
Method Article
RRNA 2'-O-Metilasyonun DNAzyme bağımlı analizi
Bu Makalede
Özet
Burada RNA DNAzyme bağımlı dekolte için bir protokol salıyoruz. Bu, RNA 2'-O-metilasyonun hızlı ve yere bağlı analizini sağlar. Bu yaklaşım, snoRNA aktivitesinin ön veya önemli değerlendirilmesi için kullanılabilir.
Özet
Kılavuz kutusu C / D küçük nükleolar RNA (snoRNA) ribozomal ve küçük nükleer RNA 2'-O-metilasyon katalize. Ancak, yüksek ökaryotlarda çok sayıda snoRNA diğer RNA türlerini promiscuously tanıyabilir ve 2'-O-metilat birden fazla hedef. Burada, DNAzymes adı verilen kısa DNA oligonükleotidlerini kullanan köklü bir yöntem kullanarak siteye özgü 2'-O-metilasyonun hızlı ve pahalı olmayan analizi için adım adım kılavuz salıyoruz. Bu DNA parçaları, RNA'yı belirli konsensüs konumlarında yönlendiren katalitik dizilerin yanı sıra DNAzyme'yi RNA hedeflerine yönlendiren değişken homoloji kollarını içerir. DNAzyme aktivitesi RNA'daki dekolte bölgesine bitişik nükleotitin 2-'O-metilasyonile inhibe edilir. Böylece, DNAzymes, sadece yarık dizisinin konsensüs ile sınırlı, snoRNA aracılı RNA 2'-O-metilasyon hızlı analizi için mükemmel araçlardır. SnoRNA snR13- ve snR47-güdümlü 2'-O-metilasyon 25S ribosomal RNA Saccharomyces cerevisiae tekniğin basitliğini göstermek ve DNAzyme bağımlı test için ayrıntılı bir protokol sağlamak için analiz.
Giriş
RNA modifikasyonları gen ekspresyonunun düzenlenmesinde önemli rol oynar. RNA 2'-O-metilasyon ve psödoürilasyon, sırasıyla kutu C / D ve kutu H / ACA küçük nükleolar RNA (snoRNA) tarafından yönlendirilir, bozulmadan RNA korumak ve onların yüksek sıralı yapıları stabilize1,2,3 . SnoRNA hedefleri ağırlıklı olarak ribozomal RNA (rRNA) ve küçük nükleer RNA'larda (snRNA) tanımlanmıştır. Ancak, yüksek ökaryotlarda, atanmış işlevleri olmayan yüzlerce snoRNA vardır ve bazıları birden fazla RNA1,4,5,6,7'yitanıyabilir. Bu nedenle, snoRNA güdümlü modifikasyonların tanımlanması ve analizine olanak sağlayan yöntemler, hücresel süreçleri yöneten mekanizmaların ortaya çıkarılmasında önemli araçlardır.
Bir kutu C / D snoRNA güdümlü putatif 2'-O-metilasyon sitesi biyoinformatik olarak tespit edilebilir ve deneysel olarak birçok teknik tarafından teyit edilebilir, RNase H-yönettiği dekolte dahil, ya da site-özel ve genom çapında yöntemler, ters transkripsiyon istihdam düşük nükleotitlerde (dNTPs) konsantrasyon8,9,10,11. Bu teknikler çok hassas ama aynı zamanda zahmetli ve pahalı, bu nedenle, ilk veya hızlı test için uygun olmayabilir. 2'-O-metilasyon bölgelerini tanımlamak için en basit ve düşük maliyetli yöntemlerden biri DNAzyme bağımlı RNA dekolte12. DNAzymes belirli pozisyonlarda RNA ennükleyolitik dekolte yeteneğine sahip kısa, tek iplikli ve katalitik aktif DNA molekülleridir. Watson-Crick baz eşleştirmesi ile RNA hedefine hibridize etmek üzere tasarlanmış değişken dizilerden oluşan korunmuş ve katalitik olarak aktif çekirdek sırası ve 5' ve 3' bağlayıcı kollardanoluşurlar (Şekil 1). Böylece, 5' ve 3' kollar katalitik sırayı belirli RNA bölgesine sunar. DNAzyme bağımlı dekolte dekolte dekolte doğrudan dekolte sitenin yukarı sında konumlandırılmış nükleotit 2'-O-metilasyon tarafından inhibe edilir12,13. Bu putatif veya bilinen RNA 2'-O-metilasyon sitelerinin analizi için DNAzymes çok pratik araçlar yapar.
