
Method Article
Трехмерная сфероидная модель для исследования опухоле-стромального взаимодействия при гепатоцеллюлярной карциноме
* Эти авторы внесли равный вклад
В этой статье
Резюме
Отсутствуют комплексные модели in vitro , которые точно повторяют соответствующие заболевания человека. В текущем исследовании представлено трехмерное (3D) создание и культура сфероидов опухолей, надежный инструмент in vitro для изучения опухолево-стромального взаимодействия при гепатоцеллюлярной карциноме человека.
Аннотация
Агрессивность и отсутствие хорошо переносимых и широко эффективных методов лечения прогрессирующей гепатоцеллюлярной карциномы (ГЦК), преобладающей формы рака печени, рационализируют ее статус второй наиболее распространенной причины смерти, связанной с раком. Доклинические модели должны быть адаптированы для повторения человеческих условий, чтобы выбрать лучших терапевтических кандидатов для клинической разработки и помочь в доставке персонализированной медицины. Трехмерные (3D) клеточные сфероидные модели показывают перспективность в качестве новой альтернативы in vitro двумерным (2D) монослойным культурам. Здесь мы описываем 3D-модель сфероида опухоли, которая использует способность отдельных клеток агрегироваться при поддержании в висячих каплях и является более репрезентативной для среды in vivo , чем стандартные монослои. Кроме того, 3D-сфероиды могут быть получены путем объединения гомотипических или гетеротипических клеток, более отражающих клеточную гетерогенность in vivo, потенциально позволяя изучать взаимодействия окружающей среды, которые могут влиять на прогрессирование и реакции на лечение. Текущее исследование оптимизировало плотность клеток для формирования 3D-гомотипических и гетеротипических сфероидов опухолей путем иммобилизации клеточных суспензий на крышках стандартных чашек Петри размером 10 см3 . Продольный анализ проводили для генерации кривых роста для гомотипических и гетеротипических сфероидов опухолей/фибробластов. Наконец, исследовано пролиферативное воздействие фибробластов (клетки COS7) и миофибробластов печени (LX2) на сфероиды гомотипической опухоли (Hep3B). Плотность посева 3000 клеток (в среде 20 мкл) успешно дала гетеротипические сфероиды Huh7/COS7, которые демонстрировали устойчивое увеличение размера до 8-го дня культивирования с последующей задержкой роста. Этот вывод был подтвержден с использованием гомотипических сфероидов Hep3B, культивируемых в кондиционированной среде LX2 (человеческая печеночная звездчатая клеточная линия) (CM). LX2 CM вызвал пролиферацию сфероидов Hep3B по сравнению с контрольными сфероидами опухоли. В заключение, этот протокол показал, что 3D-сфероиды опухолей могут использоваться в качестве простого, экономичного и предварительного инструмента in vitro для более всестороннего изучения опухолево-стромальных взаимодействий.
Введение
Глобальная заболеваемость и смертность от рака печени продолжают расти, несмотря на достижения в лечении заболеваний печени и большинства других видов рака. В 2018 году рак печени превзошел колоректальный рак и рак желудка, став второй наиболее распространенной причиной смерти, связанной с раком, во всем мире1. В 2020 году было поставлено более 9 000 000 новых диагнозов, что составляет 4,7% от общего числа случаев рака во всем мире1. Это особенно разочаровывает, учитывая, что значительные факторы риска развития ГЦК, наиболее распространенной формы рака печени, хорошо охарактеризованы2. Цирроз печени является наиболее распространенным фактором риска развития ГЦК, причем 80% случаев развиваются на фоне установленного цирроза2. Хронические заболевания печени, которые прогрессируют до цирроза и, следовательно, ГЦК, включают вирус гепатита В (HBV), вирус гепатита С (HCV), связанные с алкоголем заболевания печени (ARLD), неалкогольные жировые заболевания печени (NAFLD) - последние относятся к ожирению и сахарному диабету 2 типа (T2DM)2,3. Текущие протоколы лечения ГЦК зависят от стадии и ограничены для пациентов с прогрессирующим раком, которые чаще всего имеют плохой исход4. Были достигнуты значительные успехи в использовании ингибиторов киназы и, в последнее время, иммуноонкологические методы лечения, хотя и реалистично, приносят пользу лишь меньшинству пациентов с прогрессирующим раком печени5. Кроме того, существует опасение, что ГЦК, возникающие у пациентов с НАЖБП - наиболее быстро растущей основной причиной, на которую приходится более 50% недавно диагностированных случаев ГЦК в западных странах, могут быть более устойчивыми к терапии ингибиторами контрольных точек запрограммированной смерти 1 (PD1)6.
Были осуществлены огромные инвестиции в клинические испытания для пациентов с ГЦК, включая значительные улучшения в клинических испытаниях и их конечных точках7. После десятилетия неудач эти инвестиции начали менять возможности для пациентов. Однако реальность такова, что общая доля респондентов остается относительно низкой, причем пациенты, набранные в испытания, часто плохо представляют тех, за кем ухаживают в клиниках. Опасность заключается в том, что авансы являются дорогостоящими и приносят пользу немногим, а не многим. По мере того, как появляется все больше потенциальных методов лечения для одноразового или комбинированного применения, важно иметь доклинические модели, более прогностические реакции in vivo. Вероятно, это будут модели, которые включают дополнительные факторы, способствующие изменчивости, наблюдаемой в ответах пациентов, которые лучше отражают гетерогенность и патологическую сложность ГЦК человека8. Системы, которые воссоздают in vivo патофизиологические условия ГЦК, необходимы, чтобы помочь понять биологию эволюции, роста и прогрессирования опухоли. Существующие экспериментальные модели хронических заболеваний печени и ГЦК обычно подпадают под три основные категории: модели in vivo животного происхождения (рассмотрены в 9), культуры in vitro10 и модели ex vivo11,12. Подходы на животных широко используются для изучения хронических заболеваний печени, включая ГЦК; однако генетическая изменчивость, высокие эксплуатационные расходы и различия в иммунных системах между видами являются одними из основных ограничений для применения таких моделей9. В то время как некоторые модели ex vivo обеспечивают отличный инструмент для сосредоточения внимания на тканях человека по сравнению с другими моделями клеточных линий in vitro, доступность тканей и ограниченный ход экспериментального времени препятствуют их использованию в больших масштабах.
С другой стороны, модели клеточных линий in vitro остаются хорошим вариантом для ученых, работающих с ограниченными ресурсами, с меньшей потребностью в постоянном снабжении свежими тканями человека10. Эти модели также предоставляют инструмент, который можно использовать в качестве первого экрана, чтобы помочь с целевой проверкой выбора лекарств, прежде чем переходить к более сложным моделям in vivo. Недавняя модификация традиционных 2D монослойных культур в 3D-культуры повысила эффективность этих моделей in vitro13,14.
