
Method Article
유기체 배양을 사용하여 마우스 전립선 상피 세포의 분화 능력 평가
* 이 저자들은 동등하게 기여했습니다
요약
마우스 전립선 오르가노이드는 분화를 조절하는 메커니즘을 평가하는 유망한 맥락을 나타낸다. 이 논문은 전립선 오르가노이드를 확립하기 위한 개선된 접근법을 설명하고, (1) 오르가노이드로부터 단백질 용해액을 수집하고, (2) 전체 마운트 공초점 현미경검사법에 대한 수정 및 얼룩 오르가노이드를 수집하는 방법을 소개합니다.
초록
전립선 상피는 기저 세포와 발광 세포의 우세하게 구성됩니다. 생체 내 계보 추적은 개발, 조직 재생 및 변형 동안 마우스 전립선 기저 및 발광 세포의 분화 능력을 정의하는 데 활용되어 왔다. 그러나, 계보 추적 접근법을 사용하여 전립선 상피 분화 용량의 세포 내재 및 외인성 조절제를 평가하는 것은 종종 광범위한 번식을 필요로 하며 비용 부담이 될 수 있다. 전립선 오르가노이드 분석에서, 기저 및 발광 세포는 전립선 상피 ex 생체를 생성한다. 중요하게도, 원발성 상피 세포는 임의의 유전적 배경또는 마우스의 마우스로부터 3차원(3D) 배양으로 도금되기 전이나 후에 임의의 수의 소분자로 치료된 마우스로부터 분리될 수 있다. 분화 능력의 평가를 위한 충분한 재료는 7-10일 후에 생성된다. (1) 웨스턴 블롯에 의한 단백질 분석및 (2) 전체 마운트 공초점 현미경으로 손상되지 않은 오르가노이드의 면역 조직 화학 적 분석을 위한 기저 유래 및 발광 유래 오르가노이드의 수집은 연구원이 생체 내 분화를 평가할 수 있게 합니다 전립선 상피 세포의 용량. 조합에서 사용될 때, 이 2개의 접근은 유전 또는 약리학 조작에 응하여 전립선 기저 및 발광 세포의 분화 능력에 관하여 상보적인 정보를 제공합니다.
서문
기저 및 발광 세포는 전립선 상피1의대부분을 포함한다. 리니지 트레이싱 연구는 이러한 세포 유형이 성인 마우스2에서뚜렷한 선조에 의해 우세하게 지속된다는 것을 밝혔다; 그러나, 기저 전구체로부터의 발광 분화는 발달3,4,조직 재생5,염증6,7 및 전립선암 개시2,8을포함한 여러 맥락에서 관찰되었다. 더욱이, 신흥 데이터는 다능한 발광 선조뿐만 아니라 발광 에 전능한 선조9의존재를 지원합니다. 전이성 전립선암에서, AR 의존성 발광 계보로부터 기저 및 신경 내분비 기능을 가진 AR-무관심 혈통으로의 분화는 안드로겐 통로 억제제10,11,12에대한 내성의 점점 더 인정되는 메커니즘을 나타낸다. 따라서, 정상 생리학에서 분화가 연루됨에 따라, 암 개시 및 치료에 대한 내성, 전립선 상피 세포 분화의 주요 분자 조절기를 해명하는 것이 중요하다.
마우스 전립선 오르가노이드 모델은 전립선 상피 세포 분화를 연구하는 우아한 생체 내 문맥으로등장9,13,14. 이 분석에서, 개별 상피 세포는 1 주 안에 기저 세포와 발광 세포를 모두 포함하는 선 구조를 생성하는 3D 매트릭스로 도금됩니다. 오르가노이드 배양으로 세포를 도금하기 위한 기존 접근법은 오르가노이드를 효율적으로 생성하는 데 사용될 수 있지만, 이러한 접근법은 추가최적화(14)를필요로 한다. 전립선 오르가노이드 배양과 관련된 주목할 만한 과제는 (1) 분석으로부터 Matrigel(매트릭스 겔) 아래에 형성되는 2차원(2D) 콜로니를 제외하고, (2) 매질의 변화 동안 매트릭스 겔의 무결성을 유지하고, (3) 오르가노이드를 정확하게 계수하는 것을 포함한다. 이 논문은 마우스 전립선에서 분리된 상피 세포에서 오르가노이드를 생성하는 접근법을 간략하게 설명합니다. 설명된 접근법은 2D 콜로니의 발생을 방지하기 위해 폴리(2-하이드록세틸 메타크릴레이트)(Poly-HEMA)를 가진 코팅 판을 수반한다. 게다가, 세포는 매트릭스 젤 디스크보다는 매트릭스 젤 반지로 도금되고, 이는 매체를 바꾸고 오르가노이드를 세는 것을 덜 도전하게 합니다. 이 기술은 연구원이 어떻게 유전 변경 또는 작은 분자가 분화와 같은 중요한 프로세스를 바꾸기 전에, 또는 도중, 유기체 대형이 어떻게 소개되는지 더 쉽게 조사하는 것을 허용합니다.
전체 마운트 공초점 현미경 검사법에 의한 서쪽 얼룩 또는 면역 조직 화학 분석을 위한 전립선 오르가노이드의 수확은분화13에귀중한 기계론적인 통찰력을 제공할 수 있습니다, 그러나 그 같은 기술에 대한 오르가노이드를 준비하는 잘 확립된 프로토콜은 부족합니다. 이 원고는 (1) 단백질 용해물 또는 (2) 공초점 현미경 검사법에 대한 고정 및 염색에 대한 오르가노이드를 수확하는 방법을 설명합니다. 중요한 것은, 전립선 오르가노이드를 고정하고 염색하기 위해 기술된 접근법은 기존 방법과 관련하여 상당히 개선된다. 이들은 단면도 오르가노이드(15)에의존하는 동안, 이 원고에 기술된 방법은 견본 준비 도중 오르가노이드 손상으로부터 보호하는 것을 돕는 손상되지 않은 오르가노이드를 이용합니다. 조합에서 사용될 때, 서쪽 얼룩 및 공초점 현미경 검사법은 분화의 분자 조정기로 귀중한 통찰력을 제공할 수 있습니다. 또는 이러한 접근 방식을 사용하여 개발 및 변환과 같은 다른 프로세스를 모델링할 수 있습니다.
프로토콜
여기에 설명된 모든 방법은 로스앤젤레스 캘리포니아 대학의 기관 검토 위원회의 승인을 받았습니다.
참고: 그림 1에는이 백서에 설명된 접근 방식을 설명하는 회로도가 제공됩니다.
1. 형광 활성화 세포 선별 (FACS)을 사용하여 마우스 기저 및 발광 전립선 상피 세포를 분리 - 타이밍 : 30 분
참고: 어둠 속에서 1.3-1.5 단계를 수행합니다.
- 로손 외16에기재된 바와 같이 총 마우스 전립선으로부터 세포를 해리한 후, 세포를 FACS 튜브로 옮기고 해리매액의 100 μL에서 0.1-5 x 106 세포를 다시 중단한다(표1).
- CD45, CD31, Ter-119, EpCAM 및 CD49f : 다음과 같은 직접 공액 1 차 항체의 적절한 볼륨을 추가합니다.
