
Method Article
필라멘트 아콤유세테 헌티엘라 오만시스에서 CRISPR-Cas9 중재 게놈 편집
요약
CRISPR-Cas9 게놈 편집 시스템은 모델 및 비모델 종에서 사용되어 온 사용하기 쉬운 게놈 편집기입니다. 여기서 우리는 비 모델 필라멘트 아스피세테 곰팡이의 짝짓기 유전자에 조기 정지 코동을 도입하는 데 사용 된이 시스템의 단백질 기반 버전을 제시합니다.
초록
CRISPR-Cas9 게놈 편집 시스템은 모델 및 비모델 종의 게놈에 정밀한 변화를 도입하는 데 사용할 수 있는 분자 도구입니다. 이 기술은 유전자 녹아웃및 녹틴에서 표적 위치에 몇 가지 뉴클레오티드의 도입과 같은 보다 구체적인 변화에 이르기까지 다양한 게놈 편집 접근법에 사용될 수 있다. 게놈 편집은 유전자의 부분적인 기능적 특성화, 형질전환 유기체의 생산 및 진단 도구의 개발을 포함하여 다양한 응용 분야에 사용될 수 있다. 이전에 사용 가능한 유전자 편집 전략에 비해 CRISPR-Cas9 시스템은 새로운 종에서 쉽게 확립할 수 있으며 높은 효율성과 특이성을 자랑하는 것으로 나타났습니다. 이에 대한 주된 이유는 편집 도구가 RNA 분자를 사용하여 유전자 또는 관심서열을 표적으로 하여 표준 기본 페어링 규칙을 악용할 수 있다는 점을 감안할 때 표적 분자 설계를 간단하게 만들기 때문입니다. 다른 게놈 편집 시스템과 마찬가지로 CRISPR-Cas9 기반 방법은 또한 RNA 및 DNA 분자의 설계를 위한 양질의 서열 데이터에 대한 효율적이고 효과적인 변환 프로토콜뿐만 아니라 접근이 필요합니다. 2013년 이 시스템이 도입된 이래, 사카로미세스 세레비시아, 아라비도시스 탈리아나, 드로소필라 멜라노가스터, 머뮤스 무큘러스등 다양한 모델 종을 유전적으로 설계하는 데 사용되어 왔다. 그 후, 비 모델 종에 일하는 연구원은 시스템을 활용하고 곰팡이에 있는 이차 물질 대사, 선충 성장 및 식물에 있는 질병 저항과 같은 다양한 프로세스에 관련시킨 유전자의 연구 결과에 이용했습니다, 많은 그 외 중. 아래에 상세한 이 프로토콜은 헌티엘라 오만시스의성적 주기에 관여하는 유전자의 잘림에 대한 CRISPR-Cas9 게놈 편집 프로토콜의 사용을 설명합니다, 세라토시스티드아세아에 속하는 필라멘트 아스피세테 곰팡이.
서문
고품질, 완전히 조립된 게놈 및 전사의 가용성이 증가함에 따라 생물1의배열에서 다양한 생물학적 과정을 연구하는 능력이 크게 향상되었습니다. 이것은 두 모델 종뿐만 아니라 비 모델 종의 사실이다, 많은 생물학적 과정의 더 다양한 이해를 제공 할 수 있습니다. 이러한 종류의 데이터는 유전자 발견, 전사 네트워크의 식별 및 전체 게놈 및 전사 비교에 사용될 수 있으며, 각각은 자체 응용 분야와 함께 제공됩니다. 그러나 유전자가 예측되고 있는 동안, 화신과 결코 전에 본 적이 없는 비율로 다른 기능성 통로에, 이 유전자의 기능적인 특성은 뒤에 남아, 많은 종에 사용할 수 있는 분자 공구 키트에 의해 제한. 이는 특히 유전체 데이터가 비교적 생성이 쉽지만 추가 분자 특성화가 거의 불가능한비모델 종에대한 경우1,2.
곰팡이 종의 생물학에 중요한 특정 유전자의 기능의 부분적인 특성화는 돌연변이 균주의 현상 분석 에 이어 녹아웃 또는 녹핀 실험에 의해 달성 될 수있다3. 이 두 시스템은 최소한 변환 시스템과 유전자 편집 시스템을 포함하여 유전 공학 프로토콜의 가용성에 전적으로 의존합니다. 다양한 필라멘트 곰팡이4에서개발 된 다양한 변환 시스템이 있습니다. 생물학적 및 전기화에 의존하는 것과 같은 물리적 시스템은 트리코데르마 하르지아눔5와 아스퍼길루스 니제르6에서각각 개발되었습니다. 염화칼슘이나 아세테이트 리튬 과 같은 화학 물질을 활용하는 시스템은 Neurospora crassa7에서개발되었습니다. 마지막으로, 변형을 위한 Agrobacterium tumefaciens의 사용에 의존하는 생물학 시스템은 성공적으로 세라토시티스 albifundus8에서사용되었습니다.
다른 변환 프로토콜의 가용성과는 달리, 게놈 편집 시스템은 덜 풍부합니다. 필라멘트 곰팡이에서 수행된 많은 전통 기능 특성화 실험은 게놈3에서표적 영역 또는 유전자에 대한 상동성 의 영역에 의해 측면이 있는 선택 가능한 마커의 형태로 분할 마커 녹아웃 구조를 활용하였다. 이 방법은 상동성 지향 (HR) DNA 수리에 의존, 이는 녹아웃 구조와 관심 영역 사이의 상동성 재결합을 시설. 이러한 재결합 이벤트는 선택 가능한 마커의 서열과 관심 있는 유전자의 교체를 초래한다. 불행하게도, 이것은 Cercospora 니코티아나(10), 아스퍼길러스 퓨미가투스11 및 그로스만니아 클라비게라12를포함한 많은 종에서 성공했지만, 동종 재조합의 비율은 다른 곰팡이 종에 걸쳐 매우 가변적이며, 이는 특정 종3에서비효율적이고 때로는 사용할 수없는 프로토콜을 만듭니다.11
아연 손가락 뉴클레아제(ZFN)와 전사 활성제 와 같은 이펙터 뉴클레아제(TALENs)를 사용하는 것을 포함한 다른 게놈 편집 시스템은 특히 구체적이고 표적화된변화를만드는 능력을 감안할 때 구형 시스템에 큰 개선을 나타냈다. ZFNs와 TALENs 모두 뉴클레아제 단백질과 특정 뉴클레오티드 서열을 인식할 수 있는단백질(13)으로구성된다. 인식시, 핵은 특정 돌연변이의 도입을 용이하게 할 수 있는 이중 좌초된 DNA 브레이크를 유도합니다. 게놈 변화를 가져오기 위하여는, 뉴클레오티드 서열을 인식하는 단백질 지역은 각 실험을 위해 특별히 설계되어야 합니다. 단백질-핵산 상호작용에 의존하여 각 녹아웃 또는 녹핀 실험에 대한 표적 분자를 편집, 설계 및 생산하는 것은 어렵고 노동 집약적인14,,15이다. 이러한 과제의 예시, 극소수의 필라멘트 균류는 이러한 시스템을 사용하여 게놈 편집을 실시했습니다. 한 가지 예는 쌀 폭발 곰팡이, Magnaporthe oryzae16에서개발 된 TALENs 기반 시스템입니다.
틀림없이 게놈 편집 분야에 대한 가장 큰 혁명은 CRISPR-Cas9 시스템의 발견 및 후속 개발이었다 - RNA 분자에 의해 유도되는 엔도누셀에 의해 관심의 순서의 표적 분열을 허용하는 게놈 편집기. 이것은 CRISPR-Cas9 시스템의 주요 이점으로 단백질 핵산 상호 작용에 의존한 이전에 개발된 게놈 편집기에 큰 개선이었다 관심 있는 지역을 표적으로 하는 RNA 분자에 의존한다는 것입니다. 이는 시스템이 RNA-DNA 상호 작용에 의존하므로 각실험(15)을설계할 때 표준 기본 페어링 규칙을 악용할 수 있음을 의미합니다.
