
Method Article
Улучшенный анализ и инструменты для измерения механического ноцицепции в Drosophila Larvae
В этой статье
Резюме
Цель этого протокола состоит в том, чтобы показать, как выполнить улучшенный анализ механического ноцицепции в личинках дрозофилы. Мы используем анализ здесь, чтобы продемонстрировать, что механическая гиперчувствительность (алдиния и гипералгезия) существует в личинках дрозофилы.
Аннотация
Опубликованные анализы механического ноцицепции в Дрозофиле привели к переменным оценкам поведения. Здесь мы изготовили для использования личинки дрозофилы, индивидуальные металлические никель-титановые сплавы (нитинол) нити. Эти механические зонды похожи на нити фон Фрея, используемые у позвоночных для измерения механического ноцицепции. Здесь мы демонстрируем, как сделать и откалибровать эти механические зонды и как создать полную поведенческую дозу-ответ от подтресхолдинга (безобидный или не-вредный диапазон) до надтраста (низкий и высокий вредный диапазон) стимулов. Чтобы продемонстрировать полезность зондов, мы исследовали повреждения тканей индуцированной повышенной чувствительности в личинках дрозофилы. Механическая алдиния (гиперчувствительность к обычно безобидному механическому стимулу) и гипералгезия (преувеличенная отзывчивость к вредным механическим стимулом) еще не установлены в личинках дрозофилы. Используя механические зонды, которые обычно безобидны или зонды, которые обычно вызывают аверсивное поведение, мы обнаружили, что личинки дрозофилы развивают механическую гиперсензитизацию (как алдинию, так и гипералгезию) после повреждения тканей. Таким образом, механические зонды и анализы, которые мы здесь иллюстрируют, вероятно, будут важными инструментами для вскрытия фундаментальных молекулярно-генетических механизмов механической гиперчувствительности.
Введение
Личинки дрозофилы обладают характерным аверсивным поведением при воздействии различных вредныхраздражителей: тепловых 1,механических 2ихимических 3. Такое поведение явно отличается от нормального передвижения. Здесь мы описываем улучшенный механический анализ, который может быть использован для оценки механического ноцицепции и механической сенсибилизации.
В недавнем исследовании, мы изготовили фон Фрей-как нити с использованием нитинол провода4. Зонды, прилагающие различные силы и давление, были сделаны путем изменения длины и диаметров нитинол провода формирования каждого зонда. Механические зонды были откалиброваны и измеренные значения силы (в millinewton, mN) были преобразованы в давление (kilopascal, kPa), на основе области наконечника каждогозонда 4. Таможенное изготовление механических зондов позволило нам генерировать субтрестойное (≤200 кПа) до надтраста (225 кПа до 5318 кПа) давления, что в принципе могло бы быть полезно для изучения механической гиперчувствительности. Используя эти улучшенные механические фон Фрей-как нити, мы показали,что давление 4, в отличие от ранееисследованной силы 2,5,6 коррелирует более последовательно с аверсивной поведенческой отзывчивости в личинках дрозофилы. Улучшенный механический анализ, описанный здесь, также помог определить сохраненный сосудистый эндотелиальный фактор роста (VEGF) связанный с рецептором тирозинкиназы, сигнализирующий о пути, который регулирует механическое ноцицепция у мух икрыс 4.
Механическая алдиния и гипералгезия, два условия повышенной чувствительности, относительно недостаточно изучены в личинках дрозофилы, по сравнению с тепловыми (тепло и холод) и химическими сенсорнымимодальности 3,7,8,9,10. Это, вероятно, из-за отсутствия конкретных механических зондов, которые охватывают от безобидных стимуловдо высокого вредных диапазон 2,5,6. Обычно безобидный стимул, который вызывает типичный аверсивное поведение прокатки после drosophila личинки опыт повреждения тканей 3,7 называется allodynia. Преувеличенный подвижной ответ на обычно вредный стимул известен как гипералгезия7. Вредные стимулы определяются как те, которые вызывают повреждение тканей и могут активировать ноцицепторы11. Вредные стимулы доставлены личинки Drosophila повреждения либо барьер эпидермиса, периферических ноцицептивных сенсорныхнейронов 3,4,7, или оба.
В этой статье мы демонстрируем, как на заказ изготавливать и калибровать механические зонды типа фон Фрея, подходящие для личинок дрозофилы. Кроме того, мы показываем, как использовать эти зонды для анализа механических ноцицептивных реакций в личинках дрозофилы. Наконец, мы дополнительно продемонстрировать полезность этих зондов, используя их, чтобы продемонстрировать наличие механической гиперчувствительности, как алдиния и гипералгезия, после повреждения тканей в личинках дрозофилы (см. Представитель Результаты).
протокол
1. Конструкция механического зонда
- Вырезать каждый нитинол нити (Рисунок 1B), перпендикулярно его длинной оси, к указанной длине (Рисунок 1М-N) с помощью небольшой резак провода (Рисунок 1C). Нити бывают трех различных заранее установленных диаметров(рисунок 1B).
ПРИМЕЧАНИЕ: Указанные здесь длины являются руководством для достижения указанного приблизительного давления, используя аналогичный протокол для строительства крепления. В конечном счете, независимо от длины нити вырезать, и глубина отверстия в горе, нити должны быть измерены / откалиброваны на балансе, чтобы получить точное значение силы / давления. - Изучите кончик нити под стереомикроскопом, чтобы убедиться, что острые или нерегулярные края не остаются, поскольку они могут привести к повреждению тканей кожи личинок и помешать калибровке.
- Вручную разгньте острые края механического зонда с помощью заточки камня до тех пор, пока не сохраняются резкие неровности(рисунок 1D).
- Сделайте отверстие к концу деревянной палочки эскимо(рисунок 1E) спомощью подкожной иглы (см. Таблицу материалов). Вставьте иглу по крайней мере на полпути через высоту палочки эскимо(рисунок 1E). Это создает камеру для вставки нити нитинола.
- Нанесите древесный клей на одну нитиноловую нить(рисунок 1F)и вставьте нить с клеем в игольчатый слот в деревянной палочке эскимо(рисунок 1G). Дайте высохнуть в течение 5 ч.