- 20 분 동안 빛으로부터 보호 된 얼음에 배양하십시오.
참고: 얼룩이 없고 단일 염색된 대조군을 위해 총 해리 된 세포의 10 %를 사용하는 것이 좋습니다. 이러한 컨트롤은 분류에 대한 올바른 보정 및 전압을 설정하는 데 필요합니다. - 각 샘플에 해리 매체 1 mL을 첨가하여 항체 칵테일을 담금질합니다. 실온(RT)에서 800 x g에서 5분 동안 원심분리하여 세포를 펠렛하고 흡인하여 상류자를 제거한다.
- 1 μg/mL 4', 6-디아미디노-2-페닐린달(DAPI)을 함유하는 해리매의 적절한 부피(1 x 106세포당 250 μL)로 세포를 재중단시. FACS로 진행합니다. 마우스 기저 및 발광 전립선 상피 세포의 격리를 입증하는 유세포 분석 플롯은 도 2에예시되어 있다.
2. 1 차 마우스 오르가노이드 문화에 정렬 된 전립선 상피 세포를 도금 - 타이밍 : 2-3 H (폴리 HEMA 코팅 플레이트 준비 제외)
참고: 플레이트는 매트릭스 겔 아래 웰의 표면에 2D 콜로니 형성을 방지하기 위해 폴리 HEMA로 코팅된다. 폴리-HEMA 코팅 플레이트를 1일 전에 준비하여 정렬된 기저 또는 발광 전립선 상피 세포를 마우스 오르가노이드 배양으로 도금합니다. 감소된 성장 인자 매트릭스 겔의 1 mL aliquots를 해동, 이하 매트릭스 겔로 지칭, 얼음에 2 시간 전에 단계 2.1. Y-27632(ROCK 억제제)는 2.1단계 직전에 마우스 오르가노이드 매체에 첨가되어야 한다. 얼음에 2.1-2.8 단계를 수행합니다.
- 800 x g에서 원심분리에 의해 5 mL 둥근 바닥 튜브에서 세포를 4°C에서 5분 동안 펠렛하고 상월체를 흡인한다.
- 500 μL의 마우스 오르가노이드 매체로 세포 펠릿을 세척(표 2)14.
- 펠렛 세포를 800 x g에서 4°C에서 5분 동안 원심분리하고 상월체를 흡인한다.
- 1,000 세포 /μL의 세포 밀도에서 마우스 오르가노이드 매체에 다시 중단하십시오.
- 마스터 믹스를 준비하려면 마우스 오르가노이드 매체에 부유한 상피 세포를 매트릭스 겔과 혼합하여 25% 세포/매질 및 75% 매트릭스 겔을 포함하는 최종 혼합물을 생성합니다. 기저 세포는 일반적으로 100-2,000 세포/80 μL의 농도로 도금되는 반면, 발광 세포는 일반적으로 2,000-10,000 세포/80 μL의 농도로 도금됩니다. 도금 된 셀의 밀도는 예상 재료 수집 일 및 원하는 다운스트림 응용 프로그램에 따라 다릅니다.
참고: 마스터 믹스 준비 5분 전에 적절한 크기의 튜브를 식힙니다. 매트릭스 겔이 취급하는 동안 경화되지 않도록 하려면 매트릭스 겔을 새 튜브로 옮기기 전에 매트릭스 겔을 3-4회 파이펫팅하여 파이펫 팁을 냉각시키는 것이 중요합니다. - 24웰 플레이트의 웰당 매트릭스 젤/셀 혼합물 80 μL을 첨가합니다. 폴리 HEMA 코팅과의 직접적인 접촉을 피하면서 우물의 하반부에 액적을 피펫팅하는 것이 좋습니다. 매트릭스 겔을 추가한 후, 플레이트를 소용돌이돌려 매트릭스 겔/셀 혼합물이 우물 의 테두리 주위에 고리를 형성할 수 있도록 한다.
- 24웰 플레이트를 매트릭스 겔이 부분적으로 경화되도록 10분 동안 37°C 5%CO2 인큐베이터 우측에 놓습니다.
참고: 인큐베이터에 24웰 플레이트를 배치한 직후 37°C에서 마우스 오르가노이드 매체를 온난화하기 시작한다. - 10 분 동안 배양 한 후, 24 웰 플레이트를 거꾸로 뒤집고 매트릭스 젤이 완전히 굳어질 수 있도록 추가로 50 분 동안 배양하십시오.
- 350 μL의 미리 온난한 마우스 오르가노이드 매체를 각 우물의 중앙에 떨어뜨립니다.
참고: 매트릭스 겔의 무결성을 유지하기 위해, 매염을 첨가하면서 매트릭스 겔 링을 피하는 것이 중요하다. - 매체를 첨가한 후, 24웰 플레이트를 37°C 5%CO2 인큐베이터로 되돌린후.
3. 마우스 오르가노이드 미디어 보충 — 타이밍: 24웰 플레이트당 10-15분
참고: 기존 미디어는 48시간마다 새로운 매체로 교체해야 합니다. 각 미디어가 변경되기 전에 마우스 오르가노이드 미디어를 미리 따뜻하게 합니다. 보충에 사용되는 미디어에 ROCK 억제제를 추가할 필요는 없습니다.
- 매트릭스 젤 링을 피하면서 24 웰 플레이트를 45° 각도로 기울이고 p1000 파이펫을 사용하여 각 웰의 중앙에서 기존 용지를 부드럽게 제거합니다.
- 2.9단계에서와 같이 350 μL의 미리 온난마우스 오르가노이드 매체를 첨가합니다. 주요 영양소와 성장 인자의 급속한 고갈을 방지하기 위해 5 일 이상 배양 된 오르가노이드에 더 많은 양의 미디어 (최대 1 mL)를 추가하는 것이 좋습니다.
4. 웨스턴 블롯 분석을 위한 전립선 오르가노이드에서 단백질 용해물 추출 — 타이밍: 2.5-4 H
참고: 단백질 용해성 추출을 위한 오르가노이드를 수집하기 전에, 미리 온난디스파스 함유 매체를 준비하고 준비한다(표 1).
- 3.1단계에서뿐만 아니라 각 미디어에서 미디어를 제거합니다.
- 오르가노이드를 수집하기 위해, 전체 링이 빠질 때까지 매트릭스 겔 링에 직접 디스파스 함유 매체 1 mL을 피펫팅하여 매트릭스 겔을 반복적으로 폭발시키고, 1.5 mL 마이크로센트리퓨지 튜브로 옮김을 한다.
참고: 폴리-HEMA 코팅 웰과의 직접적인 접촉을 피하는 것이 중요합니다. 직접 접촉은 세포 생존에 부정적인 영향을 미칠 수있는 Poly-HEMA로 수집 된 물질의 오염을 일으킬 수 있습니다. - 1.5 mL 미세원원지 튜브(들)를 37°C 5%CO2 인큐베이터에 30분 내지 1시간 동안 배치하여 디스파스에 의해 매트릭스 겔의 완전한 소화를 허용한다.
