
Method Article
Микроманипуляции методы, позволяя анализ морфогенетических динамики и оборот цитоскелета регуляторов
В этой статье
Резюме
Мы опишем, как микро - и добавлены методы, такие как FRAP и photoactivation включить определение параметров подвижности и пространственно-временных динамики белков внутри миграции клеток. Экспериментальная отсчетов включают субцеллюлярные динамика и оборот регуляторов подвижности или базовой Цитоскелет актина.
Аннотация
Изучение пространственно-временных динамики белков может выявить их функциональное значение в различных контекстах. В этой статье это обсуждается как флуоресцентные восстановления после Фотообесцвечивание (FRAP) и photoactivation методов может использоваться для изучения пространственно-временных динамики белков в субцеллюлярные местах. Мы также показать, как эти методы позволяют простым определение различных параметров, связанных с актина цитоскелета регулирования и ячейки подвижности. Кроме того микроинъекции клеток дополнительно описывается как альтернативное лечение (потенциально предшествующий или дополнить вышеупомянутые добавлены методы) для триггера мгновенного воздействия арестовано белков на ячейку Морфология и функции. Микроманипуляции такие инъекции белков или местное применение плазматической мембраны проницаемой наркотиков или цитоскелета ингибиторов может служить мощным инструментом для записи непосредственных последствий данного лечения на поведение ячеек в одну ячейку и внутриклеточных уровне. Примером этого является здесь немедленной индукции lamellipodial выступ края ячейки путем инъекций рекомбинантных Rac1 белка, как установлено четверть века назад. Кроме того, мы предоставляем протокол для определения оборот расширенной зеленого флуоресцентного белка (EGFP)-VASP, полимеразы накаливания актина, заметно накапливается в lamellipodial советы B16-F1 клеток, используя FRAP, включая связанные данные анализ и кривой. Мы также представляем руководящие принципы для оценки показателей lamellipodial актина сети полимеризации, примером чему клетки, выражая EGFP-тегами β-миозина. Наконец даны указания о том, как исследовать показатели актина мономера мобильности в цитоплазме клетки, следуют актина включения на сайты быстрого накаливания Ассамблеи, например советы выступающих lamellipodia, с помощью photoactivation подходы. Ни один из этих протоколов ограничивается компонентами или регуляторы Цитоскелет актина, но может быть легко расширен для изучения в аналогичных моды пространственно-временных динамика и функции белков в различных различных внутриклеточных структур или функциональных контексты.
Введение
Мониторинг динамики пространственно-временных белков и других молекул в живых клетках стала важным инструментом во многих областях клеточной и молекулярной биологии. Расширенный флуоресцентной микроскопии методов, включая передачи энергии резонансной флюоресценции (ЛАДА) и лад Флуоресценция жизни изображений (Лада-FLIM), или FRAP, флуоресценции потери в Фотообесцвечивание (FLIP) и photoactivation, а также многие другие позволяют для временных и пространственных Отслеживание взаимодействий протеин протеина, конформационные изменения, а также определения кинетика диффузии и локализация различных белков в ячейку1,2. FRAP и photoactivation методы, в частности, широко применяются для изучения регуляторов актина цитоскелета и клеточной миграции. Эти методы могут применяться отдельно или в сочетании с дополнительной микроманипуляции методы, такие как микроинъекции3и включать выражение дневно меченых белков. Они позволяют производить оценку кинетики ассоциации белка актин богатых структур участвующих в ячейке миграции, например filopodia или lamellipodia, оборот белков в фокуса спайки4, или разветвленных сетей актина5. Они также дают возможность определения lamellipodial актина полимеризации ставок, оценки дисперсии мономерных актина в цитозоле, скорость субцеллюлярные актина транслокации мономер для полимеризуя филаментов актина в выступающие lamellipodia6и другие параметры.
FRAP — это метод для визуализации и количественной оценки подвижности белков в живой клетке, первоначально разработанная в 1970-х Аксельрод7. Область интереса (ROI) в ячейке, заполненный дневно меченых белков, временно подвергается лазер высокой интенсивности, достаточно, чтобы вызвать отбеливания Флюорофор молекул, присутствующих в этом регионе данный короткий период времени. Небеленой, дневно помечены белки, расположенные за пределами ROI во время отбеливания, диффузный и проникнуть отбеленной региона в зависимости от их пространственно-временных динамика, вызывающих перемещение молекул photobleached с течением времени. Скорость восстановления флуоресцентным в беленой регионах зависит от различных факторов, включая размер и скорость распространения данной молекулы, и конечно его текучести кадров в рамках предполагаемого связанные отбеленной структуры. Таким образом растворимые белки будет посредничать восстановления флуоресцентным в беленой ROI быстро путем диффузии, а белки, тесно связанные с структуры, такие как фокуса спайки, будет иметь больше оборот раз, как их флуоресценции восстановления будет зависеть на диффузии растворимых фракции белков и диссоциации Ассоциация кинетики связанные структуры дроби. Флуоресценции восстановления обычно приобретается и количественно вплоть до начального уровня до отбеливатель интенсивности флуоресценции. Однако это не происходит, если является частью первоначального флуоресценции интенсивности принадлежит к так называемой неподвижной фракция, которая не может быть пополнен путем диффузии или пополнения очень медленными темпами по сравнению с большинство молекул, включая мобильные фракция. Чтобы определить скорость оборота белка, генерируются FRAP кривых, представляющий степень восстановления флуоресценции со временем. От этих кривых восстановления можно рассчитать среднее раза белка восстановления. Создание кривой подходит средний FRAP данных, и следовательно математического анализа, это также можно вывести ли средняя текучесть мобильных фракции представляет собой совокупность одной однородность населения молекул, или же она состоит из два или более субпопуляций молекул, перебирая на дифференциальных ставок. Помимо оценки белка текучесть количественные подходы, отслеживание восстановления photobleached регионов в lamellipodia также можно разрешить для точной количественной оценки параметров подвижности lamellipodial как Ретроградная потока, выступ, и тарифы полимеризацию миозина. Таким образом FRAP представляет собой универсальный инструмент для применяться для оценки различных параметров в пределах структуры живых клеток.
Photoactivation-это метод, используемый для отслеживания диффузии и подвижности белков или молекул, возникая от назначенного сотовой местоположения. Техника использует, например, вариант Зеленый флуоресцирующий белок одичал тип (КГВ), первоначально разработанный Паттерсон и Lippincott-Шварц8, которые мутировали в манере, которая позволяет его флуоресценции сильно возрасти после воздействия ультрафиолетового (УФ) света (около 400 Нм; здесь, 405 нм). Как описано в Паттерсон и др., одичал типа КГВ хромофоры существуют как смешанное население нейтральных фенолов и анионными фенолятам, которые производят крупные поглощения пика примерно 397 Нм и незначительные один на 475 Нм, соответственно. После облучения белка с ультрафиолетовым светом население подвергается photoconversion, смещается в сторону анионной форме. Когда возбуждается 488 нм, белок photoconverted/photoactivated экспонатов вывозимому увеличение в флуоресцировании, недостаточно на практике различие между активирована и неактивированные GFP благодаря высокой внутренней фон флуоресценции. Однако снижение интенсивности фона было достигнуто путем введения единого аминокислоты мутации в последовательность GFP (гистидин замещение позиции 203). Полученный T203H мутант, известный также как photoactivatable-GFP (ПА-GFP) характеризуется значительным снижением поглощения незначительные пик, который после облучения с УФ-излучения увеличивается почти в 100 раз, когда впоследствии возбужденные 488 нм свет. Следовательно гиперэкспрессия PA-GFP-тегами белков является широко используется подход, который позволяет определение диффузии и подвижность молекул в клетках. Мы ранее подали ПА-GFP-тегами актина, чтобы определить скорость рассеяния мономеров актина цитозольной районов, позволяя не только изучение их мобильности в цитозоле, но и уровень их включения в выступающие lamellipodial актина сети6. Более последние литературе также описывает роман, Фото кабриолет белки, которые в принципе могут быть использованы в аналогичных моды, но укрывательство потенциальное преимущество быть видны уже до фото преобразования. Примеры для этой группы флуоресцентных белков включают Dendra2 и mEos29,10,11,12.
В этой статье мы объясним методологии microinjecting клеток с белками. Мы также объяснить, как эта техника может сочетаться с FRAP, Фотообесцвечивание белков, участвующих в регулировании Цитоскелет актина и подвижности, и каким образом могут быть получены FRAP кривых и половина время восстановления мобильных фракций. Кроме того мы предоставляем пример того как FRAP техника может использоваться для определения ставок актина полимеризации lamellipodial сетей. Мы также предоставляем инструкции и советы о том, как выполнять photoactivation экспериментов, которые могут быть использованы для определения цитозольной мобильность мономерных актина и ставки актина включения в lamellipodia. Эти методы, конечно же, являются не только ограничены для отслеживания компонентов Цитоскелет актина, но после оптимизации или потенциально требуется умеренной адаптации, могут широко применяться для других типов клеток, или для расследования различных белков, структур, и параметры.
протокол
1. Coverslip мытье и стерилизация
- Погружать 15 мм (диаметр) крышка очки (№ 1) в 500 мл флакон, содержащий смесь 40 мл 37% HCl и 60 мл 100% EtOH (не более 100 coverslips 100 мл моющего раствора).
Примечание: даже если свеже приобрели, coverslips необходимо строго очистить перед посевом клетки на их поверхности. Это потому, что они могут содержать тонких пленок смазка, которых макроскопически невидимы, но могут эффективно вмешиваться адгезии и соответствующие распространение живых клеток. В то время как такие фильмы могут быть эффективно удалены с растворами, содержащими кислоты или базы (см. Фишер и др. 13), мы обычно используют смесь кислоты/алкоголя, описанных выше. - Встряхните флакон, содержащий крышку очки для 30 мин на вращение шейкер. Выберите скорость, что позволяет Обложка очки, чтобы быть кружились свободно, но медленно достаточно, чтобы избежать частых ломать. Фильтрата раствора для удаления частей сломленного стекла, если повторно.
