
Method Article
Недорогая задача на ассоциацию «запах-вознаграждение» для тестов на обучение и память
В этой статье
Резюме
Задача ассоциативного обучения «запах-вознаграждение» была использована для исследования дифференциального влияния физиологических манипуляций на долговременную и кратковременную память.
Аннотация
Надежные и простые поведенческие парадигмы аппетитной, ассоциативной памяти имеют решающее значение для исследователей, интересующихся клеточными и молекулярными механизмами памяти. В данной работе описан эффективный и недорогой поведенческий протокол мышей для изучения влияния физиологических манипуляций (таких как инфузия фармакологических агентов) на скорость обучения и продолжительность памяти обоняния-вознаграждения. Репрезентативные результаты получены из исследования, изучающего дифференциальную роль активности рецепторов тирозинкиназы в кратковременной (СТМ) и долговременной памяти (ДЛМ). Самцы мышей были приучены ассоциировать награду (сахарную гранулу) с одним из двух запахов, и их память на эту связь была проверена через 2 или 48 часов. Непосредственно перед тренировкой в обонятельную луковицу (OB) вводили ингибитор рецептора тирозинкиназы (Trk) или инфузии носителей. Несмотря на отсутствие влияния инфузии на скорость обучения, блокада рецепторов Trk в OB избирательно нарушала LTM (48 ч), а не кратковременную память (STM; 2 ч). Ухудшение LTM было связано с уменьшением избирательности запаха, измеренной по продолжительности копания. Кульминация результатов этого эксперимента показала, что активация Trk-рецептора в OB является ключом к консолидации обонятельной памяти.
Введение
Механизмы формирования ассоциативной памяти ранее исследовались преимущественно на основе исследований условного рефлекса страха в одном попытке. Тем не менее, многие повседневные задачи обычно имеют более сложные схемы получения информации и зависят от повторяющихся встреч. Цель этого протокола состоит в том, чтобы обеспечить экономически эффективную поведенческую парадигму грызунов, которая используется для понимания клеточных и молекулярных механизмов многократного обучения и памяти.
Изучение запаха, которое зависит от главной обонятельной луковицы (OB), дает несколько преимуществ для изучения аппетитной памяти в нескольких испытаниях. Во-первых, OB-зависимые воспоминания имеют различную продолжительность (STM, LTM и промежуточная память1) и опираются на тот же молекулярный 2,3 и структурный механизм, что и другие элементы мозга, включая нейромодуляцию4, долгосрочное потенцирование5 и взрослый нейрогенез 6,7,8. Во-вторых, в отличие от областей более высокого порядка, таких как гиппокамп, OB-зависимые воспоминания позволяют наблюдать более прямое соответствие между манипуляциями экспериментатора с перцептивной средой и изменениями в нейронных схемах, ответственныхза обучение.. В данной статье подробно описана OB-зависимая парадигма ассоциативного обучения и памяти, которая может быть использована для изучения общих молекулярных и структурных механизмов. Он был разработан, чтобы позволить исследователям получить доступ к преимуществам обонятельного обучения для изучения клеточных и молекулярных механизмов памяти.
В нашей недавней публикации3 описанный здесь протокол был использован для демонстрации того, что консолидация аппетитного обучения запаху зависит от активации рецептора Trk в OB. В приведенном ниже протоколе также обсуждаются области, в которых поведенческая парадигма может быть скорректирована в соответствии с различными экспериментальными потребностями.
В общей сложности в этом исследовании было использовано 27 взрослых самцов мышей CD-1, возраст которых на момент канюляции составлял 8 недель. Точные распределения групп и использование набора запахов см. в разделе «Методы» нашей предыдущей публикации3. Самцы мышей использовались для того, чтобы избежать больших колебаний уровня эстрогена, потому что предыдущие исследованияпоказали , что обонятельная память усиливается при повышении уровня эстрогена. Эти мыши всегда содержались в обратном цикле свет/темнота в течение 12:12 часов и имели доступ к воде. Во время поведенческих экспериментов рацион мышей был ограничен для поддержания их на уровне ~90% от их свободного пищевого веса. Ограничение диеты начинали за 3 дня до начала поведенческого эксперимента. Как будет описано ниже, одному и тому же набору мышей представлены разные наборы запахов, чтобы достичь соответствующих уровней статистической мощности при минимизации использования животными. В разделе статистического анализа показано, как учесть случайную дисперсию, которая может быть вызвана этим.
протокол
Приведенный ниже протокол соответствует рекомендациям IACUC по уходу за животными в колледже Эрлхэм.
1. Канюляция обонятельной луковицы
ПРИМЕЧАНИЕ: Эти операции не требуют стерильной техники, так как они не требуют больших разрезов. Однако каждое заведение может отличаться в своих требованиях. Если экспериментаторы проводят эту операцию на линиях мышей с ослабленным иммунитетом, могут потребоваться дополнительные соображения. В целом, экспериментаторам рекомендуется обсудить этот протокол со своим ветеринаром и командой по уходу за животными перед использованием, а также очищать и дезинфицировать все инструменты между каждой операцией.
- Во время установки замочите канюли и винты в небольшой стакане или чашке Петри, содержащей 32% хлоргексидина, чтобы они были продезинфицированы.
- Обезболите мышей газообразным 4% изофлураном в чистом кислороде и закрепите в стереотаксическом аппарате. Во время операции следите за тем, чтобы мыши находились под давлением 1,5-2% изофлурановой анестезии, вводимой через носовой конус. Контролируйте дыхание на протяжении всей операции. Используйте глазную мазь, чтобы предотвратить сухость глаз во время пребывания под анестезией.
- После того, как мышь закрепится и больше не реагирует на сильное ущипывание задней лапы, используйте 32% хлоргексидин для растирания верхней части головы, чтобы очистить поверхность разреза.
