
Bu içeriği görüntülemek için JoVE aboneliği gereklidir. Oturum açın veya ücretsiz deneme sürümünü başlatın.
Method Article
Methyltransferases ve Kofaktör Analoglarının ile Nükleik Asitler ve Proteinler Dizi-spesifik Etiketleme
* Bu yazarlar eşit katkıda bulunmuştur
Bu Makalede
Özet
DNA ve proteinler dizi-spesifik afinite ya da DNA ya da protein Methyltransferases sentetik kofaktör analogları kullanılarak flüoresan raportör grup ile etiketlenir. Enzimler, aziridin veya çift aktif kofaktör benzerlerinin kofaktör özelliğine göre bir veya iki adım etiketleme için kullanılmaktadır.
Özet
S -Adenosyl-L-metionin (AdoMet veya SAM) bağlı metiltransferaz (MTase) DNA, RNA, proteinler ve küçük biyomoleküller belirli konumlara AdoMet aktive metil grubu transferini katalize eder. Bu doğal metilasyon reaksiyonu sentetik kofaktör analoglarını kullanarak alkilasyon reaksiyonları de çok çeşitli genişletilebilir. Bir aziridin halkası ile AdoMet reaktif sülfonyum merkezi değiştirilmesi çeşitli DNA MTases ile DNA ile birleştirilebilir kofaktörler yol açar. Bu aziridin kofaktörler adenin parçasının farklı pozisyonlarda raportör grup ile donatılmıştır ve DNA (Smiling DNA) L Abel ing nduced ethyltransferase- S Sequence özgü M kullanılabilir. Tipik bir örnek olarak, DNA MTase M.BseCI ve aziridin kofaktör 6BAz içinde olan 5'-ATCG T-3 'dizisi bir pBR322 plazmid DNA biyotinilasyon için bir protokol eldebir adım. Bir ctivated G grupların aldığı (Mtag) m ethyltransferase-yönettiği T ransfer için kullanılan AdoMet benzerlerinin başka bir sınıf doymamış alkil grupları sonuçları ile aktive edilmiş bir metil grubunun çıkarılması. Uzun yan zincirler sülfonyum merkezi ve doymamış bağ ile aktive olduğundan, bu kofaktörler çift aktif AdoMet analogları olarak adlandırılır. Bu analoglar aziridin kofaktörlerinin gibi DNA MTases için kofaktör olarak işlev, aynı zamanda RNA, protein ve küçük molekül MTases için değil sadece. Bunlar tipik olarak ikinci bir kimyasal aşama raportör grup ile etiketlenmiş olan eşsiz fonksiyonel gruplarla MTase substratların enzimatik modifikasyonu için kullanılır. Bu histon H3 proteinin floresan etiketlemesi için bir protokol örneklenmiştir. Küçük bir propargil grubu tıklama etiketleme