
Method Article
Uma tarefa de associação odor-recompensa de baixo custo para testes de aprendizagem e memória
Neste Artigo
Resumo
Uma tarefa de aprendizagem associativa com recompensa de odor foi usada para investigar os efeitos diferenciais da manipulação fisiológica na memória de longo e curto prazo.
Resumo
Paradigmas comportamentais robustos e simples de memória apetitiva e associativa são cruciais para pesquisadores interessados em mecanismos celulares e moleculares da memória. Neste artigo, um protocolo comportamental de camundongo eficaz e de baixo custo é descrito para examinar os efeitos da manipulação fisiológica (como a infusão de agentes farmacológicos) na taxa de aprendizado e na duração da memória de odor-recompensa. Resultados representativos são fornecidos a partir de um estudo que examina o papel diferencial da atividade do receptor de tirosina quinase na memória de curto prazo (STM) e de longo prazo (LTM). Os camundongos machos foram condicionados a associar uma recompensa (pellet de açúcar) a um dos dois odores, e sua memória para a associação foi testada 2 ou 48 h depois. Imediatamente antes do treinamento, um inibidor do receptor de tirosina quinase (Trk) ou infusões de veículo foram administrados no bulbo olfatório (OB). Embora não tenha havido efeito da infusão na taxa de aprendizado, o bloqueio dos receptores Trk no OB prejudicou seletivamente o LTM (48 h), e não a memória de curto prazo (STM; 2 h). O comprometimento do LTM foi atribuído à diminuição da seletividade de odor, medida pelo tempo de escavação. O ponto culminante dos resultados deste experimento mostrou que a ativação do receptor Trk no OB é a chave na consolidação da memória olfativa.
Introdução
Os mecanismos de formação de memória associativa foram investigados anteriormente predominantemente com base em estudos de condicionamento do medo de uma tentativa. No entanto, muitas tarefas mundanas geralmente têm padrões de aquisição mais complexos e dependem de encontros repetidos. O objetivo deste protocolo é fornecer um paradigma comportamental de roedor econômico que seja usado para entender os mecanismos celulares e moleculares do aprendizado apetitivo e da memória de vários ensaios.
A aprendizagem de odores que depende do bulbo olfatório principal (BO) oferece várias vantagens para o estudo da memória apetitiva multi-tentativa. Primeiro, as memórias dependentes de OB têm durações variadas (STM, LTM e memória de médio prazo1) e dependem do mesmo mecanismo molecular 2,3 e estrutural de outras partes do cérebro, incluindo neuromodulação4, potenciação de longo prazo5 e neurogênese adulta 6,7,8. Em segundo lugar, em contraste com as regiões de ordem superior, como o hipocampo, as memórias dependentes de OB permitem observações de uma correspondência mais direta entre as manipulações do ambiente perceptivo pelo experimentador e as mudanças nos circuitos neurais responsáveis pelo aprendizado 8,9,10,11 . Neste artigo, é detalhado um paradigma de memória e aprendizado associativo dependente de OB, que pode ser usado para estudar mecanismos moleculares e estruturais gerais. Foi desenvolvido para permitir que os pesquisadores acessem as vantagens do aprendizado olfativo para o estudo dos mecanismos celulares e moleculares da memória.
Em nossa recente publicação3, o protocolo aqui descrito foi usado para demonstrar que a consolidação do aprendizado do odor apetitivo é dependente da ativação do receptor Trk dentro do GO. No protocolo abaixo, também são discutidas áreas onde o paradigma comportamental pode ser ajustado para diferentes necessidades experimentais.
Um total de 27 camundongos CD-1 machos adultos, com 8 semanas de idade no momento da canulação, foram usados neste estudo. Para obter as distribuições precisas do grupo e o uso do conjunto de odores, consulte a seção de métodos de nossa publicação anterior3. Camundongos machos foram usados para evitar grandes flutuações nos níveis de estrogênio porque pesquisas anterioresmostraram que a memória olfativa é aprimorada pelo aumento dos níveis de estrogênio. Esses camundongos foram mantidos sempre em um ciclo reverso claro/escuro de 12:12 h e tiveram acesso à água. Durante os experimentos comportamentais, as dietas dos camundongos foram restritas para mantê-las em ~ 90% de seu peso livre de alimentação. A restrição alimentar começou 3 dias antes do início do experimento comportamental. Como será descrito abaixo, o mesmo conjunto de camundongos é apresentado com diferentes conjuntos de odores para atingir os níveis apropriados de poder estatístico, minimizando o uso do animal. A seção de análise estatística mostra como contabilizar a variância aleatória que pode ser introduzida por isso.
Protocolo
O protocolo abaixo segue as diretrizes de cuidados com animais da IACUC no Earlham College.
1. Canulação do bulbo olfatório
NOTA: Essas cirurgias não requerem técnica estéril, pois não requerem grandes incisões. No entanto, cada instituição pode diferir em seus requisitos. Se os experimentadores estiverem realizando esta cirurgia em cepas de camundongos imunocomprometidos, considerações adicionais podem ser necessárias. Ao todo, os experimentadores são incentivados a discutir este protocolo com seu veterinário e equipe de cuidados com os animais antes do uso e a limpar e desinfetar todas as ferramentas entre cada cirurgia.
- Durante a configuração, mergulhe as cânulas e parafusos em um pequeno béquer ou placa de Petri contendo 32% de clorexidina para mantê-los desinfetados.
- Anestesiar camundongos com isoflurano gasoso a 4% em oxigênio puro e fixá-los em um aparelho estereotáxico. Durante a cirurgia, certifique-se de que os camundongos sejam mantidos sob anestesia com isoflurano a 1,5-2% fornecida por um cone nasal. Monitore a respiração durante toda a cirurgia. Use pomada para os olhos para evitar o ressecamento dos olhos durante a anestesia.
