
Method Article
FRET tabanlı Sensör ATeam1.03YEMK kullanılarak Fare Beyninin Organotipik Doku Dilimlerinde Hücre İçi ATP'nin Görüntülenmesi
Bu Makalede
Özet
Fare ön beyninin organotipik dilim kültürlerinde genetik olarak kodlanmış FRET tabanlı sensör ATeam1.03YEMK'ın hücre tipi spesifik ekspresyonu için bir protokol tanımladık. Ayrıca, nöronlar ve astrositlerde hücresel ATP düzeylerinin dinamik görüntülemeiçin bu sensörünasıl kullanılacağını gösteriyoruz.
Özet
Merkezi sinir sisteminde nöronal aktivite (CNS) adenozin trifosfat (ATP) dökümü tarafından sağlanan hücresel enerji yüksek bir talep çağrıştırıyor. Nöronların elektriksel sinyalizasyonu ile bozulan plazma membranları arasında iyon gradyanları yeniden yüklemek için ATP'nin büyük bir kısmı gereklidir. Astrositlerin - hızlı elektrik sinyalleri üretmeseler de - nöronal aktiviteye yanıt olarak ATP üretiminin arttığını ve onlara enerji metabolitleri sağlayarak aktif nöronları desteklediklerine dair kanıtlar vardır. Farklı metabolitler için genetik olarak kodlanmış sensörlerin son gelişimi artık nöronlar ve astrositler arasındaki bu tür metabolik etkileşimlerin incelenmesini sağlamaktadır. Burada, adeno ile ilişkili viral vektörler (AAV) kullanılarak fare hipokampus ve korteksin organotipik doku dilimi kültürlerinde ATP'ye duyarlı Floresan Rezonans Enerji Transferi(FRET-) sensörü ATeam1.03YEMK'ın hücre tipi spesifik ekspresyonu için bir protokol açıklıyoruz. Ayrıca, bu sensörün hücre dışı potasyumdaki artışlar üzerine ve kimyasal iskemiin indüksiyonunu takip ederek nöronlar ve astrositlerdeki hücresel ATP düzeylerindeki değişikliklerin dinamik ölçümü için nasıl istihdam edilebildiğini (yani hücresel enerji metabolizmasının inhibisyonu) nasıl kullanabileceğini gösteriyoruz.
Giriş
Nöronların uyarıcı elektriksel aktivitesi büyük ölçüde plazma zarları arasında sodyum (Na+) ve potasyum (K+)gibi katyonların akı dayanmaktadır. Bu nedenle sinyalizasyon için bu iki iyonun elektrokimyasal degradelerinin bakımı gereklidir. Bu hücresel Na+/ K+-ATPase (NKA), her yerde ifade elektrojenik transmembran pompa tarafından gerçekleştirilir, hangi ekstrüzyon 3 Na+ hücre dışıuzaydan karşılığında hücre den extrudes , taşımadöngüsü1 başına ATP bir molekülün tüketimini gerektiren 1 . NKA ek olarak, plazma membran Ca2 +-ATPase, hücre içi Ca2 + homeostaz ve aktivite kaynaklıakını2 aşağıdaki ihracat için hayati önem taşımaktadır dahil olmak üzere birkaç diğer ATP tüketen iyon taşıyıcıları vardır. Presinaptik veziküllerde, bir vakuoler tip H+-ATPaz (v-ATPAz) bu bölmeye nörotransmitter alımı için gerekli proton gradyanı oluşturur3.
Nöronların aktivitesi böylece ATP önemli miktarda gerektirir iken4, onlar enerji depolama için önemli bir kapasite sergilemez. Bunun yerine, onlar komşu astrositler ile metabolik etkileşimleri güvenmek gibi görünüyor, beyinde büyük glikojen depoları5. Bu astrositik glikojen gerçekten nöronal enerji ihtiyaçlarının desteklenmesinde önemli bir rol oynadığı ileri sürülmüştür; ve iki hücre türleri arasında bu önerilen nöro-metabolik bağlantı önemli bir fenomen nöronal aktivite ye yanıt olarak ATP üretimini artırmak için astrositkapasitesi6,7,8. Bu hipotez, astrosit-nöron laktat mekiği olarak bilinen (ANLS), diğer çalışmalar da uyarılma yanıt olarak glikolikendi oranını artırabilir kanıt sağlamıştır çünkü9,10, nöro-glia etkileşimi çalışma için daha fazla yöntem ve yaklaşımların gerekliliğini yansıtan.
Nöro-glia metabolik etkileşimleri açıklamak için nöronlar ve astrositlerde hücresel enerji metabolizması ve ATP düzeylerinin araştırılması uzun beyin dokusunda yaşayan hücrelerde metabolit konsantrasyonlarında değişikliklerin tespiti için uygun probların eksikliği tarafından engel olmuştur. Son on yılda, ancak, atp, laktat, pirüuvat ve diğerleri için sensörler de dahil olmak üzere farklı metabolitleri için yeni araçlar ve yeni genetiği kodlanmış floresan probların geliştirilmesinde bir dalgalanma sağlamıştır11,12. Bu araçları kullanarak, hücresel ATP tüketimi ve tek hücre düzeyinde hücresel enerji düzeylerindeki değişiklikler ve bozulmamış beyindokusu13hücre tipi özel bir şekilde ilgili sorulara doğrudan hitap etmek mümkündür.
Bu çalışmada, kültürlü organotipik beyin dilimlerinin nöronlar ve astrositleri üzerinde sitosolik ATP dinamiklerini görselleştirmek için bir prosedür açıklıyoruz. Biz genetik olarak kodlanmış ATP-nanosensor ATeam1.03YEMK (14) nöronlar ve birkaç hafta hücre kültüründe hücre kültüründe tutulabilir fare beyin dilimleri astrositler hücre tipi özel ifade için adeno ilişkili viral vektörler (AAV) nasıl istihdam göstermek15. Kültürlü doku dilimlerini kapsayan glial yaranın nasıl giderilenin bir prosedürü tanımlanmıştır, bu da altındaki organotipik doku katmanlarındaki hücrelerin optik erişilebilirliğini ve görüntülenmesini geliştirir. Son olarak, ATeam1.03YEMK'nin bu hazırlıkta hücresel ATP düzeylerindeki değişikliklerin FRET tabanlı görüntülemesini gerçekleştirmek için nasıl kullanılabileceğini gösteriyoruz. Bu yöntem cerrahi beyin prosedürleri gerektirmez önemli avantajları barındırır, kültürlü beyin dilimleri sensör ve hücre tipi özgüllük yüksek düzeyde sağlar, diğer viral vektörler ile elektroporasyon veya transdüksiyon ile transfeksiyon gibi diğer yöntemlerle karşılaştırıldığında hücrelerde invazivlik veya stresi azaltarak10,16,17. Buna ek olarak, bu protokol atp14için daha düşük bağlama afinitesi sağlayan ATeam1.03 türevleri arasında diğer FRET tabanlı nanosensörler uygulanabilir.
Protokol
Bu çalışma, Heinrich Heine Üniversitesi Düsseldorf'un kurumsal yönergelerine ve Avrupa Topluluğu Konseyi Direktifi'ne (2010/63/EU) uygun olarak yürütülmüştür. Organotipik beyin dilimi kültürleri kullanılarak yapılan tüm deneyler, Heinrich Heine Üniversitesi Düsseldorf Hayvan Bakım ve Kullanım Tesisi Hayvan Refahı Ofisi'ne iletildi ve onaylandı (kurumsal hareket numarası: O50/05). Avrupa Komisyonu'nun tavsiyeleri doğrultusunda18, 10 güne kadar olan hayvanlar da kafa kesilerek öldürüldü.
1. Organotipik Beyin Dilimi Kültürlerinin (OTCs) Hazırlanması
- İşlemden bir gün önce veya en az 30 dk
- Petri kabını hazırlayın (steril koşullar altında). 6 kuyu plakasının kapağını çıkarın ve her kuyuya 800-850 μL OTC ortamı yerleştirin. Plakayı gerekli olana kadar kuvözde saklayın (37 °C, %5 CO2/95%O2).
- Yıkama Petri yemekleri (steril koşullar altında) hazırlayın. Her 30 mm Petri kabına 3 mL HBSS ekleyin. Prosedür başına toplam 5 yemek gereklidir. En az 30 dakika boyunca kuvöze (37 °C, %5 CO2/95% O2) yerleştirin .
- ACSF'yi hazırlayın (Tablo 1). Tuzlu eti glikozsuz 4 °C'de ertesi güne kadar saklayın.
