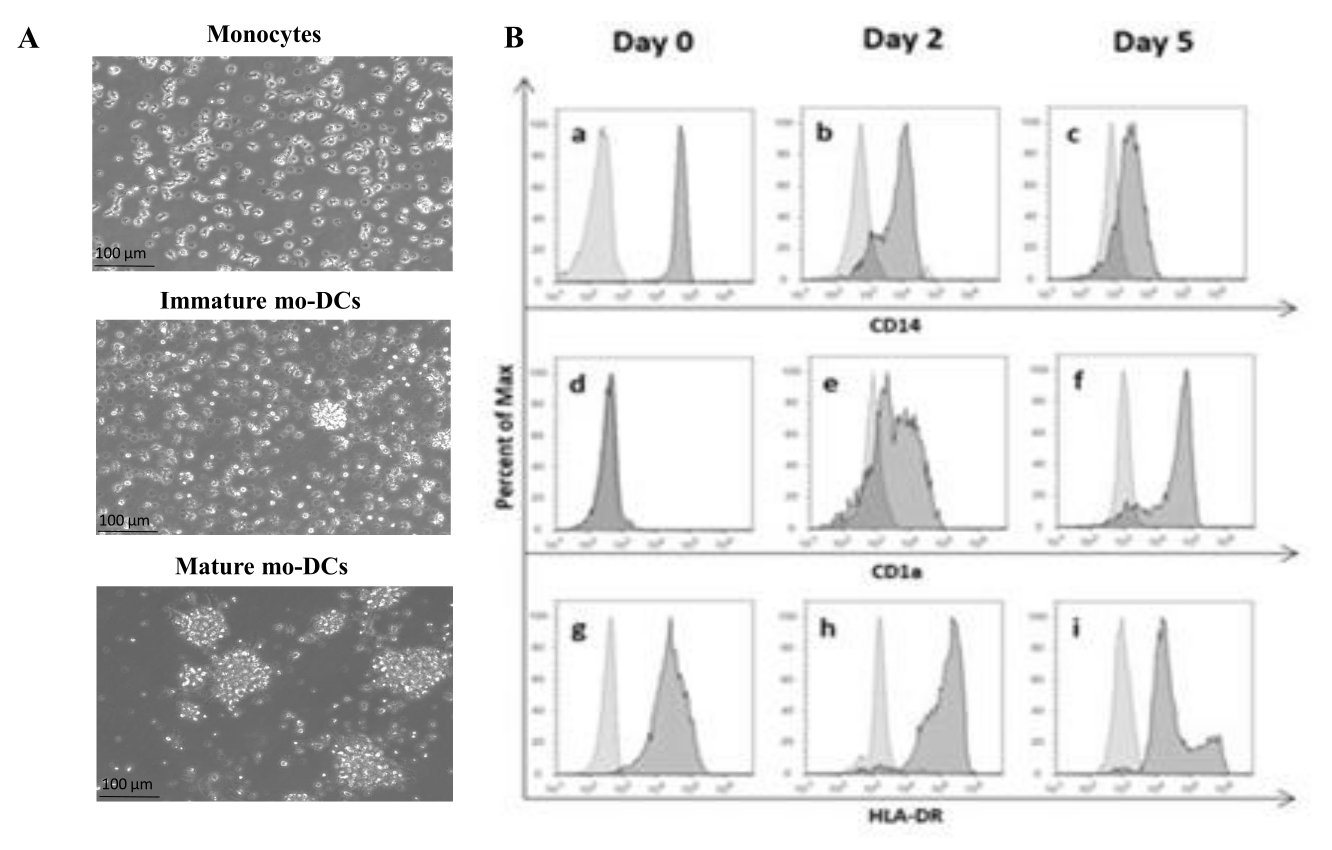
In Übereinstimmung mit dem Protokoll wurden humane PBMCs mittels Dichtegradiententrennung mit Dichtegradientenmedium aus dem Buffy Coat isoliert und gründlich gewaschen. Trypanblau wurde verwendet, um die Zählung lebensfähiger Zellen am Tag der Isolierung durchzuführen, wie zuvor in Schritt 1.4.1 beschrieben. Anschließend wurde die CD14+ Monozytenisolierung durch positive Selektion durchgeführt. Um dies zu erreichen, wurden PBMCs mit magnetischen Kügelchen inkubiert, die einen Antikörper enthielten, der das CD14-Antigen erkennt. Die ausgewählten CD14+ -Monozyten wurden in einem mit GM-CSF und IL-4 angereicherten Medium für 5-6 Tagekultiviert 27 , um sich in unreife mo-DCs zu differenzieren (Abbildung 1A). Die Reifung von mo-DCs kann durch die Anwendung eines Cocktails von Zytokinen, einschließlich IL-6, IL-1β, TNF-α und PGE235 , erreicht werden (Abbildung 1A).
Während des Differenzierungsprozesses wird erwartet, dass sich der Zellphänotyp infolge der IL-4- und GMCSF-Stimulation verändert. Die Daten zeigen, dass mo-DCs die Expression des Oberflächenmarkers CD14 verlieren, der hauptsächlich von Monozyten exprimiert wird (Abbildung 1B), und eine signifikante Expression von CD1a, einem Marker, der von humanen DCs exprimiert wird36,37. mo-DCs erhalten auch eine höhere MHC-II (HLA-DR)-Expression, ein Antigen-präsentierendes Molekül, das von menschlichen DCs und anderen Antigen-präsentierenden Zellen exprimiert wird38 (Abbildung 1B).
