
Method Article
에듀 염색에 의한 초파리 성인 난소에서 미토콘드리아 DNA 복제의 제자리 라벨에
요약
Drosophila oogenesis continues to be exceptionally useful in the study of mitochondrial proliferation and inheritance. This manuscript describes a detailed protocol used to label the replicating mitochondrial DNA (mtDNA) in Drosophila adult ovaries with 5-ethynyl-2´-deoxyuridine (EdU), which facilitates uncovering mechanisms associated with mitochondrial inheritance that were previously debatable.
초록
The mitochondrial genome is inherited exclusively through the maternal line. Understanding of how the mitochondrion and its genome are proliferated and transmitted from one generation to the next through the female oocyte is of fundamental importance. Because of the genetic tractability, and the elegant, ordered simplicity by which oocyte development proceeds, Drosophila oogenesis has become an invaluable system for mitochondrial study. An EdU (5-ethynyl-2´-deoxyuridine) labeling method was utilized to detect mitochondrial DNA (mtDNA) replication in Drosophila ovaries. This method is superior to the BrdU (5-bromo-2'-deoxyuridine) labeling method in that it allows for good structural preservation and efficient fluorescent dye penetration of whole-mount tissues.
Here we describe a detailed protocol for labeling replicating mitochondrial DNA in Drosophila adult ovaries with EdU. Some technical solutions are offered to improve the viability of the ovaries, maintain their health during preparation, and ensure high-quality imaging. Visualization of newly synthesized mtDNA in the ovaries not only reveals the striking temporal and spatial pattern of mtDNA replication through oogenesis, but also allows for simple quantification of mtDNA replication under various genetic and pharmacological perturbations.
서문
핵 게놈 외에도, 각 진핵 세포는 미토콘드리아 매트릭스에서 작은 원형 DNA의 사본의 수천을 포함하고 있습니다. 미토콘드리아 DNA (미토콘드리아)은 전자 전달계의 필수 서브 유니트를 인코딩하는 동안, 복제 및 미토콘드리아 DNA의 전사에 대한 모든 요소를 포함하는 미토콘드리아 프로테옴의 대부분은 핵 유전자에 의해 코딩된다. 상속 멘델 법칙을 따른다 핵 게놈 달리, 동물 미토콘드리아 게놈은 모계를 통하여 독점적으로 상속된다. 따라서 미토콘드리아 DNA가 증식과 여성의 난자를 통해 한 세대에서 다음 세대로 전달하는 방법을 이해하는 것이 근본적으로 중요하다. 그러나, 미토콘드리아 DNA 복제가 여성 생식계에서 조절되는 방식을 계속 논의가있다. 또한, 미토콘드리아 전송을 위해 널리 미토콘드리아 병목 이론 원시 생식 세포에서 미토콘드리아의 인구가 비교적 적은 수 두린으로 서브 샘플링되는 것을 제안g 개발 1 2. 또한, 미토콘드리아 DNA 복제 시간적 및 공간적 oogenesis시 규제 것을 의미한다. 따라서, 배아 발달 동안 미토콘드리아 DNA 복제의 시츄 검출 미토콘드리아 유전 메카니즘의 이해를 용이하게한다.
초파리 oogenesis은 미토콘드리아 DNA의 복제 및 전송을 연구하는 유전자 다루기 쉬운 시스템을 제공한다. 두 초파리 난소 각각, 산란율의 기능적 단위 ovarioles 3이라고 달걀 챔버 16-20 독립적 문자열 (도 1a 참조)가있다. 각 ovariole은 앞쪽 끝이 germarium라는 구조로 구성되어있다 oogenesis의 진보적 인 선형 조직이 포함되어 있습니다. germarium 더 뚜렷한 발달 단계에서 배아 세포를 포함하는 4 개의 영역으로 구분된다. 제 1 지역에서, 배아 줄기 세포로 알려진 C 딸 세포를 생성하는 비대칭 분할 통과ystoblasts. 영역 (2a)는 최종 분할을 완료 한 cystoblasts이 포함되어 있습니다. Cystoblasts는 부문 생산 16 셀 그룹의 네 라운드를 받아야. 16 세포는 링 운하라는 세포질 다리로 서로 연결된 상태로 유지됩니다. 다른 15 배수체 간호사 세포로서 개발 중에 만 셀들 중 하나는 난자와 같은 분화 얻어. fusome라고도 필수적인 세포질 구조는, 링 수로의 형성을 촉진 낭종 극성 간호사 난자 세포 상호 작용 4,5-를 결정하는 것이 제안되어있다. 낭종이 영역 (2b)를 향해 이동함에 따라, 낭종 구조는 germarium의 전체 폭에 걸쳐,보다 모낭 세포와 관련된다. germarium 구조는 제 신진 달걀 챔버를 포함하는 영역 (3)으로 끝난다. 그 후, 달걀 챔버 (14) 형태 학적으로 서로 다른 단계에서 ovariole을 통해 진행 germarium의 뒤쪽 끝에서 조립된다. 모세포의 성장 WH 간호사 세포에 의존무형 문화 유산 수송 단백질의 mRNA와 난자에 링 운하를 통해 endomembrane 구조 (예를 들어, 골지). 초파리 oogenesis 동안, 각 16 세포 낭종 내의 미토콘드리아의 일부가 fusome과 관련이 있음을 발견, 링 운하를 통해 이동 Balbiani 바디 (6)라는 큰 질량에 하나의 난자에 전달했다. 이 현상은 여성의 난소 통하여 미토콘드리아 상속의 품질 관리에 기여하는 것이 제안되었다.