ardından histon H3 lizin 4 (H3K4) MTase Set7 / 9 ile proteine kofaktör analog SeAdoYn aktarılırTAMRA azit ile alkynylated histon H3. Kofaktör analoglarıyla MTase aracılı etiketleme tanımlanması ve fonksiyonel çalışma MTase alt tabakalar hem de DNA genotiplemesi ve metilasyon tespiti de dahil olmak üzere pek çok verici uygulama için elverişli bir teknolojidir.
Giriş
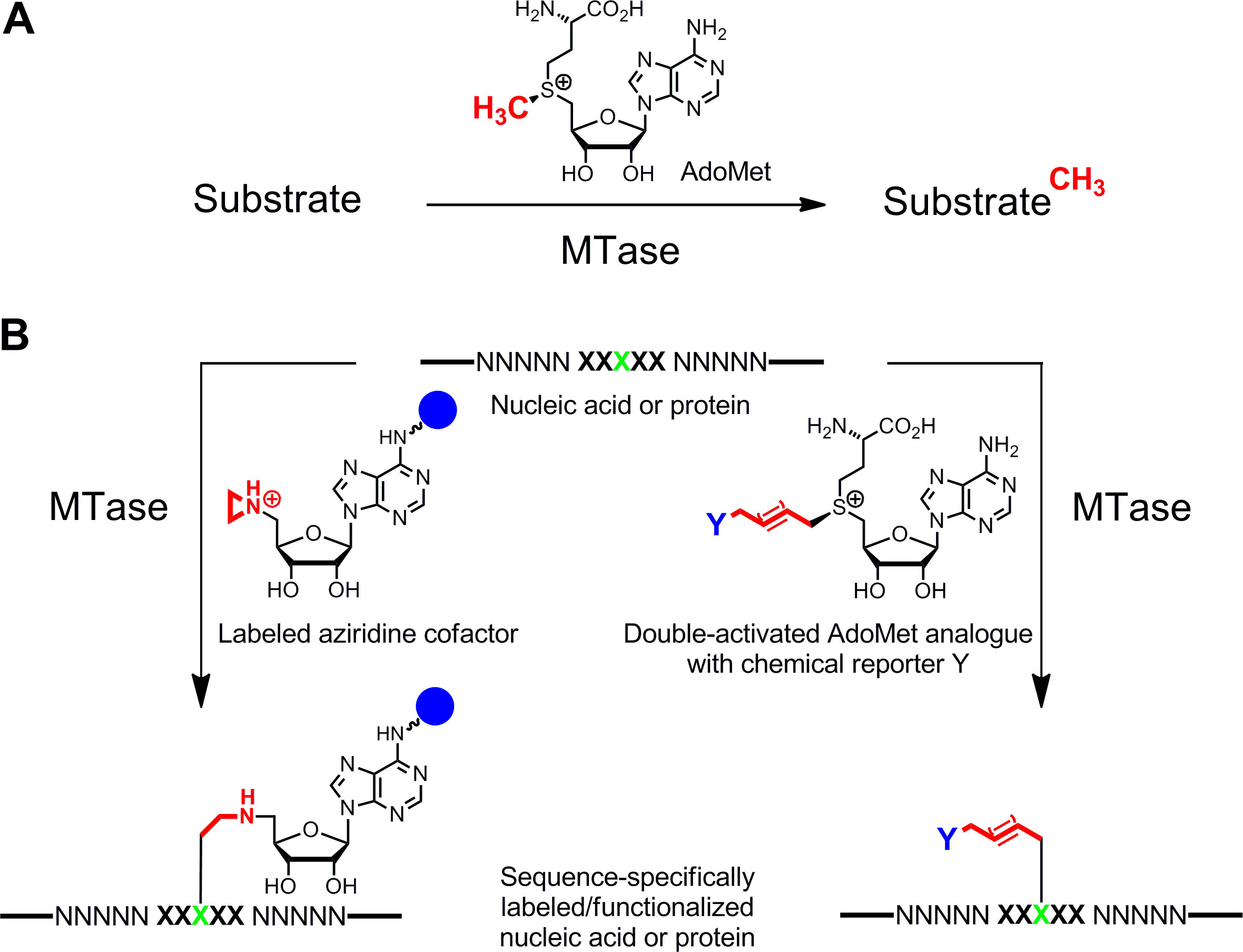
Nükleik asitler ve proteinlerin 1,2 3,4 Özgül etiketleme fonksiyonel karakterizasyonlar, tıbbi teşhis ve (nano) biyoteknoloji için önemli ilgi alanıdır. Burada S -adenosyl-L-metionin (AdoMet veya SAM) bağlı metiltransferaz (MTases) dayanır, bu biyopolimerler için enzimatik bir etiketleme yöntemi sunulmaktadır. Bu enzim sınıfının (EC 2.1.1.), Nükleik asitlerin ve proteinlerin belirli artıkların tek tek nükleofilik pozisyonlar (nitrojen, oksijen, kükürt ve karbon atomu) yönelik olarak ve doğal olarak kofaktör AdoMet (Şekil 1A) 5 aktive metil grubunu aktarır. Buna ek olarak, MTases afinite etiketleri, floroforlar veya diğer etiketler (Şekil 1B) 6 belirli etiketleme için sentetik kofaktör analoglarını kullanabilir. AdoMet analoglarının iki sınıfı geliştirilmiştir: S Sequence özgü M ethyltransferase- I Aziridin kofaktörleri Nduced L abel ing (gülümseyerek) 7 ve A ctivated G grupların aldığı m ethyltransferase-yönettiği T ransfer çift aktive AdoMet analogları (Mtag) 8.

Şekil 1.:. Metiltransferaz (MTases), DNA, RNA, proteinler ve küçük biyomoleküller dahil olmak üzere çeşitli yüzeyler için doğal kofaktör AdoMet (SAM) A. Metil grup transfer nükleik asitlerin ve proteinlerin B. Etiketleme / işlevselleştirme (NNNNN = katalize reaksiyonlar proteinler için RNA, ve amino asitler için temel DNA çiftleri, nükleotitler sentetik kofaktör analogları ile yeşil hedef tortu) ile MTase xxxxx = tanıma sekansı. Bir raportör grubu içeren Aziridin kofaktör (mavi küre)özellikle de hedef tortu (sol) ve iki aktif AdoMet analogları ile birlikte dizi adenin halkası olan bağlı bir ikinci aşamada bioorthogonal Click reaksiyonu ile etiketlenebilir, bir kimyasal raportör Y (sağ) taşıyan uzatılmış alkil zincirlerinin aktarmak yol açar. Bu rakamın büyük halini görmek için lütfen buraya tıklayınız.
