
Method Article
Um modelo esferoide tridimensional para investigar a interação tumoral-estroma em carcinoma hepatocelular
* Estes autores contribuíram igualmente
Neste Artigo
Resumo
Modelos in vitro abrangentes que recapitulem fielmente a doença humana relevante estão em falta. O presente estudo apresenta a criação e cultura de spheróides tumorais tridimensionais (3D), uma ferramenta in vitro confiável para estudar a interação tumor-estrommal no carcinoma hepatocelular humano.
Resumo
A agressividade e a falta de tratamentos bem tolerados e amplamente eficazes para o carcinoma hepatocelular avançado (CCH), a forma predominante de câncer de fígado, racionalizam sua classificação como a segunda causa mais comum de morte relacionada ao câncer. Modelos pré-clínicos precisam ser adaptados para recapitular as condições humanas para selecionar os melhores candidatos terapêuticos para o desenvolvimento clínico e auxiliar na entrega de medicamentos personalizados. Modelos eferóides celulares tridimensionais (3D) mostram a promessa como uma alternativa in vitro emergente às culturas monocamadas bidimensionais (2D). Aqui, descrevemos um modelo de esferoide de tumor 3D que explora a capacidade de células individuais de agregar quando mantido em gotículas penduradas, e é mais representativo de um ambiente in vivo do que monocamadas padrão. Além disso, os esferoides 3D podem ser produzidos combinando células homotípicas ou heterotípicas, mais reflexivas da heterogeneidade celular in vivo, potencialmente possibilitando o estudo de interações ambientais que podem influenciar a progressão e as respostas ao tratamento. A pesquisa atual otimizou a densidade celular para formar esferoides tumores 3D homotípicos e heterotípicos, imobilizando suspensões celulares nas tampas das placas padrão de 10 cm3 Petri. A análise longitudinal foi realizada para gerar curvas de crescimento para esferoides tumores/fibroblastos homotípicos versus heterotípicos. Finalmente, foi investigado o impacto proliferativo de fibroblastos (células COS7) e miofibroblasts hepáticos (LX2) em esferoides de tumor homotípico (Hep3B). Uma densidade de semeadura de 3.000 células (em 20 μL de mídia) produziu com sucesso esferoides heterotípicos Huh7/COS7, que apresentaram um aumento constante de tamanho até o dia 8 da cultura, seguido pelo retardamento do crescimento. Este achado foi corroborado usando esferoides homotípicos Hep3B cultivados em LX2 (linha celular hepática humana) meio condicionado (CM). O LX2 CM desencadeou a proliferação de esferoides Hep3B em comparação com os esferoides tumorais de controle. Em conclusão, este protocolo mostrou que os esferoides tumorais 3D podem ser usados como uma ferramenta in vitro simples, econômica e pré-tela para estudar as interações tumorais-estromas de forma mais abrangente.
Introdução
A incidência global e a mortalidade por câncer de fígado continuaram a aumentar, apesar dos avanços nos tratamentos para doenças hepáticas e da maioria dos outros tipos de câncer. Em 2018, o câncer de fígado superou o câncer colorretal e estomacal para se tornar a segunda causa mais comum de morte relacionada ao câncer em todo o mundo1. Em 2020, foram mais de 9.000.000 novos diagnósticos, representando 4,7% do total de casos de câncer em todo o mundo1. Isto é particularmente decepcionante, dado que os fatores de risco significativos para o desenvolvimento do CCH, a forma mais comum de câncer de fígado, são bem caracterizados2. A cirrose é o fator de risco mais comum para o desenvolvimento do CCH, com 80% dos casos se desenvolvendo no contexto da cirrose estabelecida2. As doenças hepáticas crônicas, que progridem para a cirrose e, consequentemente, o HCC, incluem o vírus da hepatite B (HBV), o vírus da hepatite C (HCV), a doença hepática relacionada ao álcool (ARLD), as doenças hepáticas gordurosas não alcoólicas (NAFLD) - esta última atribuída à obesidade e diabetes mellitus tipo 2 (T2DM)2,3. Os atuais protocolos de gestão do CCS são dependentes de estágio e limitados para aqueles com câncer avançado, que na maioria das vezes têm um resultado ruim4. Houve avanços significativos usando inibidores da quinase e, mais recentemente, tratamentos imuno-oncologia, embora realisticamente, beneficiam apenas uma minoria de pacientes com câncer de fígado avançado5. Além disso, há preocupação de que os HCCs decorrentes de pacientes com NAFLD - a causa básica de crescimento mais rápido, representando mais de 50% dos casos de HCC recém-diagnosticados em nações ocidentais, possam ser mais resistentes à terapia inibidora de ponto de verificação de morte programada 1 (PD1).
Houve um investimento maciço em ensaios clínicos para pacientes com CCH, incluindo melhorias significativas nos ensaios clínicos e seus pontos finais7. Após uma década de fracassos, esses investimentos começaram a mudar as oportunidades para os pacientes. No entanto, a realidade é que a proporção global de respondentes permanece relativamente pobre, com pacientes recrutados em ensaios muitas vezes mal representando aqueles atendidos nas clínicas. O perigo é que os avanços são caros e beneficiam poucos e não os muitos. À medida que mais terapias candidatas surgem para uso único ou combinação, é essencial ter modelos pré-clínicos mais preditivos de respostas in vivo. Estes são provavelmente modelos que incorporam fatores adicionais que contribuem para a variabilidade observada nas respostas dos pacientes que refletem melhor a heterogeneidade humana e a complexidade patológica8. Sistemas que recriam as condições fisiofisiológicas in vivo do HCC são necessários para ajudar a entender a biologia da evolução do tumor, crescimento e progressão. Os modelos experimentais existentes de doenças hepáticas crônicas e HCC geralmente se enquadram em três categorias principais: modelos in vivo de base animal (revisado in9), culturas in vitro10 e modelos ex vivo11,12. Abordagens baseadas em animais são amplamente utilizadas para estudar doenças hepáticas crônicas, incluindo o CCM; no entanto, a variabilidade genética, os altos custos de funcionamento e os diferentes sistemas imunológicos entre as espécies estão entre as principais limitações para a aplicação desses modelos9. Enquanto alguns modelos ex vivo fornecem uma excelente ferramenta para se concentrar em tecidos humanos em comparação com outros modelos de linha celular in vitro, a disponibilidade de tecidos e o curso de tempo experimental limitado obstruem sua utilização em grande escala.
Por outro lado, os modelos de linha de células in vitro continuam sendo uma boa opção para cientistas que trabalham com recursos limitados, com uma necessidade menor de ter um suprimento constante de tecidos humanos frescos10. Esses modelos também fornecem uma ferramenta que pode ser usada como primeira tela para ajudar na validação de alvo da seleção de medicamentos antes de seguir para modelos in vivo mais complexos. A recente modificação das culturas tradicionais de monocamadas 2D em culturas 3D melhorou a eficácia desses modelos in vitro13,14.