- 펠렛 오르가노이드를 RT에서 800 x g에서 5분 동안 원심분리하고 마이크로파이펫을 사용하여 상상체를 제거한다.
- 인산염 완충 식염수(PBS)를 오르가노이드 펠릿에 넣고 부드럽게 쓸어넘기면서 다시 돌릴 수 있습니다.
참고: 오르가노이드 펠릿을 충분히 재중단시키지 않으면 잔류 디스파스 또는 매트릭스 겔로 오르가노이드 물질의 오염이 발생할 수 있다. - RT에서 5 분 동안 800 x g에서 원심 분리에 의해 오르가노이드를 펠렛하고 마이크로 파이펫을 사용하여 상위를 제거합니다.
- 각 튜브를 드라이 아이스와 메탄올을 함유한 용액에 넣어 오르가노이드 펠릿을 빠르게 동결시. 튜브를 -80°C에서 나중에 사용할 때까지 보관하십시오. 대안적으로, 추출물 단백질은 4.6단계 바로 다음에 해내한다.
- 단백질 용해 완충액의 100 μL에서 오르가노이드 펠릿을 다시 일시 중단(표 1)포장 된 세포 볼륨의 10 μL 당. 플릭하여 다시 일시 중단합니다.
참고: 빠른 동결 후 재개하는 경우, 인산염 및 프로테아제 활성을 방지하기 위해 용해 완충액을 즉시 시료에 추가해야 하므로 -80 °C에서 샘플을 제거하기 전에 단백질 용해 완충액이 해동되도록 하십시오. - 적어도 45 분 동안 얼음에 단백질 용해 완충제에 샘플을 배양.
참고: 핵 단백질 회복의 효율성을 높이기 위해 얼음에 배양하기 전에 초음파 처리하는 것이 좋습니다. 그러나 초음파 처리는 필요하지 않습니다. 초음파 처리가 수행되지 않으면 4.10 단계로 진행하십시오.- 초음파 처리하려면 젖은 얼음에 튜브를 잠수하고 소닉 디스메브레이터의 끝을 미세 원심 분리튜브 의 외부에 부드럽게 적용하십시오. 20 kHz에서 40 s에 대한 초음파.
- 확립된 프로토콜에 따라 서부 블롯으로 진행합니다.
5. 전체 마운트 공초점 현미경 검사법에 의한 면역 조직 화학 분석을위한 전립선 오르가노이드 고정 및 염색
-
24 웰 플레이트에서 전립선 오르가노이드 수집 — 타이밍: 45-60분
참고: 공초점 현미경 검사법을 위해 처리하기 위하여 전립선 오르가노이드를 수집할 때, 그들의 구조물을 유지하기 위하여 주의하여 취급하는 것이 중요합니다. 아래의 수집 프로토콜은 격리 하는 동안 유기 체 구조의 중단을 줄이기 위해 설계 되었습니다.- 3.1단계에서뿐만 아니라 각 미디어에서 미디어를 제거합니다.
- 37°C 5%CO2 인큐베이터에서 30분 동안 500 μL의 디스파제 함유 매폐(표1)로배양하여 매트릭스 겔을 소화한다.
- 미세 원심 분리튜브에 소화 된 오르가노이드 현탁액을 수집하고 RT에서 3 분 동안 800 x g에서 원심 분리하여 오르가노이드를 펠렛하십시오.
-
전립선 오르가노이드의 전체 마운트 면역 형광 염색 — 타이밍: 3-4 일 (1-5 시간 /일)
- PBS에 4% 파라포름알데히드의 500 μL을 넣고 부드러운 흔들림으로 RT에서 2시간 동안 배양합니다.
- RT에서 800 x g에서 3 분 동안 원심 분리로 오르가노이드를 펠렛하고 상류물을 제거하고 부드러운 흔들림으로 15 분 동안 PBS 1 mL로 펠릿을 씻으십시오.
- 5.2.2 단계에서와 같이 펠릿을 2회 더 세척합니다.
- RT에서 3 분 동안 800 x g에서 원심 분리에 의해 오르가노이드를 펠렛하고 상류를 제거합니다. 차단 솔루션에 1 μg/mL DAPI를 추가합니다(표1). RT에서 2 시간 동안 배양하거나 부드러운 흔들림으로 4 °C에서 밤새 배양하십시오.
- RT에서 3 분 동안 800 x g에서 원심 분리에 의해 오르가노이드를 펠렛하고 상류를 제거합니다. 1차 항체(토끼 항-p63, 마우스 항-시토케라틴 8)를 차단 용액에 넣고 4°C에서 밤새 배양하고 부드러운 흔들림으로 배양한다.
- RT에서 3 분 동안 800 x g에서 원심 분리에 의해 오르가노이드를 펠렛하고 상류를 제거합니다. 펠릿을 1 mL의 PBS로 15분 동안 부드럽게 흔들어 주세요.
- 5.2.6 단계에서와 같이 펠릿을 2회 더 세척합니다.
- RT에서 3 분 동안 800 x g에서 원심 분리에 의해 오르가노이드를 펠렛하고 상류를 제거합니다. 이차 항체(염소 항토끼 IgG-Alexa Fluor 594, 염소 항 마우스 IgG-Alexa Fluor 488)를 차단 용액에 넣고 부드러운 흔들림으로 4°C에서 밤새 배양한다.
- RT에서 800 x g에서 3 분 동안 원심 분리로 오르가노이드를 펠렛하고 상류물을 제거하고 부드러운 흔들림으로 15 분 동안 PBS 1 mL로 펠릿을 씻으십시오.
- 5.2.9 단계에서와 같이 펠릿을 2회 더 세척합니다.
6. 전체 마운트 공초점 현미경 검사법용 스테인드 전립선 오르가노이드의 조직 정리 및 장착 — TIMING: 7 H
- RT에서 3 분 동안 800 x g에서 원심 분리에 의해 오르가노이드를 펠렛하고 상류를 제거합니다.
- PBS에 30% 자당의 1 mL을 1% 트리톤 X-100으로 넣고 RT에서 부드러운 흔들림으로 2시간 동안 배양합니다.
- RT에서 3 분 동안 800 x g에서 원심 분리에 의해 오르가노이드를 펠렛하고 상류를 제거합니다.
- PBS에 45% 자당의 1 mL을 1% 트리톤 X-100으로 넣고 RT에서 부드러운 흔들림으로 2시간 동안 배양합니다.
- RT에서 3 분 동안 800 x g에서 원심 분리에 의해 오르가노이드를 펠렛하고 상류를 제거합니다.
- PBS에 60% 자당의 1 mL을 1% 트리톤 X-100으로 넣고 RT에서 부드러운 흔들림으로 2시간 동안 배양합니다.
- RT에서 3 분 동안 800 x g에서 원심 분리하여 오르가노이드를 펠렛하고 상수의 95 %를 제거합니다.
참고: 자당의 농도가 높아지면 펠릿이 느슨해집니다. 자외선 아래에서 DAPI 염색 된 오르가노이드를 관찰하여 상층부 제거 중에 손실되지 않은지 확인하는 것이 좋습니다. - 남은 현탁액의 10-20 μL 액적을 챔버 커버슬립으로 옮기고 공초점 현미경으로 진행한다.