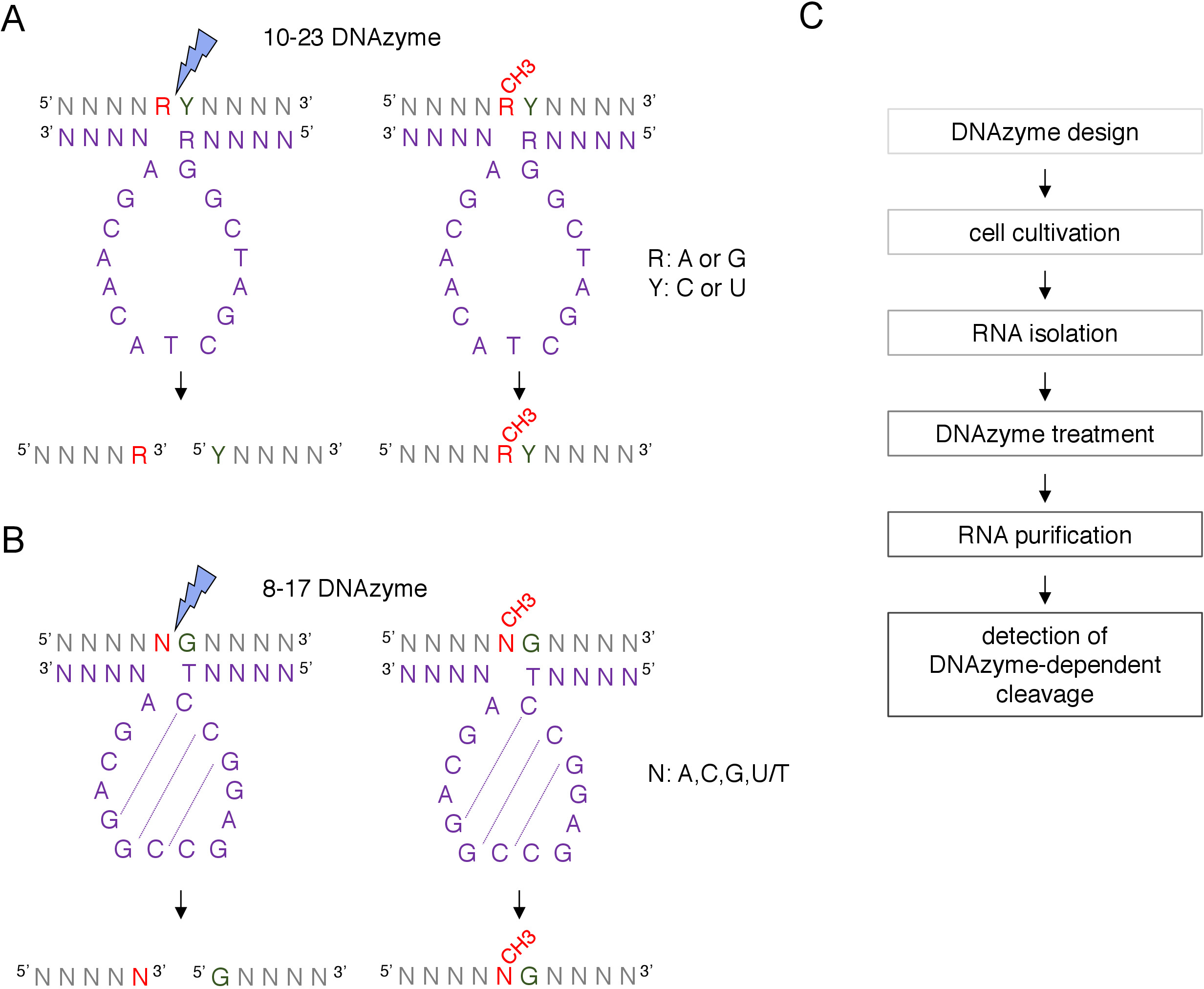
RNA modifikasyonları analizleri12için iki tip DNAzyme kullanılır. 10-23 DNAzyme(Şekil 1A)aktif dizilimi, hedeflenen RNA pürin-pirimidin (RY) dinükleotidetrafında bir döngü oluşturan ve bu iki nükleotit arasındaki bölünmeyi katalize eden 15 nükleotitten (Şekil 1A) oluşur. RNA pürin (R) DNAzyme ile baz eşleştirilmiş değildir ve DNAzyme üzerinde 2'-O-metilasyon sunar dekolte inhibe. 10-23 DNAzymes'in bağlayıcı kolları genellikle 10-15 nükleotit uzunluğundadır. İkinci DNAzyme sınıfı, 8-17 DNAzymes(Şekil 1B)14 nükleotit katalitik dizi (5'TCCGAGCCGGACGA3') içerir. Nükleotitler C2, C3 ve G4 çifti C9 G10 ve G11 ile kısa bir kök-döngü yapısı oluştururlar. 8-17 DNAzymes DNAzyme aktif dizi ilk timin ile kusurlu eşleştirilmiş herhangi bir guanin RNA yukarı cleave. Guaninin RNA nükleotit akıntısı DNAzyme ile baz la eşleşmiyor ve 2'-O-metilasyon bölünmeyi bozar. 8-17 DNAzymes dnazyme'yi kendine özgü dizilime yönlendirmek için yaklaşık 20 nükleotitten oluşan daha uzun homoloji kollarına ihtiyaç duyar.
Burada, 10-23 ve 8-17 DNAzyme bağımlı yaklaşımlar12,13 (Şekil 1C)kullanarak Saccharomyces cerevisiae rRNA 2'-O-metilasyon analizi için adım adım protokol sağlar. Bu protokol diğer organizmalar ve RNA türleri için kolayca uyarlanabilir ve bölgeye özgü RNA 2'-O-metilasyonun hızlı, ön veya ana analizleri için kullanılabilir.
Protokol
1. Suşlar, Medya ve Tampon Tarifleri
- Burada ayrıntılı olarak maya(S. cerevisiae)medya hazırlayın: YP (%1 w / v maya ekstresi, 2% w / v bakteriyolojik peptone), ve glikoz ve galaktoz stokları 20% w / v.
- Burada ayrıntılı olarak sodyum asetat (NaAc)-EDTA (AE) tampon hazırlayın: 50 mM NaAc pH 5.3 ve 10 mM EDTA.
- Burada ayrıntılı olarak 10-23 DNAzyme 4x Kuluçka Tampon hazırlayın: 24 mM Tris pH 8.0, 60 mM NaCl ve 10-23 DNAzyme 4x Reaksiyon Tampon: 200 mM Tris pH 8.0 ve 600 mM NaCl.
- Burada ayrıntılı olarak 8-17 DNAzyme 2x Reaksiyon Tampon hazırlayın: 200 mM KCl, 800 mM NaCl, 100 mM HEPES pH 7.0, 15 mM MgCl2, ve 15 mM MnCl2.
- Burada ayrıntılı olarak 10x MOPS Tampon hazırlayın: 200 mM MOPS, 50 mM NaAc, 1 mM EDTA; pH 7.0 ve 1.5x Örnek Denaturing Tampon: %50 v/v formamid, %20 v/v formaldehit, 1.5x MOPS tamponu.
- Elde S. cerevisiae suşları, BY4741 (MATa his3Δ1 leu2Δ0 met15Δ0 ura3Δ0); GAL1::SNR13 (BY4741 ama GAL1::SNR13:KANmXolarak); GAL1::SNR47 (BY4741 ama GAL1::SNR47:HIS3mXolarak). Başka herhangi bir maya suşu bu analiz için kullanılabilir.
2. DNAzyme Tasarım
- Uygun bir veritabanı kullanarak RNA ilgi veya putatif metilasyon sitesi sırasını bulun. S. cerevisiae snoRNA hedefleri için maya snoRNA veritabanını kullanın:http://people.biochem.umass.edu/fournierlab/snornadb/mastertable.php 14
- SnR13'e bağımlı site gibi metilasyon alanını bulmak için "snR13" seçeneğini belirleyin ve değiştirilmiş nükleotitin konumunu not edin (örn. snR13 güdümlü A2281 in 25S rRNA).
- Uygun veritabanını kullanarak değiştirilmiş nükleotitin yukarı ve aşağı akış dizilerini bulun. S. cerevisiae için Saccharomyces Genom Veritabanını kullanın: https://www.yeastgenome.org/
- RdN25 (25S rRNA kodlama) gibi hedef gen adını arayın.