- Передача очки покрытия на флакон, содержащий по меньшей мере 200 мл стерильной воды и инкубировать на вращение шейкер, при этом неоднократно заменяя воду до тех пор, пока кислотный запах исчезла. Для полной ликвидации следов HCL-EtOH рекомендуется несколько стирок в течение нескольких часов.
- Сухие отдельных очки покрытия на листе бумаги.
- Место крышку очки в нижней части Петри 10 см (диаметр), покрытые фильтровальная бумага и тепла сухой стерилизации. Избегайте автоклавирования, как это приведет к обложке очки, чтобы держаться вместе.
2. Лечение клеток, Transfection и посев на Coverslips
- Вырасти клетки меланомы В16-F1 мыши согласно стандартной ячейки культуры условий в среде DMEM (4,5 г/Л глюкозы), содержащий 10% плода телячьей сыворотки, глютамин 2 мм и 1% пенициллин стрептомицина при 37 ° C, 7% CO2.
- Растут NIH3T3 фибробластов клетки для микроинъекций согласно стандартной ячейки культуры условий (инкубатор культуры ткани при 37 ° C, 7% CO2) в среде DMEM (4,5 г/Л глюкозы) содержащий 10% плода бычьим сывороточным, пируват натрия 1 мм, 1 x MEM несущественные аминокислот , глютамин 2 мм и 1% пенициллин стрептомицином.
- Для transfections растут B16-F1 клетки до 100% впадения в 10 см блюдо и проход в соотношении 1:5 в 3 см (диаметр) пластиковые блюдо.
- В тот же день после того, как клетки B16-F1 разрешалось следовать для по крайней мере 6 h, transfect с 500 нг/блюдо photoactivatable ПА-GFP-актин или меткой EGFP β-актина плазмидной ДНК. Для совместного transfections PA-GFP-актина с mCherry кодирования векторов Смешайте в общей сложности 1 мкг плазмидной ДНК на 3 см блюдо.
- Transfect клетки B16-F1 с трансфекции реагента (Таблица материалов). Для 3 см блюдо, смешать 200 мкл 150 мм NaCl, содержащий 500 нг ДНК построить с 200 мкл 150 мм NaCl, содержащие 1 мкл Реагента трансфекции (т.е., ДНК (мкг): был использован реагента (мкл) соотношение 1:2).
- Инкубируйте трансфекции смесь для 20 мин при комнатной температуре (RT) и пипетки каплям на 3 см блюдо, содержащий ячейки. Аккуратно водоворот блюдо смешать и инкубировать на ночь при 37 ° C, 7% CO2.
- Подготовьте буфер покрытия Ламинин, содержащий 50 мм трис, рН 7,4 и 150 мм NaCl.
- Для ячеек, B16-F1 пальто 15 мм крышка стекла, распространяя 150 мкл Ламинин (25 мкг/мл в буфере Ламинин покрытие) и инкубировать 1 час на RT. Для NIH3T3 клеток пальто очки покрытия раствором фибронектина (25 мкг/мл в фосфат амортизированное saline (PBS)) и инкубировать 1 час на RT.
- Вымойте инкубировали Ламинин или фибронектин очки покрытия с PBS, затем аспирационная PBS и добавить 2 мл transfected клеток.
- Семя transfected клеток B16-F1 (в 1:30 отношение от вырожденная блюдо), на следующий день после трансфекции, на Ламинин покрытием coverslips. Семя NIH3T3 фибробластов (в 1:20 соотношение от вырожденная блюдо) на фибронектин покрытием coverslips.
- Позволяют клеткам выкладывают на Ламинин покрытием или фибронектин очки покрытия на ночь в инкубатор культуры ткани при 37 ° C до микроскопии. Кроме того микроскопия экспериментов может начаться в тот же день, учитывая, что клетки могут распространяться по крайней мере 2-3 ч.
3. Ассамблея микроскопии изображений камеры
- Использование тепла проводниковые алюминиевые RC-26 изображений камеры для микроскопии(рисунок 1). Смажьте силиконовой смазкой вокруг контура пластиковый уплотнитель открытия с помощью шприца (рис. 1б).
- Место покровным стеклом с клетки стороной вверх на камеры (рис. 1c).
- Поместите пластиковый уплотнитель на верхней части крышки стекла сделать безопасный уплотнение между coverslip и камеры. Исправьте пластиковый уплотнитель (по диагонали, чтобы избежать поломки coverslip) путем завинчивания Скользящие зажимы на камеры, чтобы избежать утечки (рис. 1d) среднего.
- Пипетка 37 ° C среднего подогретую микроскопии в районе Центральный. Для средних сократилось в аутофлюоресценция и таким образом оптимизирован для микроскопии используйте тот же самый рецепт в качестве питательной среды, описанные выше, но с F12-HAM вместо DMEM, дополнительно содержит 20 мм HEPES для культивирования клеток в отсутствие CO2 (Рисунок 1e).
- Вставьте тепловой извещатель в назначенный слот камеры и связать электродами камеры TC-324B автоматический регулятор температуры поддержание постоянной температуре 37 ° C (рис. 1f).
- Место небольшой каплю масла погружения на цель и разместить камеру на вершине.
- Инкубируйте в камеру с ячейками для по крайней мере 10 – 30 мин позволить им оправиться от падение температуры во время монтажа и адаптироваться к микроскопии среднего.
- До того, как инициируется микроскопии, замените питательной среды в центральный резервуар палаты (примерно 800 мкл) чтобы избежать неуместной концентрации компонентов среды и сыворотке крови за счет средних испарения. Продолжительного микроскопии сессий с открытой камеры потребует обычные изменения испарения среды.
4. микроинъекции процедура
- Пальто coverslips, подготовить клетки и собрать тепловизионной камеры, как описано выше.
- Оттепель Алиготе очищенный протеин для инъекций (обычно 10 мкл или меньше) и разбавить его с соответствующим микроинъекции буфера.
Примечание: Состав буфера может в зависимости от тип протеина и ячейки, но заботиться, чтобы использовать рН между 8.00 и 6.95 и избегать использования PBS, как большинство клеток не нравится быть введен с PBS. - Для Rac1 микроинъекции Подготовьте буфер, содержащий 100 мм NaCl, 50 мм трис-HCl рН 7,5, 5 мм2MgCl 1 DTT. Mg2 + ионы необходимы для небольших ГТФазы стабильности.
Примечание: Концентрации белка обычно варьируется от 0,1 – 1 мг/мл (максимум 2 мг/мл), в зависимости от белка, типа эксперимента и клетки типа. - Если применимо, добавьте Люминесцентную краску, таких как инертный декстрана (0,5 мкг/мл, 70 кДа) белка решение, которое может подтвердить наличие иглы потока до инъекции и разрешает документация успешно инъекции после эксперимента.
Примечание: Эксперимент здесь направлена не на после динамики вводят Rac1, который возможно только после прямого флуоресценции маркировки белка. Муфта белков с флуоресцентными красителями или фьюжн флуоресцентный белок можно, но избегать здесь, как он укрывает риск вмешательства с функцией сигнализации, в частности малых белков как Rac1 ГТФазы Ро семья (20 кДа). - Центрифуга раствор белка на 10000 x g для по крайней мере 30 минут для удаления белка агрегаты, которые могут привести к иглы засорения если присутствует в микроинъекции капилляров.
- Загрузите микроинъекции иглы (микроинъекции капилляров) с 1 мкл инъекции смеси с обратной стороны, используя кончик гибкий пипеткой наконечник/microloader.
- Если в кончик иглы присутствуют пузырьки воздуха, аккуратно нажмите иглы для того, чтобы удалить их. Действовать быстро, чтобы избежать высыхания кончика иглы, которая может привести к засорению иглы.
- Тщательно Отрегулируйте иглодержатель на устройстве микроманипуляции. Если с помощью инвертированного микроскопа для фазы контрастность изображений перед загрузкой иглы, убедитесь, что имеется достаточно места для перемещения иглы вверх и вниз без препятствует Микроскоп конденсатора.
- После завинчивания микрокапиллярной на иглодержатель, обратиться иглы с помощью устройства микроинъекции давления до translocating кончик иглы в ячейку культуры среднего давления (20 – 50 гПа фон).
Примечание: Активация давление, когда игла находится в среде приведет к среде, будучи высосанный капиллярные силы и таким образом запрещают инъекционного раствора интерес. - Положение иглы в поле зрения (при содействии с помощью малое увеличение целей). 40 X сухой цель для микроинъекции экспериментов был использован здесь.
- Макроскопически позиционировать кончик иглы в вертикальном положении по отношению к середине объектива (это ускорит нахождение кончик иглы). Используйте микроскоп с оптикой фазово контрастной переместите кончик иглы в горизонтальной плоскости по отношению к поле зрения, в оптических самолет хорошо выше слоя клеток.
Примечание: Игла будет первоначально появляются как тень в поле зрения и плоскости фокуса затем могут быть скорректированы для визуализации кончик. Найдя кончик иглы, постепенно ниже плоскости оптической следуют кончик иглы вплоть до позиции вблизи слоя клеток. - Проверьте поток иглу, переключившись на флуоресцентные канал при использовании люминесцентных декстрана и настроить поток, используя устройство давления для получения потока постоянной «фон».
Примечание: В этой статье мы опишем ручной инъекция, которая при посредничестве прорываясь через плазматическую мембрану через прикосновения клетки поверхности и нежные движения кончика иглы во время постоянной иглы потока. Это следует отличать от устройств автоматического инъекций сопровождается запрограммированных иглы снижение и иглы повышения давления во время впрыскивания, которые больше подходят для инъекций выше числа клеток, следуют более поздних популяции клеток анализ. Метод, описанный здесь оптимизирован для одной ячейки анализа путем покадровой микроскопии до, во время и после микроинъекции. - Найдите ячейку интерес и постепенно опустите иглу над ячейкой.