여기에서 자세히 설명된 CRISPR-Cas9 시스템은 단일 가이드 RNA(sgRNA), Cas9 효소 및 기증자 DNA(dDNA)17의세 가지 주요 구성 요소로 구성됩니다. sgRNA는 프로토스페이서라고 불리는 20개의 뉴클레오티드 영역뿐만 아니라스캐폴드(18)라고불리는 더 긴 영역으로 구성된다. 프로토스페이서 영역은 편집 시스템을 대상 영역으로 안내하는 데 사용되므로 각 실험에 대해 재설계되었습니다. 스캐폴드는 Cas9 효소에 물리적으로 결합하여 리보뉴클레오단백질(RNP)을 형성하고, 따라서 표적화되는 부위에 관계없이 동일하다는 RNA의 영역이다. Cas9 효소는 본영역(19)을식별하는 가이드로서 프로토스페이서를 사용하여 표적 DNA의 분열을 물리적으로 용이하게 한다. 마지막 구성 요소인 dDNA는 선택 사항이며 그 사용은 특정실험(20)에따라 달라집니다. dDNA는 Cas9 효소에 의해 절단되는 영역내로 특별히 삽입되어야 하는 서열을 항구하며, 따라서 유전자가 게놈으로 도입되고 있는 유전자 녹킨 실험또는 항생제 내성 유전자 또는 다른 선택 가능한 마커가 관심 있는 유전자를 대체하기 위해 도입되는 유전자 녹아웃 실험에 이상적입니다. dDNA는 또한 게놈으로 새로운 서열을 소개하는 것과 같은 방법으로 디자인될 수 있습니다. 예를 들어, 아래에 상세한 바와 같이, 유전자 잘림이 필요할 때 관심 유전자의 특정 부위에 인프레임 스톱 코돈이 도입될 수있다(21). 그밖 응용 프로그램은 기능성도메인(22)과같은 유전자의 특정 영역의 돌연변이를 포함하거나, 태그서열(23)의 도입을 포함한다.23
CRISPR-Cas9 시스템을 사용하는 주요 이점은 다재다능한24입니다. 이러한 적응성의 한 가지 예는 Cas9 효소가 사용되는 특정 변형 시스템에 따라 DNA, RNA 또는 단백질의 3가지 형태 중 하나에서 숙주 세포로 도입될 수 있다는 것입니다. DNA 형태로 도입될 때, cas9 유전자는 종종 선택 가능한 마커, sgRNA를 발현하는 카세트 및 필요한 경우 dDNA서열(25)을코딩하는 카세트와 함께 플라스미드에 포함된다. 이 시스템의 주요 장점은 단일 구조만 세포로 변환되어야 하며 성공적인 변환을 통해 CRISRP-Cas9 매개 게놈 편집에 필요한 모든 구성 요소가 존재한다는 것입니다. 그러나 이 메서드는 호스트 종에 대한 식 시스템의 가용성에 의존합니다. Cas9이 DNA 손상을 성공적으로 유도하려면 높은 수준에서 발현되어야 하므로 적합하고 잠재적으로 특정 특정 프로모터가 필요합니다. 이러한 프로모터가 아직 개발되지 않은 비모델 종의 경우, 이는 RNA 또는 단백질 형태로 Cas9를 도입하는 능력이 더 매력적인 옵션이 될 수 있습니다. 세포로 RNA의 도입은 RNA가 불안정하고 변환 과정에서 살아남을 수 없다는 점에서 특히 자신의 도전을 가져온다. 더욱이, DNA 또는 RNA 형태로 도입될 때, Cas9 유전자 서열은 특정 숙주시스템(17)에서사용하기 위해 코돈에 최적화될 필요가 있을 수 있다. 예를 들어, 연쇄상 구균 pyogenes에서 cas9 유전자는 포유류 숙주 세포 및 포유류 세포에서 사용하기 위해 코돈 에 최적화된 cas9 유전자에서 작동하지 않을 수 있습니다 식물 세포에서 작동하지 않을 수 있습니다. 이러한 모든 과제는 SgRNA와 함께 RNP로 조립되어 숙주세포(26,,27)로변형될 수 있는 Cas9의 단백질 형태를 사용하여 극복할 수 있다. 이 시스템은 내인성 발현 시스템이나 코돈 최적화에 의존하지 않으므로 대부분의 비모델 종에서 작동해야 합니다. 단백질 기반 시스템의 단점은 Agrobacterium-매개전송과 같은 DNA 기반 변환 시스템과 호환되지 않는다는 것입니다. 따라서, 단백질 기반 방법이 작동하려면, 프스토플라스트 또는 바이오리스트에 의존하는 것과 같은 변환 프로토콜을 사용할 수 있어야 한다. 이 RNP 기반 시스템은 필라멘트 균류, 푸사리움 옥시스포럼26 및 Mucor circinelloides27에성공적으로 사용되었습니다.
세라토시스티드아세이 가족의 일원인 헌티엘라 오마넨시스는갓 다친 우디 식물28에서흔히 발견되는 국제적인 곰팡이입니다. 고품질 게놈 및 전사체 데이터는 이 종28,,29,,30에유효하지만, 변형 또는 게놈 편집 프로토콜은 개발되지 않았습니다. 현재까지 H. omanensis에 대한 연구는 성적 주기29,,31의기본 유전 구성 요소에 초점을 맞추고있다. 이 곰팡이는 전형적인 이단성 성적 주기를 나타내며, 성적 번식은 MAT1-1과 MAT1-2 짝짓기 유형31의 분리 사이에만 발생합니다. 대조적으로, MAT1-2 분리는 밀접하게 관련된 헌티엘라 moniliformis의 독립적인 성적 번식이 가능하고 MAT1-1파트너(31)의부재시 성적 주기를 완료할 수 있다. 이러한 성적 능력의 차이는 적어도 부분적으로는 짝짓기 유전자의 큰 차이로 인해, MAT1-2-7로인해 H. omanensis가 전체 길이와 그대로 복사를 품고 있는 반면, 유전자는 H. moniliformis29,,31에서심각하게 잘린 것으로 생각된다. 이러한 유전자의 역할을 더욱 특성화하기 위해 H. omanensis의 MAT1-2-7 유전자는 H. moniliformis21에서볼 수 있는 잘림을 모방하기 위해 잘려나았다.
아래 프로토콜은 CRISPR-Cas9 게놈 편집 시스템의 단백질 기반 버전을 사용하여 H. omanensis의 변환 및 MAT1-2-7 유전자의 잘림에 대해 자세히 설명합니다. 이 프로토콜은 동종 재조합 기반 유전자 교체 및 플라스미드 계 CRISPR-Cas9 게놈 편집의 접근이 실패한 후에 개발되었다.
프로토콜
1. sgRNA의 설계 및 합성
- sgRNA의 일부를 형성할 잠재적 프로토스페이서 영역을 식별하려면 사용 중인 프로그램의 검색 기능을 사용하여 5'NGG 3'삼중항에 대한 관심 유전자를 수동으로 검색합니다. PAM 시퀀스로 이러한 세 쌍둥이를 노를 테고.
참고: 잠재적인 PAM 및 프로토스페이서 시퀀스를 검색하고 인음표하는 다양한 소프트웨어 프로그램을 사용할 수 있습니다.- 확인된 각 PAM 시퀀스의 20bp 업스트림을 선택하고 이러한 시퀀스를 잠재적 프로토스페이서로 추가합니다.
- 단일 프로토스페이스를 결정하려면 다음 필터링 단계를 수행하여 품질이 낮은 시퀀스를 폐기합니다.
- 잠재적 프로토스페이서의 특이성을 테스트하려면 PAM 서열과 프로토스페이서를 단일 시퀀스로 결합하여 전체 게놈에 대한 BLASTn 쿼리로 사용합니다. 대상영역(도 1A)을제외한 게놈 내의 모든 영역과 유사성을 보이는 프로토스페이스를 폐기한다.
- RNA 분자가 Cas9 효소에 결합하기 위한 올바른 3D 구조로 접을 수 있도록, Cas9 효소에 특이적인 프로토스페이서 및 스캐폴드 서열을 포함하는 결합된 서열을 생성한다.
- 결합된 프로토스페이서-스캐폴드 서열을 RNA 이차 구조 예측도구(재료표)에업로드한다.
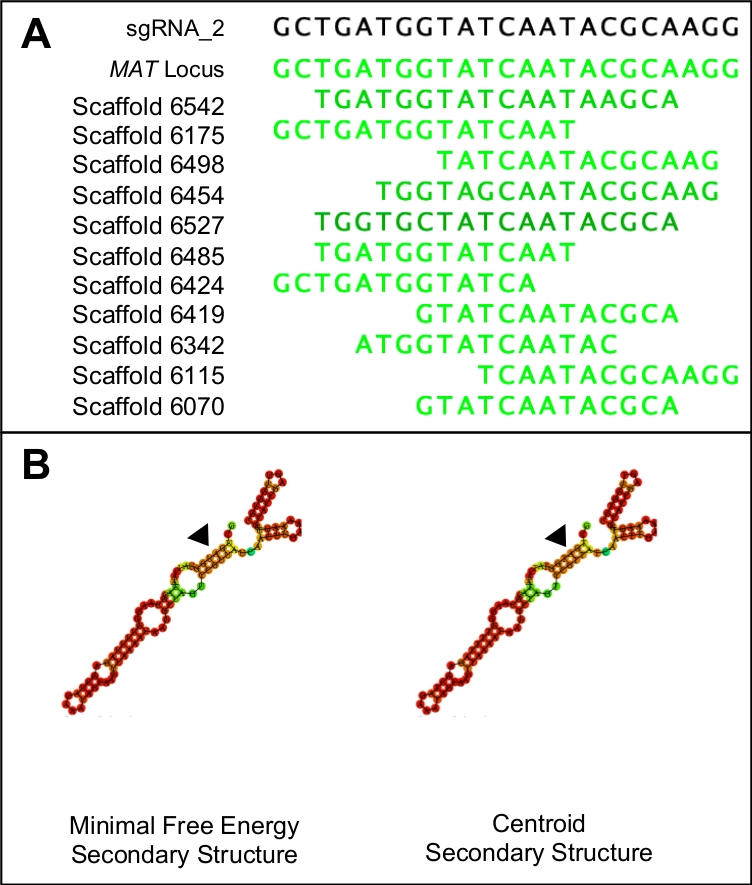
- 최소한의 자유 에너지와 중심보조 구조를 비교하여 결과를 분석합니다.