- Далее растирают макушку головы в ростральном к каудальном направлении лидокаином (местная анальгезия).
- Сильно надавите, чтобы сделать один разрез по средней линии чистым лезвием скальпеля.
- С помощью дрели, прикрепленной к стереотаксику, просверлите два отверстия над обонятельными луковицами для направляющей канюли (26 G) с координатами AP +5,0 мм, ML +/-0,75 мм по отношению к брегме.
- Просверлите два отверстия над образованием мозжечка.
ПРИМЕЧАНИЕ: Расположение этих винтов не обязательно должно быть точным, убедитесь, что они симметричны вдоль средней линии. - Вставьте винты в два отверстия над образованием мозжечка и используйте тканевый клей, чтобы прикрепить эти винты к черепу.
- С помощью стереотаксика вставьте направляющую канюлю (26 G) в отверстия, просверленные над обонятельными луковицами (шаг 1.6). Опустите канюлю DV на 1,0 мм.
- Смешайте стоматологический цемент в чашке Петри. С помощью небольшого металлического совка медленно нанесите стоматологический цемент вокруг канюли. Дайте ему высохнуть в течение 5 секунд. Затем снимите рычаги стереотаксического устройства, стараясь при этом не вытащить направляющую канюлю. Продолжайте насыпать зубной цемент на весь разрез до тех пор, пока не образуется небольшой колпачок (см. рисунок 1A).
- В этот момент вставьте фиктивные заглушки в направляющую канюлю, чтобы предотвратить закупорку.
- Сразу после операции введите кетопрофен (0,2 мг/кг) и физиологический раствор (200 мкл) для уменьшения боли и регидратации. Дайте мышам размягченную пищу или гидрогель после операции. Не оставляйте мышей без присмотра, пока они не придут в сознание, чтобы сохранить лежачее положение за грудиной.
ПРИМЕЧАНИЕ: С этого момента мыши также размещаются поодиночке. - Вводите физиологический раствор (200 μл) и кетопрофен (0,2 мг/кг) один раз в день в течение двух дней после операции.
- В течение 2 дней и до 5 дней (по мере необходимости) после операции взвешивайте мышей и следите за их весом. Если вес не возвращается к дооперационному уровню в течение двух-трех дней, проконсультируйтесь с ветеринаром о подходящем способе кормления.
- Дайте мышам восстановиться не менее 7 дней до начала поведенческой тренировки.
2. Задача ассоциативного различения
- Инфузий
- Вводите мышам специфичные для OB инфузии ингибитора рецептора тирозинкиназы, K252a (50 мкМ; 5% ДМСО в физрастворе) или носителя (5% ДМСО в физрастворе) следующим образом.
- Вводите инфузию в акушерский кабинет мышей двусторонне. Введите 2,0 мкл конечного объема на каждую луковицу со скоростью инфузии 0,2 мкл/мин и общим временем инфузии 10 мин с помощью насоса с двумя инжекторами.
ПРИМЕЧАНИЕ: Время проведения манипуляции может быть скорректировано в зависимости от типа проводимого поведенческого исследования. Точные этапы инфузии специфичны для каждого инжекторного насоса и приведены в руководстве производителя. - Обязательно оставьте инжекторы внутри канюль примерно на 5 минут после доставки, чтобы подавить обратный поток и способствовать диффузии. (т.е. планируйте в общей сложности 15 минут на каждую инфузию).
- Вводите инфузию в акушерский кабинет мышей двусторонне. Введите 2,0 мкл конечного объема на каждую луковицу со скоростью инфузии 0,2 мкл/мин и общим временем инфузии 10 мин с помощью насоса с двумя инжекторами.
- Вводите мышам специфичные для OB инфузии ингибитора рецептора тирозинкиназы, K252a (50 мкМ; 5% ДМСО в физрастворе) или носителя (5% ДМСО в физрастворе) следующим образом.
- Наборы запахов.
- Разбавьте все одоранты в легком минеральном масле до парциального давления 1,0 Па с использованием заранее рассчитанного соотношения, основанного на давлении паров (табл. 1).
- Используйте 5 отдельных пар запахов из таблицы 1 (цифры в таблице указывают объем в мкл для смешивания с 50 мл минерального масла на 1,0 Па).
- Чтобы приготовить ароматизированный песок для использования во время поведенческих этапов (разделы 2.3 и 3), смешайте 400 μл одоранта 1,0 Па из шага 2.2.2 на каждые 100 г игрового песка.
- Формирование
ПРИМЕЧАНИЕ: Мыши должны подвергаться формовке в течение 10 дней, как описано ниже.- Приведите мышей в процедурный кабинет и обрабатывайте их по 10 минут в день в течение первых двух дней после восстановления после операции.
- На 3-й день поместите в домашние клетки мышей чашку Петри, наполненную песком с ароматом (+)-лимонена, и наполните ее примерно 10 гранулами сахарозы, каждая массой 5 мг.
ПРИМЕЧАНИЕ: Используйте 1,0 Па (+)-лимонен (смешайте 102 мкл с 50 мл минерального масла) в качестве вознаграждаемого запаха и простое минеральное масло (разбавитель для тестовых запахов) в качестве невознаграждаемого запаха. Также рекомендуется выбирать мономолекулярные одоранты, поскольку они, скорее всего, будут новыми для мышей. - Пополните и песок, и гранулы на 4-й день.
- На 5-й и 6-й дни приучите мышей к специально изготовленному поведенческому аппарату, поместив их в аппарат и позволив им исследовать пространство (рис. 1B,C). Изготовьте аппарат с использованием стандартной домашней клетки, а из полиэтилена (метилметакрилата) соорудите две крышки и черную центральную перегородку. Убедитесь, что крышки и центральная перегородка на 1-2 см больше, чем домашняя клетка.