- Когда все готово для microinject, опустите иглу постепенно в направлении района perinuclear клетки, с помощью тонкой шестерни микроманипулятор джойстик, сохраняя клетки в фокусе.
- Микроинъекции нежно коснуться плазматической мембраны клетки, которые могут достаточно, чтобы проникнуть в клетки, или помощи переходных плодного очень нежный крана на Микроскоп setup.
Примечание: Белая точка на кончик иглы будет указано время контакта с плазматической мембраны; После разрыва мембраны, запечатывает кончик иглы, сопровождается нежный поток инъекционного раствора в клетку. - Остановить процесс инъекции, как только поток в ячейку видимой (в идеале в пределах 0.3 s) путем перемещения вверх кончик иглы в среду. При использовании люминесцентных декстрана, успешно инъекции могут быть немедленно документально флуоресценции.
- При желании, инициировать захвата изображений в промежуток времени до или после микроинъекции.
Примечание: Местное применение наркотиков или ингибиторы могут быть выполнены на все шаги здесь, за исключением микроинъекции событие само по себе. Для местного применения диффузии активные молекулы могут быть под контролем давления потока и документально подтверждено флуоресценции, и кончик иглы могут располагаться на нужной высоте. Примеры местных приложений экспериментов увидеть например, малые и Rottner14 или Kaverina et al. 15 - После микроинъекции подождать, пока эффект белка происходит. Для различных белков и в зависимости от ожидаемого результата время инкубации может варьироваться. Для небольших ГТФазы Rac1 ответ lamellipodium формирования могут быть начатые в течение 1 мин или менее, но занимает около 10-15 мин в среднем в полной мере развивать (рис. 1g, h).
- Судья жизнеспособность клеток после микроинъекции.
Примечание: Неуместным или вредных инъекции может вызвать повреждения клеток, который часто сопровождается неспецифической клеток край опровержения или разрыв плазматической мембраны.- Избегайте, poking через верхней и нижней плазматической мембраны, которая может возникнуть для инъекций в плоской сотовой регионах.
Примечание: Инъекции томов должны быть сведены к минимуму (в идеале < 5% объема сотовых) и обычно будет в диапазоне femtoliter. Необходимых объемов закачки может также управляться изменения концентрации, но обратите внимание, что для белков, концентрации > 2 мг/мл может стать непрактичным ввиду частых иглы засорения. Однако это также зависит от качества и поведение очищенный протеин; например, инъекции дневно в сочетании актина осложняется зависит от концентрации и неизбежным полимеризации в кончик иглы, и поэтому она редко выполняется сегодня (см. Малые и др. 16).
- Избегайте, poking через верхней и нижней плазматической мембраны, которая может возникнуть для инъекций в плоской сотовой регионах.
- Раньше, во время или после эффект микроинъекции, FRAP или photoactivation могут быть выполнены на той же ячейки (см. раздел 5 и 6).
5. FRAP процедура
- Transfect тип ячейки интерес (здесь клетки B16-F1) с плазмидной ДНК кодирования дневно тегами протеин интереса (здесь, используется с тегами EGFP версия β-актина). Семя клетки на Ламинин покрытием coverslips (шаг 2.10).
- Соберите тепловизионные камеры (раздел 3).
- Используйте следующие параметры для lamellipodial региона Фотообесцвечивание: 65 МВт мощность лазера (переменная по экспериментальной установки и лазерного источника); Диаметр луча лазера 10 пикселей; 1 мс отбеливатель выдержки времени/пиксель; время экспозиции GFP 500 мс; интервал времени 1500 мс. Экспериментальные результаты в этом документе были проведены с целью apochromatic 100 X 1.4NA.
- Выполняйте Лазерная калибровка для обеспечения точности в размеры области photobleached. До калибровки переместить поле зрения в районе не хватает любой сигнал клетки/флуоресценции и наблюдать изображение на дисплее.
- Выберите цель масштаб, нажав на кнопку соответствующего увеличения и уменьшения мощности лазера (3 – 5 МВт) в «группа | Интенсивность» меню. Чтобы инициировать ручной калибровки на Visiview программного обеспечения (v2.1.4), выберите «Настройка | FRAP» меню и нажмите на кнопку «Калибровка | Меню Настройка руководство». Убедитесь, что лазер можно выделить как острый точка. Если нет, либо переориентировать или настроить лазерного оборудования.
- Выполните калибровку направляя лазер заранее определенного программного обеспечения X-Y координаты вручную. Это заставляет программного обеспечения как конкретно лазер для определяемых пользователем региона для текущего масштаба.
- Прежде чем спровоцируют лазер, переключитесь на канал GFP и начать приобретение промежуток время изображения.
- Вручную извлечь региона photobleached на канале GFP, во время просмотра на дисплее.
- Инициировать Фотообесцвечивание ручной триггером 405 нм лазер, по крайней мере 3-4 кадры после начала загрузки изображений. Приобретение кадры до Фотообесцвечивание необходим для нормализации изображения в более поздних анализа данных.
6. Photoactivation процедура
Примечание: Программное обеспечение, Микроскоп установки и настройки, за исключением мощности лазера, аналогичны FRAP. В photoactivation, важное отличие по сравнению с FRAP, это значительно ниже, чем мощность 405nm лазерный, занятых Фотообесцвечивание должны быть использованы, чтобы активировать ПА-GFP без одновременно Фотообесцвечивание его.
- Совместное transfect тип ячейки интерес (B16-F1 клетки здесь; см. шаг 2,5) с плазмида ДНК кодирования ПА-GFP-актина и другой дневно меченых белков (например, mCherry или mCherry-Lifeact).
Примечание: В большинстве случаев, mCherry положительных клеток также будет положительным для PA-GFP-актина вектора, последний обычно не видны на канале GFP до photoactivation. Чтобы повысить вероятность, что mCherry положительных клеток также являются позитивными для PA-GFP, используйте трансфекции соотношении 1:2 mCherry:PA-GFP-миозина. После этого протокола, более 90% клеток, выражая mCherry отображается успешной активации PA-GFP-миозина. - Клетки семян B16-F1 на Ламинин покрытием coverslips (шаг 2.10).
- Соберите тепловизионные камеры (раздел 3).
- Перед запуском photoactivation экспериментов, при необходимости выполните калибровку лазера для выбранной цели (шаг 5.4-5.6).
- Задайте изображение приобретение GFP/488 нм 500 мс экспозиции и 1500 мс интервал времени (в зависимости от экспериментальный дизайн).
-
Отрегулируйте настройки программного обеспечения для получения двойного канала или трехканальный покадровой фильмы, маркировка на площади «Волны серии» и выбрав нужное количество каналов в «приобретать | Длина волны» меню. Рекомендуется, что фильмы промежуток времени приобретаются с Фазовый контраст и GFP каналы.
- При необходимости также включают mCherry канала; Однако подвергая клетки с слишком много света может побудить фотоповреждения. Это можно было бы избежать с кислородом падальщики например Oxyrase17, хотя эффективное лечение требует герметизации камеры клеток.
- Найти transfected клеток на канале mCherry.
- Прежде чем спровоцируют лазер, начать приобретение промежуток время изображения и вручную рисовать региона photoactivated на канале контраст фаза, во время просмотра на дисплее.
- Инициировать photoactivation ручной триггером 405 нм лазер (интенсивность установить между 5 – 15 МВт от «группа | Меню интенсивности»), по крайней мере 3-4 кадры после начала приобретения изображений.
7. анализ данных и представление результатов FRAP
Примечание: Метод представил используется для изучения оборот белка, накопление на участках Ассамблеи динамических актина, в данном случае VASP, который связывает с сайтов адгезии и советы выступающих lamellipodia. Мы анализируем ее оборот на кончик lamellipodium, но те же принципы анализа может применяться для расследования оборот VASP или любых других белков и других внутриклеточных отсеков.
- Откройте покадровой фильмы, производный от Visiview на Метаморф программного обеспечения. В этой статье была использована Метаморф v7.8.10.
- Производные значения интенсивности для photobleached регионов, вручную изложением соответствующих регионах на Метаморф. Рисовать фигуры на кончике lamellipodium, который охватывает весь или часть области photobleached и вручную скорректировать свою позицию на последующих кадрах при необходимости (т.е., если выступающие края), в того, чтобы отслеживать изменения в интенсивности lamellipodial соответствующих компонент во время перемещения подсказка.
- Для коррекции фона и Фотообесцвечивание приобретения анализировать регионов внутри и вне клетки. Смотрите Рисунок 2 для представителей регионов измеряемых интенсивностей.
- Пока выбран ROI, извлечь его значения интенсивности на Метаморф с помощью меню «мера | Область измерения». Убедитесь, что выбраны параметры «Время» и «Средней интенсивности» в меню «Настройка». Нажмите «Открыть журнал» и выберите «Динамический обмен данными». Нажмите «OK», чтобы открыть таблицу Excel и нажмите кнопку «Открыть журнал», чтобы вставить Метаморф значений в Excel.
Примечание: Эти значения используются для создания кривых восстановления флуоресценции. - Для создания кривых восстановления флуоресцирования на кончик lamellipodium photobleached регионов (нормализованное региона интенсивности до Фотообесцвечивание), применить следующее уравнение:
 Уравнение 1
Уравнение 1
где: FRAPTn -photobleached региона света для каждого кадра интерес после Фотообесцвечивание; ИзTn региона интенсивности, принятые вне клетки (фон) для каждого кадра интереса следит Фотообесцвечивание; INSTn -два в среднем внутри региона интенсивности для каждого кадра интерес после Фотообесцвечивание (используется для нормализации для приобретения Фотообесцвечивание со временем); FRAPT-1 является интенсивность региона photobleached до Фотообесцвечивание; T-1 является интенсивность региона, принятых вне клетки (фон) до Фотообесцвечивание; и модулейT-1 два среднем внутри региона интенсивности для каждого кадра интерес до Фотообесцвечивание. - Для каждого кадра раз интерес используйте уравнение 1 для получения кривой восстановления флуоресценции, содержащий все сроки должны расследоваться. Продолжительность времени строго зависит от белков под следствием. Когда неизвестно, выполняют предварительные эксперименты приобрести текучести белка.