- Калибруйте каждый механический зонд, прижимая его к шкале до тех пор, пока механический зонд неизгибается (рисунок 1H-L). Это точка максимальной силы, которая может быть записана в граммах. В зависимости от диаметров нити (предварительно установленных) и длин (определяемых пользователем) может быть сформирован полный спектр сил и давления.
- Преобразовать массу, записанную в шаге 1.6, чтобы заставить в millinewton (mN) с помощью формулы f и ma (Сила равна массе, умноженной гравитационным ускорением). f: сила; м: масса; a: гравитационное ускорение (9,8 м/с2)(рисунок 1М).
- Наконец, преобразовать рассчитанную силу в давление (сила/область) в килопаскаль (kPa), разделив измеренную силу на площадь поверхности кончика нити(рисунок 1M). Чтобы рассчитать площадь, переобразуйте диаметр различных нитинолных нитей с дюймов (0,04, 0,06" и 0,08") в сантиметры. Затем,2 (где, r и радиус нити нитинола) определяет область (см. Рисунок 1M). Подготовка нескольких зондов с использованием нитей разного диаметра и длины будет генерировать полный набор, охватывающий отзывчивый диапазон для личинок дрозофилы (образец, показанный на рисунке 1N).
ПРИМЕЧАНИЕ: Проверьте каждый механический зонд по крайней мере каждые 3-4 недели. Когда давление отклоняется более чем на ± 3% от первоначальной меры, новый механический зонд должен быть изготовлен.
2. Подготовка личинок
- Поднимите контрольный штамм(w1118) потомство личинок или личинок, содержащих трансгены ppk-Gal4>UAS-mCD8-GFP (для визуализации повреждения сенсорных нейронов) на стандартной пище в инкубаторе 25 градусов по Цельсию. Как правило, запасы обычно поддерживаются при 18 градусов по Цельсию, но оба родителя и личинки потомство растут при 25 градусов по Цельсию на стандартной кукурузной муки пищи для экспериментов.
ПРИМЕЧАНИЕ: Взрослые мухи (пять самцов и десять самок, 1:2 соотношение) хранятся в флаконах мухи, чтобы яйцо откладывая, около 24 ч. Время после откладки яиц (AEL) начинается с того времени, когда взрослые удаляются. - Соберите третью личинку instar, после приблизительно 96 ч откладки яиц, мягко брызг воды из-под крана в мягкую пищу мухи, содержащую личинки. Блуждающие личинки, которые покинули пищу, или которые когда-либо передние или задние spiracles, слишком велики / старые для этого анализа. Вторые личинки instar (длина менее 4 мм) слишком малы.
- Вылейте содержимое мягкой мухи пищи в чистую стандартную чашку Петри (100 мм х 15 мм).
- Используя миппы, сортируют середину третьей instar, среднего размера, личинки (см. Рисунок 2A) от небольших (вторая instar и в начале третьей instar) или больше (поздно или блуждающих третьей instar) личинок. Рекомендуется нежная манипуляция с типсами, чтобы избежать повреждения личинок тканей.
ПРИМЕЧАНИЕ: Передача с использованием типсов основана главным образом на напряжении воды, а не путем оказания давления на личинки с лезвиями типсов. Альтернативой использованию типсов для маневрирования личинок являются мягкие кисти краски. С помощью любого инструмента, пользователь должен практиковать передачу животных, чтобы не вызвать непреднамеренные повреждения тканей, которые могут осложнить поведенческие измерения. - Перенесите личинки третьей звезды, используя типсы, в небольшую чашку Петри (30 мм х 15 мм), содержащую небольшую вилку мухи, увлажненной водой при комнатной температуре. Храните личинки в маленькой чашке Петри до тех пор, пока не будут проведены эксперименты, но не дольше 20 мин.
ПРИМЕЧАНИЕ: Как правило, передача 20-30 личинок в пищевой штепсельной вилки даст достаточное количество в течение 20 минут поведенческих анализов.
3. Механический анализ ноцицепции
- Поместите личинку в середине третьей звезды (с использованием типсов) на тонкую площадку из черного или темного винила под ярким полем стереомикроскопа. Темный цвет обеспечивает контраст, который улучшает визуализацию личинки. Предпочтительно иметь свободно подвижный кусок темного винила, потому что он позволяет пользователю выровнять личинку, не касаясь или повребев ей.
- Положите оптические огни волокна между объективами цели микроскопа и черной или темной виниловой площадкой; это позволит адекватное высоко контрастное освещение для просмотра личинки.
- Откажитесь от личинок, которые не проявляют нормального передвижения после переноса на площадку. Они могут вмешиваться в нормальную ноцицептивную поведенческую реакцию. Для нормального передвижения, см. Видео 1.
- Протрите прочь, используя бумажное полотенце, любой избыток воды вокруг личинки, которые могут привести к личинки плавать на виниловой площадке.
- Ориентируй личинку, перемещая темную виниловую подушечку. Голова/рот личинки должны указывать влево, если вы правша, и наоборот, если вы левша(рисунок 2A-B).
- Применить выбранный механический зонд, как правило, для 1-2 с, на задней спинной стороне личинки примерно в брюшной сегмент A8 (см. рисунок 2B), пока зонд изгибы и вызывает ранее измеренное количество давления (Рисунок 2C). Важно, чтобы зонд прижимается к спинной поверхности личинки и сжимает личинки в основную площадку в точке контакта зонда.
ПРИМЕЧАНИЕ: В точке контакта между кончиком нити нитинола и спинной кутикулы-эпидермиса, зонды ниже 2300 кПа, в основном согнуть без проникновения кутикулы и основных тканей. Такие зонды редко влияют на личиноховуюсмертность 4. При более высоком давлении (>5000 кПа) зонды как согнуть, а иногда и проникнуть кутикулы и лежащих в основе тканей. Прокол личинок ухудшает выживаемость личинок4 и, при наблюдении, эти личинки, как правило, отбрасываются из поведенческого анализа. - Завехать поведенческий ответ для каждой личинки. Положительный ноцицептивный ответ(Видео 2) указывается, если личинка показывает полный рулон 360 "вдоль оси своего тела в пределах 3 с. Другие ответы (попытка поворота, быстрое ползание и шевелить) считаются отрицательными для целей этого анализа.