- Depois que o mouse estiver preso e não responder mais a um beliscão firme do pé traseiro, use clorexidina a 32% para esfregar o topo da cabeça para limpar a superfície da incisão.
- Em seguida, esfregue o topo da cabeça na direção rostral para caudal com lidocaína (analgesia tópica).
- Pressione com firmeza para fazer uma única incisão na linha média com uma lâmina de bisturi limpa.
- Usando uma broca acoplada ao estereotáxico, faça dois furos sobre os bulbos olfatórios para a cânula guia (26 G) usando coordenadas AP +5,0 mm, ML +/- 0,75 mm em relação ao bregma.
- Faça dois furos sobre a formação cerebelar.
NOTA: A localização desses parafusos não precisa ser precisa, certifique-se de que eles sejam simétricos ao longo da linha média. - Coloque os parafusos nos dois orifícios sobre a formação cerebelar e use um adesivo de tecido para prender esses parafusos ao crânio.
- Use o estereotáxico para inserir a cânula guia (26 G) nos orifícios perfurados sobre os bulbos olfatórios (passo 1.6). Abaixe a cânula DV 1,0 mm.
- Misture o cimento dentário em uma placa de Petri. Use uma pequena colher de metal para empilhar lentamente o cimento dental ao redor da cânula. Deixe secar por 5 s. Em seguida, remova os braços do dispositivo estereotáxico, tomando cuidado para não puxar a cânula guia para fora no processo. Continue empilhando o cimento dentário em toda a incisão até formar uma pequena tampa (ver Figura 1A).
- Coloque plugues fictícios na cânula guia neste ponto para evitar o bloqueio.
- Imediatamente após a cirurgia, injete cetoprofeno (0,2 mg/kg) e solução salina (200 μL) para reduzir a dor e reidratar. Forneça comida amolecida ou hidrogel aos camundongos após a cirurgia. Não deixe os camundongos sozinhos até que eles recuperem a consciência para manter a decúbito esternal.
NOTA: Os ratos também são alojados individualmente a partir deste ponto. - Injete solução salina (200 μL) e cetoprofeno (0,2 mg/kg) uma vez ao dia durante dois dias após a cirurgia.
- Por 2 dias e até 5 dias, (conforme necessário) após a cirurgia, pese os camundongos e monitore seus pesos. Se o peso não retornar aos níveis pré-operatórios dentro de dois ou três dias, consulte o veterinário sobre o método apropriado de alimentação.
- Permita que os ratos se recuperem por pelo menos 7 dias antes de iniciar o treinamento comportamental.
2. Tarefa de discriminação associativa
- Infusões
- Administre infusões específicas para OB do inibidor do receptor de tirosina quinase, K252a (50 μM; 5% de DMSO em solução salina) ou veículo (5% de DMSO em solução salina) aos camundongos da seguinte forma.
- Administre a infusão no OB dos camundongos bilateralmente. Injete 2,0 μL de volume final por bulbo com taxa de infusão de 0,2 μL/min e 10 min de tempo total de infusão usando uma bomba injetora dupla.
NOTA: O tempo da manipulação pode ser ajustado dependendo do tipo de estudo comportamental que está sendo realizado. As etapas exatas de infusão são específicas para cada bomba injetora e fornecidas no manual do fabricante. - Certifique-se de deixar os injetores dentro das cânulas por cerca de 5 minutos após o parto para inibir o refluxo e promover a difusão. (ou seja, planeje um total de 15 minutos para cada infusão).
- Administre a infusão no OB dos camundongos bilateralmente. Injete 2,0 μL de volume final por bulbo com taxa de infusão de 0,2 μL/min e 10 min de tempo total de infusão usando uma bomba injetora dupla.
- Administre infusões específicas para OB do inibidor do receptor de tirosina quinase, K252a (50 μM; 5% de DMSO em solução salina) ou veículo (5% de DMSO em solução salina) aos camundongos da seguinte forma.
- Conjuntos de odores.
- Dilua todos os odores em óleo mineral leve a uma pressão parcial de 1,0 Pa usando uma proporção pré-calculada com base na pressão de vapor (Tabela 1).
- Use os 5 pares de odores separados da Tabela 1 (os números na tabela indicam o volume em μL para misturar 50 mL de óleo mineral para 1,0 Pa).
- Para preparar a areia perfumada a ser usada durante as etapas comportamentais (Seção 2.3 e 3), misture 400 μL do odorante 1.0 Pa da Etapa 2.2.2 para cada 100 g de areia de brincar.
- Moldar
NOTA: Os camundongos devem ser submetidos à modelagem por um período de 10 dias, conforme descrito abaixo.- Traga os ratos para a sala de procedimentos e manuseie-os por 10 minutos por dia durante os primeiros dois dias após a recuperação da cirurgia.
- No dia 3, coloque uma placa de Petri cheia de areia perfumada com (+) -limoneno nas gaiolas domésticas dos camundongos e preenchida com cerca de 10 pastilhas de sacarose, cada uma com 5 mg de massa.
NOTA: Use 1.0 Pa (+)-limoneno (misture 102 μL em 50 mL de óleo mineral) como o odor recompensado e óleo mineral puro (o agente diluente para os odores de teste) como o odor não recompensado. A escolha de odores monomoleculares também é recomendada, pois é mais provável que sejam novos para camundongos. - Reabasteça a areia e os pellets no dia 4.
- Nos dias 5 e 6, aclimate os camundongos ao aparato comportamental feito sob medida, colocando-os no aparato e deixando-os explorar o espaço (Figura 1B, C). Faça o aparelho usando uma gaiola doméstica padrão e poli (metacrilato de metila) para construir duas tampas e uma divisória central preta. Certifique-se de que as tampas e a divisória central sejam 1-2 cm maiores que a gaiola doméstica.