Modelos in vitro 3D podem recapitular características críticas vistas em condições normais e patológicas humanas. Em condições fisiológicas, a transdução de sinal é iniciada através de crosstalk celular e interação com outras moléculas de tecido conjuntivo, ou seja, as proteínas da matriz extracelular (ECM), formando uma rede de interação 3D15,16. O tumor evolui de forma esférica 3D durante a transformação maligna, para a qual oxigênio e nutrientes são facilmente abundantes em interfase não tumoral/tumor. Ao mesmo tempo, as condições hipóxiis predominam no núcleo tumoral. Essa heterogeneidade na disponibilidade de nutrientes resulta na ativação de sinalização espacialmente distinta e vias metabólicas que regulam a tumorigênese. Essas condições são mal recapituladas nas culturas convencionais de monocamadas 2D14, nas quais as células crescem em plástico de cultura rígida de forma fisiologicamente irrelevante. As células cancerígenas também se comunicam com outras células não parenchímicas, a fonte primária de ECM, crescimento e sinalização de invasão dentro do microambiente tumoral. Ao contrário das culturas 2D, modelos in vitro 3D podem fornecer uma plataforma mais adequada para estudar essa interação tumoral-estroma.
Os modelos 3D são amplamente utilizados no campo HCC, e variam na forma como o microcú tecido é formado18,19,20,21,22,23. A maioria desses modelos utilizava as placas de ligação ultra-baixa18,19,20,21,22 ou trans-poços23 no processo de formação de esferoides. O protocolo descrito introduz a técnica de gotícula suspensa como um modelo de esferoides de tumor 3D in vitro alternativo, livre de plástico e econômico. Isso pode facilitar a avaliação dos papéis paracrino e autocrino dos fibroblastos na proliferação das células tumorais em formato 3D.
Protocolo
1. Preparação celular
- Realize todos os experimentos em condições estéreis no gabinete de segurança microbiológica de fluxo laminar classe II (ver Tabela de Materiais).
- Ligue o capô e deixe a estabilização do fluxo de ar.
- Pulverize completamente a superfície do capô interior com 70% de etanol para eliminar qualquer possível contaminação dos usuários anteriores.
- Prepare 5% de uma solução desinfetante em um copo de vidro de 500 mL. Descarte qualquer supernascedor celular ou detritos celulares dentro da solução.
- Limpe minuciosamente todos os micropipettos e caixas de ponta com 70% de etanol.
- Prepare a mídia de cultura celular fresca complementando a média de alta glicose da Eagle (DMEM) modificada de Dulbecco com 10% de soro bovino fetal desativado de calor (FBS), penicilina/estreptomicina de 1% (100 unidade/mL de penicilina e 100 μg/mL de estreptomicina) e 2 mM L-glutamina. Para a linha de células estelares hepáticas LX2, reduza a concentração do suplemento FBS para 2%.
- Mídia de cultura quente antes de iniciar o experimento.
- Pegue as linhas de células tumorais Hein7 e Hep3B HCC e as linhas celulares do fibroblasto COS7 e LX2 de seu rack de armazenamento em nitrogênio líquido. Rapidamente descongelam células criopreservadas.
- Células descongeladas diluídas com 2 mL de mídia de cultura fresca. Centrífugas a 200 x g por 4 min a temperatura ambiente. Descarte o supernatante e resuspense a pelota de célula em 1 mL de mídia de cultura quente fresca.
- Células de sementes em frasco de cultura celular T75. Incubar as células em uma incubadora de cultura celular em 5% de CO2 a 37 °C em 95% condições umidificadas até que as células atinjam 60%-70% de confluência.
2. Coleção celular
- Aspirar mídia de cultura e lavar células três vezes com soro fisco tampão de fosfato (PBS).
- Adicione 2 mL de trippsina pré-aquecida 1x para descolar células aderentes a partir da parte inferior dos frascos T75. Incubar a 37 °C em uma incubadora por 4 min.
- Inativar trippsina adicionando 4 mL de mídia de cultura completa. Colete as células de suspensão celular e centrífuga a 200 x g por 4 min a temperatura ambiente. Descarte o supernatante e resuspenque as células em mídia de cultura fresca de 4 mL.
3. Contagem celular
- Vórtice suavemente a suspensão celular para garantir a distribuição homogênea das células no tubo centrífuga.
- Usando uma pipeta de 10 μL, misture 10 μL de suspensão celular com 10 μL de azul Trypan. Pipete suavemente a mistura quatro vezes para garantir a coloração completa da superfície da célula externa com o corante.
- Conte o número de células usando um hemócito.
- Primeiro, coloque uma mancha de cobertura sobre a área de contagem do hemótmetro antes de carregar a suspensão da célula manchada.
- Coloque a ponta da pipeta contendo a suspensão celular no sulco V do hemócito. Expulse suavemente o conteúdo da ponta para o slide de contagem.
- Deixe o chorume se contentar por alguns minutos antes de consertá-lo no estágio do microscópio para contagem de células.
NOTA: Para evitar a dupla contagem, conte apenas as células dos dois lados do quadrado grande. - Conte em células sobrepostas a decisão superior ou direita e evite aquelas sobreposições da parte inferior ou da decisão esquerda.
- Calcule o número total de células.
NOTA: Número de celular por ml =
4. Coleta de mídia condicionada ao fibroblasto (CM)
- Aspirar mídia de cultura e lavar as células LX2 três vezes com PBS.
- Adicione 2 mL de trippsina pré-aquecida 1x para descolar células aderentes a partir da parte inferior dos frascos T75. Incubar o frasco a 37 °C em uma incubadora por 4 min.
- Inativar trippsina adicionando 4 mL de mídia de cultura completa. Colete as células de suspensão celular e centrífuga a 200 x g por 4 min a temperatura ambiente. Conte as células de acordo com o passo 3.
- Sementes 1 x 106 LX2 células em 10 cm3 pratos para 48 h a 37 °C.
- Recolher o fibroblast CM depois de 48 h. Centrifugar a 200 x g por 4 min a temperatura ambiente para pelotar qualquer célula flutuante. Filtro estéril do CM usando um filtro de 0,22 μm fixado na parte inferior das seringas de 20 mL.
- Colete o CM sobrenante. Aliquot o CM em tubos de 2 mL e armazene a -80 °C para novas aplicações.
NOTA: A solução pode ser armazenada por 6 meses a -80 °C.
5. Validação de densidades celulares para esferoides perfeitos
- Aspirar mídia de cultura e lavar as linhas celulares HCC três vezes com PBS.
- Adicione 2 mL de trippsina pré-aquecida 1x para descolar células aderentes a partir da parte inferior dos frascos T75. Incubar a 37 °C em uma incubadora por 4 min.
- Inativar trippsina adicionando 4 mL de mídia de cultura completa. Colete as células de suspensão celular e centrífuga a 200 x g por 4 min a temperatura ambiente.
- Conte as células de acordo com o passo 3.
