
Method Article
Получение пятерки с 3C: захват хромосомной конформации для студентов
В этой статье
Резюме
Здесь мы подробно представляем адаптацию метода захвата конформации хромосом (3C) с акцентом на участие и обучение студентов.
Аннотация
Захват конформации хромосом (3C) является мощным инструментом, который породил семейство подобных методов (например, Hi-C, 4C и 5C, называемых здесь методами 3C), которые предоставляют подробную информацию о трехмерной организации хроматина. Методы 3C использовались в широком спектре исследований, от мониторинга изменений в организации хроматина в раковых клетках до выявления контактов энхансеров с промоторами генов. В то время как многие исследования, использующие эти методы, задают большие вопросы по всему геному со сложными типами образцов (например, анализ отдельных клеток), что часто упускается из виду, так это то, что методы 3C основаны на основных методах молекулярной биологии, которые применимы к широкому кругу исследований. Решая узконаправленные вопросы организации хроматина, этот передовой метод может быть использован для улучшения исследовательской и преподавательской лабораторной работы студентов. В этом документе представлен протокол 3C и представлены адаптации и акценты для реализации в первую очередь в учебных заведениях бакалавриата в области исследований и преподавания в бакалавриате.
Введение
Геном организма содержит не только все гены, необходимые для функционирования, но и все инструкции о том, как и когда их использовать. Это делает регулирование доступа к геному одной из важнейших функций клетки. Существует множество механизмов контроля функции генов; однако на базовом уровне регуляция генов сводится к способности регуляторных транскрипционных факторов (транс-факторов) связываться с их специфическими последовательностями ДНК (цис-регуляторные последовательности). Это не врожденная способность; Вместо этого он управляется организацией/структурой генома в ядре, которая контролирует доступность/воздействие цис-регуляторных последовательностей на транс-факторы 1,2,3. Если транс-факторы не могут найти свои цис-регуляторные последовательности, то транс-факторы не могут выполнять свои регуляторные задачи. Это сделало понимание того, как геномы организованы в ядре, важным источником исследований.
Широко признано, что во время интерфазы эукариотические хромосомы в ядре занимают свой собственный домен, прикрепленный к ядерной пластинке и ядерному матриксу (рис. 1), что делает хромосому больше похожей на кусок пиццы, а не на лапшу на тарелке спагетти. Хромосомы частично конденсируются белок-ДНК-взаимодействиями (хроматин), которые скручивают и петляют части хромосомы. С помощью электронной микроскопии, трехмерной флуоресцентной гибридизации ДНК in situ (FISH) и методов мечения ДНК (т.е. флуоресцентного и искусственного метилирования ДНК) было обнаружено, что неактивные домены хроматина плотно упакованы вдоль ядерной периферии 4,5,6, в то время как части активного, менее конденсированного хроматина находятся внутри ядра 7,8,9, Статья 10. Эти эксперименты обеспечивают широкоугольное представление о динамике хромосом, но мало что делают для фиксации изменений, происходящих локально вокруг генных промоторов, наблюдаемых в исследованиях ДНКазы11,12 и нуклеосомы13,14,15.
Ключом к раскрытию динамики хроматина с более высоким разрешением стала разработка метода 3D-картирования хромосом 3C. Сам метод 3С состоит из четырех основных этапов: сшивание хроматина, расщепление хроматина рестрикционными ферментами, лигирование хроматина и очистка ДНК (рис. 2). Новые искусственные фрагменты ДНК, генерируемые этим процессом, могут быть охарактеризованы, чтобы выявить тесную физическую связь между линейно удаленными частями ДНК16. Метод 3С стал основой для создания нескольких побочных методов, которые используют начальные этапы 3С, чтобы задавать более широкие вопросы по всему геному (например, Hi-C, 4C, ChIP-C). Это семейство методов 3C определило, что хромосомы организованы в несколько дискретных единиц, называемых топологически ассоциированными доменами (TAD). TAD кодируются в геноме и определяются хроматиновыми петлями, окруженными незакольцованными границами16,17,18,19. Границы TAD поддерживаются двумя эволюционно консервативными и повсеместно распространенными факторами, включая фактор связывания CCCT (CTCF) и когезия, которые предотвращают взаимодействие петель внутри отдельных TAD16,20. Петли опосредованы взаимодействием трансфакторов с их регуляторными последовательностями, а также CTCF и когезией21.
Хотя многие исследования с использованием технологий 3C задают широкие вопросы по всему геному и используют сложные методы сбора образцов, формулировка метода 3C основана на основных методах молекулярной биологии. Это делает 3C интригующим для развертывания как в исследовательских, так и в учебных лабораториях. Метод 3C может быть использован для небольших сфокусированных вопросов и по своей сути гибок для увеличения или уменьшения масштаба (отдельные гены22, хромосомы16 и / или геномы18) в зависимости от направленности и направления задаваемых вопросов. Этот метод также был применен к широкому спектру модельных систем 7,16,19,23 и доказал свою универсальность в использовании. Это делает 3C отличным методом для студентов, поскольку студенты могут получить опыт работы с общими методами молекулярной биологии, а также получить ценный опыт в ответах на направленные вопросы.
Здесь представлен адаптированный протокол для подготовки библиотеки 3С, основанный на ранее опубликованных протоколах24,25,26,27. Этот протокол был оптимизирован примерно для 1 × 107 ячеек, хотя он генерировал библиотеки 3C всего с 1 × 105 ячеек. Этот протокол оказался универсальным и использовался для создания библиотек 3C из эмбрионов рыбок данио, клеточных линий рыбок данио-рерио и молодых взрослых (YA) Caenorhabditis elegans (круглый червь). Протокол также должен быть подходящим для клеточных линий млекопитающих и, при дальнейшей адаптации, дрожжей.
Цель этих адаптаций — сделать 3C более доступным для студентов. Были приняты меры по использованию методов, аналогичных тем, которые могут быть выполнены в учебной лаборатории бакалавриата. Метод 3C предоставляет студентам множество возможностей для обучения, чтобы изучить основные методы молекулярной биологии, которые принесут пользу их развитию на стенде, в классе и в их начинаниях после окончания учебы.
протокол
1. Дизайн грунтовки
ПРИМЕЧАНИЕ: Инструменты проектирования праймеров 3C доступны в Интернете28. Кроме того, студенты могут разработать индивидуальные буквари (см. Ниже).
- Идентификация мест расположения грунтовки
- Откройте браузер генома UCSC (http://genome.ucsc.edu/), выберите исследуемый организм и выполните поиск в области генома, подлежащей оценке, с помощью 3C.
- В следующем окне активируйте трек ферментов на вкладке « Сопоставление и секвенирование» под браузером, нажав « Ферменты Restr».
- Введите ферменты рестрикции , которые будут использоваться, и установите режим отображения на упаковку. Нажмите « Отправить».
- В разделе « Вариация» и «Повторы» убедитесь, что для параметра RepeatMasker задано значение «Плотность».
- Используя сайты ограничений в качестве ориентиров, определите интересующие сайты, не входящие в замаскированные области повторения (черные полосы). Выделите 300 пар оснований (bp), обрамляющих как восходящую, так и нисходящую последовательность, окружающую место ограничения, щелкнув позицию (самая верхняя дорожка) и перетащив ее на желаемую длину (~ 600 bp).