Sialinsäuregehalt auf der Zelloberfläche
Die Behandlung von mo-DCs mit Sialidase reduziert den Sialinsäuregehalt auf der Oberfläche von mo-DCs, was durch die Färbung mit Lektinen bestätigt werden kann, bei denen es sich um Proteine handelt, die in der Lage sind, an Kohlenhydrate zu binden39. Da das verwendete Enzym sowohl α2,3- als auch α2,6-verknüpfte Sialinsäuren von der Zelloberfläche entfernt, wurden mo-DCs mit PNA gefärbt, das T-Antigen-Galβ1-3GalNAcα1-Ser/Thr erkennt, sowie mit MAA- und SNA-Lektinen, die an α2,3- bzw. α2,6-Sialsäuren binden. Die Wirksamkeit der Sialidase-Behandlung wurde mittels Durchflusszytometrie und konfokaler Mikroskopie evaluiert (Abbildung 2). Wie in Abbildung 2A gezeigt, verringerte die Sialidase-Behandlung signifikant die MAA- und SNA-Bindung und erhöhte gleichzeitig die PNA-Färbung. Die Abnahme der SNA-Färbung nach Sialidase-Behandlung wurde durch konfokale mikroskopische Analysen bestätigt, die eine signifikant reduzierte SNA-Färbung an der Zelloberfläche zeigten (Abbildung 2B).
Funktionelle Charakterisierung von Sialidase-behandelten mo-DCs
Um zu beurteilen, wie sich die Behandlung mit Sialidase auf die Funktionen von mo-DC auswirkt, wurde der Reifungsgrad von mo-DCs nach der Behandlung mit Sialidase untersucht. Wie in Abbildung 3A dargestellt, führt die Sialidase-Behandlung zu einer signifikanten Steigerung der Expression der Antigen-präsentierenden Moleküle MHC I und MHC II sowie der Expression der kostimulatorischen CD80- und CD86-Moleküle. Um die Wirkung der Sialinsäureentfernung auf die Fähigkeit von mo-DCs, T-Zellantworten zu induzieren, zu bewerten, wurden mit Sialidase behandelte mo-DCs, die mit Tumorzelllysaten beladen waren, verwendet, um autologe T-Zellen zu primen (Abbildung 3). Als nächstes wurde das Profil der resultierenden T-Zellen basierend auf ihrer Fähigkeit, das Th1-Zytokin IFN-γ zu sezernieren, charakterisiert. Wie in Abbildung 3B gezeigt, sezernierten die T-Zellen, die durch Sialidase-behandelte Mo-DCs geprimt wurden, im Vergleich zu T-Zellen, die durch vollständig sialylierte mo-DCs geprimt wurden, signifikant höhere Konzentrationen von IFN-γ. Diese Ergebnisse deuten darauf hin, dass Sialidase-behandelte mo-DCs eine verbesserte Fähigkeit haben, autologe T-Zellen zu primen.
Lebensfähigkeit der Zellen
Nach der Behandlung mit Sialidase wurde ein Viabilitätstest durchgeführt, um sicherzustellen, dass die Behandlung nicht zytotoxisch für die Zellen war. Nach der Behandlung wurden mo-DCs mit 7-AAD und Annexin V gefärbt, um nicht lebensfähige und apoptotische Zellen zu detektieren, und mittels Durchflusszytometrie analysiert (Abbildung 4). Die Daten zeigen keinen signifikanten Unterschied in der Zelllebensfähigkeit zwischen unbehandelten (Abbildung 4, linkes Bild) und Sialidase-behandelten Zellen (Abbildung 4, rechtes Bild).

Abbildung 1: Differenzierung isolierter Monozyten in mo-DCs. (A) CD14+-Monozyten wurden aus Buffy Coats isoliert und bei einer Konzentration von 1,3 x 106 Zellen/ml bei 37 °C kultiviert. Die Monozyten wurden in einem Medium differenziert, das mit 750 U/ml IL-4 und 1.000 U/ml GM-CSF angereichert war. Mikroskopische Analyse der Morphologie von Monozyten, die am Tag 0 aus dem menschlichen Buffy-Fell isoliert wurden (Bild oben). Unreife mo-DCs; Die Zellen wurden über einen Zeitraum von 5 Tagen mit IL-4 und GM-CSF differenziert (mittleres Bild). Gereifte mo-DCs wurden unter Verwendung von IL-6-, IL-1β-, TNF-α- und PGE2-Zytokinen für 24 h erhalten (unteres Bild). Maßstabsbalken: 100 μm. (B) Die Zellen wurden während des gesamten Differenzierungszeitraums an Tag 0, Tag 2 und Tag 5 mittels Durchflusszytometrie analysiert. Zur Charakterisierung von Zelloberflächenmarkern wurden folgende Antikörper verwendet: (a-c) CD14; (D-F) CD1a und (g-i) HLA-DR (MHC-Klasse II). Die Abbildung zeigt repräsentative Histogramme von mindestens drei unabhängigen Assays. Panel (B) wurde gegenüber Videira et al.40, Patent WO2017002045A1, modifiziert. Bitte klicken Sie hier, um eine größere Version dieser Abbildung anzuzeigen.
{kind=link}

Abbildung 2: Sialidase-Behandlung von humanen mo-DCs zur Entfernung von α2,6- und α2,3-verknüpften Sialinsäuren von der Zelloberfläche. (A) Analyse von mo-DCs mittels Durchflusszytometrie mittels Lektinfärbung, um die Wirksamkeit der Sialidase-Behandlung zu testen. Humane Mo-DCs wurden mit Sialidase (graue Balken) behandelt oder unbehandelt gelassen (weiße Balken) und mit SNA-Lektin (Erkennung von [2,6]-Sialinsäuren), MAA-Lektin (Erkennung von [2,3]-Sialinsäuren) und PNA-Lektin (Erkennung von T-Antigen-Galβ1-3GalNAcα1-Ser/Thr) gefärbt. Die Werte stellen die mittlere Fluoreszenzintensität (MFI) von mindestens drei unabhängigen Assays dar. Die statistische Signifikanz wurde mit einem zweiseitigen gepaarten t-Test (*P < 0,05 oder ***P < 0,0001) bestimmt, der sich auf die Differenz zwischen den unbehandelten und den mit Sialidase behandelten DCs bezieht. Die Behandlung mit Sialidase verringerte die MAA-Bindung und erhöhte die PNA-Färbung in humanen mo-DCs, was auf die Entfernung von α(2,3)-verknüpften Sialinsäuren zurückzuführen ist. Die Entfernung von α(2,6)-verknüpften Sialinsäuren nach Sialidase-Behandlung wurde durch eine Abnahme der SNA-Färbung nachgewiesen. (B) Konfokalmikroskopie von mo-DCs, die mit Sialidase behandelt und auf Deckgläsern zur Beobachtung vorbereitet wurden. Eine Reihe von Z-Stapel-Bildern wurde von verschiedenen Zellen gesammelt und verarbeitet, um die mittlere Färbeintensität einzubeziehen. Maßstabsleisten: 20 μm. Panel (A) wurde von Silva et al.30 modifiziert; Panel (B) wurde von Silva et al.29 modifiziert. Bitte klicken Sie hier, um eine größere Version dieser Abbildung anzuzeigen.