- Приготовьте одну чашку Петри из песка с ароматом лимонена, а другую — из песка, содержащего минеральное масло. После акклиматизации на 5-й и 6-й день поместите обе посуды с песком в испытательную камеру и смешайте 10 гранул сахарозы с посудой с ароматом лимонена, чтобы она послужила наградой. Поместите каждую мышь в тестовую камеру на 10 минут и дайте ей свободно исследовать и съесть гранулы награды.
- На 7-й день познакомьте мышей с сокращенной версией заключительной процедуры тестирования, поместив в поведенческий аппарат чашки, содержащие аромат лимонена и песок с ароматом минерального масла. На этот раз включают в себя центральный разделитель.
- Положите одну награду на песок с ароматом лимонена и поместите мышь в комнату для отдыха.
- После того, как мышь будет помещена в камеру отдыха, поднимите центральную перегородку, чтобы позволить мыши войти в испытательную камеру для исследования и копания в заполненной песком посуде. Верните мышь в камеру отдыха либо после того, как она достанет гранулу награды, либо по истечении 5 минут.
- Повторите этот процесс в общей сложности 10 попыток для каждой отдельной мыши. Уравновесьте размещение вознаграждаемого блюда слева или справа, используя генератор случайных чисел. Между испытаниями нет периодов отдыха.
- На 8-й день повторите испытания 7-го дня, но также постепенно закапывайте гранулу все глубже и глубже в песок.
ПРИМЕЧАНИЕ: Большинство мышей должны начать искать невидимые гранулы вознаграждения к10-му испытанию на 8-й день. - На 9-й день увеличьте количество попыток до 20 для каждой мыши с полным глубоким закапыванием гранул сахарозы и введением мышей в испытательную камеру на 1 минуту за испытание. Пусть мыши покопаются в обеих посудах для получения награды.
- На 10-й день повторите 20 испытаний для каждой мыши, но если они углубятся в блюдо без награды, прежде чем они закопаются в блюде с наградой, а затем начните следующее испытание. Дайте мышам, которые первыми закопались в миску с вознаграждением (с ароматом лимонена), достать гранулу награды, прежде чем отправить их обратно в камеру для отдыха.
3. Обучение и тестирование
ПРИМЕЧАНИЕ: Как только мыши начнут надежно копать в поисках невидимых, пахнущих гранул вознаграждения, эксперимент можно начинать.
- Тренировка
ПРИМЕЧАНИЕ: Фаза обучения начинается через два дня после завершения формирования и состоит из 20 попыток для каждой мыши. Перед началом обучения немедленно проводите инфузии интрабульбарного препарата/носителя (подробнее об инфузии см. в разделе 2.1) и начинайте обучение сразу после инфузий.- Поместите мышь в камеру отдыха.
- Поместите две тарелки с песком, пахнущим парой нового запаха, в испытательную камеру, где в одну из тарелок будет зарыта гранула награды.
- Как только испытательная камера будет готова, поднимите непрозрачный барьер и введите мышь в тестовую камеру. Немедленно верните мышь в камеру для отдыха, если мышь сначала закопается в невознаграждаемую посуду (запишите эти испытания как «0»). Если мышь первой впитает полезный запах, дайте ей достать гранулу и верните ее в камеру для отдыха. Запишите эти испытания как «1». Если испытание длится 1 минуту без того, чтобы мышь забрала награду, отправьте мышь обратно в камеру отдыха.
- Вымойте и наполните посуду и начните следующее испытание. Повторите то же самое в течение 20 попыток.
- Тестирование
Примечание: Тестирование памяти может быть проведено в любое время, представляющее интерес для исследователя. В этом эксперименте две отдельные группы мышей были протестированы через 2 ч (STM) или 48 ч (LTM) после обучения, учитывая интерес к дифференциальному влиянию K252a на STM и LTM.- Проведите тестирование с использованием тех же запахов и процедуры, которые описаны для тренинга (раздел 3.1).
ПРИМЕЧАНИЕ: В зависимости от исследовательского вопроса, исследователю может потребоваться включить контрольные группы. Например, в ранее опубликованном эксперименте изучалось влияние блокады Trk-рецепторов на консолидацию памяти3. Таким образом, контрольной группе вводили K252a до 48-часового тестирования, чтобы показать, что эффекты не были связаны с вмешательством в извлечение.
- Проведите тестирование с использованием тех же запахов и процедуры, которые описаны для тренинга (раздел 3.1).
- Статистический анализ
ПРИМЕЧАНИЕ: Синтаксис SPSS 22.0 для каждого шага представлен в виде дополнительного файла в качестве примера.- Выполняйте статистический анализ с помощью линейного анализа смешанных эффектов. В отличие от ANOVA, линейные модели смешанных эффектов могут лучше учитывать случайные эффекты и повторяющиеся измерения.
- Рассчитайте зависимую меру: «правильная пропорция». Вспомните из пункта 3.1.3, что «1» присваивалось испытаниям, в которых мышь первой вдыхала полезный запах, и «0», если мышь первой копалась в неприятном запахе. Усреднить каждые пять попыток для создания четырех пробных блоков (ТБ; например, пробный блок 1 или ТБ1 был средним значением испытаний 1-5, пробный блок 2 или ТБ 2 был средним значением испытаний 6-10 и т.д.).
- Установите независимые переменные или фиксированные эффекты в виде групп лекарств (K252a или Vehicle; . 2.1.1) и пробные блоки (из. 3.3.1). В приведенных ниже репрезентативных результатах указаны переменные, используемые для каждого анализа.
- Включайте отдельную мышь и набор запахов, вложенных в мышь, в качестве «случайных эффектов», чтобы компенсировать внутренние поведенческие различия у мышей и любые эффекты использования нескольких наборов запахов.