- Отпустите кнопку мыши, и появится всплывающее окно. Обратите внимание на геномное местоположение, нажмите « Увеличить» и дайте браузеру перестроиться на выбор.
- Наведите указатель мыши на вкладку « Вид » на верхней ленте браузера и в раскрывающемся меню выберите DNA.
- Оставьте вариант по умолчанию, обратите внимание на обозначение замаскированных последовательностей (есть возможность сделать их N). Нажмите «Получить ДНК».
- Выделенная последовательность будет отображаться в окне. Скопируйте и вставьте эту последовательность в Primer3 (https://primer3.ut.ee/).
- Используя настройки Primer3 по умолчанию, сгенерируйте праймеры, нажав « Выбрать праймеры».
- В результатах обратите внимание на расположение фермента рестрикции и выберите праймеры, которые создают продукт ПЦР 200-500.н . с сайтом рестрикции , расположенным посередине.
- Следуя этим шагам, разработайте учебники по тестированию 1 килобазы (КБ), 2 КБ, 5 КБ, 10 КБ и 20 КБ с интересующих сайтов.
- Чтобы спроектировать праймеры входного контроля, повторите эти шаги для участка, находящегося на расстоянии 1-2 кБ от интересующего участка, на котором отсутствует узел ограничения, что означает, что он никогда не обрезается.
- Валидация функционального праймера
- Чтобы проверить функциональность праймера, настройте реакции ПЦР с использованием титрованных концентраций праймера и очищенной геномной ДНК. С помощью количественных или полуколичественных средств определите, создают ли грунтовки ожидаемый продукт.
- Перепроектируйте учебники, не прошедшие проверку.
2. День 1
ПРИМЕЧАНИЕ: Протокол может быть приостановлен (заморожен при -20 ° C) после сшивания хроматина и после сбора ядер. Шаги занимают, в среднем, 5-6 часов у студентов.
- Коллекция ядер C. elegans (адаптировано из Han et al.29)
- Хроматиновая сшивка
- Соберите 5,000 червей YA в 30 мл среды M9 в конической пробирке объемом 50 мл и вращайте при 400 × г в течение 2 мин при комнатной температуре (RT).
- Промойте гранулы червяка в 3 раза больше с помощью M9, чтобы удалить бактерии.
- Удалите надосадочную жидкость, ресуспендируйте гранулу червя в 47,3 мл M9, содержащей 2,7 мл 37% формальдегида (2% окончательного), и инкубируйте с перемешиванием (качанием или нутированием) в течение 30 мин при RT.
ВНИМАНИЕ: Обращайтесь с формальдегидом осторожно и работайте под вытяжным шкафом с надлежащими средствами индивидуальной защиты (лабораторный халат СИЗ, надлежащие перчатки и средства защиты глаз). Формальдегид является раздражителем, который поражает глаза, нос, горло и легкие. - Отжим червей при 400 × г в течение 2 мин при РТ.
- Удалите надосадочную жидкость и ресуспендируйте гранулу червя в 50 мл 1 М глицина. Закручивают червей при 400 × г в течение 2 мин при RT.
- Коллекция ядер
- Ресуспендировать червячную гранулу в 6 мл охлажденного NP-буфера (50 мМ HEPES при pH 7,5, 40 мМ NaCl, 90 мМ KCl, 2 мМ ЭДТА, 0,5 мМ EGTA, 0,1% Tween 20, 0,2 мМ DTT, 0,5 мМ спермидин, 0,25 мМ спермина, 1x полный ингибитор протеазы).
- Перенесите червячную суспензию в 7 мл свободно сидящего Dounce на льду. Сбросьте пробу в 15 раз и подержите на льду 5 минут.
- Перенесите червячную суспензию в плотно прилегающий Dounce на льду объемом 7 мл. Выдавите пробу в 20 раз и подержите на льду 5 минут.
- Перенесите червячную суспензию в чистую коническую пробирку объемом 15 мл и добавьте буфер NP в общей сложности до 10 мл (примерно 4 мл).
- Встряхните червячную суспензию на высокой настройке в течение 30 с. Инкубируйте образец на льду в течение 5 минут.
- Повторите предыдущий шаг.
- Отжим червячную суспензию при 100 × г в течение 5 мин при 4 °С.
- Перенесите надосадочную жидкость в свежую коническую пробирку объемом 15 мл.
- Проверьте образец на наличие червячного мусора. Визуализируйте 10 мкл образца с помощью светового микроскопа. Если присутствует червячный мусор, отжимают образец при 2000 × г в течение 5 мин при 4 °C, ресуспендируют гранулу в свежем 10 мл NP-буфера и снова вращают образец при 100 × г в течение 5 мин при 4 °C. Выбросьте гранулы, проверьте надосадочную жидкость на наличие мусора червей и повторяйте до тех пор, пока в образце не будет очищен мусор от червей.
ПРИМЕЧАНИЕ: В качестве альтернативы, червячный мусор также может быть удален путем процеживания образца через сетчатое фильтр для клеток 40 мкм (6x), а затем через сетчатое фильтр для клеток 20 мкм (6x). - Если в образце нет червячного мусора, возьмите 5 мкл образца, добавьте 5 мкл метилового зеленого пиронина (ядра будут синего цвета) и подсчитайте ядра с помощью гемоцитометра.
- Отжим образца при 2 000 × г в течение 5 мин при 4 °C
- Удалите надосадочную жидкость, поместите образцы на лед и продолжайте; в качестве альтернативы можно заморозить ядра и хранить при температуре -80 °C.
- Хроматиновая сшивка
- Переваривание хроматина
- Ресуспендируют ядра в 450 мкл чистой воды. Перенесите ядра в чистую микроцентрифужную пробирку объемом 1,5 мл.
- К сшитому образцу добавьте 60 мкл 10-кратного буфера фермента рестрикции DpnII и хорошо перемешайте.
ПРИМЕЧАНИЕ: Можно использовать другие рестрикционные ферменты, которые все еще разрезают сшитую ДНК. - Добавьте 15 мкл 10% додецилсульфата натрия (SDS) для проникновения ядер и инкубируйте с перемешиванием (качанием или нутированием) при 37 ° C в течение 1 часа.
ВНИМАНИЕ: SDS является раздражителем и токсичен при проглатывании или всасывании через кожу. Обращайтесь с осторожностью и используйте соответствующие СИЗ (лабораторный халат, подходящие перчатки и средства защиты глаз). - Гасят SDS, добавляя 75 мкл 20% Triton X-100 и инкубируя при перемешивании при 37 °C в течение 1 ч.
- Возьмите 10 мкл из образца в качестве непереваренного контроля. Хранить при температуре 4 °C.
- Добавьте 400 U DpnII и инкубируйте при 37 ° C в течение ночи при перемешивании.
3. День 2
ПРИМЕЧАНИЕ: В среднем студентам требуется 5 часов, чтобы выполнить эти шаги.
- Переваривание хроматина
- Добавьте дополнительно 200 ЕД DpnII и инкубируйте образцы в течение 4 ч при 37 °C с перемешиванием, чтобы обеспечить полное разложение сшитого образца.