- Pipeta diferentes densidades das linhas de células tumorais (12000, 6000, 3000, 1500, 1000, 750, 500, 250 e 125 células) em 20 μL de mídia na superfície interior de uma tampa de 10 cm3 de uma placa de Petri.
- Adicione 10 mL de PBS estéril ao fundo do prato para fornecer condições úmidas para o processo de formação de esferoides.
- Inverta a tampa do prato de 10 cm3 para permitir que a mídia, incluindo a suspensão celular, paire sobre um ambiente úmido. Deixe as gotículas penduradas por 3 dias.
- Tire imagens dos esferoides com ampliação de 50x usando um microscópio invertido 3 dias depois de pendurar as gotículas originais.
6. Tumor heterotípico/esferoides estrobomais
- Suspenda 1500 células HCC Huh7 com 1500 células fibroblastos de mamíferos COS7 (proporção 1:1) em gotículas penduradas para formar esferas. Adicione 10 mL de PBS estéril ao fundo do prato para fornecer condições úmidas para os esferoides.
- Inverta a tampa do prato de 10 cm3 para permitir que a mídia, incluindo a suspensão celular, paire sobre um ambiente úmido. Deixe as gotículas penduradas por 3 dias.
- Tire imagens dos esferoides usando um microscópio invertido do 3º dia até o 10º dia da cultura.
- Coloque a folha de 10 cm3 no palco do microscópio. Ajuste a ampliação do microscópio em 50x para todos os esferoides.
- Abra o software de microscópio no computador conectado e ajuste seu foco para ter uma imagem clara de cada esferoide. Use a Ferramenta de Captura no software de microscópio para salvar as imagens adquiridas.
7. Esferoides Hep3B homotípicos em LX2 CM
- Suspenda 3000 células Hep3B HCC nas gotículas penduradas para formar esferas. Adicione 10 mL de PBS estéril ao fundo do prato para fornecer condições úmidas para os esferoides.
- Inverta a tampa do prato de 10 cm3 para permitir que a mídia, incluindo a suspensão celular, paire sobre um ambiente úmido. Deixe as gotículas penduradas por 3 dias.
- Transfira os esferoides hep3B em 20 μL de CM frescos de células LX2 em gotículas penduradas.
NOTA: Use pontas de pipeta de 200 μL não filtradas estéreis no processo de transferência de esferoides para evitar qualquer interrupção ou ferimento nos esferoides formados. Isto também é para eliminar quaisquer células residuais que permanecem desapegadas ao esferoide único principal.- Use uma pipeta de 20 μL autoclavada para o processo de transferência. Ajuste o volume da pipeta para 2 μL. Fixar a ponta da pipeta à pipeta.
- Inverta a tampa do prato de 10 cm3 no qual os esferoides foram formados. Fixar a tampa no palco de um microscópio leve. Ajuste o foco fino do microscópio para tornar cada esferoide visível.
- Esvazie cuidadosamente o ar da micropipette pressionando o botão do êmbolo. Insira a ponta da pipeta na gota, incluindo o esferoide, a ser transferida. Aproxime-se muito do esferoide sem tocá-lo com a ponta.
- Solte suavemente a pressão no botão do êmbolo para permitir a sucção do esferoide na ponta da micropipette em 2 μL de mídia.
- Transfira o esferoide para uma nova gotícula pendurada em um novo prato de 10 cm3 , tendo nova mídia/mídia/tratamento condicionado.
NOTA: Certifique-se de que todos os esferoides sejam transferidos com sucesso para a nova antena de 10 cm3 usando o microscópio de luz.
- Tire imagens dos esferoides a 50x de ampliação usando um microscópio invertido desde o dia da transferência (dia 3) até o 7º dia de cultura em LX2 CM.
8. Cálculo do volume esferoide
- Atribua um identificador numérico único para cada esferoide para que imagens de esferoides combinados possam ser capturadas diariamente.
- Analise imagens dos esferoides em crescimento usando um pacote de software de análise de imagens.
- Abra cada imagem esferoide dentro do pacote de software. Usando a ferramenta de seleção Mão Livre e delineie cada esferoide. No botão 'Analisar ' suspenso, selecione Definir medição e, em seguida, Área. Pressione ok.
- Desenhe manualmente um círculo em torno de cada esferoide. Uma vez que a esfera é circulada, pressione Ctrl + M para permitir que o programa calcule a área esferoide em Pixels. Converta a área do esferoide em um volume.
NOTA: Volume de spheroidmm3 =0,09403 ×
- Calcule a alteração no volume de spheroid em relação ao seu volume no primeiro dia de captura de imagem.
NOTA: Isto é para normalizar o volume esferoide para o volume inicial e melhorar a precisão dada a variação natural no tamanho inicial.
Resultados
Células cultivadas em um formato 3D de várias camadas refletem com mais precisão a complexidade do microambiente tumoral do que as culturas 2D convencionais24,25. Anteriormente, muitos estudos completaram a mídia de cultura esferoides com diferentes mitogênios e fatores de crescimento26 para iniciar a formação de esferoides. Neste estudo, no entanto, a adição de fibroblastos, ou seu CM, fornece mitogênios essenciais e fatores de crescimento para acelerar o crescimento esferoide.