{kind=link}

Abbildung 3: Sialidase-Behandlung von mo-DCs, die eine höhere Expression von Reifungsmarkern induziert. (A) mo-DCs, die mit Sialidase behandelt wurden, zeigten einen höheren Reifungsphänotyp als vollständig sialylierte mo-DCs. Mittels Durchflusszytometrie wurde die Expression verschiedener Reifungsmarker untersucht. Die mo-DCs wurden mit Sialidase für 1 h bei 37 °C behandelt; Die Diagrammwerte stellen die mittlere Fluoreszenzintensität (MFI) (Mittelwert ± SEM) von mindestens drei unabhängigen Assays dar. Statistisch signifikante Unterschiede wurden mit einem t-Test (*P < 0,05, **P < 0,01) berechnet, der sich auf den Unterschied zwischen den unbehandelten und den mit Sialidase behandelten Bedingungen bezieht. (B) Desialylierte humane Mo-DCs, die mit ganzen Tumorantigenen beladen waren, induzierten spezifische T-Zell-Antworten. Die mo-DCs wurden mit Sialidase für 1 h bei 37 °C behandelt oder unbehandelt gelassen, gefolgt von der Beladung mit MCF-7-Lysaten (TL) als Quelle für Ganztumorzellantigene. Die Co-Kultur zwischen mo-DCs und autologen T-Zellen wurde für 4-8 Tage in Gegenwart von IL-2 (10 U/ml) durchgeführt. T-Zellen, die mit desialylierten mo-DCs geprimt wurden, zeigten eine signifikant höhere Sekretion des Th1-Zytokins IFN-γ. Nach T-Zell-Stimulation mit mo-DCs wurden die in die Co-Kultur-Überstände sezernierten Zytokine mittels ELISA gemessen (n = 7). Die Diagrammwerte stellen die Konzentration (pg/mL) (Mittelwert ± SEM) dar. Statistisch signifikante Unterschiede wurden mit einem t-Test (*P < 0,05) berechnet. Die Abbildung wurde von Silva et al.30 modifiziert. Bitte klicken Sie hier, um eine größere Version dieser Abbildung anzuzeigen.
{kind=link}

Abbildung 4: Mangelnder Einfluss der Sialidase-Behandlung auf die Lebensfähigkeit humaner Mo-DCs. Unbehandelte mo-DCs (linkes Bild) und Sialidase-behandelte mo-DCs (rechtes Bild) wurden einer Doppelfärbung mit Annexin V und 7-AAD unterzogen, und die Färbung wurde mittels Durchflusszytometrie analysiert. Die Daten zeigten keinen signifikanten Unterschied in der Zelllebensfähigkeit zwischen den unbehandelten und den mit Sialidase behandelten Zellen, was darauf hindeutet, dass mo-DCs eine Sialidase-Behandlung tolerieren können und lebensfähig bleiben, um ihre immunologische Funktion auszuüben. Die Abbildung wurde von Silva et al.30 modifiziert. Bitte klicken Sie hier, um eine größere Version dieser Abbildung anzuzeigen.
{kind=link}
Diskussion
Isolierung von Monozyten
Dieses Manuskript beschreibt ein Protokoll zur Erzeugung von mo-DCs aus humanen isolierten Monozyten CD14+ (Abbildung 1A), gefolgt von einer Sialidase-Behandlung, um den Sialinsäuregehalt auf der Oberfläche dieser Zellen zu reduzieren.
Es gibt verschiedene Möglichkeiten, menschliche DCs zu gewinnen, z. B. direkt aus peripherem Blut oder Gewebe oder durch Differenzierung von Vorläufern wie Stammzellen oder Monozyten. Die Gewinnung von DCs, die von Monozyten unterschieden werden, die aus peripherem Blut isoliert wurden, ist aufgrund der Leichtigkeit, hohe Mengen an Monozyten zu erhalten, im Vergleich zu anderen DC-Quellenweitaus einfacher 41. Um jedoch einen hohen Prozentsatz an isolierten Monozyten zu erhalten, müssen alle Protokollschritte sorgfältig befolgt werden. Zum Beispiel kann das Dichtegradientenmedium für die Zellen toxisch sein, und um den Zelltod zu verhindern, muss man einen längeren Zellkontakt mit dem Dichtegradientenmedium vermeiden und die Zellen gründlich waschen. Die Zellmanipulation muss so schnell wie möglich erfolgen, um den Verlust der Lebensfähigkeit der Zellen zu vermeiden. Aus PBMCs können Monozyten durch Positivselektion mit der Methode der magnetisch aktivierten Zellsortierung (MACS) isoliert werden, die eine geeignete Technologie ist, um eine hohe Anzahl von Monozyten zu erhalten. Darüber hinaus besitzen mo-DCs, die aus MACS-isolierten Monozyten gewonnen werden, im Vergleich zu anderen Monozytenselektionsmethoden eine größere Fähigkeit, die Anti-Tumor-T-Zellaktivität zu stimulieren42. In diesem Protokoll wurden die Monozyten nach der Isolierung über einen Zeitraum von 5-6 Tagen mit IL-4 und GM-CSF inkubiert, um die Differenzierung in unreife mo-DCs zu erreichen (Abbildung 1). Die Ergebnisse zeigten, dass sich die isolierten Monozyten morphologisch (Abbildung 1A) und phänotypisch (Abbildung 1B) zu unreifen mo-DCs differenzierten. Darüber hinaus verloren die mo-DCs während der Differenzierung die Expression von CD14-Markern und erhielten die Expression von CD1a und MHC-II (Abbildung 1B), die für die Antigenpräsentation in T-Zellen erforderlich sind.