{kind=link}
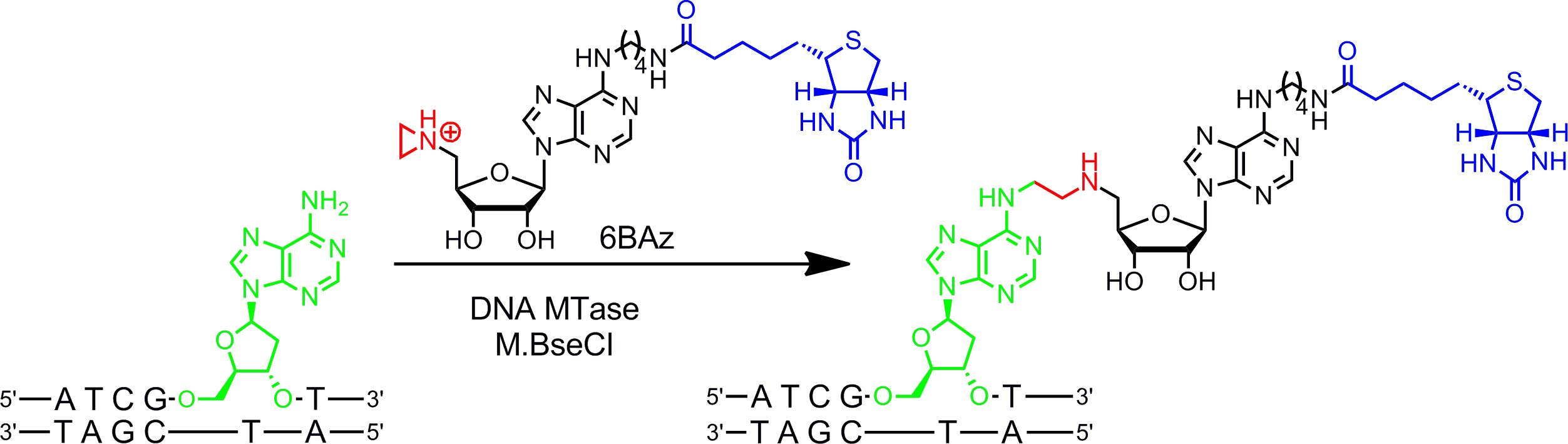
Aziridin kofaktör DNA MTases en iyi çalışır. Bir azot atomu 9 (veya bunun bir N 10,11 -mustard) yerine, sülfonyum merkezi olarak bir reaktif grup ile üç unsurlu bir halka ihtiva etmektedir. Bu nitrojen atomu protonlanma DNA ile bütün kofaktör bağlanması kovalent neden hedef nükleotid nükleofilik saldırı aziridin halkası aktive eder. Adenin halkasına raportör grupları takılarak aziridin kofaktörler (bir aşamada DNA etiket DNA MTases ile kombinasyon halinde de kullanılabilir g> Sol Şekil 1B) 7,12. 15 (adenin halkasının 6 konumuna bağlı biyotin ile aziridin kofaktör) ve Bacillus stearothermophilus'dan adenin özgü DNA MTase (M.BseCI) 16 (Şekil 2 - Bu 6BAz 13 DNA biyotinilasyon için detaylı olarak gösterilmektedir ) DNA'nın Bir adım etiketleme aziridin kofaktörlerinin yoluyla: protokol bölümüne 2 bkz. Haemophilus heamolyticus gelen M.BseCI (5'-ATCG T-3 'tanıma sekansı), yani Thermus aquaticus (M.TaqI, 5'-TCG bir -3 DNA MTases'), ek olarak (M.HhaI, 5 '-G Cı GC-3') ve Spiroplasma (M.SssI gelen 5'-C G-3 '), başarılı bir şekilde 6BAz 17 ile DNA biotinylate için kullanılmıştır. Ayrıca, aziridin kofaktörler tek aşamalı floresan DNA etiketleme 18,19 için kullanılabilecektir.
ontent "fo: keep-together.within-sayfa =" always ">
Şekil 2:. M.BseCI ve 6BAz ile DNA sekansa özel tek-aşamalı biyotinilasyon DNA MTase M.BseCI çift bükümlü DNA dizisi 5'-ATCG T-3 'tanır ve doğal olarak, ikinci adenin amino grubunun metile Kalıntı (yeşil) AdoMet kullanarak. Aziridin kofaktör 6BAz ile reaksiyona ders değiştirilir ve M.BseCI hedef adenin ile biyotin (mavi) dahil olmak üzere tüm kofaktör birleştirilmesi ile spesifik DNA biyotinilasyonunu sırası yol açar. Bu rakamın büyük halini görmek için lütfen buraya tıklayınız.
{kind=link}
Çift aktif AdoMet analogları doymamış yan zincirleri yerine sülfonyum merkezinde bir metil grubunu (Şekil 1B genişletilmiş içeren , sağ) 20. sülfonyum merkezine β-konumunda doymamış çift veya üçlü bağ elektronik konjugatif istikrar ile geçiş devlet içinde olumsuz etkileri sterik dengeler. Sülfonyum merkezi ve doymamış bağ hem de enzimatik transferi için yan zinciri etkinleştirmek için, bu kofaktör çift aktive AdoMet analogları adlandırılmıştır. Tipik haliyle, bir ikinci aşamada 8,21 olarak kemo-seçici etiketleme, amino, alkin ve asit grupları gibi benzersiz kimyasal grupların (kimyasal muhabir) ile yan zincirleri aktarmak için kullanılır. RNA ve protein ek etiketleme sağlayan 28 - Genel olarak, iki aktif AdoMet analogları RNA MTases 22,23 DNA MTases 8,20,21 için değil, aynı zamanda yardımcı faktörler ve protein MTases 24 gibi yalnızca işlev görebilir. Ancak, genişletilmiş yan zincirler sterik daha bir metil grubu daha zorlu ve mühendislik sık sık bir protein MTase aktif siteleri büyütme vardırtr verimli aktarım hızları elde etmek için gerekli. Bakır kimyasal değişiklikler aşağıdaki için enzimatik transferi sırasında geçiş devletin 1. Sabitleme ve 2. reaktif kolu: Bu sorunun başka bir çözüm, küçük bir propargil terminali alkin iki fonksiyonu vardır grup (üç karbon) ile bir AdoMet analog kullanmak için katalize azid-alkin sikloadisyonu (CuAAC) kimyasını tıklayın. Bu Oluşan proparjilik AdoMet analog 29 nötr veya hafif bazik koşullar altında ve sadece sınırlı kullanımı oldukça kararsız olduğu ortaya çıktı. Bu dezavantajı selenyum ile kükürt atomu yerine sabit olabilir. Elde edilen kofaktör 5 '- [(Se) [(3 S) -3-amino-3-karboksipropil] prop-2-ynylselenonio] -5'-deoksiadenozin (SeAdoYn, Şekil 3) ile, vahşi tip DNA tarafından kabul edilir, RNA, ve protein MTases 30 - birçok durumda protein mühendisliği ihtiyacını ortadan kaldırmaz 32. Bu floresan pro ile tarif edilmektedir histon H3 lizin ile tein etiketleme 4 (H3K4) MTase Set7 / 9 33 (: çift aktif kofaktör aracılığıyla İki aşamalı protein etiketleme Şekil 3, protokol bölümüne 3).