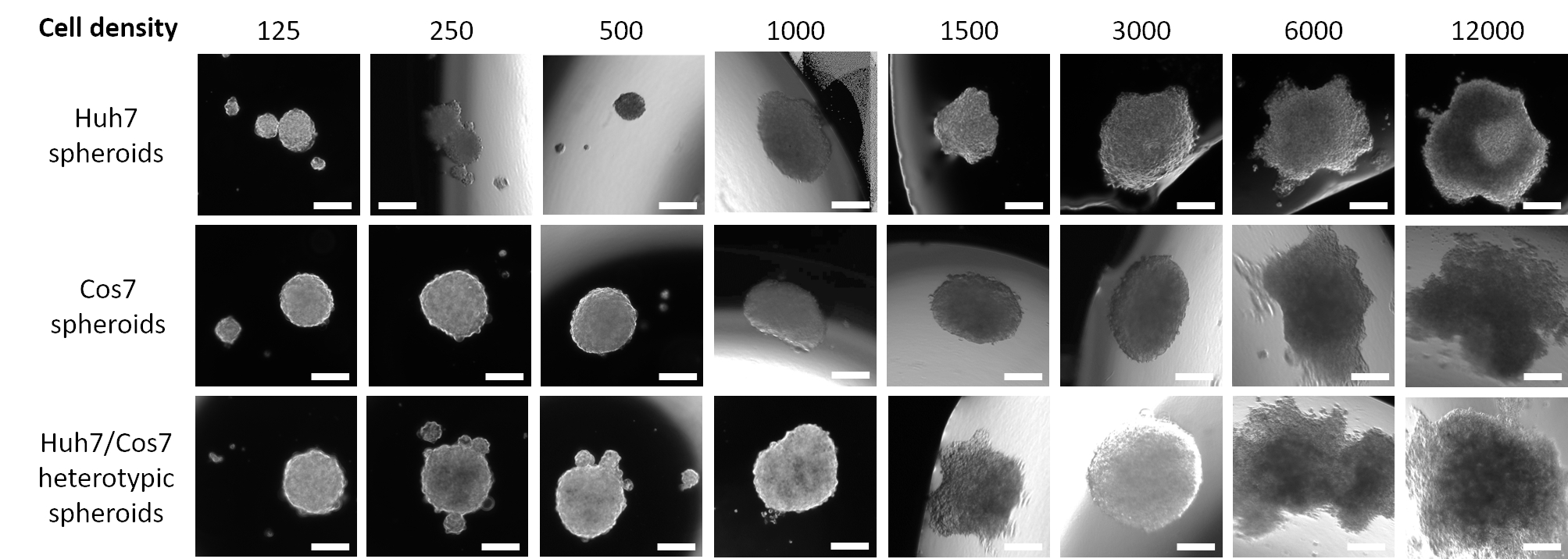
A Figura 1 retrata dados de um estudo piloto no qual as células HCC humanas Huh7 foram semeadas em uma densidade descendente de semeaduras a partir de 12.000 até 125 células em 20 μL de mídia fresca por 3 dias. Uma densidade de semeadura de 12.000 células produziu esferoides com uma forma assimétrica, que não foi corrigida mesmo depois de reduzir pela metade a densidade de semeadura celular. No entanto, esferoides criados a partir de 3000 células pareciam mais arredondados (Figura 1, linha superior). Uma redução adicional da concentração celular não conseguiu formar esferas únicas (125, 250, 500 esferas de densidade celular) nem teve uma aparência esférica regular (1000 e 1500 spheroids de densidade celular). Vale ressaltar que muitas pequenas esferas foram formadas a uma densidade de 500 células tumorais; no entanto, apenas um pequeno esferoide foi capturado em ampliação de 50x (Figura 1, linha superior). O mesmo protocolo otimizado foi aplicado à linha celular do fibroblasto renal primaz COS7 (Figura 1, linha média). A suspensão de 125, 250 e 500 fibroblastos COS7 em gotículas penduradas de 20 μL resultou em um esferoide arredondado com vários esferoides menores, enquanto maiores densidades celulares (1000, 1500 e 3000) formaram esferoides semi-arredondados únicos. Semelhante aos esferoides tumorais Huh7, maiores suspensões celulares (6.000 e 12.000 células/esfera) resultaram na formação de agregados celulares irregulares e, portanto, maiores concentrações celulares foram descartadas (Figura 1, linha média). Baixas densidades de suspensões de células heterotípicas Huh7/COS7 (125, 250 e 500 células/esfera) geraram um único esferoide com vários esferoides flutuantes ou semi-conectados (Figura 1, linha inferior). Esferoides heterotípicos de 1000 e 1500 células foram semi-arredondados, enquanto uma densidade de semeadura de 3000 células (1500 por tipo de célula) deu um esferoide 3D arredondado. Como observado anteriormente, densidades celulares mais elevadas resultaram na formação de agregados em vez de esferoides bem definidos (Figura 1, linha inferior). Em conclusão, 3000 esferoides de densidade celular foram arredondados, semelhantes aos tumores HCC humanos, e foram adaptados para outros experimentos. A colheita das suspensões celulares como gotículas penduradas por 3 dias foi suficiente para promover a formação de esferoides.
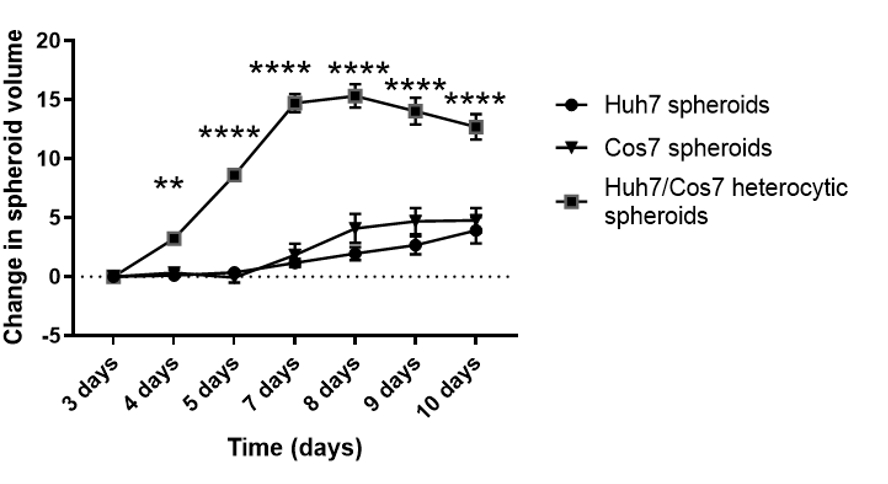
Após otimizar a melhor concentração celular e ponto de tempo para a formação de esferoides no contexto atual, o impacto proliferativo das linhas de células tumorais e fibroblastos foi avaliado longitudinalmente. As figuras 2 e a Figura 3 mostram esferoides heterotípicos Huh7/COS7 (3000 números totais de células por esferoide) monitorados do dia 3 até o dia 10. Esferoides Homotípicos Huh7 e COS7 (1500 células cada) serviram como controles. A partir do 4º dia, os esferoides heterotípicos cresceram em uma forma redonda ideal em comparação com os esferoides homotípicos (Figura 2). O volume inicial do esferoide heterotípico era maior que o de cada esferoide homofóbico. O crescimento dos esferoides foi calculado como a mudança em seu volume em relação ao volume inicial no dia da formação de esferoides para evitar a variação normal no volume esferoide (dia 3, Figura 3). Os esferoides heterotípicos apresentaram inicialmente uma fase de crescimento rápido a partir do dia 4 até o dia 7, seguido por uma fase mais lenta de crescimento no dia 8 (Figuras 2, linha superior e Figura 3). O volume esferoide diminuiu nos dias 9 e 10, possivelmente refletindo o esgotamento de nutrientes ou um núcleo hipóxico e morte celular.
Em contraste, os esferoides homotípicos Huh7 e COS7 cresceram a uma taxa muito mais lenta (Figura 2, linhas médias e inferiores; Figura 3). Os esferoides homotípicos exibiram uma curva de crescimento relativamente estática até o quinto dia de cultura (dois dias após a formação esferoide). A partir do dia 6, os esferoides homotípicos começaram a mostrar um aumento gradual em sua curva de crescimento, embora a uma taxa significativamente menor do que a dos esferoides heterotípicos (Figura 3). Em conclusão, os esferoides heterotípicos tumor/fibroblasto crescem a uma taxa maior do que os esferoides homotípicos sugerindo que o contato direto das células tumorais e dos fibroblastos aumenta o tamanho dos esferoides tumorais.