- Prepare uma placa de Petri de areia perfumada com limoneno e outra com areia contendo óleo mineral. Após a aclimatação nos dias 5 e 6, coloque os dois pratos de areia na câmara de teste e misture 10 pellets de sacarose no prato com aroma de limoneno para servir como recompensa. Coloque cada rato na câmara de teste por 10 minutos e deixe explorar e consumir livremente os pellets de recompensa.
- No dia 7, apresente aos ratos uma versão resumida do procedimento final de teste, colocando os pratos contendo areia perfumada com limoneno e óleo mineral no aparato comportamental. Desta vez, inclua o divisor central.
- Coloque uma única recompensa em cima da areia com aroma de limoneno e coloque o rato na câmara de descanso.
- Uma vez que o mouse tenha sido colocado na câmara de repouso, levante o divisor central para permitir que o mouse entre na câmara de teste para investigar e cavar os pratos cheios de areia. Retorne o mouse para a câmara de repouso depois de recuperar o projétil de recompensa ou após 5 minutos.
- Repita esse processo para um total de 10 tentativas para cada camundongo individual. Contrabalanceie a colocação do prato recompensado no lado esquerdo ou direito, empregando um gerador de números aleatórios. Não há períodos de descanso entre as tentativas.
- No dia 8, repita as tentativas do dia 7, mas também enterre progressivamente o pellet cada vez mais fundo na areia.
NOTA: A maioria dos ratos deve estar cavando para a pelota de recompensa invisível até o10º teste no dia 8. - No dia 9, aumente o número de tentativas para 20 tentativas para cada camundongo com o enterro profundo completo do pellet de sacarose e introduzindo os camundongos na câmara de teste por 1 minuto por tentativa. Deixe os ratos cavar em ambos os pratos para a recompensa.
- No dia 10, repita as 20 tentativas para cada rato, mas se eles cavarem no prato não recompensado antes de cavar no prato recompensado, comece o próximo teste. Permita que os ratos que primeiro cavaram o prato recompensado (com aroma de limoneno) recuperem o pellet de recompensa antes de enviá-los de volta para a câmara de descanso.
3. Treinamento e testes
NOTA: Uma vez que os ratos tenham começado a cavar de forma confiável para as pelotas de recompensa invisíveis e com odor, o experimento pode começar.
- Formação
NOTA: A fase de treinamento começa dois dias após a conclusão da modelagem e consiste em 20 tentativas para cada mouse. Antes do treino, administre imediatamente as perfusões intrabulbares de fármaco/veículo (ver secção 2.1 para obter detalhes sobre a perfusão) e inicie o treino imediatamente após as perfusão.- Coloque um rato na câmara de descanso.
- Coloque dois pratos de areia perfumados com um novo par de odores na câmara de teste, onde uma pelota de recompensa é enterrada em um dos pratos.
- Quando a câmara de teste estiver pronta, levante a barreira opaca e introduza o mouse na câmara de teste. Retorne imediatamente o mouse para a câmara de repouso, se o mouse cavar primeiro no prato não recompensado (registre essas tentativas como um "0"). Se o rato cavar o odor gratificante primeiro, deixe-o recuperar o pellet e devolvê-lo à câmara de repouso. Registre esses testes como um "1". Se o teste durar 1 minuto sem que o mouse recupere a recompensa, envie o mouse de volta para a câmara de descanso.
- Limpe e reabasteça os pratos e comece o próximo teste. Repita o mesmo para 20 tentativas.
- Teste
NOTA: O teste de memória pode ser realizado em qualquer duração de interesse para o pesquisador. Neste experimento, dois grupos separados de camundongos foram testados 2 h (STM) ou 48 h (LTM) após o treinamento, dado o interesse na influência diferencial de K252a em STM e LTM.- Realizar testes usando os mesmos odores e procedimentos descritos para o treinamento (Seção 3.1).
NOTA: Dependendo da questão de pesquisa, o pesquisador pode precisar incluir grupos de controle. Por exemplo, no experimento publicado anteriormente, os efeitos do bloqueio do receptor Trk na consolidação da memória foram estudados3. Portanto, um grupo de controle foi infundido com K252a antes do teste de 48 horas para mostrar que os efeitos não eram devidos à interferência na recuperação.
- Realizar testes usando os mesmos odores e procedimentos descritos para o treinamento (Seção 3.1).
- Análise estatística
NOTA: Sintaxe do SPSS 22.0 para cada etapa fornecida como um arquivo suplementar como exemplo.- Realize análises estatísticas usando análise linear de efeitos mistos. Ao contrário das ANOVAs, os modelos lineares de efeitos mistos podem explicar melhor os efeitos aleatórios e as medidas repetidas.
- Calcule a medida dependente: "proporção correta". Lembre-se de 3.1.3 que um "1" foi atribuído a testes em que o rato cavou o odor recompensador primeiro, e um "0" se o rato cavou primeiro no odor não recompensador. Média a cada cinco tentativas para criar quatro blocos de tentativa (TB; por exemplo, bloco de tentativa 1 ou TB1 foi a média das tentativas 1-5, bloco de tentativa 2 ou TB 2 foi a média das tentativas 6-10 e assim por diante).
- Defina as variáveis independentes ou efeitos fixos como grupos de medicamentos (K252a ou Veículo; Seção 2.1.1) e blocos de teste (de 3.3.1). Nos resultados representativos abaixo, as variáveis usadas para cada análise são especificadas.
- Inclua o mouse individual e o conjunto de odores aninhados no mouse como "efeitos aleatórios" para compensar as diferenças comportamentais intrínsecas nos camundongos e quaisquer efeitos do uso de vários conjuntos de odores.
- Execute uma transformação logit na proporção correta.