참고 : 이상적인 프로토 스페이서 후보는 동일한 최소한의 자유 에너지와 중심 보조 구조를 해야합니다. 두 보조 구조는 5개의 링 구조에 의해 중단되는 세 개의 줄기 루프로 구성되어야 합니다. 또한 구조는 프로토스페이서(도1B)를나타내는 영역을 제외하고 전체 구조에 걸쳐 높은 결합 확률(빨간색으로 표시)을 나타내야 한다. 실제 에너지 값은 관련이 없습니다.
- 대상 지역에 가장 가까운 후보를 선택하여 위의 필터링 단계를 통과한 후보중 최종 후보를 선택합니다.
- sgRNA를 단일 RNA 분자로 합성하려면, 사용되는 특정 Cas9 효소와 호환되는 sgRNA 합성 키트(재료 의표)를사용한다(예를 들어, 연쇄상 구균 pyogenes로부터의 Cas9).
참고: 다음 단계는 사용되는 sgRNA 합성 키트에 의존할 수 있다. 다른 키트가 사용되는 경우 제조업체의 지침을 따르십시오. 대안적으로, sgRNA는 미리 합성된 주문할 수 있다. RNA로 작업할 때 핵이 없는 시약과 일회용을 사용하십시오.- 위의 1.3 단계에서 선택한 20 bp 프로토 스페이서가 5'끝에 G를 항구하지 않으면이 영역에 G를 추가하십시오.
- T7 프로모터 시퀀스를 대상 시퀀스의 5'끝에 추가합니다. 이 시퀀스는 표준이며 5'TTCTAATAACGACTCACTATAG 3'입니다.
- 대상 시퀀스의 3'끝에 14 nt 겹침 시퀀스를 추가합니다. 이 시퀀스는 키트에 특화되어 있으며 여기에 사용되는 키트에 대한 5'GTTTTAGAGCTAGA 3'입니다.
- 생성된 단편 5' TTCTAATACGACTCACTATAG(N)20 GTTTTAGAGCTAGA를 주문하고, (N)20은 선택한 프로토스페이서를 나타내는(재료의 표)를나타냅니다.
- 제조업체의프로토콜(재료표)에따라 실온에서 다음과 같은 시약을 결합합니다: 물 2μL, 반응 완충제 10μL, 합성 프로토스페이스서 서열 5μL, 0.1 M DTT의 1μL 및 전사 효소 2 μL.
- 이 용액을 37°C에서 30분 동안 배양하고 얼음으로 옮습니다.
- 물 30μL과 DNase I의 2 μL을 넣고 37°C에서 15분 동안 혼합하고 배양합니다.
- 결과 sgRNA를 2% 아가로즈 젤에 시각화합니다.
2. sgRNA의 체외 분열 능력 테스트
참고: 이 단계는 선택 사항이지만 권장됩니다.
- 선택한 sgRNA가 표준 DNA 폴리머라아제를 사용하여 영역을 표적으로 하고 증폭시키는 사이트의 서열을 수용하는 단편을 증폭시키는 설계 프라이머.
참고: 가능하면, 대상 부위의 골짜기는 표준 아가로즈 젤에서 서로 쉽게 구별할 수 있는 매우 다른 크기의 두 개의 조각을 생성하는 방식으로 프라이머를 설계한다. - 상기 합성된 바와 같이 30nM sgRNA로 구성된 용액을 배양함으로써 결합된 sgRNA-Cas9 리보뉴클레오단백질(RNP)을 조립하여, 30nM Cas9 단백질, 10x 반응 완충제 및 10μL의 물 10분 동안 10μL을 10분 동안 조립한다.
- 3nM의 최종 농도에 대한 표적 영역의 PCR 생성물을 RNP 용액에 첨가하여 sgRNA의 골짜기 능력을 시험한다.
- 용액을 37°C에서 15분 동안 배양합니다.
- 단백질 효소 K 3μg와 RNase 2 μg를 용액에 추가하여 분열 반응을 멈추고 실온에서 10분 동안 배양합니다.
- 결과 DNA 단편을 2% 아가로즈 젤에 시각화합니다. 상기 sgRNA는 젤상에서 예상되는 크기의 두 개의 밴드가 관찰되는 경우 생체 내 실험에 적합하다.
3. dDNA의 설계 및 합성
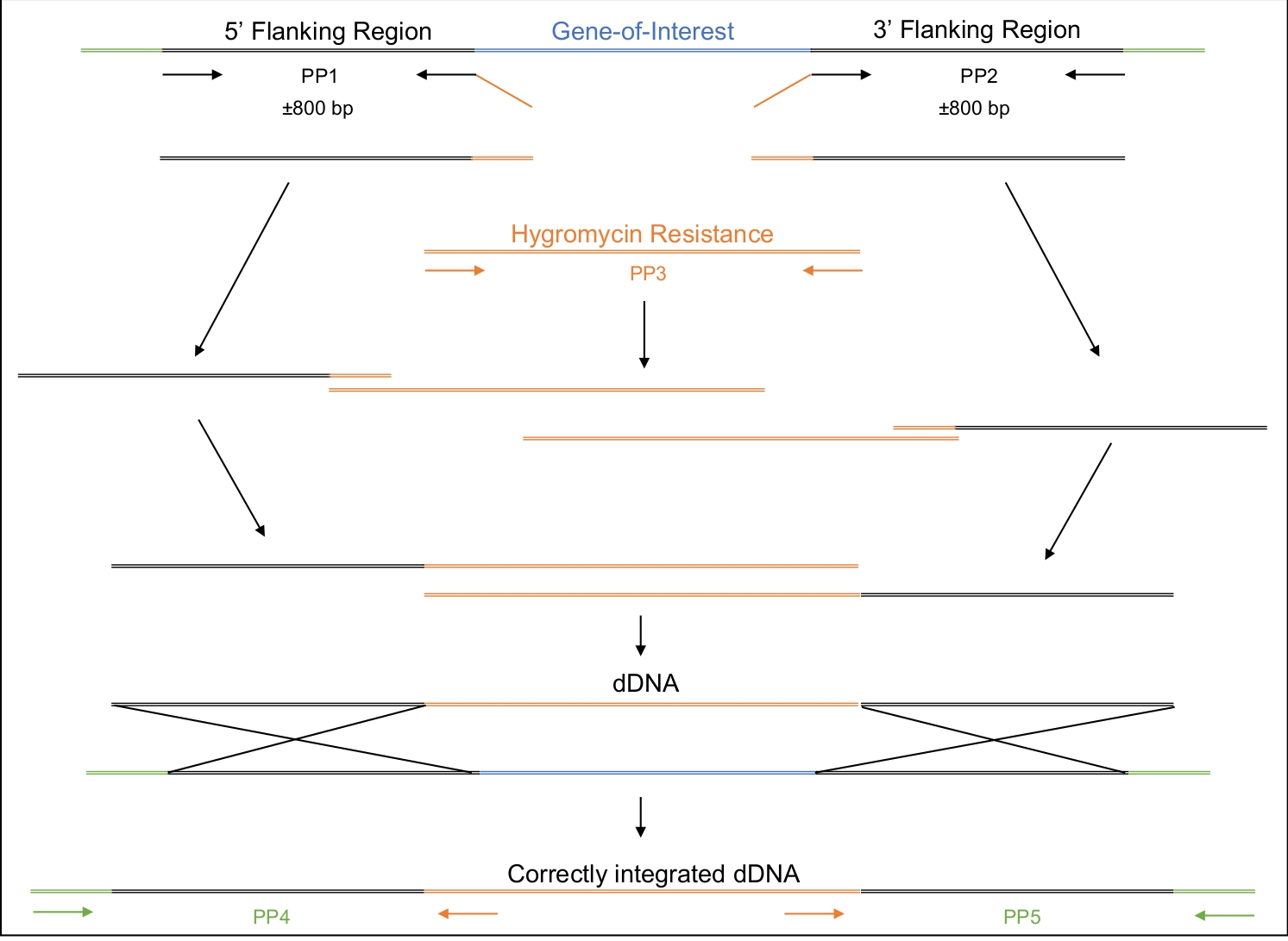
- 표적화되는 게놈 영역과 중간 부위에 상보성을 가지는 3개의 영역-5' 및 3'영역으로 구성된 dDNA를설계(재료표, 도 2).
참고: 다른 특정 시퀀스를 이 선택 가능한 마커에도 추가할 수 있습니다. 이러한 특정 실험을 위해, 스톱 코돈 서열(5'TGA 3')이 선택 가능한 마커 바로 앞에 첨가되어 인프레임 스톱 코돈이 유전자에 도입되었다. dDNA는 미리 합성된 주문할 수 있습니다. 대안적으로, dDNA는 아래 와 같이 단계적으로, 겹치는 PCR 접근 방식을 사용하여 증폭되고 조립될 수 있다.- 5'와 3'측면 영역의 약 800 bp를 증폭하는 프라이머를 설계합니다. 선택 가능한 마커 시퀀스에 상호 보완적인 20nt 의 시퀀스를 5' 영역의 역 프라이머및 3' 영역의 전방 프라이머에 추가합니다.
- 선택 가능한 마커를 증폭시키는 설계 프라이머는 증폭된 제품이 관심 있는 종에서 작동하는 것으로 알려진 프로모터뿐만 아니라 저항 유전자를 항구하도록 보장합니다.