Şekil 3:. Set7 / 9 histon H3 diziye özel iki-aşamalı floresan etiketleme, SeAdoYn ve TAMRA azid proteini MTase Set7 / 9 doğal AdoMet ile histon H3 lizin 4 amino grubunun (H3K4, yeşil) bileşi. Çift aktif kofaktör ile SeAdoYn MTase lisin kalıntıya küçük bir propargil grubu (kırmızı) aktarır. bağlı uç üçlü bağ daha sonra seçici azid türevlendirilmemiş TAMRA (tetrametilrodamin, mavi) florofor ile bioorthogonal click reaksiyonunun (bakır ile katalize edilen azit-alkin siklo, CuAAC) değiştirilir.yük / 52.014 / 52014fig3highres.jpg "target =" _ blank "> bu rakamın daha büyük bir versiyonunu görmek için burayı tıklayınız.
Protokol
1. Genel Talimatlar
- Mağaza aziridin kofaktör (DMSO içinde) 6BAz ve protein MTase -80 ° C'de Set7 / 9 ve çift aktif kofaktör SeAdoYn ve DNA MTase M.BseCI -20 ° C'de (% 50 gliserol) da dahil olmak üzere diğer tüm reaktif maddeler.
- Iyonu giderilmiş su içinde söndürme katsayısına ε 269nm (6BAz) = 16,000 cm -1 M 1 ve 260 nm ε (SeAdoYn) = 15.400 cm -1 M-1 kullanılarak UV / Vis spektroskopisi yoluyla 6BAz ve SeAdoYn konsantrasyonunu belirlemek. Tükenme katsayısı varsa 280 nm'de absorpsiyon yoluyla doğrudan, Bradford tahlili ile MTases konsantrasyonunu belirlemek veya.
- Enzim aktivitesinin kaybını önlemek için yoğun pipetle veya vorteks kabarcıkları oluşturarak önlemek için deneyin. Bunun yerine, yavaşça yukarı ve aşağı pipetleme karıştırın.
- DMSO içindeki stok solüsyonlarından aziridin kofaktörleri eklerken deneyde nihai DMSO konsantrasyonu daha az olduğundan emin olmak% 5 daha. Her DNA ile spesifik olmayan reaksiyonları önlemek için, deney tamponu içerisinde 10 mM magnezyum tuzlarıdır.
- Asidik stok çözümleri çift aktif kofaktörlerine eklerken pH değişiklikleri önlemek ve tahlil çözeltinin pH değeri önemli bir değişiklik olmadığından emin olmak için küçük hacimli (yüksek konsantre stok çözümleri) kullanın. , Gerekli bakır iyonu kompleks ile click reaksiyonunun etkilediği için bu deney tamponu içinde tioller, örneğin β-merkaptoetanol veya ditiyotreitol (DTT), kaçının.
Aziridin kofaktörlerinin yoluyla DNA'nın 2. Bir adım Etiketleme
- Sekans spesifik Metiltransferaz Bağlı Etiket ing M.BseCI DNA MTase ve aziridin kofaktör 6BAz ile plazmid DNA (gülümser).
- 20 ° C'de kofaktör çözeltisi çözülme ve buz üzerinde tepkime karışımları hazırlamak.
- Denemesine ek olarak herhangi bir spesifik olmayan modifikasyonlar ve An ve # görselleştirmek için bir "Kofaktör" kontrolünü gerçekleştirmek8220; Enzim "kontrol, MTase hazırlık, doğal kofaktör AdoMet serbest olduğundan emin olmak için.
- 10x değişiklik tamponu deney karışımı 2 ul için (100 mM Tris-HCl, içeren 100 mM MgCl2, 20 mM β-merkaptoetanol, pH 7.4) içinde, pBR322 (0.5 ug / ul), 10 eq 2 ul. DNA tanıma sekansı (pBR322 1 tanıma sekansı) ve 20 ul toplam hacim içinde 60 uM nihai konsantrasyona kadar aziridin kofaktör 6BAz başına M.BseCI. Kofaktör ve DNA MTase son ekleyin.
NOT: β-merkaptoetanol, toksik korozif ve çevreye zarar vermektedir. - "Kofaktör" kontrol için yerine 6BAz deiyonize su ekleyin yerine M.BseCI ve "enzim" kontrolü için deiyonize su ekleyin.
- Yavaşça yukarı ve aşağı pipetleme çözüm karıştırın.
- 1 saat boyunca 55 ° C'de inkübe edin.
- Santrifüj kısaca tüplerinin altındaki tüm sıvıyı toplamak için.
- Kısıtlama-modifikasyon tahlil DNA modifikasyonu doğrulamak için.
- 80 ul deiyonize su ve 3.3 ul (100 mM Tris-HCl, 50 mM MgCl2, 1 M NaCl, 1 mg / ml sığır serumu albümini, pH 8.0 ihtiva eder) 10 ul 10x R.TaqI tampon karıştırılmasıyla bir çözelti hazırlayın kısıtlama endonükleaz Thermus aquaticus (artma) (R.TaqI, 10 U / ul). Son adımda artma eklemek için emin olun.