- "Sıra" sekmesinden, 10-23 DNAzyme titrezi ve 20 nükleotit upstream (5' kol) ve 8-17 DNAzyme için metilasyon alanının aşağı (3'kol) kullanırken metilasyon alanının yukarı (5' kol) yukarı (5' kol) ve aşağı (3'kol) metilasyon alanının yukarı (5' kol) ve aşağı (3 'kol) seçin.
- 5' ve 3' koldan oluşan tamamlayıcı diziler oluşturun.
- Böğür 10-23 veya 8-17 DNAzyme katalitik dizili 5' ve 3' kolların tamamlayıcı dizileri.
- Tedarikçiden normal bir DNA oligonükleotid olarak Sipariş DNAzyme.
3. S. cerevisiae Büyüme Koşulları
NOT: S. cerevisiae BY4741 suş türevleri kullanıldı, hangi ya SNR13 veya SNR47 snoRNA ekspresyonu indükleyici GAL1 organizatörü tahrik edilir. Sentezlerini indüklemek veya inhibe etmek için, galaktoz(GAL1-bağımlıtranskripsiyon) veya glikoz(GAL1bağımlı transkripsiyon kapalı) içeren orta hücreler büyür. Bir kontrol olarak, galaktoz veya glikoz üzerinde yetiştirilen yabani türü (BY4741) kullanın.
- Uygun bir ortamda ve koşullarda maya suşları büyümek. GAL1::SNR13 ve GAL1::SNR47 suşlarının yanı sıra İzojenik yabani tip suşları analiz etmek için, yp medianın 50 mL'sinde %2 glikoz (YPD) veya orta üstel faza 30 °C'de galaktoz (YPGal) ile hücre yetiştirin.
- Santrifüj hücreleri 1.000 x g, 4 °C'de 3 dk.
- Supernatant atın ve pelet tutun.
- Hücre peletlerini sıvı nitrojen de dondurun ve -80 °C'de saklayın.
DİkKAT: Sıvı nitrojen ciddi kriyojenik yanıklara neden olabilir. Her zaman koruyucu giysiler giyin ve güvenlik önlemleri alın.
NOT: Hücre peletleri -80 °C'de 1 aya kadar saklanabilir. Protokol gerekirse burada duraklatılabilir.
4. RNA İzolasyon15
NOT: RNA'yı yalıtmak için en uygun yöntemi kullanın. Maya S. cerevisiaeiçin, sıcak-fenol RNA ekstraksiyon kullanılabilir.
- 1 mL buz gibi su ekleyin, peletleri yeniden askıya alın ve yeniden askıya alınan hücreleri 1,5 mL mikrotüplere aktarın.
- 4 °C'de 10 s için 20.000 x g'de santrifüj edin ve süpernatantı çıkarın.
- 400 μL'lik AE arabelleği ekleyin ve hücreleri yeniden askıya alın.
NOT: 4.4-4.15 adımları aksi belirtilmedikçe oda sıcaklığında gerçekleştirilir. - %10 SDS'nin 40 μL'si ve 400°L asit fenol (pH 4.5) ekleyin.
DİkKAT: Fenol toksik tir ve bir duman başlığı altında ele alınmalıdır. Fenol ile çalışırken her zaman laboratuvar önlüğü, koruyucu eldiven ve gözlük takın. Atıkları kurumsal düzenlemelere göre bertaraf edin. - 20 s için girdap tarafından iyice karıştırın.
- 10 dk için 65 °C'de kuluçka. Her 2 dakika, yavaşça açın ve basınç serbest bırakmak için tüp kapatın ve fazları karıştırmak için 2-3 kez tüp çevirmek.
- Tüpleri -80 °C'ye aktarın ve 10 dakika kuluçkaya yatırın.
- Oda sıcaklığında 5 dakika için, 20.000 x gbank ve santrifüj tüpler defrost.
- Üst fazı 400 μL asit fenol içeren yeni bir tüpe aktarın:kloroform:izoamyl alkol (25:24:1). İnterfazı bozmayın.
DİkKAT: Kloroform toksiktir ve duman kaputunun altında ele alınmalıdır. Kloroform la çalışırken her zaman laboratuvar önlüğü, koruyucu eldiven ve gözlük takın. Atıkları kurumsal düzenlemelere göre bertaraf edin. - Oda sıcaklığında 10 dakika boyunca 20.000 x g 30 s ve santrifüj için girdap tarafından iyice karıştırın.
- Üst fazı (~400 μL) 400 μL kloroform içeren yeni bir tüpe aktarın.
- Oda sıcaklığında 5 dakika boyunca 20.000 x g 30 s ve santrifüj için girdap tarafından iyice karıştırın.
- Üst fazı (~300-350 μL) 1 mL EtOH ve 40 μL 7,5 M amonyum asetat (NH4AC) içeren yeni bir tüpe aktarın. Tüpü birkaç kez çevirerek karıştırın.
- -80 °C'de 2 saat veya geceleme -20 °C'de kuluçkaya yatırın.
NOT: Yordam burada duraklatılmış olabilir. - Santrifüj 20.000 x g, 4 °C'de 10 dk. Küçük, beyaz bir RNA pelet tüpün altında görünür hale gelecektir.
- Peleti rahatsız etmemek için boru lar oluşturarak EtOH'u çıkarın.
- Oda sıcaklığında 5 dakika için 20.000 x g%70 EtOH ve santrifüj 1 mL ekleyin.
- Pipetleme ile %70 EtOH'u çıkarın.
- 15 s için 20.000 x g santrifüj ve 2-20 μL pipet ile kalan EtOH çıkarın.
- Tüp 5 dakika boyunca bankta açık bırakın RNA pelet kuru.
NOT: RNA peleti kuruduğunda rengini beyazdan saydama değiştirir. - RNA peletini 30 μL RNase/DNase içermeyen H2O'da yeniden askıya alın, tüpü hemen buzun üzerine aktarın ve mikrospektrofotometredeki RNA konsantrasyonu ölçün.
- Örnekleri -20 °C'de dondurun.
NOT: RNA -20 °C'de 1 aya kadar ve -80 °C'de 1 yıla kadar saklanabilir. Yordam burada duraklatılmış veya doğrudan bir sonraki adıma devam edilebilir.
5. DNAzyme Sindirim
-
10-23 DNAzyme sindirim
- 1,5 mL'lik tüplerde 5 g RNA, 200 pmol 10-23 DNAzyme (2 μL 100 mM stok çözeltisi) ve 2,5 μL 4x 10-23 Kuluçka Tamponu toplam hacminde 10 μL'lik bir kuluçka karışımı hazırlar.