- Для расчета половина время восстановления, вставьте значения кривой восстановления флуоресценции с соответствующим временем (в секундах) в сюжет Сигма (v.12) и выполнять кривой с использованием «динамический Fit мастер | Экспоненциальный рост на максимум» инструмент. Выберите моно экспоненциальное (сингл, 3 параметров) или Би экспоненциальное (double, 4 параметры) функции, в зависимости от лучших кривой.
- Используйте следующую формулу для моно экспоненциальной функции:
 Уравнение 2
Уравнение 2
- Используйте следующую формулу для Би экспоненциальной функции:
 Уравнение 3
Уравнение 3
- Вставить параметры «b» и «d», производного от Сигма участок (уравнение 2 или 3 уравнение) в Excel для вычисления половина время восстановления. Применяются следующие уравнения:
 Уравнение 4
Уравнение 4
или Уравнение 5
Уравнение 5
- Когда моно экспоненциальная функция приводит к точной кривой, применяются только 4 уравнением.
- Когда моно экспоненциальная функция не приводит к хорошей кривой, применять Би экспоненциальной формулы, решая уравнение 4 и 5 уравнение. Рассмотрим результате два раза восстановления как представляющие два различных белковых фракций: быстро и медленно обмена фракции, соответственно.
8. Определение скорость полимеризации Lamellipodial актина, FRAP
- Определить скорость полимеризации lamellipodial актина, B16-F1 клетки с меткой EGFP β-актина, transfect и photobleach в lamellipodial регионе (шаг 5.9) с помощью 1,5 интервал времени s и 500 мс GFP экспозиции.
- В Метаморф, откройте покадровой фильмы, приобретенные от Visiview и калибровки пикселей/мкм соотношение согласно цели используется «мера | Средство калибровки расстояния».
- Играть покадровой кино и остановить его на раме, когда lamellipodial флуоресценции восстановления, которая течет назад к ламель как линия, достиг ламели и никаких дальнейших сзади потока могут быть отслежены.
- Измерьте расстояние в мкм между кончик lamellipodium и задней восстановленные флуоресценции. Это расстояние соответствует сумме Ретроградная потока и выступ расстояния.
- Кроме того чтобы отделить выступ с ретроградным потока, Марк lamellipodial наконечник с линии один кадр перед Фотообесцвечивание. Использование линии как ссылку на точку в последующих кадрах сослаться в исходное положение кончика lamellipodium во время Фотообесцвечивание; точкой отсчета может использоваться для измерения расстояния выступ и ретроградным потока.
- Обратите внимание на время (в секундах), необходимых для восстановления флуоресценции после Фотообесцвечивание происходят. Время может быть рассчитано вручную из частоты кадров или визуализированы Метаморф через «мера | Инструмент измерения региона».
- Получить скорость полимеризации актина, используя следующее уравнение (с некоторыми параметрами уравнения, основанный на инструментальных замерах Метаморф от шагов, 8.4 и 8.6):
 Уравнение 6
Уравнение 6
где актина полимеризации составляет в мкм/мин, ретроградная потока расстояние в мкм, lamellipodial расстояние выступа — в мкм, и время указывается в секундах.
9. Анализ белков диффузии и мобильности по Photoactivation
Примечание: Представленные здесь метод описывает анализ актина мономера мобильности, используя photoactivation актина, сливается с PA-GFP, как проиллюстрировано визуализации и количественной оценки белка диффузии через цитозоль.
- Для измерения диффузии актина photoactivatable от цитозольной региона, а также накопление в пределах региона lamellipodial, используйте Метаморф для определения интенсивности с течением времени в следующих регионах (показано на рисунке 3 ): цитозольной photoactivated региона (ПА); lamellipodial регион, в котором photoactivated белки, как ожидается, накапливаются с течением времени (Лам); область за пределами ячейки, используемые для нормализации флуоресценции фон (ЗАМЕНЕН).
- При определении мобильности актина в цитозоле, измеряют собственный цитозольной регионов (см. рис. 3, регионов R1-R5). Обратите внимание, что приобретение Фотообесцвечивание не может быть определено в Аналогичным образом FRAP, за счет увеличения фокуса - и в конечном итоге клеток общесистемной флуоресценции после активации.
- Передача значения интенсивности для всех регионов от Метаморф в электронную таблицу Excel, как описано в шаге 7.4.
- Для изучения скорость перемещения актина photoactivatable от цитозольной региона photoactivation или темпы его инкорпорации в пределах региона lamellipodial (оба представлены как процент интенсивности цитозольной photoactivated региона в момент времени 0) , генерировать флуоресценции кривых из данных в шаге 9.3. Применяются следующие уравнения:
 Уравнение 7
Уравнение 7 Уравнение 8
Уравнение 8
где: ПАTn является интенсивность цитозольной photoactivated региона для каждого кадра интерес после photoactivation; ЛамTn является интенсивность lamellipodial региона для каждого кадра интерес после photoactivation; ИЗTn интенсивность региона, принятых вне клетки (фон) для каждого кадра интереса следит photoactivation; ПАT-1 является интенсивность цитозольной photoactivated региона до photoactivation; ЛамT-1 является интенсивность lamellipodial региона до photoactivation; T-1 является интенсивность региона, принятых вне клетки (фон) до photoactivation; ПАT0 является интенсивность цитозольной photoactivated региона в момент времени 0 (то есть, первый кадр после photoactivation); и внеT0 интенсивность региона, принятых вне клетки (фон) в момент времени 0 (то есть, первый кадр после photoactivation). - При необходимости для лучшей визуализации данных, нормализовать кривые интенсивности 0 путем вычитания интенсивность первый кадр после photoactivation от каждого последующего кадра.
Примечание: Следующий метод анализа (шаги 9,6 – 9,8) также позволяет расчета цитозольной дисперсии photoactivated актина в цитозоль. - Измерения интенсивности для цитозольной регионов, которые расположены последовательно дистально от активации региона.
- Представлять интенсивности этих регионов как процент интенсивности photoactivated региона в момент времени 0, примените уравнение 8, где lamellipodial интенсивности заменяются интенсивности для каждого цитозольной ROI. Размер и количество регионов может варьироваться в зависимости от ячейки размером и разгон расстояние измеряется.
- Для получения количественной значение скорости photoactivated белка, проникнув каждого цитозольной региона, вставьте Сигма сюжет (аналогично FRAP анализа в разделе 7) значения кривой роста интенсивности флуоресценции для каждого региона и время, используйте Уравнение 2 и уравнение 4 получают половину времени интенсивности флуоресценции, достигнув плато. Сравните значения1/2 t между различных экспериментальных групп.
Результаты
Рисунок 1 g h показывают фазы контрастность изображения NIH3T3 фибробластов клетки до и 10 минут после микроинъекции Rac1, который является небольшой ГТФазы Ро семьи, способны индуцировать формирование lamellipodia через его взаимодействия с волной комплекса. Ячейку сначала визуализируется перед микроинъекции (рис. 1g), чтобы подтвердить свою жизнеспособность и морфологии, например, отсутствие lamellipodia. В 10 мин после микроинъекции ячейки явно изменилась его морфологии, который ожидается от этого лечения и указывает на успешное впрыска (рис. 1h).
Для простоты и ясности мы далее обеспечивают образцовую результаты анализа FRAP и photoactivation в клетках, которые не были microinjected в дополнительно.