| Salines ve Medya - Formülasyon | ||||
| Adı | Kısaltma | Kompozisyon | Konsantrasyon [mM] | Yorum |
| Yapay Beyin Omurilik Sıvısı Çözümü | ACSF | Nacl | 125 | %5 CO2/95% O2, pH 7.4 ile kabarcıklı |
| Kartal | 2.5 | Her zaman kullanmadan hemen önce glikoz ekleyin. | ||
| CaCl2 | 2 | Glikoz ile bir günden fazla saklamayın | ||
| MgCl2 | 1 | ~310 mOsm/L | ||
| NaH2PO4 | 1.25 | |||
| NaHCO3 | 26 | |||
| Glikoz | 20 | |||
| Deneysel ACSF | E-ACSF | Nacl | 136 | %5 CO2/95% O2, pH 7.4 ile kabarcıklı |
| Kartal | 3 | Her zaman kullanmadan hemen önce glikoz ekleyin. | ||
| CaCl2 | 2 | Glikoz ile bir günden fazla saklamayın | ||
| MgCl2 | 1 | ~320 mOsm/L | ||
| NaH2PO4 | 1.25 | |||
| NaHCO3 | 24 | |||
| Glikoz | 5 | |||
| Laktat | 1 | |||
| Kimyasal İskemi Çözeltisi | Bdt | Nacl | 136 | %5 CO2/95% O2, pH 7.4 ile kabarcıklı |
| Kartal | 3 | ~318 mOsm/L | ||
| CaCl2 | 2 | |||
| MgCl2 | 1 | |||
| NaH2PO4 | 1.25 | |||
| NaHCO3 | 24 | |||
| 2, 2-Deoksiglukoz | 2 | |||
| NaN3 | 5 | |||
| 8 mM potasyum ACSF | 8 mM K+ ACSF | Nacl | 128 | %5 CO2/95% O2, pH 7.4 ile kabarcıklı |
| Kartal | 8 | ~320 mOsm/L | ||
| CaCl2 | 2 | |||
| MgCl2 | 1 | |||
| NaH2PO4 | 1.25 | |||
| NaHCO3 | 24 | |||
| Glikoz | 5 | |||
| Laktat | 1 | |||
| Hepes-tamponlu ACSF | H-ACSF | Nacl | 125 | NaOH ile pH 7.4'e ayarlandı |
| Kartal | 3 | Her zaman kullanmadan hemen önce glikoz ekleyin | ||
| CaCl2 | 2 | Glikoz ile bir günden fazla saklamayın | ||
| MgSO4 | 2 | ~310 mOsm/L (sakaroz ile ayarlanmış) | ||
| NaH2PO4 | 125 | |||
| Hepes | 25 | |||
| Glikoz | 10 | |||
| Hanks'in Dengeli Tuz Çözeltisi | HBSS | Sigma (katalog numarası H9394). | ||
| Dulbecco Fosfat-Tamponlu Tuzlu | DPBS | Gibco (katalog numarası 14287-080) | ||
| Organotipik Kültür Orta | OTC orta | ısı yalıtımlı at serumu | 20% | 34°C, % 5 CO2, pH 7.4, culturing durumu altında |
| Mem | 79% | ~320 mOsm/L | ||
| L-glutamin | 1 | |||
| Insülin | 0.01 mg/mL | |||
| Nacl | 14.5 | |||
| MgSO4 | 2 | |||
| CaCl2 | 1.44 | |||
| Askorbik asit | 0.00125 % | |||
| D-glikoz | 13 |
Tablo 1: Çözüm kompozisyonu.
- Hazırlık gününde, ACSF glikoz ekleyin, buz üzerine yerleştirin ve en az 30 dakika için% 95 O2/ 5% CO2 ile köpürmeye başlamak 7.4 pH sonuçlandı.
- Diseksiyon ve dilimleme
- Kurban fare (BalbC, her iki cinsiyet) doğum sonrası gün 6-8 hızlı decapitation tarafından ve buz gibi ACSF içeren bir cam Petri çanak baş yerleştirin.
- Burun kemiğinin arka ucuna kadar arkadan deri keserek kafatası ortaya çıkar. Sonra, dikkatle bir cerrahi makas kullanarak foramen magnum kafatası kesilmiş ve beyin ortaya çıkarmak.
NOT: Prosedürün kurumun yönergelerine uygun olduğunu doğrulayın. - Beyni çıkarın ve ACSF ile dolu buz gibi petri kabında bir filtre zarına yerleştirin.
- Hemisferleri ayırın ve 45° açıyla bir parazitergital kesim yapın. Vibratom doku aşamasında bir yarımküreyi süper yapıştırıcı ile düzeltin. Doku bloğunu hemen buz gibi ACSF içeren vibratom banyosuna aktarın (%5 CO2/95% O2ile kabarcıklandı). Sonunda dokuyu hizala. Dilimleme kadar buz gibi ACSF ikinci yarımkürede tutun.
- 250-400 μm.'de dilimleme 250-400 μm. Dilimleme hayvan başına yaklaşık 12 dilim (400 μm: ~7 dilim) verecektir.
- Dilimi kestikten sonra(Şekil 1A),hipokampal oluşumu tipik morfolojik görünümüne göre tanımlayın(Şekil 1) ve hipodermik iğneler kullanarak izole edin (23 gauge, 1"), serebral korteksin hipokampusa bitişik olan kısmını tutarak.
NOT: Kültür sırasında neokorteksin korunması hipokampus bütünlüğünü korumaya yardımcı olur. Ancak, hipokampus izole edilebilir ve gerekirse korteks olmadan kültürlü. - Tüm dilimler toplanana kadar dilimi ısıtmalı, ACSF (34 °C, %5 CO2/95% O2)ile kabarmış bir kafesin üzerine yerleştirin.
- Dilimleri steril koşullarda devam etmek için laminar akış kabinine aktarın.

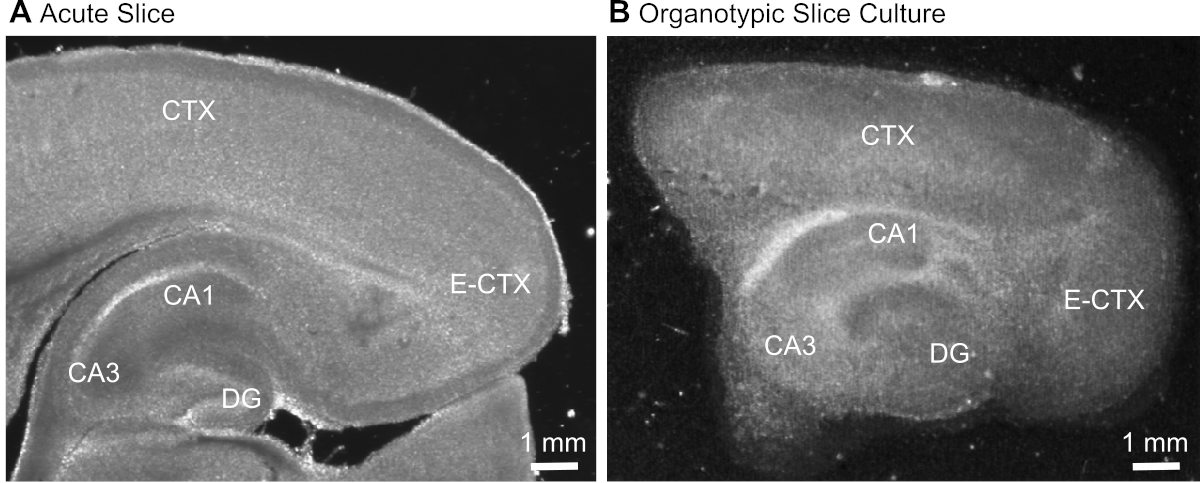
Şekil 1: Akut ve organotipik beyin dilimi preparatlarının temsili iletim görüntüleri. Akut izole parasagittal beyin diliminin(A)ve 12 gün boyunca kültürde tutulan parasagittal organotipik beyin diliminin karşılaştırılması(B) geniş alan transluminasyon mikroskopisi kullanılarak. DG = dentat girus; CA1/3 = Hipokampus'un CA1/CA3 bölgesi; E-CTX = Entorinal korteks; CTX = (neo-) korteks. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.
{kind=link}
2. Dilimleri Culturing
- Acsf'deki dilimleri ters steril cam Pasteur pipeti kullanarak steril Hank'in tuz çözeltisi ile doldurulmuş önceden ısıtılmış Petri kaplarından birine yavaşça aktarın.
NOT: Dokunun sterilizasyonu seyreltme ile sağlanır (adım 3.2). Petri kabına mümkün olduğunca az ACSF aktarın. - Pipetdeğiştirin ve ikinci Petri kabına dilimleri aktarın. İşlemi genel olarak 5 kez tekrarlayın. Aşağıdaki Petri yemeklerine mümkün olduğunca az HBSS aktarın.
- Bir dilimi kültür ekinin üstüne bir er teker yavaşça yerleştirin. Her dilim için işlemi tekrarlayın. Pipetteki türbülanslardan kaçının ve dilimin Pasteur pipetinin ucuna inmesini bekleyin. Bir tek bir membran üzerine 4 dilim kadar yerleştirebilir.
- İnce bir ipucu kullanarak hank'in çözeltisini kesici ucun üstünden dikkatlice çıkarın.
- Kültürleri(Şekil 2)deney gününe kadar gaz (carbogen, %95 O2 /5% CO2)ile 37 °C'de sıvı arasındaki arabirimde bir kuluçka makinesinde tutun. Orta yı 2-3 günde bir değiştirin.

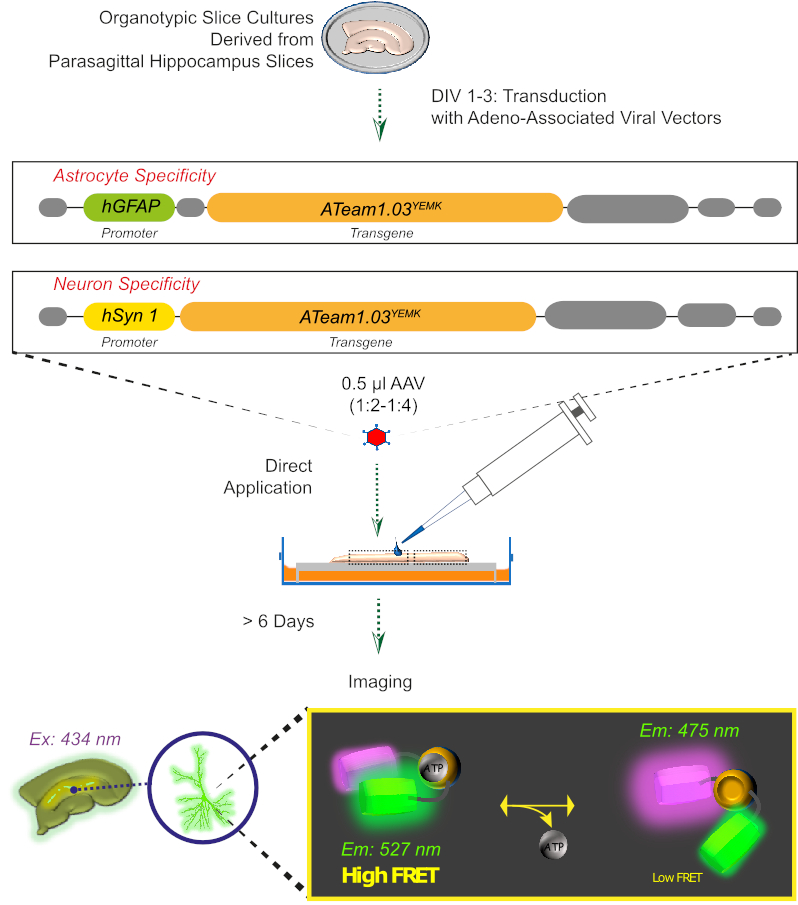
Şekil 2: Genetik olarak kodlanmış sensör ATeam1.03YEMKkullanılarak kültürlü organotipik beyin dilimlerinde FRET tabanlı ATP görüntüleme prensibi. Bu çalışmada sunulan protokolün şematik gösterimi. Kısaca, parasagittal organotipik dilimler, 1-3 gün için kültürlü, ya içeren bir adeno ilişkili viral vektör ile transduced, astrosit özgü hGFAP- veya nöron özgü hSyn-promoter ve ATeam1.03YEMKifade için sırası . Bu vektörlerin seyreltilmiş aliquots (1:2-1:4) doğrudan en az 6 gün daha culturing koşulları altında muhafaza edilir bir dilim, üstüne uygulanır. Hücre içi ATP düzeylerindeki değişiklikler daha sonra sensörü 434 nm'de heyecanlandırarak ve aynı anda 527 (alıcı) ve 475 (donör) nm'de floresan emisyon elde ederek sensörü ifade eden hücrelerde görselleştirilebilir. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.