- Tüpleri 95 °C'de belirlenen kuru bir ısı bloğuna aktarın ve 3 dakika kuluçkaya yatırın.
- Tüpleri hemen buzüzerinde aktarın ve 5 dakika kuluçkaya yatırın.
- Kısa bir süre aşağı dön ve tüpleri buza geri koy.
- 20 U RNase inhibitörü ekleyin (örn. 0.5 μL RiboLock RNase inhibitörü).
- Tüpleri 25 °C'ye ayarlanmış kuru bir ısı bloğuna yerleştirin ve 10 dakika kuluçkaya yatırın.
- Bu arada, 4x 10-23 Reaksiyon Tamponunun 5 μL'sini 300 mM MgCl2 ve 1 μL H2O.'yu 37 °C'ye ayarlanmış kuru bir blokta yerleştirerek 1,5 μL'lik bir tüpte reaksiyon karışımı hazırlayın.
- Kuluçka karışımını 37 °C'ye ayarlanmış kuru bir ısı bloğuna aktarın ve önceden ısıtılmış reaksiyon karışımından 10°L ekleyin.
- Reaksiyonu 37 °C'de 1 saat kuluçkaya yatırın.
- Tüpleri buzüzerinde aktarın ve adım 5.3.1'e geçin.
-
8-17 DNAzyme sindirim
- Toplam hacmi 6 μL olan 5 μg RNA içeren 1,5 mL'lik bir mikrotüp hazırlayın.
- 8-17 DNAzyme 400 pmol (100 mM stok tan 4 μL) ile 1,5 mL mikrotüp hazırlayın. Tüpü buzda tut.
- Tüpleri 95 °C için ayarlanmış kuru bir ısı bloğuna ve 2 dk kuluçkaya yatırın.
- RNA örneğini buz üzerinde hareket ettirin.
- 5 s için DNAzyme ile tüp aşağı spin ve 10 dakika için 25 °C'de kuluçka.
- Aynı zamanda 10 μL 2x 8-17 Reaksiyon Tamponu ve 25 °C'de kuluçkaya yatan 1,5 mL'lik bir tüp hazırlayın.
- DNAzyme ile tüpe önceden ısıtılmış 2x Reaksiyon Tamponu 10 μL ekleyerek bir reaksiyon karışımı hazırlayın.
- Reaksiyon karışımının 14 μL'sini RNA ile tüpe aktarın ve 20 U RNase inhibitörü ekleyin.
- Reaksiyonu 2 5 °C'de 2 saat kuluçkaya yatırın.
- Tüpü buz üzerinde aktarın ve RNA arınma sınamaya geçin (adım 5.3.1).
-
RNA arınma
- Reaksiyon tüpüne 350 μL su ve 400 μL kloroform ekleyin, 30 s'lik girdap la karıştırın ve oda sıcaklığında 5 dakika boyunca 20.000 x g'de santrifüj de karıştırın.
- Üst fazı (~300-350 μL) 1 mL EtOH, 40 μL 7,5 M NH4AC ve 1 μL glikojen (10 μg/μL) içeren yeni bir tüpe aktarın. Tüpü birkaç kez çevirerek karıştırın.
- -80 °C'de 2 saat veya geceleme -20 °C'de kuluçkaya yatırın.
NOT: Yordam burada duraklatılmış olabilir. - 4,15'ten 4,21'e kadar olan adımları yineleyin.
- RNA peletini 10 μL RNase/DNase içermeyen H2O'da yeniden askıya alın ve tüpleri hemen buzüzerinde aktarın.
- Örnekleri -20 °C'de dondurun.
NOT: RNA bir aya kadar -20 °C'de ve 1 yıla kadar -80 °C'de saklanabilir. Prosedür burada duraklatılmış veya RNA elektroforez ini devam edebilir.
6. RNA Elektroforez
- Sprey elektroforez ekipmanları (tank, tepsi, tarak) ile 1% SDS, 15 dakika bekletin ve ddH2O bol durulayın.
- 127,5 mL ddH2O'da 1,5 g agarose'u mikrodalgada ısıtarak eritin.
- Agarose çözeltisine 15 mL 10x MOPS ve 7,5 mL %37 formaldehit ekleyin (toplam hacim 150 mL'dir).
DİkKAT: Formaldehit toksiktir ve bir duman başlığı altında ele alınmalıdır. Formaldehit ile çalışırken her zaman laboratuvar önlüğü, koruyucu eldiven ve gözlük takın. Atıkları kurumsal düzenlemelere göre bertaraf edin. - Agarose çözeltisine uygun miktarda jel lekesi ekleyin (örn. 15 μL SYBR Güvenli DNA jeli lekesi). Iyice karıştırın ve tepsiye agarose dökün.
- Hemen jel bir tarak yerleştirin.
- Duman kaputunun altında 45 dakika bekletin. Hafif hassasiyetli jel lekesi kullanırken tepsiyi alüminyum folyo ile kaplayın.
- 1x MOPS tampon600 mL hazırlayın.
-
RNA numune hazırlama
- 1,5 mL'lik bir tüpte, sindirilmiş ve saflaştırılmış RNA numunesinin 10 μL'sini, 5 μL'lik Numune Denatüre Tampon'u ve 0,5 μL'lik 6x Yükleme Boyasını birleştirin.
DİkKAT: Formamide toksik tir ve duman kaputunun altında ele alınmalıdır. Formamid ile çalışırken her zaman laboratuvar önlüğü, koruyucu eldiven ve gözlük takın. Atıkları kurumsal düzenlemelere göre bertaraf edin. - 5 dk. 70 °C'de kuluçkaya yatan RNA örnekleri, numuneleri buz üzerinde aktarın. 5 dakika kuluçka.
- Jeli yüklemeden önce kısa bir süre aşağı dön.
- 1,5 mL'lik bir tüpte, sindirilmiş ve saflaştırılmış RNA numunesinin 10 μL'sini, 5 μL'lik Numune Denatüre Tampon'u ve 0,5 μL'lik 6x Yükleme Boyasını birleştirin.
- Elektroforez tankına jel koyun ve 1x MOPS tampon ile doldurun. Her numunenin (15°L) tüm hacmini jel üzerine yükleyin. Bromofenol mavisi jel uzunluğunun 2/3'üne ulaşana kadar 80 V'da çalıştırın.
- Seçilen jel lekesini (örneğin, UV transilluminator) tespit etmek için uygun bir görüntüleyici kullanarak jeli görüntüleyin.