- Выполните логит-преобразование для правильной пропорции.
ПРИМЕЧАНИЕ: Правильная пропорция не является непрерывной, несвязанной зависимой переменной. Таким образом, это нарушает два предположения для линейных моделей. Поэтому выполняется логит-преобразование. - Использование оценочных предельных средних для проведения апостериорных тестов значимых взаимодействий, выявленных полной моделью; Множественные попарные сравнения должны быть скорректированы при апостериорном тестировании. Обычно используются Bonferroni или Šidák.
Результаты
Как уже было описано, этот протокол позволяет исследователям оценить влияние некоторых манипуляций на обучение, STM и LTM. Примеры результатов работы Tong et al, 20183 представлены здесь. Полученные результаты подтверждают гипотезу о том, что блокада рецепторов Trk избирательно ингибирует LTM, но не обучение или STM.
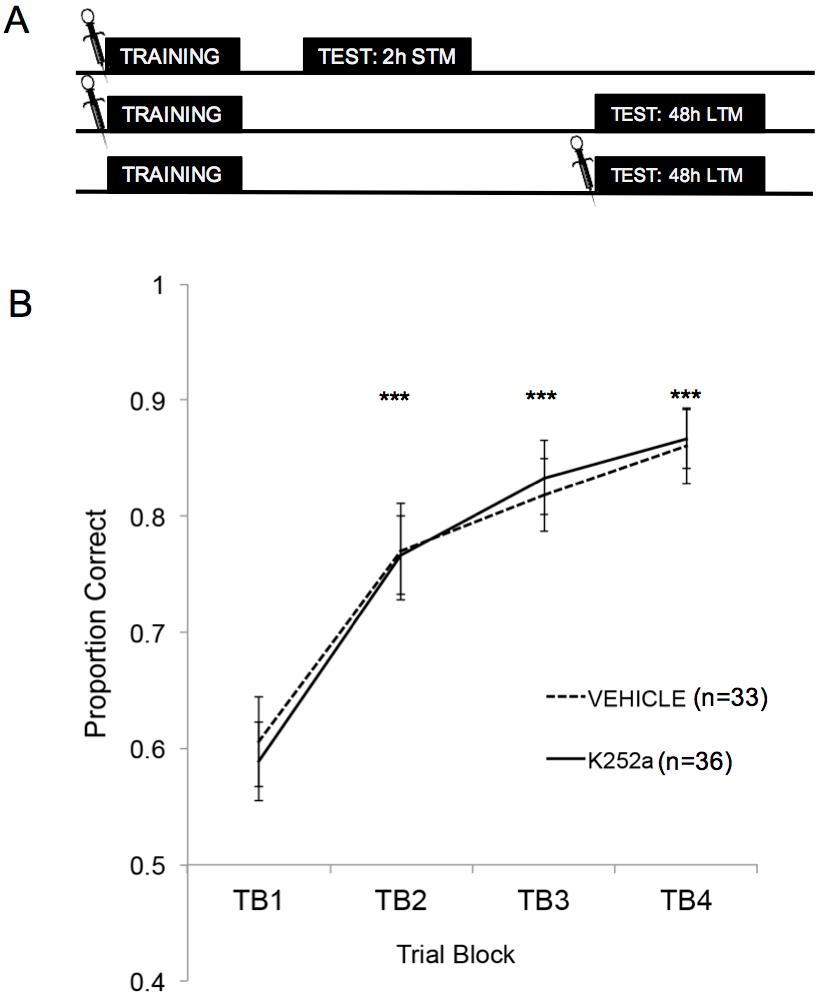
На рисунке 2A показаны схемы обучения, теста STM и теста LTM. Во-первых, было показано, что инфузии K252a не влияют на скорость обучения ассоциации «запах-вознаграждение». На рисунке 2B (раздел 3.1) показана скорость обучения как K252a, так и групп транспортных средств из программы обучения. Статистический анализ с использованием линейной смешанной модели был проведен с двумя фиксированными эффектами: лекарственной группой и пробным блоком (ТБ); Мышь и набор запахов, вложенные в мышь, были случайными эффектами. Данные тестирования (раздел 3.2) не были включены в анализ. Значимый основной эффект наблюдался в пробном блоке (F(3, 183,692) = 43,735, p < 0,001), но отсутствовал эффект от группы препаратов (F(1, 85,685) = 0,132, p = 0,717) и не было значимого взаимодействия (F(3, 183,692) = 0,111, p = 0,954). Апостериорные испытания с использованием корректировки Шидака подтвердили, что K252a и группы транспортных средств не отличались ни на одном из испытательных блоков во время обучения (p > 0,05 для всех сравнений). TB2, TB3 и TB4 были значительно выше, чем TB1 во всех сравнениях (p ≤ 0,001 во всех случаях), демонстрируя, что обе группы успешно усвоили ассоциацию запах-вознаграждение к концу 20 обучающих испытаний.