NOTA: A proporção correta não é uma variável dependente contínua e não ligada. Assim, viola duas suposições para modelos lineares. Portanto, a transformação logit é executada. - Usar médias marginais estimadas para realizar testes post hoc em interações significativas identificadas pelo modelo completo; Múltiplas comparações de pares devem ser corrigidas em testes post hoc. Bonferroni ou Šidák são normalmente usados.
Resultados
Conforme descrito, este protocolo permite que os pesquisadores avaliem a influência de alguma manipulação na aprendizagem, STM e LTM. Os resultados da amostra de Tong et al, 20183 são apresentados aqui. Os resultados apóiam a hipótese de que o bloqueio do receptor Trk inibe seletivamente o LTM, mas não o aprendizado ou o STM.
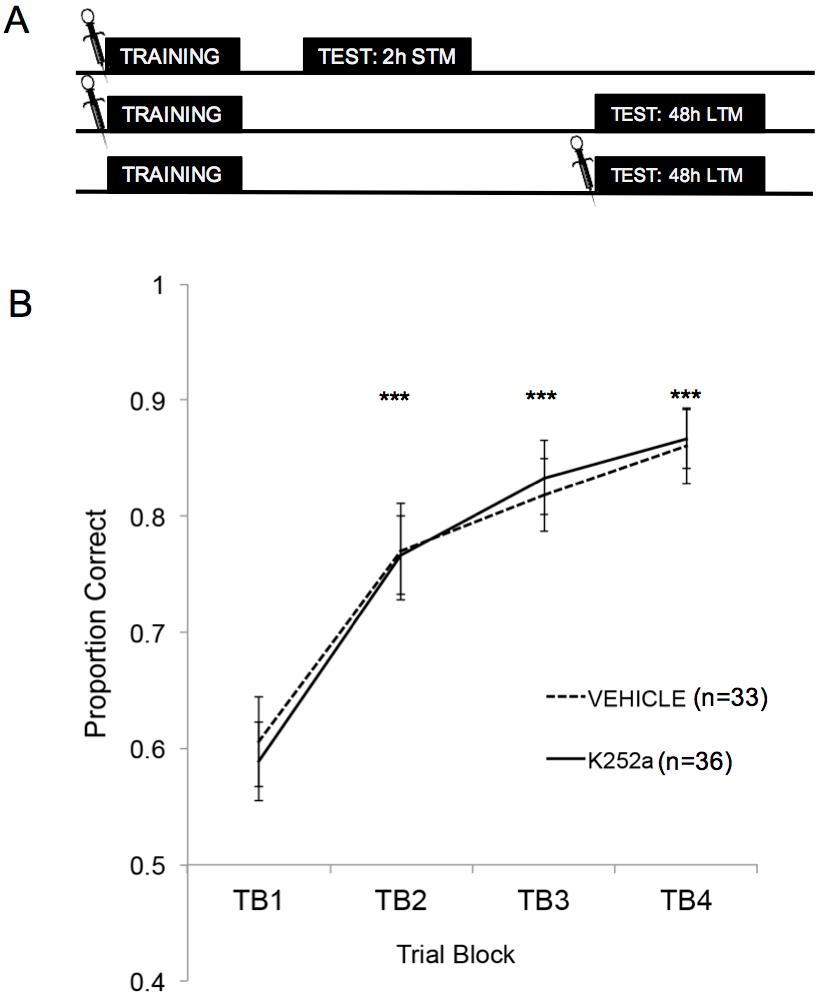
A Figura 2A mostra os esquemas de treinamento, teste STM e teste LTM. Primeiro, foi demonstrado que as infusões de K252a não afetaram a taxa de aprendizado de uma associação odor-recompensa. A Figura 2B (Seção 3.1) mostra a taxa de aprendizado dos grupos K252a e de veículos do Treinamento. A análise estatística usando um modelo linear misto foi executada com dois efeitos fixos, grupo de drogas e bloco experimental (TB); rato e conjunto de odor aninhados dentro do rato eram efeitos aleatórios. Os dados do teste (Seção 3.2) não foram incluídos na análise. Um efeito principal significativo foi observado no bloco experimental (F (3, 183,692) = 43,735, p < 0,001), mas nenhum efeito do grupo de drogas (F (1, 85,685) = 0,132, p = 0,717) e nenhuma interação significativa (F (3, 183,692) = 0,111, p = 0,954). Os testes post hoc, utilizando o ajuste de Šidák, confirmaram que os grupos K252a e veículo não diferiram em nenhum dos blocos de ensaio durante a formação (p > 0,05 para todas as comparações). TB2, TB3 e TB4 foram significativamente maiores do que TB1 para todas as comparações (p ≤ 0,001 em todos os casos), demonstrando que ambos os grupos aprenderam com sucesso a associação odor-recompensa ao final de 20 tentativas de treinamento.