미토콘드리아 DNA 합성의 검출은 세포의 DNA에 표지 된 DNA의 전구 물질의 결합에 의존한다. 전통적으로, 티미 딘, 5- 브로 모 -2'- 데 옥시 우리 딘의 뉴 클레오 사이드 유사체 (BrdU의)은 조직 배양 세포에서 미토콘드리아 7,8 복제 라벨을 사용 하였다. 그러나, 항체 - 기초의 BrdU 라벨링 특히 전체 마운트 조직 염색을 위해, 몇 가지 제한을 나타낸다. BrdU의 라벨의 하나의 주요 단점은 노출 DNA 변성을 필요로한다는 것이다BrdU 양성 에피토프이를 방지 BrdU의 항체에 의해 검출 될 수 있도록. 시편은, 시편의 구조를 중단하고 다음 염색 절차 9 복잡 수 화학 물질 (예를 들면, 염산 또는 메탄올과 아세트산의 혼합물)의 DNase로, 열, 또는 분해 등의 가혹한 변성 조건 하에서 처리되어야 10.
여기서, 대안 티미 딘 유사체 5-에 티닐 -2'- 데 옥시 우리 딘 (EDU)은 초파리 성인 난소의 미토콘드리아 DNA 복제에 라벨을 사용한다. 이 방법은 빠르고 매우 민감하다. EDU는 쉽게 DNA 복제시 세포 DNA에 통합됩니다. 다음 검출은 "클릭"반응에 기초하는 구리 (I)은 단말기 알킨 기 및 형광 지드 (10) 사이의 공유 결합 반응을 촉매 반응. 반응이 시험편의 변성이 필요하지 않기 때문에 양호한 구조적 보존을 허용한다. 염료 (A)의 또 사이즈zide는 1/500 전체 실장 조직 빠르고 효과적인 침투를 가능하게하는 항체 분자 (10)의 그 것이다. 우리는 초파리 oogenesis 동안 미토콘드리아 DNA 복제를 감지하는이 방법을 사용하고 복제에 의존하는 미토콘드리아 DNA 선택적 상속 메커니즘을 제안하는 우리를 인도 초파리의 난소 (11)의 germarium 지역에서 눈에 띄는 공간 패턴을 발견했다. 우리는 여기에서 초파리의 난소에서 미토콘드리아 DNA 복제의 듀 표시에 대한 자세한 프로토콜을 제시한다. (: 너도 알면서 T300I 산) (11)뿐만 아니라 잠재적 인 미토콘드리아 막을 분산하고 잠재적으로 미토콘드리아 DNA 복제를 방해 미토콘드리아 uncouplers, 다양한 치료 프로토콜의 응용 프로그램을 설명하기 위해, 우리는 또한 미토콘드리아 DNA a를 미토콘드리아 DNA 돌연변이 초파리에서 복제를 테스트했다.
프로토콜
1. 조직 수집 및 해부
- 건조 효모를 포함하는 각각의 유리 병에서, 문화 (10) 성인 여성 2 ~ 3 일 동안 10 명의 남성으로 날아갑니다.
참고 : 잘 먹인 여성 파리를 유지하는 것은 전반적인 품질과 난소의 생산 수율을 향상시키고 박리를 촉진 할 것이다. - 여성은 이산화탄소 (CO 2) 패드 플라이에 날아 마취.
- 입체경에서 RT 매체 해부 패드 (10 % 소 태아 혈청 (FBS)으로 보충 슈나이더의 초파리 배지) 몇 방울을 놓는다. 살아 있고 건강한 조직을 유지하기 위해 매체의 모든 해부 절차를 실시한다.
- 살찐 여성이 낮은 흉부에서 날카로운 미세 코 집게 비행 잡아. 복부에 조직이 노출 될 때까지 파리의 극단적 인 후방에서 부드럽게 예인선 집게의 또 다른 세트를 사용합니다. 다른 조직 (예를 들어 내장)에서 두 개의 난소를 분리합니다.
참고 : 각 난소 표시 작성이다 불투명 한 구조를16 ~ 20 부착 ovarioles의 라. - 시약의 침투를 강화 떨어져을 당겨 몇 번 ovariole 각각의 사이에 포셉의 팁을 전달하여 난소를 엽니 다. 다음 절차 중 조직의 손실을 최소화하기 위해 난소 내에서 연결된 ovarioles를 유지합니다.
- 10 % FBS와 슈나이더의 초파리 배지 500 μl를 함유하는 1.5 ml의 마이크로 원심 튜브에 즉시 전송 난소.
- 해부를 반복하고 각 microcentrifuge 관에서 10 ~ 15 난소를 수집합니다.
2. 에듀 라벨
- 각 튜브의 매체를 대기음, 7 μM의 aphidicolin을 포함하는 10 % FBS와 슈나이더의 초파리 매체의 500 μL로 교체합니다.
주 : Aphidicolin는 미토콘드리아 DNA의 복제 (7), (12)에 영향을주지 않으면 서 DNA 폴리머 라제 α의 억제 핵 DNA 합성을 차단하는 데 사용된다. aphidicolin 농도까지 증가시킬 수있다70 μM 더 나은 억제를 달성했다. DMSO에 3-30 밀리미터 aphidicolin의 원액 최대 6 주 동안 -20 ℃에서 어둠 속에서 저장 될 수있다. - 매체를 변경하는 동안 그들이 솔루션에서 포장되어 있는지 확인 건강한 난소를 유지합니다. 단계 2.6을 통해 10 % FBS와 슈나이더의 초파리 매체를 사용합니다.
- 부드러운 회전 벤치 탑 로커에 실온에서 3 시간 동안 난소를 품어.
- 약물 치료, 예를 들어 미토콘드리아 uncoupler 보닐 시안화 -4- 트리 플루오 로메 톡시 페닐 (FCCP) 들어 aphidicolin 처리 2 시간 후 배지에 약물을 적절한 농도 (예를 들면, 10 μM FCCP)를 추가한다. 또 다른 1 시간 동안 배양을 계속합니다.