3D-модели in vitro могут повторять критические особенности, наблюдаемые в нормальных и патологических состояниях человека. В физиологических условиях сигнальная трансдукция инициируется посредством клеточных перекрестных помех и взаимодействия с другими молекулами соединительной ткани, а именно белками внеклеточного матрикса (ECM), образуя сеть 3D-взаимодействия15,16. Опухоль развивается в 3D-сферической форме во время злокачественной трансформации, для которой кислород и питательные вещества легко обильны в неопухолевой / опухолевой интерфазе. При этом в ядре опухоли преобладают гипоксические состояния. Эта неоднородность в доступности питательных веществ приводит к активации пространственно различных сигнальных и метаболических путей, которые регулируют опухолевый генез. Эти условия плохо повторяются в обычных 2D-монослойных культурах14, в которых клетки растут на жесткой культуральной пластике физиологически нерелевантным образом. Раковые клетки также общаются с другими непаренхимальными клетками, основным источником ECM, роста и инвазии, сигнализирующих в микроокружении опухоли. В отличие от 2D-культур, 3D-модели in vitro могут обеспечить более подходящую платформу для изучения этого опухолево-стромального взаимодействия17.
3D-модели широко используются в области HCC, и они различаются по способу формирования микроткани18,19,20,21,22,23. В большинстве этих моделей в процессе формирования сфероидов использовались либо сверхнизкие связующие пластины18,19,20,21,22, либо транс-скважины23. Описанный протокол вводит метод висячих капель в качестве альтернативной, не содержащей пластика и экономически эффективной 3D-модели сфероидной опухоли in vitro. Это может облегчить оценку паракринной и аутокринной ролей фибробластов в пролиферации опухолевых клеток в 3D-формате.
протокол
1. Клеточная подготовка
- Провести все эксперименты в стерильных условиях в шкафу микробиологической безопасности ламинарного потока II класса (см. Таблицу материалов).
- Включите капот и обеспечьте стабилизацию воздушного потока.
- Тщательно опрыскайте внутреннюю поверхность вытяжки 70% этанолом, чтобы исключить любое возможное загрязнение от предыдущих пользователей.
- Приготовьте 5% дезинфицирующего раствора в стеклянном стакане объемом 500 мл. Выбросьте любой клеточный супернатант или клеточный мусор внутри раствора.
- Тщательно очистите все микропипетки и коробки с наконечниками с 70% этанолом.
- Приготовьте свежие питательные среды для культивирования клеток, дополнив модифицированную среду Eagle (DMEM) Dulbecco высокой глюкозой с 10% дезактивированной теплом фетальной бычьей сывороткой (FBS), 1% пенициллина / стрептомицина (100 единиц / мл пенициллина и 100 мкг / мл стрептомицина) и 2 мМ L-глутамина. Для печеночной звездчатой клеточной линии LX2 уменьшите концентрацию добавки FBS до 2%.
- Нагревайте питательные среды перед началом эксперимента.
- Возьмите линии опухолевых клеток Huh7 и Hep3B HCC и клеточные линии фибробластов COS7 и LX2 из их стойки для хранения в жидком азоте. Быстро размораживает криоконсервированные клетки.
- Разбавить размороженные клетки 2 мл свежей питательной среды. Центрифужные ячейки при 200 х г в течение 4 мин при комнатной температуре. Выбросьте супернатант и повторно суспендируйте гранулу клетки в 1 мл свежей теплой питательной среды.
- Семенные клетки в колбе для культивирования клеток Т75. Инкубируют клетки в инкубаторе клеточной культуры в 5% CO2 при 37 °C в 95% увлажненных условиях до тех пор, пока клетки не достигнут 60%-70% слияния.
2. Коллекция ячеек
- Аспирировать культуральную среду и трижды промыть клетки фосфатным буферным физиологическим раствором (PBS).
- Добавьте 2 мл предварительно нагретого 1x трипсина, чтобы отделить адгезивные клетки от нижней части колб T75. Инкубировать при 37 °C в инкубаторе в течение 4 мин.
- Инактивировать трипсин путем добавления 4 мл полной питательной среды. Соберите ячейку суспензии и центрифуги при 200 х г в течение 4 мин при комнатной температуре. Выбросьте супернатант и повторно суспендируйте клетки в свежих питательных средах объемом 4 мл.
3. Подсчет клеток
- Осторожно вращайте клеточную суспензию, чтобы обеспечить однородное распределение клеток в трубке центрифуги.
- Используя пипетку 10 мкл, смешайте 10 мкл клеточной суспензии с 10 мкл трипанового синего. Аккуратно выложите смесь вверх и вниз четыре раза, чтобы обеспечить полное окрашивание наружной поверхности клетки красителем.
- Подсчитайте количество клеток с помощью гемоцитометра.
- Во-первых, поместите крышку над областью подсчета гемоцитометра перед загрузкой окрашенной клеточной суспензии.
- Поместите наконечник пипетки, содержащий клеточную суспензию, в V-образную канавку гемоцитометра. Осторожно выталкивайте содержимое наконечника в счетный слайд.
- Оставьте суспензию на пару минут, прежде чем зафиксировать ее на стадии микроскопа для подсчета клеток.
ПРИМЕЧАНИЕ: Чтобы избежать двойного счета, подсчитывайте ячейки только с двух сторон большого квадрата. - Подсчитывайте ячейки, перекрывающие верхнее или правое постановление, и избегайте тех, которые перекрывают нижнюю или левую границу.
- Рассчитайте общее количество ячеек.
ПРИМЕЧАНИЕ: Количество ячеек на мл =
4. Сбор кондиционированных сред фибробластов (СМ)
- Аспирируйте культуральную среду и трижды промывайте клетки LX2 PBS.
- Добавьте 2 мл предварительно нагретого 1x трипсина, чтобы отделить адгезивные клетки от нижней части колб T75. Инкубировать колбу при 37 °C в инкубаторе в течение 4 мин.
- Инактивировать трипсин путем добавления 4 мл полной питательной среды. Соберите ячейку суспензии и центрифуги при 200 х г в течение 4 мин при комнатной температуре. Подсчитайте ячейки в соответствии с шагом 3.
- Семена 1 х 106 LX2 клеток в посуде 10 см3 в течение 48 ч при 37 °C.
- Собирают фибробласт СМ через 48 ч. Центрифуга при 200 х г в течение 4 мин при комнатной температуре гранулирует любые плавающие ячейки. Стерильная фильтрация СМ с помощью фильтра 0,22 мкм, закрепленного на дне шприцев объемом 20 мл.
- Соберите супернатант CM. Aliquot CM в пробирки по 2 мл и храните при -80 °C для дальнейшего применения.
ПРИМЕЧАНИЕ: Раствор можно хранить в течение 6 месяцев при -80 °C.
5. Проверка плотности клеток для идеальных сфероидов
- Аспирируйте питательные среды и промывайте клеточные линии ГЦК три раза PBS.
- Добавьте 2 мл предварительно нагретого 1x трипсина, чтобы отделить адгезивные клетки от нижней части колб T75. Инкубировать при 37 °C в инкубаторе в течение 4 мин.