- Возьмите аликвоту 10 мкл из образцов в качестве контроля пищеварения.
- Инактивируйте фермент рестрикции путем инкубации образца в течение 20 мин при 65 ° C (или в соответствии с инструкциями производителя). Перейдите к шагу 2.1.3.
- Если фермент не может быть инактивирован при нагревании, добавьте 80 мкл 10% SDS и инкубируйте образец в течение 30 минут при 65 ° C. Затем добавьте 375 мкл 20% Triton X-100 и перемешайте, перемешивая. Инкубируйте образец в течение 1 ч при 37 °C и переходите к шагу 2.2.
- Лигирование хроматина
- Переложите образец в чистую коническую пробирку объемом 50 мл, отрегулируйте объем образца до 5,7 мл с молекулярным H2O и перемешайте путем завихрения.
- Добавьте 700 мкл 10-кратного буфера T4-лигазы и перемешайте, перемешивая.
- Добавьте 60 ЕД ДНК-лигазы Т4 и перемешайте, перемешивая.
- Инкубировать в течение ночи при температуре 16 °C.
4. День 3
ПРИМЕЧАНИЕ: В среднем студентам требуется 15-30 минут, чтобы выполнить эти шаги. После ночной инкубации образцы можно заморозить.
- Переваривание белка и обратная сшивка
- Добавьте 30 мкл протеиназы К (10 мг / мл) в образец 3C и инкубируйте при 65 ° C в течение ночи с перемешиванием. Добавьте 5 мкл протеиназы К (10 мг / мл) к непереваренному и контролю пищеварения и инкубируйте при 65 ° C в течение ночи с перемешиванием.
5. День 4
ПРИМЕЧАНИЕ: В среднем студентам требуется 4-5 часов, чтобы выполнить эти шаги.
- Очистка библиотеки 3С
- Добавьте 30 мкл РНКазы А (10 мг / мл) к образцу 3C и перемешайте. Инкубировать образец в течение 45 мин при 37 °C.
- Добавьте 7 мл фенол-хлороформа в образец и перемешайте, встряхивая.
ВНИМАНИЕ: Фенол-хлороформ раздражает кожу и глаза и может вызвать ожоги, если контакт не лечить. Фенол-хлороформ следует использовать в вытяжном шкафу с надлежащей защитой глаз и перчатками (нитриловыми). - Центрифугируйте образец в течение 15 мин при дозе 3,270 × г при ЛТ. Соберите водную фазу и переложите в чистую коническую пробирку объемом 50 мл. Добавьте равные объемы хлороформа и перемешайте образец, встряхивая.
- Центрифугируют образец в течение 15 мин при 3,270 × г при ЛТ. Соберите водную фазу и переложите в чистую коническую пробирку объемом 50 мл. Добавьте 7,5 мл молекулярного H2O, 35 мл 100% этанола и (необязательно) 7 мкл гликогена (1 мг / мл). Перемешайте встряхиванием и инкубируйте при температуре -80 °C, пока образец не замерзнет.
ПРИМЕЧАНИЕ: Для замерзания образца может потребоваться 1 час или более. Здесь протокол можно поставить на паузу.- Пока образец 3C замерзает, очистите контрольные образцы. К контрольным образцам отрегулируйте объем до 500 мкл с помощью молекулярной воды и добавьте 2 мкл смеси РНКазы А (10 мг / мл), щелкнув пробиркой.
- Ненадолго открутите, чтобы собрать образец на дне пробирки. Инкубировать образцы в течение 45 мин при 37 °C.
- К контрольным образцам добавляют 1 мл фенолхлороформа и перемешивают, встряхивая пробирки. Центрифугируют образец в течение 15 мин при дозе 3,270 × г при ЛТ. Соберите водную фазу контрольной группы и поместите в чистую микроцентрифужную пробирку объемом 1.5 мл.
- Добавьте равные объемы хлороформа и перемешайте образец, встряхивая. Центрифугируют образец в течение 15 мин при 3,270 × г при ЛТ. Соберите водную фазу контрольной группы и поместите в чистую микроцентрифужную пробирку объемом 1.5 мл.
- Добавьте 1 мл 100% этанола и 2 мкл гликогена (1 мг/мл). Смешать встряхиванием и выдерживать при температуре −80 °C в течение 30 мин.
- Центрифугируют образец в течение 15 мин при 3 270 × г при 4 °C. Удалите надосадочную жидкость и добавьте 750 мкл охлажденного 70% этанола.
- Центрифугируют образец в течение 10 мин при 3 270 × г при 4 °C. Удалите надосадочную жидкость и высушите образцы на воздухе. Ресуспендируют гранулу в 50 мкл молекулярного H2O. Заморозьте или перейдите к шагу 6.
- Центрифугируют образец в течение 60 мин при 3 270 × г при 4 °C. Удалите надосадочную жидкость и добавьте 10 мл охлажденного 70% этанола. Разрушите и разбейте гранулы ДНК, встряхнув образец для смешивания.
- Центрифугируют образец в течение 30 мин при 3 270 × г при 4 °C. Удалите надосадочную жидкость и дайте образцу частично высохнуть на воздухе при ЛТ. Ресуспендируют гранулу в 150 мкл 10 мМ трис-HCl (рН 7,5) пипеткой вверх и вниз. Это «Библиотека 3C».
ПРИМЕЧАНИЕ: Заморозьте образец или продолжите анализ.
6. День 5
ПРИМЕЧАНИЕ: В среднем студентам требуется 1-2 часа, чтобы выполнить эти шаги.
- Определение качества образца по стандартной кривой контрольной грунтовки
- Количественно определите концентрацию ДНК для образца 3C, а также для всех контрольных групп и скорректируйте образцы до 30 мкг/мкл (если позволяют концентрации). Последовательно разбавляют образцы два раза четыре раза, в результате чего получается пять разведений для каждого образца (1x, 0.5x, 0.25x, 0.125x, 0.0625x).
ПРИМЕЧАНИЕ: Если количественное определение не может быть легко выполнено, последовательно разбавьте образцы, как указано выше, и продолжайте. - Настройте ПЦР-реакции для 3С, геномный контроль и переваренный контроль для каждого разбавленного образца с контрольными праймерами: 1 мкл ДНК, 10 мкл 5-кратного реакционного буфера, 1 мкл 10 мМ dNTP, 1 мкл контрольного праймера 10 мМ (прямого и обратного смешанного), 1 мкл Taq-полимеразы и 36 мкл воды.
- Следуя инструкциям к программному обеспечению, относящемуся к аппарату ПЦР, настройте программу ПЦР со стандартной кривой, используя условия циклирования следующим образом: 30 с при 98 °C; 30 циклов по 5 с при 98 °C, 5 с при 60 °C и 10 с при 72 °C; 1 мин при 72 °C; Выдерживать 4 °C.
- Используя программное обеспечение, сгенерируйте стандартные кривые для образцов. После того, как программное обеспечение сгенерирует кривую, обратите внимание на эффективность ПЦР и значение R2 .