ПРИМЕЧАНИЕ: Личинки стимулировали с подтюхом механическим стимулом (200 кПа) не вызывают типичный ноцицептивный или подвижного ответа (Видео 3). Некоторые личинки действительно проявляли быструю перемотку вперед или легкие сенсорные реакции, такие как изменения в направлении движения. - Отбросьте личинку и подготовьте следующую для анализа, повторяя шаги 3.1 до 3.7.
- Повторите шаги 3.1-3.7 до достижения желаемого количества личинок (для каждого зонда здесь использовалось от трех до шести наборов n и 10 личинок).
ПРИМЕЧАНИЕ: При использовании механических зондов более низкого давления (174-462 кПа), анализ займет больше времени на личинку. Это потому, что кончик длинных нитей колеблется больше, что делает его труднее тыкать личинки в центре сегмента A8. Практика необходима с этими зондами.
4. Конфокальная микроскопия для оценки нейрональной морфологии
- Поместите личинку (генотипа ppk-Gal4>UAS-mCD8-GFP для обозначения сенсорных нейронов), ранее стимулируемых нитиноловой нитью, в камеру эфиризации внутри банки Coplin, содержащей стакан 10 мл с ватным тампоном, пропитанным 1 мл диэтилового эфира. Пусть личинка сидит в камере в течение 5 минут.
ПРИМЕЧАНИЕ: Подробный протокол для эфиризации представлен в предыдущем исследовании, опубликованном нашей группой12. - Аккуратно промыть личинку из эфирной камеры в небольшую чашку Петри.
- Готовы один слайд микроскопа, два небольших крышки (22 х 22 мм), и один длинный обложки (22 х 54 мм) (см. Таблицу материалов).
- Добавьте небольшие капли эфира: раствор масла (1:5 соотношения этилового эфира к раствору галоуглеродного масла, см. таблицу материалов)к обоим концам горки, затем поместите небольшие крышки поверх маленьких капель. Это расположение создает небольшой пробел пространства, где личинка может поместиться.
ПРИМЕЧАНИЕ: Нажмите на небольшие крышки против слайда микроскопа, пока не будет трудно скольжения. - Добавьте несколько капель эфира: масляный раствор на середине слайда микроскопа, а затем поместите личинку, используя миппы, на центр слайда микроскопа (между небольшими крышками). Убедитесь, что антеропостерная ось личинки параллельна короткой стороне слайда и что спинная сторона обращена вверх.
- Обложка личинок с длинной крышкой размещены на верхней части личинки и два небольших coverslips.
ПРИМЕЧАНИЕ: Щедро нажмите длинный обложки, пока личинка почти плоская. - Сегмент изображений A8 личинки с помощью конфокального микроскопа (см. Таблицу материалов)с использованием лазерной длины волны 488 (GFP).
ПРИМЕЧАНИЕ: Изображение личинки сразу же, потому что анестезия через эфир будет исчезать быстро (5-10 мин) и личинка проснется и двигаться, что осложнит дальнейшее изображение. - Захват изображения с разрешением 1024 х 1024 пикселей с использованием 20-х числовой диафрагмы (NA) 0,7 сухой объектив цели в 1x зум, размер шага 1,5 мкм.
5. Количественная оценка повреждений тканей
- Соберите и преобразуйте изображения стека серии «З» из раздела 4.8 в единую проекцию (уплощение нескольких изображений, сделанных на разных координационных плоскостях, в одно композитное изображение). Это может быть выполнено с использованием коммерчески доступного программного обеспечения (например, Olympus Fluoview) или любой эквивалентной платформы с открытым исходным кодом, например, Fiji/Image J. Сохранить единую проекцию в формате TIFF.
- Откройте программу анализа изображений Fiji/ImageJ.
- Нажмите на файл, из бара меню, и выберите Открыть из окна, которое отображается.
- Выберите сохраненную единую проекцию изображения, сохраненную в формате TIFF, для анализа.
- Нажмите на Edit, из бара меню, и выберите опцию Инверт из окна, которое отображается.
- Нажмите на изображение, из бара меню, затем выберите Настроить, из окна, которое отображается, и, наконец, выбрать яркость / контрастность вариант.
- Выберите опцию Freehand Shape из бара инструментов для измерения области зазора (если таковые имеются).
- Нажмите на Analyzeиз бара меню и выберите опцию «Мера». Это будет отображать область зазора или раны.