{kind=link}
3. Aadeno ile Ilişkili Viral Vektörile ATP Sensörlerinin İfadesi (Şekil 2)
NOT: Genetiği değiştirilmiş organizmaların kullanımı için tüm gereksinimleri karşılamak için emin olun!
- Viral vektörü işlemek için, aliquot adeno ile ilişkili viral vektörler (AAV2/5) 1-2 μL'de tekrarlanan donma ve erimeyi önler. Aliquots'u -80 °C'de saklayın.
- Vektörle temas eden tüm kullanılmış artık malzemeleri atmak için steril tezgahın üzerine %10 çamaşır suyu içeren bir şişe yerleştirin.
- Transdüksiyon için, 2-3 μL DPBS ile vektörün 1 μL'lik seyreltilmesini hazırlayın. Stok çözeltileri normalde mL başına 1012 viral genom (vg/mL) büyüklüğünde fiziksel bir titrek sergiler.
- Kültürlü bir dilim içeren bir kesici ucu steril kaputa aktarın.
- Dokuya dokunmadan seyreltilmiş vektörün 0,5 μL'sini doğrudan her dilimin üstüne uygulayın.
NOT: Doku derin tabakalarında daha iyi ifade 1-3 gün in vitro (DIV) kültürlü dilimler transducing tarafından elde edilir. Eski kültürlerin transdüksiyon uymak çevredeki glial ykar veya nöronlarda düşük bir ifade hücrelerin baskın bir ifade neden olabilir, sırasıyla. - Son olarak, dilimleri kuvöze geri yerleştirin ve en az 6 gün daha saklayın. Transdüksiyon gününde ortamı değiştirmeyin.
4. Glial Yaranın Çıkarılması (Şekil 3)
- Bir denemeye başlamadan hemen önce, kültürlü dilimler içeren bir kesici ucu steril kaputa aktarın ve 1 mL OCT orta veya MEM içeren 30 mm'lik bir tabağa yerleştirin.
- Tabağı stereoskopun altına yerleştirin ve dilimin yüzeyine odaklanın.
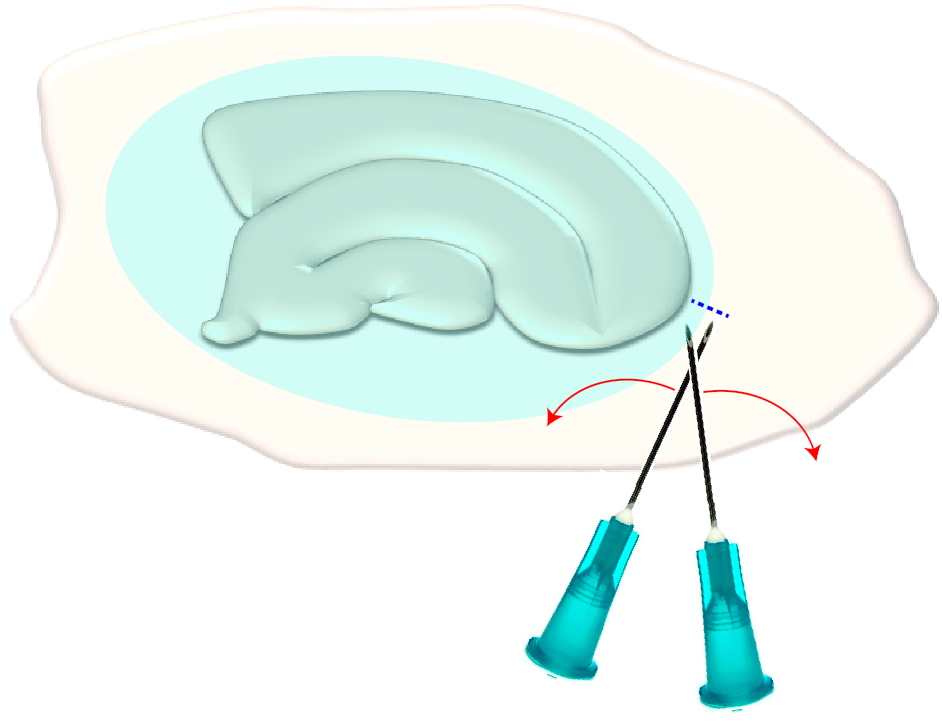
- Seçilen dilimin dar kenarlarında kısa bir çapraz kesme yapmak için iki steril hipodermik iğne (23 G, 1") kullanın(Şekil 3). Bu yordam, glial yaranın oluşturduğu yüzeydeki gerilimi serbest bırakarak geri çekilmesine neden olur ve böylece altta yatan katmanları açığa çıkarır (Bkz. Şekil 5).
NOT: İlk doku tabakası (glial skar) esas olarak reaktif astrositler tarafından oluşturulur. Geniş alan görüntülemede, bu yoğun doku tabakası ışığın ek saçılmasına neden olacak ve bulanık görüntülere yol açacaktır. Yara izi kaldırılması böylece uygun organotipik doku içeren derin katmanları, daha iyi görünürlük elde etmek için avantajlıdır. Bu nedenle, sadece kenarına ve sadece dilim hazırlanması üst tabakasında bu kesim gerçekleştirmek için dikkatli olun ve altındaki doku zarar değil. Otcs'de glial skar lı verilerile skarsız OTC'lerden elde edilen veriler arasındaki farkları gözlemlemedik (veriler gösterilmedi). - Hazırlanan dilimi kesici uçtan çıkarın. Bu amaçla, steril bir neşter kullanın ve membran düz paralel kesim yaparak çıkarmak, ortasında dilim ile bir kare veya üçgen oluşturarak, cımbız ile membran kenarları tutarken. Kesici uçek dilimler barındırıyorsa, orijinal plakaya ve kuluçka makinesine geri aktarın. Ortamın yüzey gerilimi membran yüzeyine sızmasını önleyecektir.

Şekil 3: Glial yaranın mekanik olarak çıkarılmasının şematik çizimi. Şekil glial bir yara izi (mavimsi elipsoid) ile kaplı bir hipokampal dilim kültürü gösterir. Bir kez kültürün en küçük kutup ve glial yara (mavi kesikli çizgi) kenarında iki şırınga iğneuçları kesme, yara bir kenara çevirmek olacaktır. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.
{kind=link}
5. FRET tabanlı ATP Görüntüleme (Şekil 4)
- Deneyden önce, 7,4 pH elde etmek için en az 30 dakika boyunca %95 O2/5%CO2 ile E-ACSF hazırlayın ve kabar. Monokromatörün floresan ışık kaynağını (Xenon lambası) açın (Şekil 4). Kuvözdeki dilimi çıkarmadan hemen önce perfüzyonu başlatın.
NOT: Tüm deney boyunca tuzlu su labirlikte %95 O2 ve %5 CO2 ile kabardırın. - Dilimi, peristaltik pompa kullanılarak sürekli olarak yeni karbogenated E-ACSF ile perfüzyonlanmış deneysel bir hazneye aktarın(Şekil 4). Sonra, bir ızgara ile dilim düzeltin. Odayı mikroskop aşamasına yerleştirin ve perfüzyon sistemini bağlayın. Perfüzyon için gaz geçirmez laboratuvar tüpü önerilir.
NOT: Deneyler, deney tasarımına bağlı olarak oda sıcaklığında veya fizyolojik sıcaklığa yakın bir yerde yapılabilir. Dokunun hareketi ve/veya kesme stresindeki değişikliklerden kaynaklanan odak değişikliklerini önlemek için perfüzyon akışının stabilitesini ve güvenilirliğini kontrol edin. Bizim ve diğer birçok laboratuvar tarafından kullanılan dilim leme için standart perfüzyon hızları 1,5-2,5 mL/dk'dır.

Şekil 4: FRET görüntüleme kurulumunun konfigürasyonu. (A) Farklı bileşenlerin şematik illüstrasyonu ve FRET görüntüleme kurulumu için gerekli mekansal düzenlemesi. Düzenleme, ışık kaynağı olarak kenon lambası olan bir monokromatör, dik sabit kademeli mikroskop (1), görüntü ayırıcı sistemi (2), zaman atlamalı kayıt için dijital CCD veya CMOS kamera (3) ve kararlı sabit perfüzyon için uyarlanmış deneysel bir banyodan (4) oluşmaktadır. Banyo perfüzyonu ayarlanabilir akış hızına sahip peristaltik bir pompa ile gerçekleştirilir. (B) Deneysel çalışma alanının görüntüsü. FRET görüntüleme kurulumu, x/y-çeviri aşamasına sahip titreşim sönümlü bir masaya monte edilmiştir ve deneysel banyo gömülüdür. Sayılar: bakınız (A). (C) Monokromatöründen dijital kameraya ışık yolunun şematik görünümü. Belirtilen farklı filtreler ve dikroik ayna konumudur. Sayılar: bakınız (A). Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.