Sonuçlar
RRNA modifikasyonlarının analizinde DNAzyme bağımlı dekolte nin faydası son zamanlarda snoRNA olgunlaşma13bağlamında gösterilmiştir. DNAzyme bağımlı tsay 5'-end pre-snoRNA işleme eksikliği 2'-O-metilasyon düzeyleri 2'-O-metilasyon düzeylerini etkilediğini göstermek için kullanılmıştır 25S ve 18S rRNA S. cerevisiae13.
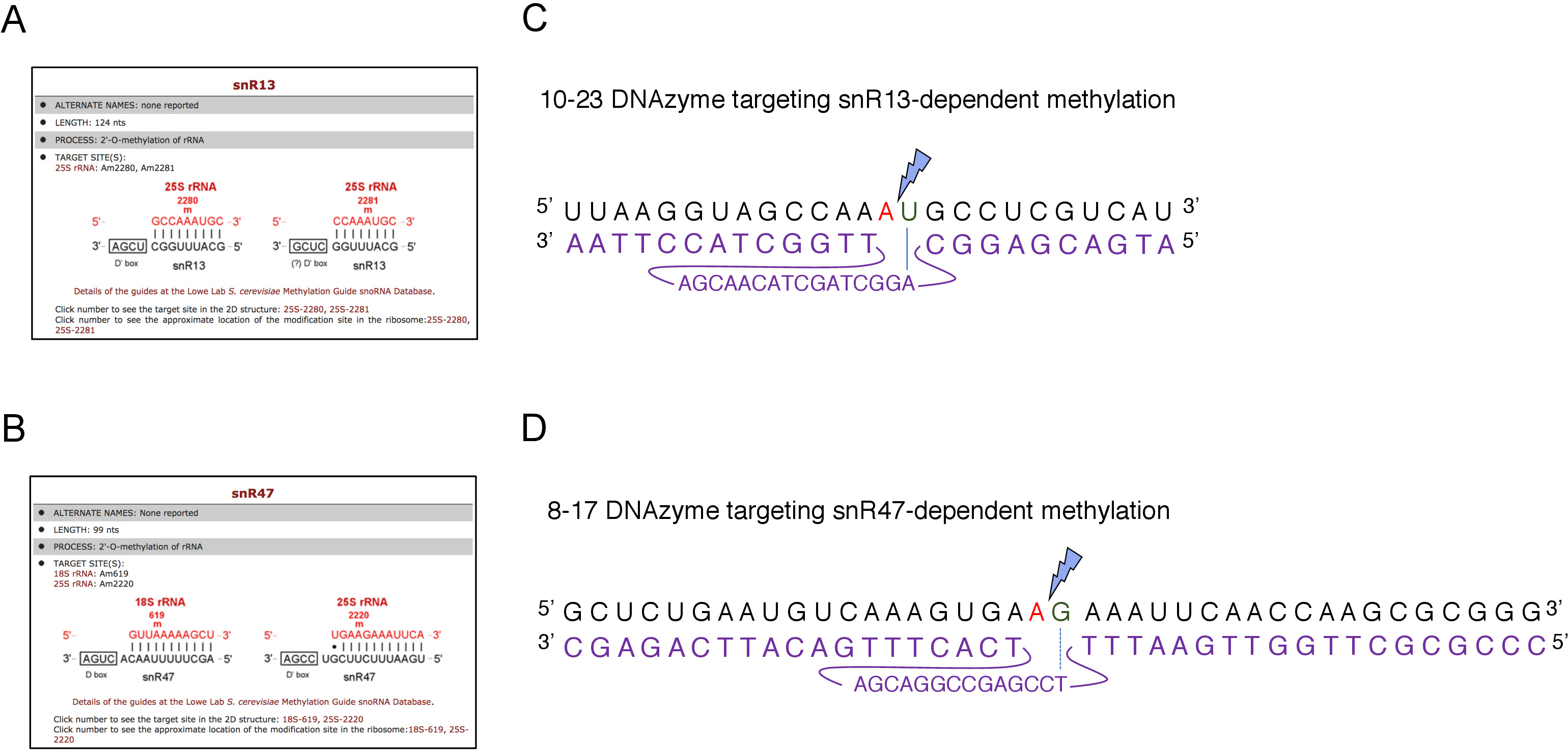
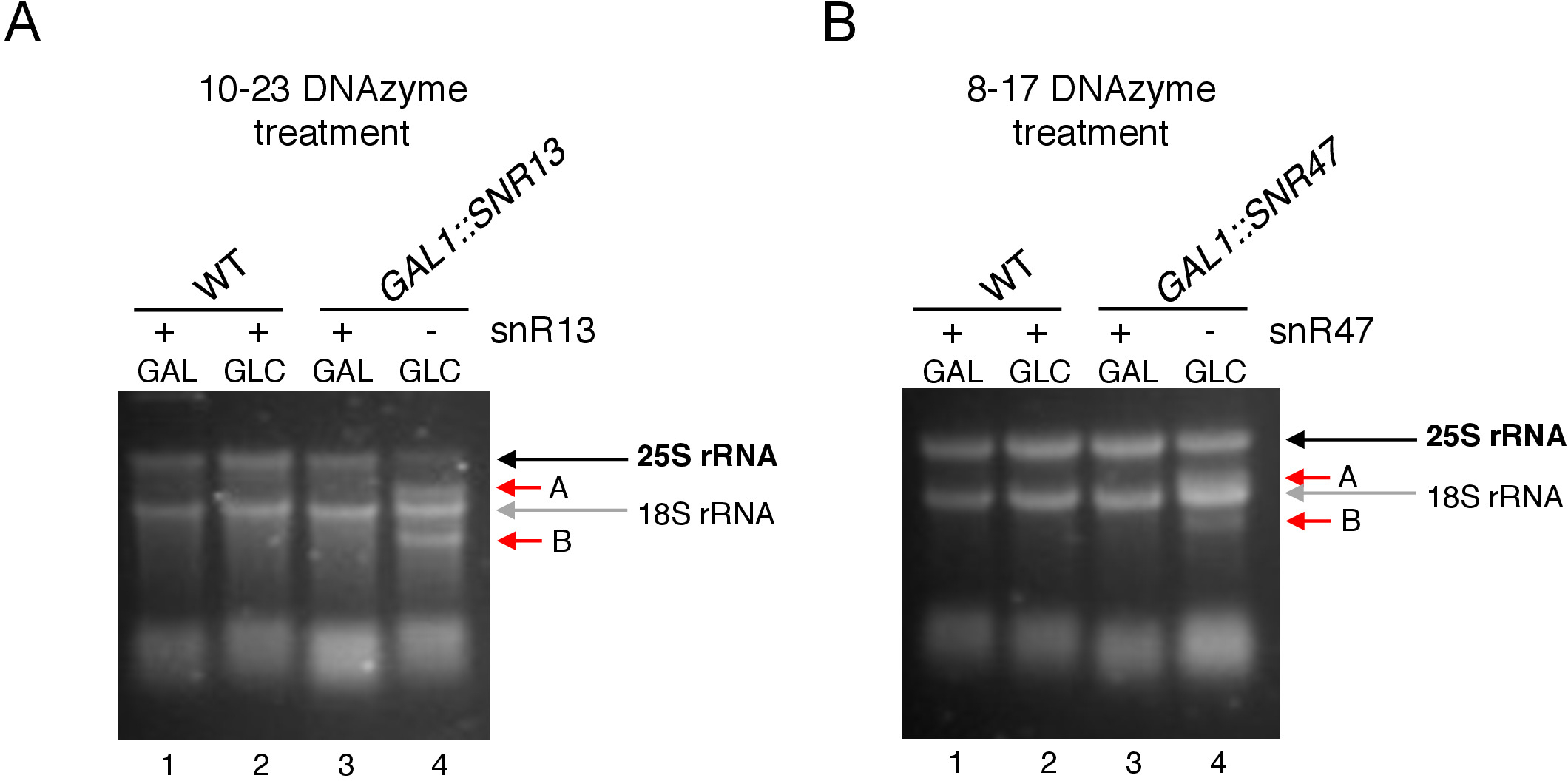
Burada, tekniğin etkinliğini ve basitliğini göstermek için indükleyici bir snoRNA transkripsiyon sistemi kullandık. Kutu C/D snR13, 25S rRNA'da iki pozisyonda metilasyon kılavuzları, adenin 2281(Şekil 2A). Bu nükleotiti urasil takip eder, bu da 10-23 DNAzyme ile bölünmek tekonsusu dinükleotid (RY) oluşturur. Kutu C/D snR47 ayrıca 25S rRNA'da iki nükleotitin metilasyonunu yönlendirir (Şekil 2B). Pozisyon 2220 adenin bir guanin kalıntısı tarafından takip edilir ve bu dinükleotit bir 8-17 DNAzyme tarafından yarık olabilir. SnR13 veya snR47 snoRNA sentezini indüklemek veya inhibe etmek için, snR13 veya SNR47 genlerinin indüklen GAL1 promotörupstream'ini ve galaktoz içeren orta bölgelerde ki ekili hücreleri (GAL1-bağımlı transkripsiyon) veya glikoz(GAL1-bağımlı transkripsiyon kapalı). Daha sonra, GAL1 izole RNA::SNR13 hücreleri 10-23 DNAzyme snR13 bağımlı sitede 25S rRNA cleave için tasarlanmış ile inkübe edildi, nükleotidler arasında 2281 ve 2282(Şekil 2C). GAL1'den RNA::SNR47 suşu nükleotitler arasında 8-17 DNAzyme hedefleme snR47 bağımlı site ile tedavi edildi 2220 ve 2221 (Şekil 2D). Bir kontrol olarak, yabani tip BY4741 suşu ya galaktoz veya glikoz üzerinde büyüyen RNA her iki DNAzymes ile inkübe edildi. DNAzyme ile tedavi edilen RNA'nın elektroforezi, GAL1::SNR13 ve GAL1::SNR47 suşlarının galaktoz (GAL) üzerinde büyüyen 25S rRNA'nın bozulmadan kaldığını ortaya koymuştur (Şekil 3A,B; şeritler 3). Buna karşılık, RNA GAL1 izole::SNR13 ve GAL1::SNR47 