- Инактивировать трипсин путем добавления 4 мл полной питательной среды. Соберите ячейку суспензии и центрифуги при 200 х г в течение 4 мин при комнатной температуре.
- Подсчитайте ячейки в соответствии с шагом 3.
- Пипетка различной плотности от опухолевых клеточных линий (12000, 6000, 3000, 1500, 1000, 750, 500, 250 и 125 клеток) в 20 мкл среды на внутренней поверхности 10 см3 крышки чашки Петри.
- Добавьте 10 мл стерильного PBS на дно тарелки, чтобы обеспечить влажные условия для процесса образования сфероидов.
- Переверните крышку тарелки размером 10 см3 , чтобы среда, включая клеточную суспензию, висела над влажной средой. Оставьте висячие капли на 3 дня.
- Делайте снимки сфероидов с 50-кратным увеличением с помощью перевернутого микроскопа через 3 дня после подвешивания оригинальных капель.
6. Гетеротипические опухолевые/стромальные сфероиды
- Суспендировать 1500 клеток Huh7 HCC с 1500 клетками фибробластов млекопитающих COS7 (соотношение 1:1) в висячих каплях для формирования сфер. Добавьте 10 мл стерильного PBS на дно тарелки, чтобы обеспечить влажные условия для сфероидов.
- Переверните крышку тарелки размером 10 см3 , чтобы среда, включая клеточную суспензию, висела над влажной средой. Оставьте висячие капли на 3 дня.
- Делайте снимки сфероидов с помощью инвертированного микроскопа с 3-го по 10-й день посева.
- Поставьте тарелку объемом 10 см3 на ступень микроскопа. Отрегулируйте увеличение микроскопа в 50 раз для всех сфероидов.
- Откройте программное обеспечение микроскопа на подключенном компьютере и отрегулируйте его фокус, чтобы иметь четкое изображение каждого сфероида. Используйте инструмент capture Tool на программном обеспечении микроскопа для сохранения полученных изображений.
7. Гомотипические сфероиды Hep3B в LX2 CM
- Подвешивайте клетки Hep3B HCC 3000 в подвесных каплях, чтобы сформировать сферы. Добавьте 10 мл стерильного PBS на дно тарелки, чтобы обеспечить влажные условия для сфероидов.
- Переверните крышку тарелки размером 10 см3 , чтобы среда, включая клеточную суспензию, висела над влажной средой. Оставьте висячие капли на 3 дня.
- Перенос сфероидов Hep3B в 20 мкл свежего СМ из клеток LX2 в подвесных каплях.
ПРИМЕЧАНИЕ: Используйте стерильные автоклавные нефильтрованные наконечники пипеток объемом 200 мкл в процессе переноса сфероидов, чтобы избежать каких-либо нарушений или травмированных сфероидов. Это также делается для устранения любых остаточных клеток, которые остаются непривязанными к основному одиночному сфероиду.- Используйте автоклавную пипетку объемом 20 мкл для процесса передачи. Отрегулируйте объем пипетки до 2 мкл. Прикрепите наконечник пипетки к пипетке.
- Перевернуть крышку тарелки объемом 10 см3 , на которой образовались сфероиды. Закрепите крышку на сцене светового микроскопа. Отрегулируйте тонкую фокусировку микроскопа, чтобы сделать каждый сфероид видимым.
- Осторожно очистите воздух от микропипетки, нажав кнопку плунжера. Вставьте наконечник пипетки в каплю, включая сфероид, подлежащую переноске. Подойдите очень близко к сфероиду, не касаясь его наконечником.
- Осторожно отпустите давление на кнопку плунжера, чтобы обеспечить всасывание сфероида в наконечник микропипетки в среде 2 мкл.
- Переведите сфероид в новую каплю, висящую на новой тарелке размером 10 см3 , имеющую новые носители/кондиционированные носители/обработку.
ПРИМЕЧАНИЕ: Убедитесь, что все сфероиды успешно перенесены на новую тарелку размером 10 см3 с помощью светового микроскопа.
- Делайте снимки сфероидов с 50-кратным увеличением с помощью инвертированного микроскопа со дня переноса (день 3) до 7-го дня культуры в LX2 CM.
8. Расчет объема сфероида
- Назначьте уникальный числовой идентификатор для каждого сфероида, чтобы изображения из соответствующих сфероидов можно было захватывать ежедневно.
- Анализируйте изображения растущих сфероидов с помощью программного пакета для анализа изображений.
- Откройте каждое сфероидное изображение в программном пакете. С помощью инструмента « Выделение от руки » и контур каждого сфероида. В раскрывающемся списке Анализ выберите Задать измерение, а затем Область. Нажмите OK.
- Вручную нарисуйте круг вокруг каждого сфероида. Как только сфера обведена, нажмите Ctrl + M , чтобы позволить программе вычислить сфероидную область в пикселях. Преобразуйте площадь сфероида в объем.
ПРИМЕЧАНИЕ: Объем сфероидмм3 =0,09403 ×
- Рассчитайте изменение объема сфероида относительно его объема в первый день захвата изображения.
ПРИМЕЧАНИЕ: Это делается для нормализации объема сфероида до начального объема и повышения точности с учетом естественного изменения начального размера.
Результаты
Клетки, культивируемые в многослойном 3D-формате, более точно отражают сложность микроокружения опухоли, чем обычные 2D-культуры24,25. Ранее многие исследования дополняли сфероиды культуральной среды различными митогенами и факторами роста26, чтобы инициировать образование сфероидов. В этом исследовании, однако, добавление фибробластов или их КМ обеспечивает необходимые митогены и факторы роста для ускорения роста сфероидов.