- Количественно определите концентрацию ДНК для образца 3C, а также для всех контрольных групп и скорректируйте образцы до 30 мкг/мкл (если позволяют концентрации). Последовательно разбавляют образцы два раза четыре раза, в результате чего получается пять разведений для каждого образца (1x, 0.5x, 0.25x, 0.125x, 0.0625x).
- Определение концентрации ДНК
- Чтобы определить концентрацию ДНК образцов 3C, сравните образцы с образцом геномной ДНК известной концентрации. Разбавьте образцы до 30 нг/мкл и последовательно разбавьте геномный контроль с 10 нг/мкл до 0,01 нг/мкл в два этапа, чтобы создать стандартную кривую.
ПРИМЕЧАНИЕ: Если количественная оценка не может быть легко выполнена, разбавьте образец 1:10 и продолжайте. - Настройте реакции ПЦР для 3C, геномного контроля и переваренного контроля для каждого разбавленного образца с контрольными праймерами: 1 мкл ДНК, 10 мкл 5-кратного реакционного буфера, 1 мкл 10 мМ dNTP, 1 мкл контрольного праймера 10 мМ (прямого и обратного смешивания), 1 мкл Taq-полимеразы и 36 мкл воды.
- Следуя инструкциям к программному обеспечению, предназначенному для ПЦР-машины, настройте стандартную программу ПЦР-графики, используя следующие условия циклирования: 30 с при 98 °C; 30 циклов по 5 с при 98 °C, 5 с при 60 °C и 10 с при 72 °C; 1 мин при 72 °C; Выдерживать 4 °C.
- Используя программное обеспечение, сгенерируйте стандартные кривые для образцов, нанеся образцы 3C на кривую. Сравните положение содержания образца 3C со стандартной кривой, созданной реакциями геномной ДНК известной концентрации, и отрегулируйте образцы 3C до 30 нг / мкл.
- Чтобы определить концентрацию ДНК образцов 3C, сравните образцы с образцом геномной ДНК известной концентрации. Разбавьте образцы до 30 нг/мкл и последовательно разбавьте геномный контроль с 10 нг/мкл до 0,01 нг/мкл в два этапа, чтобы создать стандартную кривую.
- Определение наличия или отсутствия хроматинового взаимодействия
- Настройте реакции кПЦР с 30 нг образца 3C, геномного контрольного образца и переваренного контрольного образца: 1 мкл ДНК, 10 мкл 5-кратного реакционного буфера, 1 мкл 10 мМ dNTP, 1 мкл 10 мМ праймера (прямого и обратного смешанного), 1 мкл Taq-полимеразы и 36 мкл воды.
ПРИМЕЧАНИЕ: Реакции должны быть настроены с использованием контрольного праймера, тестовых праймеров для отдельных геномных локусов и желаемых комбинаций тестовых праймеров (сайт «a» вперед и сайт «b» назад), используемых для определения взаимодействия хроматина (рис. 3). - Следуя инструкциям к программному обеспечению ПЦР-машины, настройте программу ПЦР следующим образом: 30 с при 98 °C; 40 циклов по 5 с при 98 °C, 5 с при 60 °C и 10 с при 72 °C; 1 мин при 72 °C; Выдерживать 4 °C.
- После того, как ПЦР была запущена, осмотрите график амплификации. Убедитесь, что реакции ПЦР демонстрируют экспоненциальное усиление, удваиваясь каждый цикл.
ПРИМЕЧАНИЕ: Реакции, которые не показывают экспоненциального усиления, не могут быть проанализированы дальше. - Следуя инструкциям к программному обеспечению ПЦР-аппарата, определите порог эксперимента с кПЦР.
- Экспортируйте значения Ct для всех образцов.
- Определите эффективность сбраживания, используя значения Ct тестового праймера (TP) и контрольного праймера (CP) из непереваренного контрольного (UC) и переваренного контрольного (DC) образцов, используя уравнение (1).
Процент переваривания = 100 - (1)
(1) - Эти значения должны быть в пределах 80%-90% переваривания. Запишите эти значения (рис. 4A).
- Определите относительное взаимодействие хроматина, используя комбинацию тестового праймера (TPC) и контрольного праймера (CP) Ct из непереваренного контрольного (UC) и 3C (3C) образцов, используя уравнение (2).
Хроматиновое взаимодействие = 100 - (2)
(2) - Нанесите значения для каждого образца на гистограмму для каждой комбинации тестового праймера.
- Сравните сигнал образцов 3C с контрольными образцами, чтобы определить, имеет ли образец 3C обогащение определенного взаимодействия хроматина над контрольными образцами. Образцы 3C, которые показывают обогащение по сравнению с контролем, могут считаться условно положительными и требуют проверки с помощью секвенирования Сэнгера (рис. 4B).
ПРИМЕЧАНИЕ: При анализе данных графика важно помнить, что если не используется «контрольный шаблон» (см. обсуждение), содержание реакций (т.е. от одного набора праймеров к другому) не может быть сравнено.
- Настройте реакции кПЦР с 30 нг образца 3C, геномного контрольного образца и переваренного контрольного образца: 1 мкл ДНК, 10 мкл 5-кратного реакционного буфера, 1 мкл 10 мМ dNTP, 1 мкл 10 мМ праймера (прямого и обратного смешанного), 1 мкл Taq-полимеразы и 36 мкл воды.
- Идентификация продуктов 3C
- Запустите продукты ПЦР на 1,5% агарозном геле.
- Визуализируйте гель для правильного размера продуктов ПЦР с помощью УФ-светового короба или системы документирования геля.
ПРИМЕЧАНИЕ: Хорошо построенные библиотеки 3C будут содержать широкий спектр фрагментов ДНК, и, несмотря на предыдущую проверку праймеров на специфичность, реакции ПЦР из библиотек 3C могут содержать много полос (рис. 5A). Это не мешает анализу библиотек 3С. - Исключите полосы продукта, соответствующие ожидаемому размеру фрагмента, и выполните гелевую экстракцию продуктов ПЦР, следуя инструкциям коммерческого набора или самодельному протоколу, изложенному ниже.
ВНИМАНИЕ: Ультрафиолетовый свет является известным канцерогеном, и следует соблюдать большую осторожность, чтобы ограничить время воздействия ультрафиолетового света. Следует носить защиту глаз, устойчивую к ультрафиолетовому излучению, защиту от образцов, перчатки и лабораторный халат.- Чтобы сконструировать самодельный картридж для очистки, проткните иглой отверстие в дне пробирки объемом 0,5 мл.
- Упакуйте небольшое количество ваты из ватного тампона на дно пробирки объемом 0,5 мл, заполнив не более половины тюбика.
- Поместите пробирку 0.5 мл в пробирку 1.5 мл; Убедитесь, что меньшая трубка лежит на кромке большей трубки, а не в трубке.
- Аккуратно разрежьте фрагмент геля на более мелкие кусочки и поместите его в пробирку картриджа объемом 0,5 мл.
- Поместите сборку в морозильную камеру с температурой −20 °C на 5 минут. Отжимайте сборку в течение 3 мин при 13 000 × g при RT.
- Храните пробирку объемом 1,5 мл с извлеченной ДНК в буфере и утилизируйте пробирку объемом 0,5 мл, содержащую остатки агарозы.