hücreleri glikoz üzerinde büyüyen (GLC) ilgili DNAzymes tarafından sindirilmiştir(Şekil 3A,B; lanes 4). Her iki olguda da 25S rRNA bandı azaldı ve 5' ve 3' kesme dekolte ürünleri (A ve B) gözlendi. Bu gal1::SNR13 ve GAL1::SNR47 suşları, 25S rRNA 2'-O-methylated snR13- veya snR47-güdümlü sitelerde galaktoz bir karbon kaynağı olarak kullanılan ve bu snoRNA ifade edildi gösterir. SnR13 veya snR47 ekspresyonu DNAzyme bağımlı dekolte için izin verilen glikoz üzerinde kapatıldı zaman 25S rRNA metilasyon eksikliği. Yabani tip numuneler için RNA sindirimi gözlenmemiştir(Şekil 3A,B; şerit 1 ve 2), snR13 ve snR47'nin ifadesi galaktoz/glikozdan bağımsız dır. Bu nedenle, rRNA normalde metillenmiş ve dnazymes aktivitesine karşı çok dirençliidi.
Genel olarak, bizim deney 10-23(Şekil 3A) ve 8-17(Şekil 3B) DNAzymes kutu C / D snR13 veya snR47 yokluğu ile ilişkili dekolte aktivitesi gösterir, açıkça bu snoRNA sorumlu olduğunu gösteren 25S rRNA 2'-O-metilasyon belirli yerlerde.

Şekil 1: DNAzymes ve RNA yüzeyleri. (A) 10-23 DNAzymes bir pürin-pirimidin (RY) RNA dinükleotid cleave. RNA'daki R DNAzyme ile eşleşmiyor, Y ise DNAzyme'deki R tabanına tamamlayıcıdır. RNA'da pürinin (R) metilasyon DNAzyme bağımlı dekolte bastırır. (B) 8-17 DNAzymes, DNAzyme katalitik dizideki ilk timinle kusurlu bir şekilde eşleştirilmiş guaninin RNA'sını yukarı doğru besler. Guaninden önceki nükleotit eşlenmez ve metilasyon DNAzyme bağımlı dekolte korur. RNA gri (metilasyon alanı dışında) gösterilir, DNAzyme mor olarak gösterilir. N = herhangi bir nükleotit, R = pürin: adenin veya guanin, Y = pirimidin: sitozin veya urasil; CH3- RNA metilasyon gösterir. DNAzyme etkin dizileri içindeki baz eşleşmesi noktalı çizgilerle işaretlenir. Mavi bir şimşek dekolte alanını işaretler. (C) DNAzyme bağımlı bir analizin adımlarını gösteren bir akış şeması. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.