- 또는 약물없이 매체를 포함 aphidicolin를 제거합니다. 간단히 두 번 매체와 난소를 (aphidicolin이 필요하지 않습니다) 헹군다.
- 1 ml의 매체를 포함하는 10 μM 에듀 7 μM의 aphidicolin를 추가하고 incubatin 계속2 시간 동안 실온에서 g. -20 ° C에서 (DMSO)에 10 mM의 듀 원액을 저장합니다.
- 에듀과 aphidicolin를 포함하는 매체를 제거합니다. 3 분 각각에 대해 두 번 (aphidicolin없이) 매체를 씻으십시오.
3. 조직 고정 및 Permeabilization
- 인산 완충 식염수 (PBS, pH 7.4의) 4 % 파라 포름 알데히드 용액을 제조 하였다.
참고 : 사전 득점 앰플에 저장 상용 가능한 파라 포름 알데히드를 사용했다. 새로운 앰플을 열고 사용하기 전에 4 %로 희석.
주의 : 포름 알데히드는 독성이; 그것은 피부와 눈 보호 흄 후드에서 처리되어야한다. - 부드러운 회전 실온에서 20 분 동안 4 % 파라 포름 알데히드와 난소를 수정합니다.
- 정착액을 제거하고 5 분마다 시간 동안 PBS에서 3 % BSA 1 ㎖에 두 번 난소을 씻는다.
- 조직을 Permeabilize 하시려면 위해, 세척 용액을 제거하고 PBS에서 0.5 % 트리톤 X-100 1 ㎖를 추가한다. 실온에서 20 분 동안 인큐베이션.
- permeabilization 솔루션을 제거하고 1 ㎖에 두 번 세척PBS에서 3 % BSA의.
4. 에듀 검출
주 : 듀 검출 "클릭"화학을 기반으로하는 구리 (I)는 EDU의 말단 알킨 그룹 형광 지드 추가 [3 + 2] 고리 화 (13), 촉매 반응 및 형광 분자는 후속 검출을 실시한다. 쉽게 비 촉매의 Cu (II) 종에 산화 구리 (I), 이후, 반응을 촉매하는 데 필요하며, 이는 구리 (I)을 얻었다 반응계에서의 Cu (II), 황산 줄이는 것을 권장한다. 즉, 구리 (II), 황산은 아스코르브 산 (본원에서 "첨가물 버퍼")를 생성하기 위해 구리 (I) 등의 환원제의 존재 하에서 사용된다.
- 실험 전에 (알렉사 플 루어 488 아 지드, 알렉사 플 루어 "염료 아 지드"이하로 명명 된 바람직한 형광에 따라 555 아 지드 또는 다른 사람), 에듀 반응 버퍼와 에듀 버퍼 첨가제 알렉사 플 루어 아 지드의 작업 용액을 제조 제조업체에 따라명령.
- 70 ㎕의 DMSO에 염료 지드를 재구성한다. 최대 1 년 -20 ° C에서 작동하는 솔루션을 저장합니다.
- 탈 이온수로 10 배 에듀 반응 버퍼를 희석하여 신선한 에듀 반응 버퍼를 준비합니다.
참고 : 사용 후 4 ° C에서 나머지 1 배 솔루션을 저장합니다. 1 배 솔루션은 최대 6 개월 동안 안정하다. - 완전 탈 이온수 2 ㎖ 중의 분말을 용해하여 듀 버퍼 첨가제의 10 배 스톡 용액을 만든다.
참고 :이 원액 1 년까지에서 -20 ° C에 대한 안정적입니다. 상기 용액이 갈색을 개발하면이 저하하고 폐기되어야한다.
- 신선한마다 EDU 반응 칵테일을 준비하고, 준비 15 분 이내에 사용한다. 재현 가능한 결과를 달성하기 위해, 반응 성분이 같은 비율을 유지하고 있는지 확인.
- 신선한 1 배 EDU는 탈 이온수 (단계 4.1.3에서 제조)를 10 배 원액 1:10 희석 첨가제 솔루션을 버퍼링합니다. 이 S 사용같은 날 olution.
- 하기의 성분을 조합하여 듀 반응 칵테일 1 ml의 준비 : 860 μL 1X 듀 반응 완충액 40 μL에 CuSO 4 2.5 μL 염료 지드 100 ㎕의 1X 듀 버퍼 첨가제.
주 : 재료가 최적의 성능을 보장하기 위해 추가되는 것이 중요하다.
- 3 % BSA와 PBS를 제거하고 각 튜브에 EDU 반응 칵테일의 0.5 ml를 추가합니다. 부드러운 회전을 실온에서 30 분 동안 인큐베이션. 빛으로부터 보호 샘플을 유지합니다.
- 에듀 반응 칵테일을 제거하고 PBS에서 3 % BSA 1 ㎖로 한 번 씻는다.
- 1 ml의 PBS로 한번 샘플을 세척한다.
5. 항체 라벨링
참고 : 에듀 염색 후 항체 라벨링을 수행합니다. 추가 염색이 요구되지 않는 경우, 하나는 직접 장착 및 영상을 진행할 수 있습니다. 모든 샘플들이 다음 절차에서 빛으로부터 보호하는 것이 중요하다.
- 세척 용액을 제거합니다. 30 분 동안 PBS에서 0.2 % BSA, 0.1 % 트리톤 X-100을 함유하는 차단 용액에 1 ml의 난소를 차단.
- 차단 솔루션을 제거하고 차단 솔루션에 희석 차 항체 (항체 α 예를 들어, 마우스 ATP 합성 효소 서브 유닛, 1 : 1,000 희석)로 교체합니다. 4 ° CO / N에서 어둠 속에서 품어.
- 솔루션 3 회, 10 분마다 차단의 1 ml의 샘플을 씻으십시오. 백그라운드를 최소화하기 위해, 30 분마다 두 번 더 세척 하였다. 차단 솔루션을 제거합니다.