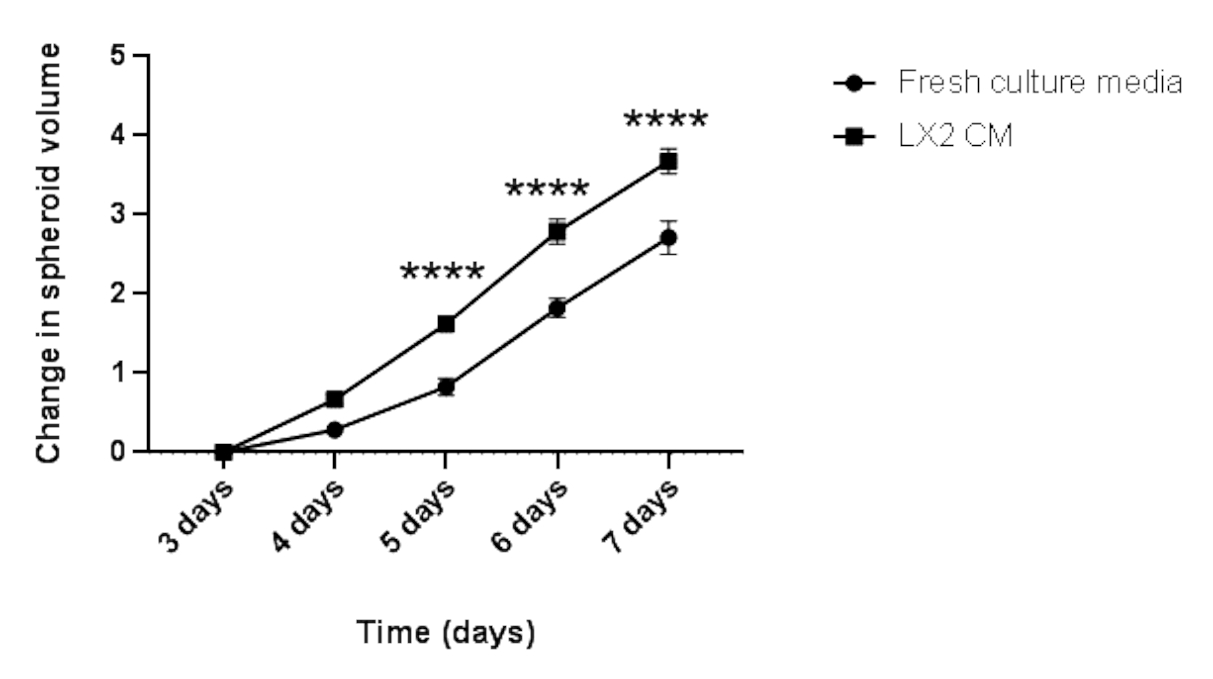
Finalmente, para validar os achados acima e estudar o impacto paracrítico das células mesenquimais na proliferação de esferoides HCC em um contexto relacionado ao fígado, os esferoides hep3B HCC de 3 dias de idade foram cultivados em mídia fresca regular ou CM a partir de células estelares hepáticas LX2 (Figura 4 e Figura 5) por mais 4 dias. À primeira vista, 3000 células Hep3B formaram esferoides perfeitamente arredondados após 3 dias (Figura 4). Os spheróides hep3B mostraram proliferação contínua em mídia fresca do dia 3 até o dia 7 (Figura 4, linha superior). A taxa de crescimento foi aprimorada quando os spheróides Hep3B foram mantidos em LX2 CM (Figura 4, linha inferior). Esta conversão dependente da mídia no crescimento esferoide hep3B apresentou significância estatística do dia 4 até o final do experimento (Figura 5), sugerindo uma proliferação movida por fibroblastos de esferoides tumorais.
Em conclusão, este estudo modificou com sucesso as culturas esferoides 3D existentes para explorar o crosstalk entre células tumorais de HCC e diferentes linhas celulares de fibroblasto e investigar o significado proliferativo dessa interação celular direta e indireta (Figura 6). Uma caracterização adicional dos esferoides formados é necessária para melhorar a forma como as células tumorais interagem com o microambiente circundante.

Figura 1: Otimização da densidade celular ideal para a formação de esferoides. As colunas representam diferentes densidades de semeadura de células, e as linhas representam esferoides formados a partir de células Huh7 (linha superior), células COS7 (linha média) e células heterotípicas Huh7/COS7 (linha inferior). As imagens representam esferoides formados após a suspensão de 125, 250, 500, 1000, 1500, 3000, 6000 e 12.000 células (da esquerda para a direita) em 20 μL de mídia por 3 dias. O estudo piloto incluiu dois esferoides por condição, e as imagens foram tiradas em 50x de ampliação, barra de escala = 200 μm. Por favor clique aqui para ver uma versão maior desta figura.
{kind=link}

Figura 2: Crescimento esferoide heterotípico versus homotípico. As colunas representam o dia em que imagens esferoides foram tiradas, e as linhas mostram imagens representativas para esferoides formados a partir de células Huh7 (linha superior), células COS7 (linha média) e células Huh7/COS7 (linha inferior). As imagens representam os esferoides formados após a suspensão de 3000 células de cada condição (homotípica Huh7, COS7, ou os esferoides heterotípicos Huh7/COS7) de diferentes pontos de tempo da esquerda para a direita. O experimento incluiu 10 esferoides por condição, e as imagens foram tiradas a 50x de ampliação, barra de escala = 200 μm. Por favor, clique aqui para ver uma versão maior desta figura.
{kind=link}

Figura 3: Análise longitudinal do crescimento esferoide heterotípico versus homotípico. O gráfico mostra a curva de crescimento dos esferoides Heterotípicos Huh7/COS7 versus esferoides homotípicos Huh7 e COS7. Os dados são apresentados como média ± s.e.m; n = 10 esferoides independentes. ** p < 0,01; p < 0,001. Clique aqui para ver uma versão maior desta figura.
{kind=link}

Figura 4: Crescimento de esferoides Hep3B Homotípicos em LX2 CM. As colunas representam o dia em que imagens esferoides foram capturadas, e as linhas mostram imagens representativas de esferoides Hep3B cultivados em mídia de cultura DMEM fresca (linha superior) ou LX2 CM (linha inferior). O experimento incluiu sete esferoides por condição (mídia fresca ou LX2 CM), e as imagens foram tiradas a 50x de ampliação, barra de escala = 200 μm. Por favor clique aqui para ver uma versão maior desta figura.
{kind=link}

Figura 5: Análise longitudinal do crescimento esferoide hep3B homotípico. O gráfico mostra a curva de crescimento dos spheróides Hep3B em mídia de cultura DMEM fresca ou LX2 CM. Os dados são apresentados como média ± s.e.m; n = 7 esferoides independentes. p < 0,001. Clique aqui para ver uma versão maior desta figura.
