
Method Article
Оценка дифференциации потенциала мыши простаты эпителиальных клеток с использованием органоидной культуры
* Эти авторы внесли равный вклад
В этой статье
Резюме
Органоиды простаты мыши представляют многообещающий контекст для оценки механизмов, регулирующих дифференциацию. Эта статья описывает улучшенный подход к установке органов простаты, и вводит методы (1) собирать лизат белка из органоидов, и (2) исправить и пятно органоидов для цельномонтажковой микроскопии.
Аннотация
Эпителий простаты состоит преимущественно из базальных и светящихся клеток. Отслеживание линии vivo было использовано для определения дифференциации способности базальных и светящихся клеток мыши простаты во время развития, регенерации тканей и трансформации. Тем не менее, оценка клеточных и внутренних регуляторов эпителиальной дифференциации простаты с использованием подхода отслеживания линий часто требует обширного разведения и может быть экономически запретительным. В органоидном ассиде простаты базальные и светящиеся клетки генерируют простатический эпителий ex vivo. Важно отметить, что первичные эпителиальные клетки могут быть выделены из мышей любого генетического фона или мышей, обработанных любым количеством мелких молекул до или после, покрытие в трехмерной (3D) культуры. Достаточно материала для оценки дифференциации создается через 7-10 дней. Сбор базально-полученных и светящихся органоидов для (1) анализа белка с помощью западного побелки и (2) иммуногистохимического анализа нетронутых органоидов с помощью целой конфокальной микроскопии позволяет исследователям оценить дифференциацию ex vivo элициальных клеток простаты. При использовании в сочетании, эти два подхода обеспечивают дополнительную информацию о дифференциации способности базальных и светящихся клеток простаты в ответ на генетические или фармакологические манипуляции.
Введение
Базальные и светящиеся клетки составляют большинство эпителия простаты1. Исследования отслеживания линий показали, что эти типы клеток преимущественно самоуверенны отдельными прародителями во взрослой мыши2; однако, дифференциация светила от базальных прародителей наблюдалась в нескольких контекстах, включая развитие3,4,регенерацию тканей5,воспаление6,7 и начало рака простаты2,8. Кроме того, новые данные подтверждают существование многопотентных световых прародителей, а также светящихся прародителей9. При метастатическом раке предстательной железы дифференциация от АР-зависимой линии светящегося света до AR-безразличной линии с базальными и нейроэндокринными особенностями представляет собой все более ценимый механизм устойчивости к ингибиторам андрогенных путей10,11,12. Поэтому, поскольку дифференциация вовлечена в нормальную физиологию, инициация рака и устойчивость к терапии, выяснение ключевых молекулярных регуляторов дифференциации эпителиальных клеток простаты имеет решающее значение.
Мышь простаты органоидной модель юала стала элегантным контексте ex vivo для изучения дифференциации эпителиальной клетки простаты9,13,14. В этом расследовании, отдельные эпителиальные клетки покрываются в 3D-матрицы, где они генерируют железистые структуры, содержащие как базальные и светящиеся клетки в течение 1 недели. В то время как существующие подходы к покрытию клеток в органоидную культуру могут быть использованы для эффективного генерации органоидов, эти подходы требуют дальнейшей оптимизации14. Известные проблемы, связанные с культивированием органов простаты включают (1) за исключением двухмерных (2D) колоний, которые образуются под Matrigel (матричный гель) от анализа, (2) поддержание целостности матричного геля во время медиа-изменений, и (3) подсчета органоидов точно. В этой статье излагается подход к генерации органоидов из эпителиальных клеток, изолированных от простаты мыши. Описанный подход предполагает покрытие пластин с поли (2-гидроксиэтил метакрилат) (Поли-HEMA) для предотвращения возникновения 2D колоний. Кроме того, клетки покрываются в матричной гель кольцо, а не матричного геля диск, что делает изменение средств массовой информации и подсчета органоидов менее сложным. Эти методы позволяют исследователям более легко исследовать, как генетические изменения или небольшие молекулы, введенные до или во время органоидного образования, изменяют ключевые процессы, такие как дифференциация.
Сбор простаты органоидов для западной подья или иммуногистохимический анализ с помощью цельномонтажной конфокальной микроскопии может обеспечить ценное механистическое понимание дифференциации13,но устоявшихся протоколов для подготовки органоидов для таких методов не хватает. Данная рукопись описывает подходы к сбору органоидов для (1) сбора белкового лизата, или (2) фиксации и окрашивания для конфокальной микроскопии. Важно отметить, что подход, описанный для фиксации и окрашивания органов простаты значительно улучшилось по сравнению с существующими методами. Хотя они полагаются на секционирование органоидов15, метод, описанный в этой рукописи использует нетронутые органоиды, который помогает защитить от повреждения органоидов во время подготовки образца. При использовании в комбинации, Западная поместье и конфокальная микроскопия может обеспечить ценную информацию о молекулярных регуляторов дифференциации. Кроме того, эти подходы могут быть использованы для моделирования других процессов, таких как разработка и трансформация.
протокол
Все методы, описанные здесь, были одобрены Институциональным наблюдательным советом Калифорнийского университета в Лос-Анджелесе.