- 고충실도 DNA폴리머라제(재료표)를사용하여 3개의 dDNA 영역을 증폭시한다. 편집중인 유기체의 gDNA로부터 5' 및 3' 영역을 증폭시합니다. 관련 소스에서 선택 가능한 마커를 증폭합니다.
- 단일 반응에서 증폭된 5' 영역을 선택 가능한 마커와 결합하고, 장거리, 고충실도 DNA 폴리머라제를 사용하여 전체 영역을 증폭시한다.
- 두 번째 단일 반응에서 증폭된 3' 영역을 선택 가능한 마커와 결합하고, 장거리, 고충실도 DNA 폴리머라제를 사용하여 전체 영역을 증폭시한다.
- 마지막으로, 두 개의 선행 PCR 제품을 단일 반응으로 결합하고 전체 dDNA 서열을 장거리, 고충실도 DNA 폴리머라아제로 증폭시다.
- 1% 아가로즈 젤에 DNA 단편을 시각화합니다. 두 개 이상의 단편이 생성되는 경우, 겔 정화 키트를 사용하여 젤에서 올바르게 크기의 조각을 정화한다.
4. 상류세포 추출
- 코니아를 생산하기 위해, 1cm x 1cm 미셀리아 덮인 한천 블록이 있는 500mL 플라스크에서 신선한 2% 맥아 추출물 국물(MEB)의 200mL를 접종한다.
참고 : 모든 곰팡이가 코니디아를 생산할 수있는 것은 아닙니다. 이 경우, 균주도 사용할 수 있습니다. 이것은 전형적으로 프로토콜에서 더 멀리 lysing 효소의 더 높은 농도를 요구할 것입니다.- 25°C에서 25°C에서 흔들리는 인큐베이터에서 액체 배양을 배양하여 24-48h에 대해 120 rpm에서 흔들어 보수합니다.
참고 : 이 인큐베이션 시간과 온도는 H. omanensis에최적화되었습니다. 이것은 다른 종에 최적화될 필요가 있을 것입니다. - 코니디아를 수확하려면; 멸균 실험실 천(예를 들어, 미라클로스)의 층을 통해 액체 배양을 걸러내고, 10분 동안 4°C에서 3,220x g에서 50mL 원심분리기 튜브 및 원심분리기로 편성 현탁액을 전달한다. 상부체를 폐기합니다.
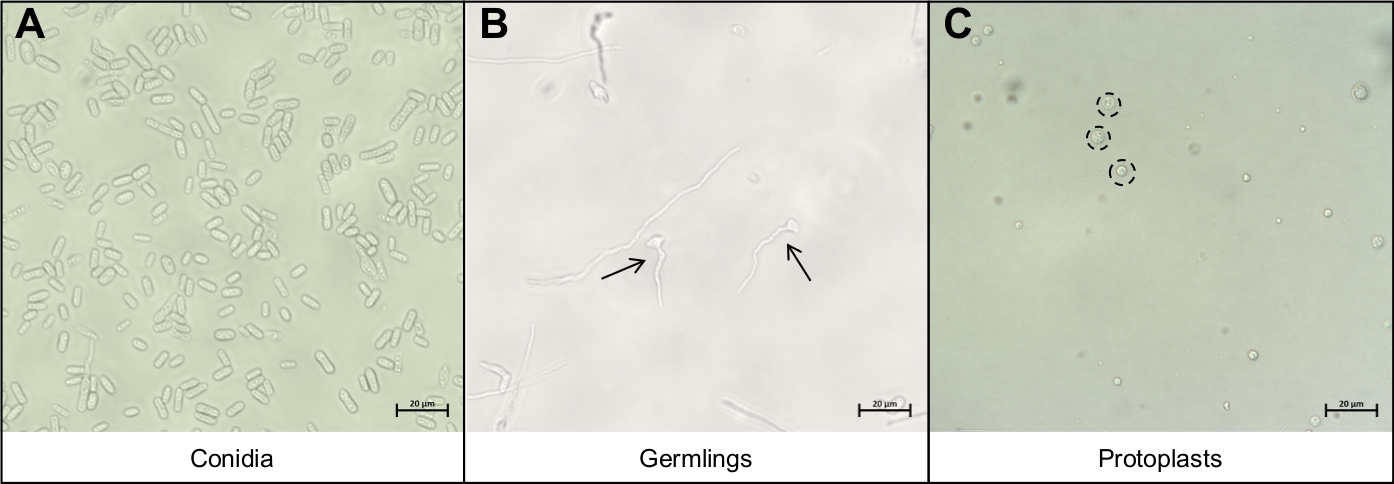
- 코니디아 용액의 5mL의 물과 파이펫 10 μL에서 코니아를 현미경 슬라이드에 재중단하고 커버슬립으로 덮습니다. 코니아만 복구되었는지 확인하기 위해 40배배율 미만의 복합 현미경을 사용하여시각화(그림 3A).
- 500 mL 플라스크에서, 재중단 코니디아의 총 부피와 신선한 1 % MEB의 200 mL을 접종.
- 25°C에서 흔들리는 인큐베이터에서 액체 배양을 120 rpm에서 최대 12시간 동안 흔들어 배양합니다.
참고 : 이 잠복기는 H. omanensis에최적화되었습니다. 이것은 다른 종에 최적화될 필요가 있을 것입니다. - 세균을 수확하려면 액체 배양을 50mL 원심분리기 튜브및 원심분리기로 4°C에서 3,220 x g에서 10분 동안 이송합니다. 상부체를 폐기합니다.
- 1 M 소르비톨의 최대 10 mL에서 세균을 다시 중단합니다.
- 배아용용액의 파이펫 10 μL은 현미경 슬라이드에 덮고 커버슬립으로 덮습니다. 40배율 미만의 복합 현미경을 사용하여 배율만 회수되었는지 확인합니다(그림3B).
참고: 프로토콜은 여기에서 일시 중지할 수 있습니다. -80°C에서 1M 소비톨에 세균을 저장합니다.
- 25°C에서 25°C에서 흔들리는 인큐베이터에서 액체 배양을 배양하여 24-48h에 대해 120 rpm에서 흔들어 보수합니다.
- 젊은 세균의 세포벽을 lyse및 프토플라스트를 방출하려면, 멸균 50mL 플라스크에서 다양한 농도에서 리스 효소의 9mL에 세균 현탁액의 1mL을 추가합니다.
참고: 다른 효소 농도와 인큐베이션 시간이 사용되며 표 1에서찾을 수 있습니다. 효소와 농도는 또한 곰팡이에 따라 달라질 가능성이 있으며 각 종에 최적화해야합니다.- 25°C에서 흔들리는 인큐베이터에서 포자 효소 용액을 배양하여 80 rpm에서 2-3시간 동안 흔들어 보시합니다.
- 멸균 실험실 천층을 통해 프톱서스트 용액을 걸레로 걸이고 10분 동안 4°C에서 1,810 x g에서 원심분리로 프톱서스트를 수집합니다. 상부체를 폐기합니다.
참고: 전형체는 세포벽이 없는 세포이므로 기계적 중단에 매우 민감합니다. 특히 파이펫팅 시 신중하게 처리해야 합니다. - STC 버퍼의 200 μL(재료의 표)에서프토플라스트 펠릿을 조심스럽게 재중단합니다.
- 프로토프스트 용액의 피펫 10 μL은 현미경 슬라이드에 덮고 커버슬립으로 덮습니다. 40배율 미만의 복합 현미경을 사용하여 전형만 회수되었는지 확인합니다(도3C).
- 혈전계를 사용하여 위의 단계에서 생성된 프톱서의 수를 계산하고 계산합니다. 프로토프스트 용액을 약 5 x 106 프토플라스트가 들어 있는 알리쿼트에 희석합니다.
참고: 프로토콜은 여기에서 일시 중지할 수 있습니다. STC 버퍼에 -80°C에서 프톱라스트를 저장합니다.
5. 프토플라스트 및 PEG 지원 변환 및 변혁적 복구
- 변환을 시작하려면 약 5 x 106 프톱라스트를 RNP 용액의 단일 부피와 dDNA 단편의 약 6 μg를 결합합니다.
참고: 프토플라스트는 기계적 중단에 매우 민감합니다. 특히 파이펫팅 시 신중하게 처리해야 합니다.- 파이펫을 사용하여, 천천히 프로토프스트용액에 30% PTC 용액의 1mL을 드립하고 실온에서 20분 동안 용액을 배양한다.
참고: 이 단계는 민감하고 매우 중요한 단계입니다. 갓 준비된 PTC 솔루션을 사용하고 가능한 한 느리고 균등하게 셀 위에 용액을 떨어뜨려 세포 표면에 소수성 층을 생성해야 합니다. - 프로토플라스트 용액과 파이펫에 5mL의 삼투성 제어 매체(OCM)를 넣고 천천히 부드럽게 파이펫을 추가하여 용액이 완전히 혼합되도록 합니다.
- 25°C에서 흔들리는 인큐베이터에서 프로토프스트 용액을 하룻밤 사이에 80rpm에서 흔들어 보배합니다.
- 파이펫을 사용하여, 천천히 프로토프스트용액에 30% PTC 용액의 1mL을 드립하고 실온에서 20분 동안 용액을 배양한다.
- 변형된 격리를 선택하려면 솔루션을 5개의 빈 60mm 배양 판으로 나눕니다.