Результаты
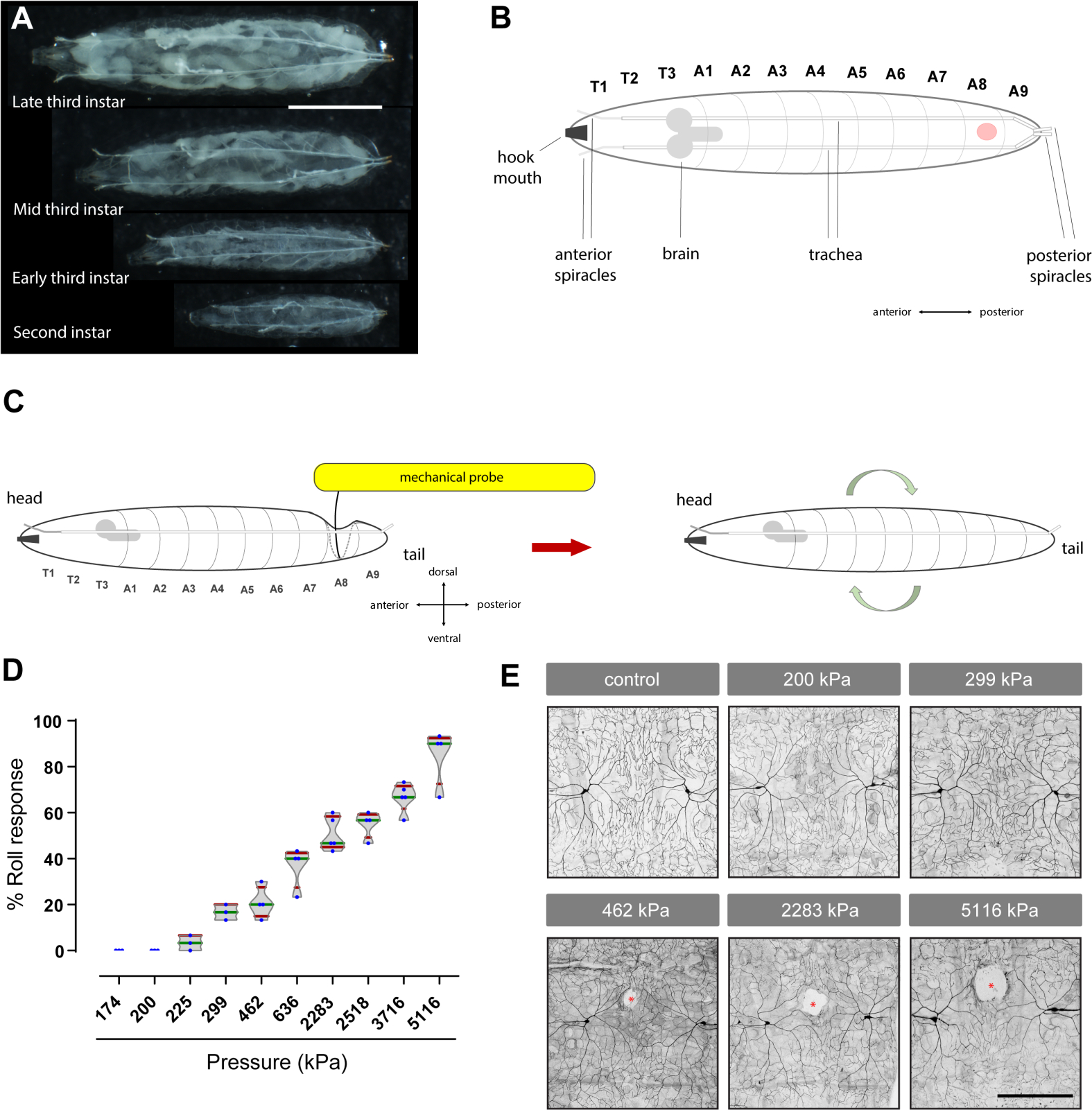
Мы разработали индивидуальные механические зонды, используя нитиноловые нити (Рисунок 1A,N), чтобы вызвать механически-вызванных поведения и генерируется полная поведенческая кривая реакции дозы с использованием как безобидные и вредные механические зондыразличной интенсивности ( Рисунок 2D), демонстрируя, что эти зонды могут быть использованы для изучения базовых (при отсутствии травм) механическое ноцицепция.
Результаты нашего поведенческого анализа показали, что зонды, оказывающие давление ниже 200 кПа (1,57 мН)(рисунок 1М),при применении к личинкам дрозофилы, не провоцируют обратный скользящий ответ(рисунок 2D и видео 3). Как и ожидалось, эти подтресхолдинговые или незнусовые механические зонды (175 кПа или 200 кПа) не вызвали видимых повреждений нейрональнойткани (рисунок 2E). Поскольку они не вызывают повреждений, такие зонды могут быть полезны для оценки механической алдынии (гиперчувствительность к обычно незнусным механическим стимулам). И наоборот, надтресхолдовые или вредные зонды (от 462 кПа до 5116 кПа) вызвали увеличенную поведенческуюреакцию (рисунок 2D) в дозе зависимойманере – с более высоким давлением, вызывая более сильные поведенческие реакции. Как и ожидалось, suprathreshold механическое давление также индуцированных доза-зависимых тканей повреждения периферических сенсорных нейронов себя(рисунок 2E). Измеренная область повреждения тканей (в2 м 2 ± стандартное отклонение), взятая из четырех личинок для каждой группы, составила: 2051,03 ± 703,81 (462 кПа), 5102,2. 29 ± 1 004,67 (2 283 кПа) и 12 238,83 ± 3724,11 (5116 кПа). Таким образом, давление больше или равно 462 кПа (63 мН), которые вызывают аверсивный скользящий ответ (в 25% или более личинок) и вызывают видимые повреждениянейрональной ткани ( рисунок 2E), может быть целесообразным для изучения механической гипералгезии (гиперчувствительность к обычно вредных механических стимулов). Ноцицептивные механические зонды (≥462 кПа) всегда вызывают повреждение тканей (n No 10, оцениваются качественно), но не всегда провоцируют обратную реакцию подвижного состава.