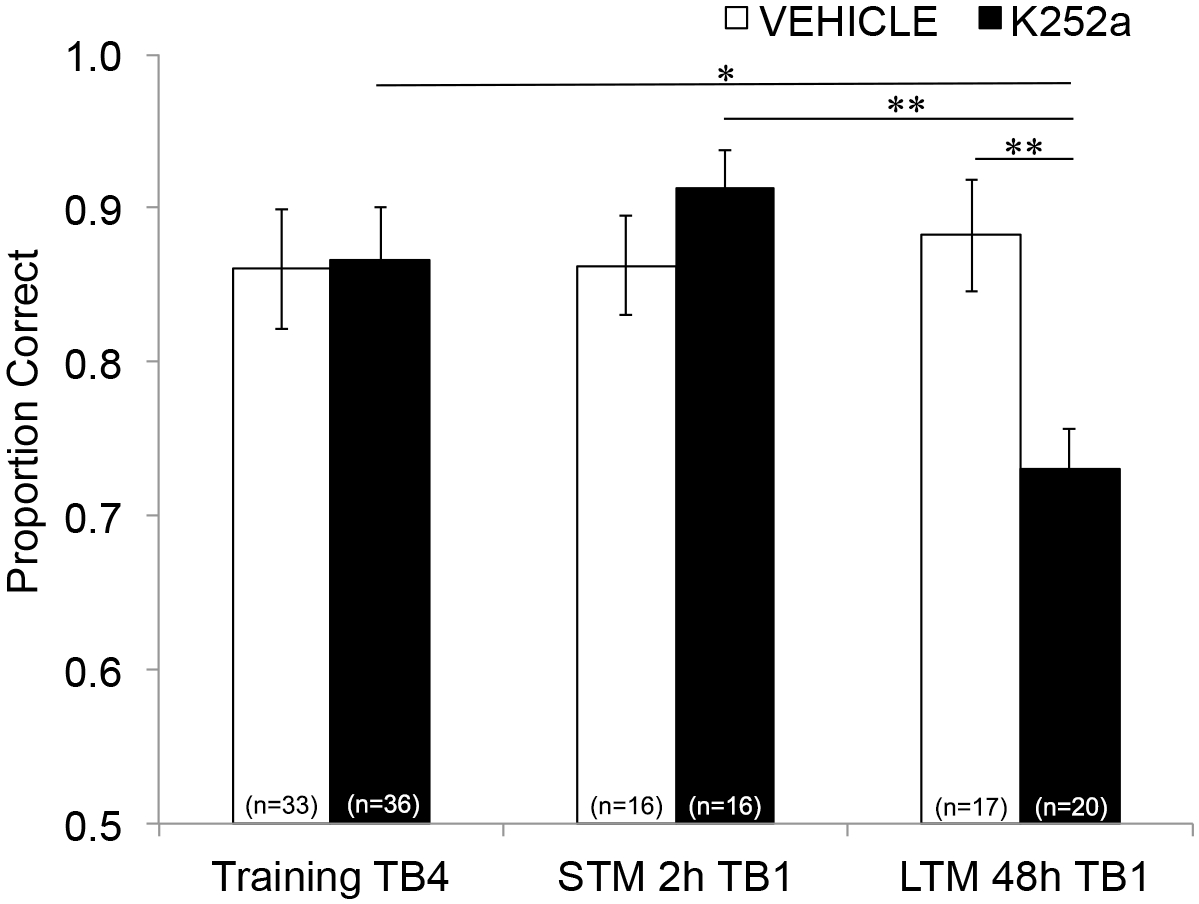
Em seguida, para examinar os efeitos da infusão no STM e LTM, a mesma análise foi executada e incluiu dados do Teste (Seção 3.2). Mostrou uma interação significativa entre o grupo de medicamentos e o bloco de ensaio (F(2, 77,558) = 4,043, p = 0,021), sem efeitos principais significativos do grupo de medicamentos (F(1, 55,629) = 1,438, p = 0,236) ou bloqueio de ensaio (F(2, 69,979) = 1,360, p = 0,263). A fim de examinar especificamente a memória, comparações post hoc em pares com correção de Šidák, compararam as diferenças entre o último bloco de teste de Treinamento (Seção 3.1) e o primeiro bloco de teste de Teste (Seção 3.2) 2 (STM) ou 48 horas (LTM) depois. Para camundongos com infusão de veículo, as comparações mostraram retenção da memória associativa em 2 e 48 h após o treinamento (p > 0,05 para todas as comparações com o desempenho do treinamento). Para camundongos com infusão de K252a, o primeiro bloco experimental do teste de 2 horas (STM) não diferiu do último bloco experimental de treinamento (p > 0,05); no entanto, seu desempenho de memória foi significativamente menor após 48 h (p = 0,018). Além disso, a memória no teste de 48 horas foi significativamente reduzida em comparação com a memória no teste de 2 horas (p = 0,009) e com o desempenho do grupo veículo no teste de 48 horas (p = 0,006). Não houve diferença no STM entre camundongos infundidos com veículo e K252a (p = 0,356). Juntos, os resultados mostram que a inibição de K252a dos receptores Trk no bulbo olfatório interrompe seletivamente a memória de odor de longo prazo, mas não de curto prazo (Figura 3).
| Conjunto de odores | Odor 1 | Odor 2 |
| 1 | ácido pentanóico | ácido butanóico |
| 225.1 | 63.6 | |
| 2 | hexanal | heptanal |
| 11.1 | 35.3 | |
| 3 | acetato de propila | acetato de butilo |
| 3.1 | 10.9 | |
| 4 | 2-octanona | 2-heptanona |
| 87.4 | 28.7 | |
| 5 | pentanol | hexanol |
| 37.2 | 127.3 |
Tabela 1: Volumes de mistura para conjuntos de odores. Cada linha mostra dois odores que podem ser usados como um par para as etapas comportamentais. Por exemplo, para usar o primeiro "conjunto de odores", faça as misturas de ácido pentanóico e butanóico. Os números na tabela indicam volume em μL para misturar 50 mL de óleo mineral para concentração de 1,0 Pa de cada odor. Durante o treinamento e o teste, uma placa de Petri seria perfumada com ácido pentanóico, a outra com ácido butanóico.