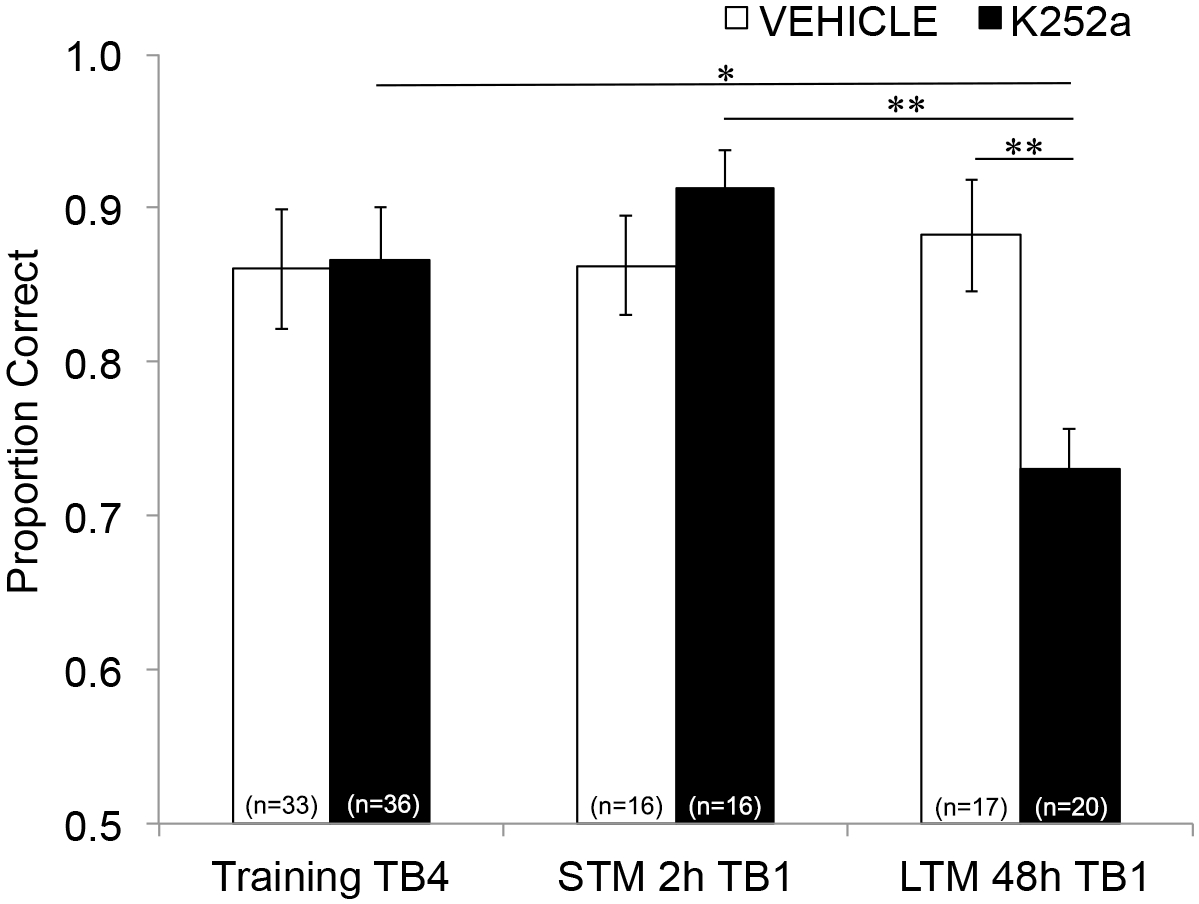
Затем, чтобы изучить влияние инфузии на STM и LTM, был проведен тот же анализ, который включал данные тестирования (раздел 3.2). Оно показало значимое взаимодействие между группой препарата и блоком исследования (F(2, 77,558) = 4,043, p = 0,021), при этом не было значимых основных эффектов группы препаратов (F(1, 55,629) = 1,438, p = 0,236) или блока исследования (F(2, 69,979) = 1,360, p = 0,263). Для того, чтобы конкретно исследовать память, апостериорные парные сравнения с коррекцией Шидака сравнивали различия между последним пробным блоком Тренировки (Раздел 3.1) и первым пробным блоком Тестирования (Раздел 3.2) либо через 2 (STM), либо через 48 часов (LTM). У мышей, получавших инфузионные носители, сравнения показали сохранение ассоциативной памяти как через 2, так и через 48 часов после тренировки (p > 0,05 для всех сравнений с эффективностью тренировки). Для мышей, получавших K252a, первый пробный блок 2-часового теста (STM) не отличался от последнего пробного блока Training (p > 0,05); Тем не менее, их производительность памяти была значительно ниже через 48 ч (p = 0,018). Кроме того, память на 48-часовом тесте была значительно снижена по сравнению с памятью на 2-часовом тесте (p = 0,009) и по сравнению с производительностью группы транспортных средств на 48-часовом тесте (p = 0,006). Не было никакой разницы в STM между мышами, получавшими носители и K252a (p = 0,356). В совокупности результаты показывают, что ингибирование K252a рецепторов Trk в обонятельной луковице избирательно нарушает долгосрочную, но не кратковременную память обоняния (рис. 3).
| Набор запахов | Запах 1 | Запах 2 |
| 1 | пентановая кислота | Бутановая кислота |
| 225.1 | 63.6 | |
| 2 | гексановый | гептанал |
| 11.1 | 35.3 | |
| 3 | пропилацетат | бутилацетат |
| 3.1 | 10.9 | |
| 4 | 2-октанон | 2-гептанон |
| 87.4 | 28.7 | |
| 5 | пентанол | гексанол |
| 37.2 | 127.3 |
Таблица 1: Объемы смешивания для наборов запахов. В каждой строке отображаются два запаха, которые можно использовать в качестве пары для поведенческих шагов. Например, чтобы использовать первый «набор запахов», сделайте смеси пентановой и бутановой кислот. Цифры в таблице указывают объем в μл для смешивания с 50 мл минерального масла для концентрации 1,0 Па каждого запаха. Во время тренировок и испытаний одна чашка Петри ароматизировалась пентановой кислотой, другая — бутановой кислотой.

