
Method Article
Simultaneous Whole-cell Recordings from Photoreceptors and Second-order Neurons in an Amphibian Retinal Slice Preparation
In This Article
Summary
We describe the preparation of thin retinal slices from aquatic tiger salamanders (Ambystoma tigrinum) and explain how we use these slices to study synaptic processing in the retina by obtaining dual whole-cell voltage clamp recordings from photoreceptors and second-order horizontal and bipolar cells.
Abstract
One of the central tasks in retinal neuroscience is to understand the circuitry of retinal neurons and how those connections are responsible for shaping the signals transmitted to the brain. Photons are detected in the retina by rod and cone photoreceptors, which convert that energy into an electrical signal, transmitting it to other retinal neurons, where it is processed and communicated to central targets in the brain via the optic nerve. Important early insights into retinal circuitry and visual processing came from the histological studies of Cajal1,2 and, later, from electrophysiological recordings of the spiking activity of retinal ganglion cells - the output cells of the retina3,4.
A detailed understanding of visual processing in the retina requires an understanding of the signaling at each step in the pathway from photoreceptor to retinal ganglion cell. However, many retinal cell types are buried deep in the tissue and therefore relatively inaccessible for electrophysiological recording. This limitation can be overcome by working with vertical slices, in which cells residing within each of the retinal layers are clearly visible and accessible for electrophysiological recording.
Here, we describe a method for making vertical sections of retinas from larval tiger salamanders (Ambystoma tigrinum). While this preparation was originally developed for recordings with sharp microelectrodes5,6, we describe a method for dual whole-cell voltage clamp recordings from photoreceptors and second-order horizontal and bipolar cells in which we manipulate the photoreceptor's membrane potential while simultaneously recording post-synaptic responses in horizontal or bipolar cells. The photoreceptors of the tiger salamander are considerably larger than those of mammalian species, making this an ideal preparation in which to undertake this technically challenging experimental approach. These experiments are described with an eye toward probing the signaling properties of the synaptic ribbon - a specialized synaptic structure found in a only a handful of neurons, including rod and cone photoreceptors, that is well suited for maintaining a high rate of tonic neurotransmitter release7,8 - and how it contributes to the unique signaling properties of this first retinal synapse.
Protocol
1. Retinal Slices Preparation
- Assemble the chamber (design illustrated in Figure 1). Place two beads of vacuum grease, spaced ~8-10 mm apart, across the recording chamber to form a channel for superfusate and in which to embed the retinal slices. Add a second bead of grease a few millimeters farther out beyond each of these two beads of grease to act as a levee and limit spillover. Place a small triangular piece of KimWipe at the end of the chamber to ensure fluid contact with the reference electrode.
- Press a piece of nitrocellulose membrane (~ 5 x 10 mm; 0.8 μm pores) flat against the glass microscope slide into two small beads of vacuum grease. Avoid putting grease directly beneath the center of the nitrocellulose membrane, as this can prevent the retina from adhering.
- To prepare the tissue slicer, break a double edge razor blade into 4 pieces and attach one to the slicing arm. Cut a thin slice of nitrocellulose membrane to ensure that the cutting edge of the razor blade lays flat against the recording chamber and therefore cuts cleanly through the nitrocellulose membrane.
- Keep a small beaker of amphibian saline solution (Table 1) on ice at the dissection station.
- Euthanize salamander by decapitation. Hemisect the head sagitally and pith through the spinal cord. Place half of the head on a piece of cotton moistened with amphibian saline atop a linoleum block. The other half of the head can be wrapped with a moist paper towel and stored at 4 °C for use later in the day.
- Enucleate the eye. Using small Vannas scissors, cut the skin connecting the eye to the surrounding orbit. After freeing the front of the eye from the surrounding orbital tissue, pull the eye forward and slide the scissors under the eye to cut through the eye muscles and optic nerve, freeing the eye from the orbit.
- Place the enucleated eye on a bed of cotton on the linoleum block. Discard the half head. Trim excess orbital fat from the back of the eye.
- Make a small incision in the center of cornea with a sharp surgical blade. Remove the cornea by sliding fine Vannas scissors into the incision and extending the cut radially out toward the ora serrata. Cut circumferentially around the ora serrata by rotating the linoleum block or the cotton between cuts.