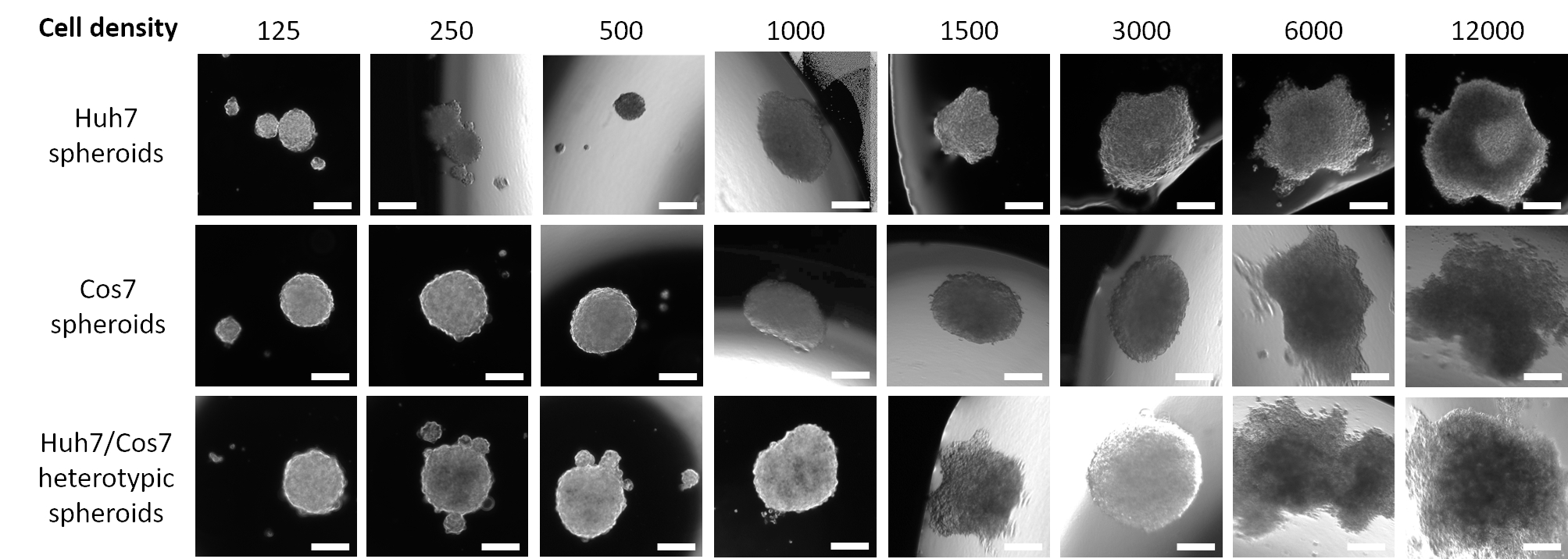
На рисунке 1 показаны данные пилотного исследования, в котором клетки HCC человека Huh7 были посеяны в нисходящей плотности посева, начиная с 12 000 до 125 клеток в свежих средах объемом 20 мкл в течение 3 дней. Плотность посева в 12 000 клеток дала сфероиды с асимметричной формой, которая не была исправлена даже после сокращения вдвое плотности посева клеток. Однако сфероиды, созданные из 3000 клеток, оказались более округлыми (рисунок 1, верхний ряд). Дальнейшее снижение концентрации клеток не привело к образованию единичных сфер (125, 250, 500 сфер клеточной плотности) и не имело правильного сферического вида (сферы клеточной плотности 1000 и 1500). Стоит упомянуть, что многие мелкие сферы образовались при плотности 500 опухолевых клеток; однако только один небольшой сфероид был захвачен при 50-кратном увеличении (рисунок 1, верхний ряд). Тот же оптимизированный протокол был применен к клеточной линии фибробластов приматов приматов COS7 (рисунок 1, средний ряд). Суспендирование 125, 250 и 500 фибробластов COS7 в висячих каплях 20 мкл приводило к образованию одного округлого сфероида с несколькими меньшими сфероидами, в то время как более высокие плотности клеток (1000, 1500 и 3000) образовывали одиночные полукругленные сфероиды. Подобно сфероидам опухоли Huh7, более высокие клеточные суспензии (6000 и 12000 клеток / сфера) приводили к образованию нерегулярных клеточных агрегатов, и, следовательно, более высокие концентрации клеток были отброшены (рисунок 1, средний ряд). Низкая плотность гетеротипических суспензий клеток Huh7/COS7 (125, 250 и 500 клеток/сфера) порождала один сфероид с несколькими плавающими или полуприсоединенными сфероидами (рисунок 1, нижний ряд). 1000- и 1500-клеточные гетеротипические сфероиды были полукруглыми, в то время как плотность посева 3000 клеток (1500 на тип клеток) давала округлый 3D-сфероид. Как отмечалось ранее, более высокая плотность клеток приводила к образованию агрегатов, а не четко определенных сфероидов (рисунок 1, нижний ряд). В заключение, 3000 сфероидов клеточной плотности были округлены, подобно опухолям ГЦК человека, и были адаптированы для дальнейших экспериментов. Культивирования клеточных суспензий в виде висячих капель в течение 3 дней было достаточно, чтобы способствовать образованию сфероидов.
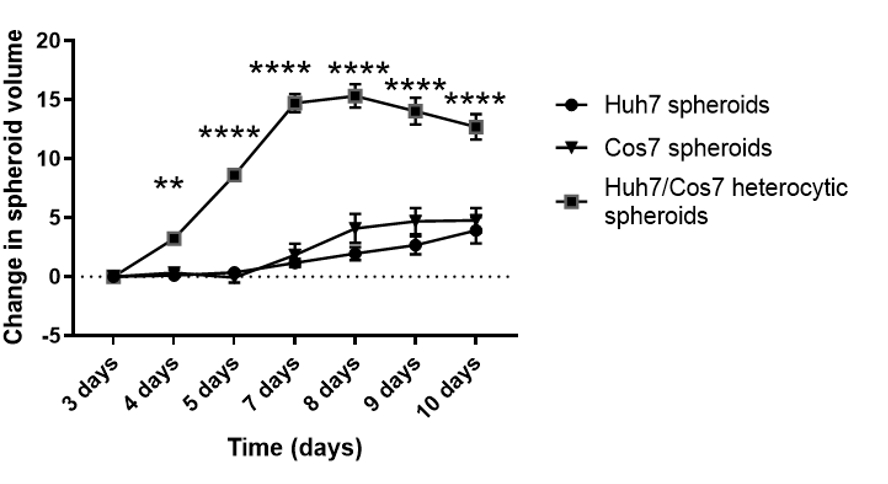
После оптимизации наилучшей концентрации клеток и временной точки для образования сфероидов в текущем контексте пролиферативное воздействие кокультурирования опухолевых и фибробластных клеточных линий оценивали продольно. На рисунках 2 и 3 показаны гетеротипические сфероиды Huh7/COS7 (3000 общих чисел клеток на сфероид), контролируемые с 3-го по 10-й день. Гомотипические сфероиды Huh7 и COS7 (по 1500 клеток в каждой) служили в качестве контроля. Начиная с 4-го дня, гетеротипические сфероиды росли в идеальной округлой форме по сравнению с гомотипическими сфероидами (рисунок 2). Первоначальный объем гетеротипического сфероида был больше, чем у каждого гомотипического сфероида. Рост сфероидов рассчитывали как изменение их объема относительно исходного объема в день образования сфероидов, чтобы избежать нормального изменения объема сфероидов (день 3, рис. 3). Гетеротипические сфероиды первоначально показали фазу быстрого роста, начиная с 4-го по 7-й день, за которой следовала более медленная фаза роста на 8-й день (рисунки 2, верхний ряд и рисунок 3). Объем сфероидов уменьшался на 9 и 10 день, возможно, отражая истощение питательных веществ или гипоксическое ядро и гибель клеток.
Напротив, гомотипические сфероиды Huh7 и COS7 росли гораздо медленнее (рисунок 2, средний и нижний ряды; Рисунок 3). Гомотипические сфероиды демонстрировали относительно статическую кривую роста до пятого дня культивирования (через два дня после образования сфероидов). Начиная с 6-го дня, гомотипические сфероиды начали демонстрировать постепенное увеличение своей кривой роста, хотя и со значительно более низкой скоростью, чем у гетеротипических сфероидов (рисунок 3). В заключение, гетеротипические сфероиды опухоли / фибробластов растут с более высокой скоростью, чем гомотипические сфероиды, предполагая, что прямой контакт опухолевых клеток и фибробластов увеличивает размер сфероидов опухоли.