참고: 커버슬립 조각은 스페이서로서 사용할 액적의 양쪽에 배치될 수있다(도 4C). 이들은 표지슬립이 물방울 위에 놓일 때 오르가노이드가 붕괴되는 것을 방지합니다.
결과
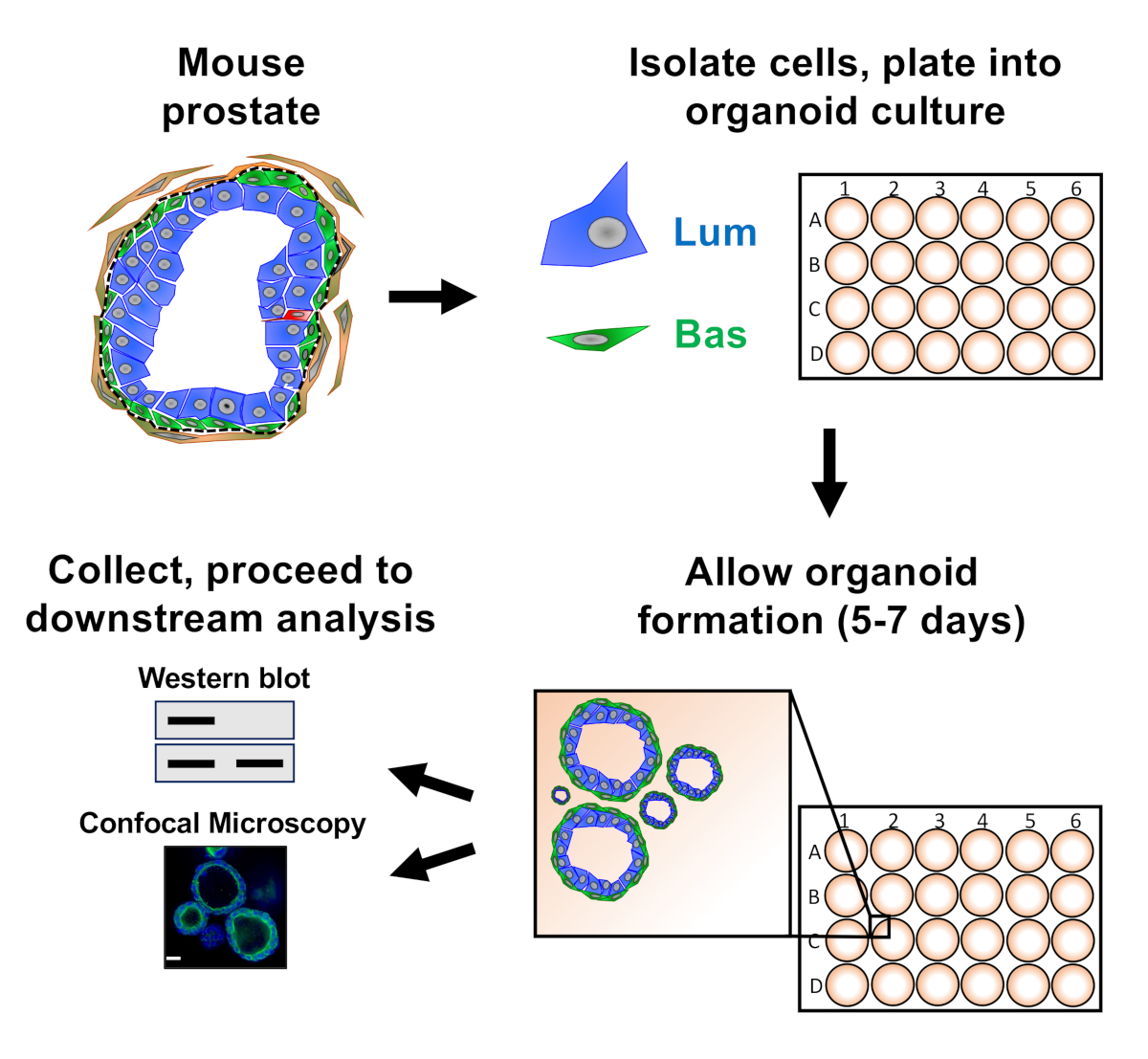
전립선 상피 세포는 마우스 오르가노이드 배양으로 도금되어 오르가노이드를 형성하며, 이는 하류 분석을 위한 준비 전에 수확된다(그림1).
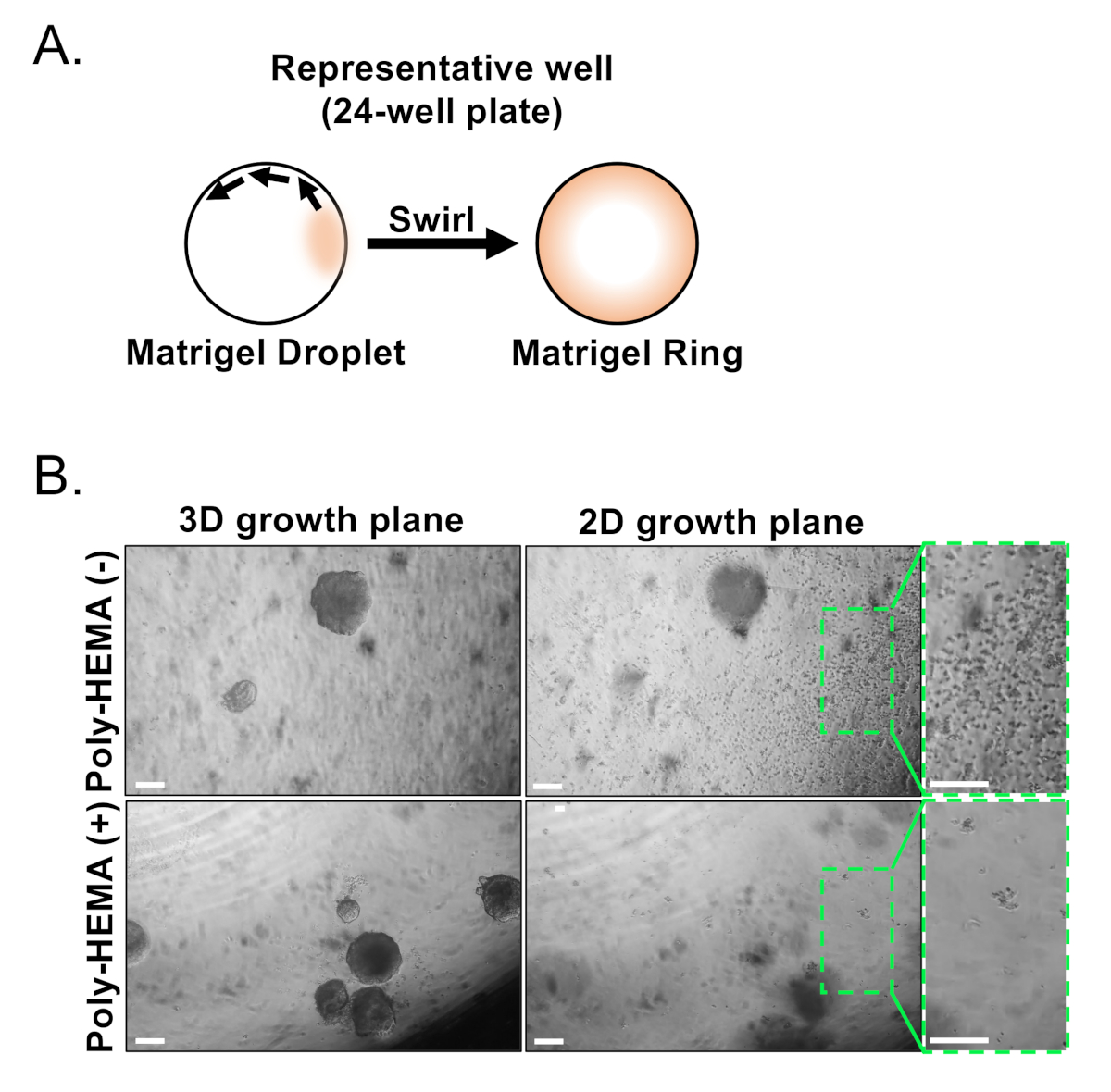
기저 및 발광 상피 세포는 FACS를 사용하여 단리된다. DAPI+ 세포를 배제하고 린+세포(CD45, CD31, Ter119)를 고갈한 후, 기저 및 광도 세포는 EpCAM 및 CD49f의 차분발 발현에 기초하여 구별된다(도2). 유기 체양 배양으로 플레이트 전립선 기저 및 발광 세포를 설명하는 접근 방식은 수반 : (1) 매트릭스 겔 링으로 세포를 도금, 및 (2) 폴리 HEMA와 우물코팅. 고리로 도금은 이전에 Agarwal 외9에기술되었습니다. 이러한접근법(그림 3A)을활용하여 연구자들은 매질의 둘레를 따라 서거노이드를 보다 쉽게 카운트하면서 매트릭스 겔을 보다 쉽게 피할 수 있다(3단계). 폴리-HEMA를 가진 코팅 우물은 망막 오르가노이드(17)에있는 2D 식민지 대형을 방지하기 위하여 보였습니다; 그러나, 이 접근법은 전립선 오르가노이드 모델에서 활용되지 않았다. 중요한 것은, 폴리-HEMA(표3)로코팅웰을 하여 오르가노이드 형성을 방해하지 않고 2D 콜로니의 발생을 제거한다(도3B). 이러한 변형은 전립선 오르가노이드 분석법의 기능을 확장합니다.