- 실온에서 2 시간 동안 : (200 희석 예를 들어, 염소 항 - 마우스 알렉사 플 루어 568 차 항체, 1) 차단 솔루션에 희석 차 항체와 함께 품어.
주 : 듀에 연결된 하나 이상의 다른 색상을 사용한다. - 단계 5.3에서 수행되는 세척 단계를 반복합니다.
- 1 ml의 PBS로 씻어 한번 세제를 제거합니다.
6. 장착 및 이미지
- 조심스럽게 모든 PBS와 IMM 제거ediately 50 μL가 (또는 DAPI없이) 매체를 장착와 난소를 다룹니다.
참고 : ovarioles가 서로 분리되는 동안 강화하지 않는 장착 매체를 사용합니다. 난소 주일까지 4 ℃에서 장착 가능 매체에 저장 될 수있다. - 피펫 팁의 단부를 절단하고, 현미경 슬라이드 상에 조심스럽게 난소 전송할 사용.
- 실체 현미경 아래에서 날카로운 미세 코 집게와 완전히 분리 된 각각의 ovariole. 후방 끝 단계 14 성숙 계란 챔버에 연결 조직을 제거합니다. 젊은 계란 챔버 앞쪽 끝과 투명에 있습니다.
- 이들이 서로 겹치지 않도록 집게 계란 챔버 정렬.
- 천천히 샘플의 상단에 # 1.5 유리 커버 슬립을 낮 춥니 다. 설치 매체는 실온에서 몇 시간 동안 중합 할 수 있습니다. 투명 매니큐어로 밀봉합니다.
- 이미지는 촬영에 앞서 4 ° C에서 다음 날, 또는 저장소를 슬라이드.
- 공 초점 마일에서 시각화63X 기름 침지 목적으로 croscope. 입체 Z - 스택 이미지를 캡처합니다.
참고 : EDU는 상피 칼집에서 나중에 단계 달걀 챔버 및 배경 형광의 신호 강한이 있습니다. 특정 germarium에 EDU 라벨을 조사 할 때, 하나에 의해 중복 또는 다른 조직 또는 그 이후의 단계 달걀 챔버에 가까운 germariums을 피해야한다.
결과
위의 프로토콜은 초파리 oogenesis 동안 미토콘드리아 DNA 복제를 나타내는 미토콘드리아 (그림 1B-C)과 관련된 반점 구조의 시각화 할 수 있습니다. ATP 합성 효소 알파 서브 유닛 (그림 2)에 대한 염색하여 표시 미토콘드리아와 지역화 듀 puncta에. 관찰 된 신호는 검증, 에티 디움 브로마이드 (11), 미토콘드리아 DNA 복제 (14)에 대한 억제제로 치료 난소에 존재했다 이들 puncta의 mtDNA.Aphidicolin는 미토콘드리아 DNA 복제 (그림 2)에 영향을주지 않고 핵 DNA 염색을 억제하는 데 사용 된 복제 참으로 레이블입니다. aphidicolin 치료하지 않으면, 강렬한 에듀 신호는 핵 라벨 및 미토콘드리아 DNA의 puncta의 거의 (그림 3) 검출되었다. 그러나 aphidicolin의 존재하에 핵의 혼입을 대폭 감소시키고, 미토콘드리아와 관련된 많은 puncta에 관찰되었다.
이후 germarium 계란 챔버 (그림 1B)의 미토콘드리아 DNA 복제의 높은 수준이 있습니다. 그러나, 특히, 미토콘드리아 DNA 복제는 germarium의 공간 패턴을 표시. 듀 puncta의의 숫자로 나타낸 바와 같이, 거기에 germarium의 영역 (1)의 미토콘드리아 DNA 복제의 중간 수준이지만, 거의 영역 (2a)에서 더 듀 통합 (그림 1C). 낭종이 germarium에서 영역 (2b)까지 아래로 이동함에 따라, 미토콘드리아 DNA 복제를 재개 영역 (2b)의 후방 낭종에 듀 puncta의 개수는 영역 (2a) (도 1C)에서보다 훨씬 높았다. 후 리 타이 샤오 (HTS) 단백질에 의해 스테인드 구체적으로, 집중적 인 듀 통합은 링 운하와 fusome 구조 주위에 집중했다. 미토콘드리아는 germarium의 영역 (3)에 높은 수준으로 복제 보관하고 (도 1C)
과 특이의 관계를 설명하기 위해IC 유전자 또는 미토콘드리아 증식 치료는 초파리 난소 유전자 조작 또는 약품 처리를 실시 할 수있다. 우리는 FCCP, 미토콘드리아 막 잠재력을 발산 고전 미토콘드리아 protonophore, 서로 다른 농도의 난소를 처리 하였다. 대조군으로서, DMSO는 미토콘드리아 DNA의 복제 (도 4a)에 아무런 영향을 미치지 않았다. FCCP의 높은 용량 (10 μM)는 거의 완전히 germarium (그림 4D)를 통해 미토콘드리아 DNA 복제를 고갈. 그럼에도 불구하고, FCCP (2 또는 5 μM)의 낮은 농도는 지역 2B, 3 미토콘드리아 중단에 더 민감 제안, 영역 (b) 및 (3) (그림 4B-C)의 영역 1에서 미토콘드리아 DNA 복제에 약간의 영향 만 억제 복제를 가지고, 또는 그들은 상대적으로 느린 복제 속도론을 유지한다. 상기 결과는 미토콘드리아 DNA의 복제 미토콘드리아 활성과 관련된 것으로 나타났다. 특히, germarium의 다른 지역은 미토콘드리아 꼬마 도깨비에 다르게 반응airment.