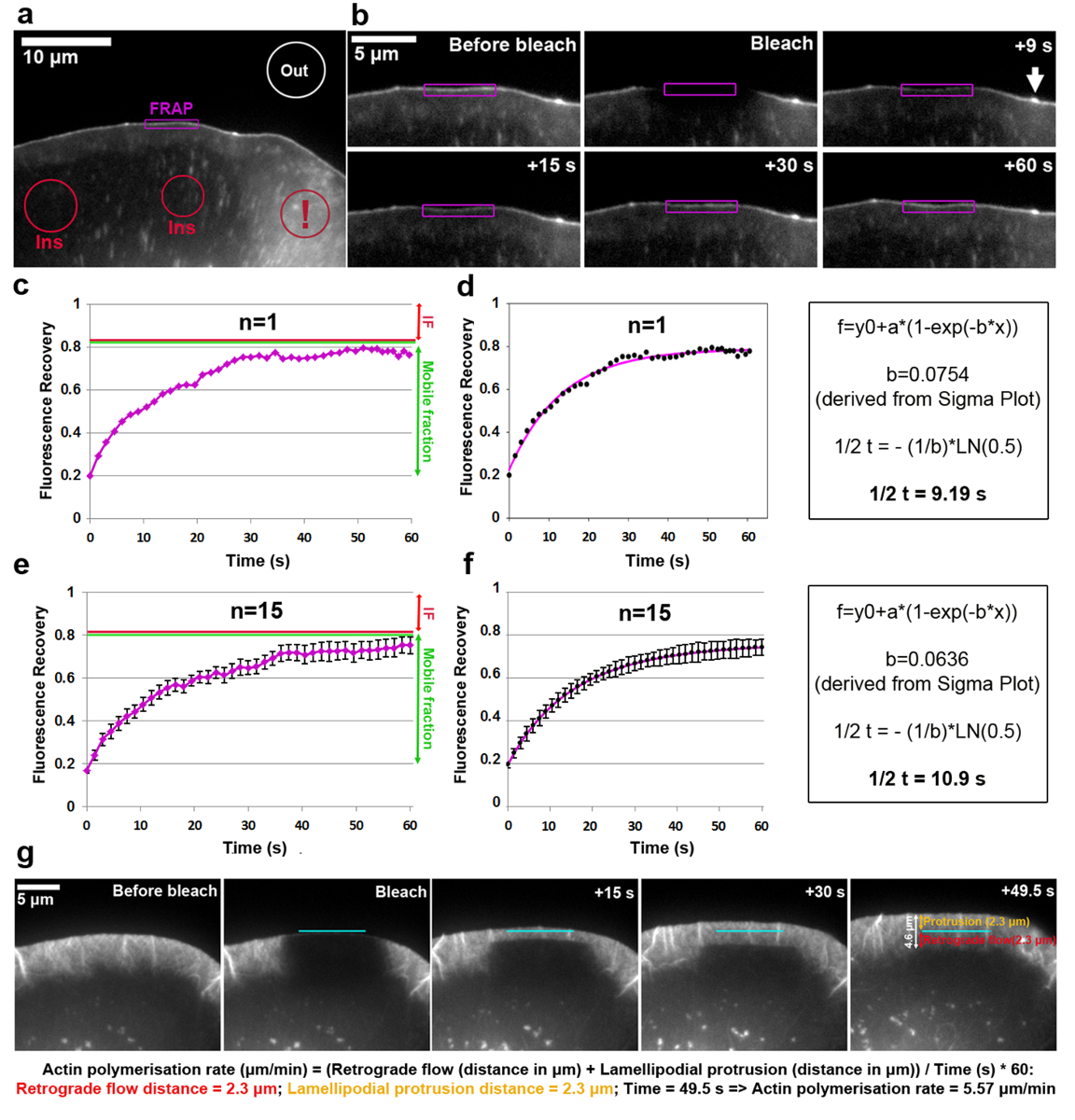
Анализ товарооборота с тегами EGFP VASP на кончик lamellipodium показан на рисунке 2a-f. Обратите внимание, что VASP Кроме предназначен для зарождающейся и фокуса спайки, малые и удлиненные точек в ячейку интерьера18,19. Интенсивность флуоресценции lamellipodial региона с четкой VASP накопления на кончике был беленой и для каждой сроки, измеряется контуру Руа до, во время и после отбеливания как lamellipodium выступает вперед. Как беленый EGFP VASP белки являются время перезапуска, Неотбеленная молекул на этих сайтах, постепенное восстановление флуоресценции наблюдается (рис. 2b). Кривая восстановления FRAP полученные таким образом и нормализуется до отбеливатель интенсивности (выражается в 1) можно увидеть на рис. 2c. Фотообесцвечивание эффективность может меняться и было около 20% от стоимости до отбеливания в этом примере, как определено из значения в t0 (первый кадр после Фотообесцвечивание). Увеличение флуоресценции достигает плато в примере, показанном на примерно 80% флуоресценции до отбеливания. В статической структуры в ходе время эксперимента, например фокуса адгезии разница между предварительной отбеливатель интенсивности и плато флуоресценции достигли после восстановления определяется как неподвижные дроби (если, красная стрелка на рисунке 2 c, e), тогда как количество флуоресцирования восстановленные между время отбеливания и полного восстановления определяется как мобильные дроби (зеленый двунаправленной стрелки в рисунке 2c, e). Обратите внимание, что в динамично меняющейся структуры как кончик lamellipodium, проанализированы здесь, степень IF может не только представляют собой неподвижным молекул, но также являются производными от сокращения выступ скорости, как интенсивность EGFP-VASP, как известно, зависит от этого параметра18. Чтобы вычислить время восстановления, форму кривой был создан Сигма сюжет (рис.d). В этом случае значение параметра «b», извлечены из решения уравнения 2 равно 0.0754, которая применяется к логарифмическая функция (уравнение 4) результаты в примерно половине времени восстановления 9.19 s (Рисунок 2d , правой панели), который является сравнительно быстро в этой конкретной ячейке по сравнению с среднем ранее опубликованы5. Необходимо отметить, что половина время восстановления иногда может значительно отличаться от ячейки к ячейке в пределах той же самой популяции. Таким образом для получения представительной результатов, мы рекомендуем, определяя этот параметр как среднее из клеток, по крайней мере 15-20. Чтобы проиллюстрировать степень дисперсии, арифметических середин EGFP VASP восстановления в среднем от 15 клеток для каждый момент времени были созданные (Рисунок 2e), и средняя кривая вписывается создается и отображается в аналогичных моды (Рисунок 2 f).
Степень полимеризации lamellipodial актина сети включает в себя сумму вперед сети протрузии и ретроградным потока. FRAP может применяться для измерения скорости полимеризации актина transfecting клетками (в данном случае B16-F1) с меткой EGFP β-актина и Фотообесцвечивание выступающие lamellipodial региона (рис. 2g). Для анализа lamellipodial актина сети полимеризации флуоресценции восстановления после отбеливания EGFP-тегами β-актина оценивается с течением времени. Как полимеризации актина мономеров прогрессирует колючая концы филаментов актина lamellipodial (которые все указывают к передней20) сеть является постоянно перемещаются назад и продвигается вперед, ставки которых могут быть легко полученные через восстановление поляризации флуоресценции после Фотообесцвечивание. Флуоресценции восстановление lamellipodium завершена, как только беленой зоны достиг переходной зоне между задней частью lamellipodium и ламелей, которая характеризуется меньшей плотности более горизонтально образное накаливания расслоений переворачивая гораздо медленнее, чем то, что наблюдается в lamellipodium. Как показано на рисунке 2g, флуоресценции восстановления могут быть визуализированы как горизонтальные края и течет обратно к ламелей, которая позволяет измерение расстояний протрузии и ретроградным потока (индивидуально представлены линии в правой панели Рисунок 2g как оранжевый и красный двунаправленной стрелки, соответственно).
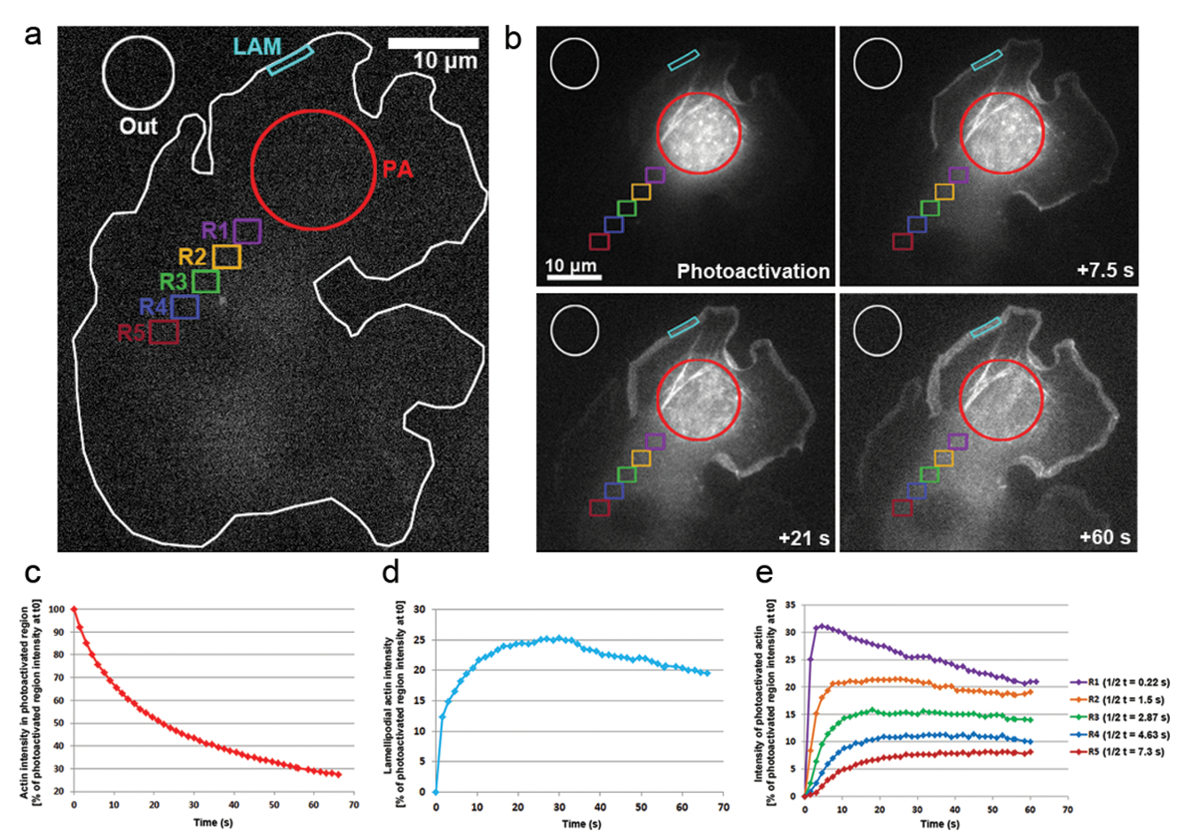
Мы также применили photoactivation в клетках B16-F1 transfected с PA-GFP-актина для отслеживания мобильности актина мономеров в цитозоле и скорость их включения в выступающие lamellipodia. Как показано на рис. 3, б, цитозольной регион был photoactivated под воздействием лазера 405 нм, в то время как изображения были приобретены на GFP канала каждые 1,5 s для визуализации распределения GFP-меткой, актина photoactivated. Photoactivated GFP-актина можно увидеть диффундирующих из цитозольной региона в рисунке 3b. Скорость снижения интенсивности флуоресценции в регионе photoactivated цитозолевая представлена как процент первоначального интенсивности в t0 (первый кадр после photoactivation; Рисунок 3 c). Photoactivated актина также интегрируется в советы lamellipodia, где новые актина мономеров добавляются к растущей колючая концы удлинения нитей актина во время выступа. Чтобы оценить lamellipodial включения, мы измерили интенсивности флуоресценции со временем двухмерный контур или региона около 5 мкм в ширину и 1 мкм в высоту; регион был постоянно повторно позиционируется на кончике lamellipodium как он торчали. Актин инкорпорации была представлена как процент интенсивности флуоресценции photoactivated цитозолевая региона в t0 (рис.d). По мере удлинения нитей актина новых мономеров актина были включены на lamellipodial фронте. Часть этих актина мономеров ковшах взята из цитозольной бассейн, где мономеров были photoactivated. Это приводит к быстрому увеличению флуоресценции в lamellipodia в первые 20 сек после photoactivation. Как новых мономеров добавляются на фронт lamellipodial, ранее включены актина мономеров потока с нитями к ламель Ретроградная потоком. Со временем ROI полностью заполнены с флуоресцентные мономеров и плато в флуоресцировании достигается (рис.d). Постепенное падение в флуоресцировании затем наблюдается, когда, после распространения photoactivated мономеров всей ячейки, не photoactivated актина мономеров чаще повторно добавляются к lamellipodial фронта. Это снижение в флуоресцировании найдет новое плато, которое будет достигнуто как только баланс всей ячейки между photoactivated и не photoactivated мономеров достигается (данные не показаны).