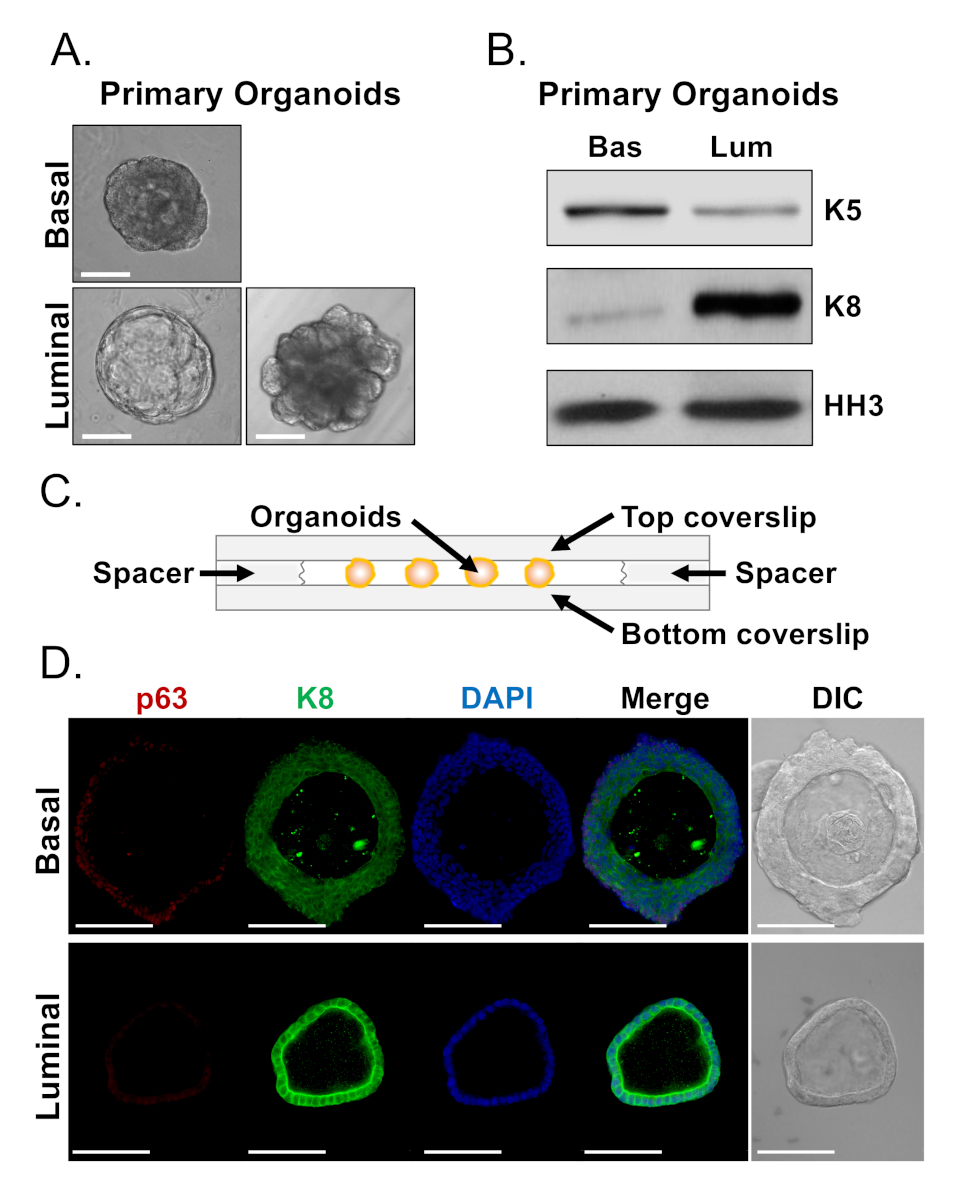
기저 및 발광 세포는 뚜렷한 형태와 오르가노이드를 형성한다(그림 4A). 대부분의 기저 유래 오르가노이드는 배양 7일 후 크기(100-300 μm 직경)와 유사하지만, 발광 유래 오르가노이드는 상당한 이질성(30-450 μm 직경)을 나타낸다. 또한, 대부분의 기저 유래 오르가노이드는 다층 상피에 둘러싸인 루멘을 포함(그림 4A, 상단),반면 발광 유래 오르가노이드는 중공에서 형태학 범위, 단층 상피고체로, 운하화하지 않는 세포의 다층 코드(그림 4A, 아래쪽). 위에서 설명한 접근법은 하류 분석을 위한 오르가노이드를 준비하기 위해(단계 4, 5), 이러한 표현형 차이가 계보 마커 발현의 차이를 반영하는지 여부를 조사하는 데 사용되었다. 서양 얼룩 분석은 기저 및 발광 유래 오르가노이드가 기저 및 발광 1 차 세포와 관련된 특징을 유지하는 것으로 나타났습니다. 기저 유래 오르가노이드는 기저 마커 시토케라틴 5(K5)의 상부를 발현하는 반면, 발광 유래 오르가노이드는 광마커 사이토케라틴 8(K8)의 상부를 발현한다(도4B). 기저 및 발광 마커는 모두 벌크 집단에서 기저 및 발광 유래 오르가노이드에서 검출되었으며, 아마도 분화의암시(도 4B).