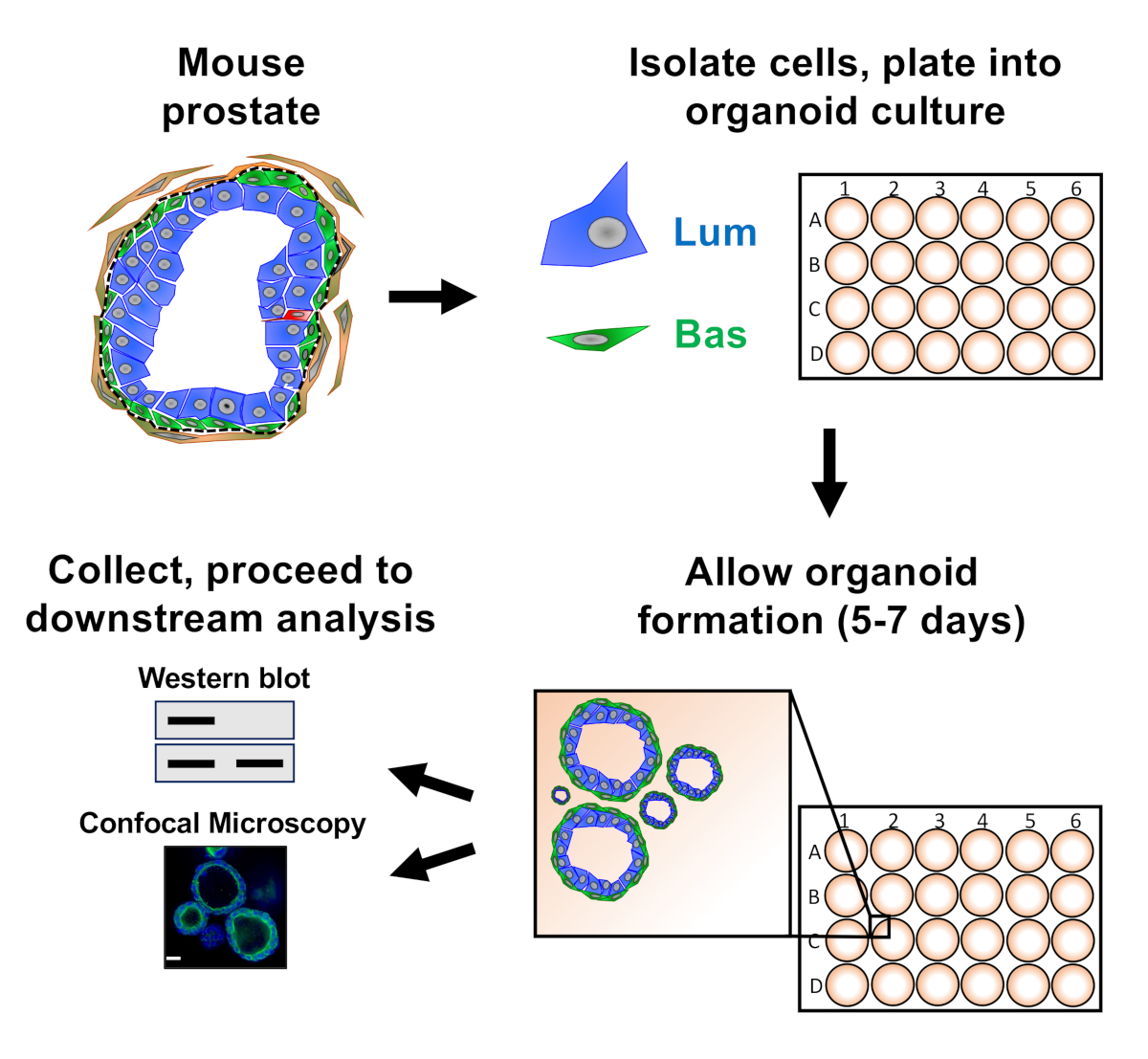
ПРИМЕЧАНИЕ: Схема, иллюстрирующая подходы, описанные в документе, приведена на рисунке 1.
1. Изолирующая мышь базальной и люминециальной простаты эпителиальных клеток с использованием флуоресценции активированной сортировки клеток (FACS) - ВРЕМЯ: 30 Мин
ПРИМЕЧАНИЕ: Выполните шаги 1.3-1.5 в темноте.
- После того, как клетки отослали клетки от общей простаты мыши, как описано в Lawson et al.16,перенесите клетки в трубки FACS и повторно отрептуем 0,1-5 х 106-6-х клеток в 100 ЗЛ диссоциационных носителей (Таблица 1).
- Добавьте соответствующий объем следующих непосредственно спряженых первичных антител: CD45, CD31, Ter-119, EpCAM и CD49f.
- Инкубировать на льду, защищенном от света, в течение 20 мин.
ПРИМЕЧАНИЕ: Рекомендуется использовать 10% от общего числа разъединенных ячеек для неокрашенных и одноцветных элементов управления. Эти элементы управления необходимы для установки правильной компенсации и напряжения для сортировки. - Утолить антитела коктейль, добавив 1 мл диссоциации средств массовой информации для каждого образца. Пелле клетки центрифугации при температуре 800 х г в течение 5 мин при комнатной температуре (RT) и удалить супернатант, аспирируя.
- Повторное приостановку клеток в соответствующем объеме (250 л на 1 х 106 ячеек) диссоциационных носителей, содержащих 1 мкг/мл 4',6-диамидино-2-фенилиндол (DAPI). Переход к FACS. Поток цитометрии участков, демонстрирующих изоляцию мыши базальных и светящихся эпителиальных клеток простаты иллюстрируются на рисунке 2.
2. Покрытие Сортированные эпителиальные клетки простаты в первичную мышь органоидной культуры - ВРЕМЯ: 2-3 H (Исключая Поли-HEMA покрытием плиты Подготовка)
ПРИМЕЧАНИЕ: Плиты покрыты Поли-HEMA для предотвращения 2D образования колонии на поверхности хорошо под матричной гель. Подготовка Поли-HEMA покрытием пластин 1 день до покрытия отсортированных базальных или светящихся эпителиальных клеток простаты в мыши органоидной культуры. Оттепель 1 мл aliquots пониженного фактора роста матричного геля, далее называют матричного геля, на льду 2 ч до шага 2.1. Y-27632 (ингибитор ROCK) следует добавлять в органоидные средства массовой информации мыши непосредственно перед шагом 2.1. Выполните шаги 2.1-2.8 на льду.
- Пеллет клетки в 5 мл круглых нижних труб центругирование при 800 х г в течение 5 мин при 4 кв и аспирировать супернатант.
- Вымойте клеточные гранулы в 500 л мыши органоидных носителей (Таблица 2)14.
- Пелле клетки центрифугации при 800 х г в течение 5 мин при 4 кв и аспирировать супернатант.
- Повторное в мышиных органоидных носителях при плотности клеток в 1000 клеток/ЗЛ.
- Чтобы подготовить мастер смеси, смешать эпителиальные клетки приостановлено в мыши органоидных носителей с матричным гелем для создания окончательной смеси, которая содержит 25% клеток / носителей и 75% матричного геля. Базальные клетки, как правило, покрываются концентрацией 100-2000 клеток/80 л, в то время как светящиеся клетки, как правило, покрываются концентрацией 2000-10 000 клеток/80 л. Плотность клеток, покрытых, варьируется в зависимости от дня ожидаемого сбора материала и желаемого применения вниз по течению.
ПРИМЕЧАНИЕ: Охладите соответствующе размер трубки (ы) для ожидаемого объема мастер смеси 5 мин до мастер-микс подготовки. Для обеспечения матричного геля не затвердевает при обработке, очень важно, чтобы охладить кончик пипетки, пайпетируя матричный гель 3-4 раза до переноса его в новую трубку. - Добавьте 80 qL матричной гель/клеточной смеси на скважину 24-хорошо пластины. Рекомендуется прокладка капли на нижнюю половину стены скважины, избегая при этом прямого контакта с покрытием Poly-HEMA. После добавления матричного геля, закружить пластины, чтобы матричной гель / клеточная смесь, чтобы сформировать кольцо вокруг обода колодца.