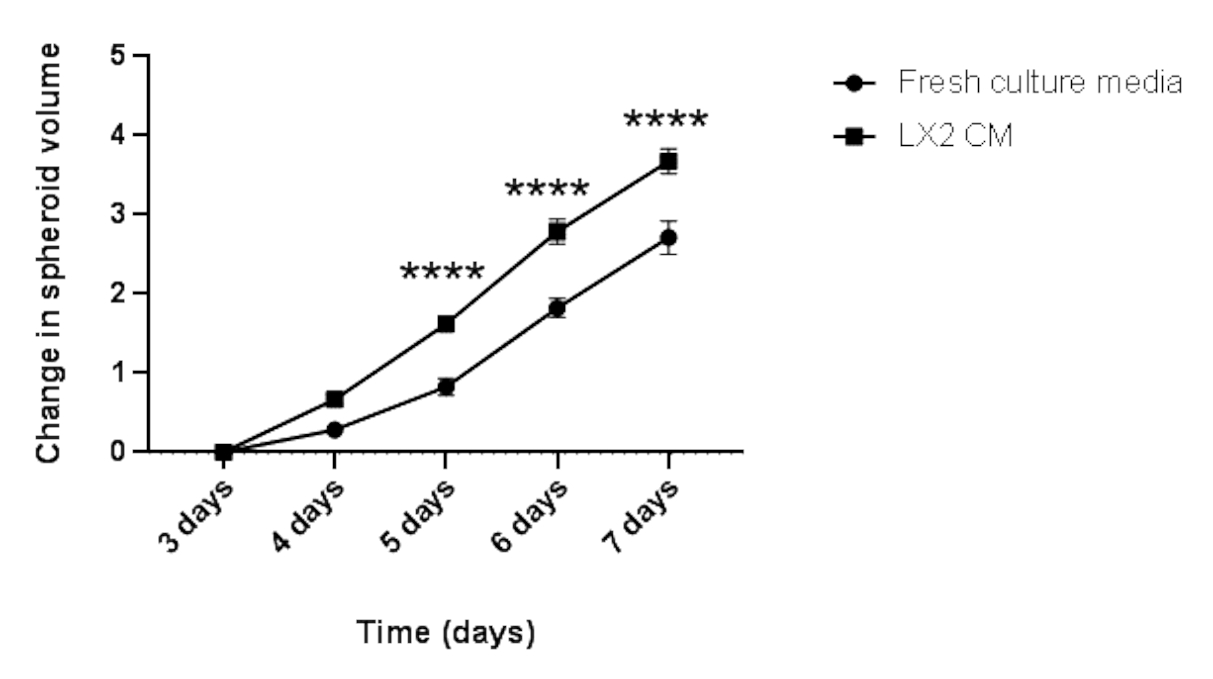
Наконец, чтобы подтвердить вышеуказанные результаты и изучить паракринное влияние мезенхимальных клеток на пролиферацию сфероидов ГЦК в контексте, связанном с печенью, 3-дневные гомотипические сфероиды Hep3B HCC выращивали в обычных свежих средах или CM из печеночных звездчатых клеток LX2 (Рисунок 4 и Рисунок 5) в течение дополнительных 4 дней. На первый взгляд, 3000 hep3B клеток образовали идеально округлые сфероиды через 3 дня (рисунок 4). Сфероиды Hep3B показали постоянное размножение в свежих средах с 3 по 7 день (рисунок 4, верхний ряд). Скорость роста была увеличена, когда сфероиды Hep3B поддерживались в LX2 CM (рисунок 4, нижний ряд). Это медиазависимое преобразование в росте сфероидов Hep3B продемонстрировало статистическую значимость с 4-го дня до конца эксперимента (рисунок 5), предполагая пролиферацию опухолевых сфероидов, вызванную фибробластами.
В заключение, это исследование успешно модифицировало существующие 3D-сфероидные культуры, чтобы использовать перекрестные помехи между опухолевыми клетками ГЦК и различными клеточными линиями фибробластов и исследовать пролиферативное значение этого прямого и косвенного клеточного взаимодействия (рисунок 6). Дальнейшая характеристика образовавшихся сфероидов необходима для улучшения взаимодействия опухолевых клеток с окружающей микросредой.

Рисунок 1: Оптимизация оптимальной плотности клеток для образования сфероидов. Столбцы представляют различную плотность заполнения ячеек, а строки представляют сфероиды, образованные из ячеек Huh7 (верхняя строка), ячеек COS7 (средняя строка) и гетеротипических ячеек Huh7/COS7 (нижняя строка). Изображения представляют сфероиды, образующиеся после приостановки 125, 250, 500, 1000, 1500, 3000, 6000 и 12000 клеток (слева направо) в среде 20 мкл в течение 3 дней. Пилотное исследование включало два сфероида на каждое условие, и изображения были сделаны с 50-кратным увеличением, шкала = 200 мкм. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}

Рисунок 2: Гетеротипический и гомотипический рост сфероидов. Столбцы представляют день, когда были сделаны сфероидные изображения, а строки показывают репрезентативные изображения для сфероидов, сформированных из ячеек Huh7 (верхняя строка), ячеек COS7 (средний ряд) и ячеек Huh7 / COS7 (нижний ряд). Изображения представляют сфероиды, образующиеся после приостановки 3000 клеток из каждого состояния (гомотипические Huh7, COS7 или гетеротипические сфероиды Huh7 / COS7) различных временных точек слева направо. Эксперимент включал 10 сфероидов на каждое условие, и изображения были сделаны с 50-кратным увеличением, шкала = 200 мкм. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}

Рисунок 3: Продольный анализ гетеротипического и гомотипического роста сфероидов. На графике показана кривая роста гетеротипических сфероидов Huh7/COS7 по сравнению с гомотипическими сфероидами Huh7 и COS7. Данные представляются в виде средних ± т.е.m; n = 10 независимых сфероидов. ** p < 0,01; p < 0,001. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}

Рисунок 4: Рост гомотипических сфероидов Hep3B в LX2 CM. Столбцы представляют день, когда были сделаны изображения сфероидов, а строки показывают репрезентативные изображения сфероидов Hep3B, культивированных в свежих питательных средах DMEM (верхний ряд) или LX2 CM (нижний ряд). Эксперимент включал семь сфероидов на каждое условие (свежие носители или LX2 CM), а изображения были сделаны с 50-кратным увеличением, шкала = 200 мкм. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}

Рисунок 5: Продольный анализ гомотипического роста сфероидов Hep3B. На графике показана кривая роста сфероидов Hep3B в свежих культуральных средах DMEM или LX2 CM. Данные представлены в виде среднего ± s.e.m; n = 7 независимых сфероидов. p < 0,001. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}

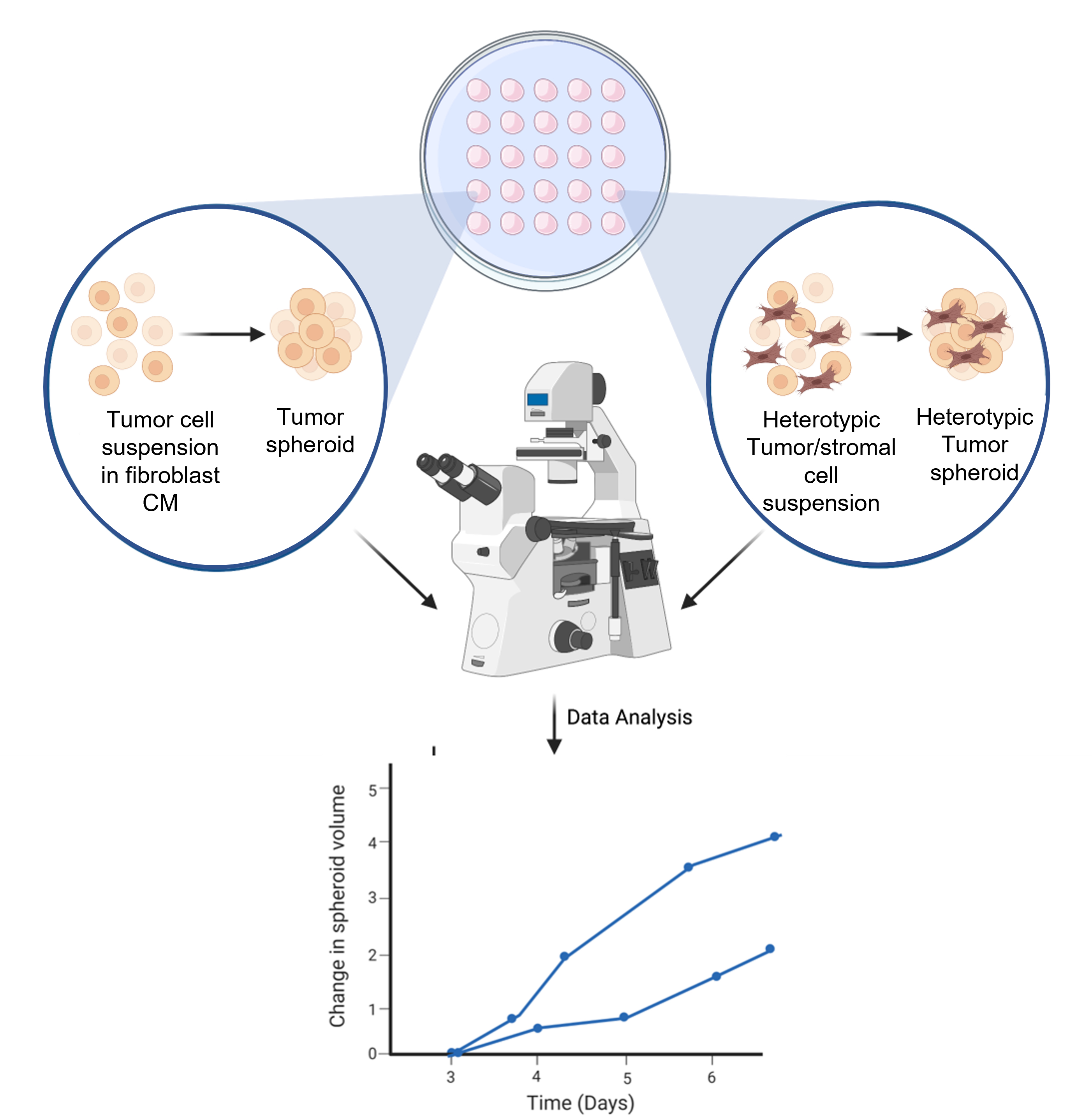