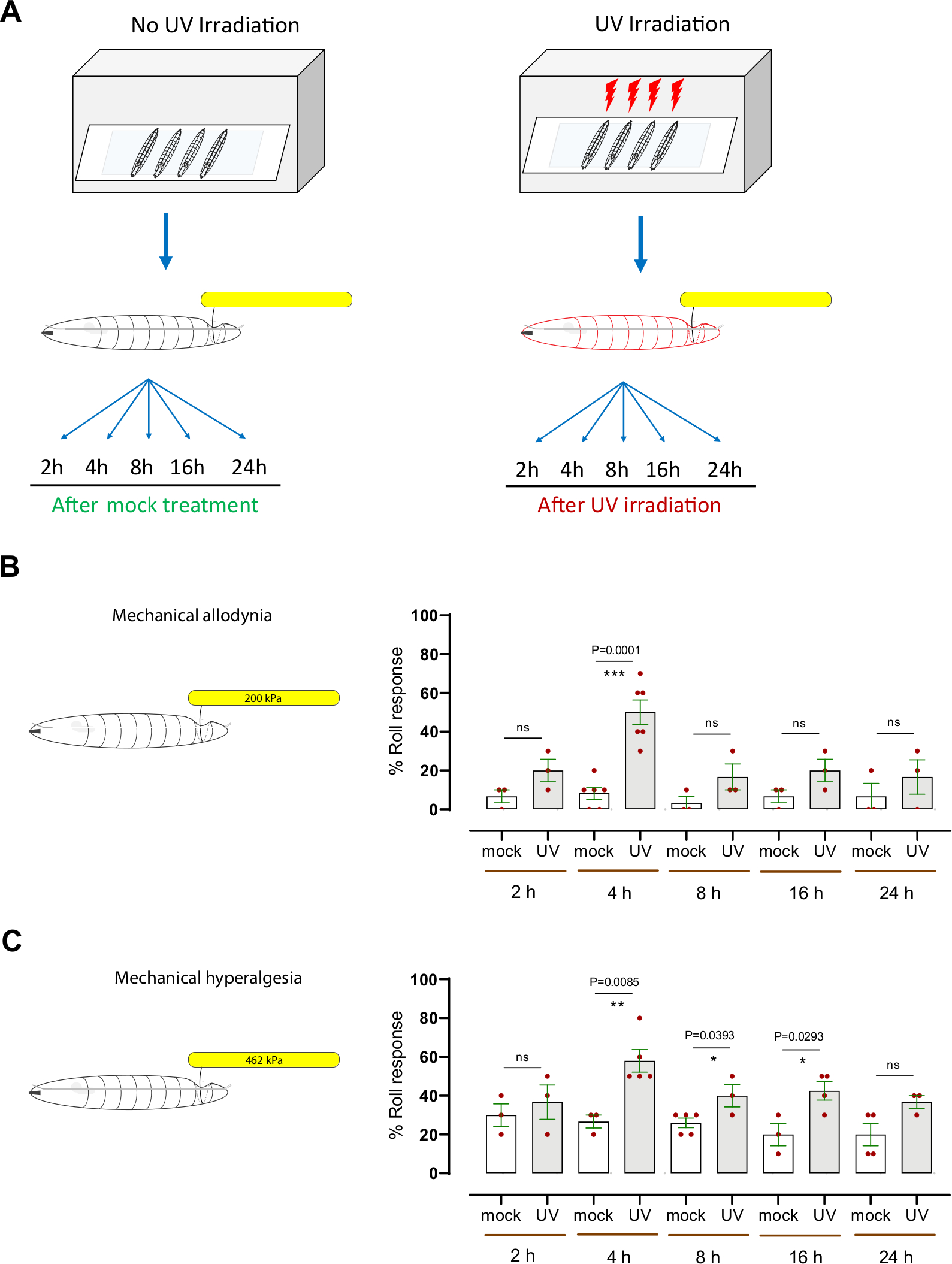
Для оценки механической гиперчувствительности (алдиния и гипералгезия), мы использовали устоявшуюся модель личинки Drosophila ноцицептивной сенсибилизации, которая использует ультрафиолетовое излучение (УФ) для индуцированияповреждения тканей 7,12. Этот анализ помог вскрыть генетические и клеточные механизмы тепловой ноцицептивнойгиперчувствительности 8,9,10,13,14,15. Чтобы определить, вызывает ли УФ-обработка механическую алодинию, личинкитретьейзвезды (w1118)были облучены или облучены УФ(15-20 мДж/см2)(рисунок 3A). Затем личинки были протестированы поведенчески на 2 ч, 4 ч, 8 ч, 16 ч и 24 ч после обработки с обычно подтресхолд механическим зондом (200 кПа, 1,57 мН). Приблизительно 20% личинок ответили уже через 2 ч после УФ-лечения, в то время как 50% ответили на 4 ч, по сравнению с 6,6% и 8,3% макет УФ-облученных животных, соответственно (Рисунок 3B). Это указывает на то, что уф-индуцированной повреждения тканей вызывает механическую алодинию на 4 ч после облучения. В более поздние моменты времени (8 ч, 16 ч и 24 ч) поведенческая реакция УФ-обработанных личинок была в диапазоне от 16%до 20% респондентов (среднее значение n 3-6 наборов по 10 личинок каждая), немного увеличилась (но не статистически значимой) по сравнению с макет-облученной контрольной группой (в диапазоне 3%-6% ответчиков, среднее значение n 3-6 наборов по 10 личинок каждая)(рисунок 3B).
Для исследования механической гипералгезии было использовано давление надтресхолда (462 кПа, 3,63 мН), которое обычно вызывает аверсивную реакцию подвижного состава в 20%личинок (рисунок 2D)и вызывает повреждениенейронной ткани (рисунок 2E). Мы применили 462 кПа зонд на спинной стороне личинок с или без УФ-индуцированной повреждения тканей(рисунок 3A). Мы обнаружили, что личинки, исследованные на 4 ч, 8 ч и 16 ч после УФ-лечения показали значительное увеличение обратной реакции прокатки, с 4 ч является пиком поведенческой гиперчувствительности (60% реагировать); макет УФ-облученных животных показал 27% аверсивного ответа(рисунок 3C). Как и в случае механической ародинии, поведенческая реакция на 8 ч, 16 ч и 24 ч УФ-обработанных животных (в диапазоне 36%-42%) статистически неотличима от необработавных личинок (в диапазоне от 20%до 26%). Larvae на конце третьего этапа instar действительно показывают небольшое снижение базовой поведенческой реакции по сравнению со средней третьей стадией instar. Мы предполагаем, что это может быть либо увеличение размера личинок(рисунок 2A) или увеличение толщины кутикулы, покрывающей тело. Этот факт может объяснить, почему на более позднем этапе развития УФ-обработка не вызывает большей механической сенсибилизации, как наблюдается 4 ч после УФ-лечения.
В совокупности наши результаты показывают, что личинки дрозофилы развивают как механическую алодинию, так и механическую гипералгезию после повреждения тканей, вызванных УФ-излучением. Пиковое время механической алодинии и гипералгезии такое же, 4 ч после УФ-лечения; однако, механическая гипералгезия имеет более выраженный височный хвост, поскольку она возвращается к исходной линии медленнее по сравнению с механической алодинии.