- After cutting all the way around the eye, remove the cornea and lens by pulling them out of the side of the eyecup. Move the resulting eyecup onto a hard surface of the linoleum block moistened with amphibian saline solution. Cut it into thirds with a sharp razor blade, using a fine sawing motion to ensure that you have cut all the way through the sclera.
- Place one or two pieces of eyecup on the nitrocellulose membrane with the retinal surface facing down. Submerge the remaining pieces with additional saline and place them in the refrigerator at ~4 °C.
- Gently press the piece of eyecup against the nitrocellulose membrane with fine forceps. Submerge the nitrocellulose membrane and eyecup piece with several drops of cold amphibian saline and blot at the edges with KimWipe to help the retina to adhere. Once again, submerge the eyecup and nitrocellulose membrane with several drops of cold amphibian saline solution and peel away the sclera/choroid/retinal pigment epithelium to isolate the retina (which can appear pink due to the presence of unbleached rhodopsin). If necessary, cut the optic nerve to free the retina.
- If the retina has not adhered tightly, drain away the saline with a KimWipe to pull the retina more firmly down onto the nitrocellulose membrane. Replace the saline. Repeat, if necessary.
- Fill the chamber with cold amphibian saline and transfer it to the stage of the tissue slicer. Slice the retina and nitrocellulose membrane into thin strips, working from one end to the other by turning the vernier micrometer in 125 μm increments. Press the razor blade gently but firmly through the retina and nitrocellulose membrane.
- Transfer the retinal slices by moving strips of nitrocellulose membrane to the main channel of the recording chamber. Lift a strip of membrane free and then hold it in place while moving the chamber beneath it, being sure to keep the slices submerged. Embed the edges of the nitrocellulose membrane in the strips of vacuum grease, rotating them 90 degrees to view the retinal layers.
- Press the nitrocellulose membrane flat against the glass surface. Even if there is not retina on every piece, place strips of nitrocellulose membrane at regular intervals (~1 mm apart) along the entire length of the perfusion channel to help break up the surface tension and improve fluid flow.
2. Paired Whole-cell Recordings
- After all of the slices have been transferred, move the recording chamber to the stage of an upright, fixed stage microscope and attach the reference electrode lead. Focus on the slices using a long-working distance, water immersion, 40-60X objective. The microscope should be placed on an air table to dampen vibrations and enclosed in a Faraday cage to reduce electrical interference.
- Superfuse the slices continuously at a rate of 1 ml/min with amphibian saline solution bubbled with 100% O2. Connect the suction, making sure that the inflow and outflow are balanced. Outflow can be regulated by rotating the beveled end of the suction needle or by moving the KimWipe at the end of the chamber closer or farther away from the outflow needle. The preparation can be kept at room temperature or cooled with a Peltier device or by simply setting an ice pack on the microscope stage.
- Examine slices under dim or infra-red light and identify a pair of cells - a photoreceptor (rod or cone) and nearby horizontal or bipolar cell - to target for whole-cell recording. Rods can be identified by their large cell bodies and prominent rod-like outer segments (Figure 2A). Cones are smaller than rods and have small tapered outer segments. Bipolar cell and horizontal cell somas are in the outermost row of cell bodies in the inner nuclear layer (INL; Figures 2B and 2C).
- Before preparing slices, use a pipette puller to manufacture micropipettes from borosilicate glass (1.2 mm outer diameter, 0.95 mm inner diameter with a glass filament). The tip of each micropipette should be ~1-2 microns in diameter.
- Using a non-metallic filling needle (e.g. one manufactured from a 1 cc syringe or a MicroFil), fill pipettes with the intracellular solution (Table 1) and attach to the electrode holder.
- Elevate the microscope objective slightly. Position the photoreceptor pipette beneath the objective and then lower it so that the tip is positioned just above the slices. Repeat with the second pipette.
- Adjust any offset in the baseline current level on the amplifier. Check the pipette resistance with a 5-10 mV depolarizing pulse. We typically use pipettes that range from 10-15 MΩ, the result of the long taper of the shaft and low osmolarity of the amphibian pipette solutions. With higher osmolarity mammalian solutions, these same pipettes exhibit resistance values of ~8-12 MΩ. While we have used larger tip diameters with resistance values of 3-4 MΩ in amphibian solutions, the advantages provided by a lower access resistance are offset by a greater difficulty in sealing onto cell membranes and a more rapid rundown of calcium currents and other second messenger-sensitive responses.