Diese Isolierung und Differenzierung von Monozyten in mo-DCs sind Einschränkungen dieses Protokolls. Der Isolierungsprozess ist ein sensibler Schritt, der sorgfältig und schnell durchgeführt werden muss, um den Zelltod zu vermeiden, und dieser Schritt muss auch jedes Mal durchgeführt werden, wenn mo-DCs für ein neues Experiment benötigt werden. Der Differenzierungsprozess dauert 5-6 Tage, was eine Schwierigkeit bei der Anwendung dieser Methode für Hochdurchsatzanalysen darstellt. Nichtsdestotrotz sind die Isolierungsmethode und die Verwendung von Zytokinen zur Differenzierung von mo-DCs nützlich, um eine hohe Anzahl von funktionellen mo-DCs in vitro zu Versuchszwecken zu erzeugen. Die in diesem Protokoll erzeugten mo-DCs sind in der Lage, sich einer Sialidase-Behandlung, Durchflusszytometrie, ELISA, konfokaler Mikroskopie usw. zu unterziehen, was die Bedeutung und Nützlichkeit dieser Methode unterstreicht30.
Behandlung mit unreifen mo-DCs und Sialidase
Sialidasen sind essentiell für die Regulation der Sialylierung und sind für die Entfernung von Sialinsäuren aus den Glykanen der Zelloberfläche verantwortlich. In mo-DCs führt die Sialinsäureentfernung durch Sialidase zur Reifung dieser Zellen, was die Antigen-Kreuzpräsentation und die anschließende T-Zell-Aktivierung und Anti-Tumor-Aktivität erhöht30.
Unreife humane mo-DCs weisen im Vergleich zu reifen mo-DCs einen hohen Gehalt an α(2,6)- und α(2,3)-verknüpften Sialinsäurenauf der Zelloberfläche auf 27 im Vergleich zu reifen mo-DCs31,43. Darüber hinaus verbessert die Entfernung von Sialinsäuren durch Behandlung von mo-DCs mit Sialidase die Reifung der DCs 28,30,31. Die für dieses Experiment ausgewählte Sialidase stammte aus dem Bakterium Clostridium perfringens. Aber auch andere Organismen produzieren Sialidasen, wie das Bakterium Streptococcus pneumoniae, Vibrio cholerae oder Salmonella typhimurium44, der Blutegel Macrobdella decora45 und sogar Homo sapiens46, und auch Sialidasen aus diesen Organismen werden experimentell verwendet. Jede Sialidase hat jedoch unterschiedliche Substratspezifitäten. Darüber hinaus kann die Verwendung des Sialidase-Enzyms seine Grenzen haben. So kann beispielsweise die Manipulation von mo-DCs während der Behandlung diese Zellen weiter stimulieren. Darüber hinaus müssen die Menge an Sialidase und die Inkubationszeiten basierend auf der Art der verwendeten Zellen und ihrer Sialinsäurezusammensetzung optimiert werden. Die Sialinsäureentfernung ist kein dauerhafter Effekt, sondern ein vorübergehendes Phänomen, da die Zelle ihren Sialinsäuregehalt an der Zelloberfläche wiederherstellt. Neben der Sialidase gibt es noch andere Methoden, um die Sialinsäuremoleküle an der Oberfläche von Zellen zu reduzieren, wie z.B. die Verwendung von Sialyltransferase-Inhibitoren, Gen-Knockouts von Sialyltransferase-Genen oder die metabolische Blockade von Sialinsäure mit Sialinsäuremimetika47,48,49. Nichtsdestotrotz können diese Methoden unterschiedliche Auswirkungen auf Zellen haben, und neben der Desialylierung muss auch die Lebensfähigkeit der Zellen berücksichtigt werden. Die Sialidase-Enzymbehandlung ist eine praktische Methode, um Sialinsäuren auf der Zelloberfläche effektiv und vorübergehend zu entfernen und gleichzeitig die Lebensfähigkeit der Zellen zu erhalten.
In dieser Arbeit wurde Sialidase zu den unreifen mo-DCs in einer Konzentration von 500 mU/5 x 106 Zellen/ml zugegeben, und die Zellen wurden bei 37 °C für 60 min inkubiert. Die Behandlung wurde mit RPMI-1640 ohne Serum durchgeführt, um die Lebensfähigkeit der Zellen zu erhalten und Wechselwirkungen zwischen den im Serum vorhandenen sialylierten Molekülen zu vermeiden30. Die Behandlung mit Sialidase kann neben RPMI auch mit anderen Puffern durchgeführt werden, z. B. mit 50 mM Natriumacetat, pH 5,1 (im Fall von C. perfingens sialidase) oder PBS50,51,52. Nichtsdestotrotz ist RPMI-1640 das gebräuchlichste Nährmedium für DCs, da es während des Eingriffs konstante Versuchsbedingungen aufrechterhält, die Induktion der Reifung vermeidet und den Stress reduziert, der durch Sialidase-Puffer oder PBS53,54,55,56 verursacht werden kann. Nach der Inkubation mit Sialidase ist es wichtig, die Zellen gründlich mit einem mit Serum angereicherten Medium zu waschen, um sicherzustellen, dass die Enzymreaktion gestoppt wurde. Das Vorhandensein von sialylierten Molekülen im Serum konkurriert als Substrate für Sialidase und gewährleistet so einen schnellen Reaktionsstopp.