- Поместите 24-хорошо пластины в 37 c 5% CO2 инкубатор правой стороны в течение 10 минут, чтобы матричный гель частично затвердеть.
ПРИМЕЧАНИЕ: Начните нагревать органоидные средства мыши при 37 градусах Цельсия сразу после размещения 24-хорошей пластины в инкубаторе. - После инкубации в течение 10 минут, переверните 24-ну хорошую тарелку вверх дном и инкубировать еще 50 минут, чтобы матричной гель полностью затвердеть.
- Добавьте 350 л предварительно разогретых органоидных медиа-носителей мыши, которые скатятся в центр каждого колодца.
ПРИМЕЧАНИЕ: Для поддержания целостности матричного геля, очень важно, чтобы избежать матричного геля кольцо при добавлении средств массовой информации. - После добавления средств массовой информации, вернуть 24-хорошо пластины в 37 C 5% CO2 инкубатора.
3. Пополнение мышь органоидных средств массовой информации - ВРЕМЯ: 10-15 Мин за 24-хорошо плиты
ПРИМЕЧАНИЕ: Существующие носители должны быть заменены свежими носителями каждые 48 ч. Перед каждой сменой носителя, предварительно теплые мыши органоидных носителей. Не обязательно добавлять ингибитор ROCK в средства, используемые для пополнения.
- Наклоните 24-колодую пластину под углом 45 градусов и аккуратно удалите существующие носители из центра каждого колодца, используя пипетку p1000, избегая при этом кольца матричного геля.
- Добавьте 350 л предварительно разогретых органоидных носителей мыши, как и в шаге 2.9. Рекомендуется добавлять больший объем носителей (до 1 мл) в органоиды, культивированные в течение более 5 дней, чтобы предотвратить быстрое истощение ключевых питательных веществ и факторов роста.
4. Извлечение белка лизата из органов простаты для анализа западной подошвы - ВРЕМЯ: 2,5-4 H
ПРИМЕЧАНИЕ: Перед сбором органоидов для экстракции белка лизат, подготовить и предварительно теплой диспазы-содержащих средств массовой информации (Таблица 1).
- Удалите носители с каждого из них, а также в шаге 3.1.
- Для сбора органоидов, неоднократно взрыва матричного геля путем pipetting 1 мл диспаз-содержащих носителей непосредственно на матричного геля кольцо, пока все кольцо выбили, и передать 1,5 мл микроцентрифуг трубки.
ПРИМЕЧАНИЕ: Очень важно избегать прямого контакта с скважинами с покрытием Poly-HEMA. Прямой контакт может привести к загрязнению собранного материала поли-HEMA, что может негативно повлиять на выживание клеток. - Поместите микроцентрифугную трубку 1,5 мл (ы) в инкубатор 37 градусов по Цельсию 5% CO2 в течение 30 мин до 1 ч, чтобы обеспечить полное переваривание матричного геля путем диспази.
- Пелле органоиды центрифугирование при 800 х г в течение 5 мин на RT и удалить супернатан с помощью микропипетля.
- Добавить фосфат-буферный солен (PBS) в органоидные гранулы и resuspend, мягко щелкая.
ПРИМЕЧАНИЕ: Неспособность достаточно восстановить органоидные гранулы может привести к загрязнению органоидного материала с остаточной диспазы или матричного геля. - Пелле органоидов центрифугированием при 800 х г в течение 5 мин на РТ и удалите супернатанс с помощью микропипети.
- Быстро заморозить органоидные гранулы, поместив каждую трубку в раствор, содержащий сухой лед и метанол. Храните трубку (ы) до будущего использования при -80 градусах Цельсия. Кроме того, экстракт белка лизат сразу же после шага 4.6.
- Повторите органоидные гранулы в 100 л буфера лиза белка(таблица 1) на 10 л объема упакованных клеток. Флик, чтобы приостановить.
ПРИМЕЧАНИЕ: При возобновлении после быстрой заморозки убедитесь, что буфер лиза белка размораживается до удаления образцов из -80 градусов по Цельсию, так как буфер лисиса должен быть немедленно добавлен в образцы, чтобы предотвратить фосфатазу и активность протеазы. - Инкубировать образцы в буфере лиза белка на льду, по крайней мере 45 мин.
ПРИМЕЧАНИЕ: Рекомендуется снотировать сядок перед инкубации на льду для повышения эффективности восстановления ядерного белка; однако, sonication не требуется. Если звукование не выполняется, приступайте к шагу 4.10.- Чтобы сонести, погрузить трубки во влажный лед и аккуратно нанесите кончик звукового дисмбератора на внешнюю часть микроцентрифуговой трубки. Sonicate для 40 s при 20 кГц.
- Переходке к западному помету в соответствии с установленными протоколами.
5. Фиксация и окрашивание органов простаты для иммуногистохимического анализа с помощью конфокальной микроскопии
-
Сбор органов простаты из 24-колодцев - ВРЕМЯ: 45-60 мин
ПРИМЕЧАНИЕ: При сборе органов простаты для обработки конфокальной микроскопии, очень важно обращаться с ними с осторожностью, чтобы сохранить их структуру. Протокол сбора ниже предназначен для уменьшения нарушения органоидной структуры во время изоляции.- Удалите носители с каждого из них, а также в шаге 3.1.
- Переварить матричного геля путем инкубации с 500 зл и содержащих средства массовой информации(Таблица 1) в течение 30 минут в 37 C 5% CO2 инкубатора.
- Соберите переваренные органоидные суспензии в микроцентрифуговой трубке и гранулы органоидов центрифугированием при 800 х г в течение 3 мин на РТ. Удалите супернатант.
-
Цельномонтаж иммунофлуоресцентного окрашивания органов простаты - ВРЕМЯ: 3-4 дня (1-5 ч/день)
- Добавьте 500 л 4% параформальдегида в PBS и инкубировать в течение 2 ч на RT с нежной тряской.
- Гралет органоиды центрифугации при 800 х г в течение 3 мин на RT, удалить супернатант, и мыть гранулы с 1 мл PBS в течение 15 минут с нежной встряхивания.
- Вымойте гранулы, как в шаге 5.2.2 для дополнительных два раза.
- Пелле органоидов центрифугированием при 800 х г в течение 3 мин на РТ и удалите супернатант. Добавить 1 мкг/мл DAPI в блокирующем растворе(таблица 1). Инкубировать в течение 2 ч на RT или в качестве альтернативы на ночь при 4 градусах Цельсия с нежной тряской.