Рисунок 1: Разработка инструмента, похожего на фон Фрея, для оценки механического ноцицепции личинок дрозофилы. (A) Изображение механического зонда, используемого для изучения механического ноцицепции в личинках дрозофилы. (B)Нити нитинола и их относительные диаметры показаны в относительном масштабе. (C)Изображение резака диагональной проволоки, используемого для разрезания нитинолных нитей. (D)Сглаживание острых краев вырезанной нити нитинола с заточками камня. (E) Гиподермическая игла используется, чтобы сделать отверстие в деревянной ручкой палки эскимо зонда. Кончик иглы должен достичь по крайней мере половины высоты ручки палкой для безопасной вставки нити. (F-G) Прикрепление нити нитинола путем склеивания в деревянную ручку палочки эскимо с отверстием вставки. (H-L) Калибровка механических зондов путем нажатия их на шкалу. (M) Значения силы (в mN) и давления (в kPa), генерируемые различными механическими зондами. Длина каждой нити нитинола, используемой для построения зондов (P1-P10; P: зонд) подробно описан в сантиметрах (см). (N)Изображение полного набора механических зондов, от 174 кПа до 5116 кПа. Пожалуйста, нажмите здесь, чтобы просмотреть большую версию этой цифры.
{kind=link}

Рисунок 2: Механический анализ ноцицепции: фон Фрей-как нити генерировать дозу реакции кривой аверсивного поведения прокатки и вызвать повреждение тканей сенсорных нейронов. (A) Фотографии различных стадий (второй и третий instar) личинок Drosophila. Шкала бар: 2 мм. (B) Мультфильм спинного зрения третьей instar личинки Drosophila. Красная точка указывает на брюшной сегмент, где применяется механический зонд. Т: грудной сегмент; A: брюшной сегмент. Другие анатомические ориентиры помечены. (C) Мультфильм анализа: механический зонд применяется к спинной стороне личинки, пока он не изгибается против поверхности ниже, а затем проводится в течение 2 с. Если давление достаточно высокое, это вызывает обратный скользящий ответ при освобождении. (D)Реакция поведенческой дозы; каждая синяя точка представляет собой процент личинок, которые ответили, с обратным прокатки, к механической стимуляции в набор из 10 животных. Скрипка участок процента аверсивного подвижного поведения, вызванного различными механическими зондами. kPa: килопаскаль. Участки коробок представляют медианы (зеленые), усы (красные) представляют собой 10-й и 90-й процентили. (E)Повреждение тканей: Третий instar личинок (генотипа ppk-Gal4>UAS-mCD8-GFP для обозначения ноцицептивных сенсорных нейронов) были исследованы на спинном сегменте A8 с указанным давлением. Затем были исследованы флуоресцентно помеченные парные сенсорные нейроны класса ddaC IV (через спинную середину) (см. разделы 4 и 5). Белые области (красные звездочки) представляют собой зазоры или повреждения тканей. Шкала бар: 100 мкм. В панели B личинка отображается в спинной вид, в то время как в C это боковой вид. Механические зонды, прижатые к спинной стороне кутикулы-эпидермиса личинки, производят депрессию, похожую на карман в точке соприкосновения кончика зонда и прилегающих районов. Твердая черная линия, изогнутая к брюшной стороне, находится в верхней части кармана, в то время как пунктирной серой боковой линией представляет собой боковую сторону и нижнюю часть кармана. Пожалуйста, нажмите здесь, чтобы просмотреть большую версию этой цифры.
{kind=link}

Рисунок 3: Механическая повышенная чувствительность после уф-повреждений. (A)Схема экспериментального дизайна для проверки сенсибилизации. Середина третьей instar были макет лечение (не УФ) или УФ облученных. Механический анализ ноцицепции был затем выполнен в разных точках времени (2 ч, 4 ч, 8 ч, 16 ч и 24 ч) после инсценировки лечения или облучения. (B) Механическая алодиния: Процент личинок, экспонирующих обратный прокат после зондирования с обычно субтрестойкой или незнусным механическим стимулом (200 кПа, 1,57 мН) в указанные моменты времени после макетного лечения или УФ-облучения. (C)Механическая гипералгезия: Процент личинок, экспонирующих обратный прокат после зондирования с обычно надтресхолдом или вредным механическим стимулом (462 кПа, 3,63 мН) в указанные моменты времени после макетной обработки или УФ-облучения. Бары ошибок указывают на среднее количество q/- SEM. Для статистического анализа использовалсядвуххвостый неспареный т-тест: < 0,05 евро, < 0,01; ns: не является значительным. Каждая красная точка, в панелях B и C, представляет собой среднее пропорции 10 личинок, n 3-6 наборов на точку времени /условие. Пожалуйста, нажмите здесь, чтобы просмотреть большую версию этой цифры.
{kind=link}
Видео 1: Нормальное движение личинок дрозофилы. Пожалуйста, нажмите здесь, чтобы загрузить это видео.
Видео 2: Незвука механическая стимуляция личинок дрозофилы. Пожалуйста, нажмите здесь, чтобы загрузить это видео.
Видео 3: Подтресурс механической стимуляции личинок дрозофилы. Пожалуйста, нажмите здесь, чтобы загрузить это видео.
Обсуждение
Мы модифицировали установленный механическийанализ 1,2,16 сиспользованием индивидуальных механических зондов, изготовленных из нитинолных нитей. Этот металлический сплав позволяет нам использовать нити меньшего диаметра, соответствующие размеру личинок дрозофилы. Рыболовные линии на основе монофиламентов доминировали в области мухи механического ноцицепции на сегодняшний день2,5,6,16. Наши нити нитинола поддерживают свою форму и измеряют давление в течение примерно 3-5 месяцев (по нашему опыту). Изменяя длину и диаметр нитинол нитей, пользователь может генерировать широкий спектр давлений, охватывающих от подтресхолдинга до почти полного подвижного ответа. В частности, сделать подтресхолдинг зондов проще с меньшим диаметром нитинол нити. Используя эти зонды, мы обнаружили, что давление, а не сила, вызывает более последовательные ноцифенсивные поведенческие реакции4. Мы демонстрируем здесь, используя устоявшуюся УФ-индуцированной ноцицептивноймодели сенсибилизации 7,10,13, что эти нити также являются полезным инструментом для изучения механической гиперчувствительности-алдодиния и гипералгезия.