우리는 기저 유래 오르가노이드에서 계보 마커 발현을 특성화하고 형태학적으로 뚜렷한 발광 유래 오르가노이드가 그대로 유기체를 염색하고 공초점 현미경 검사법을 수행함으로써 마커 발현에 차이가 있는지 여부를 결정하려고했습니다(그림 4D). 기저 유래 오르가노이드는 기저 마커 p63의 높은 수준을 발현하는 외부 층을 가진 다층 상피와 발광 마커 K8 (p63hi,K8mid)및 P8의 검출 가능한 수준 및 높은 수준의 K8 (p63lo,K8hi)의내부 층을 포함하였다(도 4D, 상단). K8에 대해 단층 발광 유래 오르가노이드의 모든 세포가 긍정적으로 염색되었지만, 선택된 세포만 핵 p63(그림4D, 아래)을함유하였다. 이 데이터는 서양 얼룩 또는 공초점 현미경 검사법에 의하여 분석을 위한 organoids를 수확하고 준비하는 접근을 확인하고 그것으로 분화를 포함하여 중요한 세포 프로세스를 공부하는 organoid 분석결과의 기능을 확장합니다.

그림 1: 수집 및 분석을 위한 전립선 오르가노이드를 생성하는 회로도 예시 워크플로우. 총 마우스 전립선은 해리되고 기저 및 발광 전립선 상피 세포는 확립된프로토콜(18,19)을통해 형광 활성화 세포 선별에 의해 단리된다. 마우스 오르가노이드 및 매트릭스 겔의 혼합물에서 현탁된 기저 또는 발광 세포는 매트릭스 겔 링으로 도금된다. 배양의 5 7 일 후에, 유기노이드는 서쪽 얼룩 또는 공초점 현미경 검사법에 의하여 분석을 위해 수확됩니다. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
{kind=link}

도 2: 형광 활성화 세포 선별(FACS)을 사용하여 마우스 기저 및 발광 전립선 상피 세포의 분리. 마우스 전립선에서 해리 된 세포는 DAPI로 염색되어 죽은 세포와 살아있는 것을 구별하고 표면 항체를 구별하여 FACS 이전에 기저와 발광 세포를 구별합니다. 왼쪽 = DAPI- 셀에 게이트. FSC-A = 정방향 분산. 중앙 = 린에 게이트- 세포 (CD45로,CD31로,Ter119로). SSC-A = 측면 분산. 오른쪽 = 기저 세포 (Bas) (EpCAMhi,CD49fhi),루미날 셀 (Lum) (EpCAMhi,CD49f중간). 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
{kind=link}

그림 3: 마우스 전립선 오르가노이드의 확립. (a)24웰 플레이트의 웰에서 매트릭스 겔 링을 생성하는 도식예 접근법. (B)대표적인 위상 대조 이미지 오르가노이드(3D 성장 평면) 및 2차원 콜로니(2D 성장 평면)는 전립선 상피 세포를 코팅되지 않은(Poly-HEMA(-)) 또는 코팅(Poly-HEMA(+)) 24웰 플레이트로 도금한 후 7일 후에 형성되었다. 2D 성장 평면 내의 박스영역은 오른쪽에 확대됩니다. 배율 막대 = 200 μm. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
{kind=link}

그림 4: 웨스턴 블롯 및 전체 마운트 공초점 현미경 검사법에 의한 전립선 오르가노이드의 계보 마커 발현 분석. (A)7일 배양 후 기저 유래(상단) 및 발광 유래(아래) 오르가노이드의 대표적인 위상 대조 이미지. 스케일 바 = 100 μm.(B)배양 5일 후 기저 유래(Bas) 및 발광 유래(Lum) 오르가노이드의 서양 블롯 분석. 기저 마커, 시토케라틴 5(K5) 및 발광 마커, 시토케라틴 8(K8) 및 로딩 제어, 히스톤 H3(HH3)에 대한 염색. (C)스페이서와 챔버 커버 슬립을 보여주는 회로도. (D)대표적인 차등 간섭 콘트라스트(DIC) 및 기저 유래(상부) 및 발광 유래(하단) 오르가노이드의 면역형광 이미지는 7일 배양 후. p63 (빨강), K8 (녹색) 및 DAPI (파란색)에 대 한 염색 을 개별적으로 병합 합니다. 배율 막대 = 100 μm. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
{kind=link}
| 조리법 | |
| 디스파스 함유 미디어 | 고급 DMEM F12에서 1 mg/mL 디스파제 + 10 μM ROCK 억제제. 0.22 μm 필터를 사용하여 필터 살균. |
| 해리 매체 | RPMI 1640에서 10 % FBS + 1 x 페니실린 - 스트렙토 마이신. 0.22 μm 필터를 사용하여 필터 살균. |
| 단백질 라기스 완충제 | RIPA 완충제 + 인산염 억제제 + 프로테아제 억제제 |
| 차단 솔루션 | 0.2% 트리톤 X-100으로 PBS에서 10% FBS |
표 1: 주요 솔루션 준비에 대한 지침입니다.
| 구성 요소 | 농도 |
| B-27 | 1x (50x 농축액에서 희석) |
| 글루타맥스 | 1x (100x 농축액에서 희석) |
| N-아세틸-L-시스테인 | 1.25 mM |
| 노르모신 ("것") | 50 μg/mL |
| 재조합 인간 EGF, 동물 무료 | 50 ng/mL |
| 재조합 인간 노긴 | 100 ng/mL |
| R-스폰딘 1-컨디셔닝 미디어 | 10% 컨디셔닝 된 미디어 |
| A83-01 | 200 nM |
| Dht | 1 nM |
| Y-27632 디하이드로클로라이드 (ROCK 억제제) | 10 μM |
| 고급 DMEM/F-12 | 베이스 미디어 |
| R-spondin 1-컨디셔닝된 매체는 드로스트, 외13에기재된 바와 같이 생성된다. 모든 성분을 첨가한 후 0.22 μm 필터를 사용하여 마우스 오르가노이드 매체를 살균합니다. ROCK 억제제는 오가노이드의 문화 확립 및 통과 중에만 첨가됩니다. | |
표 2: 마우스 오르가노이드 의 제조를 위한 지침.