Figura 1: Colocação da canulação e aparelho comportamental. (A) Mostra a posição relativa da cânula, tampa de cimento dentário e parafusos na cabeça do mouse. Observe que as agulhas da cânula alcançam os dois bulbos olfatórios, o próprio pedestal é embutido na tampa do cimento dentário. Os parafusos são colocados em dois orifícios perfurados no crânio sobre a formação cerebelar. Os parafusos não tocam o cérebro em si, mas atuam como uma âncora caudal para a capa de cimento dental. A figura mostra o tamanho relativo para fazer a tampa de cimento. (B) Mostra o aparato comportamental montado. O corpo é uma gaiola típica de rato. As tampas com orifícios para ar eram feitas de plexiglass. O divisor central também é feito de plexiglass preto. As tampas devem ser feitas de plexiglass pesado o suficiente para que possam atuar como um trato para que o divisor central seja levantado e colocado durante os testes. (C) Mostra um lado do aparato comportamental. Observe que o divisor central é um pouco mais alto que a gaiola para facilitar o levantamento. Placas de Petri de areia podem ser colocadas ao longo da borda. Clique aqui para ver uma versão maior desta figura.
{kind=link}

Figura 2: Desenho do estudo e resultados de aprendizagem. (A) Mostra o esquema do desenho do estudo. Observe que os grupos STM e LTM eram independentes (ou seja, diferentes grupos de camundongos). Os símbolos no início dos blocos indicam o momento em que as infusões foram administradas. (B) Mostra a proporção correta para as Provas 1-20 durante o Treinamento. Os resultados indicam que os grupos K252a e veículo não diferiram em sua taxa de aprendizado (inclinação das linhas). As barras de erro representaram o erro padrão da média (EPM). Os asteriscos mostram aumentos significativos na proporção correta em comparação com TB1 para as coortes de veículos e K252a (p ≤ 0,001 para todas as comparações). Esta figura é adaptada de Tong et al. 2018 com permissão3. Clique aqui para ver uma versão maior desta figura.
{kind=link}

Figura 3: Efeitos diferenciais do bloqueio do receptor Trk no STM e LTM. Mostra a proporção correta para o Bloco de Teste 4 de Treinamento e o Bloco de Teste 1 do Teste STM e LTM. Ou seja, um modelo linear misto é usado para comparar a proporção correta durante o último bloco de tentativa da fase de treinamento (Figura 2; Treinamento-TB4) para aqueles durante os primeiros blocos de teste (Teste-TB1) de teste de memória de curto prazo (teste de 2 horas) e longo prazo (teste de 48 horas). O modelo linear misto teve dois efeitos fixos: grupo de drogas e bloco de tentativa (Training-TB4, STM-TB1, LTM-TB1). Os efeitos aleatórios foram mouse e odor set aninhado dentro do mouse. Comparações post hoc descobriram que os camundongos K252a tinham LTM significativamente prejudicado (comparação com o treinamento TB4; p = 0,018), mas não STM (p > 0,05). O desempenho do LTM por camundongos com infusão de K252a também foi significativamente menor do que o STM de camundongos com infusão de K252a (p = 0,009) e menor do que o LTM de camundongos veículo (p = 0,006). As barras de erro representavam o SEM. Esta figura é adaptada de Tong et al. 2018 com permissão3. Clique aqui para ver uma versão maior desta figura.
{kind=link}

Arquivo Suplementar: Sintaxe usada para a análise estatística. Clique aqui para baixar este arquivo.
Discussão
O condicionamento do medo de uma tentativa é um poderoso protocolo comportamental para estudar a dinâmica molecular e celular da memória, mas grande parte do aprendizado natural é incremental e melhor modelado por meio de um paradigma como o descrito acima. A inibição dos receptores Trk no BO impediu a consolidação da memória olfativa em um paradigma de aprendizagem apetitiva multi-tentativa, como mostrado anteriormente por nosso grupo3. A descoberta abre novos caminhos para a pesquisa sobre o tempo diferencial de mecanismos moleculares, como neurotrofinas, na aprendizagem apetitiva e aversiva.
Este experimento consistiu em duas partes críticas: (1) a canulação e (2) a tarefa de discriminação associativa (subdividida em modelagem, treinamento e teste). Os experimentadores podem adaptar este protocolo à sua questão de pesquisa específica. Por exemplo, estávamos interessados principalmente no OB e este protocolo estabelecido pode ser facilmente aplicado a outros estudos de OB. Para experimentadores com outras regiões de interesse, será importante validar os locais de infusão em um estudo piloto. Os experimentadores também podem precisar considerar a taxa de difusão, a penetração espacial e a duração da bioatividade do que estão infundindo.
As etapas de modelagem descritas no protocolo foram amplamente utilizadas pelos autores deste protocolo. Aderir a eles conforme descrito parece ser importante para que os ratos aprendam a tarefa em tempo hábil. Usando outras linhas do tempo, os autores observaram mais variação entre os camundongos em sua familiaridade com a tarefa e isso significou treinamento adicional para levar todos os camundongos a um critério para testes com odores experimentais. Para treinamento e teste, o pesquisador tem flexibilidade, dependendo de seus interesses de pesquisa, com o número de testes, a concentração dos odores experimentais e a semelhança dos odores entre si. Recomendamos o uso de vários conjuntos de odores quando possível, como descrevemos, a fim de reduzir o número de animais usados para o experimento. Consulte a seção Análise Estatística para obter orientações sobre como considerar o uso de vários conjuntos de ordens na análise final. Em princípio, a semelhança do par de odores pode ser variada para ajustar a dificuldade da discriminação. No estudo publicado anteriormente do nosso grupo3, os pares de odores consistem em dois odores do mesmo grupo funcional, mas diferiam um do outro por um comprimento de carbono. Essas discriminações são mais difíceis do que pares que diferem em dois ou mais comprimentos de carbono, mas mais fáceis do que enantiômeros (por exemplo, (+)-limoneno e (-)-limoneno). Odores de diferentes grupos funcionais são altamente diferentes perceptivamente. Cleland et al13 discutem mais variações de estímulos e seus efeitos em parâmetros específicos de aprendizagem.