- While applying slight positive pressure, position the post-synaptic pipette so that it contacts the horizontal or bipolar cell body. Then position the presynaptic pipette so that it contacts the cell body of a rod or cone photoreceptor. Recordings appear to be more stable when pipette tips contact the inner segment rather than the soma, especially in cones.
- While monitoring the resistance, release the positive pressure on the post-synaptic pipette. Sometimes, the release of positive pressure is sufficient to form a gigaohm seal. If not, apply gentle suction with a 1 ml syringe or by mouth. After the tip resistance has grown to > 100 MΩ, apply a holding potential of -60 mV. After obtaining a gigaohm seal, null out any pipette capacitance transients and repeat the sealing procedure for the photoreceptor pipette, applying a holding potential of -70 mV.
- Rupture the patch by using your mouth or a syringe to apply suction to each cell in turn. Rods, cones, and bipolar cells will typically rupture with gentle suction. Obtaining whole-cell configuration with a horizontal cell may require greater suction (i.e. with a 3 cc syringe) in combination with strong quick voltage pulses delivered with the "zap" feature of the patch clamp amplifier. Rupture of the membrane and establishment of whole-cell configuration will be evident by the appearance of whole-cell capacitance transients.
- Confirm identity of the post-synaptic cell physiologically by applying a light flash and delivering a series of voltage steps from -120 to +40 mV in 20 mV increments (Figures 3 and 4).
- To assess if the pair of cells are synaptically connected, deliver a brief (25-100 msec), 60 mV step depolarization to the photoreceptor (to -10 mV, near the peak of the L-type voltage-gated calcium current) and look for post-synaptic currents in the second order neuron (Figure 5). A strong depolarizing step should evoke a fast, transient inward post-synaptic current in the post-synaptic horizontal or OFF bipolar cell caused by a burst of vesicle release from the cone (Figure 5).
Results
Representative traces of light responses from neurons in vertical slices of salamander retina are shown in Figure 3. The cone, horizontal cell, and OFF bipolar cell all display an outward current in response to light onset. The prominent inward current following the light flash in the horizontal and bipolar cell recordings is caused by the increased release of glutamate from photoreceptors as they depolarize at light offset. The ON bipolar cell responds with an inward current at light onset resulting from a sign-inverting metabotropic glutamate receptor signaling cascade and activation of TRPM1 channels9. Horizontal cells and bipolar cells can be distinguished from one another by their I-V relationships (Figure 4). Horizontal cells typically have a linear or inwardly rectifying I-V and low input resistance (<500 MΩ; Figure 4A) while bipolar cells have a high input resistance (0.5-2 GΩ) and outwardly-rectifying I--V (Figure 4B). Figure 5 shows representative results from recordings of a cone-horizontal cell pair (Figure 5A) and cone-OFF bipolar cell pair (Figure 5B). In each, depolarizing the cone to -10 mV from a holding potential of -70 mV evoked a voltage-gated calcium current in the cone and fast inward EPSC in the horizontal or bipolar cell. This strong stimulus is sufficient to empty the readily-releasable pool of ~20 vesicles from each synaptic ribbon, resulting in an EPSC of ~47 pA/ribbon10. The horizontal cell EPSC in Figure 5A was 232 pA, suggesting that it received 5 ribbon contacts from the presynaptic cone. A similar estimate from the 178 pA EPSC in the off bipolar cell (Figure 5B) suggests that it received 4 ribbon contacts from the presynaptic cone.