{kind=link}

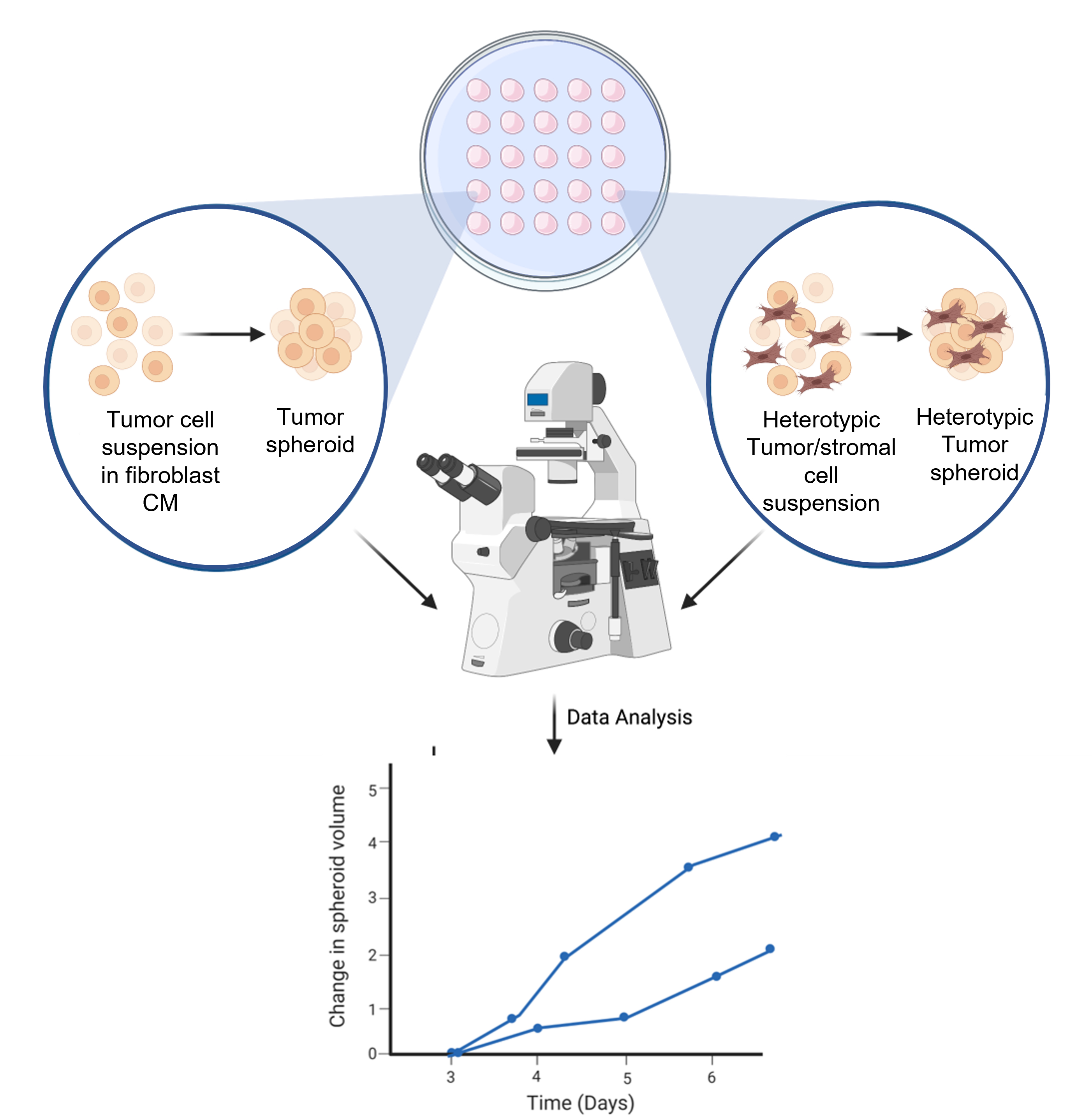
Figura 6: Representação esquemática do processo de formação de esferoides. A suspensão da célula é pipeta na tampa interna de 10 cm3 Petri-prato. A tampa é invertida e mantida por 3 dias para permitir a formação de esferoides homotípicos ou heterotípicos. Imagens esferoides são tiradas a 50x de ampliação. A figura é criada usando uma ferramenta de ilustração científica baseada na Web (ver Tabela de Materiais). Clique aqui para ver uma versão maior desta figura.
{kind=link}
Discussão
O contexto em que as linhas celulares experimentais crescem influencia seu perfil de expressão genética, análise de caminhos e critérios funcionais. Por exemplo, nas células cancerígenas de mama, a coordenação entre diferentes vias oncogênicas só é retida se as células cancerígenas forem cultivadas em conformação 3D27. Genes desregulamentados em melanoma 3D e esferoides de câncer de mama, mas não nas células monocamadas, são mais relevantes para o tumor humano in vivo28,29. Por exemplo, os níveis de integrin β1 em células epiteliais mamárias e contrapartes tumorais foram inferiores aos níveis cultivados em um formato 2D29. Além disso, os fibroblastos em estruturas 3D tendem a migrar distintamente em termos de morfologia e velocidade em comparação com os cultivados em plástico30. Além disso, a rigidez mecânica do substrato de crescimento ativa caminhos específicos que incentivam a transformação maligna das células epiteliais normais através da desregulamentação da quinase extracelular regulada por sinal (ERK)/Rho em células epiteliais31. Esses fatores favorecem a transição da cultura 2D convencional para os modelos esferoides 3D, uma vez que as culturas 3D estão mais de acordo com a doença humana.
O presente estudo utilizou ferramentas e suprimentos de laboratório convencionais para produzir um modelo de esferoide de tumor 3D. Os esferoides formados responderam aos sinais de proliferação provenientes de fibroblastos, como mostra seu crescimento significativamente maior. Este modelo pertence à categoria de esferoides tumorais multicelulares. Existem três outras categorias de esferoides tumorais 3D, incluindo tumoroses32, esferas tumorais derivadas de tecido33,34 e esferoides multicelulares organotipos35. Em esferoides tumorais multicelulares, as células tumorais são suspensas em condições de baixa adesão para permitir que elas se agregem juntas para formar esferas sem tocar no fundo do vaso cultural36. Várias abordagens foram empregadas para fornecer esta técnica independente de ancoragem, que vai desde sistemas rotativos, técnicas de sobreposição líquida e placas em forma de U de fixação ultra-baixa não revestidas37. No HCC, a maioria das culturas esferoides in vitro usam o acessório ultra-baixo de 24 ou 96 placas para formar esferoides arredondados18,19,20,21,22. Esta técnica, no entanto, não é rentável e não exclui qualquer contato acidental celular-plástico. Outros sistemas usam trans-poços23 ou fatias de fígado12 para produzir microsacsúgenos hepáticos. O uso de tecidos humanos no trabalho translacional é o padrão-ouro, mas nem sempre disponível ou acessível por muitos grupos de pesquisa. A abordagem atual beneficiou-se da teoria das gotículas penduradas. A suspensão celular é adicionada para que os esferoides tumorais sejam formados em uma interface de ar líquido acessível para formar esferas arredondadas únicas38. As vantagens do uso dessa técnica são a falta de qualquer contato celular-plástico e a facilidade de estudar o crosstalk autocrino e paracrino entre as células tumoral e fibroblasto. Este modelo foi validado ainda em outro estudo decifrando a importância do TREM2 não parenchímal na proteção do fígado do desenvolvimento do HCC39. Este trabalho mostrou que o volume de spheroids Hep3B e PLC/PRF5 foi maior no controle LX2 CM versus TREM2-overexpressing LX2 CM de forma dependente de WNT39.