{kind=link}

Şekil 2: 25S rRNA'da snR13 ve snR47 bağımlı metilasyon bölgelerini hedefleyen DNAzymes. (A, B) SnR13 bağımlı(A) ve snR47 bağımlı (B) metilasyon sitelerini 25S rRNA (C) 25S rRNA dizisini çevreleyen snR13 bağımlı metilasyon alanı (A2281)ve 10-23 DNAzyme (mor olarak gösterilmiştir) gösteren tarayıcı ekran görüntüleri A2281 ve U2282arasında RNA cleave için tasarlanmıştır. U2282 DNAzyme aktif dizisinden ilk nükleotit (mavi bir çizgi ile işaretlenmiş) ile bir çift oluştururken A2281 DNAzyme ile eşleştirilmiş değildir. Mavi bir şimşek dekolteyi işaretler. (D) 25S rRNA dizisi çevreleyen snR47 bağımlı metilasyon alanı (A2220) ve 8-17 DNAzyme (mor gösterilir) A2220 ve G2221arasında RNA cleave için tasarlanmıştır. G2221 kusurlu timin ile eşleştirilmiş ise Bir2220 DNAzyme ile hibridize edilmez (kesik çizgi ile gösterilir). Mavi bir şimşek dekolte alanını işaretler. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.
{kind=link}

Şekil 3: 10-23 ve 8-17 DNAzyme bağımlı tahlil kullanılarak 25S rRNA'nın bölgeye özgü 2'-O-metilasyonanalizi. (A) 10-23 DNAzyme kullanılarak snR13 bağımlı 25S rRNA metilasyon analizi. (B) 8-17 DNAzyme kullanılarak snR47-bağımlı 25S rRNA metilasyon analizi. RNA, denagüle ağarose jelinde lekelenme görüntülenmiş. Dekolte ürünleri A ve B kırmızı oklarla işaretlenir. WT = yabani tip zorlanma; GAL = galaktoz, GLC = glikoz. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.
{kind=link}
Tartışmalar
DNAzyme bağımlı sindirim siteye özgü RNA 2'-O-metilasyon12,13analiz etmek için basit ve hızlı bir yöntem olarak kullanılabilir. Dekolte bölgesinin nükleotit yukarı metilatif değilse DNAzymes CLEAVE RNA. Diğer yaklaşımların aksine, RNase H-yönettiği sindirim, alkali bozulma veya düşük nükleotit konsantrasyonu içinde ters transkripsiyon kantitatif PCR veya sıralama8,10,11 takip ,16, DNAzyme yaklaşım basit bir DNA oligonükleotid ve herhangi bir moleküler biyoloji laboratuvarında mevcut temel reaktifler gerektirir. Ayrıca, DNAzymes kutu H / ACA snoRNA12aracılık RNA psödouridylation analiz etmek için benzer bir şekilde kullanılabilir , hangi onları snoRNA hedefleri çalışmada çok yönlü araçlar yapar.
DNAzyme bağımlı yaklaşımlar sadece dekolte site konsensüs dizileri ile sınırlıdır17. 10-23 DNAzymes 2'-O-metilasyon sadece RY dinükleotidin R pozisyonunda analiz etmek için kullanılabilirken, 8-17 DNAzymes guanin akıntısında bulunan nükleotitmodifikasyonunu tanır. Sonuç olarak, dinükleotidler guanin-adenin (GA), adenin-adenin (AA), pirimidin-adenin (YA) ve pirimidin-pirimidin (YY) ilk nükleotit 2'-O-metilasyon gibi değişiklikler analiz edilemez. Ayrıca, DNAzyme bağımlı dekolte12 düşük verimlilik düşünülmelidir. Bazı DNAzymes neredeyse tamamen RNA cleave rağmen(Şekil 3B),birçok DNAzymes sadece kısmen hedeflerini sindirmek(Şekil 3B). Verimlilik dekolte alanını çevreleyen sıraya bağlı olabilir. Örneğin, aynı nükleotit in enine ait RNA bölgeleri DNAzyme etkin dizisinin doğru konumlandırılmasını etkileyebilir. Ayrıca, güçlü ikincil yapı oluşturan RNA bölgeleri yeniden melezleyebilir ve DNAzyme'yi hedef sıraya bağlayabilir. Bu sorunların üstesinden gelmek için, 10-23 DNAzyme ve RNA substrat ısıtma ve soğutma döngüleriuygulanabilir 18.
RRNA'nın 2'-O-metilasyonunu araştırmak için DNAzyme yaklaşımını kullandık. N6-metyladenozin19gibi diğer RNA modifikasyonlarını analiz etmek için de bu tekniği kullanabilirsiniz. Ribozomal RNA, bolluğu nedeniyle elektroforez ile analiz edilebilir ve dekolte ürünleri UV ışığı altında görselleştirilebilir. Ancak, bu RNA Polimeraz II-oluşturulan kodlama RNA 'lar (mRNA) ve kodlama olmayan RNA'lar (ncRNA) gibi daha az bol RNA'lar için geçerli değildir. Bu RNA'lar genellikle agarose veya poliakrilamid jellerde RNA boyama ile doğrudan tespit edilemez. Bu gibi durumlarda, DNAzyme bağımlı dekolte Kuzey blotting tarafından görselleştirilmiş olabilir, dolaylı PCR / nicel PCR tarafından tespit veya polimeraz (örneğin, KlenTaq DNA Polimeraz) ile kantitatif PCR tarafından analiz 2′-O-metillenmiş RNA ayırt yeteneğine sahip metillenmemiş RNA20,21.
Açıklamalar
Yazarların açıklayacak bir şeyi yok.
Teşekkürler
Biz makalenin eleştirel okuma için Maya Wilson ve Aneika Leney teşekkür ederiz. Bu çalışma, Wellcome Trust ve Royal Society (200473/Z/16/Z) tarafından ortaklaşa finanse edilen Sir Henry Dale Bursu tarafından desteklenmiştir.