- 2.1.7 Her tüpe R Taql tampon 10x 2 ul ve yukarıdaki (2.2.1) çözelti 28 ul ekle.
- Yavaşça yukarı ve aşağı pipetleme çözüm karıştırın.
- 30 dakika boyunca 65 ° C'de inkübe edin.
- Santrifüj kısaca tüplerinin altındaki tüm sıvıyı toplamak için.
- Streptavidin ile Elektromobilite kayma deneyi (EMSA) fonksiyonel değişiklik doğrulamak için.
- Her bir tüp (2.2.5) 25 ul çıkarın ve streptavidin m göre bir streptavidin çözeltisi (1 mM, 2.4 ulonomer 100 mM Na 2 HPO 4 içeren streptavidin tamponu, 100 mM NaCI, pH 7.5; Toplam biyotin 4 eşdeğer). Kalan tüpler streptavidin tamponu 2.4 ul ekleyin.
- 1 saat boyunca 37 ° C'de tüm tüpler inkübe edin.
- Agaroz jel elektroforez ile analizi.
- Her tüpe 6x yükleme tamponu (% 0.25 bromofenol mavisi,% 30 gliserol) içinde 5 ul ekle.
- Yavaşça çözüm karıştırın.
- Yük (bir 10,000X stok çözeltisinden 1x GelRed içeren 0.5x TBE tamponu içinde% 1 agaroz), bir agaroz jeli, kuyu içine her bir numune için 10 ul.
- Yaklaşık 80 V ile 0.5x TBE tamponu jel çalıştırın. 1 saat.
- Bir filtre (540 ± 50 nm) ile donatılmış bir CCD kamera ile UV tablosu (312 nm) DNA bantları gözünüzde canlandırın.
NOT: UV ışık Gözleri ve cildi zarar vermektedir.
Çift Aktif kofaktörlerinin aracılığıyla 3. İki aşamalı Protein Etiketleme
- Methyltransfhiston H3 lizin 4 etiketleme (modifikasyon adım) için Set7 / 9 ve çift-aktif kofaktör SeAdoYn ile Aktif Gruplar (Mtag) Devri silmek-yöneltti.
- Bileşenleri çözülme ve buz üzerinde reaksiyon karışımları hazırlayın. NOT: Her zaman SeAdoYn bozulmasını önlemek için soğutulmuş tutun.
- Denemesine ek olarak herhangi bir spesifik olmayan değişiklikler görselleştirmek için bir "Kofaktör" kontrol gerçekleştirmek ve bir "enzim" kontrol, flüoresan probun spesifik olmayan reaksiyonlar ortaya çıkarmaktır.
- Bir deney çözeltisi (20 ul) modifikasyon tamponu (50 mM Tris-HCl,% 5 gliserol, pH 8.5), 10 uM histon H3, uM Set7 / 9 10 ve 600 uM SeAdoYn (selenyum hem de epimerler karışımı) ihtiva eden hazırlayın. Son adımda daha sonra maddeyi ve MTase ekleyin.
- "Kofaktör" kontrol için 3.1.3 gibi bir tahlil çözelti hazırlamak ve sentetik kofaktör ile rekabet etmek 60 mM AdoMet ekleyin. "Enzim" kontrol için yerine SeAdoYn deiyonize su ekleyin.
- Yavaşça yukarı ve aşağı pipetleme çözüm karıştırın. PH şerit (- 10 pH aralığı 5) üst alanında her solüsyonun 1 ul ekleyerek pH değerini kontrol edin.
- 2 saat süre ile 37 ° C'de inkübe edilir.
- Bu arada, bir% 12 SDS-poliakrilamid jel hazırlamak (çalışma jelde: 357 mM Bis-Tris pH 6,5-6,8,% 0.1 (a / h) APS ağırlık,% 0.04 (h / h) TEMED ve% 12 akrilamid / bisakrilamid 37.5: 1 ; yükleme jel: 357 mM Bis-Tris 6,5-6,8 pH,% 0.1 APS (w / v),% 0.04 (h / h) TEMED,% 5 akrilamid / bisakrilamid 37.5: 1).
NOT: Akrilamid / bisakrilamid tehlikeli toksik ve sağlıktır. Bu işlem sırasında eldiven giyin.
- Bakır-katalize azid-alkin sikloadisyonu (CuAAC) (etiketleme adım) üzerinden histon H3 alkinylated lizin 4 Kimyasal etiketleme.
- Sadece modifikasyonu tepkime sonuna kadar 3 mM CuSO 4, 3 mM tris (3-hidroksipropil-triazolilmetil) amin (THPTA), 250 mM sodyum askorbat ve 6 mM TAMRA azit içeren 5x tıklama karışımı hazırlamak20 ul toplam hacim.
- CuAAC başlatmak ve değiştirme reaksiyonu durdurmak için her bir tüpe, taze hazırlanmış 5x tıklama karışımı 5 ul ekle.
- Yukarı ve aşağı pipetleme hafifçe karıştırın.
- Floroforun fotoğraf beyazlatma önlemek için ışık alüminyum folyo ile tüm tüpleri koruyun.
- 1 saat boyunca 20 ° C'de inkübe edilir.
- Protein yağış ücretsiz TAMRA floroforun fazlalığını ortadan kaldırmak için.
- 34 - bedava TAMRA floroforun yoğun olarak-jel floresan floresan etiketli histon H3 outshining önlemek için, proteinler (3.3.4 3.3.2) yağış fazlalığı floroforu çıkarın.