Uma grande limitação deste protocolo é que leva muito mais tempo para ser realizado em comparação com tarefas automatizadas de aprendizado associativo, onde vários animais podem ser testados em paralelo. Para um determinado teste, um pesquisador levaria pelo menos 20 minutos para completar 20 tentativas para um camundongo. No entanto, é essa falta de automação que significa que o protocolo é mais acessível financeiramente, uma prioridade para muitas instituições. É importante ressaltar que, no caso deste experimento, descobriu-se que este protocolo é altamente tratável e eficaz para o treinamento de pesquisadores de graduação com interesses em neurociência comportamental. Em particular, esses alunos desenvolvem fortes habilidades de manejo de animais, além dos benefícios usuais da participação em pesquisas.
Os pesquisadores interessados em adotar esse paradigma podem variar vários parâmetros. Aparentemente, as manipulações farmacológicas dos mecanismos são diversas, e esse protocolo comportamental pode ser usado com técnicas quimiogenéticas ou várias outras formas de manipular vias moleculares e celulares (por exemplo, optogenética). O próprio paradigma pode ser ajustado para variar o tipo de aprendizado e memória testados. Por exemplo, os pesquisadores podem ajustar a semelhança dos dois odores apresentados para controlar a taxa de aprendizado. Em nosso estudo3, os pares de odores consistem em dois odores do mesmo grupo funcional, mas diferiam um do outro por um comprimento de carbono. Essas discriminações são mais difíceis do que pares que diferem em dois ou mais comprimentos de carbono, mas mais fáceis do que os enantiômeros (por exemplo, (+)-limoneno e (-)-limoneno). Odores de diferentes grupos funcionais são altamente diferentes perceptivamente. Cleland ecols.13 discutem mais variações de estímulos e seus efeitos em parâmetros específicos de aprendizagem, concluindo que odores mais semelhantes são mais difíceis de discriminar e, portanto, levam mais tempo para seremaprendidos14. Essas manipulações também afetariam a força das memórias. Nesse sentido, os pesquisadores podem estar interessados em testar a memória em diferentes momentos após o aprendizado. Por exemplo, dois estudos 15,16 examinaram o papel do BDNF na persistência do LTM para uma tarefa de aprendizagem aversiva de uma tentativa. BDNF é um ligante de TrkB. Os estudos mostraram que a infusão de oligonucleotídeos anti-BDNF antisense no hipocampo 12 horas após o aprendizado bloqueou o LTM 7 dias depois, mas não 2 dias depois. Este estudo mostra que a escala de tempo dos mecanismos moleculares após o aprendizado inicial desempenha papéis interessantes e ainda a serem compreendidos no LTM. Este artigo descreve um protocolo comportamental que permitiria a investigação dessas escalas de tempo. Outros parâmetros de interesse para aplicação futura incluem o modelo de camundongo utilizado. Por exemplo, seria interessante substituir camundongos machos (que têm uma neurobiologia significativamente melhor estudada) 17 por camundongos fêmeas em estudos futuros para examinar variações na taxa de aprendizado, STM e LTM, já que as fêmeas de mamíferos têm maior sensibilidade e seletividade a odores do que os machos18. Obviamente, modelos de doenças de roedores também podem ser usados de forma eficaz com este protocolo.
Divulgações
Os autores deste artigo não tinham interesses financeiros concorrentes.
Agradecimentos
O projeto foi apoiado pelo Scantland Summer Collaborative Research Gift e pelo Stephen and Sylvia Tregidga Burges Endowed Research Fund. Os autores gostariam de agradecer aos colegas do Departamento de Psicologia e do Programa de Neurociência do Earlham College por seu apoio e orientação.
Materiais
| Name | Company | Catalog Number | Comments |
| Double guide cannula | PlasticsOne | C235GS-5-1.5/SPC | Custom order |
| (-)-limonene | Sigma-Aldrich | 218367-50G | |
| (+)-limonene | Sigma-Aldrich | 183164-100ML | |
| 2-hetanone | Sigma-Aldrich | 537683 | |
| 2-octanone | Sigma-Aldrich | O4709 | |
| 5mg sucrose pellets | Test Diet | 1811560 | Custom size. Used for rewards |
| Butanoic acid | Sigma-Aldrich | B103500 | |
| butyl acetate | Sigma-Aldrich | 402842 | |
| Dental Cement Powder (Coral) | A-M Systems | 525000 | |
| Dental Cement Solvent | A-M Systems | 526000 | |
| Double connector assembly | PlasticsOne | C232C | |
| Double dummy cannula | PlasticsOne | C235DCS-5/SPC dummy dbl | Custom order |
| Double injector | PlasticsOne | C235IS-5/SPC | Custom order |
| Drill | Kopf Instruments | Model 1474 High Speed Stereotaxic Drill | This drill requires an additional "adaptor" piece in order to fit certain drill bits. We get by this problem by wrapping the drill bit with lab tape to increase the circumference of the drill it to fit. This may not be an option for surgeries requiring sterile technique. |
| Eye Ointment | Purchase from local pharmacy | ||
| Figure 1 illustration software | BioRender | ||
| heptanal | Sigma-Aldrich | W254002 | |
| hexanal | Sigma-Aldrich | 115606 | |
| hexanol | Sigma-Aldrich | H13303 | |
| Infusion pump model 11 | Harvard Apparatus | 4169D | Used pumps available via American Instrument Exchange |
| Isoflurane | Santa Cruz Animal Health | sc-363629Rx | Vet prescription needed for order |
| K252a | Sigma-Aldrich | K2015 | Mixed to 50uM in DMSO (5%) |
| Ketoprofen | Allivet | 25920 | Vet prescription needed for order |
| Lidocaine | Aspercreme | Purchased from Amazon | |
| Mounting Screws | PlasticsOne | 00-96 X 3/32 | |
| Mouse Anesthesia Mask | Kopf Instruments | Model 907 Mouse Anesthesia Mask | Used with the stereotaxic to allow oxygen and anesthesia while mouse in stereotax |
| Mouse Nose Adaptor | Kopf Instruments | Model 926 Mouse Adaptor | Used with the stereotaxic to allow for head of mouse to be secured. |
| Novalsan | Jeffers | 41375 | |
| Pentanoic acid | Sigma-Aldrich | 240370 | |
| pentanol | Sigma-Aldrich | 138975 | |
| Petri dish glass bottoms | VWR | 10754-804 | |
| Polycarbonate Café bottoms | Ancare | N10PCSEC | Use normal housing cages and custom fit a track in the middle to act as the track for an opaque plexiglass divider |
| propyl acetate | Sigma-Aldrich | 537438 | |
| Quikrete Premium Play Sand | Purchase from local hardware store | ||
| Saline | Insight Needles | N/A | Sterile saline for drug mixing |
| Stereotaxic apparatus | Kopf Instruments | Model 902 Small Animal Stereotaxic Instrument | |
| Testing chamber | Ancare | N10PCSEC | Our testing chambers are modified using the regular mouse housing cage. The manuscript details what was done. |
| Vetbond Tissue Adhesive | 3M | Purchased from Amazon |
Referências
- Grimes, M. T., Smith, M., Li, X., Darby-King, A., Harley, C. W., Mclean, J. H. Mammalian Intermediate-Term Memory: New Findings in Neonate Rat. Neurobiology of Learning and Memory. 95 (3), 385-391 (2011).