- Очищенная ДНК может быть отправлена на секвенирование по Сэнгеру с использованием прямых и обратных праймеров для фрагмента 3C.
- Идентификация хроматиновых контактов с помощью Блата
- По завершении секвенирования определите, соответствуют ли образцы стандартам контроля качества, указанным в отчете о секвенировании.
- Для последовательностей, передающих контроль качества, откройте файлы .seq и .ab1 (трассировка) в программе редактирования последовательности, такой как Another Plasmid Editor (ApE), проверьте файл .ab1 на наличие четких базовых пиков вызова и отредактируйте пики, неправильно вызванные в файле .seq.
- Используя отредактированный SEQ-файл, найдите сайт DpnII. В зависимости от результата последовательности, это должно быть на полпути к сообщенной последовательности.
- Чтобы определить, является ли последовательность ожидаемой целевой последовательностью, выделите сайт DpnII и дополнительные 30-50.н. восходящей последовательности, соответствующие прямому праймеру геномных локусов, протестированных на 3C. Выполните поиск Блата этой последовательности по целевым видам. Убедитесь, что геномные локусы совпадают с локусами праймера.
- Повторите описанный выше шаг для последовательности обратного праймера, убедившись, что возвращенный геномный локус совпадает с локусом обратного праймера.
Результаты
В результате этой процедуры будет получен один экспериментальный образец 3C и два контрольных образца (непереваренный и переваренный). С использованием этих трех образцов была проведена кПЦР. На основе этих результатов была рассчитана эффективность сбраживания (уравнение 1) и зарегистрирована (таблица 1). Из этих расчетов было определено, что образец 3C имел эффективность пищеварения примерно 88% (среднее значение таблицы 1) по семи протестированным геномным локусам.
Затем образцы были протестированы на наличие дальних контактов хроматина между различными геномными локусами с использованием комбинаций локус-специфических праймеров (табл. 2) и кПЦР. Используя эти результаты, было рассчитано содержание продукта относительно контрольного набора праймера (уравнение 2) и построено график для сравнения (рис. 4). Эти данные показали, что 8 из 10 реакций были условно положительными для дальних взаимодействий.
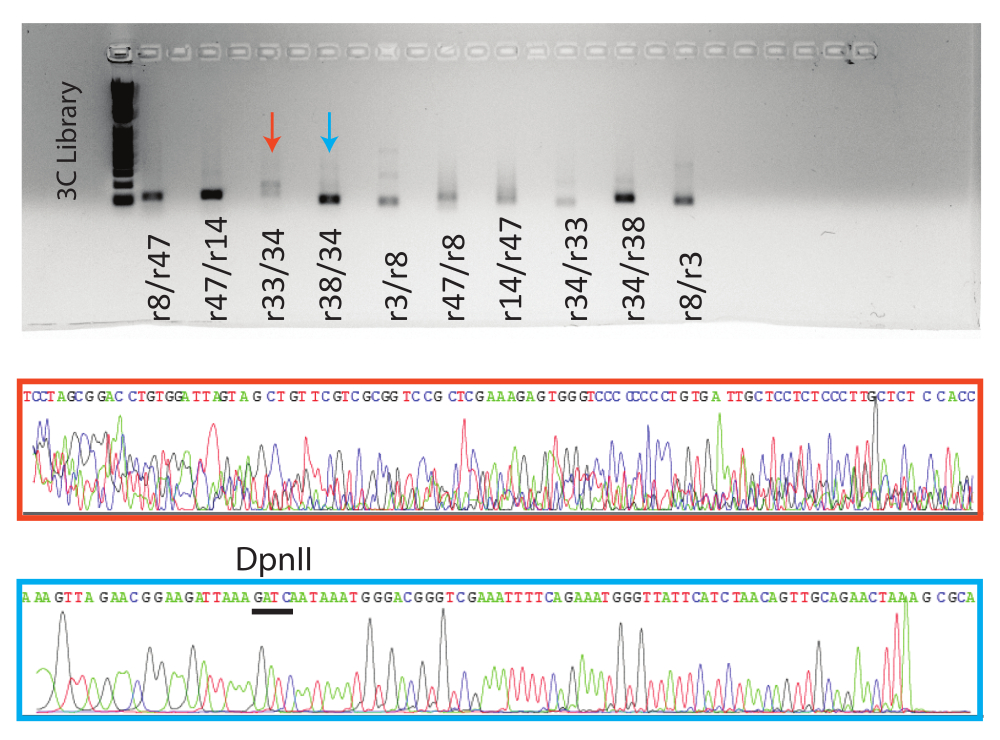
Затем реакции ПЦР запускали на агарозном геле. Ожидаемый продукт ПЦР для восьми условных положительных результатов и одной отрицательной реакции был очищен гелем и отправлен на секвенирование по Сэнгеру. Приведены результаты репрезентативных положительных (синяя стрелка, синяя рамка) и отрицательных (красная стрелка, красное прямоугольнико) реакций (рис. 5).

Рисунок 1: Структура хромосомы в ядре. Гипотетическая организация хромосом внутри ядра. а) ядерная оболочка, черные линии; (B) ядерная пластинка оранжевого цвета; (C) гетерохроматин, уплотненные линии; (D) эухроматин, свободные петли. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}

Рисунок 2: Схема протокола 3C. Отдаленные части линейных хромосом (синий, желтый и розовый) сближаются в ядре через регуляторные петли. (A) Петлевые структуры опосредованы транскрипционными факторами (серый круг и черная звезда); Эти взаимодействия сохраняются за счет химического сшивания. (B) Петли разрываются через ферментативное расщепление (черные линии). (C) Отдаленные кусочки хроматина перевязываются вместе через липкие концы, созданные в процессе пищеварения. (D) ДНК очищается от белка. (E) Последовательность внутри фрагментов идентифицируется и сопоставляется с геномом. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}

Рисунок 3: Схема грунтовки 3С. Праймеры 3C разработаны вокруг сайтов рестрикции на разных расстояниях от интересующего геномного местоположения. Розовые стрелки представляют экспериментальные наборы праймеров, окружающие сайт DpnII. Экспериментальные праймеры могут быть смешаны и сопоставлены для оценки петли хроматина в этой области. Оранжевые праймеры представляют собой отрицательный контроль без сайта DpnII. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}

Рисунок 4: Репрезентативные данные кПЦР для эксперимента 3C. Относительное обилие наборов тестовых праймеров 3C. (A) Контрольный график, показывающий относительное содержание продукта в непереваренном контроле (синий цвет), переваренном контроле (серый) и образце 3C (оранжевый). b) эксперимент 3С; относительное содержание продукта из комбинаций тестовых праймеров в непереваренном контроле (синий), переваренном контроле (серый) и образце 3C (оранжевый). Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}

Рисунок 5: Визуализация продуктов 3C qPCR. Сверху гель с конечными продуктами qPCR с указанными образцами. Внизу, репрезентативные файлы трассировки последовательности Сэнгера для указанных реакций. Оранжевый образец является примером ложноположительного результата (см. рис. 4, r38/34), а синий образец является примером истинного положительного результата с сайтом DpnII, указанным над трассировкой. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}
| Место | % переваривания |
| Р8 | 86.98 |
| Р3 | 88.44 |
| Р38 | 89.64 |
| Р34 | 87.55 |
| Р33 | 87.85 |
| Р14 | 86.97 |
| Р47 | 89.45 |
Таблица 1: Расчетная эффективность пищеварения.