| Amphibian saline | Presynaptic pipette | Postsynaptic pipette | |
| NaCl | 116 mM | 3.5 mM | 3.5 mM |
| KCl | 2.5 mM | ||
| CaCl2 | 1.8 mM | 1 mM | 1 mM |
| MgCl2 | 0.5 mM | 1 mM | 1 mM |
| HEPES | 10 mM | 10 mM | 10 mM |
| Glucose | 5 mM | ||
| Cs-glutamate* | 40 mM | ||
| Cs-gluconate** | 50 mM | 90 mM | |
| Tetraethylammonium Chloride | 10 mM | 10 mM | |
| ATP-Mg | 9 mM | 9 mM | |
| GTP | 0.5 mM | 0.5 mM | |
| EGTA | 5 mM | 5 mM | |
| pH*** | 7.8 | 7.2 | 7.2 |
| Osmolarity**** | 245 mOsm | 240 mOsm | 240 mOSM |
*Cs-glutamate is made by neutralizing 40 mM L-glutamic acid with 40 mM CsOH in the pipette solution. ** A 1 M stock of Cs-gluconate is made by neutralizing a solution of CsOH with 45-50% D-gluconic acid. ***The pH should be adjusted with NaOH for the extracellular solution and CsOH for the pipette solutions. **** Keeping the osmolarity of the pipette solutions just below that of the extracellular solution prevents cell swelling and enhances longevity of the recording.
Table 1. Components and parameters for the standard intracellular and extracellular solutions used in this protocol.

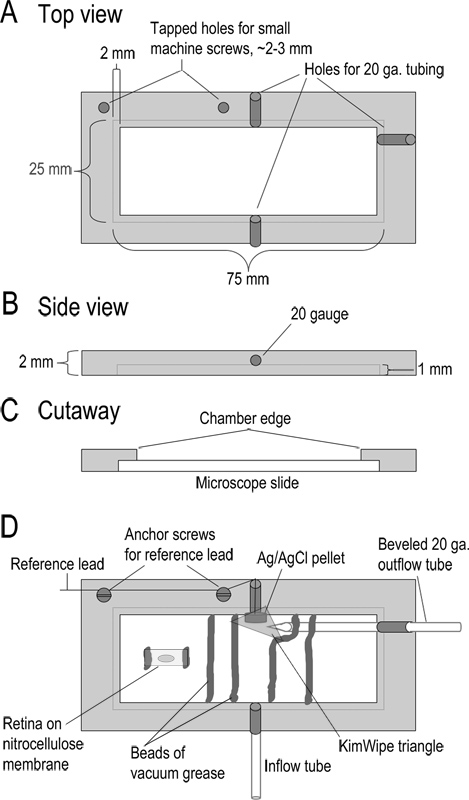
Figure 1. Recording chamber design. (A-C) Top, side, and cutaway views of the recording chamber showing dimensions. The chamber is machined from a 2 mm thick piece of acrylic. (D) Assembled chamber. Superfusate enters the chamber through a 10 cm length of Teflon tubing (inflow tube; type 24LW). Superfusate is removed by applying mild suction to a beveled 20 gauge metal tube at the other side of the chamber. Adjacent to this output tube is a Ag/AgCl pellet reference electrode. The lead from this reference electrode is connected to the reference input of the headstage. A small triangle of KimWipe is placed over the reference electrode to keep it in contact with the solution and regulate solution flow into the outflow tube. The base of the chamber is formed by placing a glass microscope slide into the recessed edges of the chamber. The slide is held in place with a bead of vacuum grease. Strips of nitrocellulose membrane with retinal slices are embedded in beads of vacuum grease that form a channel for solution to flow. Prior to making slices, a 5 x 10 mm piece of nitrocellulose membrane is affixed to the chamber with two small beads of vacuum grease. A piece of the eyecup is placed vitreal side down on this nitrocellulose membrane and lifted away once the retina adheres. Click here to view larger figure.
{kind=link}

Figure 2. Dye-filled cell pairs in vertical slice preparation. A) Images of a rod and synaptically-coupled horizontal cell that were filled with contrasting fluorescent dyes introduced via the patch pipette during simultaneous whole-cell recording. Lucifer yellow (2 mg/ml) was included in the rod pipette solution (yellow) and sulfarhodamine B (1 mg/ml) was included in the horizontal cell pipette solution (purple). Fluorescent images were captured using a spinning disc confocal microscope (Perkin Elmer Ultraview LCI) equipped with a cooled CCD camera (Orca ER) and mounted to the fixed stage microscope (Nikon E600 FN with 60X, 1.0 NA water immersion objective). These images were overlaid on bright field images of the corresponding retinal slices using Adobe Photoshop. B) Images of a cone and synaptically-coupled OFF bipolar cell. The bipolar cell axon terminal ramifies in the outermost (S1) sublamina of the inner plexiform layer near the border with the inner nuclear layer. C) Images of a cone and synaptically-coupled horizontal cell. Note that the cone terminals are considerably larger than the axon terminal of the rod. Although horizontal cells can be identified by the oblong shape of their cell bodies, cell bodies of ON- and OFF-type bipolar cells in the INL cannot be easily distinguished prior to recording. However, one can target displaced cone-driven OFF bipolar cells which have cell bodies in the ONL and can be distinguished from cones by the absence of inner and outer segments. The scale bar is 20 μm.