그림 1. 미토콘드리아 DNA의 초파리 oogenesis 동안 복제. 초파리 ovariole의 (A) 다이어그램 및 germarium의 확대 전망. 계란 챔버의 연속적인 발달 단계를 후부, 오른쪽, 앞쪽 왼쪽에서 ovariole을 보여줍니다. germarium에서 fusome (적색), 배아 줄기 세포 (파란색), 미토콘드리아 (녹색), (위치, fusome 구조 미토콘드리아 클러스터 인식 파선) 미래 난자, 낭종 (복숭아) 네 발달 영역 현상은 도시 . (B) HTS-RC, 링 운하와 fusome 마커에 대한 야생형 듀에 의해 표시 ovariole과 항체의 대표 Z - 스택 투영합니다. aphidicolin의 존재에서, EDU는 미토콘드리아 DNA (화살촉) 및 핵 (화살표)로 통합되었다. 스케일 바, 10 μm의.(C) B의 박스 영역에 설명 germaria의 확대 전망. 네 가지 발달 영역이 표시됩니다. 스케일 바, 5 μm의 11. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
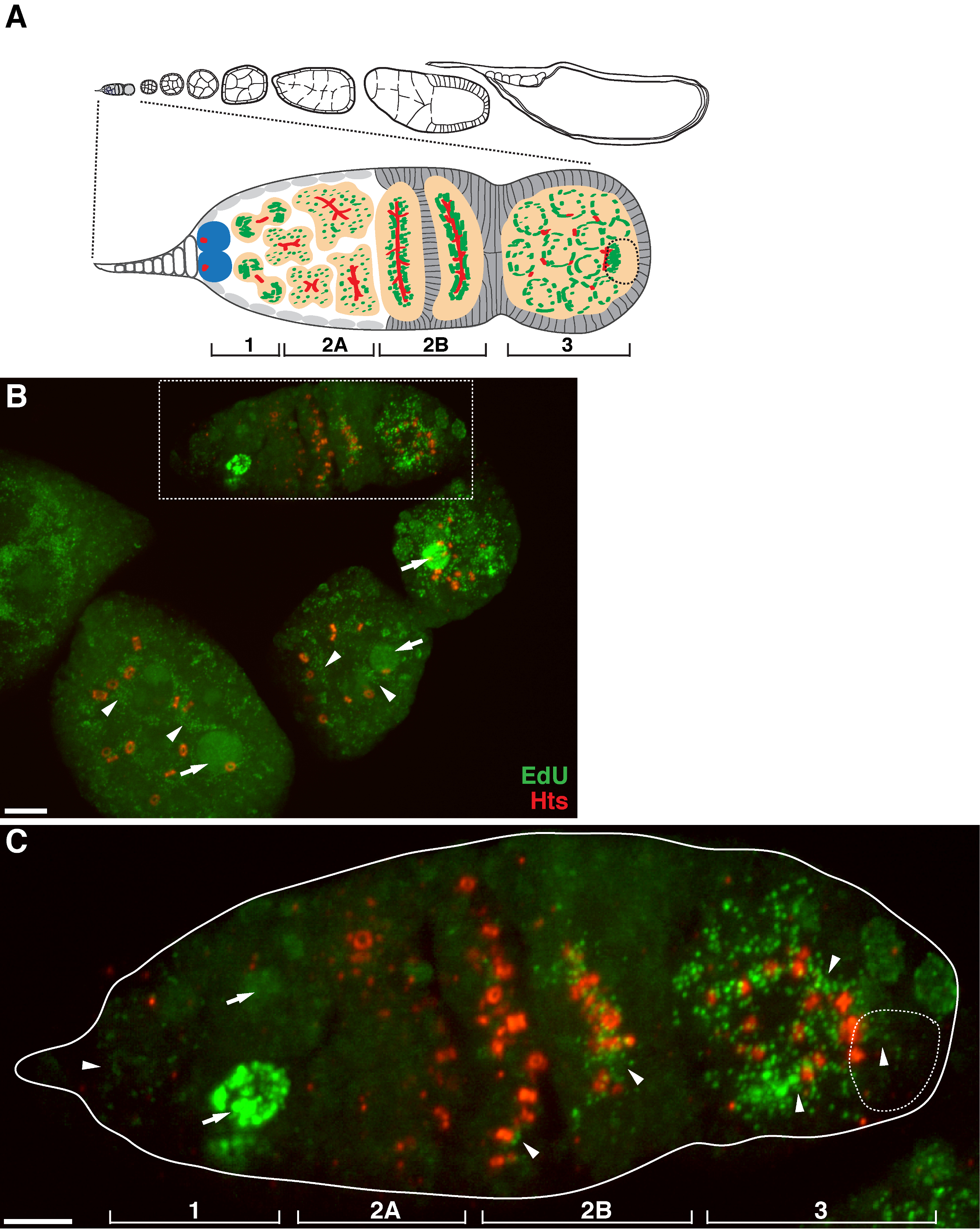{kind=link}

aphidicolin 치료와 듀 통합 시각화 초파리 germarium 그림 2. 미토콘드리아 DNA 복제 (A) -. (D) 에듀의 결합을 나타내는 야생형 germarium 대표 공 촛점 섹션 (녹색, B), 미토콘드리아, ATP 합성 효소 알파 서브 유닛 염색으로 표시 DNA 중합 효소-α 억제제 aphidicolin와 사전 배양과 DAPI 염색 (파란색, D)으로 표시 (적색, C) 및 핵. (A &# 39 ;-D ')의 박스 영역의 확대 된 이미지 (A) 듀 통합 (B을 보여주는'), 미토콘드리아 (C ') 및 핵 (D'). aphidicolin의 존재하에 핵 도입 (화살표)은 감소되고, 많은 puncta의 미토콘드리아 (화살촉) 내에 국부적 하였다. 스케일 바, 10 μm의. 그림 (11)에서 수정되었습니다. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
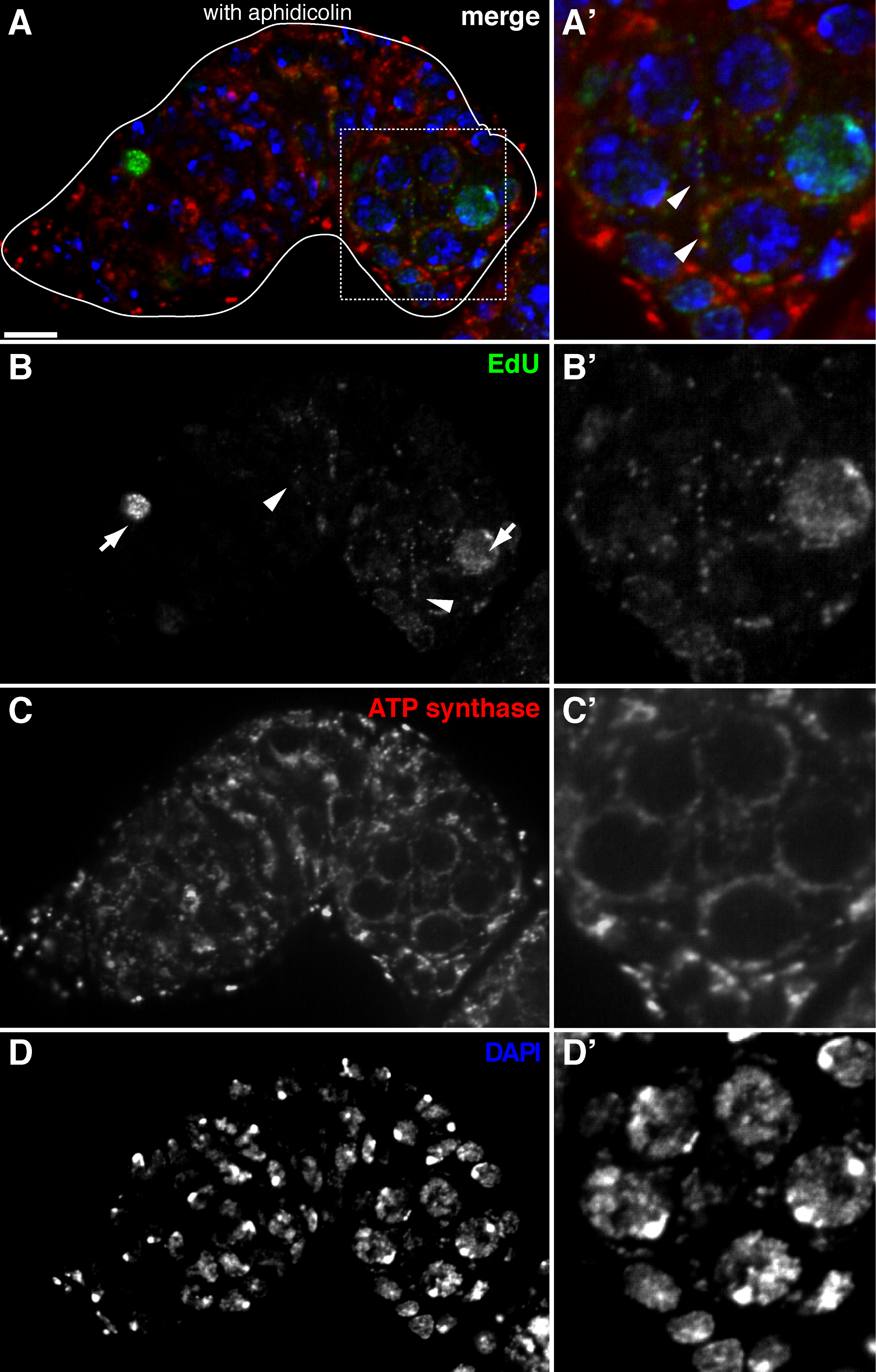{kind=link}