| Имя | Последовательность | Геномные локусы | ||
| r3 FWD | ACGCAAGTAAAATTCTGGTTTTTGACC | chrX:11361475 | ||
| r3 RVS | TTTCCTGAGCTCTAACCATGTTTGC | chrX:11361561 | ||
| r38 FWD | TTACTTCTGAAGTAATCTTTTCTTATCCCC | chrX:5859700 | ||
| Р38 РВС | AGACGAGCTGATTAAAAGTAGTTGAG | chrX:5859775 | ||
| r34 FWD | ATTTGTGGATTGCGTGGAGACG | chrX:5429702 | ||
| r34 RVS | AATAATCCTCTTAACAAACGTGGCC | chrX:5429777 | ||
| r33 FWD | AAGAGTTGTCCAAAATAAATTGAGCTAAC | chrX:6296704 | ||
| r33 RVS | TTCAGAAAAGTAAACTTTGACTTGGAACG | chrX:6296807 | ||
| r14 FWD | AATTATCGATTTTTCCATCGCGCAG | chrX:8036367 | ||
| r14 RVS | ATTTCAATGAAAATGTAAAATGTTCCTTCTC | chrX:8036427 | ||
| r47 FWD | ATCTAGACTTGATAATATTTGTGTGTCCTC | chrX:9464939 | ||
| r47 RVS | AAGTTCTGCAACTGTTAGATGAATAACAC | chrX:9465064 | ||
| r8 FWD | ГАГААТГТТТТТГТТГТГААААКТТГ | chrX:11094257 | ||
| r8 РВС | TTACGAAATTTGGTAGTTTTGGACC | chrX:11094362 | ||
| Контрольная грунтовка FWD | CAATCGTCTCGCTCACTTGTC | chrX:7608049 | ||
| Контрольная грунтовка РВС | GATGTGAGCAACAAGGCACC | chrX:7608166 | ||
Таблица 2: Праймеры для репрезентативного эксперимента 3С.
Обсуждение
3C - это мощный метод, основанный на базовых молекулярных методах. Именно эта основа фундаментальных инструментов делает 3C такой интригующей техникой для использования со студентами. С таким количеством недавних исследований, наблюдающих динамику хроматина в таком широком масштабе, использование этих результатов для разработки узконаправленного эксперимента на одном гене или геномной области может создать уникальный и эффективный эксперимент в студенческих исследованиях. Часто подобные эксперименты считаются слишком сложными для студентов, но при тщательном планировании они легко достижимы. Важно отметить, что анализы, предназначенные для исследования соединений хроматина, захваченных библиотекой 3C, могут варьироваться от полуколичественной конечной ПЦР до полногеномного секвенирования. Фактически, данные из первой статьи3C 16 были получены с помощью кПЦР. Этот широкий спектр анализов может быть использован, потому что все технологии 3C производят один и тот же продукт - библиотеку фрагментов ДНК, представляющих 3D-связи в ядре.
Здесь представлена адаптация более гибкого и удобного протокола, который лучше подходит для студентов-исследователей. Периоды паузы, перечисленные выше, подразумевают ночные задержки; Однако эти паузы могут длиться в выходные дни, а в случае клеток и ядер - в течение нескольких недель. Наиболее важным фактором является то, когда работа будет выполнена. Часто в протоколах есть чувствительные ко времени шаги, когда приостановка невозможна. За пределами нескольких точек (день 1 и день 2) есть много мест, где можно остановить и заморозить образец. Это имеет решающее значение при работе со студентами, где графики и сроки лабораторных работ должны быть гибкими. В дополнение к включению этих остановок в протокол, студентам рекомендуется работать в парах или даже в небольших группах по три или четыре человека. Группы хорошо подходят для этого протокола, так как учащиеся могут поддерживать друг друга и создавать систему друзей, чтобы все работали безопасно. Лабораторная работа также доставляет больше удовольствия другим участникам. В группах студенты также могут работать над различными вопросами, ориентированными на организацию хроматина, выполняя при этом один и тот же протокол. Таким образом, даже когда студенты работают над отдельными проектами, протокол связывает их усилия, и благодаря этому они могут поддерживать друг друга.
Другие адаптации предназначены для обхода того факта, что определенные специализированные инструменты и оборудование не обязательно можно найти во всех учебных заведениях. К этому оборудованию относятся, помимо прочего, термоциклеры для кПЦР, системы документирования геля и нанообъемные спектрометры. Действительно, эти элементы оборудования удобны, но не являются обязательным требованием. Здесь классический метод 3C также описан в части дизайна грунтовки; Это включает в себя идентификацию интересующего геномного локуса и, исходя из этого, оценку других геномных локусов для точек контакта хроматина, находящихся дальше. Этот метод также хорошо работает, если используется опубликованный набор данных, такой как набор данных с использованием Hi-C, где идентифицируются известные положительные (соединяющие) и отрицательные (несоединяющиеся) локусы. Разработка экспериментов с использованием этих опубликованных наборов данных является еще одной отличной адаптацией для учебных лабораторий, поскольку шанс на успешную идентификацию хроматиновых соединений обычно выше. Кроме того, исследовательская статья может обсуждаться в классе и использоваться в качестве справочного материала.
Этот протокол использует модифицированный подход qPCR для визуализации формирования продукта 3C. Элементы управления необходимы для успеха техники 3C. В каждом эксперименте используются как контрольные образцы, так и контрольные элементы праймера, чтобы определить завершение процедуры 3C. Контрольные образцы включают непереваренный контроль (геномная ДНК) и переваренный контроль. Непереваренный контроль определяет базовый сигнал для наборов праймеров и используется с контролем сшитого сбраживания для определения эффективности сбраживания. Ожидается, что произойдет падение продукта для любых праймеров, направленных через сайт рестрикции. Сравнение этого значения с непереваренным контролем дает представление о том, насколько хорошо образец был переварен.
Праймеры для ПЦР включают контрольный праймер и тестовые праймеры. Контрольный праймер представляет собой набор праймеров, который находится рядом с анализируемой областью генома и не содержит сайта рестрикции. Это обеспечивает базовый уровень для определения обилия тестовых праймерных продуктов ПЦР. Тестовые праймеры представляют собой прямые и обратные праймеры, которые обрамляют сайт рестрикции для определенного геномного локуса, представляющего интерес (рис. 3). Реакции с использованием этих наборов праймеров сравниваются для определения эффективности сбраживания, так как содержание продукта должно снизиться, если место ограничения было разрезано. При определении организации хроматина один тестовый праймер из одного локуса соединяется с другим тестовым праймером из другого геномного локуса, чтобы определить, находятся ли эти два локуса близко друг к другу в 3D-пространстве. В этом случае ожидается, что продукт ПЦР будет найден только с использованием образца 3C в качестве шаблона.