Мобильность актина мономеров в цитозоле была получена путем измерения интенсивности флуоресценции в регионах одинакового размера, позиционируется дистально от региона photoactivated (примером на рисунке 3 регионов цветом помечены R1-R5). Как показано на рисунке 3e, интенсивности флуоресценции в каждом из этих регионов постепенно снижается от cytosolically photoactivated региона, как часть photoactivated актина мономеров становится все более разбавленный с Неактивированные мономеров (то есть, не-лампа дневного света). Кроме того, пик флуоресценции достигается позже: более отдаленные измеренных регион расположен из региона photoactivated, тем больше время, которое требуется для актина мономеров для диффундируют в этих регионах. Представитель значения для степени актина мономера проникновения в каждом регионе могут быть получены путем количественной оценки половины времени достижения плато флуоресценции. Более отдаленные, региона, тем больше времени он принимает для актина photoactivated для диффузного в него, и поэтому требуется больше времени для флуоресцентных плато быть достигнуто, в конечном итоге приводит к более высокому значению t1/2 (eрис. 3).

Рисунок 1 : Imaging процедуры палата Ассамблеи и микроинъекции. () изображений камеры компоненты. (b) Силиконовая смазка тщательно смазывают вокруг открытия пластиковый уплотнитель. (c) coverslip располагается ячейка-стороной вверх в центр тепловизионные камеры открытия. (d) безопасной печати устанавливается позиционирования пластиковый уплотнитель на вершине coverslip и затянув стороне зажимы. (e) микроскопия среднего накапаны в камере слот. (f) тепловизионная камера располагается на микроскопа, электродов и тепловой извещатель связаны с нагревательным элементом предварительно задать до 37 ° C, и клетки могут адаптировать для по крайней мере 30 минут до начала микроскопии. В этом примере микроскопа также оснащен микроманипулятор для выполнения микроинъекций и иглы микроинъекции погружается в средних, охватывающих слоя клеток в тепловизионной камере. (g) NIH3T3 фибробластов клетки визуализируется перед микроинъекции фазово контрастной микроскопии. Красного Креста в perinuclear отсеке указывает местоположение будущего микроинъекции, который соответствует высоким цитоплазматических региона из-за близости к громоздких ядра. (h) 10 мин после микроинъекции с Rac1, клетка реагирует на видных формирования lamellipodia по периметру всей ячейки (обозначены зелеными стрелками). Пожалуйста, нажмите здесь, чтобы посмотреть большую версию этой фигуры.
{kind=link}

Рисунок 2: FRAP позволяет определить ставки белка оборот или lamellipodial полимеризации актина. () представитель пример ячейки B16-F1, выражая EGFP-VASP до Фотообесцвечивание lamellipodial региона, как указано. По-разному окрашенных контуров/формы помечены для указания, какие регионы были рассмотрены для измерения интенсивности флуоресценции со временем. Примечание красный контур отмечены восклицательным знаком, который этикетки цитозольной региона, расположенные в области, содержащей несколько пузырьков и поверхности оборками ячейку. Динамические области, как это следует избегать для выбора регионов флуоресценции ссылки, как они характерны сильные краткосрочные колебания флуоресценции, потенциально вызывая неточные результаты. (b) Lamellipodial региона EGFP VASP выражая ячейки до и после Фотообесцвечивание. Восстановления флуоресцентного сигнала после Фотообесцвечивание в регионе отмечен в фиолетовый визуализируется с течением времени. Стрелка указывает кончик microspike, обогащенные для VASP вероятно из-за высокой плотности нитей актина сверхвысоких там19. кривой восстановления (c) пример FRAP, как производные от количественной оценки интенсивности флуоресценции photobleached lamellipodium (пурпурный контур) в б. красные и зеленые линии справа показывают, соответственно, неподвижных и передвижных фракций. (d) A соответствует кривой восстановления FRAP в c (левая панель) и пример метода расчета, используемых для получения половину времени восстановления (правая панель). (e) пример восстановления FRAP кривой производным от усреднения кривых восстановления флуоресценции 15 клеток, с SEM баров, указывающее степень изменчивости в выборочной совокупности. (f) кривая fit производным от усреднения подходит кривая восстановления FRAP 15 клеток (левая панель) и пример метода расчета, используемых для получения половину времени восстановления (правая панель). (g) промежуток времени панели выступающих lamellipodium ячейку B16-F1, выражая EGFP-тегами β-актина до и после отбеливания lamellipodial региона как указано, следуют флуоресценции восстановления в lamellipodium со временем. На правой панели, измеренная на выступ и ретроградным расстояния предоставляются (в оранжевый и красный, соответственно). Расчеты в изображение панели показывают, как сумма протрузии и ретроградным расстояния используются для получения скорость полимеризации lamellipodial актина сети. Пожалуйста, нажмите здесь, чтобы посмотреть большую версию этой фигуры.
{kind=link}

Рисунок 3: Photoactivation ПА-GFP-актина на мономера, отслеживание всей ячейки. () A репрезентативного примера B16-F1 ячейки, выражая ПА-GFP-актина прежде чем спровоцируют photoactivation цитозолевая региона как обозначается красным кружком (PA). Чтобы указать, какие регионы были рассмотрены для измерения интенсивности флуоресценции со временем помечены по-разному цветные контуры. (b) Иллюстрация временного распределения ПА-GFP-актина ниже photoactivation. Обратите внимание постепенное сокращение флуоресценции в photoactivated, цитозольной региона (красный кружок), как photoactivated актина диффундирует вдали от него. Из-за их распространение в передней и сборки в сети photoactivated актина мономеров постепенно усиливаются в lamellipodia (голубой региона) и в цитозоле (различных цветных областей) в зависимости от расстояния и времени моды. (c) представителя, временное снижение флуоресценции в регионе photoactivated цитозолевая (красный контур в b). (d) височные изменения в интенсивности флуоресценции в регионе lamellipodial (голубой контур в b). (e) кривых представитель временных изменений в интенсивности флуоресценции цитозольной регионов (цветом в b) благодаря позиционирования в переменной расстояниях от района photoactivation. Обратите внимание, как плато раза достижения флюоресценция (указывается в легенду справа) увеличить расстояние от данного региона в области photoactivation, вероятно, соотнося с увеличенной раз, необходимые для распространения актина мономеров в соответствующего региона. Пожалуйста, нажмите здесь, чтобы посмотреть большую версию этой фигуры.
{kind=link}
Обсуждение
Здесь мы обсуждаем важнейшие шаги в описанных в этой статье, и как они могут быть оптимизированы для использования в различных экспериментальных условиях.
Микроинъекции является метод, который может быть применен для мониторинга в клетках мгновенные эффекты от введения экзогенных белки, ингибиторы или наркотики. Это может быть особенно выгодным для определения функции белков в трудных для transfect типов клеток или в ситуациях, когда долгосрочные выражение не желателен. Необходимо отметить, что выживание некоторых типов клеток варьируется в зависимости от внеклеточного матрикса, который они посеяны на. Наиболее эндотелиальной эпителия, типов или фибробластоподобных клеток, даже небольшие, как рыбы кератиноцитах (см. Данг et al. 21 и Андерсон и крест22) могут быть успешно введены. Однако есть исключения, такие как B16-F1 клетки посеян на Ламинин, которые составляют систему отличную модель миграции клеток, но несовместимы с инъекций на этот тип субстрата для неизвестной причины. Для NIH3T3 клетки фибробластов мы регулярно проводить инъекции на фибронектин субстрата, а также добавлены дополнительные методы, такие как FRAP (даже с photoactivation; отображаются для B16-F1 клетки здесь) может выполняться одинаково хорошо в эти фибробласты (см. например, Köstler и др. 3). необходимо также учитывать, что различные белки, согласно их функциональных свойств и целей эксперимента, может принимать различное количество времени, чтобы вызвать изменения, изменяясь от секунд до часа. Преимуществом метода является, что дозировка/концентрация экзогенных агента может контролироваться более точно на уровне отдельной ячейки, чем например, при использовании плазмида transfection. Кроме того флуоресцентных меток белка не является необходимостью гарантировать свое присутствие в ячейке, которая может повысить гибкость, если требуется одновременный многоканальный визуализация других белков, дневно тегами. Микроинъекции может быть особенно полезен для анализа мгновенные эффекты специфических белков или белковых смесей на динамических изменений морфологии клеток или цитоскелета (например, Данг и др. « 21 пример мгновенного воздействия на миграции, Arp2/3 комплекса ингибитора Арпинский). Недостатком метода является его инвазивность, которая может вызвать повреждение клеток или влиять на морфологию клеток. Таким образом важным соображением при выполнении микроинъекций является мониторинг жизнеспособность клеток. Метод, представленный здесь зависит от ручного манипулирования. В условиях, проверенных на совместимость с успешным инъекции, например фибробластов, растущих на фибронектин субстрат протокол ручной инъекций, описанные здесь позволяет почти 100% успеха; Это важно, когда сочетание этот подход с сложных и длительных последующих экспериментов, включая видео микроскопии или FRAP, как ранее опубликованные3. Это не исключает, что иногда, отдельные клетки могут страдать от микроинъекции событие, которое может быть безопасно признаны резкие изменения контраста ядро и цитоплазму, следуют Ретракция края ячейки. Таких случаях экспериментальных исключены и таким образом не считаются для дальнейшего анализа.