{kind=link}
- İletim ışığını kullanarak kültürlü dilimi odak landırın. Deneylerin yapılacağı alanı belirleyin (örnek: Hipokampus'un CA1 bölgesi). Görüntüleme denemelerine başlamadan önce, dilimlerin tuzlu koşullara uyum sağlaması için en az 15 dakika bekleyin. Deneysel kurulumun yapılandırması için Şekil 4'ebakınız.
- Kamerayı ve görüntüleme yazılımını açın. Ardından, uygun filtre küpünü seçin.
- 435/17 nm (~435 nm) de donör floresan protein (eCFP) heyecanlandırmak. Pozlama süresini 40 ila 90 ms arasında ayarlayın.
NOT: Dilimlerin floresan ışığa güçlü bir şekilde maruz kalmasına fototoksik etkilerneden olabilir. - 435 nm'de uyarma hem 475 nm (eCFP; donör) hem de 527 nm (Venüs; kabul edici) emisyona neden olur. Floresan emisyonuna emisyon farkını 500 nm'ye bir emisyon görüntü ayırıcıile bölün ve donör ve kabul edilebilir floresanyı daha fazla izole etmek için 483/32 ve 542/27 bant geçiş filtrelerini çalıştırın. Güçlü ifade dedektörlerin doygunluğu ile sonuçlanabilir. Bu durumda, uyarma yoğunluğunu azaltmak için nötr bir yoğunluk filtresi kullanabilirsiniz.
- Arka plan çıkarma için hücresel floresan yoksun görünen bir ilgi alanı (RoI) seçin. Sonra, HÜCRE cisimlerini delineating ROI oluşturmak.
- Görüntü edinme sıklığını ve genel kayıt süresini ayarlayın. Uzun (>30 dk) deneylerde fototoksisiteyi önlemek için 0.2-0.5 Hz'lik bir kazanım sıklığı önerilir.
- Kaydı başlatın. Bu hazırlık istikrarı sağlamak için temel koşullar altında en az 5 dakika kaydetmek için tavsiye edilir.
NOT: Gerekirse kayıt sırasında hücrenin odağı ayarlayın. - Hücre içi ATP'de değişiklikler emdirmek için perfüzyon tüpünü standart ACSF'den metabolik inhibitörler içeren bir saline aktarın (örneğin BDT, Bkz. Tablo 1 ve aşağıya bakınız). Alternatif olarak, aktif nöronlardan potasyum salınımını taklit etmek için yüksek potasyum konsantrasyonu ile bir tuzlu kullanın.
NOT: Banyo perfüzyonu ile uygulama küresel tüm hazırlık üzerinde hareket nispeten yavaş bir süreçtir. Yeni çözümün deneysel banyoya girmeye başladığı zamana dikkat edin. Oda ile tuzlu hazne arasındaki mesafeye ve perfüzyonun hızına bağlı olarak gecikme süresi göz önünde bulundurulmalıdır.
6. Hücresel ATeam Floresan Yüksek Çözünürlüklü Dokümantasyon
- Kayıtlardan hemen sonra, dilim kültürünü içeren kayıt odasını konfokal lazer tarayın mikroskobuna aktarın.
NOT: Özel ilgi yiy. Olası fotohasar nedeniyle, bu adımı yalnızca denemelerden sonra gerçekleştirin. Dokümantasyon amacıyla, E-ACSF'yi H-ACSF ile değiştirebiliriz. Bu nedenle, bir perfüzyon sistemi mutlaka gerekli değildir. - Verilen optik yapılandırmada mümkün olan en yüksek z çözünürlüğünde z-yığınları alın.
- Görüntü çözünürlüğünü artırmak için bir deconvolution algoritması uygulayın.
Sonuçlar
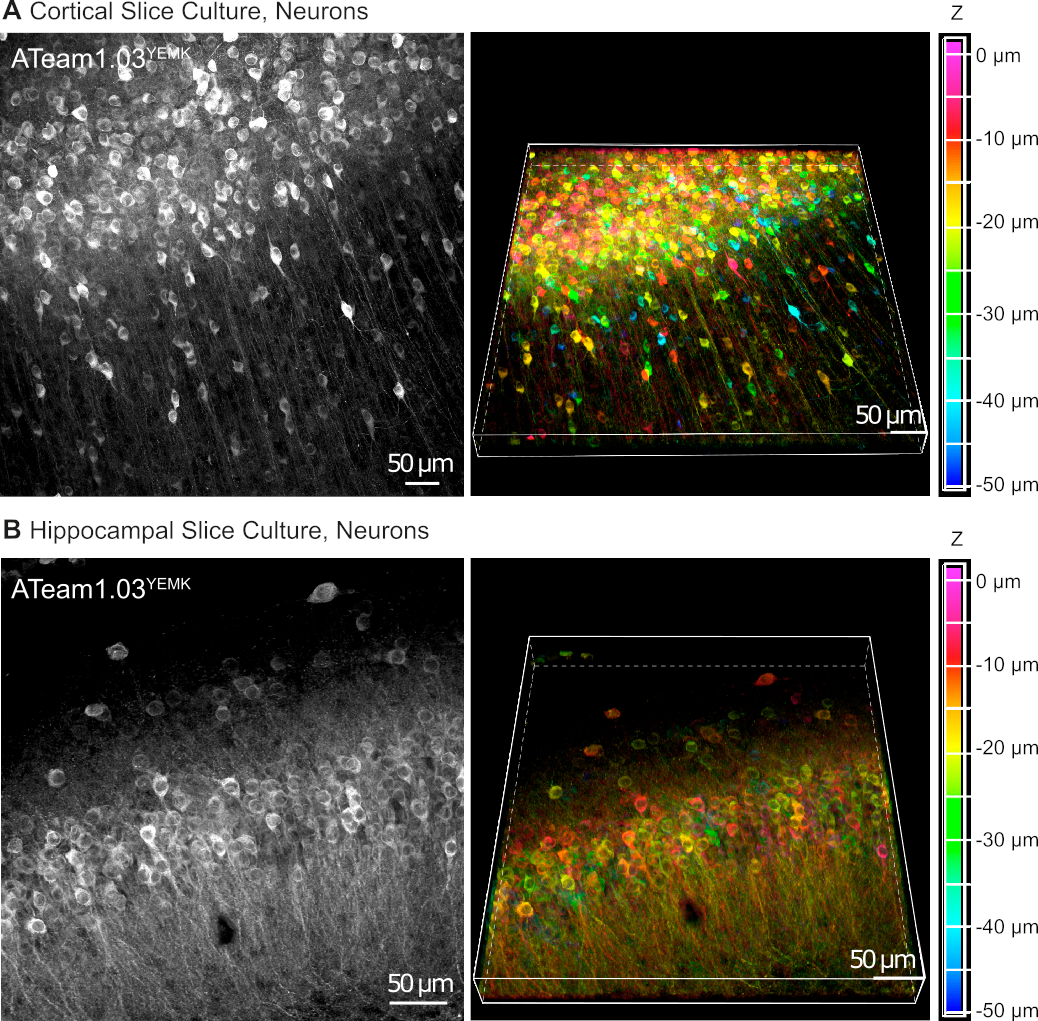
AAV vektörleri, canlı doku içindeki hücrelerdeki yabancı genleri seçici olarak ifade etmek için güvenilir bir araçtır16. ATeam1.03YEMK'ın sıralı kasetini ve belirli bir organizatörü içeren ATV'lerin doğrudan uygulanması, seçilen hücre tipinde sensörün yüksek bir şekilde ifade edilebilmelidir. DIV 14'te (transdüksiyondan ~10 gün sonra), insan sinapsin organizatörü altında ATeam'i ifade eden nöronlar, dilim yüzeyinin 50 μm altında, kültürlü doku dilimlerinin neokorteksinde yüksek yoğunlukta bulunurlar(Şekil 5A). Hipokampusta karşılaştırılabilir sonuçlar elde edilebilir (Şekil 5B).

Şekil 5: Kültürlü parasagittal organotipik beyin dilimlerinde ATeam1.03YEMK'yi ifade eden nöronların görselleştirilmesi. Soldaki görüntüler, kortikal dokunun43optik kesitinin (her biri 1,05 μm) ve(B)70 optik kesitinin (her biri 0,6 μm) genişletilmiş odak projeksiyonlarına karşılık gelmektedir. Sağdaki görüntüler aynı projeksiyonun ses görünümünü temsil eder. Hücreler, sağdaki renk skalası ile belirtildiği gibi dilim yüzeyine göre derinliklerine göre renk kodludur. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.