- Grimes, M. T., Harley, C. W., Darby-King, A., Mclean, J. H. PKA Increases in the Olfactory Bulb Act as Unconditioned Stimuli and Provide Evidence for Parallel Memory Systems: Pairing Odor with Increased PKA Creates Intermediate- and Long-Term, but not Short-Term, Memories. Learning and Memory. 19 (3), 107-115 (2012).
- Tong, M. T., Kim, T. Y. P., Cleland, T. A. Kinase Activity in the Olfactory Bulb is Required for Odor Memory Consolidation. Learning & Memory. 25 (5), 198-205 (2018).
- Devore, S., Lee, J., Linster, C. Odor Preferences Shape Discrimination Learning in Rats. Behavioral Neuroscience. 127 (4), 498-504 (2013).
- Gao, Y., Strowbridge, B. W. Long-Term Plasticity of Excitatory Inputs to Granule Cells in the Rat Olfactory Bulb. Nature Neuroscience. 12 (6), 731-733 (2009).
- Bath, K. G., Mandairon, N., et al. Variant Brain-Derived Neurotrophic Factor (Val66Met) Alters Adult Olfactory Bulb Neurogenesis and Spontaneous Olfactory Discrimination. Journal of Neuroscience. 28 (10), 2383-2393 (2008).
- Lazarini, F., Lledo, P. M. Is Adult Neurogenesis Essential for Olfaction. Trends in Neurosciences. 34 (1), 20-30 (2011).
- Mandairon, N., Peace, S., Karnow, A., Kim, J., Ennis, M., Linster, C. Noradrenergic Modulation in the Olfactory Bulb Influences Spontaneous and Reward-Motivated Discrimination, but not the Formation of Habituation Memory. European Journal of Neuroscience. 27 (5), 1210-1219 (2008).
- Guérin, D., Peace, S. T., Didier, A., Linster, C., Cleland, T. A. Noradrenergic Neuromodulation in the Olfactory Bulb Modulates Odor Habituation and Spontaneous Discrimination. Behavioral Neuroscience. 122 (4), 816-826 (2008).
- Moreno, M. M., Bath, K., Kuczewski, N., Sacquet, J., Didier, A., Mandairon, N. Action of the Noradrenergic System on Adult-Born Cells Is Required for Olfactory Learning in Mice. Journal of Neuroscience. 32 (11), 3748-3758 (2012).
- Vinera, J., Kermen, F., Sacquet, J., Didier, A., Mandairon, N., Richard, M. Olfactory Perceptual Learning Requires Action of Noradrenaline in the Olfactory Bulb: Comparison with Olfactory Associative Learning. Learning and Memory. 22 (3), 192-196 (2015).
- Dillon, T. S., Fox, L. C., Han, C., Linster, C. 17β-estradiol Enhances Memory Duration in the Main Olfactory Bulb in CD-1 Mice. Behavioral Neuroscience. 127 (6), 923 (2013).
- Cleland, T. A., Narla, V. A., Boudadi, K. Multiple Learning Parameters Differentially Regulate Olfactory Generalization. Behavioral Neuroscience. 123 (1), 26 (2009).
- Cleland, T. A., Morse, A., Yue, E. L., Linster, C. Behavioral Models of Odor Similarity. Behavioral Neuroscience. 116 (2), 222-231 (2002).
- Bekinschtein, P., Cammarota, M., Igaz, L., Bevilaqua, L., Izquierdo, I., Medina, J. Persistence of Long-Term Memory Storage Requires a Late Protein Synthesis- and BDNF- Dependent Phase in the Hippocampus. Neuron. 53 (2), 261-277 (2007).
- Bekinschtein, P., et al. BDNF is Essential to Promote Persistence of Long-Term Memory Storage. Proceedings of the National Academy of Sciences. 105 (7), 2711-2716 (2008).
- Andreano, J., Cahill, L. Sex influences on the neurobiology of learning and memory. Learning and Memory. 16 (2009), 248-266 (2009).
- Kass, M. D., Czarnecki, L. A., Moberly, A. H., Mcgann, J. P. Differences in Peripheral Sensory Input to the Olfactory Bulb Between Male and Female Mice. Scientific Reports. 7 (1), (2017).
Reimpressões e Permissões
Solicitar permissão para reutilizar o texto ou figuras deste artigo JoVE
Solicitar PermissãoExplore Mais Artigos
This article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. Todos os direitos reservados