Важно отметить, что даже проверенные праймеры имеют тенденцию к выходу из строя (рис. 4: набор праймеров r14). Кроме того, продукты ПЦР часто идентифицируются в контрольных реакциях и в реакциях, в которых соединение хроматина не прогнозируется (например, переваренный контроль, поскольку он не лигирован). Эти экземпляры секвенированы и либо не проходят проверку качества Сэнгера, либо возвращаются без определенной последовательности (рис. 5). Кроме того, традиционные эксперименты 3C генерируют «контрольный шаблон», несшитый, переваренный и лигированный образец ДНК, который представляет все возможные лигированные фрагменты, которые могут быть получены с заданным количеством ДНК. «Контрольный шаблон» играет важную роль в сравнении интенсивности сигналов кПЦР между двумя геномными локусами, чтобы определить, представляет ли сигнал истинное взаимодействие или просто случайную ассоциацию. Создание «контрольного шаблона» может быть проблематичным, так как большая часть анализируемого хроматина должна быть захвачена в виде искусственной хромосомы и обработана вместе с образцами 3C. Обеспечение такой конструкции может оказаться неосуществимым, и ее создание может выйти за рамки семестрового проекта. Из-за этих сложностей мы предлагаем использовать контрольную грунтовку. Контрольный праймер не заменяет всю функциональность «шаблона управления», но все же предоставляет возможность анализировать данные для определения «присутствия» или «отсутствия».
При проведении кПЦР важно использовать равные количества образца. Это должно быть определено, даже если используется наноспектрофотометр, такой как нанокапля, путем создания стандартной кривой из геномной ДНК известной концентрации и подгонки образцов 3C к этой линии. Эти количества должны быть зарегистрированы и использованы в последующих ПЦР. Также важно качество реакции ПЦР. По мере проведения ПЦР в кПЦР содержание продукта измеряется с помощью флуоресценции и регистрируется. Эта запись доступна в графике усиления. После завершения программы важно проверить график усиления и убедиться, что реакции (за исключением элементов управления без шаблона) имеют три фазы: базовую, экспоненциальную и фазу плато/насыщения. Важно проверить, что реакции имеют экспоненциальную фазу, в частности для установки порога (см. ниже). Кроме того, для серийно разбавленных образцов должен наблюдаться сдвиг значений Ct в соответствии с разбавлением образца (самая высокая концентрация будет иметь самые низкие значения Ct, а самая низкая концентрация будет иметь самые высокие значения Ct). Образцы, которые не отражают это изменение на графике амплификации, требуют нового разбавления или указывают на более серьезную проблему с образованием образца 3C. Наконец, при создании стандартной кривой программное обеспечение ПЦР вычисляет эффективность ПЦР и значение R2 . Эффективность ПЦР должна быть больше 90%, а значение R2 должно быть больше 0,99. Если какое-либо из этих условий не выполняется, вполне вероятно, что что-то не так с образцом или праймерами ПЦР.
После кПЦР процент переваривания и наличие взаимодействий 3C можно рассчитать с помощью кПЦР Ct для каждой реакции. Чтобы определить их, сначала необходимо установить порог реакции ПЦР. Обычно это делается с помощью программного обеспечения, поставляемого с машиной qPCR. Установка порога определит концентрацию продукта ПЦР, которая будет использоваться для сравнения значений Ct образца. Порог должен делить пополам кривые амплификации реакций ПЦР в экспоненциальной фазе амплификации. Можно сравнивать только реакции ПЦР с экспоненциальной амплификацией (в данном случае контрольные праймеры и реакции тестового праймера), так как это единственный способ убедиться, что реакции амплифицируют ДНК с одинаковой скоростью и могут быть точно сравнены. При анализе графиков 3C условно положительные реакции идентифицируются как реакции с большим количеством продукта по сравнению с контрольными образцами, геномным контролем и контролем пищеварения (рис. 4B). Однако эти образцы должны быть дополнительно проверены с использованием секвенирования Сэнгера после очистки геля продукта ПЦР.
После секвенирования по Сэнгеру образцы, прошедшие контроль качества, могут быть проанализированы с помощью Blat. Цель этого анализа состоит в том, чтобы определить, имеет ли образец последовательность обоих целевых геномных локусов, фланкирующих сайт рестрикции (DpnII в случае этого протокола). Если обе последовательности идентифицированы, то фрагмент 3C можно считать валидированным. Если результаты Блата не возвращают ожидаемую последовательность, это может указывать на то, что один или оба праймера не являются оптимальными, что приводит к ложноположительному результату кПЦР. Файлы трассировки для ложноположительных выборок будут иметь неопределенные базовые пики, а отчеты Seq будут содержать в основном «n» базовых вызовов.
Валидация по Сэнгеру имеет важное значение, так как возможны ложные срабатывания при формировании артефактного продукта ПЦР. Эти ложноположительные результаты могут быть идентифицированы, когда продукты секвенирования не имеют ожидаемой целевой последовательности или сайта DpnII, характерного для правильного фрагмента 3C (рис. 5). Секвенирование фрагментов ПЦР также предоставляет еще одну точку данных для эксперимента и дает понять студентам, что метод 3C идентифицирует отдаленные геномные локусы, которые собираются вместе в 3D-пространстве внутри ядер.
Метод 3C предоставляет студентам множество основополагающих молекулярных методов в гибкой и простой процедуре. Этот метод 3C также является отправной точкой для других методов 3C, которые включают секвенирование следующего поколения (NGS). Эти типы экспериментов могут познакомить студентов с важными аспектами биоинформатики и основаны на основных принципах, изложенных здесь. Опыт и участие студентов являются ключом к их успеху и развитию в качестве молодых ученых. Предоставляя эти возможности, студенты могут укрепить свое понимание основных принципов, одновременно укрепляя уверенность в себе для решения передовых методов и вопросов.
Раскрытие информации
Авторы заявляют об отсутствии конфликта интересов.
Благодарности
Эта работа была частично поддержана Премией институционального развития Род-Айленда (IDeA) Сетью передового опыта в области биомедицинских исследований от Национального института общих медицинских наук Национальных институтов здравоохранения в рамках гранта No P20GM103430 и Центра здоровья и поведенческих наук Брайанта.