Figure 3. Light-evoked currents recorded under voltage clamp from four different retinal neurons in response to a bright 500 msec flash of white light. The cone (A), horizontal cell (B), and off bipolar cell (C) all responded to light with an outward current. (D) The on bipolar cell responded to the same light stimulus with an inward current. The baseline noise exhibited by the off bipolar cell (C) reflects a continual release of synaptic vesicles in darkness that diminishes when photoreceptors hyperpolarize in light. Responses from these four cells were obtained in separate recordings using slices prepared under white light. The intensity of the white light stimulus used in these examples produced responses in horizontal and bipolar cells equivalent to 580 nm photon flux of 1 x 105 photons/sec/μm2.

Figure 4. Current-voltage (I-V) relationships of horizontal and off bipolar cells. (A) Top panel, membrane currents evoked by a series of 150 msec voltage steps from -120 to +40 mV applied in 20 mV increments (bottom panel) to a horizontal cell. Horizontal cells typically have a low input resistance and linear or inwardly rectifying I-V relationships. (B) I-V relationship of an off bipolar cell in response to a series of voltage steps. Bipolar cells have a higher input resistance and an outwardly rectifying I-V, due to the activation of voltage-gated potassium currents.

Figure 5. Examples of paired recording data. (A) Recording of the cone voltage-gated calcium current (Cone ICa) in response to a 100 msec step to -10 mV from a holding potential of -70 mV (Cone Vm). Leak and capacitance transients were subtracted using a P/8 leak subtraction protocol. A fast excitatory post-synaptic current (EPSC; HC PSC) was recorded simultaneously from a horizontal cell, showing that these two cells are synaptically coupled. (B) An EPSC was recorded in an off bipolar cell in response to the same stimulus in another cone.
Discussion
The retinal slice preparation has proven very useful for analyzing the circuitry and mechanisms employed by the retina to process visual information. The ability to obtain whole cell recordings simultaneously from pre- and post-synaptic neurons has been particularly helpful in this endeavor. Paired whole cell recordings are much easier to achieve with slices than with flat-mount retina preparations because the different retina layers are exposed. Moreover, because of their large retinal neurons, salamanders have a long history as a retinal preparation and therefore provide a particularly well-characterized model system.
With practice, healthy slices of salamander retina can be prepared regularly. A few key steps can make the difference between success and failure. 1) Make sure the razor blade is mounted on the tissue slicer so that it lays flat against the glass surface and slices cleanly though both the tissue and underlying nitrocellulose membrane. If you have made a clean cut through the nitrocellulose membrane, you should hear a faint click as the razor blade strikes the surface of the glass slide. 2) Make sure the retina has adhered to the nitrocellulose membrane. Otherwise, the retina can float away from the membrane during any step of the procedures. 3) Do not expose cut slices to air, as this will damage many of the superficial cells. 4) Make sure the slices and nitrocellulose membrane lie flat against the glass slide so that the retinal layers are apparent under the dissecting microscope. 5) Balance the rates of superfusate inflow and outflow in order to avoid overflowing the recording chamber. This prevents sudden changes in solution levels, which can cause abrupt tissue movements. 6) Select a healthy pair of cells close to one another. Cells with smooth cytoplasm are healthier than cells with grainy cytoplasm. Cells deeper in the slice are more likely to retain intact synaptic contacts. 7) Make sure the pipette tip has not broken or brushed against other tissue or debris on the way down to the cells. 8) Check the pipette resistance to ensure that it is not clogged with debris or a bubble, both of which can make it difficult to obtain a quality whole-cell recording.