| 폴리 헤마 코팅 플레이트 를 준비하기위한 프로토콜 | |
| 1 | 50 mL 98 % EtOH에 0.25 g의 폴리 헤마를 추가하십시오. 쉐이커에 37°C에서 폴리 헤마를 용해시다. 이 프로세스는 적어도 4 시간이 걸립니다. |
| 2 | 0.22 μm 필터를 사용하여 폴리 HEMA를 살균하는 필터. |
| 3 | 24웰 플레이트의 웰당 폴리-HEMA 용액 200 μl을 첨가합니다. |
| 4 | 폴리-HEMA를 첨가한 후 24웰 플레이트에서 뚜껑을 제거하고 용액이 하룻밤 사이에 증발할 수 있도록 하십시오. |
| 5 | 각각 잘 PBS로 두 번 씻고 최종 세척 후 보관하기 전에 우물이 완전히 건조되었는지 확인하십시오. 참고: 세척 중 폴리-HEMA 코팅을 방해하면 상피 세포를 유기체 배양으로 도금하면 2차원 성장에 기여할 수 있습니다. 폴리 헤마 코팅 웰의 손상을 방지하려면 세척 하는 동안 파이펫 팁과 직접 접촉 하지 마십시오. 폴리-HEMA 코팅 웰의 무결성은 파이펫 팁에 의해 폴리-HEMA가 긁히지 않는 한 그대로 유지됩니다. |
| 6 | 폴리-HEMA 코팅 플레이트는 최대 2주 동안 4°C에서 보관할 수 있다. 참고: 보관 전에 파라필름에 포장플레이트를 포장하면 오염 위험이 줄어듭니다. |
표 3: 폴리 HEMA 코팅 플레이트의 제조를 위한 프로토콜.
토론
전립선 상피 세포 분화는 정상 전립선 생물학2,3,4,5,6,7 및 질병 생물학8,10,11,12모두에 연루되어 있다. 그러나 이 프로세스의 마스터 레귤레이터는 정의되지 않은 상태로 유지됩니다. 전립선 상피 세포 분화의 주요 조절기를 식별하는 것은 그것을 모델링하기 위하여 잘 확립된 문맥의 부재 때문에 부분적으로 어려웠습니다. 2D 단층 배양은 분화11,12를모델링하는데 사용될 수 있지만, 이러한 맥락은 복잡한 전립선 미세환경을 재화하지 못한다. 또한, 모델 차별화에 생체 내 컨텍스트는 조작하기 어렵기 때문에 기계론적 연구에 자신을 빌려주지 않습니다. 따라서, 조작하기 쉬운의 식별, 아직 생리학적으로 관련된 맥락, 분화를 연구하는 것은 중요하다.
전립선 오르가노이드 모델은 기저에서 발광 분화에 대한 기저체가 발생하는 우아한 생체 내 맥락을 나타낸다. 전립선 오르가노이드를 확립하는 방법은 잘 확립된 14; 그러나 이러한 메서드의 추가 최적화가 필요합니다. 또한 분석을 위해 전립선 오르가노이드를 수확하고 준비하는 접근법은 명확하게 설명되지 않습니다. 이 논문은 마우스 전립선에서 유기체 배양으로 분리된 판상피 세포에 대한 접근법을 설명합니다. 이 접근법은 (1) 오르가노이드 형성 동안 2D 콜로니의 발생을 방지하고, (2) 미디어 보충 동안 매트릭스 젤에 대한 중단의 위험을 감소시키고, (3) 오르가노이드를 보다 효과적으로 계산할 수 있게 한다. 또한,이 원고는 서양 얼룩 분석, 또는 전체 마운트 공초점 현미경 검사법에 대한 준비를위한 오르가노이드를 수확하는 접근 방식을 설명합니다. 중요한 것은, 공초점 현미경 검사법을 위한 오르가노이드를 준비하는 데 이용된 접근은 그것의 기간을 통해 오르가노이드의 손상의 본래 구조를 유지합니다, 이는 심상 취득의 앞에 오르가노이드 손상을 감소시킵니다. 전부, 기술된 접근은 전립선 오르가노이드 분석의 기능을 확장합니다.
특히, 기저 및 발광 세포의 오르가노이드 형성 능력은 각각의 인구를 격리하는 데 사용되는 방법, 및 배양 조건에 의해 모두 변경될 수 있습니다. 이 분석에서 사용된 오르가노이드 배양 조건은 카르타우스 외13에의해 처음 기술되었다. 카르타우스 외. 기저 세포는 더 높은 오르가노이드 형성 능력을 가지고 있다고보고 한 반면 (15%) 발광 세포(1%)13,Chua 등은 뚜렷한 절연 방법 및 배양 조건을 사용하여 발광 세포(0.2-0.3%)가 있다고 보고했습니다. 기저세포(0.03%)보다 오르가노이드 형성 능력이20%더 높다. 전반적으로, Karthaus 등. 에 의해 기술된 방법은 기저 및 발광 세포 둘 다를 위한 더 높은 organoid 형성 비율로 이끌어 내고, 아마 기저 및 발광 세포를 격리하기 위하여 이용된 접근에 있는 다름을반영합니다 13,발광 세포에서 organoid 대형에 대하여 편견하는 배양 조건과는 반대로. 이 원고에 기술된 프로토콜이 다능한 발광 선조또는 헌신적인 발광선조로부터의발광 오르가노이드 형성을 선호하는지 여부는 불분명합니다 9. 시기 와 비용 금지, 생체 내 계보 추적 연구는 유기 성 분석에서 해명 뚜렷한 전립선 상피 계보와 관련된 선조 기능을 검증하는 데 사용할 수 있습니다.
발달, 분화 및 변환과 같은 프로세스는 전립선 생물학과 관련이 없을 뿐만 아니라 뇌, 폐, 내장, 췌장 및 간을 포함한 다른 조직의 생물학과도 관련이 있습니다. 설명된 방법은 전립선뿐만 아니라 광범위한 조직에서 이러한 과정을 연구하기 위해 오르가노이드 모델의 활용을 용이하게합니다.
공개
저자는 공개 할 것이 없다.
감사의 말
PDC와 JMG는 루스 L. 커슈슈타인 국가 연구 서비스 상 GM007185에 의해 지원됩니다. JAD는 T. 하슨과 사울 마르티네즈 장학금에 수여 (R25GM055052)의 국립 국립 연구소의 일반 의학 연구소에 의해 지원됩니다. ASG는 스피처 가족 재단 기금과 길 엔다우먼트의 지원을 받고 있습니다. 이 작품은 미국 암 학회 (RSG-17-068-01-TBG), 국방부 (W81XWH-13-1-0470), 마가렛 E에 의해 지원되었다. 초기 의학 연구 신탁, NIH/NCI (전립선암의 P50CA092131/UCLA SPORE), 로즈 힐스 재단, UCLA의 존슨 종합 암 센터, 브로드 줄기 세포 연구 센터, 임상 및 중개 과학 연구소 및 비뇨기과 종양학 연구소의 지원.