Charakterisierung von Oberflächenmarkern durch Durchflusszytometrie und konfokale Mikroskopie
Für die Bestimmung des Sialinsäureprofils haben wir in Protokollabschnitt 3 die Lektinfärbung verwendet, gefolgt von Durchflusszytometrie und konfokaler Laser-Scanning-Mikroskopie. Für die Zellfärbung wurden in beiden Fällen die Lektinkonzentrationen und Inkubationsbedingungen optimiert, um eine Zellagglutination und einen Zelltod zu vermeiden. Es ist wichtig, die Inkubation bei 4 °C in Puffern durchzuführen, die mindestens 2 % FBS oder BSA enthalten, um eine unspezifische Bindung der Lektine zu vermeiden. In diesem Protokoll wurde RPMI-1640 mit 10 % FBS verwendet, um konstante Versuchsbedingungen aufrechtzuerhalten und Zellstress zu vermeiden. In Bezug auf die konfokale Mikroskopie ist die Fixierung der Zellen vor der Färbung unerlässlich, um die Morphologie zu erhalten, die Autolyse zu verhindern und die Antigenität aufrechtzuerhalten.
Die Analyse des mo-DC-Phänotyps mittels Durchflusszytometrie zeigte, dass mit Sialidase behandelte mo-DCs im Vergleich zu MMA- und SNA-Lektinen eine signifikant höhere Menge an PNA-Lektin aufwiesen, die nach der Sialidase-Behandlung abnahm (Abbildung 2A). Wie erwartet, nahm die PNA-Färbung zu, da PNA nicht-sialylierte Antigene erkennt, im Gegensatz zu MAA und SNA, die direkt an α2,3- bzw. α2,6-Sialinsäuren binden30. Diese Färbung bestätigt die effektive Entfernung von Sialinsäuren von der Zelloberfläche unter Verwendung dieses Protokolls. Eine weitere Methode, die zur Validierung der Behandlung und zur Analyse des Sialinsäuregehalts auf der Zelloberfläche verwendet werden kann, ist die Lektinfärbung, gefolgt von konfokaler Mikroskopie, wie in Abbildung 2B dargestellt.
Neben den erstgenannten Beispielen gibt es alternative Ansätze zur Bewertung und Charakterisierung des Sialinsäuregehalts, wie z.B. die Lektinsondierung mittels Western Blotting. Alternative Sialinsäure-spezifische Lektine sind ebenfalls erhältlich, wie z. B. Siglecs, eine Gruppe von Lektinen, die eine deutliche Präferenz für Sialinsäuretypen und -bindungen haben57. Neben der Verwendung von Lektinen in beiden Techniken (Durchflusszytometrie, Mikroskopie oder Western Blot) ist es auch möglich, den Sialinsäuregehalt mit Antikörpern zu charakterisieren. Zum Beispiel können α2,8-Sialinsäuren durch Antikörper wie Klon 735 bestimmt werden, der spezifisch für Polysialinsäure58 ist. Darüber hinaus können Zellen nach der Behandlung mit Sialidase funktionell auf ihre biologische oder therapeutische Wirksamkeit getestet werden, indem ihr Phänotyp und ihre Fähigkeit, T-Zellen zu aktivieren, bewertetwerden 40. Wie in den Beispielen gezeigt, zeigten Sialidase-behandelte mo-DCs einen höheren Reifungsphänotyp sowie eine erhöhte Expression von Antigen-präsentierenden und kostimulatorischen Molekülen.
Darüber hinaus können Sialidase-behandelte mo-DCs mit Antigenen beladen und mit T-Zellen oder anderen Zellen kokultiviert werden, um dann hinsichtlich des Phänotyps, des Zytokinsekretionsprofils oder anderer Merkmale untersucht zu werden. Im vorliegenden Beispiel zeigen die Daten, dass mit Sialidase behandelte mo-DCs mit Tumorantigenen beladen und dann zur Aktivierung von T-Zellen verwendet werden können. Tatsächlich zeigten die resultierenden T-Zellen eine erhöhte IFN-γ-Sekretion, was mit früheren Berichten über die Wirkung des Sialinsäuremangels auf die Steigerung der Fähigkeit von mo-DCs, T-Zellen zu aktivieren, übereinstimmt 27,28,29,30,31.
Zusammenfassend zeigt dieses Protokoll eine machbare, praktikable und praktische Methode zur Erzeugung von mo-DCs zur Manipulation des Sialinsäuregehalts durch Behandlung mit Sialidase. Dieses Protokoll stellt eine Methodik dar, die verschiedenen Zwecken und Anwendungen dienen kann. Diese Methode kann nicht nur eine entscheidende Rolle beim Verständnis der Rolle von Sialinsäuren bei der Reifung und Reaktion von Immunzellen spielen, sondern auch als immunmodulatorisches Werkzeug eingesetzt werden.
Offenlegungen
Die Autoren erklären, dass keine konkurrierenden finanziellen Interessen oder andere Interessenkonflikte bestehen.
Danksagungen
Die Autoren danken für die Finanzierung durch die Europäische Kommission GLYCOTwinning GA 101079417 und EJPRD/0001/2020 EU 825575; die Fundação para a Ciência e Tecnologia (FCT) Portugal im Rahmen der Zuschüsse FCT 2022.04607.PTDC, UIDP/04378/2020, UIDB/04378/2020 (UCIBIO) und LA/P/0140/2020 (i4HB). FCT-NOVA. und Stemmatters wurden auch vom Fundo Europeu de Desenvolvimento Regional (FEDER) über das Programa Operacional Regional do Norte (Norte 2020) für die SI I& DT DCMatters-Projekt (NORTE-01-0247-FEDER-047212). Wir erkennen die Einrichtung von Biolabs bei FCT-NOVA und GLYCOVID NOVA Saude an.