- Пелле органоидов центрифугированием при 800 х г в течение 3 мин на РТ и удалите супернатант. Добавить первичное антитело (кролик анти-p63, мышь анти-цитокератин 8) в блокирование раствора и инкубировать на ночь при 4 градусах Цельсия с нежной встряхивания.
- Пелле органоидов центрифугированием при 800 х г в течение 3 мин на РТ и удалите супернатант. Вымойте гранулы с 1 мл PBS в течение 15 минут с нежной встряхивания.
- Вымойте гранулы, как в шаге 5.2.6 для дополнительных два раза.
- Пелле органоидов центрифугированием при 800 х г в течение 3 мин на РТ и удалите супернатант. Добавить вторичное антитело (коза анти-кролик IgG-Alexa Fluor 594, коза анти-мышь IgG-Alexa Fluor 488) в блокирующем растворе и инкубировать ночь при 4 градусов по Цельсию с нежной тряской.
- Гралет органоиды центрифугации при 800 х г в течение 3 мин на RT, удалить супернатант, и мыть гранулы с 1 мл PBS в течение 15 минут с нежной встряхивания.
- Вымойте гранулы, как в шаге 5.2.9 для дополнительных два раза.
6. Очистка тканей и монтаж окрашенных органов простаты для целостной конфокальной микроскопии - ВРЕМЯ: 7 H
- Пелле органоидов центрифугированием при 800 х г в течение 3 мин на РТ и удалите супернатант.
- Добавьте 1 мл 30% сахарозы в PBS с 1% Triton X-100 и инкубировать в течение 2 ч на RT с нежной встряхивания.
- Пелле органоидов центрифугированием при 800 х г в течение 3 мин на РТ и удалите супернатант.
- Добавьте 1 мл 45% сахарозы в PBS с 1% Triton X-100 и инкубировать в течение 2 ч на RT с нежной встряхивания.
- Пелле органоидов центрифугированием при 800 х г в течение 3 мин на РТ и удалите супернатант.
- Добавьте 1 мл 60% сахарозы в PBS с 1% Triton X-100 и инкубировать в течение 2 ч на RT с нежной встряхивания.
- Пелле органоидов центрифугированием при 800 х г в течение 3 мин на РТ и удаляйте 95% супернатанта.
ПРИМЕЧАНИЕ: Гранулы становятся слабее, как концентрация сахарозы становится выше. Наблюдение DAPI окрашенных органоидов под ультрафиолетовым светом, чтобы подтвердить, что они не были потеряны во время удаления супернатанта рекомендуется. - Перенесите каплю оставшейся подвески на камерную крышку и перейдите к конфокальной микроскопии.
ПРИМЕЧАНИЕ: Фрагменты coverslip могут быть размещены по обе стороны от капли, которые будут использоваться в качестве прокладок(рисунок 4C). Они предотвращают коллапс органоидов, когда над каплей помещается крышка.
Результаты
Эпителиальные клетки предстательной железы покрыли в культуре органов мыши, где они образуют органоиды, которые собирают до подготовки к анализу вниз по течению(рисунок 1).
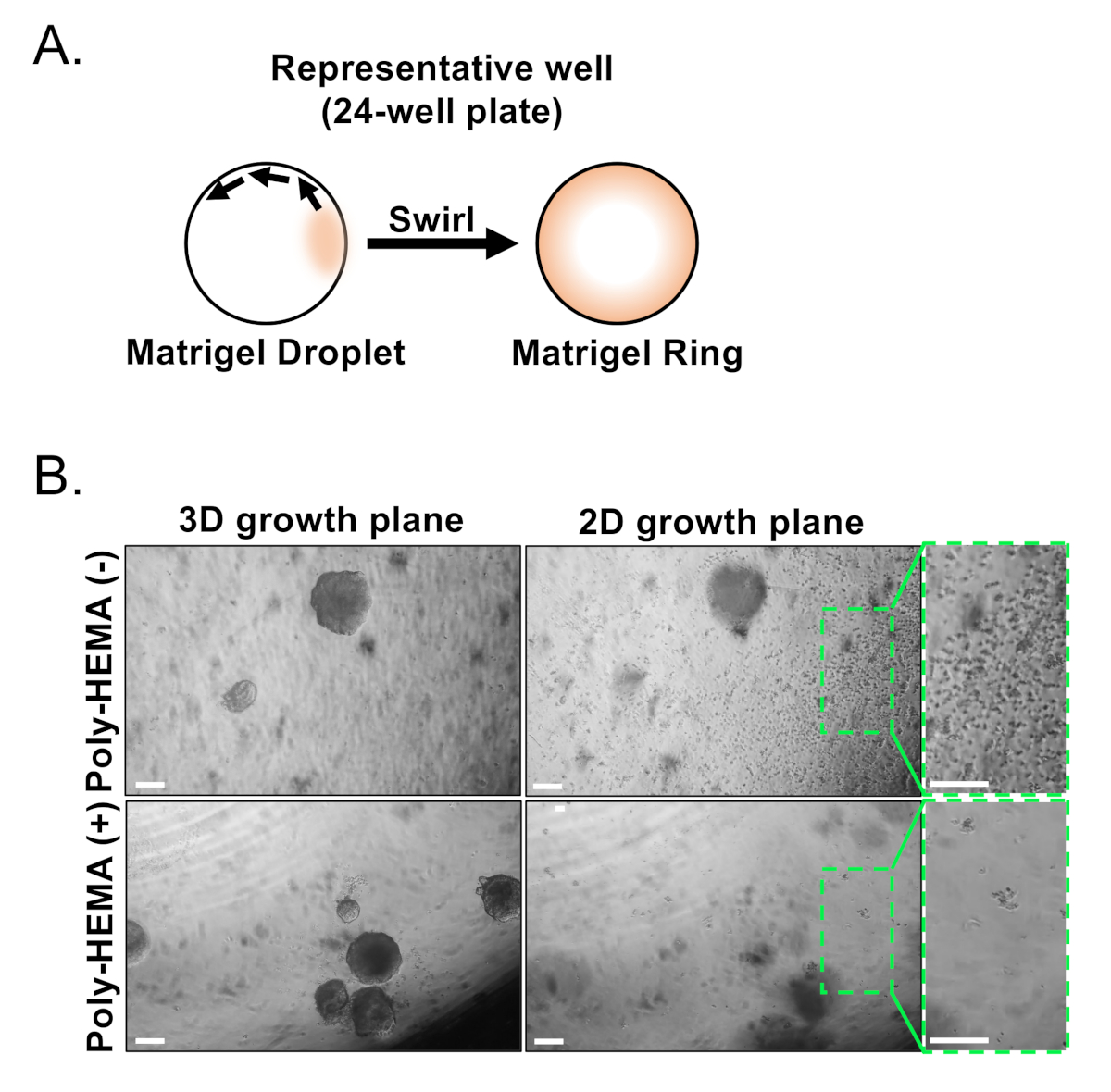
Базальные и светящиеся эпителиальные клетки изолированы с помощью FACS. После исключения DAPI- клеток и истощениялиняных клеток (CD45, CD31, Ter119), базальные и светящиеся клетки отличаются на основе дифференциального выражения EpCAM и CD49f (Рисунок 2). Подход, описанный в пластине базальных и светящихся клеток в органоидной культуры влечет за собой: (1) покрытие клеток в матричных гелевых колец, и (2) покрытие скважин с Поли-HEMA. Покрытие в кольца было ранее описано в Agarwal и др.9. Использование этого подхода(рисунок 3A) позволяет исследователям легче избежать матричного геля при пополнении средств массовой информации (Шаг 3), и легче рассчитывать органоидов, следуя окружности скважины. Покрытие скважин с Поли-HEMA было показано, чтобы предотвратить 2D колонии формирования в органоидов в женелинене17; однако этот подход не был использован в модели органоидов простаты. Важно отметить, что покрытие скважин поли-HEMA(Таблица 3) устраняет возникновение 2D колоний, не мешая органоидного образования(рисунок 3B). Эти изменения расширяют возможности органоидного ассирования простаты.