Рисунок 6: Схематическое изображение процесса формирования сфероидов. Клеточная суспензия пипетируется на внутренней крышке чашки Петри объемом 10 см3 . Крышка перевернута и хранится в течение 3 дней, чтобы обеспечить гомотипическое или гетеротипическое образование сфероидов. Сфероидные изображения делаются с 50-кратным увеличением. Рисунок создан с использованием веб-инструмента научной иллюстрации (см. Таблицу материалов). Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}
Обсуждение
Контекст, в котором растут экспериментальные клеточные линии, влияет на их профиль экспрессии генов, анализ путей и функциональные критерии. Например, в клетках рака молочной железы координация между различными онкогенными путями сохраняется только в том случае, если раковые клетки выращиваются в 3D-конформации27. Дерегулированные гены в 3D-меланоме и сфероидах рака молочной железы, но не в монослойных клетках, более актуальны для опухоли человека in vivo28,29. Например, уровни интегрина β1 в эпителиальных клетках молочной железы и опухолевых аналогах были ниже, чем уровни, выращенные в формате 2D29. Кроме того, фибробласты в 3D-структурах имеют тенденцию к отчетливой миграции с точки зрения морфологии и скорости по сравнению с теми, которые культивируются на пластике30. Кроме того, механическая жесткость ростового субстрата активирует специфические пути, которые стимулируют злокачественную трансформацию нормальных эпителиальных клеток путем дерегуляции внеклеточной сигнально-регулируемой киназы (ERK)/Rho в эпителиальных клетках31. Эти факторы способствуют переходу от обычной 2D-культуры к 3D-сфероидным моделям, поскольку 3D-культуры больше соответствуют заболеваниям человека.
В текущем исследовании использовались обычные лабораторные инструменты и расходные материалы для создания 3D-модели сфероида опухоли. Образовавшиеся сфероиды реагировали на сигналы пролиферации, поступающие от фибробластов, о чем свидетельствует их значительно увеличенный рост. Данная модель относится к категории многоклеточных сфероидов опухолей. Существует три другие категории 3D-сфероидов опухолей, включая tumorospheres32, тканевые опухолевые сферы33,34 и органотипические многоклеточные сфероиды35. В многоклеточных сфероидах опухоли опухолевые клетки суспендируются в условиях низкой адгезии, чтобы они могли объединяться вместе, образуя сферы, не касаясь дна культурального сосуда36. Для обеспечения этой независимой от крепления техники было использовано несколько подходов, начиная от вращающихся систем, методов жидкого наложения и непокрытых U-образных пластин со сверхнизким креплением37. В HCC большинство сфероидных культур in vitro используют сверхнизкие прикрепленные 24- или 96-луночные пластины для формирования округлых сфероидов18,19,20,21,22. Этот метод, однако, не является экономически эффективным и не исключает случайного контакта между клетками и пластиком. Другие системы используют транс-колодцы23 или срезы печени12 для производства микротканей печени. Использование человеческих тканей в трансляционной работе является золотым стандартом, но не всегда доступно или доступно многим исследовательским группам. Нынешний подход опирается на теорию висящих капель. Клеточную суспензию добавляют таким образом, что сфероиды опухоли образуются в доступном интерфейсе жидкость-воздух с образованием одиночных округлых сфер38. Преимуществами использования данной методики являются отсутствие какого-либо клеточно-пластического контакта и легкость изучения аутокринных и паракринных перекрестных помех между опухолевыми и фибробластными клетками. Эта модель была дополнительно подтверждена в другом исследовании, расшифровывающем важность непаренхимального TREM2 в защите печени от развития ГЦК39. Эта работа показала, что объем сфероидов Hep3B и PLC/PRF5 был выше в контроле LX2 CM по сравнению с TREM2-сверхэкспрессирующим LX2 CM в зависимости от Wnt39.