Materialien
| Name | Company | Catalog Number | Comments |
| 15 mL conical tube | AstiK’s | CTGP-E15-050 | Obtaining Monocyte-derived Dendritic Cells; Treatment of Cells with Sialidase |
| 24-well plate | Greiner Bio-one | 662 160 | Obtaining Monocyte-derived Dendritic Cells; Treatment of Cells with Sialidase |
| 50 mL conical tube | AstiK’s | CTGP-E50-050 | Obtaining Monocyte-derived Dendritic Cells |
| 7-Aminoactinomycin D (7-AAD) | BioLegend | 420404 | Obtaining Monocyte-derived Dendritic Cells |
| Annexin V | Immunotools | 31490013 | Obtaining Monocyte-derived Dendritic Cells |
| Attune Acoustic Focusing Flow Cytometer | Thermo Fisher Scientific | Determination of Sialic Acid Profile; Maturation Profiling of mo-DCs | |
| BSA | Sigma - Aldrich | A3294-100G | Obtaining Monocyte-derived Dendritic Cells; Determination of Sialic Acid Profile |
| CD14 (Monoclonal TÜK4) | Miltenyi Biotec | 130-080-701 | Obtaining Monocyte-derived Dendritic Cells |
| CD80 | Immunotools | 21270803 | Maturation Profiling of mo-DCs |
| CD86 | Immunotools | 21480863 | Maturation Profiling of mo-DCs |
| Cell counting slides and trypan blue | EVE | EVS-050 | Obtaining Monocyte-derived Dendritic Cells |
| Centrifuge | Eppendorf | 5430 R | Obtaining Monocyte-derived Dendritic Cells; Treatment of Cells with Sialidase; Determination of Sialic Acid Profile; Maturation Profiling of mo-DCs |
| Density gradient medium (Histopaque) | Sigma - Aldrich | 10771-100ML | Obtaining Monocyte-derived Dendritic Cells |
| EDTA | Gibco, ThermoFisher | 15400054 | Obtaining Monocyte-derived Dendritic Cells |
| Elisa kit (IFN-γ) | Immunotools | 31673539 | Maturation Profiling of mo-DCs |
| EVE automated cell count | NanoEntek | 10027-452 | Obtaining Monocyte-derived Dendritic Cells |
| Fetal bovine serum (FBS) | Gibco | 10500064 | Obtaining Monocyte-derived Dendritic Cells; Treatment of Cells with Sialidase; Determination of Sialic Acid Profile |
| Granulocyte-macrophage colony-stimulating factor (GM-CSF) | Miltenyi Biotec | 130-093-864 | Obtaining Monocyte-derived Dendritic Cells |
| Human CD14 microbeads (Immunomagnetic beads) | Miltenyi Biotec | 130-050-201 | Obtaining Monocyte-derived Dendritic Cells |
| Interleukin (IL)-1β | Sigma - Aldrich | I9401 | Obtaining Monocyte-derived Dendritic Cells |
| Interleukin (IL)-4 | Miltenyi Biotec | 130-093-919 | Obtaining Monocyte-derived Dendritic Cells |
| Interleukin (IL)-6 | Sigma - Aldrich | SRP3096 | Obtaining Monocyte-derived Dendritic Cells |
| L-glutamine | Gibco | A2916801 | Obtaining Monocyte-derived Dendritic Cells |
| LS column and plunger | Miltenyi Biotec | 130-042-401 | Obtaining Monocyte-derived Dendritic Cells |
| Maackia amurensis (MAA) lectin (MAA lectin - Biotinylated) | Vector labs | B-1265-1 | Determination of Sialic Acid Profile |
| MHC-I (HLA-ABC) | Immunotools | 21159033 | Maturation Profiling of mo-DCs |
| MHC-II (HLA-DR) | Immunostep | HLADRA-100T | Maturation Profiling of mo-DCs |
| Microtubes | AstiK’s | PCRP-E015-500 | Obtaining Monocyte-derived Dendritic Cells; Treatment of Cells with Sialidase; Determination of Sialic Acid Profile; Maturation Profiling of mo-DCs |
| Neuraminidase (Sialidase) | Roche | 11585886001 | Treatment of Cells with Sialidase |
| Non-essential amino acids (NEAA) | Gibco | 11140-050 | Obtaining Monocyte-derived Dendritic Cells |
| Paraformaldehyde (PFA 2%) | Polysciences Europe | 25085-1 | Determination of Sialic Acid Profile; Maturation Profiling of mo-DCs |
| Paraformaldehyde (PFA 4%) | Biotium | 22023 | Determination of Sialic Acid Profile |
| Pasteur pipettes | Labbox | PIPP-003-500 | Obtaining Monocyte-derived Dendritic Cells |
| Peanut (Arachis hypogaea) Agglutinin (PNA) lectin (PNA lectin - FITC) | Vector labs | FL-1071 | Determination of Sialic Acid Profile |
| Penicillin/streptomycin | Gibco | 15140163 | Obtaining Monocyte-derived Dendritic Cells |
| Phosphate Buffered Saline (PBS) | NZYTech | MB18201 | Obtaining Monocyte-derived Dendritic Cells; Treatment of Cells with Sialidase; Determination of Sialic Acid Profile; Maturation Profiling of mo-DCs |
| Prostaglandin E2 (PGE2) | Sigma - Aldrich | P0409 | Obtaining Monocyte-derived Dendritic Cells |
| RBC lysis buffer | BioLegend | 420302 | Obtaining Monocyte-derived Dendritic Cells |
| RPMI-1640 medium (containing 11.1 mM glucose) | Gibco | 31870074 | Obtaining Monocyte-derived Dendritic Cells; Treatment of Cells with Sialidase; Determination of Sialic Acid Profile |
| Sambucus nigra lectin (SNA lectin - Biotinylated) | Vector labs | B-1305-2 | Determination of Sialic Acid Profile |
| Sambucus nigra lectin (SNA lectin - FITC) | Vector labs | FL-1301-2 | Determination of Sialic Acid Profile |
| Sodium pyruvate | Thermofisher | 11360-070 | Obtaining Monocyte-derived Dendritic Cells |
| SpectroMax190 | Molecular Devices | Maturation Profiling of mo-DCs | |
| Streptavidin-PE | BioLegend | 405203 | Determination of Sialic Acid Profile; Maturation Profiling of mo-DCs |
| Tetramethylbenzidine (TMB) | Sigma - Aldrich | T0440 | Maturation Profiling of mo-DCs |
| Tumour necrosis factor-α (TNF-α) | Sigma - Aldrich | H8916 | Obtaining Monocyte-derived Dendritic Cells |
| Zeiss LSM710 confocal microscope | Zeiss | Determination of Sialic Acid Profile |
Referenzen
- Varki, A., Gagneux, P. Multifarious roles of sialic acids in immunity. Annals of the New York Academy of Sciences. 1253, 16-36 (2012).