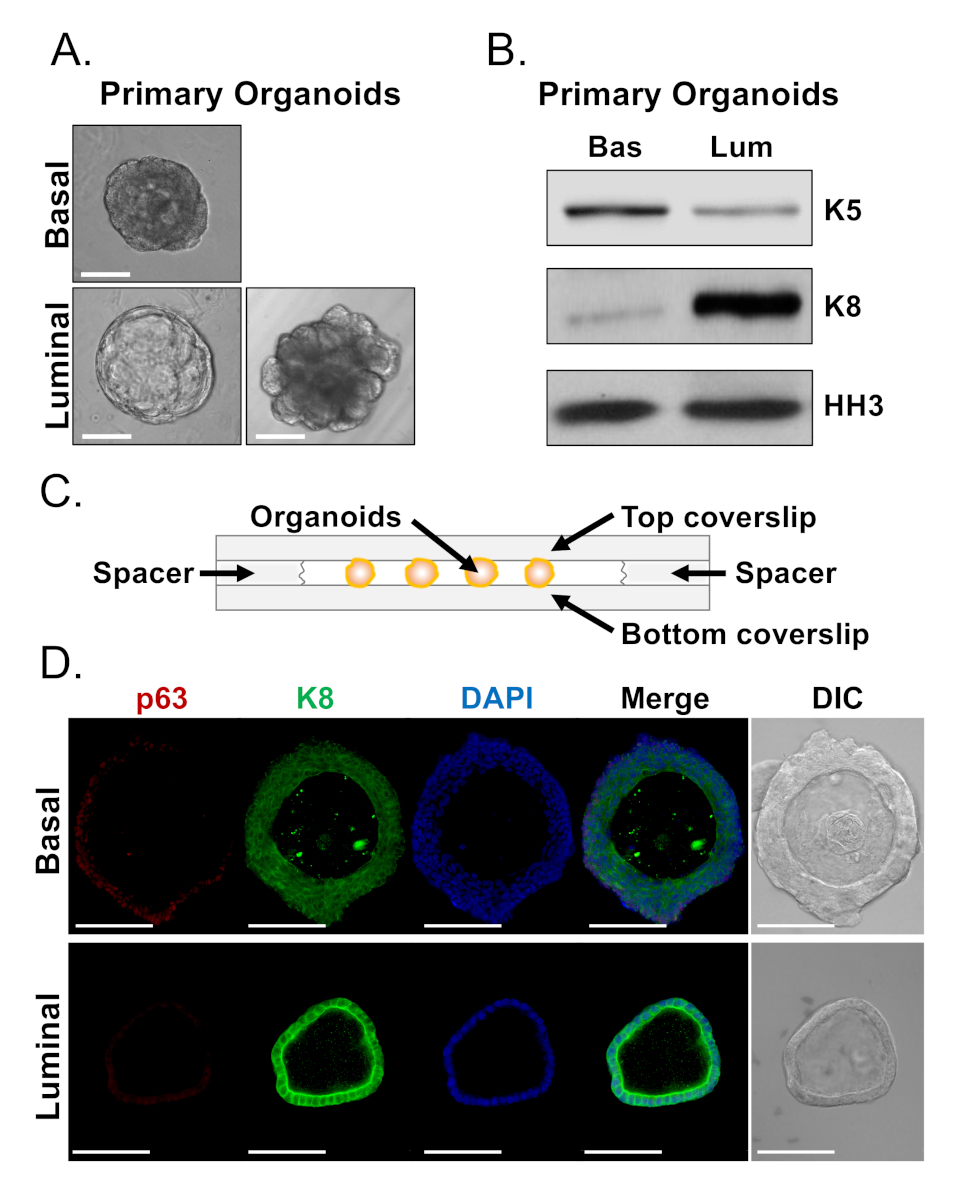
Базальные и светящиеся клетки образуют органоиды с различными морфологиями(рисунок 4А). В то время как большинство органоидов базально-производной являются похожими по размеру (100-300 мкм диаметр) после 7 дней в культуре, органоиды, полученные из света, обладают значительной неоднородностью (диаметр 30-450 мкм). Кроме того, большинство базально-полученных органоидов содержат люмены, окруженные многослойным эпителием(рисунок 4А, верхняя),в то время как органоиды, полученные из светила, варьируются в морфологии от полого, с однослойным эпителием до твердых, с многослойными шнурами клеток, которые не канализируются(рисунок 4А, внизу). Описанные выше подходы для подготовки органоидов для анализа ниже по течению (Шаги 4, 5) были использованы для исследования того, отражают ли эти фенотипические различия различий в выражении маркера линии. Западный анализ поблуня показал, что базальные и светиловые органоиды сохраняют особенности, связанные с базальными и светящимися первичными клетками. Органоиды, полученные из базального производства, выражают более высокие уровни базального маркера цитокератин 5 (K5), в то время как органоиды, полученные из светила, выражают более высокие уровни цитокератина светящегося светового маркера 8 (K8)(рисунок 4B). Оба базальных и светящихся маркеров были обнаружены в базальных и светящихся полученных органоидов в массовой популяции, возможно, наводящий на дифференциацию (Рисунок 4B).