Malzemeler
| Name | Company | Catalog Number | Comments |
| Chemicals | |||
| Acid phenol | SIGMA | P4682 | |
| Agarose | VWR | A2114 | |
| Ammonium acetate | SIGMA | A1542 | |
| Chlorophorm | Fisher scientific | 10293850 | |
| DNase/RNase free water | Fischer Scientific | 10526945 | |
| DNAzyme | Integrated DNA Technology | Custom oligo DNA | |
| EDTA | SIGMA | E9884 | |
| Ethanol Absolute | Fisher scientific | 10437341 | |
| Formaldehyde | Sigma | F8775 | |
| Formamide | sigma | F9037 | |
| Galactose | SIGMA | G0750 | |
| Gel Loading Dye | Thermo Fisher Scientific | R0611 | |
| Glucose | SIGMA | G7021 | |
| Glycogen | Thermo Fisher Scientific | R0561 | |
| HEPES | SIGMA | H3375 | |
| Isoamyl | SIGMA | W205702 | |
| KCl | SIGMA | P9333 | |
| MgCl2 | SIGMA | M8266 | |
| MnCl2 | SIGMA | 244589 | |
| MOPS | SIGMA | M1254 | |
| NaCl | SIGMA | S7653 | |
| Oxoid Peptone Bacteriological | Thermo Fisher Scientific | LP0037 | |
| Oxoid Yeast Extract Powder | Thermo Fisher Scientific | LP0021 | |
| RiboLock RNase Inhibitor (40 U/µL) | Thermo Fisher Scientific | EO0382 | |
| SDS | SIGMA | 74255 | |
| Sodium acetate trihydrate | SIGMA | S8625 | |
| SYBR Safe DNA Gel Stain | Thermo Fisher Scientific | S33102 | |
| Tris base | SIGMA | TRIS-RO | |
| Name | Company | Catalog Number | Comments |
| Equipment | |||
| 1.5 mL microtubes | Sarstedt | ||
| 152VR5C01M -80°C freezer | Thermo Fisher Scientific | ||
| 250 mL Erlenmeyer flasks | Cole-Parmer | ||
| 50 mL conical tubes | Sarstedt | ||
| Combicup VX200 vortex | Appleton Woods | ||
| DS-11 microspectrophotometer | Denovix | ||
| Electrophoresis chamber (20 cm tray) | SIGMA | ||
| FiveEasy F20 pH meter | Appleton Woods | ||
| Gel documentation system | Syngene | ||
| Heraeus Fresco 21 micro centrifuge | Fisher Scientific | ||
| Megafuge 8R centrifuge with rotator suitable for 50 mL conical tubes | Fisher Scientific | ||
| Mini Fuge Plus mini centrifuge | Starlab | ||
| Mixer HC thermal block | Starlab | ||
| OLS26 Shaking Water Bath | Grant | ||
| PowerPac power supplier | BioRad |
Referanslar
- Dieci, G., Preti, M., Montanini, B. Eukaryotic snoRNAs: a paradigm for gene expression flexibility. Genomics. 94 (2), 83-88 (2009).
- Watkins, N. J., Bohnsack, M. T. The box C/D and H/ACA snoRNPs: key players in the modification, processing and the dynamic folding of ribosomal RNA. Wiley Interdisciplinary Review RNA. 3 (3), 397-414 (2012).
- Kufel, J., Grzechnik, P. Small Nucleolar RNAs Tell a Different Tale. Trends in Genetics. , (2018).
- Li, T., Zhou, X., Wang, X., Zhu, D., Zhang, Y. Identification and characterization of human snoRNA core promoters. Genomics. 96 (1), 50-56 (2010).
- Jorjani, H., et al. An updated human snoRNAome. Nucleic Acids Research. 44 (11), 5068-5082 (2016).
- Hubbard, T. J., et al. Ensembl 2009. Nucleic Acids Research. 37 (Database issue), D690-D697 (2009).
- Makarova, J. A., Kramerov, D. A. SNOntology: Myriads of novel snoRNAs or just a mirage?. BMC Genomics. 12, 543 (2011).
- Yu, Y. T., Shu, M. D., Steitz, J. A. A new method for detecting sites of 2'-O-methylation in RNA molecules. RNA. 3 (3), 324-331 (1997).
- Decatur, W. A., Liang, X. H., Piekna-Przybylska, D., Fournier, M. J. Identifying effects of snoRNA-guided modifications on the synthesis and function of the yeast ribosome. Methods in Enzymology. 425, 283-316 (2007).
- Dong, Z. W., et al. RTL-P: a sensitive approach for detecting sites of 2'-O-methylation in RNA molecules. Nucleic Acids Research. 40 (20), e157 (2012).
- Birkedal, U., et al. Profiling of ribose methylations in RNA by high-throughput sequencing. Angewandte Chemie International Edition, England. 54 (2), 451-455 (2015).
- Buchhaupt, M., Peifer, C., Entian, K. D. Analysis of 2'-O-methylated nucleosides and pseudouridines in ribosomal RNAs using DNAzymes. Analytical Biochemistry. 361 (1), 102-108 (2007).
- Grzechnik, P., et al. Nuclear fate of yeast snoRNA is determined by co-transcriptional Rnt1 cleavage. Nature Communication. 9 (1), 1783 (2018).
- Piekna-Przybylska, D., Decatur, W. A., Fournier, M. J. New bioinformatic tools for analysis of nucleotide modifications in eukaryotic rRNA. RNA. 13 (3), 305-312 (2007).
- Schmitt, M. E., Brown, T. A., Trumpower, B. L. A rapid and simple method for preparation of RNA from Saccharomyces cerevisiae. Nucleic Acids Research. 18 (10), 3091-3092 (1990).
- Maden, B. E. Mapping 2'-O-methyl groups in ribosomal RNA. Methods. 25 (3), 374-382 (2001).
- Santoro, S. W., Joyce, G. F. A general purpose RNA-cleaving DNA enzyme. Proceedings of the National Academy of Science, U. S. A. 94 (9), 4262-4266 (1997).
- Hengesbach, M., Meusburger, M., Lyko, F., Helm, M. Use of DNAzymes for site-specific analysis of ribonucleotide modifications. RNA. 14 (1), 180-187 (2008).
- Sednev, M. V., et al. N(6) -Methyladenosine-Sensitive RNA-Cleaving Deoxyribozymes. Angewandte Chemie International Edition, England. 57 (6), 15117-15121 (2018).
- Aschenbrenner, J., Marx, A. Direct and site-specific quantification of RNA 2'-O-methylation by PCR with an engineered DNA polymerase. Nucleic Acids Research. 44 (8), 3495-3502 (2016).
- Lee, K. W., Bogenhagen, D. F. Assignment of 2'-O-methyltransferases to modification sites on the mammalian mitochondrial large subunit 16 S ribosomal RNA (rRNA). Journal of Biological Chemistry. 289 (36), 24936-24942 (2014).
Yeniden Basımlar ve İzinler
Bu JoVE makalesinin metnini veya resimlerini yeniden kullanma izni talebi
Izin talebiDaha Fazla Makale Keşfet
This article has been published
Video Coming Soon
JoVE Hakkında
Telif Hakkı © 2020 MyJove Corporation. Tüm hakları saklıdır