Rather than attaching the retina to nitrocellulose filter paper, some investigators embed retinas in a block of agar and use a vibratome to cut retinal slices. Although we have not tried this approach, Kim et al.11 discuss advantages of both approaches. In their experience, the agar-based approach provides a more consistent yield of flat slices with well-delineated retinal layers but the filter paper-based approach yields healthier photoreceptors.
Rods and cones are responsible for transducing light into changes in membrane potential. With paired recordings, the membrane potential of rods or cones can be manipulated directly and so the ability to generate light responses, while helpful for identifying cell types, may not be essential. We therefore often prepare slices in white light. However, even when prepared under bright illumination, salamander retinal neurons can generate large light responses as illustrated by the responses in Fig. 3. This is partly due to a relatively large reservoir of chromophore in the large outer segment volume but may also reflect the ability of Müller cells to regenerate 11-cis-retinal for cones12. To obtain fully dark-adapted light responses, one can prepare the slices under infrared illumination. For dissections under infrared light, we attach GenIII image intensifiers (Nitemate NAV3, Litton Industries, Tempe, AZ) to the oculars of the dissecting microscope and illuminate the tissue with an infrared LED flashlight. For slicing and other procedures that are not conducted under the dissecting microscope, we employ a head-mounted image intensifier. For placement of the patch pipettes, we visualize slices using an infrared-sensitive CCD camera (e.g. Watec 502H, Watec Inc., Middletown, NY) mounted to the upright, fixed-stage microscope. With these precautions, one can obtain rod responses exhibiting single photon sensitivity6, 13.
One limitation of working in retinal slices is that long cellular processes of large field retinal neurons may lose many of their dendrites during the slicing procedure. Retinal slice preparations are therefore more useful for studying the physiology of cells in which the synaptic contacts involve processes close to the cell body. Amphibian and mammalian retinas share many of the same cell types and utilize similar physiological mechanisms14-16. While salamander retina is a good model for many aspects of mammalian retina, one important difference appears to be the presence of a dedicated rod pathway in mammals that involves contacts of specialized rod bipolar cells onto AII amacrine cells14. An additional limitation of the salamander retina is the small number of genetic tools developed specifically for this species. However, antibodies and shRNA reagents that target well-conserved regions in other species can be used successfully in salamander, as can many small molecule inhibitors and peptide reagents. Additionally, with a few modifications in technique, retinal slices can be prepared from other species in which some of these tools are more readily available.
Beyond its utility for paired whole-cell recording, the salamander retinal slice preparation is also amenable to a variety of other approaches. As discussed above, retinal slices can be used to study light responses in combination with various voltage clamp protocols17. Retinal neurons can also be loaded with fluorescent dyes sensitive to Ca2+, Cl-, or Na+ introduced through the patch pipette or by bath-application15,18-20. A fluorescent peptide that binds to the synaptic ribbon21 can be introduced through the patch pipette and used for imaging the ribbon10 or, when conjugated to fluorescein, for acutely and selectively damaging the ribbon22. We have also used retinal slices in combination with quantum dots to monitor the movements of individual calcium channels at rod and cone synaptic terminals23. Thus, the vertical retinal slice is a versatile experimental preparation for studying basic synaptic mechanisms and the unique processing functions performed at the first synapse in the visual signaling pathway.
Disclosures
No conflicts of interest declared.
Acknowledgements
This work was funded by Research to Prevent Blindness and National Institutes of Health grant EY10542.
Materials
| Name | Company | Catalog Number | Comments |
| Tissue slicer | Stoelting | 51425 | |
| Double edge razor blades | Ted Pella, Inc | 121-6 | |
| Nitrocellulose membranes | Millipore | AAWP02500 | Type AAWP 0.8 mm pore |
| Borosilicate glass pipettes | World Precision Instruments | TW120F-4 | 1.2mm OD 0.95 mm ID |
| Ag/AgCl pellet | Warner | E206 | |
| MicroFil | World Precision Instruments | MF34G-5 | 34 ga. Filling needle, 67 mm long |
References
- Ramòn y Cajal, S., Thorpe, S. A., Glickstein, M. . The Structure of the Retina. , (1972).
- Piccolino, M. Cajal and the retina: a 100-year retrospective. Trends Neurosci. 11, 521-525 (1998).
- Hartline, H. K. The response of single optic nerve fibers of the vertebrate eye to illumination of the retina. Am. J. Physiol. 121, 400-415 (1938).