Мы стремились охарактеризовать выражение маркера линии в базально-полученных органоидов и определить, является ли морфологически различных светящихся полученных органоидов обладают различиями в выражении маркера путем окрашивания нетронутых органоидов и выполнения конфокальной микроскопии (Рисунок 4D). Органоиды базального производства содержали многослойный эпителий с внешними слоями, выражающими высокие уровни базального маркера p63 и умеренные уровни светящегося маркера K8 (p63hi,K8mid),и внутренних слоев без обнаруживаемых уровней p63 и высоких уровней K8 (p63lo,K8hi)(Рисунок 4D, верхний). В то время как все клетки в однослойных световых органоидов положительно окрашенных положительно для K8, только избранные клетки содержали ядерные p63(Рисунок 4D, дно). Эти данные подтверждают подходы к сбору урожая и подготовке органоидов для анализа западной помарки или конфокальной микроскопии и тем самым расширяют возможности органоидного анализа для изучения ключевых клеточных процессов, включая дифференциацию.

Рисунок 1: Схематический иллюстрирующие рабочий процесс для генерации органов простаты для сбора и анализа. Общая простата мыши разобщена и базальных и светящихся эпителиальных клеток предстательной железы изолированы флуоресценции активированной сортировки клеток через установленные протоколы18,19. Базальные или светящиеся клетки, подвешенные в смеси органоидных носителей мыши и матричного геля, покрываются в кольца митричного геля. После 5-7 дней культуры, органоиды собирают для анализа западной помок или конфокальной микроскопии. Пожалуйста, нажмите здесь, чтобы просмотреть большую версию этой цифры.
{kind=link}

Рисунок 2: Изоляция базальных и светящихся клеток эпителия простаты с помощью флуоресценции активированной сортировки клеток (FACS). Разъединенные клетки от простаты мыши запятнаны DAPI, для того чтобы различить в реальном маштабе времени от мертвых клеток, и антитела поверхности, для того чтобы различить базальные от световых клеток, до FACS. Слева - Пригнена на ячейках DAPI- FSC-A - вперед-разброс. Центр - Пригнайна на Лин- клетки (CD45lo, CD31lo, Ter119lo). SSC-A - боковой рассеяние. Справа - Базальные клетки (Бас) (EpCAMпривет, CD49fпривет), Luminal клетки (Lum) (EpCAMпривет, CD49fсередине). Пожалуйста, нажмите здесь, чтобы просмотреть большую версию этой цифры.
{kind=link}

Рисунок 3: Создание органов простаты мыши. (A) Схематический иллюстрируя подход для создания кольца матричного геля в колодце 24-колодца пластины. (B) Представитель фазы контрастные изображения органоидов (3D плоскости роста) и двумерных колоний (2D плоскости роста) формируется 7 дней после покрытия эпителиальных клеток простаты в не-покрытие (Поли-HEMA (-)), или покрытием (Поли-HEMA (я)) 24-ну хорошо пластин. Коробок регионов в 2D плоскости роста увеличиваются справа. Шкала баров 200 мкм. Пожалуйста, нажмите здесь, чтобы просмотреть большую версию этой цифры.
{kind=link}

Рисунок 4: Анализ экспрессии маркеров линии в органоидах простаты по западной пометке и цельномонтажной конфокальной микроскопии. (A) Представитель фазы контрастные изображения базально-производных (вверху), и светило-полученных (нижних) органоидов после 7 дней культуры. Шкала бар No 100 мкм. (B) Западный анализ подьение базально-производных (Bas) и светило производных (Lum) органоидов после 5 дней культуры. Окрашивание для базального маркера, цитокератин 5 (K5) и светящийся маркер, цитокератин 8 (K8), а также контроль нагрузки, гистон H3 (HH3). (C) Схематическая иллюстрирующая камерная обложка с помощью прокладок. (D) Представитель дифференциального интерференционного контраста (DIC) и иммунофлуоресцентных изображений базальных (сверху) и светящихся (нижних) органоидов после 7 дней культуры. Окрашивание для p63 (красный), K8 (зеленый) и DAPI (синий) индивидуально и слились. Шкала баров 100 мкм. Пожалуйста, нажмите здесь, чтобы просмотреть большую версию этой цифры.
{kind=link}
| Рецепты | |
| Диспаз-содержащие средства массовой информации | 1 мг/мл диспазы и 10 мкм ингибитора ROCK в продвинутых DMEM F12. Фильтр стерилизовать с помощью 0,22 мкм фильтра. |
| Средства диссоциации | 10% FBS - 1x пенициллин-стрептомицин в RPMI 1640. Фильтр стерилизовать с помощью 0,22 мкм фильтра. |
| Буфер протеинового лиза | RipA буфера и ингибиторы фосфатазы и ингибиторы протеазы |
| Блокирующее решение | 10% FBS в PBS с 0.2% Тритон X-100 |
Таблица 1: Инструкции по подготовке ключевых решений.
| Компонент | Концентрации |
| B-27 | 1x (разбавленный из 50x концентрата) |
| ГлутаМАКС | 1x (разбавленный из 100x концентрата) |
| N-ацетил-L-цистеин | 1,25 мМ |
| Нормоцин | 50 мкг/мл |
| Рекомбинантный человек EGF, без животных | 50 нг/мл |
| Рекомбинантный человек Ноггин | 100 нг/мл |
| R-спондин 1-кондиционированные носители | 10% условные носители |
| A83-01 | 200 нм |
| Dht | 1 нм |
| Y-27632 дигидрохлорид (ингибитор ROCK) | 10 км |
| Расширенный DMEM/F-12 | Базовые носители |
| R-спондин 1-кондиционированные носители генерируется, как описано в Drost, и др.13. После добавления всех компонентов, фильтр стерилизовать мыши органоидных носителей с помощью 0,22 мкм фильтра. Ингибитор ROCK добавляется только при установлении культуры и прохождении органоидов. | |
Таблица 2: Инструкции по подготовке органов мыши.
| Протокол для подготовки поли-HEMA покрытием пластин | |
| 1 | Добавьте 0,25 г Поли-ХЕМА к 50 мл 98% EtOH. Растворите Poly-HEMA при 37 градусах По Цельсия на шейкере. Этот процесс занимает не менее 4 ч. |
| 2 | Фильтр стерилизовать Poly-HEMA с помощью фильтра 0,22 мкм. |
| 3 | Добавьте 200 л раствора Poly-HEMA на скважину 24-хорошо пластины (ы). |
| 4 | Удалите крышку (ы) из 24-хорошо пластины (ы) после добавления Poly-HEMA и позволяют раствору испаряться на ночь. |
| 5 | Вымойте каждый колодец дважды с PBS и обеспечить скважины полностью сухой до хранения после окончательной стирки. ПРИМЕЧАНИЕ: Нарушение поли-HEMA покрытие во время стирки может способствовать 2-мерного роста при покрытии эпителиальных клеток в органоидной культуры. Чтобы предотвратить повреждение скважин с поли-ХЕМА, избегайте прямого контакта с наконечником пипетки во время мытья. Целостность Poly-HEMA покрытием скважин останется нетронутым, если Поли-HEMA соскребли на кончике пипетки. |
| 6 | Пластины с поли-ХЕМА с покрытием могут храниться при 4 градусах Цельсия в течение двух недель. ПРИМЕЧАНИЕ: Упаковка пластин в парафильме перед хранением снизит риск заражения. |
Таблица 3: Протокол для подготовки поли-HEMA покрытием пластин.