- 75 ul metanol, 18.8 ul kloroform ve kısa bir süre için her eklemeden sonra her tüp ve vorteks deiyonize su 50 ul ekle. 5 dakika boyunca 16,000 x g'de santrifüjleyin. Protein içeren arayüz katmanı, bozmadan üst faz çıkarın.
- Kalan faz 56.3 ul Metanol In 5 dakika boyunca 16.000 xg'de her tüp, girdap ve santrifüj protein pelet. Süpernatantı. Pelet yıkamak için bu adımı tekrarlayın.
- Tüy bırakmayan bir doku ile açık tüpler Kapak ve 15 saat kurumaya bırakın - 30 dakika.
- SDS PAGE yoluyla analizi.
- (20 ul SDS yükleme tamponu (50 mM Tris-HCI,% 2.5 (ağ / hac) SDS,% 10 (h / h) gliserol, 320 mM β-merkaptoetanol ve% 0.05 3.3.4 çökeltilir proteinleri çözülür w / V) bromfenol mavisi, pH 6.8). Tamamen bir pipet ile tüplerin duvarları durulama pelet çözmek için emin olun.
- 10 dakika boyunca 95 ° C'de inkübe edin ve bunları 20 ° C'ye kadar soğumaya bırakın.
- Santrifüj kısaca tüplerinin altındaki tüm sıvıyı toplamak için.
- Bir SDS poliakrilamid jeli (3.1.7) oyuklarına Her numunenin bütün miktarı yerleştirin. 50 mM MOPS, 50 mM Tris-X (Tris-bazlı), 5 mM EDTA,% 0.1 kullanarak (a / h) elektroforez için tampon olarak SDS aktarıldı.
- Yaklaşık 120 V ile jel çalıştırın. 90 dakika.
- Bir filtre (50 nm ± nm 540) ile donatılmış bir CCD kamera ile UV tablosu (312 nm) üzerinde-jel floresan gözünüzde canlandırın.
NOT: UV ışık Gözleri ve cildi zarar vermektedir.
Sonuçlar
Aziridin kofaktörlerinin yoluyla DNA'nın Bir adım Etiketleme
Bu örnek, reaksiyon, çift kollu bir 5'-ATCG T-3 'dizisi içindeki ikinci adenin kalıntısı değiştirir ve pBR322 plazmidi (Şekil 4A) ilgili bir tanıma sitesi vardır DNA MTase M.BseCI ile gerçekleştirilir. Plazmid etiketleme test etmek için, pBR322 kısıtlama endonükleaz (artma) R.TaqI (5'-TCGA-3 ') ile tehdit edildi. R.TaqI M.BseCI sitede yer alan bunlar...
Tartışmalar
DNA MTases ve aziridin cofactors (gülümseyen DNA) ile DNA Bir adım etiketleme sağlam bir yöntemdir, ancak deney planlarken bazı yönleri dikkate alınmalıdır.
Aziridin kofaktör: M.BseCI DNA etiketleme için 6BAz konsantrasyonu 60 uM idi. Diğer DNA MTases kullanırken kofaktör konsantrasyonu düşük 20 gibi uM DNA MTase M.TaqI 19 kullanılmıştır, örneğin konsantrasyonları optimize edilmelidir. Düşük 6BAz konsantrasyonları streptavidin (tahlil...
Açıklamalar
The authors disclose the following competing financial interest: E.W. is inventor on related patents.
Teşekkürler
The authors thank Kerstin Glensk for preparing the MTases M.BseCI and Set7/9 and gratefully acknowledge funding by the Excellence Initiative of the German Federal and State Governments and RWTH Aachen University. The authors are happy to provide 6BAz and SeAdoYn or other cofactor analogues for collaborative research.
Malzemeler
| Name | Company | Catalog Number | Comments |
| 6BAz | Synthesized according to Weinhold et al., Patent number US 8,129,106, published March 6, 2012. | ||
| β-Mercaptoethanol | Serva | 28625 | |
| Acetic acid | Fisher Scientific | 10304980 | |
| Acrylamide/Bis Solution, 37.5:1 | Serva | 10688 | |
| UltraPure Agarose | Invitrogen | 16500100 | |
| Ammonium persulfate (APS) | Serva | 13375 | |
| Bis-Tris | Gerbu | 1304 | |
| Boric acid | Gerbu | 1115 | |
| Bromophenol blue Na salt | Serva | 15375 | |
| Copper(II) sulfate | Aldrich | C1297 | |
| Chloroform | Fisher Scientific | 10020090 | |
| Coomassie Brilliant Blue | Serva | 17525 | |
| EDTA disodium salt | Gerbu | 1034 | |
| Ethanol | Merck | 100983 | |
| GelRed (10,000x in water) | Biotium | 41003 | |
| Glycerol (99.5%) | Gerbu | 2006 | |
| FastRuler Low Range DNA Ladder | Thermo Scientific | SM1103 | |
| Histone H3 | Expression plasmid obtained from Dr. Philipp Voigt and Prof. Danny Reinberg; expression and isolation according to T. J. Richmond et al., J. Mol. Biol. 1997, 272, 301-311. | ||
| M.BseCI | Expression plasmid obtained from Dr. Michael Kokkinidis; expression and isolation according to Kapetaniou et al., Acta Cryst. 2006, F63, 12-14. | ||
| Methanol | Fisher Scientific | 10675112 | |
| Magnesiumchloride hexahydrate | J.T. Baker | 4003 | |
| MOPS | Gerbu | 1081 | |
| Sodium chloride | Gerbu | 1112 | |
| pH strip (Neutralit) | Merck | 1,095,330,001 | |
| pBR322 | Thermo Scientific | SD0041 | |
| R.TaqI (10 u/µl) | Thermo Scientific | ER0671 | |
| SeAdoYn | Synthesized according to Willnow et al., ChemBioChem 2012, 13, 1167-1173. | ||
| Set7/9 | Expression plasmid obtained from Prof. Danny Reinberg, expression and isolation according to D. Reinberg et al., Genes Dev.2002, 16, 479-489. | ||
| Streptavidin | Gerbu | 3058 | |
| (+)-Sodium L-ascorbate | Sigma Life Science | A7631 | |
| SDS Granular | Gerbu | 1833 | |
| di-Sodium hydrogenphosphate | Merck | 106,586 | |
| TAMRA azide | Synthesized according to reference 30: Willnow et al., ChemBioChem 2012, 13, 1167-1173. | ||
| TaqI buffer (10x) | Thermo Scientific | B28 | |
| N,N,N',N'-Tetramethylethylenediamine (TEMED) | Acros Organics | 42058 | |
| Tris-HCl | Gerbu | 1028 | |
| Tris-X (TRIS-base) | Gerbu | 1018 | |
| Tris(3-hydroxypropyltriazolyl-methyl)amine (THPTA) | Sigma-Aldrich | 762342 |
Referanslar
- Gottfried, A., Weinhold, E. Sequence-specific covalent labelling of DNA. Biochem. Soc. Trans. 39, 623-628 (2011).