Материалы
| Name | Company | Catalog Number | Comments |
| 37% Formaldehyde | Millapore-Sigma | F8775 | |
| 100% Ethanol | Millapore-Sigma | E7023 | |
| CaCl2 | MP Biomedical | 215350280 | |
| chloroform | Millapore-Sigma | C0549 | |
| cOmplete, EDTA-free Protease Inhibitor Cocktail | Millapore-Sigma | COEDTAF-RO | mixed to 50x in water. Diluted to 1x in Sucrose buffer and GB buffer fresh |
| Dithiothreitol (DTT) | Millapore-Sigma | D0632 | 1 M stock diluted to 500 µM in Sucrose buffer and GB buffer fresh |
| DpnII | NEB | R0543M | |
| Glycerol | Millapore-Sigma | G9012 | |
| glycine | Millapore-Sigma | G8898 | |
| glycogen | Millapore-Sigma | 10901393001 | |
| HEPES | Millapore-Sigma | H3375 | |
| KCl | Millapore-Sigma | P3911 | |
| KH2PO4 | Millapore-Sigma | P5655 | |
| methyl green pyronin | Millapore-Sigma | HT70116 | |
| MgAc2 | Thermoscientific | 1222530 | |
| Na2HPO4 | Millapore-Sigma | S5136 | |
| NaCl | Millapore-Sigma | S9888 | |
| phenol-chloroform | Millapore-Sigma | P3803 | |
| Pronase | Millapore-Sigma | 11459643001 | |
| Proteinase K | IBI Scientific | IB05406 | |
| qPCR Ready mix (Phire Taq etc) | Millapore-Sigma | KCQS07 | |
| RNase A | Millapore-Sigma | R6148 | |
| Sodium Acetate | Millapore-Sigma | S2889 | |
| sodium dodecyl sulfate (SDS) | Millapore-Sigma | L3771 | |
| Sucrose | Millapore-Sigma | S0389 | |
| T4 DNA Ligase | Promega | M1804 | |
| Tris-HCl | Millapore-Sigma | 108319 | |
| Triton X-100 | Millapore-Sigma | T9284 | |
| Trypsin-EDTA | Millapore-Sigma | T4049 |
Ссылки
- McBryant, S. J., Adams, V. H., Hansen, J. C. Chromatin architectural proteins. Chromosome Research. 14 (1), 39-51 (2006).
- Nalabothula, N., et al. The chromatin architectural proteins HMGD1 and H1 bind reciprocally and have opposite effects on chromatin structure and gene regulation. BMC Genomics. 15, 92(2014).
- John, S., et al. Chromatin accessibility pre-determines glucocorticoid receptor binding patterns. Nature Genetics. 43 (3), 264-268 (2011).
- Fawcett, D. W. On the occurrence of a fibrous lamina on the inner aspect of the nuclear envelope in certain cells of vertebrates. The American Journal of Anatomy. 119 (1), 129-145 (1966).
- Reddy, K. L., Zullo, J. M., Bertolino, E., Singh, H. Transcriptional repression mediated by repositioning of genes to the nuclear lamina. Nature. 452 (7184), 243-247 (2008).
- Finlan, L. E., et al. Recruitment to the nuclear periphery can alter expression of genes in human cells. PLoS Genetics. 4 (3), e1000039(2008).
- Chambeyron, S., Da Silva, N. R., Lawson, K. A., Bickmore, W. A. Nuclear re-organisation of the Hoxb complex during mouse embryonic development. Development. 132 (9), 2215-2223 (2005).
- Chambeyron, S., Bickmore, W. A. Chromatin decondensation and nuclear reorganization of the HoxB locus upon induction of transcription. Genes and Development. 18 (10), 1119-1130 (2004).
- Mahy, N. L., Perry, P. E., Gilchrist, S., Baldock, R. A., Bickmore, W. A. Spatial organization of active and inactive genes and noncoding DNA within chromosome territories. The Journal of Cell Biology. 157 (4), 579-589 (2002).
- Mahy, N. L., Perry, P. E., Bickmore, W. A. Gene density and transcription influence the localization of chromatin outside of chromosome territories detectable by FISH. The Journal of Cell Biology. 159 (5), 753-763 (2002).
- Li, B., Carey, M., Workman, J. L. The role of chromatin during transcription. Cell. 128 (4), 707-719 (2007).
- Chen, A., Chen, D., Chen, Y. Advances of DNase-seq for mapping active gene regulatory elements across the genome in animals. Gene. 667, 83-94 (2018).
- Hughes, A. L., Rando, O. J. Mechanisms underlying nucleosome positioning in vivo. Annual Review of Biophysics. 43, 41-63 (2014).
- Weiner, A., Hughes, A., Yassour, M., Rando, O. J., Friedman, N. High-resolution nucleosome mapping reveals transcription-dependent promoter packaging. Genome Research. 20 (1), 90-100 (2010).
- Hughes, A. L., Jin, Y., Rando, O. J., Struhl, K. A functional evolutionary approach to identify determinants of nucleosome positioning: A unifying model for establishing the genome-wide pattern. Molecular Cell. 48 (1), 5-15 (2012).
- Dekker, J., Rippe, K., Dekker, M., Kleckner, N. Capturing chromosome conformation. Science. 295 (5558), 1306-1311 (2002).
- van Berkum, N. L., Dekker, J. Determining spatial chromatin organization of large genomic regions using 5C technology. Methods in Molecular Biology. 567, 189-213 (2009).
- Smith, E. M., Lajoie, B. R., Jain, G., Dekker, J. Invariant TAD boundaries constrain cell-type-specific looping interactions between promoters and distal elements around the CFTR locus. American Journal of Human Genetics. 98 (1), 185-201 (2016).
- Crane, E. E. Two inputs into C. elegans dosage compensation: Chromosome conformation and the miRNA-specific argonaute ALG-2. University of California, Berkeley. , PhD thesis (2012).
- Bell, A. C., West, A. G., Felsenfeld, G. The protein CTCF is required for the enhancer blocking activity of vertebrate insulators. Cell. 98 (3), 387-396 (1999).
- Cuadrado, A., et al. Specific contributions of cohesin-SA1 and cohesin-SA2 to TADs and polycomb domains in embryonic stem cells. Cell Reports. 27 (12), 3500-3510 (2019).
- Sarro, R., et al. Disrupting the three-dimensional regulatory topology of the Pitx1 locus results in overtly normal development. Development. 145 (7), (2018).
- Tolhuis, B., et al. Interactions among polycomb domains are guided by chromosome architecture. PLoS Genetics. 7 (3), e1001343(2011).
- Splinter, E., de Wit, E., van de Werken, H. J. G., Klous, P., de Laat, W. Determining long-range chromatin interactions for selected genomic sites using 4C-seq technology: From fixation to computation. Methods. 58 (3), 221-230 (2012).
- Fernández-Miñán, A., Bessa, J., Tena, J. J., Gómez-Skarmeta, J. L. Chapter 21 - Assay for transposase-accessible chromatin and circularized chromosome conformation capture, two methods to explore the regulatory landscapes of genes in zebrafish. Methods in Cell Biology. 135, 413-430 (2016).
- Dekker, J. The three "C" s of chromosome conformation capture: controls, controls, controls. Nature Methods. 3 (1), 17-21 (2006).
- Hagège, H., et al. Quantitative analysis of chromosome conformation capture assays (3C-qPCR). Nature Protocols. 2 (7), 1722-1733 (2007).
- Lajoie, B. R., van Berkum, N. L., Sanyal, A., Dekker, J. My5C: Webtools for chromosome conformation capture studies. Nature Methods. 6 (10), 690-691 (2009).
- Han, M., Wei, G., McManus, C. E., Hillier, L. W., Reinke, V. Isolated C. elegans germ nuclei exhibit distinct genomic profiles of histone modification and gene expression. BMC Genomics. 20 (1), 500(2019).
Перепечатки и разрешения
Запросить разрешение на использование текста или рисунков этого JoVE статьи
Запросить разрешениеСмотреть дополнительные статьи
This article has been published
Video Coming Soon
Авторские права © 2025 MyJoVE Corporation. Все права защищены