Рисунок 1: Расположение канюляции и поведенческий аппарат. (A) Показывает взаимное расположение канюли, зубного цементного колпачка и винтов к головке мыши. Обратите внимание, что иглы канюли достают до двух обонятельных луковиц, сама тумба заделывается в колпачок из стоматологического цемента. Винты вставляются в два отверстия, просверленных в черепе над образованием мозжечка. Винты не касаются самого мозга, но они действуют как каудальный якорь для колпачка стоматологического цемента. На рисунке показан относительный размер для изготовления цементной крышки. (Б) Показывает собранный поведенческий аппарат. Тело представляет собой типичную мышиную домашнюю клетку. Крышки с отверстиями для воздуха изготавливались из оргстекла. Центральная перегородка также выполнена из черного оргстекла. Крышки должны быть изготовлены из достаточно тяжелого плексигласа, чтобы они могли служить трактом для подъема и опускания центральной перегородки во время испытаний. (В) Показывает одну сторону поведенческого аппарата. Обратите внимание, что центральный разделитель немного выше каркаса для удобства подъема. По краю можно расположить чашки Петри с песком. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этой цифры.
{kind=link}

Рисунок 2: Дизайн исследования и результаты обучения. (A) Показывает схему плана исследования. Обратите внимание, что группы STM и LTM были независимыми (т.е. разными группами мышей). Символы в начале блоков указывают на время, когда проводились вливания. (B) Показывает пропорцию, правильную для испытаний 1-20 во время обучения. Результаты показывают, что K252a и группы транспортных средств не различались по скорости обучения (наклону линий). Полосы погрешностей представляли собой стандартную погрешность среднего значения (SEM). Звездочки показывают значительное увеличение правильной пропорции по сравнению с TB1 как для транспортных средств, так и для когорт K252a (p ≤ 0,001 для всех сравнений). Этот рисунок адаптирован из Tong et al. 2018 с разрешения3. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этой цифры.
{kind=link}

Рисунок 3: Дифференциальные эффекты блокады Trk-рецепторов на STM и LTM. Показывает правильную пропорцию для пробного блока 4 обучения и пробного блока 1 тестирования STM и LTM. То есть, линейная смешанная модель используется для сравнения пропорции, правильной во время последнего пробного блока фазы обучения (рисунок 2; Training-TB4) во время первых пробных блоков (Testing-TB1) кратковременного (2-часовой тест) и долгосрочного (48-часовой тест) тестов памяти. Линейная смешанная модель имела два фиксированных эффекта: группу лекарств и пробный блок (Training-TB4, STM-TB1, LTM-TB1). Случайными эффектами были мышь и набор запахов, вложенных в мышь. Апостериорные сравнения показали, что у мышей K252a значительно ухудшился LTM (сравнение с обучением TB4; p = 0,018), но не STM (p > 0,05). Производительность LTM мышей, посаженных K252a, также была значительно ниже, чем STM мышей, посаженных в K252a (p = 0,009), и ниже, чем LTM мышей-носителей (p = 0,006). Полосы погрешностей представляли SEM. Этот рисунок адаптирован из Tong et al. 2018 с разрешения3. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этой цифры.
{kind=link}

Дополнительный файл: Синтаксис, используемый для статистического анализа. Пожалуйста, нажмите здесь, чтобы скачать этот файл.
Обсуждение
Обусловливание страха при одной попытке является мощным поведенческим протоколом для изучения молекулярной и клеточной динамики памяти, но большая часть естественного обучения является инкрементальной и лучше всего моделируется с помощью парадигмы, подобной описанной выше. Ингибирование рецепторов Trk в OB предотвращало консолидацию обонятельной памяти в парадигме добровольного обучения, как было показано ранее в нашей группе3. Это открытие открывает новые возможности для исследований дифференциального времени молекулярных механизмов, таких как нейротрофины, в аппетитном и аверсивном обучении.
Этот эксперимент состоял из двух важнейших частей: (1) канюляции и (2) задачи ассоциативного различения (подразделяющейся на формирование, обучение и тестирование). Экспериментаторы могут адаптировать этот протокол к своему конкретному исследовательскому вопросу. Например, нас в первую очередь интересовала ОВ, и этот установленный протокол может быть легко применен к другим исследованиям ОВ. Для экспериментаторов с другими областями, представляющими интерес, будет важно проверить места инфузии в пилотном исследовании. Экспериментаторам также может потребоваться учитывать скорость диффузии, пространственное проникновение и продолжительность биологической активности того, что они вводят.
Этапы формирования, изложенные в протоколе, широко используются авторами этого протокола. Их соблюдение в соответствии с описанием имеет важное значение для того, чтобы мыши своевременно освоили задачу. Используя другие временные шкалы, авторы наблюдали большую вариативность среди мышей в их знакомстве с задачей, и это означало дополнительное обучение, чтобы привести всех мышей к критерию для тестирования с экспериментальными запахами. Для обучения и тестирования исследователь обладает гибкостью, в зависимости от своих исследовательских интересов, в отношении количества испытаний, концентрации экспериментальных запахов и сходства запахов друг с другом. Мы рекомендуем использовать несколько наборов запахов, когда это возможно, как мы описали, чтобы уменьшить количество животных, используемых для эксперимента. В разделе «Статистический анализ» приведены рекомендации по учету использования нескольких наборов ордеров в окончательном анализе. В принципе, сходство пары запахов можно варьировать, чтобы скорректировать сложность различения. В ранее опубликованном исследовании нашей группы3 пары запахов состоят из двух ароматантов одной функциональной группы, но отличающихся друг от друга на одну длину углерода. Эти различения сложнее, чем у пар, отличающихся двумя или более длинами углерода, но легче, чем у энантиомеров (например, (+)-лимонен и (-)-лимонен). Одоранты из разных функциональных групп сильно различаются в восприятии. Cleland et al. 13 обсуждают другие вариации стимулов и их влияние на конкретные параметры обучения.