도 3 미토콘드리아 DNA 복제는 거의 aphidicolin 처리없이 초파리 germarium에서 검출 (A) -. (D) 듀 혼입 (녹색)에 나타낸 야생형 germarium 대표 촛점 부는 미토콘드리아는로 표시DNA를 중합 효소-α 억제제 aphidicolin의 부재 DAPI 염색 (청색)로 표지 ATP 합성 효소 알파 서브 유닛 염색 (적색) 및 핵. (A'-D ') 확대 된 (A)의 박스 영역의 이미지 듀 통합 (B을 보여주는'), 미토콘드리아 (C ') 및 핵 (D'). aphidicolin없이 강렬한 에듀 신호 라벨 핵 (화살표)과 미토콘드리아 DNA의 puncta의 거의 검출되었다. 스케일 바, 10 μm의. 그림 (11)에서 수정되었습니다. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
{kind=link}

도 4 미토콘드리아 uncoupler는 미토콘드리아 DNA의 복제를 저해. 야생형 germarium를 나타내는 대표 Z 스택 프로젝트 DMSO 처리(A) 또는 제 2 μM (B), 5 μM (C), 듀 혼입 중에 10 μM (D)의 농도에서 미토콘드리아 uncoupler FCCP. 2 μM FCCP (B)로 처리 영역 (b)의 손상 에듀 라벨을 참고하고, 지역 2B, 5 μM의 FCCP 치료 (3) 모두 (상자에 설명). 네 가지 발달 영역이 표시됩니다. 화살표, 핵 DNA; 화살촉, 미토콘드리아 DNA. 스케일 바, 10 μm의. 그림 (11)에서 수정되었습니다. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
{kind=link}
토론
EDU 라벨링 신규 합성 DNA에 혼입 및 뉴 클레오 시드 유사체의 염색에 기반 증식 세포에서 DNA 합성을 검출하기위한 신규하고 효과적인 방법이다. 이 방법은 빠르고 매우 민감한 점에서의 BrdU 라벨링 방법보다 우수하다. 더욱 중요한 것은 10. 역사적 BrdU의 라벨링에 우수한 대안으로, 듀 표시는의 S-단계에서 핵 DNA 복제를 연구에 사용 된, 조직 (9)를 장착 전체적으로 양호한 구조적 보전 효율적 듀 염색 침투를 허용 세포주기. Aphidicolin는 S 단계 12-15에서 핵 DNA 복제의 주요 효소 인 DNA 폴리머 라제 α의 억제제이다. 미토콘드리아 DNA의 복제 처리 aphidicolin 둔감하다 DNA 폴리머 라제 γ에 의해 수행된다. 따라서, 이전에와 듀 배양 동안 aphidicolin 치료는 크게 핵 DNA에 듀 설립을 억제 하였다. 그것은해야적절하게 저장, 핵 DNA 복제를 억제 aphidicolin의 효능은 우리의 손에 변수 인 경우 aphidicolin 몇 주 동안 안정 수도있다. 추가 데이터 분석에서 강한 핵 DNA 표지와 ovarioles 또는 달걀 챔버는 제외한다.