Обсуждение
Дифференциация эпителиальной клетки простаты была вовлечена в нормальную биологию простаты2,3,4,5,6,7 и биологию болезни8,10,11,12; однако главные регулирующие органы этого процесса остаются неопределенными. Выявление ключевых регуляторов дифференциации эпителиальных клеток простаты было трудно отчасти из-за отсутствия устоявшихся контекстов для ее моделирования. В то время как 2D монослойная культура может быть использована для моделирования дифференциации11,12,этот контекст не может резюмировать сложную микросреду простаты. Кроме того, в контекстах vivo для моделирования дифференциации не поддаются механистическим исследованиям, поскольку они являются сложными для манипулирования. Таким образом, определение легко манипулировать, но физиологически-соответствующий контекст, для изучения дифференциации имеет решающее значение.
Модель органоидов простаты представляет собой элегантный контекст ex vivo, где, как сообщается, происходит базальная дифференциация светящихся. Методы установления органов простаты хорошо установлены14; однако необходима дальнейшая оптимизация этих методов. Кроме того, четко не описаны подходы к сбору и подготовке органов простаты для анализа. Эта статья описывает подход к пластине эпителиальных клеток простаты, изолированных от мыши простаты в органоидной культуры. Такой подход позволяет исследователям (1) предотвратить появление 2D колоний во время органоидного образования, (2) снизить риск нарушения матричного геля во время пополнения мультимедиа, и (3) более эффективно считать органоиды. Кроме того, в этой рукописи излагаются подходы к сбору органоидов для подготовки к анализу западной подевой или всемонта конфокальной микроскопии. Важно отметить, что подход, используемый для подготовки органоидов для конфокальной микроскопии поддерживает нетронутую структуру органоидов через его продолжительность, что уменьшает повреждение органоидов до приобретения изображения. В целом, описанные подходы расширяют возможности органоидного ассирования простаты.
Примечательно, что органоиднообразующая способность базальных и светящихся клеток может быть изменена как методами, используемыми для изоляции соответствующих популяций, так и культурными условиями. Органоидные условия культуры, используемые в этом ассее, были впервые описаны Karthaus et al.13. В то время как Karthaus et al. сообщили, что базальные клетки обладают более высокой органоидной способностью формирования (15%) чем светящиеся клетки (1%)13, Чуа и др., используя различные методы изоляции и культурные условия, сообщили, что светящиеся клетки (0,2-0,3%) имеют более высокую органоидно-образующую способность, чем базальные клетки (0,03%)20. В целом, методы, описанные Karthaus et al., приводят к более высоким органоидным темпам формирования как для базальных, так и для светящихся клеток, что, вероятно, отражает различия в подходе, используемом для изоляции базальных и светящихся клеток13,в отличие от культурных условий, которые направлены против органоидного образования из светящихся клеток. Остается неясным, является ли протокол, описанный в этой рукописи способствует образованию светящихся органоидов от многопотентных светящихся прародителей, или совершенные светящиеся потомки9. Хотя своевременно и экономически запретительным, in vivo линии отслеживания исследований могут быть использованы для проверки прародителей особенности, связанные с различными эпителиальных линий простаты выяснял в органоидных исследования.
Такие процессы, как развитие, дифференциация и трансформация, имеют отношение не только к биологии простаты, но и имеют отношение к биологии других тканей, включая мозг, легкие, кишечник, поджелудочную железу и печень. Описанные методы облегчают использование органоидной модели для изучения этих процессов не только в предстательной железе, но и в широком диапазоне тканей.
Раскрытие информации
Авторам нечего раскрывать.
Благодарности
PDC и JMG поддерживаются Рут Л. Киршштейн Национальной исследовательской службы премии GM007185. JAD поддерживается Национальным институтом общих медицинских наук Национальных институтов здравоохранения (R25GM055052), присуждаемой Т. Хассону и стипендией Саула Мартинеса. ASG поддерживается Фондом Фонда семьи Спитцерии и Фондом Джилл. Эта работа была поддержана Американского онкологического общества (RSG-17-068-01-TBG), Министерство обороны (W81XWH-13-1-0470), Маргарет Е. Ранние медицинские исследования Trust, NIH / NCI (P50CA092131/UCLA SPORE в рак предстательной железы), Роуз Хиллз Фонд, и поддержка со стороны UCLA в Jonsson Всеобъемлющий онкологический центр, Широкий центр стволовых клеток, клинической и трансляционной научно-исследовательский институт, и Институт урологической онкологии.