В текущем исследовании фибробласты были смешаны с опухолевыми клетками до образования сфероидов, чтобы исследовать пролиферативное воздействие этой кокультуры. Другие добавили фибробласты, эндотелиальные клетки и иммунные клетки с суспензией опухолевых клеток после формирования сфероида опухоли для изучения миграции стромальных клеток в опухоль40,41. Текущая гетеротипическая сфероидная модель показала картину роста, аналогичную ранее сообщенным моделям и исходной опухоли in vivo; Сфероиды показывают экспоненциальный рост, за которым следует фаза замедленного роста, скорее всего, из-за истощения питательных веществ и расширения некротического ядра42. Вместо того, чтобы добавлять факторы роста и митогены, чтобы помочь образованию сфероидов, в этом исследовании использовался более физиологический подход, поскольку эти факторы роста были получены от прямого контакта с фибробластами или их секретомом в ФИбробласте КМ. Важно отметить, что клетки LX2 могут быть дополнительно активированы путем обработки TGFβ1 или PDGF43. Клетки LX2 обрабатывали 10 нг/мл TGFβ1 в течение 48 ч перед удалением среды для различных экспериментальных условий. Клетки LX2 промывали три раза PBS (чтобы исключить любой прямой эффект TGFβ1), а затем добавляли свежие среды еще в течение 24 ч перед сбором КМ. CM, собранный из TGFβ1-стимулированного LX2, индуцировал рост сфероидов Hep3B с более высокой скоростью, чем CM из контрольного LX2 (данные не показаны). Эта гибкая система может поддаваться кокультурам различных раковых клеток плюс/минус фибробласты, а также линиям, полученным от пациента. Он также может предложить скрининговый анализ со средней пропускной способностью для лекарств, которые могут быть быстро приняты и вставлены в конвейер трансляционного обнаружения лекарств для информирования о дозировке для исследований in vivo.
Ограничением текущего исследования является использование увековеченных клеточных линий в образовании сфероидов, а не свежеизолированных опухолевых клеток ГЦК и связанных с раком фибробластов. Другим ограничением является сравнение роста гомотипического сфероида Hep3B между CM из LX2 и в свежих средах, а не CM из других нефибробластных клеточных линий. Последнее ограничение может быть устранено путем генетической модификации гена, представляющего интерес для фибробластов, с последующим применением КМ дикого типа по сравнению с генетически модифицированным фибробластом к гомотипическим сфероидам.
Раскрытие информации
FO является директором и акционером Fibrofind limited.
Благодарности
MYWZ финансируется Государственным секретарем по вопросам бизнеса, энергетики и промышленной стратегии и премией Ньютона 2020 года в рамках официальной помощи в целях развития Великобритании «ODA» и фонда Ньютона. SS поддерживается стипендией Cancer Research UK Advanced Clinician Scientist C53575/A29959. SS, FO и HR получают финансирование в рамках HUNTER, финансируемое через партнерство между Cancer Research UK, Fondazione AIRC и Fundacion Cientifica de la Asociacion Espanola Contra el Cancer.
Материалы
| Name | Company | Catalog Number | Comments |
| 1x Trypsin | Sigma Aldrich | 59429C | |
| 2 mL tubes | Eppendorf | ||
| Biorender | Biorender.com | Online | |
| Class II laminar flow BioMAT2 hood | Medair Technologies | ||
| Dulbecco’s modified Eagle’s medium (DMEM) | Biosera | M-D1107 | |
| Excel sofware | Microsoft office 365 | ||
| Graphpad prism sofware | GraphPad software | ||
| Heat deactivated fetal bovine serum (FBS) | life tech | 105000064 | |
| Image J software | Fiji | ||
| L-glutamine | Sigma Aldrich | G7513-100ml | |
| Penicillin/streptomycin (100 unit/ml penicillin and 100 μg/ml streptomycin) | Sigma Aldrich | P0781-100ml | |
| Petri dish 90 mm, triple vented | Greiner | 633175 | |
| Trypan blue | Sigma Aldrich | T8154 | |
| Virkon™ Broad Spectrum Disinfectant, 5kg Bucket | Rely+On™ | SCI-129999 | |
| Ziess inverted microscope | Ziess |
Ссылки
- Ferlay, J., et al. Cancer statistics for the year 2020: an overview. International Journal of Cancer. , (2021).
- Llovet, J. M., et al. Hepatocellular carcinoma. Nature Reviews Disease Primers. 7 (1), 6 (2021).
- Reeves, H. L., Zaki, M. Y., Day, C. P. Hepatocellular carcinoma in obesity, Type 2 diabetes, and NAFLD. Digestive Diseases & Sciences. 61 (5), 1234-1245 (2016).
- Forner, A., Reig, M., Bruix, J. Hepatocellular carcinoma. Lancet. 391 (10127), 1301-1314 (2018).
- Reig, M., et al. Diagnosis and treatment of hepatocellular carcinoma. Update of the consensus document of the AEEH, AEC, SEOM, SERAM, SERVEI, and SETH. Medicina Clinica (Barc). 156 (9), 1-30 (2021).
- Pfister, D., et al. NASH limits anti-tumour surveillance in immunotherapy-treated HCC. Nature. 592 (7854), 450-456 (2021).
- Llovet, J. M., et al. Trial design and endpoints in hepatocellular carcinoma: AASLD consensus conference. Hepatology. 73, 158-191 (2021).
- Llovet, J. M., Hernandez-Gea, V. Hepatocellular carcinoma: reasons for phase III failure and novel perspectives on trial design. Clinical Cancer Research. 20 (8), 2072-2079 (2014).
- Brown, Z. J., Heinrich, B., Greten, T. F. Mouse models of hepatocellular carcinoma: an overview and highlights for immunotherapy research. Nature Reviews Gastroenterology and Hepatology. 15 (9), 536-554 (2018).
- Nikolic, M., Sustersic, T., Filipovic, N. In vitro models and on-chip systems: Biomaterial interaction studies with tissues generated using lung epithelial and liver metabolic cell lines. Frontiers in Bioengineering and Biotechnology. 6, 120 (2018).
- Clark, A. M., Ma, B., Taylor, D. L., Griffith, L., Wells, A. Liver metastases: Microenvironments and ex-vivo models. Experimental Biology and Medicine. 241 (15), 1639-1652 (2016).