미토콘드리아 DNA의 복제를 용이하게 또한 세포질 총 부피로 정규화 듀 puncta의 수를 카운트함으로써 미토콘드리아 DNA 복제의 수준을 정량화하기위한 간단한 방법을 수득 세포질에서 puncta에 같이 시각화 될 수있다. 이미징 소프트웨어는 자동으로 큰 데이터 세트의 계산 분석에 특히 유용 현미경 이미지에서 듀 puncta의 식별에 적용될 수있다. 핵 DNA 복제의 불완전 억제 염색체에 별개의 궤적에서 듀 통합으로 이어질 및 핵 내부 puncta에로 표시 할 수 있기 때문에,주의 사항,주의해야한다. 또한 다른 경우, 에듀의 강도는 repli에 혼입배경과 소음이 높은이 될 수있는 동안 상호간의 미토콘드리아 DNA는 약한 될 수 있습니다. 따라서, 자동 이미지 분석에 대한 개별 매개 변수는 신중하게 정의되어야한다. 또한 이미지가 적절한 듀 puncta의 식별되어 있는지 확인하기 위해 훈련 된 눈으로 검사하는 것이 좋습니다.
초파리 oogenesis 동안 새로 합성 된 미토콘드리아의 시각화 미토콘드리아 DNA 복제가 약리 유전 또는 섭동 다양한 실시 파리의 실험을 수행하여, 생리적 또는 병리 적 상태에 의해 규제되는 방법을 조사 할 기회를 제공한다. 너도 알면서 T300I 11 : 이전의 연구에서, EDU의 통합 분석은 미토콘드리아 DNA 돌연변이 초파리 산에서 수행되었다. 또한, 미토콘드리아 막 잠재력을 방해하기 위해, 난소은 미토콘드리아 uncoupler FCCP 또는 2,4- 디 니트로 페놀 (DNP) 이전 에듀에 통합의 다양한 농도로 처리 하였다. 실험 목적에 따라약물의 특성은 다른 방법이 효과적인 전달을 위해 채택 될 수 있습니다. 성인 초파리를 들어, 약물이 증기 (예를 들면, 에탄올 및 코카인)로 표현 될 수있다 (16, 17) 또는 약물이 신속하게 본체 (18)에 걸쳐 확산되어 복부에 주입 될 수있다. 가장 일반적인 방법은 약물 즉석 식품 또는 수크로오스 / 약물로 포화 된 여과지에 첨가된다는 것이다. 예를 들어, 미세 소관 조립체 콜히친의 억제제, 난소 절제술 19 전 2-3 일 동안은 파리에 공급 하였다. 따라서, 상기 약물 전달 방법을 평가하고 적절한 농도를 선택하는 것이 중요하다.
초파리의 난소에서 미토콘드리아 DNA 복제의 성공 이미징을 보장하기 위해 몇 가지 중요한 단계는 신중하게 실행해야합니다. 제일, 해부 및 에듀 통합 동안 난소의 생존과 건강을 유지하는 것은 필수적이다 (1 단계와 2 단계). FBS와 슈나이더의 초파리 배지 RT로 가온 할 필요사용하기 전에. 하나는 계란 챔버 및 해부 도구 또는 피펫 팁 사이의 직접 접촉을 최소화해야한다. 난소는 탈수를 방지하기 위해 솔루션에서 모든 시간을 몰입해야한다. 조직이 쉽게 발생할 수 있습니다 잘못 취급하면 희미하거나 더 형광 신호합니다. 조직 마운팅 공정 중에 ovarioles들은 서로 분리되어야하며, 현미경 슬라이드에 퍼져. 계란 챔버가 서로 위에 쌓인 않았는지 확인하십시오. 우리는 단 14 이상으로 계란 챔버가 작은 듀 통합을 표시 것으로 나타났습니다. 또한, 때문에 큰 크기의 후반 단계 달걀 챔버는 종종 이웃 젊은 달걀 챔버의 초점이 맞지 원인. 따라서 후반 단계 달걀 챔버 폐기하는 것이 좋습니다.
여기에서 우리는 초파리 성인 난소에서 미토콘드리아 DNA를 복제 표시에 대한 자세한 프로토콜을 제공합니다. 이 방법은 다양한 유전 적 및 약리학 섭동하에 미토콘드리아 DNA 복제의 간단한 정량 허용발달 미토콘드리아 생물 및 미토콘드리아 DNA 상속을 기본 메커니즘을 해부하는 데 유용 할 것입니다.
공개
The authors have nothing to disclose.
감사의 말
We thank K. Delaney for comments on the manuscript. This work was supported by the National, Heart, Lung, and Blood Institute (NHLBI) Intramural Program.