{kind=link}
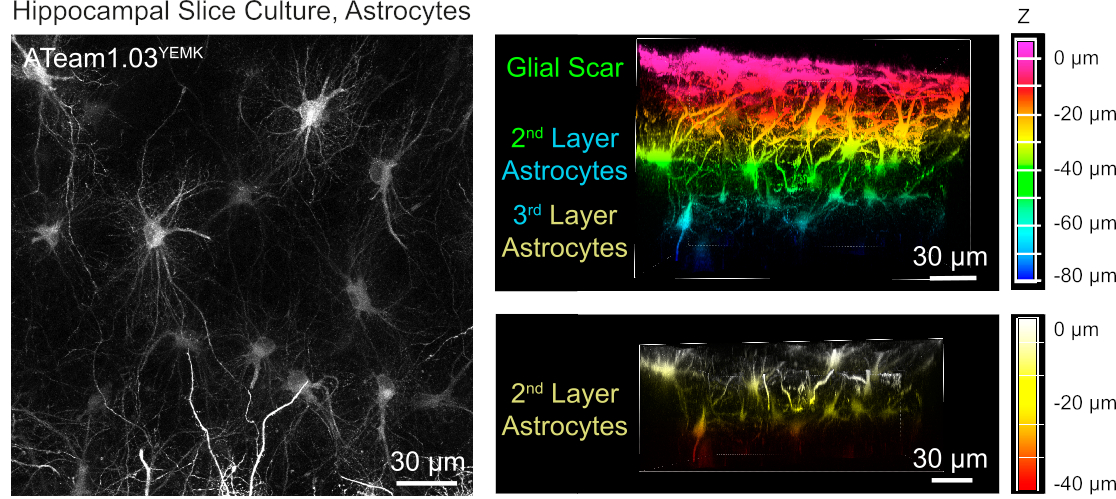
Astrositlerde ATP düzeylerinin ölçülmesi için, ATeam1.03YEMK insan glial fibrillary asidik protein (GFAP) organizatörü kontrolü altında ifade edilir. Bu da hem neokortekste hem de kültürlü doku dilimlerinin hipokampusundaki hücrelerin etkin transdüksiyonuna yol açar(Şekil 6). Özellikle, dilim preparatlarının yüzeyine göre derinliğe bağlı olarak iki farklı morfolojik fenotip ayırt edilebilir. Birinci yüzeysel tabakada hücreler, ağırlıklı olarak yüzeye paralel olarak düzenlenmiş kalın birincil süreçler ile karakterize edilir. Bu hücreler güçlü bir şekilde örtüşen etki alanları sergilerler ve görünüşe göre reaktif astrositlerden oluşan yoğun bir örgü oluştururlar(Şekil 6). Daha derin katmanlarda (yüzeyden 30-60 μm), transduced astrositler büyük ölçüde küresel etki alanları oluşturan ince hücresel süreçler sergilerler ve morfolojileri daha önce bildirilen19,20,21 (Şekil 6)yerinde astrositlere benzer. Derin tabaka astrositlerin daha iyi transdüksiyonu ve bu derin katmanlara daha iyi optik erişim elde etmek için, glial skar dokusu Adım 4'te açıklandığı gibi çıkarılabilir.

Şekil 6: Kültürlü parasagittal organotipik beyin dilimlerinde ATeam1.03YEMK'yi ifade eden astrositlerin görselleştirilmesi. Soldaki görüntü, 191 optik kesitten (her biri 0,45 μm) oluşan genişletilmiş bir odak projeksiyonuna karşılık gelir. İllüstrasyon amacıyla, glial yara astrositprojeksiyon dışında tutuldu. Sağdaki görüntüler, glial yaranın çıkarılmasından önce ve sonra aynı projeksiyonun ses görünümünü temsil eder. Hücreler, sağdaki renk ölçeklerinde belirtildiği gibi dilim yüzeyine göre derinliklerine göre renk kodludur. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.
{kind=link}
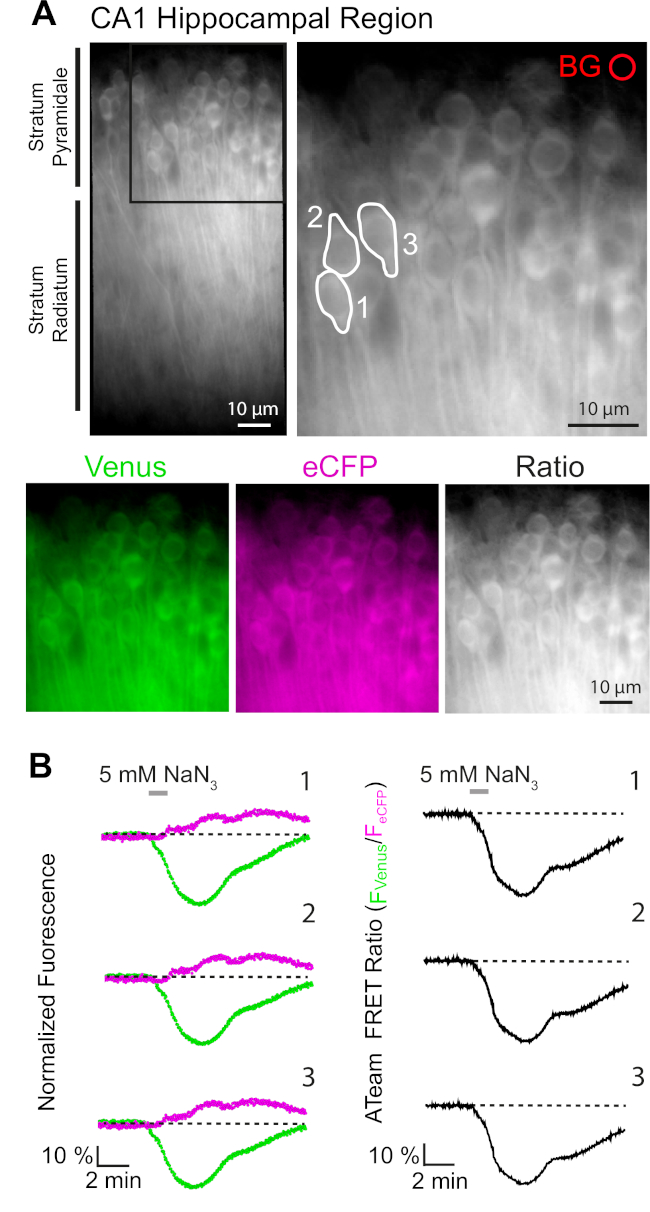
ATeam1.03YEMK'ın başarılı bir şekilde ifade edileb1/400r, kullanılan organizatöre bağlı olarak nöronveya astrositlerdeki ATP düzeylerindeki değişikliklerin dinamik ölçümüne olanak tanır (yukarıya bakın). Deneyler sürekli E-ACSF ile perfüzyonlu deneysel bir banyoda yapıldı (%95 O2/5%CO2ile kabarmış). Hipokampal nöronlarda ATeam1.03YEMK'yi ifade eden organotipik dilimlerde, kayda başlamadan önce ilgi bölgeleri (ROI) seçilmiştir ve piramidal hücrelerin somata'sını temsil eder(Şekil 7A). Ayrıca, arka plan çıkarma için bir bölge seçilmiştir (Şekil 7A). Venüs'ün emisyon ve eCFP floresansı bu ROI'ların her biri için ayrı ayrı toplanmış ve zaman içinde floresan emisyon düzeyi olarak gösterilmiştir(Şekil 7B). Floresan'ı kontrol koşulları altında birkaç dakika boyunca kontrol altında kaydettikten sonra, hücre metabolizması dilim preparatını glikozsuz saline maruz bırakarak inhibe edildi ve 5 mM sodyum azid (NaN3)bir dakika süreyle eklendi (Şekil 7B). Bu manipülasyon, Venüs 'ün (527 nm) azalması ve eCFP (475 nm) emisyon artışı ile FRET çiftinin emisyon yoğunluğunda(Şekil 7B, sol paneller) zıt değişikliklere yol açarak, bu yönde bir artışa neden oldu. Venüs'ün floresan emisyonunu eCFP (FVenüs/FeCFP)ile bölerek FRET oranının hesaplanması, hücre içi ATP düzeylerindeki göreceli değişiklikleri yansıtan sinyallerle sonuçlandı, sözde "ATeam FRET oranı"(Şekil 7B, sağ panel). Kaydedilen tüm nöronlarda (N = 5 dilimdeki n = 70 hücre), NaN3 ATeam FRET oranında geri dönüşümlü bir azalmaya neden olarak hücre içi ATP düzeylerinde hücre içi atp düzeylerinde hücresel metabolizmanın inhibisyonu sırasında geri dönüşümlü bir düşüşe neden oldu.

Şekil 7: Zaman atlamalı ATeam FRET oranı görüntüleme gösterimi. (A) Sol üst: Nöronlarda ATeam1.03YEMK ifade kültürlü bir organotypic hipokampal dilim in ca1 bölgesinin piramit tabaka ve stratum radiatum geniş alan floresan görüntüsü. Sağ üst: Solda belirtildiği gibi kutulu bölümün genişletilmiş görünümü. Beyaz çizgiler ilgi bölgeleri (ROI) 1-3 ca1 piramidal nöronların hücre organları temsil eden(B)analiz için seçilen. BG, arka plan düzeltmesi için seçilen YG'yi temsil eder. Alt: Venüs floresan emisyon (yeşil), eCFP (mor) ve Venüs/eCFP oranını temsil eden sözde renkli görüntüler. (B) RoIs 1-3'te nöronal hücre cisimlerini temsil eden zaman atlamalı kayıt (bkz. A). Soldaki izler Venüs (yeşil) ve eCFP (macenta) floresan emisyonnormalize göstermektedir. Sağdaki izler ilgili ATeam FRET oranını gösterir. 1 dakika boyunca ekstrasellüler glikoz yokluğunda 5 mM NaN3 ile perfüzyonun ATeam FRET oranında geri dönüşümlü bir azalmaya neden olduğunu ve hücre içi ATP konsantrasyonunda bir düşüş olduğunu unutmayın. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.