Материалы
| Name | Company | Catalog Number | Comments |
| µ-Dish 35 mm, high | ibidi | 81156 | |
| 16% Paraformaldehyde | Thermo Fisher Scientific | 50-980-487 | |
| 4’,6-diamidino-2-phenylindole (DAPI) | Thermo Fisher Scientific | D1306 | |
| A83-01 | Tocris | 2939 | |
| Advanced DMEM/F-12 | Thermo Fisher Scientific | 12634010 | |
| APC/Cy7 anti-mouse CD326 (Ep-CAM) Antibody, 100 μg | BioLegend | 118218 | |
| B-27 Supplement (50x), Serum Free | Thermo Fisher Scientific | 17504044 | |
| Complete Protease Inhibitor Cocktail | Sigma | 11836145001 | |
| (DiHydro)testosterone (5α-Androstan-17β-ol-3-one) | Sigma | A-8380 | |
| Dispase II, Powder | Thermo Fisher Scientific | 17-105-041 | |
| Fetal Bovine Serum (FBS) | Sigma | F8667 | |
| FITC anti-mouse CD31 Antibody (0.5 mg/mL, 50 μg) | BioLegend | 102405 | |
| FITC anti-mouse CD45 Antibody (0.5 mg/mL, 50 μg) | BioLegend | 103107 | |
| FITC anti-mouse TER-119/Erythroid Cells Antibody (0.5 mg/mL, 50 μg) | BioLegend | 116205 | |
| Goat anti-mouse IgG-Alexa Fluor 488 | Invitrogen | A28175 | |
| Goat anti-rabbit IgG-Alexa Fluor 594 | Invitrogen | A11012 | |
| GlutaMAX | Thermo Fisher Scientific | 35050061 | |
| Halt Phosphatase Inhibitor | Thermo Fisher Scientific | 78428 | |
| Matrigel GFR Membrane Matrix | Corning | CB-40230C | |
| Mouse anti-cytokeratin 8 | BioLegend | 904804 | |
| N-acetyl-L-cysteine | Sigma | A9165 | |
| Normocin | Thermo Fisher Scientific | ant-nr-1 | |
| PE anti-human/mouse CD49f Antibody | BioLegend | 313612 | |
| Penicillin-Streptomycin (10,000 U/mL) | Thermo Fisher Scientific | 15-140-122 | |
| Poly(2-hydroxyethyl methacrylate) (Poly-HEMA) | Sigma | P3932-25G | |
| Rabbit anti-p63 | BioLegend | 619002 | |
| Radioimmunoprecipitation assay (RIPA) | Thermo Fisher Scientific | PI89901 | |
| Recombinant Human EGF, Animal-Free | PeproTech | AF-100-15 | |
| Recombinant Human Noggin | PeproTech | 120-10C | |
| RPMI 1640 Medium, HEPES (cs of 10) | Thermo Fisher Scientific | 22400105 | |
| Sonic Dismembrator | Thermo Fisher Scientific | FB120 | |
| Sucrose | Sigma | S0389-500G | |
| Triton X-100 | Sigma | X100-5ML | |
| Y-27632 dihydrochloride (ROCK inhibitor) | Selleck Chemical | S1049-50MG |
Ссылки
- Kwon, O. J., Xin, L. Prostate epithelial stem and progenitor cells. American Journal of Clinical and Experimental Urology. 2 (3), 209-218 (2014).
- Choi, N., Zhang, B., Zhang, L., Ittmann, M., Xin, L. Adult Murine Prostate Basal and Luminal Cells Are Self-Sustained Lineages that Can Both Serve as Targets for Prostate Cancer Initiation. Cancer Cell. 21 (2), 253-265 (2012).
- Ousset, M., Van Keymeulen, A., et al. Multipotent and unipotent progenitors contribute to prostate postnatal development. Nature Cell Biology. 14 (11), 1131-1138 (2012).
- Wang, J., et al. Symmetrical and asymmetrical division analysis provides evidence for a hierarchy of prostate epithelial cell lineages. Nature Communications. 5, 1-13 (2014).
- Wang, Z. A., Mitrofanova, A., et al. Lineage analysis of basal epithelial cells reveals their unexpected plasticity and supports a cell-of-origin model for prostate cancer heterogeneity. Nature Cell Biology. 15 (3), 274-283 (2013).
- Kwon, O. J., Zhang, B., Zhang, L., Xin, L. High fat diet promotes prostatic basal-to-luminal differentiation and accelerates initiation of prostate epithelial hyperplasia originated from basal cells. Stem Cell Research. 16 (3), 682-691 (2016).
- Kwon, O. J., Zhang, L., Ittmann, M. M., Xin, L. Prostatic inflammation enhances basal-to-luminal differentiation and accelerates initiation of prostate cancer with a basal cell origin. Proceedings of the National Academy of Sciences. 192 (3), 997-999 (2014).
- Stoyanova, T., et al. Prostate cancer originating in basal cells progresses to adenocarcinoma propagated by luminal-like cells. Proceedings of the National Academy of Sciences. 110 (50), 20111-20116 (2013).
- Agarwal, S., Hynes, P. G., et al. Identification of Different Classes of Luminal Progenitor Cells within Prostate Tumors. Cell Reports. 13 (10), 2147-2158 (2015).
- Ku, S. Y., et al. Rb1 and Trp53 cooperate to suppress prostate cancer lineage plasticity, metastasis, and antiandrogen resistance. Science. 355 (6320), 78-83 (2017).
- Mu, P., et al. SOX2 promotes lineage plasticity and antiandrogen resistance in TP53- and RB1-deficient prostate cancer. Science. 355 (6320), 84-88 (2017).
- Bishop, J. L., et al. The Master Neural Transcription Factor BRN2 Is an Androgen Receptor-Suppressed Driver of Neuroendocrine Differentiation in Prostate Cancer. Cancer Discovery. 7 (1), 54-71 (2016).
- Karthaus, W. R., et al. Identification of multipotent luminal progenitor cells in human prostate organoid cultures. Cell. 159 (1), 163-175 (2014).
- Drost, J., Karthaus, W. R., et al. Organoid culture systems for prostate epithelial and cancer tissue. Nature Protocols. 11 (2), 347-358 (2016).
- McCray, T., Richards, Z., Marsili, J., Prins, G. S., Nonn, L. Handling and Assessment of Human Primary Prostate Organoid Culture. Journal of Visualized Experiments. (143), e59051(2019).
- Lawson, D. A., Xin, L., Lukacs, R. U., Cheng, D., Witte, O. N. Isolation and functional characterization of murine prostate stem cells. Proceedings of the National Academy of Sciences of the United States of America. 104 (1), 181-186 (2007).
- Chen, H. Y., Kaya, K. D., Dong, L., Swaroop, A. Three-dimensional retinal organoids from mouse pluripotent stem cells mimic in vivo development with enhanced stratification and rod photoreceptor differentiation. Molecular vision. 22, 1077-1094 (2016).
- Liu, X., et al. Low CD38 Identifies Progenitor-like Inflammation-Associated Luminal Cells that Can Initiate Human Prostate Cancer and Predict Poor Outcome. Cell Reports. 17 (10), 2596-2606 (2016).
- Lukacs, R. U., Goldstein, A. S., Lawson, D. A., Cheng, D., Witte, O. N. Isolation, cultivation and characterization of adult murine prostate stem cells. Nature protocols. 5 (4), 702-713 (2010).
- Chua, C. W., Shibata, M., et al. Single luminal epithelial progenitors can generate prostate organoids in culture. Nature Cell Biology. 16 (10), 951-961 (2014).
Перепечатки и разрешения
Запросить разрешение на использование текста или рисунков этого JoVE статьи
Запросить разрешениеСмотреть дополнительные статьи
This article has been published
Video Coming Soon
Авторские права © 2025 MyJoVE Corporation. Все права защищены