자료
| Name | Company | Catalog Number | Comments |
| Schneider’s Drosophila medium | Invitrogen | 21720-024 | |
| FBS | Invitrogen | 10100-147 | |
| Pair of Dumont #5 Forceps | Fine Science Tools | 11252-20 | |
| Aphidicolin | Sigma | A0781 | Aliquot after dissolving in DMSO. Avoid repetitive thawing and freezing. Protect from light. May be used within 6 weeks after dissolving. |
| FCCP | Sigma | C2920 | |
| DMSO | Sigma | D2650 | |
| Paraformaldehyde, 16% EM grade | Electron Microscopy Sciences | 15710 | Formaldehyde is toxic; it should be handled in a fume hood with skin and eye protection. |
| PBS | KD Medical | RGF-3190 | |
| BSA | Sigma | A7030 | |
| Triton X-100 | Sigma | T9284 | |
| Click-iT EdU Alexa Fluor 488 Imaging Kit | Invitrogen | C10337 | EdU, CuSO4, Alexa Fluor 488 azide, EdU reaction buffer and Edu buffer additive are included |
| Mouse ATP synthase subunit α antibody, (15H4C4) | MitoSciences | Ab14748 | 1:1,000 dilution |
| Mouse Hts antibody (clone RC) | Developmental Studies Hybridoma Bank (DSHB) | hts RC | 1:1,000 dilution |
| Goat anti-mouse Alexa Fluor 568 secondary antibody | Invitrogen | A-11004 | 1:200 dilution |
| Vectashield mounting medium with DAPI | Vector Laboratories | H-1500 | |
| Glass coverslips, #1.5 22 mm x 22 mm | Fisher Scientific | 12-541-B | |
| Microscope slide | Fisher Scientific | 22-038-103 | |
| Nail polish | Elf | Many of the pigments used in nail polishes are fluorescent and leach into specimens. Only clear nail polish should be used. |
참고문헌
- Taylor, R. W., Turnbull, D. M. Mitochondrial DNA mutations in human disease. Nat Rev Genet. 6 (5), 389-402 (2005).
- Wallace, D. C., Chalkia, D. Mitochondrial DNA genetics and the heteroplasmy conundrum in evolution and disease. Cold Spring Harb Perspect Biol. 5 (11), a021220(2013).
- Spradling, A. C. The development of Drosophila melanogaster. , Cold Spring Harbor Lab. Press. (1993).
- Riechmann, V., Ephrussi, A. Axis formation during Drosophila oogenesis. Curr Opin Genet Dev. 11 (4), 374-383 (2001).
- Lin, H., Yue, L., Spradling, A. C. The Drosophila fusome, a germline-specific organelle, contains membrane skeletal proteins and functions in cyst formation. Development. 120 (4), 947-956 (1994).
- Cox, R. T., Spradling, A. C. A Balbiani body and the fusome mediate mitochondrial inheritance during Drosophila oogenesis. Development. 130 (8), 1579-1590 (2003).
- Davis, A. F., Clayton, D. A. In situ localization of mitochondrial DNA replication in intact mammalian cells. J Cell Biol. 135 (4), 883-893 (1996).
- Iborra, F. J., Kimura, H., Cook, P. R. The functional organization of mitochondrial genomes in human cells. BMC Biol. 2, (2004).
- Rakic, P. Neurogenesis in adult primate neocortex: an evaluation of the evidence. Nat Rev Neurosci. 3 (1), 65-71 (2002).
- Salic, A., Mitchison, T. J. A chemical method for fast and sensitive detection of DNA synthesis in vivo. Proc Natl Acad Sci U S A. 105 (7), 2415-2420 (2008).
- Hill, J. H., Chen, Z., Xu, H. Selective propagation of functional mitochondrial DNA during oogenesis restricts the transmission of a deleterious mitochondrial variant. Nat Genet. 46 (4), 389-392 (2014).
- Lentz, S. I., et al. Mitochondrial DNA (mtDNA) Biogenesis: Visualization and Duel Incorporation of BrdU and EdU Into Newly Synthesized mtDNA In Vitro. Journal of Histochemistry & Cytochemistry. 58 (2), 207-218 (2010).
- Tornoe, C. W., Christensen, C., Meldal, M. Peptidotriazoles on solid phase: [1,2,3]-triazoles by regiospecific copper(i)-catalyzed 1,3-dipolar cycloadditions of terminal alkynes to azides. J Org Chem. 67 (9), 3057-3064 (2002).
- Horwitz, H. B., Holt, C. E. Specific inhibition by ethidium bromide of mitochondrial DNA synthesis in physarum polycephalum. J. Cell Biol. 49, 546-553 (1971).
- Huberman, J. A. New views of the biochemistry of eucaryotic DNA replication revealed by aphidicolin, an unusual inhibitor of DNA polymerase alpha. Cell. 23 (3), 647-648 (1981).
- McClung, C., Hirsh, J. Stereotypic behavioral responses to free-base cocaine and the development of behavioral sensitization in Drosophila. Curr Biol. 8 (2), 109-112 (1998).
- Moore, M. S., et al. Ethanol intoxication in Drosophila: Genetic and pharmacological evidence for regulation by the cAMP signaling pathway. Cell. 93 (6), 997-1007 (1998).
- Dzitoyeva, S., Dimitrijevic, N., Manev, H. Gamma-aminobutyric acid B receptor 1 mediates behavior-impairing actions of alcohol in Drosophila: adult RNA interference and pharmacological evidence. Proc Natl Acad Sci U S A. 100 (9), 5485-5490 (2003).
- Koch, E. A., Spitzer, R. H. Multiple effects of colchicine on oogenesis in Drosophila: induced sterility and switch of potential oocyte to nurse-cell developmental pathway. Cell Tissue Res. 228 (1), 21-32 (1983).
재인쇄 및 허가
JoVE'article의 텍스트 или 그림을 다시 사용하시려면 허가 살펴보기
허가 살펴보기This article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. 판권 소유