- Kuffler, S. W. Discharge patterns and functional organization of mammalian retina. J. Neurophysiol. 16, 37-68 (1953).
- Werblin, F. S. Transmission along and between rods in the riger salamander retina. J. Physiol. 280, 449-470 (1978).
- Wu, S. M. Synaptic connections between neurons in living slices of the larval tiger salamander retina. J. Neurosci. Meth. 20, 139-149 (1987).
- Heidelberger, R., Thoreson, W. B., Witkovsky, P. Synaptic transmission at retinal ribbon synapses. Prog. Retin. Eye Res. 24, 682-720 (2005).
- Schmitz, F. The making of synaptic ribbons: how they are built and what they do. Neuroscientist. 15, 611-624 (2009).
- Morgans, C. W., Brown, R. L., Duvoisin, R. M. TRPM1: the endpoint of the mGluR6 signal transduction cascade in retinal ON-bipolar cells. Bioessays. 32, 609-614 (2010).
- Bartoletti, T. M., Babai, N., Thoreson, W. B. Vesicle pool size at the salamander cone ribbon synapse. J. Neurophysiol. 103, 419-423 (2010).
- Kim, M. H., Vickers, E., von Gersdorff, H. Patch-clamp capacitance measurements and Ca2+ imaging at single nerve terminals in retinal slices. J. Vis. Exp. (59), e3345 (2012).
- Wang, J. S., Estevez, M. E., Cornwall, M. C., Kefalov, V. J. Intra-retinal visual cycle required for rapid and complete cone dark adaptation. Nat. Neurosci. 12, 295-302 (2009).
- Thoreson, W. B., Tranchina, D., Witkovsky, P. Kinetics of synaptic transfer from rods and cones to horizontal cells in the salamander retina. Neuroscience. 122, 785-798 (2003).
- Wu, S. M. Synaptic organization of the vertebrate retina: general principles and species-specific variations: the Friedenwald lecture. Invest. Ophthalmol. Vis. Sci. 51, 1263-1274 (2010).
- Babai, N., Thoreson, W. B. Horizontal cell feedback regulates calcium currents and intracellular calcium levels in rod photoreceptors of salamander and mouse retina. J. Physiol. 587, 2353-2364 (2009).
- Babai, N., Morgans, C. W., Thoreson, W. B. Calcium-induced calcium release contributes to synaptic release from mouse rod photoreceptors. Neuroscience. 165, 1447-1456 (2010).
- Thoreson, W. B., Burkhardt, D. A. Contrast encoding in retinal bipolar cells: current vs. voltage. Vis. Neurosci. 20, 19-28 (2003).
- Thoreson, W. B., Bryson, E. J., Rabl, K. Reciprocal interactions between calcium and chloride in rod photoreceptors. J. Neurophysiol. 90, 1747-1753 (2003).
- Cadetti, L., Bryson, E. J., Ciccone, C. A., Rabl, K., Thoreson, W. B. Calcium-induced calcium release in rod photoreceptor terminals boosts synaptic transmission during maintained depolarization. Eur. J. Neurosci. 23, 2983-2990 (2006).
- Luo, J., Boosalis, B. J., Thoreson, W. B., Margalit, E. A comparison of optical and electrophysiological methods for recording retinal ganglion cells during electrical stimulation. Curr. Eye Res. 37, 218-227 (2012).
- Zenisek, D., Horst, N. K., Merrifield, C., Sterling, P., Matthews, G. Visualizing synaptic ribbons in the living cell. J. Neurosci. 24, 9752-9759 (2004).
- Snellman, J., Mehta, B., Babai, N., Bartoletti, T. M., Akmentin, W., Francis, A., Matthews, G., Thoreson, W. B., Zenisek, D. Acute destruction of the synaptic ribbon reveals a role for the ribbon in vesicle priming. Nat. Neurosci. 14, 1135-1141 (2011).
- Mercer, A. J., Chen, M., Thoreson, W. B. Lateral mobility of presynaptic L-type calcium channels at photoreceptor ribbon synapses. J. Neurosci. 31, 4397-4406 (2011).
Reprints and Permissions
Request permission to reuse the text or figures of this JoVE article
Request PermissionThis article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. All rights reserved