Предыдущие исследования с использованием механических зондов, изготовленных из лески, привели к определеннойизменчивости в поведенческой отзывчивости 2,6,16,17. Это может быть причиной нескольких факторов. Во-первых, поскольку давление является важной переменной, полировка кончика нити так, что она округляется и не имеет каких-либо острых краев имеет решающее значение. Во-вторых, отчетность значения давления, а не только силы имеет важное значение для воспроизводимости экспериментов, потому что различные механические зонды, которые генерируют аналогичные силы могут вызвать разрозненноедавление 4. В-третьих, очень важно применять только одну механическую стимуляцию на личинку с помощью вредных зондов, потому что такие зонды производят дозозависимыеповреждения тканей на эпидермальных 4 и сенсорных нейрональных уровнях(рисунок 2E). Второй или последующий вредный механический стимул, после повреждения тканей был вызван, может предположительно ухудшить функцию пострадавших периферических сенсорных нейронов и вызвать измененную поведенческую реакцию. В другом исследовании личинки стимулировали дважды с вредными механическими зондами в основномотображается повышенная поведенческая реакция 5, предполагая развитие острой механической сенсибилизации (гипералгезия), которые могут возникнуть в результате повреждения тканей, спровоцированных первым вредным механическим стимулом. И наоборот, другиеавторы 6 сообщили смешанные (увеличение или снижение) поведенческой реакции, указывая, что измененная поведенческая реакция может быть связано с повреждением / дисфункции нейронной ткани. Стимулирование каждой личинки только один раз устраняет возможную дисперсию в поведенческих реакций в результате сенсибилизации или повреждения тканей. В-четвертых, мы механически стимулировали сегмент A8, который является более задним, чем предыдущие исследования (предпочтительные области A3-A4)2,5,16. Зонды между 3900 кПа и 5300 кПа применяется либо сегмент A2 или A8 не показывают каких-либо поведенческих различий4. Кроме того, A8, по сравнению с A2-A4, легче стимулировать с помощью механических зондов, которые генерируют более низкое давление (<300 кПа), потому что личинка тоньше в этой области и, следовательно, легче сжимается. Другие исследования показали, что вредные механической стимуляции задней части личинки (поставляется жесткой булавкой насекомых, состоявшейся с типсами) в основном вызвали вперед передвижения, а не обратный или подвижногоответа 18. Эта различная поведенческая реакция может быть вызвана различиями в свойствах использованных материалов (сгибаемая нитиноловая нитья против неудержимого насекомого штифта) или различным давлением, поставляемым личинкам (значение давления булавки насекомого не сообщалось).
Развитие механического анализа ноцицепции для личинок дрозофилы позволило области обнаружить, что различные механические сенсорные ионные каналы и нейронные цепиопоясыв механическое ноцицепцию 5,6,16,17. Однако изучение механической гиперчувствительности (алдения и гипералгезия) запаздывание, по сравнению с сенсибилизацией других сенсорных условий –тепла 7,8,10,13,14,холода9ихимической 3. Это отставание может быть отчасти связано с отсутствием подходящих механических зондов, которые могут генерировать полный диапазон реакции, охватывающий подтяза к надтрастанию давления. Особое значение, особенно для оценки механической алодинии, имеют подпорные зонды, которые не вызывают аверсивного подвижного ответа от невредимых личинок. Значение наших усовершенствованных механических зондов заключается в том, что они могут быть изготовлены, чтобы охватить безобидные стимулы (субтресхолд 174 кПа-200 кПа) или низкий и высокий вредный диапазон (надтресхолд 225 кПа до 5116 кПа). Здесь мы демонстрируем с помощью нитинола фон Фрей, как нити, что личинки дрозофилы развивать как механические алодинии и механической гипералгезии после УФ облучения. Механическая сенсибилизация показывает некоторые различия по сравнению с тепловой сенсибилизацией. Как начало, так и пик механической сенсибилизации приходится на более ранние (4 ч) по сравнению с тепловой (тепловой) сенсибилизацией (8 ч для гипералгезии и 24 ч для алодинии)7. Кроме того, механическая алодиния и гипералгезия являются сопутствующими (оба пика на 4 ч). Кроме того, в то время как теплочувствительность (алдиния и гипералгезия) разрешает полностью в болеепоздние моменты времени 7, механическая гиперчувствительность выставлены длинный хвост, который остался немного выше базового уровня. Холодная сенсибилизация в Дрозофиле включает в себя переключательв холодном поведении 9 и появление новых холодных вызванных поведения- явление, которое не наблюдается с механической стимуляции. Эти различия в начале, продолжительности и наблюдаемом поведении свидетельствуют о том, что каждый сенсорный модальности может контролироваться различными сигнальными путями. Сочетание анализа сенсибилизации, описанного здесь, с мощными генетическими инструментами, доступными в Дрозофиле, должно позволить точное генетическое вскрытие наблюдаемой механической гиперчувствительности (алдения и гипералгезия).
Раскрытие информации
Авторов нечего раскрывать.
Благодарности
Мы благодарим Томаса Вана за разработку прототипа нитей фон Фрея, Патрика Дж. Хуанга за улучшение механического анализа зондов, Центра запасов Bloomington Drosophila Stock Center для управления(w1118) и ppk-Gal4>UAS-mCD8-GFP, а также сотрудников лаборатории Галко за критическое прочтение рукописи. Эта работа была поддержана R21NS087360 и R35GM126929 в MJG.