- 각 배양판에 30 μg/mL hygromycin B로 보충된 OCM 한천 10mL을 넣고 각 플레이트를 천천히 회전하여 완전히 섞습니다.
- 40 μg/mL hygromycin B로 보충된 삼투압 제어 매체 한천10mL을 추가하기 전에 한천의 첫 번째 층을 설정합니다.
- 단일 격리가 한천의 두 층을 통해 성장하는 것을 볼 수있을 때까지 한고의 두 번째 층이 25 °C에서 배양하고 배양할 수 있도록하십시오.
- 성공적으로 변형된 분리물을 회수하기 위해 40 μg/mL hygromycin B로 보충된 한천 층을 통해 성장할 수 있는 개별 분리물을 50μg/mL hygromycin B(MEA-50)로 보충한 신선한 몰트 추출물 한천(MEA) 플레이트로 이송합니다.
- 5일 동안 25°C에서 신선한 배양을 배양하여 매일 성장을 확인합니다. 이 매체에서 지속적인 성장을 할 수있는 문화는 성공적으로 변형되었으며 추가 연구에 사용할 수 있습니다.
6. dDNA의 통합 및 안정성 확인
- dDNA가 표적 부위에서 게놈에 통합되었음을 확인하기 위해 예측된 5' 및 3'인서트사이트(그림 2)를측면으로 하는 설계 프라이머를 설계한다.
- 이 두 프라이머 세트와 고충실도 DNA 폴리머라제를 사용하여 두 개의 PCR을 수행합니다. 두 PCR 모두 예상 크기와 서열의 앰플리턴을 산출하는 경우 dDNA가 대상 영역에 성공적으로 통합되었습니다. 그런 다음, dDNA의 안정적인 통합을 위해 돌연변이 얼룩을 평가한다.
- dDNA가 게놈에 안정적으로 통합되어 식물 성장 중에 유지될 것임을 확인하기 위해 미디어 전송 테스트를 수행한다.
- MEA-50 배지에서 미아-50 배지에 분리되는 활발하게 성장하는 돌연변이에서 미약MEA 배지로 균천으로 덮인 한 덩어리를 전송한다. 3 일 동안 25 °C에서 배양하십시오.
- MEA 배지에서 성장하는 격리된 식소에서 MEA-50 배지로 미셀리아 로 덮인 한도를 전송합니다. 3 일 동안 25 °C에서 배양하십시오.
- 이 과정을 반복, 적어도 4 라운드 에 대한 보충에서 보충 매체로 적극적으로 성장하는 mycelia를 전송합니다.
참고: 많은 전송 후 MEA-50 배지에 대한 지속적인 성장이 가능한 경우 dDNA는 게놈에 안정적으로 통합되어 식물 성장을 통해 유지될 수 있습니다. 돌연변이 얼룩은 통합 된 dDNA의 단일 복사본의 존재를 위해 평가 될 수있다.
- dDNA가 단일 위치에서 게놈에 통합되었는지 확인하기 위해 남부 블롯 분석을 수행한다.
- 제조업체의 프로토콜에 따라 HindIII 및 EcoRI 제한 효소를 사용하여 각 돌연변이 균주에서 총 30 μg의 gDNA를 소화합니다.
참고: 제한 효소의 선택은 연구원에게 달려 있지만, 제한 효소의 인식 부위가 dDNA 서열에 존재하지 않는지 확인하십시오. - 소화된 gDNA를 0.75% 아가로즈 젤로 분리하고 표준 절차를 이용하여 DNA를 나일론 막에전달한다(32).
- dDNA 서열을 표적으로 하는 프로브를 사용하여 멤브레인을 혼성화로 피사체.
- dDNA의 짧은 (300 bp) 영역을 증폭하기 위해 프라이머를 설계합니다.
- 이러한 프라이머를 사용하여 PCR DIG 라벨링 믹스를 사용하여 프로브를 합성합니다.
- 표준절차(32)를사용하여 멤브레인 혼성화, 치료 및 시각화를 위해 새로 합성된 프로브를 사용합니다. 각 차선에서 단일 밴드만 볼 경우 dDNA는 게놈의 단일 위치에만 존재합니다. 돌연변이 균주는 이제 추가 현상 분석 및 기능 특성화 실험에 사용할 수 있습니다.
- 제조업체의 프로토콜에 따라 HindIII 및 EcoRI 제한 효소를 사용하여 각 돌연변이 균주에서 총 30 μg의 gDNA를 소화합니다.
7. 돌연변이 균주의 현상 분석
- MAT 유전자의 중단이 연구중인 곰팡이의 성적 능력에 영향을 미쳤는지 여부를 결정하기 위해 짝짓기 실험을 수행합니다.
참고: 이 단계는 연구되는 특정 유전자 및 종에 달려 있습니다. 이 경우, 표적화되는 유전자는 성적 번식에 관여하는 것으로 생각되고 따라서 짝짓기 테스트가 수행되었다. 유전자가 무성 생식에 관여하는 것을 예를 들면, 그 때 양심적인 생산 같이 측정될 수 있었습니다.- 돌연변이 균주의 이단 기능을 테스트하기 위해, 돌연변이 균주뿐만 아니라 반대 짝짓기 유형의 변형으로 신선한 MEA 배지를 공동 접종한다. H. omanensis의경우, 플레이트의 뚜껑을 닫아 두지만 밀봉되지 않고 실온에서 7 일 동안 배양하십시오. 성적 구조의 생산에 대한 시각적 평가.
- 돌연변이 균주의 호모탈릭 기능을 테스트하기 위해, 돌연변이 변형으로 신선한 MEA 배지를 접종한다. H. omanensis의경우, 플레이트의 뚜껑을 닫아 두지만 밀봉되지 않고 실온에서 7 일 동안 배양하십시오. 성적 구조의 생산에 대한 시각적 평가.
- MAT 유전자의 중단이 연구중인 곰팡이의 성장 속도에 영향을 미쳤는지 여부를 결정하기 위해 성장 속도 실험을 수행합니다.
- 대형 멸균 파이펫 팁의 뒷면을 한천에 삽입하여 돌연변이 및 야생형 균주의 배양의 활발해지는 가장자리에서 균천으로 덮인 한천 플러그를 만듭니다.
- 이 천 플러그와 신선한 MEA 매체를 접종. 각 배양 유형당 적어도 세 개의 복제가 이루어지도록 합니다.
- 20°C에서 3일간 성장한 후, 두 개의 수직 직경에서 성장을 측정합니다.
- 야생형 및 돌연변이 균주의 데이터를 비교합니다.
결과
상술한 프로토콜은 비모델 아스코피세테, H. 오만시스로부터짝짓기 유전자로 조기 정지 코돈의 도입을 용이하게 했다. 이 과정은 CRISPR-Cas9 게놈 편집 시스템의 버전을 활용하고 이 프로토콜에서 가장 중요한 단계 중 하나는 고품질 sgRNA의 설계 및 합성입니다. 도 1은 이 분자가 A) 특히 관심 유전자를 표적으로 하고 Cas9 단백질과 결합하기 위해 게놈 및 B)의 다른 부위와 거의 유사성을 보이는 방식으로 설계되었는지를 보여줍니다. sgRNA는 또한 효과적으로 표적 영역을 c떠나고 할 수 있어야 합니다. 표적 영역의 분열을 표적으로 하고 허용하는 sgRNA의 능력은 시험관 내에서실시되었고, 예상 크기의 두 가지 제품을 산출하였다.
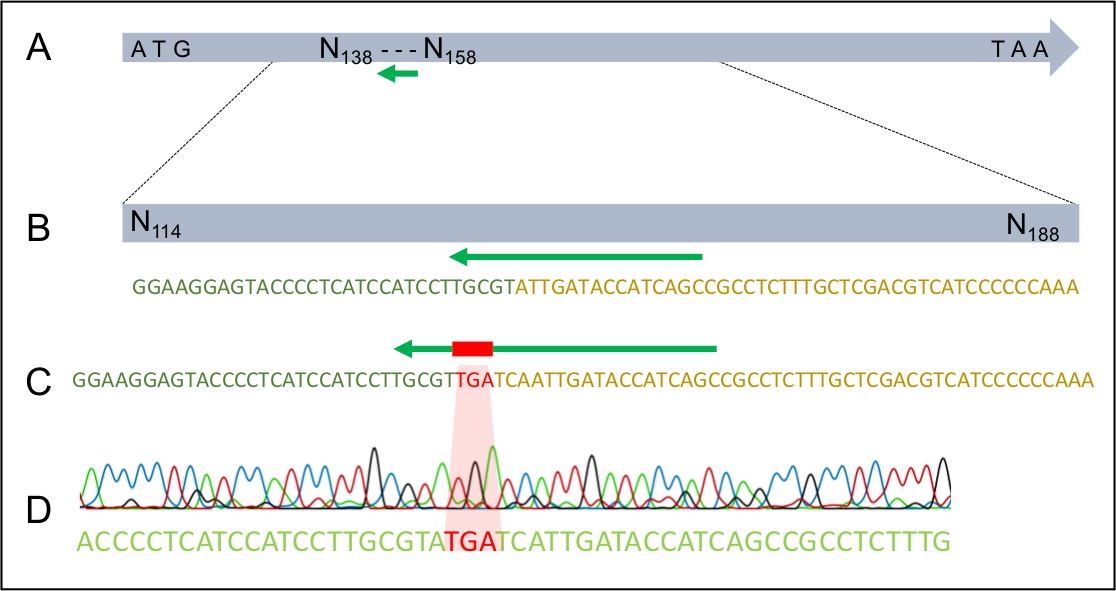
성공적인 변환이 이루어지면 dDNA가 예상 된 장소에서 한 번만 게놈에 통합되었는지 확인하는 것이 중요합니다. 그림 2는 삽입 부위를 대상으로 하는 PCR 프라이머의 설계를 보여 주며, 이는 올바른 통합 사이트에 대한 잠재적 인 변압제를 선별하는 데 사용할 수 있습니다. 5' 및 3' 삽입 부위 측면에 있는 프라이머를 설계하면 dDNA가 올바른 영역에 삽입되는 경우에만 증폭이 가능합니다. 도 4는 조기 정지 코돈이 정확한 판독 프레임으로 MAT1-2-7 유전자로 도입되어 유전자가 H. moniliformis와유사한 방식으로 잘려낼 수 있도록 하는 것을 보여줍니다. 더욱이, 남부 블롯 분석은 dDNA 구조가 게놈의 단일 부위에만 통합되었다는 것을 보여주었다.