No presente estudo, os fibroblastos foram misturados com células tumorais antes da formação esferoide para investigar o impacto proliferativo dessa co-cultura. Outros adicionaram fibroblastos, células endoteliais e células imunes com suspensão de células tumorais após formarem um esferoide tumoral para estudar a migração de células estrômicas para o tumor40,41. O modelo esferoide heterotípico atual mostrou um padrão de crescimento semelhante aos modelos relatados anteriormente e ao tumor in vivo original; esferoides mostram crescimento exponencial seguido de uma fase de crescimento retardada, provavelmente devido ao esgotamento dos nutrientes e ao alargamento do núcleo necrosado42. Em vez de adicionar fatores de crescimento e mitogênios para ajudar a formação de esferoides, este estudo utilizou uma abordagem mais fisiológica, tendo esses fatores de crescimento a partir do contato direto com os fibroblastos ou seu secretome no fibroblast CM. É essencial mencionar que as células LX2 podem ser ativadas ainda mais pelo tratamento com TGFβ1 ou PDGF43. As células LX2 foram tratadas com 10 ng/mL TGFβ1 por 48 h antes de remover a mídia para diferentes configurações experimentais. As células LX2 foram lavadas três vezes com PBS (para excluir qualquer efeito TGFβ1 direto), e então a mídia fresca foi adicionada por mais 24 horas antes de coletar o CM. O CM coletado do LX2 estimulado pelo TGFβ1 induziu o crescimento dos spheroids hep3B a uma taxa maior do que o CM do controle LX2 (dados não mostrados). Este sistema flexível pode se prestar a co-culturas de diferentes células cancerosas mais/menos fibroblastos, bem como linhas derivadas do paciente. Também pode oferecer um ensaio de triagem de rendimento médio para drogas que poderiam ser rapidamente adotadas e inseridas em um pipeline de descoberta de drogas translacionais para informar a dosagem para estudos in vivo.
Uma limitação do presente estudo é o uso das linhas celulares imortalizadas na formação esferoide em vez de células tumorais HCC recém-isoladas e fibroblastos associados ao câncer. Outra limitação é comparar o crescimento do esferoide Hep3B homotípico entre CM de LX2 e em mídia fresca em vez de CM de outras linhas celulares não-fibroblastos. Esta última limitação poderia ser tratada pela modificação genética de um gene de interesse nos fibroblastos seguido pela aplicação do CM do tipo selvagem versus fibroblasto geneticamente modificado para esferoides homotípicos.
Divulgações
A FO é diretora e acionista da Fibrofind Limited.
Agradecimentos
O MYWZ é financiado pelo Secretário de Estado para Negócios, Energia e Estratégia Industrial e pelo Prêmio Newton 2020 como parte do fundo "ODA" e do Fundo Newton. A SS é apoiada pela bolsa cientista clínico avançado C53575/A29959 da Cancer Research UK Advanced Clinician Scientist. SS, FO e RH recebem financiamento como parte da HUNTER, financiada por meio de uma parceria entre a Cancer Research UK, Fondazione AIRC e Fundacion Cientifica de la Asociacion Espanola Contra el Cancer.
Materiais
| Name | Company | Catalog Number | Comments |
| 1x Trypsin | Sigma Aldrich | 59429C | |
| 2 mL tubes | Eppendorf | ||
| Biorender | Biorender.com | Online | |
| Class II laminar flow BioMAT2 hood | Medair Technologies | ||
| Dulbecco’s modified Eagle’s medium (DMEM) | Biosera | M-D1107 | |
| Excel sofware | Microsoft office 365 | ||
| Graphpad prism sofware | GraphPad software | ||
| Heat deactivated fetal bovine serum (FBS) | life tech | 105000064 | |
| Image J software | Fiji | ||
| L-glutamine | Sigma Aldrich | G7513-100ml | |
| Penicillin/streptomycin (100 unit/ml penicillin and 100 μg/ml streptomycin) | Sigma Aldrich | P0781-100ml | |
| Petri dish 90 mm, triple vented | Greiner | 633175 | |
| Trypan blue | Sigma Aldrich | T8154 | |
| Virkon™ Broad Spectrum Disinfectant, 5kg Bucket | Rely+On™ | SCI-129999 | |
| Ziess inverted microscope | Ziess |
Referências
- Ferlay, J., et al. Cancer statistics for the year 2020: an overview. International Journal of Cancer. , (2021).
- Llovet, J. M., et al. Hepatocellular carcinoma. Nature Reviews Disease Primers. 7 (1), 6 (2021).
- Reeves, H. L., Zaki, M. Y., Day, C. P. Hepatocellular carcinoma in obesity, Type 2 diabetes, and NAFLD. Digestive Diseases & Sciences. 61 (5), 1234-1245 (2016).
- Forner, A., Reig, M., Bruix, J. Hepatocellular carcinoma. Lancet. 391 (10127), 1301-1314 (2018).
- Reig, M., et al. Diagnosis and treatment of hepatocellular carcinoma. Update of the consensus document of the AEEH, AEC, SEOM, SERAM, SERVEI, and SETH. Medicina Clinica (Barc). 156 (9), 1-30 (2021).
- Pfister, D., et al. NASH limits anti-tumour surveillance in immunotherapy-treated HCC. Nature. 592 (7854), 450-456 (2021).
- Llovet, J. M., et al. Trial design and endpoints in hepatocellular carcinoma: AASLD consensus conference. Hepatology. 73, 158-191 (2021).
- Llovet, J. M., Hernandez-Gea, V. Hepatocellular carcinoma: reasons for phase III failure and novel perspectives on trial design. Clinical Cancer Research. 20 (8), 2072-2079 (2014).
- Brown, Z. J., Heinrich, B., Greten, T. F. Mouse models of hepatocellular carcinoma: an overview and highlights for immunotherapy research. Nature Reviews Gastroenterology and Hepatology. 15 (9), 536-554 (2018).
- Nikolic, M., Sustersic, T., Filipovic, N. In vitro models and on-chip systems: Biomaterial interaction studies with tissues generated using lung epithelial and liver metabolic cell lines. Frontiers in Bioengineering and Biotechnology. 6, 120 (2018).
- Clark, A. M., Ma, B., Taylor, D. L., Griffith, L., Wells, A. Liver metastases: Microenvironments and ex-vivo models. Experimental Biology and Medicine. 241 (15), 1639-1652 (2016).