자료
| Name | Company | Catalog Number | Comments |
| µ-Dish 35 mm, high | ibidi | 81156 | |
| 16% Paraformaldehyde | Thermo Fisher Scientific | 50-980-487 | |
| 4’,6-diamidino-2-phenylindole (DAPI) | Thermo Fisher Scientific | D1306 | |
| A83-01 | Tocris | 2939 | |
| Advanced DMEM/F-12 | Thermo Fisher Scientific | 12634010 | |
| APC/Cy7 anti-mouse CD326 (Ep-CAM) Antibody, 100 μg | BioLegend | 118218 | |
| B-27 Supplement (50x), Serum Free | Thermo Fisher Scientific | 17504044 | |
| Complete Protease Inhibitor Cocktail | Sigma | 11836145001 | |
| (DiHydro)testosterone (5α-Androstan-17β-ol-3-one) | Sigma | A-8380 | |
| Dispase II, Powder | Thermo Fisher Scientific | 17-105-041 | |
| Fetal Bovine Serum (FBS) | Sigma | F8667 | |
| FITC anti-mouse CD31 Antibody (0.5 mg/mL, 50 μg) | BioLegend | 102405 | |
| FITC anti-mouse CD45 Antibody (0.5 mg/mL, 50 μg) | BioLegend | 103107 | |
| FITC anti-mouse TER-119/Erythroid Cells Antibody (0.5 mg/mL, 50 μg) | BioLegend | 116205 | |
| Goat anti-mouse IgG-Alexa Fluor 488 | Invitrogen | A28175 | |
| Goat anti-rabbit IgG-Alexa Fluor 594 | Invitrogen | A11012 | |
| GlutaMAX | Thermo Fisher Scientific | 35050061 | |
| Halt Phosphatase Inhibitor | Thermo Fisher Scientific | 78428 | |
| Matrigel GFR Membrane Matrix | Corning | CB-40230C | |
| Mouse anti-cytokeratin 8 | BioLegend | 904804 | |
| N-acetyl-L-cysteine | Sigma | A9165 | |
| Normocin | Thermo Fisher Scientific | ant-nr-1 | |
| PE anti-human/mouse CD49f Antibody | BioLegend | 313612 | |
| Penicillin-Streptomycin (10,000 U/mL) | Thermo Fisher Scientific | 15-140-122 | |
| Poly(2-hydroxyethyl methacrylate) (Poly-HEMA) | Sigma | P3932-25G | |
| Rabbit anti-p63 | BioLegend | 619002 | |
| Radioimmunoprecipitation assay (RIPA) | Thermo Fisher Scientific | PI89901 | |
| Recombinant Human EGF, Animal-Free | PeproTech | AF-100-15 | |
| Recombinant Human Noggin | PeproTech | 120-10C | |
| RPMI 1640 Medium, HEPES (cs of 10) | Thermo Fisher Scientific | 22400105 | |
| Sonic Dismembrator | Thermo Fisher Scientific | FB120 | |
| Sucrose | Sigma | S0389-500G | |
| Triton X-100 | Sigma | X100-5ML | |
| Y-27632 dihydrochloride (ROCK inhibitor) | Selleck Chemical | S1049-50MG |
참고문헌
- Kwon, O. J., Xin, L. Prostate epithelial stem and progenitor cells. American Journal of Clinical and Experimental Urology. 2 (3), 209-218 (2014).
- Choi, N., Zhang, B., Zhang, L., Ittmann, M., Xin, L. Adult Murine Prostate Basal and Luminal Cells Are Self-Sustained Lineages that Can Both Serve as Targets for Prostate Cancer Initiation. Cancer Cell. 21 (2), 253-265 (2012).
- Ousset, M., Van Keymeulen, A., et al. Multipotent and unipotent progenitors contribute to prostate postnatal development. Nature Cell Biology. 14 (11), 1131-1138 (2012).
- Wang, J., et al. Symmetrical and asymmetrical division analysis provides evidence for a hierarchy of prostate epithelial cell lineages. Nature Communications. 5, 1-13 (2014).
- Wang, Z. A., Mitrofanova, A., et al. Lineage analysis of basal epithelial cells reveals their unexpected plasticity and supports a cell-of-origin model for prostate cancer heterogeneity. Nature Cell Biology. 15 (3), 274-283 (2013).
- Kwon, O. J., Zhang, B., Zhang, L., Xin, L. High fat diet promotes prostatic basal-to-luminal differentiation and accelerates initiation of prostate epithelial hyperplasia originated from basal cells. Stem Cell Research. 16 (3), 682-691 (2016).
- Kwon, O. J., Zhang, L., Ittmann, M. M., Xin, L. Prostatic inflammation enhances basal-to-luminal differentiation and accelerates initiation of prostate cancer with a basal cell origin. Proceedings of the National Academy of Sciences. 192 (3), 997-999 (2014).
- Stoyanova, T., et al. Prostate cancer originating in basal cells progresses to adenocarcinoma propagated by luminal-like cells. Proceedings of the National Academy of Sciences. 110 (50), 20111-20116 (2013).
- Agarwal, S., Hynes, P. G., et al. Identification of Different Classes of Luminal Progenitor Cells within Prostate Tumors. Cell Reports. 13 (10), 2147-2158 (2015).
- Ku, S. Y., et al. Rb1 and Trp53 cooperate to suppress prostate cancer lineage plasticity, metastasis, and antiandrogen resistance. Science. 355 (6320), 78-83 (2017).
- Mu, P., et al. SOX2 promotes lineage plasticity and antiandrogen resistance in TP53- and RB1-deficient prostate cancer. Science. 355 (6320), 84-88 (2017).
- Bishop, J. L., et al. The Master Neural Transcription Factor BRN2 Is an Androgen Receptor-Suppressed Driver of Neuroendocrine Differentiation in Prostate Cancer. Cancer Discovery. 7 (1), 54-71 (2016).
- Karthaus, W. R., et al. Identification of multipotent luminal progenitor cells in human prostate organoid cultures. Cell. 159 (1), 163-175 (2014).
- Drost, J., Karthaus, W. R., et al. Organoid culture systems for prostate epithelial and cancer tissue. Nature Protocols. 11 (2), 347-358 (2016).
- McCray, T., Richards, Z., Marsili, J., Prins, G. S., Nonn, L. Handling and Assessment of Human Primary Prostate Organoid Culture. Journal of Visualized Experiments. (143), e59051(2019).
- Lawson, D. A., Xin, L., Lukacs, R. U., Cheng, D., Witte, O. N. Isolation and functional characterization of murine prostate stem cells. Proceedings of the National Academy of Sciences of the United States of America. 104 (1), 181-186 (2007).
- Chen, H. Y., Kaya, K. D., Dong, L., Swaroop, A. Three-dimensional retinal organoids from mouse pluripotent stem cells mimic in vivo development with enhanced stratification and rod photoreceptor differentiation. Molecular vision. 22, 1077-1094 (2016).
- Liu, X., et al. Low CD38 Identifies Progenitor-like Inflammation-Associated Luminal Cells that Can Initiate Human Prostate Cancer and Predict Poor Outcome. Cell Reports. 17 (10), 2596-2606 (2016).
- Lukacs, R. U., Goldstein, A. S., Lawson, D. A., Cheng, D., Witte, O. N. Isolation, cultivation and characterization of adult murine prostate stem cells. Nature protocols. 5 (4), 702-713 (2010).
- Chua, C. W., Shibata, M., et al. Single luminal epithelial progenitors can generate prostate organoids in culture. Nature Cell Biology. 16 (10), 951-961 (2014).
재인쇄 및 허가
JoVE'article의 텍스트 или 그림을 다시 사용하시려면 허가 살펴보기
허가 살펴보기더 많은 기사 탐색
This article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. 판권 소유