프로토콜의 성공은 돌연변이 균주의 현상 분석시 확인되었다. MAT1-2-7 중단 실험의 경우, 2개의 독립적인 돌연변이 균주가 개발되었다. 두 분리체에서 식물성 방사형 증가율이 현저히 감소하여 새로운 짝짓기유전자(도 5)의자극효과를 시사하였다. 더욱이, 돌연변이 분리는 성적 포자를 생성하지 않은 미성숙 한 성적 구조를 생산, 성적 주기를 완료 할 수 없었다(그림 5). 이것은 잠복의 며칠 안에 전체 성적 주기를 완료 한 야생 형 분리(그림5)와는 대조적이었다.

그림 1: 적합한 sgRNA 후보 선택.
(A)적합한 sgRNA는 게놈의 표적 영역과 유사성만 갖는다(이 경우 MAT 메뚜기 서열에 의해 표시). (B)적합한 sgRNA는 동일한 최소한의 자유 에너지 및 중심 보조 구조를 가지며, 3개의 줄기 루프와 5개의 고리가 기본 스텝 루프에 들어설 것이다. 또한 구조의 대부분은 높은 구속력 확률(어두운 주황색과 빨간색으로 표시)을 가지며, 더 낮은 구속력 확률은 프로토스페이서 영역(검은 색 삼각형으로 표시)에서 볼 수 있어야 합니다. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
{kind=link}

그림 2: dDNA의 설계, 증폭 및 조립.
제1 및 제2 프라이머 쌍(PP1 및 PP2)은 관심 유전자의 약 800bp 업스트림(5') 및 800bp 하류(3')를 증폭시키는 데 사용됩니다. PP1의 역프라이머와 PP2의 전방 프라이머에는 히그로마이신 저항 카세트에 대한 호모로지 영역이 포함됩니다. 세 번째 프라이머 쌍은 전체 히그로마이신 저항 카세트를 증폭시합니다. 단계적 방식으로, 다양한 앰플리콘은 전체 dDNA까지 조립되며, 5' 영역, 히그로마이신 저항 카세트 및 3'영역으로 구성된, 조립된다. 세포로 변형될 때, dDNA는 Cas9 효소가 절단하도록 지시될 지역에서 재결합하여 관심 있는 유전자를 히그로마이신 저항 카세트로 대체해야 합니다. PP4 및 PP5는 dDNA가 적절한 위치에서 게놈에 올바르게 삽입되었는지 여부를 결정하는 데 사용할 수 있다. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
{kind=link}

그림 3: 프토플라스트 추출 프로토콜 동안 중요한 다른 세포 유형입니다.
(A)코니디아는 프로토콜의 시작 재료로 사용된다. 이러한 코니디아는 발아하고(B)젊은 세균이 될 때까지 성장할 수 있습니다. 젊은 세균의 이상적인 성장 단계는 두 개의 검은 화살표에 의해 표시됩니다. (B)에서 본 다른B균원가 는 저하에 너무 성숙하고 사용해서는 안됩니다. 프로토콜의 마지막 단계는(C)둥근 프토플라스트의 방출이며, 검은 색, 점선 원으로 표시된다. 이 세포는 더 이상 세포벽을 가지고 있지 않으며 따라서 기계적 중단에 매우 민감합니다. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
{kind=link}

도 4: TGA 스톱 코돈의 성공적인 통합을 H. 오마넨시스의 MAT1-2-7 유전자로 성공적으로 통합한다.
(A)전신 H. 오만증 MAT1-2-7 유전자, 녹색 화살표로 표시된 sgRNA 표적 부위. (B) H. 오만시스 MAT1-2-7 유전자 내의 sgRNA 표적 부위의 확대 회로도. (C) H. 오마넨시스의 MAT1-2-7 유전자에 동종에 의해 측면에 있는 정지 코돈을 보여주는 dDNA 영역의 확대 회로도. (D) Sanger 서열 크로마토그램은 MAT1-2-7 유전자에 스톱 코돈의 성공적인 통합을 나타낸다. 윌슨 외 202021에서수정 . 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
{kind=link}

그림 5: (A) 야생형 분리및 (B) 돌연변이 분리 사이의 표현성 차이.
각 패널의 처음 세 이미지는 두 개의 격리 유형의 성적 능력의 차이를 보여줍니다. 야생형 분리는 성재생 중 성숙한 아스코마타를 형성하지만, 목의 끝에서 포자의 발산으로 완성되지만, 돌연변이 분리는 성적인 포자를 생성하지 않는 미성숙한 성적 구조만형성한다. 각 패널의 네 번째 이미지는 두 분리 유형의 성장률과 형태학의 차이를 보여줍니다. 야생형 분리는 훨씬 더 빠르게 증가하고 더 많은 공중 균증으로 증가하지만, 돌연변이는 느리게 표시하고 한천 내에 잠겨 있습니다. 윌슨 외 202021에서수정 . 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
{kind=link}
| 반응 | 효소 농도 | 성능 저하 시간 |
| A | 1.250 mg/mL | 180분 |
| B | 1.875 mg/mL | 180분 |
| C | 2.500 mg/mL | 150분 |
| D | 3.750 mg/mL | 150분 |
| 전자 | 4.375 mg/mL | 120분 |
| F | 5.000 mg/mL | 120분 |
표 1: 트리코데르마 하르지안눔의 용액을 분해하는 세균/미셀리아 용액의 분해. 다른 효소 농도 는 다른 잠복기에 해당, 더 긴 배양을 필요로 하는 낮은 농도와.
토론
H. omanensis의 성공적인 변환 및 MAT1-2-7 유전자의 편집을 위한 프로토콜은 hygromycin B21에저항을 위한 유전자와 함께 프레임 내 조기 정지 코동을 도입하여 입증되었다. 이것은 CRISPR-Cas9 게놈 편집 시스템의 단백질 기지를 둔 버전을 사용하여 달성되었습니다. 이 실험은 sgRNA의 체외 전사, dDNA의 PCR 기반 조립 및 이 두 핵산의 공동 변환을 수반하며, 이 두 핵산은 H. omanensis로부터 추출된 프토프러스트로 시판되는 Cas9 효소를 결합했습니다.
다른 많은 분자 도구의 가용성에 의존하는 다른 프로토콜과 달리, 위에서 설명한 프로토콜은 분자 도구 상자가 여전히 상당히 제한된 종에서 성공적으로 사용될 수 있다21. 프로토콜은 확립된 변환 시스템 및 NGS 데이터의 가용성, 바람직하게는 전체 게놈 서열에만 의존합니다. 효과적인 변환 시스템은 이 것을 사용할 수 없는 종에서 일부 최적화를 취할 수 있지만, 다양한 종에 사용할 수 있는 많은 다른 프로토콜이 있다. 더욱이, 게놈 데이터는 종의 가장 모호한조차도 점점 더 유효해지고 있고 이미 존재하지 않는 경우에 de novo를 생성하기 쉽게 되고 있습니다.
프로토콜의 길이를 감안할 때 수정을 도입할 수 있고 문제 해결이 필요한 단계가 많이 있습니다. 이것은 종 특정 것으로 간주되는 단계의 특히 사실이다. 예를 들어, 실험에 중요한 세포 유형을 생성하기 위해 특정 온도 및 특정 시간 길이에서 수행해야 하는 이 프로토콜에는 많은 인큐베이션 단계가 있습니다. 따라서 이러한 단계는 종별 최적화가 필요합니다. 가능하면, 특정 세포 또는 성장 단계의 현미경 사진은 다른 종(Figure 1)으로이 프로토콜을 전달하는 것을 돕기 위해 제공되었습니다. 프루프를 방출하기 위해 곰팡이 세포의 세포벽을 저하시키는 데 사용되는 효소의 종류와 농도는 또한 연구중인 곰팡이의 종에 특이할 것이다. 이 프로토콜에서는, Fusarium verticillioides33같이 종에 있는 프루프의 추출을 위해 다른 효소 조합이 요구되는 동안, 다른 효소 조합에서 용해 효소의 단 하나 근원이 이용됩니다. 이 단계는 전적으로 세포벽의 화학적 메이크에 달려 있으며 따라서 종에 따라 최적화될 필요가 있습니다.