{kind=link}
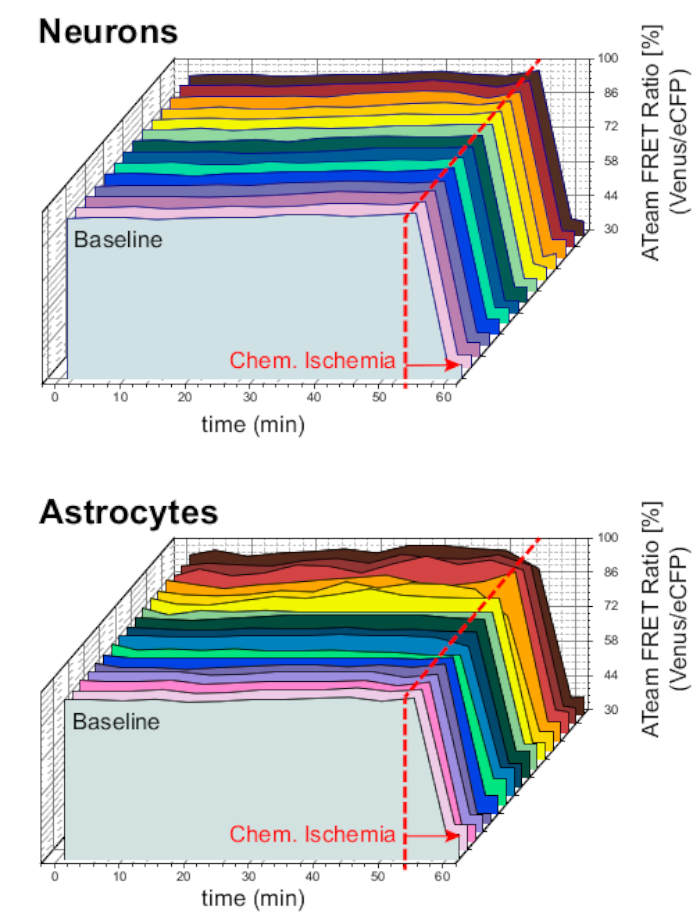
Uzun süreli deney koşullarında preparatın ve sensörün stabilitesini sağlamak için, nöronlarda veya astrositlerde ATeam'i ifade eden dilimler uzun süre ACSF ile sürekli olarak perfüzyona maruz kalmıştır (>50 dk; n = 12 hücre, Her biri 3 beyinden N = 3 OTC). Bu koşullar altında, ATeam FRET oranı değişmedi (Şekil 8). Preparatların metabolik inhibitörler içeren BDT'ye teşhir edilmesi ("Kimyasal iskemi", tablo 1'ebakınız), buna karşılık, yukarıda da görüldüğü gibi ATeam FRET oranında beklenen düşüşe yol açmıştır.

Şekil 8: ATeam kullanan temel deneyler. Nöronlarda (üstte) ve astrositlerde (altta) temel koşullar altında 14 farklı hücrede uzun süreli ATeam FRET oranı görüntüleme. Veriler diğer deneysel verilerle karşılaştırılabilir koşullar altında alınmıştır. Her ölçümün sonunda, okla belirtildiği gibi BDT ile perfüzyon ile kimyasal iskemi ortaya çıkarıldı. NOT: Temel ATeam FRET oranları, temel koşullar altında zaman içinde sabittir. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.
{kind=link}
Daha sonra, ateam1.03YEMK ifade nöronve astrositlerin yanıtlarını ekstrasellüler potasyum konsantrasyonundaki artışa analiz ettik. Stabil bir bazin oluşturduktan sonra nöronlar potasyum konsantrasyonu 3 dakika boyunca 3 ila 8 mM'ye yükseltildiği bir tuzlu luk la perfüzyona bünyemiştir(Şekil 9A). Ancak bu manipülasyon, ATeam FRET oranında (N = 5 diliminden 56 hücre) saptanabilir bir değişikliğe yol alamayabilir bir durumdur. Sensörün ATP düzeylerindeki bir değişikliğe tepki göstermesini sağlamak için, dilimler BDT tarafından E-ACSF'nin değiştirilmesiyle ortaya çıkan sürekli bir kimyasal iskemi dönemine tekrar maruz kalmıştır. Kimyasal iskemi, ATeam FRET oranında yeni, kararlı bir seviyeye doğru hızlı bir düşüşe yol açarak ATP'nin 2-3 dakika sonra nominal olarak tükendiğını gösterir(Şekil 9A).

Şekil 9: Nöronlar ve astrositlerdeki ATP düzeylerindeki değişiklikleri gösteren temsili deneyler. (A,B): Soldaki görüntüler, organotipik dilimlerin hipokampal CA1 bölgesinde bulunan nöron ve astrositlerden Gelen ATeam floresansını göstermektedir. Sağdaki izler, tek bir hücre gövdesi üzerinde konumlandırılmış bir Yatırım Getirisi'nden elde edilen ATeam FRET oranının zaman atlamalı kayıtlarını temsil eder. Her iki deneyde de, dilimler ilk olarak 3 dakika boyunca hücre dışı potasyum konsantrasyonunda bir artışa maruz kalındı (bkz. bar), ardından kimyasal iskemiye son maruz ilerler. Nöronlar ekstrasellüler potasyum yüksekliğine yanıt vermezken(A),astrositler ATP bir artış ile reaksiyona (B). Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.
{kind=link}
Aynı deneysel protokol, ATeam1.03YEMK'nin astrositlerle ifade edildiği dilimlerle gerçekleştirilmiştir. Nöronlarda gözlenenin aksine, astrositler ATeam FRET oranında geri dönüşümlü bir artış la hücre dışı potasyum artışına tepki göstererek hücre içi ATP düzeylerinde bir artış olduğunu (N = 5 dilimdeki n = 70 hücre)(Şekil 9B). Kimyasal iskemi sonraki maruz kalma sonuçlandı, beklendiği gibi, ATeam FRET oranında büyük bir düşüş, hücre içi ATP nominal tükenmesi göstergesi(Şekil 9B).
Tartışmalar
Burada, ATeam1.03YEMKhücre tipi özel ifade için bir prosedür göstermek , bir FRET tabanlı, genetik olarak kodlanmış nanosensor14, fare beyninin organotipik doku dilimi kültürlerde astrosit veya nöronlarda ATP düzeylerinde değişikliklerin ölçümü için15. Örnek kayıtlarda, hücre dışı potasyum konsantrasyonundaki artışın nöronlardaki ATP konsantrasyonlarında bir değişikliğe yol açmadığını, astrositik ATP düzeylerinin ise bu manipülasyona yanıt olarak yükseldiğini gösteriyoruz. Ayrıca, sonuçlarımız hücre metabolizmasının inhibisyonu üzerine, ATeam1.03YEMK FRET oranının her iki hücre tipinde de hızla düştüğünü ve hücre içi ATP'de hızlı bir düşüş olduğunu göstermektedir.
Organotipik dilim kültürlerinde ATeam1.03YEMK ifadesi en az 7-10 gün kontrollü koşullarda kültürde doku bakımı gerektirir. Alternatif olarak, ATeam1.03YEMK da akut izole beyin dokusu dilimleri ve farelerin optik sinirlerde ATP ölçümü için istihdam edilebilir13,15. Ancak akut izole dokudaki ölçümler, transgenik hayvanların neslini veya viral vektörlerin beyne stereotaktik bir şekilde uygulanmasını, hayvan deneyleri ve sıkı hayvan bakım protokollerini içerir. Bu bağlamda, organotipik doku dilimi kültürlerde ATeam1.03YEMK ifadesi yararlı ve değerli bir alternatif22,23temsil eder. Uzun yıllardır, organotipik doku dilimi kültürleri nöral özellikleri, bağlantı ve geliştirme24,25,26çalışma için kurulmuş bir model sistemi olarak hizmet vermektedir. Onlar sadece genel doku mimarisi ve laminasyon korumak değil(Şekil 1),aynı zamanda üstün erişilebilirlik ve deneysel koşulların doğrudan kontrolü gibi hücre kültürlerinin tercihli özellikleri ev sahipliği. Organotipik doku dilimi kültürleri de rutin viral vektörler27kullanarak yabancı genleri ifade etmek için istihdam edilmektedir. Viral vektörler çeşitli beyin dokusu içine transgenler teslim bildirilmiştir16,28. Adenoviral vektörler glial hücrelerde yüksek ekspresyon indükler, ama hipokampal nöronlar16değil , ve glial reaktivite üretebilir17. Burada kullanılan Adeno ilişkili viral vektörler iyi bir alternatif olarak ortaya15, ve bunların etkinliği de in vivo29gösterilmiştir .
Esas olarak nöronal özelliklerin incelenmesinde kullanılırken, son çalışmalar da astrositlerin analizi için organotipik doku dilimi kültürleri istihdam edilebilir olduğunu ortaya kurduk. Kültürlü dilimler genellikle reaktif astrositler bir tabaka ile kaplıdır19,30 (Şekil 5),ancak astrositler daha yerli, non-reaktif morfoloji ve sitomimari daha derin katmanlarda sergiler19,30 ( Şekil5). Bu çalışmada, dış glial skar mekanik kaldırma için bir prosedür tarif, hangi uygun organotipik doku katmanları içinde yerli astrositlerin daha iyi deneysel ve optik erişilebilirlik ile sonuçlanır. Ayrıca, kaldırılması organotipik dilimlerin daha derin katmanlarında ifade etkinliğini artırır; glial yara izi çıkarılmazsa, AEV'lerin transdüksiyonu yüzeysel hücre tabakaları ile sınırlı olabilir.
Doku dilimleri üzerinde deneyler yaparken çeşitli dış mekanik faktörlerin göz önünde bulundurulması gerekir. Banyo perfüzyon hızındaki bir değişim tüm hazırlık hareketlerini tetikleyebilir ve/veya odak değişikliklerine neden olabilir ve sensör sinyalinin yapay geçici değişikliklerine neden olabilir. Ayrıca, hem astrositler ve nöronlar gibi yüksek perfüzyon oranları 32 tarafından empoze mekanik deformasyona yanıt bildirilmiştir32,33. Elimizde, güvenilir bir peristaltik pompa kullanarak, doku ve amaç (menisküs) arasında tuzlu küçük ve kararlı hacimleri korumak ile birlikte burada kullanılan perfüzyon hızı (1.5-2.5 mL/dk) temel koşullar altında istikrarlı bir FRET sinyali ile sonuçlanır; Şekil 8).
Bu çalışmada, aTeam1.03YEMK ile FRET tabanlı görüntüleme nöronlar ve astrositlerde ATP düzeylerini izlemek için istihdam edilebilir olduğunu göstermektedir. Hücresel ATP ölçümü için daha önce tanıtılan alternatif bir araç sözde luciferin-luciferase assay34,35,36,37. Bu yaklaşım, ancak, biyolüminesans görüntüleme dayanmaktadır ve sadece kısmen yüksek arka plan gürültü düzeyleri nedeniyle oldukça düşük zamansal ve mekansal çözünürlük sağlar. Son yıllarda rutin olarak kullanılan bir diğer yöntem de iyona duyarlı florofor magnezyumyeşili 38,39,40kullanarak hücre içi magnezyum konsantrasyonundaki değişikliklerin görüntülenmesiydi. Bu yaklaşım, ATP tüketiminin ortak faktörlü magnezyumsalınımına yol açan gözlemle ilgilidir. Magnezyum yeşili ile görüntüleme, atp düzeylerindeki değişikliklerin sadece ikincil bir tahminini sağlar. Ayrıca, magnezyum yeşili de hücre içi kalsiyum değişikliklere duyarlı, bu yöntem ile elde edilen sonuçları yorumlarken başka bir zorluk tanıtAn.