- Bochner, B. S., Zimmermann, N. Role of Siglecs and related glycan-binding proteins in immune responses and immunoregulation. The Journal of Allergy and Clinical Immunology. 135 (3), 598-608 (2015).
- Smith, B. A. H., Bertozzi, C. R. The clinical impact of glycobiology: Targeting selectins, Siglecs and mammalian glycans. Nature Reviews Drug Discovery. 20, 217-243 (2021).
- Schauer, R. Sialic acids as regulators of molecular and cellular interactions. Current Opinion in Structural Biology. 19 (5), 507-514 (2009).
- Manhardt, C. T., Punch, P. R., Dougher, C. W. L., Lau, J. T. Y. Extrinsic sialylation is dynamically regulated by systemic triggers in vivo. Journal of Biological Chemistry. 292 (33), 13514-13520 (2017).
- Cabral, M. G., et al. Human dendritic cells contain cell surface sialyltransferase activity. Immunology Letters. 131 (1), 89-96 (2010).
- Bordron, A., et al. Hyposialylation must be considered to develop future therapies in autoimmune diseases. International Journal of Molecular Sciences. 22 (7), 3402(2021).
- Julien, S., Videira, P. A., Delannoy, P. Sialyl-Tn in cancer: (How) did we miss the target. Biomolecules. 2 (4), 435-466 (2012).
- Munkley, J. Aberrant sialylation in cancer: Therapeutic opportunities. Cancers. 14 (17), 4248(2022).
- Dennis, J. W., Laferte, S., Waghorne, C., Breitman, M. L., Kerbel, R. S. S1-6 Branching of Asn-linked oligosaccharides is directly associated with metastasis. Science. 236 (4801), 582-585 (1987).
- Pinho, S. S., Reis, C. A. Glycosylation in cancer: Mechanisms and clinical implications. Nature Reviews Cancer. 15 (9), 540-555 (2015).
- Manni, M., Läubli, H. Targeting glyco-immune checkpoints for cancer therapy. Expert Opinion on Biological Therapy. 21 (8), 1063-1071 (2021).
- Sjögren, J., Lood, R., Nägeli, A. On enzymatic remodeling of IgG glycosylation; Unique tools with broad applications. Glycobiology. 30 (4), 254-267 (2020).
- Trastoy, B., et al. Sculpting therapeutic monoclonal antibody N-glycans using endoglycosidases. Current Opinion in Structural Biology. 72, 248-259 (2022).
- Pascoal, C., et al. Sialyl LewisX/A and cytokeratin crosstalk in triple negative breast cancer. Cancers. 15 (3), 731(2023).
- von Itzstein, M., et al. Rational design of potent sialidase-based inhibitors of influenza virus replication. Nature. 363 (6428), 418-423 (1993).
- Gray, M. A., et al. Targeted glycan degradation potentiates the anticancer immune response in vivo. Nature Chemical Biology. 16 (12), 1376-1384 (2020).
- Fernandes, Â, et al. Glycans as shapers of tumour microenvironment: A sweet driver of T-cell-mediated anti-tumour immune response. Immunology. 168 (2), 217-232 (2023).
- Togayachi, A., et al. Polylactosamine on glycoproteins influences basal levels of lymphocyte and macrophage activation. Proceedings of the National Academy of Sciences of the United States of America. 104 (40), 15829-15834 (2007).
- Park, D. D. Resident and elicited murine macrophages differ in expression of their glycomes and glycan-binding proteins. Cell Chemical Biology. 28 (4), 567-582 (2021).
- Steinman, R. M. Dendritic cells and immune-based therapies. Experimental Hematology. 24 (8), 859-862 (1996).
- Sabado, R. L., Bhardwaj, N. Directing dendritic cell immunotherapy towards successful cancer treatment. Immunotherapy. 2 (1), 37-56 (2010).
- Pardoll, D. M. The blockade of immune checkpoints in cancer immunotherapy. Nature Reviews Cancer. 12 (4), 252-264 (2012).
- Steinman, R. M., Banchereau, J. Taking dendritic cells into medicine. Nature. 449 (7161), 419-426 (2007).
- So-Rosillo, R., Small, E. J. Sipuleucel-T (APC8015) for prostate cancer. Expert Review of Anticancer Therapy. 6 (9), 1163-1167 (2006).
- Cheever, M. A., Higano, C. S. PROVENGE (Sipuleucel-T) in prostate cancer: The first FDA-approved therapeutic cancer vaccine. Clinical Cancer Research. 17 (11), 3520-3526 (2011).
- Videira, P. A., et al. Surface α2-3- and α2-6-sialylation of human monocytes and derived dendritic cells and its influence on endocytosis. Glycoconjugate Journal. 25 (3), 259-268 (2008).
- Cabral, M. G., et al. The phagocytic capacity and immunological potency of human dendritic cells is improved by α2,6-sialic acid deficiency. Immunology. 138 (3), 235-245 (2013).
- Silva, Z., et al. MHC class I stability is modulated by cell surface sialylation in human dendritic cells. Pharmaceutics. 12 (3), 249(2020).
- Silva, M., et al. Sialic acid removal from dendritic cells improves antigen cross-presentation and boosts anti-tumor immune responses. Oncotarget. 7 (27), 41053-41066 (2016).
- Crespo, H. J., et al. Effect of sialic acid loss on dendritic cell maturation. Immunology. 128, 621-631 (2009).
- Council of Europe. Guide to the Preparation, Use and Quality Assurance of Blood Components. Council of Europe. , Strasbourg, France. (2017).