이 방법은 표현 시스템에 의존하지 않는 비 모델 종을 공부하는 사람들에게 특히 중요합니다. CRISPR-Cas9 게놈 편집 시스템을 확립하는 인기 있는 방법은 선택의 세포로 변형되는 하나 또는 두 개의 플라스미드로부터 Cas9 단백질, sgRNA 및 dDNA를 발현하는 것입니다. 이 경우, Cas9는 연구되는 특정 유기체에서 높은 수준의 발현이 가능한 프로모터에 의해 발현될 필요가 있다. 일반 프로모터는 필라멘트 균류에 사용하기 위해 개발되었으며 모든 종에서 호환되지 않지만 낮은 수준의 발현을 허용하며 항생제 내성 유전자와 같은 발현에 성공적으로 사용할 수 있습니다. 그러나 이러한 프로모터는 종종 높은 수준의 발현을 허용하지 않으므로 Cas9 단백질을 표현하는 데 사용할 수 없습니다. CRISPR-Cas9 게놈 편집 시스템의 단백질 기반 버전을 사용하여 이러한 한계를 극복하고 sgRNA 및 dDNA가 이미 생산된 Cas9 효소로 세포로 공동 변환될 수 있게 합니다.
H. omanensis에서 사용하기위한이 단백질 기반 시스템의 개발은 고전적인 분할 마커 접근 방식과 플라스미드 기반 CRISPR-Cas9 시스템을 사용하여 게놈 편집에 많은 실패 한 시도 후에 왔다. 효율성은 종마다 다르지만, 분할 마커 접근법은 알터나리아교류34,,35, C. 니코티아나에(36)와36같은 다양한 종에서 100% 효율로 성공적으로 사용되었습니다. 반면, H. omanensis에서 이 시스템의 효율성은 80개 이상의 독립적인 변환 및 통합 이벤트에도 불구하고 0이었습니다. 마찬가지로, 플라스미드 기반 CRISPR-Cas9 시스템은 트리코데르마 레이제이(>93%)17과 페니실륨 크리소제눔(최대 100%)37에서높은 효율로 성공적으로 사용되고 있다. 이것은, 다시, H. omanensis에서이시스템의 유용성과는 대조적으로. Cas9 단백질의 충분한 발현은 가사 유전자에서 예측된 2개의 종 특정 프로모터를 포함하여 잠재적인 프로모터의 수를 시도에도 불구하고 H. omanensis에서 달성되지 않았습니다. 따라서 이 시스템은 전혀 사용할 수 없습니다. 그러나 CRISPR-Cas9 시스템의 단백질 기반 버전을 사용하여 많은 독립적인 변압제를 산출했으며, 그 중 두 가지는 올바른 위치에 통합 된 dDNA를 수용했습니다. 또한,이 실험은 한 번만 시도되었으며 이 시스템을 사용할 수있는 용이성을 더 한번 보여주는 성공적이었습니다.
이 프로토콜의 향후 응용 프로그램에는 세라토시스티드아세아제의 다른 종에서 최적화 및 사용이 포함됩니다. 이미,이들종30,,38,39종에 사용할 수 있는 풍부한 NGS 데이터가 있으며, 호스트특이성(40)및 성장률 및독성(41)에 관한 연구가 진행되고 있다. 이 연구 결과는 이 프로세스에서 관련되기 위하여 생각되는 유전자의 기능적인 특성에 의해 강화될 수 있습니다, 지금 변환 및 게놈 편집 프로토콜의 가용성 때문에 가능하게 될 연구.
결론적으로, 비 모델 종에서 중요한 생물학적 과정의 근본적인 유전자에 대한 철저한 조사는 광범위한 생물학적 자원과 분자 도구 키트의 존재에 의존하지 않는 사용하기 쉬운 게놈 편집 프로토콜의 가용성 덕분에 접근성이 높아지고 있습니다. 비 모델 종을 연구하는 것은 쉬워지고 모델 종에서 해명 된 표준 생물학적 과정에서 새로운 경로와 흥미로운 편차의 발견을 허용합니다.
공개
저자는 공개 할 것이 없습니다.
감사의 말
이 프로젝트는 프리토리아 대학, 과학 기술부 (DST)/국립 연구 재단 (NRF) 나무 건강 생명 공학 (CTHB)의 우수 센터에 의해 지원되었습니다. 이 프로젝트는 또한 곰팡이 유전체학의 BD 윙필드 교수의 DST /NRF SARChI 의자 (교부금 번호 : 98353)뿐만 아니라 박사 AM 윌슨의 NRF 박사 학위 (108548)에 의해 지원되었다. 보조금 소지자는 이 작품에 표현된 의견, 결과 및 결론 또는 권고사항이 연구자의 의견이며 자금 조달 기관이 이와 관련하여 어떠한 책임도 지지 않는다는 것을 인정합니다.
자료
| Name | Company | Catalog Number | Comments |
| EcoRI-HF | New England Biolabs, Ipswich, USA | R3101S | |
| EnGen Spy Cas9 NLS protein | New England Biolabs, Ipswich, USA | M0646T | Used to assemble the RNP |
| Eppendorf 5810 R centrifuge | Eppendorf, Hamberg, Germany | ||
| FastStart Taq DNA Polymerase | Sigma, St Louis, USA | 12032902001 | Standard DNA polyermase |
| GeneJET Gel Extraction Kit | ThermoFisher Scientific, Waltham, USA | K0691 | |
| HindIII-HF | New England Biolabs, Ipswich, USA | R3104S | |
| HiScribeTM T7 Quick High Yield RNA synthesis kit | New England Biolabs, Ipswich, USA | E2050S | |
| Hygromycin B from Streptomyces hygroscopicus | Sigma, St Louis, USA | 10843555001 | |
| Infors HT Ecotron Shaking Incubator | Infors AG, Bottmingen, Switzerland | ||
| LongAmp Taq DNA Polymerase | New England Biolabs, Ipswich, USA | M0323S | Long-range, high-fidelity DNA polymerase |
| Malt extract agar, 2% (MEA) | 20 g ME and 20 g agar in 1 l ddH20 | ||
| Malt extract | Sigma, St Louis, USA | 70167-500G | |
| Agar | Sigma, St Louis, USA | A5306 | |
| Malt Extract broth, 1% (MEB) | Sigma, St Louis, USA | 70167-500G | 2 g ME in 200 ml ddH20 |
| Malt Extract broth, 2% (MEB) | Sigma, St Louis, USA | 70167-500G | 4 g ME in 200 ml ddH20 |
| Miracloth | Merck Millipore, New Jersey, USA | 475855 | |
| Nylon membrane (positively charged) | Sigma, St Louis, USA | 11209299001 | |
| Osmotic control medium (OCM) | 0.3% yeast extract, 20% sucrose, 0.3% casein hydrolysate | ||
| Casein Hydrolysate | Sigma, St Louis, USA | 22090 | |
| Sucrose | Sigma, St Louis, USA | 84097 | |
| Yeast extract | Sigma, St Louis, USA | Y1625 | |
| Osmotic control medium (OCM) agar | Osmotic control medium (OCM) + 1% agar | ||
| Agar | Sigma, St Louis, USA | A5306 | |
| PCR DIG Labeling Mix | Sigma, St Louis, USA | 11585550910 | |
| Phusion High-Fidelity DNA Polymerase | ThermoFisher Scientific, Waltham, USA | F-530XL | High fidelity DNA polymerase |
| Plasmid pcb1004 | N/A | N/A | From: Carroll et al., 1994 |
| Presynthesized sgRNA | Inqaba Biotec, Pretoria, South Africa | Ordered as an synthesized dsDNA with specified sequence | |
| Proteinase K | Sigma, St Louis, USA | P2308 | |
| PTC Solution | 30% polyethylene glycol 8000 in STC buffer from above | ||
| Polyethylene glycol 8000 | Sigma, St Louis, USA | 1546605 | |
| RNase A | ThermoFisher Scientific, Waltham, USA | 12091021 | |
| RNAfold Webserver | Institute for Theoretical Chemistry, University of Vienna | N/A | http://rna.tbi.univie.ac.at/cgi-bin/RNAWebSuite/RNAfold.cgi |
| RNAstructure | Mathews Lab | N/A | https://rna.urmc.rochester.edu/RNAstructureWeb/Servers/Predict1/Predict1.html |
| Sorbitol, 1 M | Sigma, St Louis, USA | 1617000 | 182.17g sorbitol in 1 l ddH20 |
| STC Buffer | 20% sucrose, 50 mM Tris-HCl pH 8.00 and 50 mM CaCl2 | ||
| Calcium chloride | Sigma, St Louis, USA | 429759 | |
| Tris-HCl pH 8.00 | Sigma, St Louis, USA | 10812846001 | |
| Sucrose | Sigma, St Louis, USA | 84097 | |
| Trichoderma harzianum lysing enzymes | Sigma, St Louis, USA | L1412 | |
| Zeiss Axioskop 2 Plus Ergonomic Trinocular Microscope | Zeiss, Oberkochen, Germany |
참고문헌
- Ekblom, R., Galindo, J. Applications of next generation sequencing in molecular ecology of non-model organisms. Heredity. 107, 1-15 (2011).