Hücresel metabolitlerin doğrudan görüntüleme için genetik kodlanmış nanosensörlerin son gelişme, bu nedenle, ileriye doğru büyük bir adım temsil11,12. Hücre içi ATP36,41,42,43ölçümü için kullanılabilir birkaç farklı sensörler oluşturuldu. Bunlar arasında oranmetrik floresan ATP göstergesi "QUEEN"41 yanı sıra PercevalHR, hangi ATP algılar:ADP oranı42. İkinci sonda hücrelerin enerji durumunun incelenmesi için değerli bir araç olsa da, pH42'dekideğişikliklerin eşzamanlı olarak ölçülmesini gerektirir.
ATeam çeşitli varyantları var olan bir nanosensor, hangi - diğerleri arasında - ATP14için bağlayıcı yakınlık farklıdır. In vitro, ATeam1.03YEMK 37 °C'de 1.2 mM Kd sergiler, hipotalamus ve serebellum 34 hipokampus37 arasında değişen farklı nöronal hücre tiplerinde belirlenen hücresel ATP düzeylerine yakın, hipotalamus37,44,45. Cuvette ölçümlerinde sıcaklığın 10 °C düşürülmesi, ATeam1.03YEMK'ın ATP'ye olan bağlanma yakınlığında önemli bir azalmaya yol açarak,14numaralı oda sıcaklığında hücresel görüntüleme için ideal olmayabileceğini düşündürmektedir. Daha önceki çalışmamız15,ancak, davranış ve yanıt ATeam1.03YEMK farklı manipülasyonlar nöronlar ve astrositler ifade benzer fizyolojik ve oda sıcaklığında, sensör her iki koşulda hücre içi ATP düzeylerinin güvenilir belirlenmesine olanak sağladığını gösteren gösterdi. Buna ek olarak, daha önceki deneylerimizde ATeam1.03YEMK'ın pH duyarlılığıelealınmıştır. Düşük mM aralığındaKi Kd bir sorun sayılsa, alternatif ATeam varyantları14kullanılabilir , bunların arasında ATeam'in kırmızı vitesli varyantları ("GO-ATeam")43.
ATeam1.03YEMK kullanarak yapılan deneyler, ekstrasellüler potasyum konsantrasyonunda sadece birkaç mM (3-8 mM) artış, organotipik dilim kültüründeki astrositlerde ATeam1.03YEMK oranında geçici bir artışa yol açtığını göstermektedir. Bu gözlem önceki çalışmalarda doğrular15,46 ve açıkça astrositler aktif nöronlar tarafından atp üretiminde bir artış ile potasyum salınımına yanıt gösterir, çoğunlukla Na bir uyarılma sonucu muhtemel+/ -ATPase ve Na+/ HCO3- cotransporter, sırasıyla47,48. Bunun aksine, nöronlar bir yanıt göstermedi, hangi önceki çalışma ile uyumlu da15. Her iki hücre tipi, ancak, hızlı ve güçlü hücresel glikoliz ve mitokondriyal solunum inhibisyonu için tepki15önce gösterildiği gibi . Kimyasal iskemi koşulları altında, ATeam FRET oranları hücresel ATP nominal tükenmesi gösteren, yeni bir istikrarlı seviyeye düştü. İkinci sonuç hem nöronlar hem de astrositler de sinaptik aktivasyon veya nörotransmitterlerin uygulanması ile ek uyarılma olmadan sabit devlet koşulları altında ATP ilgili bir tüketim sergilemek düşündürmektedir. Birlikte ele alındığında, genetik olarak kodlanmış nanosensörler ile FRET tabanlı görüntüleme, aralarında ATeam1.03YEMK,farklı koşullar altında hücre içi ATP düzeyleri ve hücresel ATP tüketimi değişiklikleri sorumlu hücresel süreçleri açıklamak için değerli bir yaklaşım sağlayacaktır sonucuna varıyoruz.
Açıklamalar
Yazarlar hiçbir rakip çıkarları beyan. Yazarlar, video makalesinde kullanılan araçları üreten Nikon Microscope Solutions, Düsseldorf, Almanya tarafından açık erişimli yayın sağlayan mali destek aldı. Şirket ne burada sunulan deneylerin tasarımında, ne yürütülmesinde, ne veri işlemede, ne de el yazması yazıda yer aldı.
Teşekkürler
Yazarlar uzman teknik yardım için Claudia Roderigo ve Simone Durry teşekkür etmek istiyorum. Dr. Niklas J. Gerkau ve M.Sc. Joel Nelson'a organotipik dilim kültürlerin hazırlanmasında yardımcı olan yardımcılarından dolayı teşekkür ederiz. Yazarın laboratuvarındaki araştırmalar Alman Araştırma Birliği (DFG) tarafından finanse edildi; 2795 İçİn: Ro 2327/13-1 ve SPP 1757: Ro 2327/8-2 CRR için; ve SPP 1757: Genç Glia Start-Up rl finansman).
Malzemeler
| Name | Company | Catalog Number | Comments |
| 2-deoxyglucose | Alfa Aesar | L07338 | Non-metabolizable glucose analog |
| 36-IMA-410-019 Argon laser | Melles Griot | 488 nm wavelength argon | |
| Ascorbic acid | Carl Roth | 3525.1 | Antioxidant, Vitamin C |
| band pass filters 483/32 | AHF Analysentechnik AG | Splitter compatible emmision filter | |
| band pass filters 542/27 | AHF Analysentechnik AG | Splitter compatible emmision filter | |
| Beamsplitter T 455 LP | AHF Analysentechnik AG | Excitation dichroic mirror | |
| Beamsplitter T 505 LPXR | AHF Analysentechnik AG | Splitter dichroic | |
| Confocal laser scannig microscope C1 | Nikon Microscope Solutions | Modular confocal microscope system C1 | |
| Data processing Origin Pro 9.0.0 (64-bit) | OriginLab corporation | Scientific graphing and data analysis software | |
| D-glucose monohydrate | Caelo | 2580-1kg | |
| DPBS | GIBCO/Life | 14190250 | Dulbecco's phosphate-buffered saline |
| Eclipse E 600FN upright microscope | Nikon Microscope Solutions | ||
| Eclipse FN1 upright microscope | Nikon Microscope Solutions | ||
| Experimental chamber | custom build | Perfusion chamber for live-cell imaging | |
| EZ-C1 Silver Version 3.91 | Nikon Microscope Solutions | Imaging software for confocal microscope | |
| Hanks' Balanced Salt solution | Sigma-Aldrich | H9394 | With Phenol Red for pH monitoring |
| HERAcell 150 | Thermo Scientific | CO2 incubator HERAcell ® 150 with decontamination routine | |
| HERAsafe KS/KSP | Thermo Scientific | Safety Cabinet | |
| Horse serum | GIBCO/Life | 26050088 | Heat inactivated |
| Huygens Professional | SVI Imaging | Deconvolution software | |
| Image J 1.52i | Wayne Rasban national Institute of Health | Image processing Software available in the public domain | |
| Insulin | Sigma-Aldrich | I6634 | Insulin from bovine pancreas |
| IP serie peristaltic pump | Ismatec | High-precisionmulti-channel pump | |
| Layout software, Illustrator CS6 | Adobe | Vector graphics editor | |
| L-glutamine | GIBCO/Life | 25030024 | |
| Microm HM 650 V | Thermo Scientific | Vibration microtome. Thermo scientific discontinued the production of the device in the meantime. Any other slicer or tissue chopper siutable for slicing living tissue is fine, too. | |
| Microscope stage | custom build | ||
| Microsoft Excel 16 | Microsoft | Spreadsheet software for basic data processing | |
| Millicell culture insert | Merck Millipore | PICM0RG50 | Hydrophilized PTFE, pore size 0.4 μm |
| Minimum Essential Medium Eagle | Sigma-Aldrich | M7278 | Synthetic cell culture media |
| Monochromator Polychrome V | Thermo Scientific/FEI | Ultra fast switching monochromator | |
| NaN3 (Sodium Azide) | Sigma-Aldrich | S-8032 | Mitochondrial inhibitor (complex IV inhibitor). CAUTION: Azide is toxic. Be aware not to accidentally ingest or inhale it, and prevent ist absoption through the skin. |
| Nikon Fluor 40x / 0.80 W DIC M ∞/0 WD 2.0 | Nikon Microscope Solutions | Water Immersion Microscope Objective | |
| NIS Elements 4.50 advanced Research | Nikon Microscope Solutions | Imaging software. Upgraded version for FRET imaging | |
| ORCA-Flash4.0 | Hamamatsu Photonics | Digital CMOS camera | |
| Perfusion tubing | Pro Liquid GmbH | Tygon tubing, 1.52 x 322 mm (Wd: 0.85) | |
| Photoshop CS 6 Version 13.0 | Adobe | Image processing software | |
| Sodium L-lactate | Sigma-Aldrich | 71718-10G | |
| ssAAV-2/2-hSyn1-ATeam1.03YEMK-WPRE-hGHp(A) | ETH Zürich | v244 | Single-stranded AAV vector that induces the expression of ATeam1.03YEMK under the control of the human synapsin 1 promoter fragment hSyn1. |
| ssAAV-5/2-hGFAP-hHBbI/E-ATeam1.03YEMK-WPRE-bGHp(A) | ETH Zürich | v307 | Single-stranded AAV vector that induces the expression of ATeam1.03YEMK under the control of the human glial fibrillary acidic protein promoter fragment ABC1D. |
| WVIEW GEMINI optic system | Hamamatsu Photonics | Emission Image Splitter |
Referanslar
- Sweadner, K. J. Isozymes of the Na+/K+-ATPase. Biochimica et Biophysica Acta. 988, 185-220 (1989).