Однако, half автоматическом подход также широко используется, например используя быстрое (< 300 мс) под контролем машина иглы снижение совпадающих с увеличением давления впрыска, так, чтобы игла имеет только должен располагаться над каждой ячейкой до соответствующих инъекции. Показатель успеха half автоматическом инъекции является по определению ниже, чем ручной подход, описанный выше, просто потому что он оптимизирован для скорости, а затем анализ нескольких клеток, которые успешно пережили это лечение; Таким образом он не зависит от успешного введения отдельной ячейки. Таким образом в отличие от одноклеточного анализ, half автоматическом инъекции лучше подходят для анализа эффекты инъекций нескольких сотен клеток, например, в видео микроскопия при низком увеличении или ячейки фиксации и окраски. Независимо от используемых детальный подход микроинъекции не является конечной точкой assay, но могут быть объединены с различные методы, включая FRAP или photoactivation3.
При определении белка текучести, FRAP, интенсивность лазерного должны быть оптимизированы, в зависимости от условий установки и изображения микроскопа (увеличение, цели, и т.д., а также тип клеток, структура и флуоресцентный белок для Фотообесцвечивание). Обратите внимание, что при оптимальной лазерной мощности, эффективного отбеливания в сочетании с наименьшими фотоповреждения, чтобы избежать усадки или полное втягивание структуры в рамках анализа (например, lamellipodia или filopodia) или даже повреждения на клеточном уровне. В идеале по крайней мере 70-80% отбеливания эффективности должно быть достигнуто, хотя полный отбеливание может сдерживаться чрезвычайно быстрый оборот белка, в котором случае, все, что выше 50% также может быть приемлемым. Оптимальная мощность отбеливания для данной структуры и флуоресцентные краски должны быть проверены экспериментально, начиная от мощности низкой лазера следовали его постепенное увеличение. Конечно, любой Люминесцентную краску можно по определению быть отбеленные с лазерного света вблизи пика возбуждения (488 нм для часто используемых зеленых красителей, например FITC или EGFP). Однако лазеры с более скоро длин волны, например вблизи УФ лазеров, доставить высшие силы и таким образом может также использоваться для эффективного отбеливания часто используемых красителей. Мы регулярно используют 405 нм Лазер диода (120 МВт) для отбеливания как EGFP, так и красной краски люминесцентные, флуоресцентные (например, mCherry), хотя и с немного ниже эффективности в случае последнего (данные не показаны). Как 405 нм диод может также использоваться для photoactivation PA-GFP (см. ниже), он придает этой системы с максимальной гибкостью.
B16-F1 клеточных структур и флуоресцентные белки photobleached здесь были применены 405 нм лазерной полномочий между 65 – 100 МВт. При анализе photobleached региона, важно рассмотреть ли данная структура сохраняется в своей первоначальной форме над анализа периода времени. Например при анализе оборот белков в lamellipodia советы, следует позаботиться ли кривизны lamellipodia значительно изменяется со временем, как изменения в кривизны может привести к неточные результаты, если региона/контур проанализированы не полностью охватывают полностью структуры в каждом кадре измерений. Кроме того следует отметить, что пучки, встроенные в lamellipodia, например microspikes, может вызвать отклонения в интенсивности флуоресценции. Как показано на рисунке 2b (белая стрелка в 9 s сроки), microspike как структура расположен рядом с измеренной photobleached региона, но остается за ее пределами на протяжении периода измерения и таким образом не вызывает неточность. Для анализа белка оборота важные соображения при выборе местоположения и размера анализе регионов являются, что их флуоресценции со временем следует не поддаваться влиянию значительно изменения морфологии клеток или факторы другой чем трудно избежать приобретение Фотообесцвечивание. Например структуры, обеспечивая значительный количественный вклад анализируемой структуры не должны двигаться из измеренных региона в ходе анализа; Кроме того отношения, флуоресцентный подразделений например везикулярного структур, которые привлекают белка не следует заносить области интересов во время анализа. Для определения скорости полимеризации lamellipodial актина, следует позаботиться что анализируются не втягивания или плессировки (т.е., вверх складывания) lamellipodia, как это будет сильно влиять на точность результатов. Кроме того втягивание lamellipodial регионов может появиться как быстрое сзади транслокации, потенциально ведет к завышению ставок lamellipodial актина полимеризации. Дополнительного рассмотрения — это расстояние внутриклеточных нормализации регионов (берется как ссылку позиции для коррекции Фотообесцвечивание приобретения) от фактического положения Фотообесцвечивание, который должен быть достаточно большим, чтобы избежать прямой влияние на photobleached площадь.
При создании оптимальных условий для photoactivation конструкций, ПА-GFP-меткой, следует позаботиться чтобы избежать мгновенного отбеливания во время photoactivation. В нашей работе, лучшие результаты были получены с лазерной полномочий 5 - 10 раз ниже, чем обычно занятых для отбеливания EGFP. Для захвата изображений молекул photoactivated выдержка и временной интервал между кадрами должны оптимизированы с учетом размера регионов и структур photoactivated и проанализированы, а также потенциальной мобильности photoactivated белки для других внутриклеточных местах. Что касается всех видов флуоресценции изображений поддержание жизнеспособности клеток имеет решающее значение для получения физиологически релевантные результаты.
В принципе, зеленый и красный photoconversion флуоресцентных белков, таких как mEos или Dronpa варианты12 представляет собой столь же мощный метод следующие динамики и оборот внутриклеточных структур, таких как lamellipodium (см., например, Бёрнетт и др. 23). Преимущество последнего метода в отличие от ПА-GFP будет возможность следовать динамики белков до и после преобразования с двух различных цветов, без необходимости совместного Экспресс дополнительные красный флуоресцирующий белок. Однако в наших предварительных экспериментов, степень изменения контраста и интенсивностью флуоресцентного сигнала, достигнутые после photoactivation PA-GFP был больше по сравнению с photoconverted зонды, возможно за счет превосходной спектральных особенностей зеленый и красный флуоресцентных зондов (данные не показаны). В любом случае подробные исследования по актина накаливания оборот в ячейке края выступов например lamellipodia или Vaccinia вируса индуцированной актина хвосты пока только были опубликованы с помощью ПА-GFP производные5,6,24.
При рассмотрении какой регион ячейки для анализа после photoactivation, несколько факторов должен учитываться, которые обсуждаются, используя конкретный пример, показанный здесь (включение актина мономеров на краю после активации в цитозоле клеток), но Конечно могут быть экстраполированы различных аналогичных научных проблем. Во-первых, при измерении скорости включения lamellipodial cytosolically photoactivated белков, например, в отдельных экспериментальных условиях (как показано в Димчев et al. 6), размеры цитозольной регионов и их расстояние до краев lamellipodial должны быть сопоставимыми между экспериментальной группы. Важно также учитывать, что при photoactivating цитозолевая регионов, толщина клеток больше в позициях ближе к ядру. Активация толще сотовой регионах может привести к большее количество активированных белков, учитывая, что распределение белка, чтобы быть активированы содержанием однородно распределяется в цитозоль. И наконец выражение уровни белка, чтобы быть активированы безусловно, может быть сильно варьирует в отдельных клетках. Из всех этих соображений изменчивости важно сравнить уровень включения cytosolically активированный белков в другом месте в ячейке по отношению к общей флуоресценции, полученные после активации в конкретных регионах.
Мы описали как микроинъекции может использоваться в качестве инструмента для изучения воздействия белков на морфологию клеток и примером этого, демонстрируя мощным индукции lamellipodial структур в клетки фибробластов microinjected с NIH3T3 малых ГТФазы Rac1. Ранее мы применили эту технику, чтобы вмешиваться в функции Arp2/3 в клетках, microinjected с доменом C-терминал WCA шрам/волны3. Различные параметры в microinjected клетки могут быть проанализированы другие анализы, например FRAP или photoactivation. Мы описали, как FRAP и photoactivation могут быть использованы для изучения субцеллюлярные динамика и мобильность актина мономеров. Наша группа была использована FRAP ранее5 расследовать оборота белков локализации lamellipodia, таких как VASP, Abi, cortactin, Кофилин и укупорки белка, или для выяснения оборот компонентов в фокуса спайки в присутствии и отсутствие сигнализации4Rac. Кроме того измерения актина полимеризации ставки может быть достигнуто путем Фотообесцвечивание с тегами EGFP β-актина5, но существуют альтернативные методы. Отслеживание флуоресцентные неоднородностей, как живой клетки изображений совместимых датчиков маркировки сотовых актина нитей, например Lifeact25, также может быть занятых6,26. Преимуществом здесь является, что можно избежать гиперэкспрессия β-актина, который способен увеличить выступ края ячейки и миграции и таким образом потенциально препятствует конкретного анализа или экспериментальных вопрос (см., например, Кейдж и др. 26; Пекхам и др. 27). Однако невыгодном зонд Lifeact представляет его быстрого включения/выключения кинетики привязки к филаментов актина, так что отбеливание актина накаливания структур помечены Lifeact в клетках предоставляет информацию только на оборот зонда, но не оборот филаментов актина, которых он связывает25. Отслеживания флуоресценции неоднородностей ранее занятых6,26 обеспечивают практического компромисса, похож на широко используется отслеживание флуоресценции крапинками включены в нитевидные цитоскелета структуры ( например, лосося и Уотерман28см), но могут быть не так прямо вперед, чтобы использовать и как точно, как FRAP EGFP-тегами F-актина структур. Photoactivation был применен нами для оценки показателей учета мономерных актина выступающие lamellipodia, а также его мобильности в цитозоле, в контексте экспериментально настроило цитозольной F-актина уровней6. Методика полезна при изучении мобильность и распределение белков производным от относительно крупных областей, например цитозольной регионов. Однако изучение распределения белков производным от относительно небольших photoactivated структур; например, конусы роста может быть сложным из-за низкого числа флуоресцентных молекул активирован, слабых сигналов и таким образом отсутствие чувствительности. Потенциальных альтернативных методов photoactivation или photoconversion люминесценции (см. выше) может включать обратную FRAP, которая опирается на Фотообесцвечивание всю ячейку за исключением ROI, следуют отслеживания мобильности флуоресцентных молекул вдали от Этот регион. Метод не требует экспрессирующих photoactivatable версии белков, но всегда будет привлекать воздействием необычно высокая доза мощности лазера, потенциально вызывая нежелательных побочных эффектов, таких как фотоповреждения.