- Zohar, H., Muller, S. J. Labeling DNA for single-molecule experiments: methods of labeling internal specific sequences on double-stranded DNA. Nanoscale. 3, 3027-3039 (2011).
- Hinner, M. J., Johnsson, K. How to obtain labeled proteins and what to do with them. Curr. Opin. Biotechnol. 21, 766-776 (2010).
- Wua, Y. -. W., Goody, R. S. Probing protein function by chemical modification. J. Pept. Sci. 16, 514-523 (2010).
- Struck, A. -. W., Thompson, M. L., Wong, L. S., Micklefield, J. S-Adenosyl-methionine-dependent methyltransferases: Highly versatile enzymes in biocatalysis, biosynthesis and other biotechnological applications. ChemBioChem. 13, 2642-2655 (2012).
- Klimasauskas, S., Weinhold, E. A new tool for biotechnology: AdoMet-dependent methyltransferases. Trends Biotechnol. 25, 99-104 (2007).
- Pljevaljcic, G., Schmidt, F., Weinhold, E. Sequence-specific Methyltransferase-Induced Labeling of DNA (SMILing DNA). ChemBioChem. 5, 265-269 (2004).
- Lukinavicius, G., Lapiene, V., Stasevskij, Z., Dalhoff, C., Weinhold, E., Klimasauskas, S. Targeted labeling of DNA by methyltransferase-directed Transfer of Activated Groups (mTAG). J. Am. Chem. Soc. 129, 2758-2759 (1021).
- Pignot, M., Siethoff, C., Linscheid, M., Weinhold, E. Coupling of a nucleoside with DNA by a methyltransferase. Angew. Chem. Int. Ed. 37, 2888-2891 (1998).
- Weller, R. L., Rajski, S. R. Design, synthesis, and preliminary biological evaluation of a DNA methyltransferase-directed alkylating agent. ChemBioChem. 7, 243-245 (2006).
- Du, Y., Hendrick, C. E., Frye, K. S., Comstock, L. R. Fluorescent DNA Labeling by N-Mustard Analogues of S-adenosyl-l-methionine. ChemBioChem. 13, 2225-2233 (2012).
- Pljevaljcic, G., Schmidt, F., Scheidig, A. J., Lurz, R., Weinhold, E. Quantitative labeling of long plasmid DNA with nanometer precision. ChemBioChem. 8, 1516-1519 (1002).
- Wilkinson, S., et al. Molecular scale architecture: engineered three- and four-way junctions. Bioconjugate Chem. 19, 470-475 (2008).
- Braun, G., et al. Enzyme-directed positioning of nanoparticles on large DNA templates. Bioconjugate Chem. 19, 476-479 (2008).
- Kim, S., et al. Enzymatically incorporated genomic tags for optical mapping of DNA binding proteins. Chem. Int. Ed. 51, 3578-3581 (2012).
- Rina, M., Bouriotis, V. Cloning purification and characterization of the BseCI DNA methyltransferase from Bacillus stearothermophilus. Gene. 133, 91-94 (1993).
- Weinhold, E., Meier, T., Düfel, H., Markert-Hahn, C., Schmuck, R. Sequence-specific detection of methylation in biomolecules. US Patent. , (2012).
- Pljevaljcic, G., Pignot, M., Weinhold, E. Design of a new fluorescent cofactor for DNA methyltransferases and sequence-specific labeling of DNA. J. Am. Chem. Soc. 125, 3492-3410 (2003).
- Schmidt, F. H. -. G., Hüben, M., Gider, B., Renault, F., Teulade-Fichou, M. -. P., Weinhold, E. Sequence-specific Methyltransferase-Induced Labelling (SMILing) of plasmid DNA for studying cell transfection. Bioorg. Med. Chem. 16, 40-48 (2008).
- Dalhoff, C., Lukinavicius, G., Klimasauskas, S., Weinhold, E. Direct transfer of extended groups from synthetic cofactors by DNA methyltransferases. Nat. Chem. Biol. 2, 31-32 (2006).