- Paish, H. L., et al. A bioreactor technology for modeling fibrosis in human and rodent precision-cut liver slices. Hepatology. 70 (4), 1377-1391 (2019).
- Brassard-Jollive, N., Monnot, C., Muller, L., Germain, S. In vitro 3D systems to model tumor angiogenesis and interactions with stromal cells. Frontiers in Cell and Developmental Biology. 8, 594903 (2020).
- Pampaloni, F., Reynaud, E. G., Stelzer, E. H. The third dimension bridges the gap between cell culture and live tissue. Nature Reviews Molecular Cell Biology. 8 (10), 839-845 (2007).
- Kleinman, H. K., Philp, D., Hoffman, M. P. Role of the extracellular matrix in morphogenesis. Current Opinion in Biotechnology. 14 (5), 526-532 (2003).
- Bissell, M. J., Radisky, D. C., Rizki, A., Weaver, V. M., Petersen, O. W. The organizing principle: microenvironmental influences in the normal and malignant breast. Differentiation. 70 (9-10), 537-546 (2002).
- Ronnov-Jessen, L., Petersen, O. W., Koteliansky, V. E., Bissell, M. J. The origin of the myofibroblasts in breast cancer. Recapitulation of tumor environment in culture unravels diversity and implicates converted fibroblasts and recruited smooth muscle cells. Journal of Clinical Investigations. 95 (2), 859-873 (1995).
- Shao, H., et al. A novel stromal fibroblast-modulated 3D tumor spheroid model for studying tumor-stroma interaction and drug discovery. Journal of Visualized Experiments: JoVE. (156), e60660 (2020).
- Song, Y., et al. Activated hepatic stellate cells play pivotal roles in hepatocellular carcinoma cell chemoresistance and migration in multicellular tumor spheroids. Scientific Reports. 6, 36750 (2016).
- Pingitore, P., et al. Human multilineage 3D spheroids as a model of liver steatosis and fibrosis. International Journal of Molecular Sciences. 20 (7), (2019).
- Kozyra, M., et al. Human hepatic 3D spheroids as a model for steatosis and insulin resistance. Scientific Reports. 8 (1), 14297 (2018).
- Khawar, I. A., et al. Three dimensional mixed-cell spheroids mimic stroma-mediated chemoresistance and invasive migration in hepatocellular carcinoma. Neoplasia. 20 (8), 800-812 (2018).
- Feaver, R. E., et al. Development of an in vitro human liver system for interrogating non-alcoholic steatohepatitis. Journal of Clinical Investigations Insight. 1 (20), 90954 (2016).
- Wartenberg, M., et al. Regulation of the multidrug resistance transporter P-glycoprotein in multicellular tumor spheroids by hypoxia-inducible factor (HIF-1) and reactive oxygen species. Federation of American Societies for Experimental Biology Journal. 17 (3), 503-505 (2003).
- Chen, R., et al. Screening candidate metastasis-associated genes in three-dimensional HCC spheroids with different metastasis potential. International Journal of Clinical Experimental Pathology. 7 (5), 2527-2535 (2014).
- Venkataraman, G., et al. Preferential self-association of basic fibroblast growth factor is stabilized by heparin during receptor dimerization and activation. Proceedings of the National Academy of Sciences of the United States of America. 93 (2), 845-850 (1996).
- Bissell, M. J., et al. Tissue structure, nuclear organization, and gene expression in normal and malignant breast. Cancer Research. 59, 1757-1763 (1999).
- Ghosh, S., et al. Three-dimensional culture of melanoma cells profoundly affects gene expression profile: a high density oligonucleotide array study. Journal of Cellular Physiology. 204 (2), 522-531 (2005).
- Delcommenne, M., Streuli, C. H. Control of integrin expression by extracellular matrix. Journal of Biological Chemistry. 270 (45), 26794-26801 (1995).
- Meshel, A. S., Wei, Q., Adelstein, R. S., Sheetz, M. P. Basic mechanism of three-dimensional collagen fibre transport by fibroblasts. Nature Cell Biology. 7 (2), 157-164 (2005).
- Paszek, M. J., et al. Tensional homeostasis and the malignant phenotype. Cancer Cell. 8 (3), 241-254 (2005).
- Yu, M., et al. Cancer therapy. Ex vivo culture of circulating breast tumor cells for individualized testing of drug susceptibility. Science. 345 (6193), 216-220 (2014).
- Weiswald, L. B., et al. Newly characterised ex vivo colospheres as a three-dimensional colon cancer cell model of tumour aggressiveness. British Journal of Cancer. 101 (3), 473-482 (2009).
- Kondo, J., et al. Retaining cell-cell contact enables preparation and culture of spheroids composed of pure primary cancer cells from colorectal cancer. Proceedings of the National Academy of Sciences of the United States of America. 108 (15), 6235-6240 (2011).
- Rajcevic, U., et al. Colorectal cancer derived organotypic spheroids maintain essential tissue characteristics but adapt their metabolism in culture. Proteome Sciences. 12, 39 (2014).
- Friedrich, J., Seidel, C., Ebner, R., Kunz-Schughart, L. A. Spheroid-based drug screen: considerations and practical approach. Nature Protocols. 4 (3), 309-324 (2009).
- Friedrich, J., Ebner, R., Kunz-Schughart, L. A. Experimental anti-tumor therapy in 3-D: spheroids--old hat or new challenge. International Journal of Radiation Biology. 83 (11-12), 849-871 (2007).
- Kelm, J. M., Timmins, N. E., Brown, C. J., Fussenegger, M., Nielsen, L. K. Method for generation of homogeneous multicellular tumor spheroids applicable to a wide variety of cell types. Biotechnology & Bioengineering. 83 (2), 173-180 (2003).
- Esparza-Baquer, A., et al. TREM-2 defends the liver against hepatocellular carcinoma through multifactorial protective mechanisms. Gut. 70 (7), 1345-1361 (2021).
- Dangles-Marie, V., et al. A three-dimensional tumor cell defect in activating autologous CTLs is associated with inefficient antigen presentation correlated with heat shock protein-70 down-regulation. Cancer Research. 63 (13), 3682-3687 (2003).
- Timmins, N. E., Dietmair, S., Nielsen, L. K. Hanging-drop multicellular spheroids as a model of tumour angiogenesis. Angiogenesis. 7 (2), 97-103 (2004).
- Mayer, B., et al. Multicellular gastric cancer spheroids recapitulate growth pattern and differentiation phenotype of human gastric carcinomas. Gastroenterology. 121 (4), 839-852 (2001).
- Xu, L., et al. Human hepatic stellate cell lines, LX-1 and LX-2: new tools for analysis of hepatic fibrosis. Gut. 54 (1), 142-151 (2005).
Reimpressões e Permissões
Solicitar permissão para reutilizar o texto ou figuras deste artigo JoVE
Solicitar PermissãoExplore Mais Artigos
This article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. Todos os direitos reservados