- Paish, H. L., et al. A bioreactor technology for modeling fibrosis in human and rodent precision-cut liver slices. Hepatology. 70 (4), 1377-1391 (2019).
- Brassard-Jollive, N., Monnot, C., Muller, L., Germain, S. In vitro 3D systems to model tumor angiogenesis and interactions with stromal cells. Frontiers in Cell and Developmental Biology. 8, 594903 (2020).
- Pampaloni, F., Reynaud, E. G., Stelzer, E. H. The third dimension bridges the gap between cell culture and live tissue. Nature Reviews Molecular Cell Biology. 8 (10), 839-845 (2007).
- Kleinman, H. K., Philp, D., Hoffman, M. P. Role of the extracellular matrix in morphogenesis. Current Opinion in Biotechnology. 14 (5), 526-532 (2003).
- Bissell, M. J., Radisky, D. C., Rizki, A., Weaver, V. M., Petersen, O. W. The organizing principle: microenvironmental influences in the normal and malignant breast. Differentiation. 70 (9-10), 537-546 (2002).
- Ronnov-Jessen, L., Petersen, O. W., Koteliansky, V. E., Bissell, M. J. The origin of the myofibroblasts in breast cancer. Recapitulation of tumor environment in culture unravels diversity and implicates converted fibroblasts and recruited smooth muscle cells. Journal of Clinical Investigations. 95 (2), 859-873 (1995).
- Shao, H., et al. A novel stromal fibroblast-modulated 3D tumor spheroid model for studying tumor-stroma interaction and drug discovery. Journal of Visualized Experiments: JoVE. (156), e60660 (2020).
- Song, Y., et al. Activated hepatic stellate cells play pivotal roles in hepatocellular carcinoma cell chemoresistance and migration in multicellular tumor spheroids. Scientific Reports. 6, 36750 (2016).
- Pingitore, P., et al. Human multilineage 3D spheroids as a model of liver steatosis and fibrosis. International Journal of Molecular Sciences. 20 (7), (2019).
- Kozyra, M., et al. Human hepatic 3D spheroids as a model for steatosis and insulin resistance. Scientific Reports. 8 (1), 14297 (2018).
- Khawar, I. A., et al. Three dimensional mixed-cell spheroids mimic stroma-mediated chemoresistance and invasive migration in hepatocellular carcinoma. Neoplasia. 20 (8), 800-812 (2018).
- Feaver, R. E., et al. Development of an in vitro human liver system for interrogating non-alcoholic steatohepatitis. Journal of Clinical Investigations Insight. 1 (20), 90954 (2016).
- Wartenberg, M., et al. Regulation of the multidrug resistance transporter P-glycoprotein in multicellular tumor spheroids by hypoxia-inducible factor (HIF-1) and reactive oxygen species. Federation of American Societies for Experimental Biology Journal. 17 (3), 503-505 (2003).
- Chen, R., et al. Screening candidate metastasis-associated genes in three-dimensional HCC spheroids with different metastasis potential. International Journal of Clinical Experimental Pathology. 7 (5), 2527-2535 (2014).
- Venkataraman, G., et al. Preferential self-association of basic fibroblast growth factor is stabilized by heparin during receptor dimerization and activation. Proceedings of the National Academy of Sciences of the United States of America. 93 (2), 845-850 (1996).
- Bissell, M. J., et al. Tissue structure, nuclear organization, and gene expression in normal and malignant breast. Cancer Research. 59, 1757-1763 (1999).
- Ghosh, S., et al. Three-dimensional culture of melanoma cells profoundly affects gene expression profile: a high density oligonucleotide array study. Journal of Cellular Physiology. 204 (2), 522-531 (2005).
- Delcommenne, M., Streuli, C. H. Control of integrin expression by extracellular matrix. Journal of Biological Chemistry. 270 (45), 26794-26801 (1995).
- Meshel, A. S., Wei, Q., Adelstein, R. S., Sheetz, M. P. Basic mechanism of three-dimensional collagen fibre transport by fibroblasts. Nature Cell Biology. 7 (2), 157-164 (2005).
- Paszek, M. J., et al. Tensional homeostasis and the malignant phenotype. Cancer Cell. 8 (3), 241-254 (2005).
- Yu, M., et al. Cancer therapy. Ex vivo culture of circulating breast tumor cells for individualized testing of drug susceptibility. Science. 345 (6193), 216-220 (2014).
- Weiswald, L. B., et al. Newly characterised ex vivo colospheres as a three-dimensional colon cancer cell model of tumour aggressiveness. British Journal of Cancer. 101 (3), 473-482 (2009).
- Kondo, J., et al. Retaining cell-cell contact enables preparation and culture of spheroids composed of pure primary cancer cells from colorectal cancer. Proceedings of the National Academy of Sciences of the United States of America. 108 (15), 6235-6240 (2011).
- Rajcevic, U., et al. Colorectal cancer derived organotypic spheroids maintain essential tissue characteristics but adapt their metabolism in culture. Proteome Sciences. 12, 39 (2014).
- Friedrich, J., Seidel, C., Ebner, R., Kunz-Schughart, L. A. Spheroid-based drug screen: considerations and practical approach. Nature Protocols. 4 (3), 309-324 (2009).
- Friedrich, J., Ebner, R., Kunz-Schughart, L. A. Experimental anti-tumor therapy in 3-D: spheroids--old hat or new challenge. International Journal of Radiation Biology. 83 (11-12), 849-871 (2007).
- Kelm, J. M., Timmins, N. E., Brown, C. J., Fussenegger, M., Nielsen, L. K. Method for generation of homogeneous multicellular tumor spheroids applicable to a wide variety of cell types. Biotechnology & Bioengineering. 83 (2), 173-180 (2003).
- Esparza-Baquer, A., et al. TREM-2 defends the liver against hepatocellular carcinoma through multifactorial protective mechanisms. Gut. 70 (7), 1345-1361 (2021).
- Dangles-Marie, V., et al. A three-dimensional tumor cell defect in activating autologous CTLs is associated with inefficient antigen presentation correlated with heat shock protein-70 down-regulation. Cancer Research. 63 (13), 3682-3687 (2003).
- Timmins, N. E., Dietmair, S., Nielsen, L. K. Hanging-drop multicellular spheroids as a model of tumour angiogenesis. Angiogenesis. 7 (2), 97-103 (2004).
- Mayer, B., et al. Multicellular gastric cancer spheroids recapitulate growth pattern and differentiation phenotype of human gastric carcinomas. Gastroenterology. 121 (4), 839-852 (2001).
- Xu, L., et al. Human hepatic stellate cell lines, LX-1 and LX-2: new tools for analysis of hepatic fibrosis. Gut. 54 (1), 142-151 (2005).
Перепечатки и разрешения
Запросить разрешение на использование текста или рисунков этого JoVE статьи
Запросить разрешениеСмотреть дополнительные статьи
This article has been published
Video Coming Soon
Авторские права © 2025 MyJoVE Corporation. Все права защищены