- Lukinavicius, G., Tomkuviene, M., Masevicius, V., Klimasauskas, S. Enhanced chemical stability of AdoMet analogues for improved methyltransferase-directed labeling of DNA. ACS Chem. Biol. 8, 1134-1139 (2013).
- Motorin, Y., et al. Expanding the chemical scope of RNA:methyltransferases to site-specific alkynylation of RNA for click labeling. Nucleic Acids Res. 39, 1943-1952 (1943).
- Schulz, D., Holstein, J. M., Rentmeister, A. A chemo-enzymatic approach for site-specific modification of the RNA cap. Angew. Chem. Int. Ed. 52, 7874-7878 (2013).
- Peters, W., et al. Enzymatic site-specific functionalization of protein methyltransferase substrates with alkynes for click labeling. Angew. Chem. Int. Ed. 49, 5170-5173 (2010).
- Islam, K., Zheng, W., Yu, H., Deng, H., Luo, M. Expanding cofactor repertoire of protein lysine methyltransferase for substrate labeling. ACS Chem. Biol. 6, 679-684 (2011).
- Wang, R., Zheng, W., Yu, H., Deng, H., Luo, M. Labeling substrates of protein arginine methyltransferase with engineered enzymes and matched S-adenosyl-l-methionine analogues. J. Am. Chem. Soc. 133, 7648-7651 (2011).
- Islam, K., et al. Bioorthogonal profiling of protein methylation using azido derivative of S-adenosyl-l-methionine. J. Am. Chem. Soc. 134, 5909-5915 (2012).
- Islam, K., et al. Defining efficient enzyme-cofactor pairs for bioorthogonal profiling of protein methylation. Proc. Natl. Acad. Sci. U.S.A. 110, 16778-16783 (2013).
- Binda, O., Boyce, M., Rush, J. S., Palaniappan, K. K., Bertozzi, C. R., Gozani, O. A chemical method for labeling lysine methyltransferase substrates. ChemBioChem. 12, 330-334 (2011).
- Willnow, S., Martin, M., Lüscher, B., Weinhold, E. A selenium-based click AdoMet analogue for versatile substrate labeling with wild-type protein methyltransferases. ChemBioChem. 13, 1167-1173 (2012).
- Bothwell, I. R., et al. Se-Adenosyl-l-selenomethionine cofactor analogue as a reporter of protein methylation. J. Am. Chem. Soc. 134, 14905-14912 (2012).
- Tomkuviene, M., Clouet-d’Orval, B., Cerniauskas, I., Weinhold, E., Klimasauskas, S. Programmable sequence-specific click-labeling of RNA using archaeal box C/D RNP methyltransferases. Nucleic Acids Res. 40, 6765-6773 (2012).
- Nishioka, K., et al. Set9, a novel histone H3 methyltransferase that facilitates transcription by precluding histone tail modifications required for heterochromatin formation. Genes Dev. 16, 479-489 (2002).
- Clark, P. M., et al. Direct in-gel fluorescence detection and cellular imaging of O-GlcNAc-modified proteins. J. Am. Chem. Soc. 130, (2008).
- Lukinavicius, G., Lapinaite, A., Urbanaviciute, G., Gerasimaite, R., Klimasauskas, S. Engineering the DNA cytosine-5 methyltransferase reaction for sequence-specific labeling of DNA. Nucleic Acids Res. 40, 11594-11602 (2012).
- Neely, R. K., Dedecker, P., Hotta, J., Urbanaviciute, G., Klimasauskas, S., Hofkens, J. DNA fluorocode: A single molecule, optical map of DNA with nanometre resolution. Chem. Sci. 1, 453-460 (2010).
- Roberts, R. J., Vincze, T., Posfai, J., Macelis, D. REBASE-a database for DNA restriction and modification: enzymes, genes and genomes. Nucleic Acids Res. 38, 234-236 (2010).
- Petrossian, T. C., Clarke, S. G. Uncovering the human methyltransferasome. Mol. Cell. Proteomics. 10, 1-12 (2011).
- Kriukiene, E., et al. DNA unmethylome profiling by covalent capture of CpG sites. Nat. Commun. 4, 2190 (2013).
- Wang, R., et al. Profiling genome-wide chromatin methylation with engineered posttranslation apparatus within living cells. J. Am. Chem. Soc. 135, 1048-1056 (2013).
- Zhang, C., Weller, R. L., Thorson, J. S., Rajski, S. R. Natural product diversification using a non-natural cofactor analogue of S-adenosyl-l-methionine. J. Am. Chem. Soc. 128, 2760-2761 (2006).
- Stecher, H., et al. Biocatalytic Fiedel-Crafts alkylation using non-natural cofactors. Angew. Chem. Int. Ed. 48, 9546-9548 (2009).
- Lee, B. W. K., Sun, H. G., Zang, T., Kim, B. J., Alfaro, J. F., Zhou, Z. S. Enzyme-catalyzed transfer of a ketone group from an S-adenosylmethionine analogue: A tool for the functional analysis of methyltransferases. J. Am. Chem. Soc. 132, 3642-3643 (2010).
- Winter, J. M., et al. Expanding the structural diversity of polyketides by exploring the cofactor tolerance of an inline methyltransferase domain. Org. Lett. 15, 3774-3777 (2013).
Yeniden Basımlar ve İzinler
Bu JoVE makalesinin metnini veya resimlerini yeniden kullanma izni talebi
Izin talebiThis article has been published
Video Coming Soon
JoVE Hakkında
Telif Hakkı © 2020 MyJove Corporation. Tüm hakları saklıdır