- LS Columns. Miltenyi Biotec. , Available from: https://www.miltenyibiotec.com/US-en/products/Is-columns.html#130-042-401 (2012).
- Nair, S., Archer, G. E., Tedder, T. F. Isolation and generation of human dendritic cells. Current Protocols in Immunology. 07, Unit 7.32 (2012).
- Wu, X., Xu, F., Liu, J., Wang, G. Comparative study of dendritic cells matured by using IL-1β, IL-6, TNF-α and prostaglandins E2 for different time span. Experimental and Therapeutic Medicine. 14 (2), 1389-1394 (2017).
- Naeim, F., Nagesh Rao, P., Song, S., Phan, R. Chapter 2 - Principles of Immunophenotyping. Atlas of Hematopathology. , Academic Press. Cambridge, MA. 29-56 (2018).
- Cernadas, M., Lu, J., Watts, G., Brenner, M. B. CD1a expression defines an interleukin-12 producing population of human dendritic cells. Clinical and Experimental Immunology. 155, 523-533 (2009).
- Santambrogio, L., Strominger, J. L. The ins and outs of MHC class II proteins in dendritic cells. Immunity. 25 (6), 857-859 (2006).
- Raposo, C. D., Canelas, A. B., Barros, M. T. Human lectins, their carbohydrate affinities and where to find them. Biomolecules. 11 (2), 188(2021).
- Videira, P. A. Q., et al. Patent WO2017002045. A viable cell population, method for production and uses thereof. Portugal patent. , Universidade NOVA de Lisboa. Lisbon, Portugal. (2017).
- Bai, L., Feuerer, M., Beckhove, P., Umansky, V., Schirrmacher, V. Generation of dendritic cells from human bone marrow mononuclear cells: Advantages for clinical application in comparison to peripheral blood monocyte derived cells. International Journal of Oncology. 20 (2), 247-253 (2002).
- Marques, G. S., Silva, Z., Videira, P. A. Antitumor efficacy of human monocyte-derived dendritic cells: Comparing effects of two monocyte isolation methods. Biological Procedures Online. 20, 4(2018).
- Bax, M., et al. Dendritic cell maturation results in pronounced changes in glycan expression affecting recognition by Siglecs and galectins. Journal of Immunology. 179 (12), 8216-8224 (2007).
- Chinoy, Z. S., Montembault, E., Moremen, K. W., Royou, A., Friscourt, F. Impacting bacterial sialidase activity by incorporating bioorthogonal chemical reporters onto mammalian cell-surface sialosides. ACS Chemical Biology. 16 (11), 2307-2314 (2021).
- Chou, M. -Y., Li, S. -C., Li, Y. -T. Cloning and expression of sialidase L, a NeuAcα2→3Gal-specific sialidase from the leech, Macrobdella decora. Journal of Biological Chemistry. 271 (32), 19219-19224 (1996).
- Crespo, H. J., Lau, J. T. Y., Videira, P. A. Dendritic cells: A spot on sialic acid. Frontiers in Immunology. 4, 491(2013).
- Büll, C. Metabolic sialic acid blockade lowers the activation threshold of moDCs for TLR stimulation. Immunology & Cell Biology. 95 (4), 408-415 (2017).
- Ohmi, Y., et al. Majority of alpha2,6-sialylated glycans in the adult mouse brain exist in O -glycans: SALSA-MS analysis for knockout mice of alpha2,6-sialyltransferase genes. Glycobiology. 31 (5), 557-570 (2021).
- Chung, C., et al. Integrated genome and protein editing swaps α-2,6 sialylation for α-2,3 sialic acid on recombinant antibodies from CHO. Biotechnology Journal. 12 (2), 1600502(2017).
- Hyvärinen, S., Meri, S., Jokiranta, T. S. Disturbed sialic acid recognition on endothelial cells and platelets in complement attack causes atypical hemolytic uremic syndrome. Blood. 127 (22), 2701-2710 (2016).
- Powell, L. D., Whiteheart, S. W., Hart, G. W. Cell surface sialic acid influences tumor cell recognition in the mixed lymphocyte reaction. Journal of Immunology. 139, 262-270 (1987).
- Corfield, A. P., Higa, H., Paulson, J. C., Schauer, R. The specificity of viral and bacterial sialidases for α(2-3)- and α(2-6)-linked sialic acids in glycoproteins. Biochimica et Biophysica Acta - Protein Structure and Molecular Enzymology. 744 (2), 121-126 (1983).
- Tkachenko, N., Wojas, K., Tabarkiewicz, J., Rolinski, J. Generation of dendritic cells from human peripheral blood monocytes - Comparison of different culture media. Folia Histochemica et Cytobiologica. 43, 25-30 (2005).
- Kim, S. J., et al. Human CD141+ dendritic cells generated from adult peripheral blood monocytes. Cytotherapy. 21 (10), 1049-1063 (2019).
- Calmeiro, J., et al. In-depth analysis of the impact of different serum-free media on the production of clinical grade dendritic cells for cancer immunotherapy. Frontiers in Immunology. 11, 593363(2021).
- Stamatos, N. M., et al. LPS-induced cytokine production in human dendritic cells is regulated by sialidase activity. Journal of Leukocyte Biology. 88 (6), 1227-1239 (2010).
- Lehmann, F., Tiralongo, E., Tiralongo, J. Sialic acid-specific lectins: Occurrence, specificity and function. Cellular and Molecular Life Sciences. 63 (12), 1331-1354 (2006).
- Frosch, M., Görgen, I., Boulnois, G. J., Timmis, K. N., Bitter-Suermann, D. NZB mouse system for production of monoclonal antibodies to weak bacterial antigens: Isolation of an IgG antibody to the polysaccharide capsules of Escherichia coli K1 and group B meningococci. Proceedings of the National Academy of Sciences of the United States of America. 82 (4), 1194-1198 (1985).
Nachdrucke und Genehmigungen
Genehmigung beantragen, um den Text oder die Abbildungen dieses JoVE-Artikels zu verwenden
Genehmigung beantragenThis article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. Alle Rechte vorbehalten