Очевидно photoactivation и FRAP не может отличить ли белки движутся как мономеры, димеры или даже небольшие олигомеров и ли они перемещаются в сочетании с партнерами дополнительные привязки. Информация такого рода вместо этого могут быть получены флуоресценции корреляции спектроскопии методы29 или, альтернативно, FLIM-ладу30. Тем не менее FRAP и photoactivation представляют собой простой подходы к непосредственно оценить динамику местных и глобальных белка в клетках, независимо от того протеин интереса, субцеллюлярные расположения или типа клеток изучены.
Раскрытие информации
Авторы не имеют ничего сообщать.
Благодарности
Мы признательны для немецкого фонда научных исследований (DFG) для финансовой поддержки (Грант Nr. RO2414/5-1-KR).
Материалы
| Name | Company | Catalog Number | Comments |
| B16-F1 mouse skin melanoma cells | American Type Culture Collection, Manassas, VA | CRL-6323 | |
| NIH-3T3 cells | American Type Culture Collection, Manassas, VA | CRL-1658 | |
| DMEM 4.5g/L glucose | Life Technologies, Thermno Fisher Scientific, Germany | 41965-039 | |
| Ham’s F-12 medium | Sigma-Aldrich | N8641 | |
| Fetal calf serum (FCS) | PAA Laboratories, Linz, Austria | A15-102 | |
| Fetal bovine serum (FBS) | Sigma-Aldrich, Germany | F7524 | Lot054M3396 |
| MEM Non essential amino acids | Gibco, ThermoFisher Scientific, Germany | 11140035 | |
| L-Glumatine 200mM (100x) | Life Technolgies | 25030-024 | |
| Pen-Strep 5000 U/mL | Life technologies | 15070063 | |
| Sodium Pyruvate (100 mM) | Gibco, ThermoFisher Scientific, Germany | 11360-039 | |
| Laminin | Sigma-Aldrich | L-2020 | |
| Laminin coating buffer | Self-made: 50mM Tris ph7.4, 150mM NaCl | ||
| Fibronectin from human plasma | Roche Diagnostics, Mannheim, Germany | 11 051 407 001 | |
| Jetpei | Polyplus Transfection, Illkirch, France | 101-10N | |
| JetPei buffer | Polyplus Transfection, Illkirch, France | 702-50 | 150mM NaCl |
| PA-GFP-actin plasmid DNA | described in Koestler et al.2008 | ||
| pEGFP-actin plasmid DNA | Clontech, Mountain View, CA, USA | ||
| Rac1 protein for microinjection | Purified as GST-tagged version, and cleaved from GST prior to injection | ||
| Microinjection buffer | Self-made: 100mM NaCl, 50mM Tris-HCl ph7.5, 5mM MgCl2, 1mM DTT | ||
| Dextran, Texas Red, 70,000 MW, Lysine Fixable | Molecular Probes, Thermno Fisher Scientific, Germany | D1864 | |
| Microscope circular cover glasses 15mm, No.1 | Karl Hecht, Aisstent, Sondheim, Germany | 1001/15 | |
| Eppendorf Femtotips Microloader Tips | Eppendorf, Hamburg, Germany | 5242 956 003 | |
| Eppendorf Femtotip Microinjection Capillary Tips | Eppendorf, Hamburg, Germany | 930000035 | |
| Silicone Grease | ACC Silicones, Bridgewater, England | SGM494 | |
| Aluminium Open Diamond Bath Imaging Chamber | Warner instruments | RC-26 | |
| Automatic temperature controller | Warner Instruments | TC-324B | |
| Microscope: Axio Observer | Carl Zeiss, Jena, Germany | ||
| CoolSnap-HQ2 camera | Photometrics, Tucson, AZ | ||
| Lambda DG4 light source | Sutter Instrucment, Novato, CA | ||
| Laser source | Visitron Systems | ||
| Eppendorf FemtoJet microinjector | Eppendorf, Hamburg, Germany | With built-in compressor for pressure supply | |
| Nikon Narishige Micromanipulator system | Nikon Instruments, Japan | ||
| Visiview software v2.1.4 | Visitron Systems, Puchheim, Germany | ||
| Metamorph software v7.8.10 | Molecular Devices, Sunnyvale, CA | ||
| Sigma Plot v.12 | Systat Software Inc. |
Ссылки
- Day, R. N., Davidson, M. W. The fluorescent protein palette: tools for cellular imaging. Chem Soc Rev. 38 (10), 2887-2921 (2009).
- Ishikawa-Ankerhold, H. C., Ankerhold, R., Drummen, G. P. Advanced fluorescence microscopy techniques--FRAP, FLIP, FLAP, FRET and FLIM. Molecules. 17 (4), 4047-4132 (2012).
- Koestler, S. A., et al. Arp2/3 complex is essential for actin network treadmilling as well as for targeting of capping protein and cofilin. Mol Biol Cell. 24 (18), 2861-2875 (2013).
- Steffen, A., et al. Rac function is crucial for cell migration but is not required for spreading and focal adhesion formation. J Cell Sci. 126, Pt 20 4572-4588 (2013).
- Lai, F. P., et al. Arp2/3 complex interactions and actin network turnover in lamellipodia. EMBO J. 27 (7), 982-992 (2008).
- Dimchev, G., et al. Efficiency of lamellipodia protrusion is determined by the extent of cytosolic actin assembly. Mol Biol Cell. 28 (10), 1311-1325 (2017).
- Koppel, D. E., Axelrod, D., Schlessinger, J., Elson, E. L., Webb, W. W. Dynamics of fluorescence marker concentration as a probe of mobility. Biophys J. 16 (11), 1315-1329 (1976).
- Patterson, G. H., Lippincott-Schwartz, J. A photoactivatable GFP for selective photolabeling of proteins and cells. Science. 297 (5588), 1873-1877 (2002).
- McKinney, S. A., Murphy, C. S., Hazelwood, K. L., Davidson, M. W., Looger, L. L. A bright and photostable photoconvertible fluorescent protein. Nat Methods. 6 (2), 131-133 (2009).
- Gurskaya, N. G., et al. Engineering of a monomeric green-to-red photoactivatable fluorescent protein induced by blue light. Nat Biotechnol. 24 (4), 461-465 (2006).
- Lippincott-Schwartz, J., Patterson, G. H. Photoactivatable fluorescent proteins for diffraction-limited and super-resolution imaging. Trends Cell Biol. 19 (11), 555-565 (2009).
- Kremers, G. J., Piston, D. Photoconversion of purified fluorescent proteins and dual-probe optical highlighting in live cells. J Vis Exp. (40), (2010).
- Fischer, A. H., Jacobson, K. A., Rose, J., Zeller, R. Preparation of slides and coverslips for microscopy. CSH Protoc. 2008, 4988(2008).
- Small, J. V., Rottner, K. Actin-based Motility. Carlier, M. F. , Springer. Dordrecht. (2010).
- Kaverina, I., et al. Enforced polarisation and locomotion of fibroblasts lacking microtubules. Curr Biol. 10 (12), 739-742 (2000).
- Small, J., Rottner, K., Hahne, P., Anderson, K. I. Visualising the actin cytoskeleton. Microsc Res Tech. 47 (1), 3-17 (1999).
- Mikhailov, A. V., Gundersen, G. G. Centripetal transport of microtubules in motile cells. Cell Motil Cytoskeleton. 32 (3), 173-186 (1995).
- Rottner, K., Behrendt, B., Small, J. V., Wehland, J. VASP dynamics during lamellipodia protrusion. Nat Cell Biol. 1 (5), 321-322 (1999).
- Svitkina, T. M., et al. Mechanism of filopodia initiation by reorganization of a dendritic network. J Cell Biol. 160 (3), 409-421 (2003).
- Small, J. V., Isenberg, G., Celis, J. E. Polarity of actin at the leading edge of cultured cells. Nature. 272 (5654), 638-639 (1978).
- Dang, I., et al. Inhibitory signalling to the Arp2/3 complex steers cell migration. Nature. 503 (7475), 281-284 (2013).
- Anderson, K. I., Cross, R. Contact dynamics during keratocyte motility. Curr Biol. 10 (5), 253-260 (2000).
- Burnette, D. T., et al. A role for actin arcs in the leading-edge advance of migrating cells. Nat Cell Biol. 13 (4), 371-381 (2011).
- Humphries, A. C., et al. Clathrin potentiates vaccinia-induced actin polymerization to facilitate viral spread. Cell Host Microbe. 12 (3), 346-359 (2012).
- Riedl, J., et al. Lifeact: a versatile marker to visualize F-actin. Nat Methods. 5 (7), 605-607 (2008).
- Kage, F., et al. FMNL formins boost lamellipodial force generation. Nat Commun. 8, 14832(2017).
- Peckham, M., Miller, G., Wells, C., Zicha, D., Dunn, G. A. Specific changes to the mechanism of cell locomotion induced by overexpression of beta-actin. J Cell Sci. 114, Pt 7 1367-1377 (2001).
- Salmon, E. D., Waterman, C. M. How we discovered fluorescent speckle microscopy. Mol Biol Cell. 22 (21), 3940-3942 (2011).
- Machan, R., Wohland, T. Recent applications of fluorescence correlation spectroscopy in live systems. FEBS Lett. 588 (19), 3571-3584 (2014).
- Becker, W. Fluorescence lifetime imaging--techniques and applications. J Microsc. 247 (2), 119-136 (2012).
Перепечатки и разрешения
Запросить разрешение на использование текста или рисунков этого JoVE статьи
Запросить разрешениеСмотреть дополнительные статьи
This article has been published
Video Coming Soon
Авторские права © 2025 MyJoVE Corporation. Все права защищены