Материалы
| Name | Company | Catalog Number | Comments |
| Beaker | Fisher Scientific | 02-540C | Beaker of 10 ml of capacity. Any similar container will do. |
| Black (Arkansas) bench stone | Dan’s Whetstone | SKU: I200306B24b-HQ-BAB-622-C | Used to smoothe any irregularities of the nitinol wire tips. https://www.danswhetstone.com/product/special-extra-wide-black-bench-stone-6-x-2-1-2-x-1-2/ |
| Confocal microscope | Olympus | FV1000 | Any equivalent confocal microscope will do |
| Coplin Jar | Fisher Scientific | 08-816 | https://www.fishersci.com/shop/products/fisherbrand-glass-staining-dishes-10-slides-screw-cap/08816#?keyword=08-816 |
| Diethyl ether | Fisher Scientific | E138-500 | For anesthetizing larvae. |
| Etherization chamber | This is a homemade customized chamber. Please see details of its construction in our previous published paper12. The purpose of the etherization chamber is allow entry of diethyl ether fumes but prevent larval escape. | ||
| Fiber Optic Light Guide | Schott AG | A08575 | Schott Dual Gooseneck 23 inch |
| Forceps | Fine Science Tool | FS-1670 | For transferring larvae |
| Glue | Aleene's | N/A | Aleene's® Wood Glue, formerly called (Aleene's All-Purpose Wood Glue) https://www.aleenes.com/aleenes-wood-glue |
| Graspable holder | Loew Cornell | N/A | Loew-Cornell Simply Art Wood Colored Craft Sticks, 500 pieces. |
| Halocarbon oil 700 | Sigma | H8898-100ML | |
| Hypodermic needle 30G 1/2"L | Fisher Scientific | NC1471286 | BD Precisionglide® syringe needles, gauge 30, L 1/2 inches. Used to make a hole into the wooden holder for the nitinol wires |
| Large Petridish | Falcon | 351007 | 60 mm x 10 mm Polystyrene Petridish |
| Microscope (Zeiss) Stemi 2000 | Carl Zeiss, Inc. | NT55-605 | Any equivalent microscope will do |
| Microscope Cover Glass 22x22 | Fisher | 12-545-B | |
| Microscope Cover Glass 22x40 | Corning | 2980-224 | Tickness 1 1/2 |
| Microscope Slides | Globe Scientific Inc. | 1358Y | |
| Mini Diagonal Cutter | Fisher Scientific | S43981 | For cutting nitinol filaments |
| Nitinol filaments, Diameters: 0.004”, 0.006”, 0.008” | Mailin Co | N/A | Fifteen pieces of each diameter of 12” length were ordered. https://malinco.com/ |
| Piece of black vinyl | Office Depot | N/A | We use a small piece of vinyl cut from a binder. Dark color provides contrast. A small piece allows orientation of the larva |
| Small Petridish | Falcon | 351008 | 35 mm x 10 mm Polystyrene Petridish |
| Spatula | Fisher Scientific | 21-401-10 | Double-Ended Micro-Tapered Stainless Steel Spatula. Used to place the food in the petri dish |
| Wipes | Fisher Scientific | 06-666A | Kimpes KMTECH, Science Brand. Used to dry larvae of excess moisture. |
| W1118 | Bloomington Drosophila Stock Center | 3605 | Control strain for behavioral assays |
| ppk-Gal4>UAS-mCD8-GFP | Bloomington Drosophila Stock Center | 8749 | Strain for fluorescent labeling of class IV md neurons |
Ссылки
- Tracey, W. D., Wilson, R. I., Laurent, G., Benzer, S. Painless, a Drosophila gene essential for nociception. Cell. 113 (2), 261-273 (2003).
- Zhong, L., Hwang, R. Y., Tracey, W. D. Pickpocket is a DEG/ENaC protein required for mechanical nociception in Drosophila larvae. Current Biology. 20 (5), 429-434 (2010).
- Lopez-Bellido, R., Himmel, N. J., Gutstein, H. B., Cox, D. N., Galko, M. J. An assay for chemical nociception in Drosophila larvae. Philosophical Transactions of the Royal Society of London. Series B: Biological Sciences. 374 (1785), 20190282(2019).
- Lopez-Bellido, R., et al. Growth factor signaling regulates mechanical nociception in flies and vertebrates. Journal of Neuroscience. 39 (30), 6012-6030 (2019).
- Hu, C., et al. Sensory integration and neuromodulatory feedback facilitate Drosophila mechanonociceptive behavior. Nature Neuroscience. 20 (8), 1085-1095 (2017).
- Kim, S. E., Coste, B., Chadha, A., Cook, B., Patapoutian, A. The role of Drosophila Piezo in mechanical nociception. Nature. 483 (7388), 209-212 (2012).
- Babcock, D. T., Landry, C., Galko, M. J. Cytokine signaling mediates UV-induced nociceptive sensitization in Drosophila larvae. Current Biology. 19 (10), 799-806 (2009).
- Babcock, D. T., et al. Hedgehog signaling regulates nociceptive sensitization. Current Biology. 21 (18), 1525-1533 (2011).
- Turner, H. N., Patel, A. A., Cox, D. N., Galko, M. J. Injury-induced cold sensitization in Drosophila larvae involves behavioral shifts that require the TRP channel Brv1. PloS One. 13 (12), 0209577(2018).
- Im, S. H., et al. Tachykinin acts upstream of autocrine Hedgehog signaling during nociceptive sensitization in Drosophila. eLife. 4, 10735(2015).
- Cervero, F., Merskey, H. What is a noxious stimulus. Pain Forum. 5 (3), 157-161 (1996).
- Chattopadhyay, A., Gilstrap, A. V., Galko, M. J. Local and global methods of assessing thermal nociception in Drosophila larvae. Journal of Visualized Experiments: JoVE. (63), e3837(2012).
- Follansbee, T. L., et al. Drosophila nociceptive sensitization requires BMP signaling via the canonical SMAD pathway. Journal of Neuroscience. 37 (35), 8524-8533 (2017).
- Im, S. H., Patel, A. A., Cox, D. N., Galko, M. J. Drosophila insulin receptor regulates the persistence of injury-induced nociceptive sensitization. Disease Models & Mechanisms. 11 (5), (2018).
- Jo, J., et al. Drosophila caspase activity is required independently of apoptosis to produce active TNF/Eiger during nociceptive sensitization. Cell Death & Disease. 8 (5), 2786(2017).
- Hwang, R. Y., et al. Nociceptive neurons protect Drosophila larvae from parasitoid wasps. Current Biology. 17 (24), 2105-2116 (2007).
- Guo, Y., Wang, Y., Wang, Q., Wang, Z. The role of PPK26 in Drosophila larval mechanical nociception. Cell Reports. 9 (4), 1183-1190 (2014).
- Takagi, S., et al. Divergent connectivity of homologous command-like neurons mediates segment-specific touch responses in Drosophila. Neuron. 96 (6), 1373-1387 (2017).
Перепечатки и разрешения
Запросить разрешение на использование текста или рисунков этого JoVE статьи
Запросить разрешениеСмотреть дополнительные статьи
This article has been published
Video Coming Soon
Авторские права © 2025 MyJoVE Corporation. Все права защищены