- Clapham, D. E. Calcium signaling. Cell. 131, 1047-1058 (2007).
- Cotter, K., Stransky, L., McGuire, C., Forgac, M. Recent Insights into the Structure, Regulation, and Function of the V-ATPases. Trends in Biochemical Sciences. 40, 611-622 (2015).
- Harris, J. J., Jolivet, R., Attwell, D. Synaptic energy use and supply. Neuron. 75, 762-777 (2012).
- Brown, A. M., Ransom, B. R. Astrocyte glycogen and brain energy metabolism. Glia. 55, 1263-1271 (2007).
- Hertz, L., et al. Roles of astrocytic Na+,K+-ATPase and glycogenolysis for K+ homeostasis in mammalian brain. Journal of Neuroscience Research. 93, 1019-1030 (2015).
- Allaman, I., Belanger, M., Magistretti, P. J. Astrocyte-neuron metabolic relationships: For better and for worse. Trends in Neurosciences. 34, 76-87 (2011).
- Barros, L. F., Deitmer, J. W. Glucose and lactate supply to the synapse. Brain Research Reviews. 63, 149-159 (2010).
- Diaz-Garcia, C. M., et al. Neuronal Stimulation Triggers Neuronal Glycolysis and Not Lactate Uptake. Cell Metabolism. 26, 361-374 (2017).
- Diaz-Garcia, C. M., et al. Quantitative in vivo imaging of neuronal glucose concentrations with a genetically encoded fluorescence lifetime sensor. Journal of Neuroscience Research. 97, 946-960 (2019).
- Barros, L. F., et al. Current technical approaches to brain energy metabolism. Glia. 66, 1138-1159 (2018).
- Tantama, M., Hung, Y. P., Yellen, G. Optogenetic reporters: Fluorescent protein-based genetically encoded indicators of signaling and metabolism in the brain. Progress in Brain Research. 196, 235-263 (2012).
- Trevisiol, A., et al. Monitoring ATP dynamics in electrically active white matter tracts. eLife. 6, e24241(2017).
- Imamura, H., et al. Visualization of ATP levels inside single living cells with fluorescence resonance energy transfer-based genetically encoded indicators. Proceedings of the National Academy of Sciences of the United States of America. 106, 15651-15656 (2009).
- Lerchundi, R., et al. FRET-based imaging of intracellular ATP in organotypic brain slices. Journal of Neuroscience Research. , 1-13 (2018).
- Ehrengruber, M. U., et al. Gene transfer into neurons from hippocampal slices: comparison of recombinant Semliki Forest Virus, adenovirus, adeno-associated virus, lentivirus, and measles virus. Molecular and Cellular Neurosciences. 17, 855-871 (2001).
- Woo, J., et al. Functional Characterization of Resting and Adenovirus-Induced Reactive Astrocytes in Three-Dimensional Culture. Experimental Neurobiology. 26, 158-167 (2017).
- Close, B., et al. Recommendations for euthanasia of experimental animals: Part 2. DGXT of the European Commission. Laboratory Animals. 31, 1-32 (1997).
- Benediktsson, A. M., Schachtele, S. J., Green, S. H., Dailey, M. E. Ballistic labeling and dynamic imaging of astrocytes in organotypic hippocampal slice cultures. Journal of Neuroscience Methods. 141, 41-53 (2005).
- Lanjakornsiripan, D., et al. Layer-specific morphological and molecular differences in neocortical astrocytes and their dependence on neuronal layers. Nature Communications. 9, 1623(2018).
- Bushong, E. A., Martone, M. E., Ellisman, M. H. Examination of the relationship between astrocyte morphology and laminar boundaries in the molecular layer of adult dentate gyrus. The Journal of Comparative Neurology. 462, 241-251 (2003).
- Frotscher, M., Zafirov, S., Heimrich, B. Development of identified neuronal types and of specific synaptic connections in slice cultures of rat hippocampus. Progress in Neurobiology. 45, vii-xxviii (1995).
- Galimberti, I., et al. Long-term rearrangements of hippocampal mossy fiber terminal connectivity in the adult regulated by experience. Neuron. 50, 749-763 (2006).
- Stoppini, L., Buchs, P. A., Muller, D. A simple method for organotypic cultures of nervous tissue. Journal of Neuroscience Methods. 37, 173-182 (1991).
- Forster, E., Zhao, S., Frotscher, M. Laminating the hippocampus. Nature Reviews. Neuroscience. 7, 259-267 (2006).
- Holopainen, I. E. Organotypic Hippocampal Slice Cultures: A Model System to Study Basic Cellular and Molecular Mechanisms of Neuronal Cell Death, Neuroprotection, and Synaptic Plasticity. Neurochemical Research. 30, 1521-1528 (2005).
- Teschemacher, A. G., et al. Targeting specific neuronal populations using adeno- and lentiviral vectors: applications for imaging and studies of cell function. Experimental Physiology. 90, 61-69 (2005).
- Kantor, B., Bailey, R. M., Wimberly, K., Kalburgi, S. N., Gray, S. J. Methods for gene transfer to the central nervous system. Advances in Genetics. 87, 125-197 (2014).
- Mächler, P., et al. In Vivo Evidence for a Lactate Gradient from Astrocytes to Neurons. Cell Metabolism. 23, 94-102 (2016).
- Schreiner, A. E., Berlinger, E., Langer, J., Kafitz, K. W., Rose, C. R. Lesion-Induced Alterations in Astrocyte Glutamate Transporter Expression and Function in the Hippocampus. ISRN Neurology. 2013, 893605(2013).
- Haber, M., Zhou, L., Murai, K. K. Cooperative astrocyte and dendritic spine dynamics at hippocampal excitatory synapses. The Journal of Neuroscience: The Official Journal of the Society for Neuroscience. 26, 8881-8891 (2006).
- Neary, J. T., Kang, Y., Tran, M., Feld, J. Traumatic injury activates protein kinase B/Akt in cultured astrocytes: role of extracellular ATP and P2 purinergic receptors. Journal of Neurotrauma. 22, 491-500 (2005).
- Xia, J., et al. Neurons respond directly to mechanical deformation with pannexin-mediated ATP release and autostimulation of P2X7 receptors. The Journal of Physiology. 590, 2285-2304 (2012).
- Ainscow, E. K., Mirshamsi, S., Tang, T., Ashford, M. L., Rutter, G. A. Dynamic imaging of free cytosolic ATP concentration during fuel sensing by rat hypothalamic neurones: evidence for ATP-independent control of ATP-sensitive K+ channels. The Journal of Physiology. 544, 429-445 (2002).
- Arcuino, G., et al. Intercellular calcium signaling mediated by point-source burst release of ATP. Proceedings of the National Academy of Sciences of the United States of America. 99, 9840-9845 (2002).
- Rajendran, M., Dane, E., Conley, J., Tantama, M. Imaging Adenosine Triphosphate (ATP). The Biological Bulletin. 231, 73-84 (2016).
- Rangaraju, V., Calloway, N., Ryan, T. A. Activity-driven local ATP synthesis is required for synaptic function. Cell. 156, 825-835 (2014).
- Chatton, J. Y., Pellerin, L., Magistretti, P. J. GABA uptake into astrocytes is not associated with significant metabolic cost: implications for brain imaging of inhibitory transmission. Proceedings of the National Academy of Sciences of the United States of America. 100, 12456-12461 (2003).
- Magistretti, P. J., Chatton, J. Y. Relationship between L-glutamate-regulated intracellular Na+ dynamics and ATP hydrolysis in astrocytes. Journal of Neural Transmission (Vienna). 112, 77-85 (2005).
- Langer, J., et al. Rapid sodium signaling couples glutamate uptake to breakdown of ATP in perivascular astrocyte endfeet. Glia. 65, 293-308 (2017).
- Yaginuma, H., et al. Diversity in ATP concentrations in a single bacterial cell population revealed by quantitative single-cell imaging. Scientific Reports. 4, 6522(2014).
- Tantama, M., Martinez-Francois, J. R., Mongeon, R., Yellen, G. Imaging energy status in live cells with a fluorescent biosensor of the intracellular ATP-to-ADP ratio. Nature Communications. 4, 2550(2013).
- Nakano, M., Imamura, H., Nagai, T., Noji, H. Ca2+ Regulation of Mitochondrial ATP Synthesis Visualized at the Single Cell Level. ACS Chemical Biology. 6, 709-715 (2011).
- Mollajew, R., Toloe, J., Mironov, S. L. Single KATP channel opening in response to stimulation of AMPA/kainate receptors is mediated by Na+ accumulation and submembrane ATP and ADP changes. The Journal of Physiology. 591, 2593-2609 (2013).
- Pathak, D., et al. The Role of Mitochondrially Derived ATP in Synaptic Vesicle Recycling. The Journal of Biological Chemistry. 290, 22325-22336 (2015).
- Karus, C., Mondragao, M. A., Ziemens, D., Rose, C. R. Astrocytes restrict discharge duration and neuronal sodium loads during recurrent network activity. Glia. 63, 936-957 (2015).
- Larsen, B. R., Stoica, A., MacAulay, N. Managing Brain Extracellular K+ during Neuronal Activity: The Physiological Role of the Na+/K+-ATPase Subunit Isoforms. Frontiers in Physiology. 7, 141(2016).
- Ruminot, I., et al. NBCe1 mediates the acute stimulation of astrocytic glycolysis by extracellular K+. The Journal of Neuroscience : The Official Journal of the Society for Neuroscience. 31, 14264-14271 (2011).
Yeniden Basımlar ve İzinler
Bu JoVE makalesinin metnini veya resimlerini yeniden kullanma izni talebi
Izin talebiDaha Fazla Makale Keşfet
This article has been published
Video Coming Soon
JoVE Hakkında
Telif Hakkı © 2020 MyJove Corporation. Tüm hakları saklıdır