Одним из основных ограничений этого протокола является то, что он занимает гораздо больше времени по сравнению с автоматизированными задачами ассоциативного обучения, где несколько животных могут быть протестированы параллельно. Для данного теста одному исследователю потребуется не менее 20 минут, чтобы выполнить 20 испытаний для одной мыши. Тем не менее, именно это отсутствие автоматизации означает, что протокол более финансово доступен, что является приоритетом для многих учреждений. Важно отметить, что в случае с этим экспериментом было обнаружено, что этот протокол очень пластичен и эффективен для обучения студентов бакалавриата, интересующихся поведенческой нейронаукой. В частности, эти студенты развивают сильные навыки обращения с животными в дополнение к обычным преимуществам участия в исследованиях.
Исследователи, заинтересованные в принятии этой парадигмы, могут варьировать несколько параметров. Наиболее очевидно, что фармакологические манипуляции с механизмами разнообразны, и этот поведенческий протокол может быть использован с хемогенетическими методами или различными другими способами манипулирования молекулярными и клеточными путями (например, оптогенетикой). Сама парадигма может быть скорректирована в зависимости от типа обучаемости и проверяемой памяти. Например, исследователи могут регулировать сходство двух представленных запахов, чтобы контролировать скорость обучения. В нашем исследовании3 пары запахов состоят из двух одорантов одной функциональной группы, но отличающихся друг от друга на одну длину углерода. Эти различия сложнее, чем у пар, отличающихся двумя или более длинами углерода, но легче, чем у энантиомеров (например, (+)-лимонен и (-)-лимонен). Одоранты из разных функциональных групп сильно различаются в восприятии. Cleland et al. 13 обсуждают другие вариации стимулов и их влияние на конкретные параметры обучения, приходя к выводу, что более похожие запахи труднее различить и, следовательно, требуется больше временидля обучения. Эти манипуляции также повлияют на силу памяти. В этом ключе исследователи могут быть заинтересованы в проверке памяти в разные моменты времени после обучения. Например, в двух исследованиях 15,16 изучалась роль BDNF в персистенции LTM для задачи аверсивного обучения за одну попытку. BDNF является лигандом TrkB. Исследования показали, что инфузия анти-BDNF антисмысловых олигонуклеотидов в гиппокампе через 12 часов после обучения блокировала LTM через 7 дней, но не через 2 дня. Это исследование показывает, что временная шкала молекулярных механизмов после первоначального обучения играет интересную роль, которую еще предстоит понять. В этой статье описывается поведенческий протокол, который позволил бы исследовать эти временные масштабы. Другие параметры, представляющие интерес для будущего применения, включают используемую модель мыши. Например, было бы интересно заменить самцов мышей (которые имеют значительно лучше изученную нейробиологию)17 самками мышей в будущих исследованиях, чтобы изучить различия в скорости обучения, STM и LTM, поскольку самки млекопитающих имеют более высокую чувствительность и избирательность к запахам, чем самцымлекопитающих18. Конечно, модели болезней грызунов также можно эффективно использовать с этим протоколом.
Раскрытие информации
У авторов этой статьи не было конкурирующих финансовых интересов.
Благодарности
Проект был поддержан Scantland Summer Collaborative Research Gift и Исследовательским фондом Стивена и Сильвии Трегидга Берджес. Авторы выражают признательность коллегам из Департамента психологии и Программы неврологии в Колледже Эрлхэм за их поддержку и руководство.
Материалы
| Name | Company | Catalog Number | Comments |
| Double guide cannula | PlasticsOne | C235GS-5-1.5/SPC | Custom order |
| (-)-limonene | Sigma-Aldrich | 218367-50G | |
| (+)-limonene | Sigma-Aldrich | 183164-100ML | |
| 2-hetanone | Sigma-Aldrich | 537683 | |
| 2-octanone | Sigma-Aldrich | O4709 | |
| 5mg sucrose pellets | Test Diet | 1811560 | Custom size. Used for rewards |
| Butanoic acid | Sigma-Aldrich | B103500 | |
| butyl acetate | Sigma-Aldrich | 402842 | |
| Dental Cement Powder (Coral) | A-M Systems | 525000 | |
| Dental Cement Solvent | A-M Systems | 526000 | |
| Double connector assembly | PlasticsOne | C232C | |
| Double dummy cannula | PlasticsOne | C235DCS-5/SPC dummy dbl | Custom order |
| Double injector | PlasticsOne | C235IS-5/SPC | Custom order |
| Drill | Kopf Instruments | Model 1474 High Speed Stereotaxic Drill | This drill requires an additional "adaptor" piece in order to fit certain drill bits. We get by this problem by wrapping the drill bit with lab tape to increase the circumference of the drill it to fit. This may not be an option for surgeries requiring sterile technique. |
| Eye Ointment | Purchase from local pharmacy | ||
| Figure 1 illustration software | BioRender | ||
| heptanal | Sigma-Aldrich | W254002 | |
| hexanal | Sigma-Aldrich | 115606 | |
| hexanol | Sigma-Aldrich | H13303 | |
| Infusion pump model 11 | Harvard Apparatus | 4169D | Used pumps available via American Instrument Exchange |
| Isoflurane | Santa Cruz Animal Health | sc-363629Rx | Vet prescription needed for order |
| K252a | Sigma-Aldrich | K2015 | Mixed to 50uM in DMSO (5%) |
| Ketoprofen | Allivet | 25920 | Vet prescription needed for order |
| Lidocaine | Aspercreme | Purchased from Amazon | |
| Mounting Screws | PlasticsOne | 00-96 X 3/32 | |
| Mouse Anesthesia Mask | Kopf Instruments | Model 907 Mouse Anesthesia Mask | Used with the stereotaxic to allow oxygen and anesthesia while mouse in stereotax |
| Mouse Nose Adaptor | Kopf Instruments | Model 926 Mouse Adaptor | Used with the stereotaxic to allow for head of mouse to be secured. |
| Novalsan | Jeffers | 41375 | |
| Pentanoic acid | Sigma-Aldrich | 240370 | |
| pentanol | Sigma-Aldrich | 138975 | |
| Petri dish glass bottoms | VWR | 10754-804 | |
| Polycarbonate Café bottoms | Ancare | N10PCSEC | Use normal housing cages and custom fit a track in the middle to act as the track for an opaque plexiglass divider |
| propyl acetate | Sigma-Aldrich | 537438 | |
| Quikrete Premium Play Sand | Purchase from local hardware store | ||
| Saline | Insight Needles | N/A | Sterile saline for drug mixing |
| Stereotaxic apparatus | Kopf Instruments | Model 902 Small Animal Stereotaxic Instrument | |
| Testing chamber | Ancare | N10PCSEC | Our testing chambers are modified using the regular mouse housing cage. The manuscript details what was done. |
| Vetbond Tissue Adhesive | 3M | Purchased from Amazon |
Ссылки
- Grimes, M. T., Smith, M., Li, X., Darby-King, A., Harley, C. W., Mclean, J. H. Mammalian Intermediate-Term Memory: New Findings in Neonate Rat. Neurobiology of Learning and Memory. 95 (3), 385-391 (2011).