- Russell, J. J., et al. Non-model model organisms. BMC Biology. 15 (55), 1-31 (2017).
- Kück, U., Hoff, B. New tools for the genetic manipulation of filamentous fungi. Applied Microbiology and Biotechnology. 86, 51-62 (2010).
- Li, D., Tang, Y., Lin, J., Cai, W. Methods for genetic transformation of filamentous fungi. Microbial Cell Factories. 16 (168), 1-13 (2017).
- Lorito, M., Hayes, C. K., Di Pietro, A., Harman, G. E. Biolistic transformation of Trichoderma harzianum and Gliocladium virens using plasmid and genomic DNA. Current Biotechnology. 24, 349-356 (1993).
- Taylor, P., et al. Transformation of intact Aspergillus niger by electroporation. Bioscience, Biotechnology, and Biochemistry. 58 (12), 2224-2227 (2014).
- Dhawale, S. S., Paietta, J. V., Marzluf, G. A. A new, rapid and efficient transformation procedure for Neurospora. Current Genetics. 8, 77-79 (1984).
- Sayari, M., Van Der Nest, M. A., Steenkamp, E. T., Adegeye, O. O., Marincowitz, S. Agrobacterium-mediated transformation of Ceratocystis albifundus. Microbiological Research. 226, 55-64 (2019).
- Meyer, V. Genetic engineering of filamentous fungi- Progress, obstacles and future trends. Biotechnology Advances. 26, 177-185 (2008).
- You, B. J., Lee, M. H., Chung, K. R. Gene-specific disruption in the filamentous fungus Cercospora nicotianae using a split-marker approach. Archives of Microbiology. 191, 615-622 (2009).
- Gravelat, F. N., Askew, D. S., Sheppard, D. C. Targeted gene deletion in Aspergillus fumigatus using the hygromycin-resistance split-marker approach. Host-Fungus Interactions. 845, 119-130 (2012).
- Wang, Y., Diguistini, S., Bohlmann, J., Breuil, C. Agrobacterium-meditated gene disruption using split-marker in Grosmannia clavigera, a mountain pine beetle associated pathogen. Current Genetics. 56, 297-307 (2010).
- Wood, A. J., et al. Targeted genome editing across species using ZFNs and TALENs. Science. 333, 307 (2011).
- Mahfouz, M. M., Piatek, A., Neal, C. Genome engineering via TALENs and CRISPR/Cas9 systems: Challenges and perspectives. Plant Biotechnology. 12, 1006-1014 (2014).
- Wang, H., La Russa, M., Qi, L. S. CRISPR/Cas9 in genome editing and beyond. Annual Review of Biochemistry. 85 (1), 227-264 (2016).
- Arazoe, T., et al. Tailor-made TALEN system for highly efficient targeted gene replacement in the rice blast fungus. Biotechnology and Bioengineering. 112 (7), 1335-1342 (2015).
- Liu, R., Chen, L., Jiang, Y., Zhou, Z., Zou, G. Efficient genome editing in filamentous fungus Trichoderma reesei using the CRISPR/Cas9 system. Cell Discovery. 1, 1-11 (2015).
- Jinek, M., Chylinski, K., Fonfara, I., Hauer, M., Doudna, J. A., Charpentier, E. A programmable dual-RNA-guided DNA endonuclease in adaptive bacterial immunity. Science. 337, 816-822 (2012).
- Gasiunas, G., Barrangou, R., Horvath, P., Siksnys, V. Cas9-crRNA ribonucleoprotein complex mediates specific DNA cleavage for adaptive immunity in bacteria. Proceedings of the National Academy of Sciences. 109 (39), 2579-2586 (2012).
- Ran, F. A., Hsu, P. D., Wright, J., Agarwala, V., Scott, D. A., Zhang, F. Genome engineering using the CRISPR-Cas9 system. Cell. 8 (11), 2281-2308 (2013).
- Wilson, A. M., Wilken, P. M., Van Der Nest, M. A., Wing, M. J., Wing, B. D. The novel Huntiella omanensis mating gene, MAT1-2-7, is essential for ascomatal maturation. Fungal Genetics and Biology. 137, 103335 (2020).
- Miao, J., et al. Characterization of an N-terminal non-core domain of RAG1 gene disrupted Syrian Hamster model generated by CRISPR Cas9. Viruses. 10 (243), 10050243 (2018).
- Roberts, B., et al. Systematic gene tagging using CRISPR/Cas9 in human stem cells to illuminate cell organization. Molecular Biology of the Cell. 28 (21), 2854-2874 (2016).
- Schneider, S., Kirchner, M., Kirchner, M., Schneider, S. CRISPR-Cas: From the bacterial adaptive immune system to a versatile tool for genome engineering. Angewandte Chemie International Edition. 54 (46), 13508-13514 (2015).
- Nødvig, C. S., Nielsen, J. B., Kogle, M. E., Mortensen, U. H. A CRISPR-Cas9 system for genetic engineering of filamentous fungi. PLoS ONE. 10 (7), 1-18 (2015).
- Wang, Q., Cobine, P. A., Coleman, J. J. Efficient genome editing in Fusarium oxysporum based on CRISPR/Cas9 ribonucleoprotein complexes. Fungal Genetics and Biology. 117, 21-29 (2018).
- Nagy, G., et al. Development of a plasmid free CRISPR-Cas9 system for the genetic modification of Mucor circinelloides. Scientific Reports. 7 (16800), 1-10 (2017).
- Al-Subhi, A. M., Al-Adawi, A. O., Van Wyk, M., Deadman, M. L., Wingfield, M. J. Ceratocystis omanensis, a new species from diseased mango trees in Oman. Mycological Research. 110 (2), 237-245 (2006).
- Wilson, A. M., van der Nest, M. A., Wilken, P. M., Wingfield, M. J., Wingfield, B. D. Pheromone expression reveals putative mechanism of unisexuality in a saprobic ascomycete fungus. PLoS ONE. 13 (3), 0192517 (2018).
- van der Nest, M. A., et al. Draft genomes of Amanita jacksonii, Ceratocystis albifundus, Fusarium circinatum, Huntiella omanensis, Leptographium procerum, Rutstroemia sydowiana, and Sclerotinia echinophila. IMA Fungus. 5 (2), 472-485 (2014).
- Wilson, A. M., Godlonton, T., van der Nest, M. A., Wilken, P. M., Wingfield, M. J., Wingfield, B. D. Unisexual reproduction in Huntiella moniliformis. Fungal Genetics and Biology. 80, 1-9 (2015).
- Sambrook, J., Green, M. . Molecular cloning: A laboratory manual. , (2012).
- Ramamoorthy, V., Govindaraj, L., Dhanasekaran, M., Vetrivel, S., Kumar, K. K., Ebenezar, E. Combination of driselase and lysing enzyme in one molar potassium chloride is effective for the production of protoplasts from germinated conidia of Fusarium verticillioides. Journal of Microbiological Methods. , (2015).
- Lin, C., Yang, S. L., Wang, N., Chung, K. The FUS3 MAPK signaling pathway of the citrus pathogen Alternaria alternata functions independently or cooperatively with the fungal redox-responsive AP1 regulator for diverse developmental, physiological and pathogenic processes. Fungal Genetics and Biology. 47 (4), 381-391 (2010).
- Lin, C., Chung, K. Specialized and shared functions of the histidine kinase- and HOG1 MAP kinase-mediated signaling pathways in Alternaria alternata, a filamentous fungal pathogen of citrus. Fungal Genetics and Biology. 47 (10), 818-827 (2010).
- Choquer, M., et al. The CTB1 gene encoding a fungal polyketide synthase is required for cercosporin biosynthesis and fungal virulence of Cercospora nicotianae. Molecular Plant-Microbe Interactions. 18 (5), 468-476 (2005).
- Pohl, C., Kiel, J. A. K. W., Driessen, A. J. M., Bovenberg, R. A. L., Nygård, Y. CRISPR/Cas9 based genome editing of Penicillium chrysogenum. ACS Synthetic Biology. 5 (7), 754-764 (2016).
- van der Nest, M. A. M. A., et al. Draft genome sequences of Diplodia sapinea, Ceratocystis manginecans and Ceratocystis moniliformis. IMA Fungus. 5 (1), 135-140 (2014).
- Wingfield, B. D., et al. Draft genome sequences for Ceratocystis fagacearum, C. harringtonii, Grosmannia penicillata, and Huntiella bhutanensis. IMA Fungus. 7 (2), 317-323 (2016).
- Fourie, A., Van Der Nest, M. A., De Vos, L., Wingfield, M. J., Wingfield, B. D., Barnes, I. QTL mapping of mycelial growth and aggressiveness to distinct hosts in Ceratocystis pathogens. Fungal Genetics and Biology. 131, 103242 (2019).
- Lee, D. H., Roux, J., Wingfield, B. D., Wingfield, M. J. Variation in growth rates and aggressiveness of naturally occurring self-fertile and self-sterile isolates of the wilt pathogen Ceratocystis albifundus. Plant Pathology. 64 (5), 1103-1109 (2015).
재인쇄 및 허가
JoVE'article의 텍스트 или 그림을 다시 사용하시려면 허가 살펴보기
허가 살펴보기더 많은 기사 탐색
This article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. 판권 소유