- Grimes, M. T., Harley, C. W., Darby-King, A., Mclean, J. H. PKA Increases in the Olfactory Bulb Act as Unconditioned Stimuli and Provide Evidence for Parallel Memory Systems: Pairing Odor with Increased PKA Creates Intermediate- and Long-Term, but not Short-Term, Memories. Learning and Memory. 19 (3), 107-115 (2012).
- Tong, M. T., Kim, T. Y. P., Cleland, T. A. Kinase Activity in the Olfactory Bulb is Required for Odor Memory Consolidation. Learning & Memory. 25 (5), 198-205 (2018).
- Devore, S., Lee, J., Linster, C. Odor Preferences Shape Discrimination Learning in Rats. Behavioral Neuroscience. 127 (4), 498-504 (2013).
- Gao, Y., Strowbridge, B. W. Long-Term Plasticity of Excitatory Inputs to Granule Cells in the Rat Olfactory Bulb. Nature Neuroscience. 12 (6), 731-733 (2009).
- Bath, K. G., Mandairon, N., et al. Variant Brain-Derived Neurotrophic Factor (Val66Met) Alters Adult Olfactory Bulb Neurogenesis and Spontaneous Olfactory Discrimination. Journal of Neuroscience. 28 (10), 2383-2393 (2008).
- Lazarini, F., Lledo, P. M. Is Adult Neurogenesis Essential for Olfaction. Trends in Neurosciences. 34 (1), 20-30 (2011).
- Mandairon, N., Peace, S., Karnow, A., Kim, J., Ennis, M., Linster, C. Noradrenergic Modulation in the Olfactory Bulb Influences Spontaneous and Reward-Motivated Discrimination, but not the Formation of Habituation Memory. European Journal of Neuroscience. 27 (5), 1210-1219 (2008).
- Guérin, D., Peace, S. T., Didier, A., Linster, C., Cleland, T. A. Noradrenergic Neuromodulation in the Olfactory Bulb Modulates Odor Habituation and Spontaneous Discrimination. Behavioral Neuroscience. 122 (4), 816-826 (2008).
- Moreno, M. M., Bath, K., Kuczewski, N., Sacquet, J., Didier, A., Mandairon, N. Action of the Noradrenergic System on Adult-Born Cells Is Required for Olfactory Learning in Mice. Journal of Neuroscience. 32 (11), 3748-3758 (2012).
- Vinera, J., Kermen, F., Sacquet, J., Didier, A., Mandairon, N., Richard, M. Olfactory Perceptual Learning Requires Action of Noradrenaline in the Olfactory Bulb: Comparison with Olfactory Associative Learning. Learning and Memory. 22 (3), 192-196 (2015).
- Dillon, T. S., Fox, L. C., Han, C., Linster, C. 17β-estradiol Enhances Memory Duration in the Main Olfactory Bulb in CD-1 Mice. Behavioral Neuroscience. 127 (6), 923 (2013).
- Cleland, T. A., Narla, V. A., Boudadi, K. Multiple Learning Parameters Differentially Regulate Olfactory Generalization. Behavioral Neuroscience. 123 (1), 26 (2009).
- Cleland, T. A., Morse, A., Yue, E. L., Linster, C. Behavioral Models of Odor Similarity. Behavioral Neuroscience. 116 (2), 222-231 (2002).
- Bekinschtein, P., Cammarota, M., Igaz, L., Bevilaqua, L., Izquierdo, I., Medina, J. Persistence of Long-Term Memory Storage Requires a Late Protein Synthesis- and BDNF- Dependent Phase in the Hippocampus. Neuron. 53 (2), 261-277 (2007).
- Bekinschtein, P., et al. BDNF is Essential to Promote Persistence of Long-Term Memory Storage. Proceedings of the National Academy of Sciences. 105 (7), 2711-2716 (2008).
- Andreano, J., Cahill, L. Sex influences on the neurobiology of learning and memory. Learning and Memory. 16 (2009), 248-266 (2009).
- Kass, M. D., Czarnecki, L. A., Moberly, A. H., Mcgann, J. P. Differences in Peripheral Sensory Input to the Olfactory Bulb Between Male and Female Mice. Scientific Reports. 7 (1), (2017).
Перепечатки и разрешения
Запросить разрешение на использование текста или рисунков этого JoVE статьи
Запросить разрешениеThis article has been published
Video Coming Soon
Авторские права © 2025 MyJoVE Corporation. Все права защищены