
Method Article
Imagem de ATP intracelular em fatias organotípicas do tecido do cérebro do rato usando o sensor ateam1.03yEMK baseado em FRET
Neste Artigo
Resumo
Descrevemos um protocolo para a expressão específica do tipo celular do sensor geneticamente codificado baseado em FRET ATeam1.03YEMK em culturas organotípicas do antecérebro do rato. Além disso, mostramos como usar este sensor para imagens dinâmicas dos níveis de ATP celular em neurônios e astrócitos.
Resumo
A atividade neuronal no sistema nervoso central (SNC) evoca uma alta demanda de energia celular fornecida pela quebra do triposfato de adenosina (ATP). Uma grande parte do ATP é necessária para reinstalar gradientes de íons através de membranas plasmáticas degradadas pela sinalização elétrica dos neurônios. Há evidências de que os astrócitos - embora não gerem sinais elétricos rápidos - passam pelo aumento da produção de ATP em resposta à atividade neuronal e suportam neurônios ativos, fornecendo metabólitos de energia para eles. O recente desenvolvimento de sensores geneticamente codificados para diferentes metabólitos agora permite o estudo de tais interações metabólicas entre neurônios e astrócitos. Aqui, descrevemos um protocolo para a expressão específica do tipo celular do sensor De setores de ressonância de ressonância (FRET-) aTeam1.03YEMK em culturas organotípicas de fatia de tecido do hipocampo e córtex do mouse usando vetores virais associados ao adeno (AAV). Além disso, demonstramos como este sensor pode ser empregado para a medição dinâmica das mudanças nos níveis de ATP celular em neurônios e astrócitos após aumentos no potássio extracelular e após a indução da isquemia química (ou seja, uma inibição do metabolismo energético celular).
Introdução
A atividade elétrica excitatória dos neurônios é em grande parte baseada no fluxo de cições como sódio (Na+)e potássio (K+)através de suas membranas plasmáticas. A manutenção dos gradientes eletroquímicos destes dois íons é exigida assim para sinalizar. Isso é realizado pela celular Na+/K+-ATPase (NKA), uma bomba de transmembrana eletrogênica onipresentemente expressa, que extrude 3 Na+ fora da célula em troca de 2 K+ do espaço extracelular, exigindo o consumo de uma molécula de ATP por ciclo de transporte1. Além do NKA, existem vários outros transportadores de íons consumidores de ATP, incluindo a membrana plasmática Ca2+-ATPase, que é vital para a homeostase intracelular Ca2+ e sua exportação após o influxo induzido pela atividade2. Nas vesículas pré-sinápticas, um tipo vacuolar H+-ATPase (v-ATPase) cria o gradiente de prótons necessário para a captação de neurotransmissores neste compartimento3.
Enquanto a atividade dos neurônios, portanto, requer uma quantidade substancial de ATP4, eles não exibem uma capacidade significativa para o armazenamento de energia. Em vez disso, eles parecem confiar em interações metabólicas com os astrocitos vizinhos, as principais lojas de glicogênio no cérebro5. Tem sido sugerido que o glicogênio ascítico realmente desempenha um papel importante no apoio às necessidades de energia neuronal; e um fenômeno chave neste acoplamento neuro-metabólico proposto entre os dois tipos de células é a capacidade dos astrócitos para aumentar sua produção de ATP em resposta à atividade neuronal6,7,8. Esta hipótese, conhecida como ônibus de lactato do neurônio astrócito (ANLS), ainda está em debate, porque outros trabalhos forneceram evidências de que os neurônios também podem aumentar sua própria taxa de glicolíse em resposta à estimulação9,10,refletindo a necessidade de outros métodos e abordagens para estudar a interação neuro-glia.
Investigação do metabolismo da energia celular e níveis de ATP em neurônios e astrócitos para elucidar interações metabólicas neuro-glia tem sido dificultada pela falta de sondas adequadas para a detecção de mudanças nas concentrações de metabólitos em células vivas no tecido cerebral. A última década, no entanto, proporcionou um aumento no desenvolvimento de novas ferramentas e novas sondas fluorescentes geneticamente codificadas para diferentes metabólitos, incluindo sensores para ATP, lactato, piruvato e outros11,12. Usando essas ferramentas, agora é possível abordar diretamente questões relacionadas ao consumo de ATP celular e mudanças nos níveis de energia celular no nível de célula única e de forma específica do tipo celular no tecido cerebral intacto13.
No presente trabalho, descrevemos um procedimento para visualizar a dinâmica citossólica atp em neurônios e astrócitos de fatias de cérebro organotypic culturado. Mostramos como empregar vetores virais associados ao adeno (AAV) para expressão específica do tipo célula do atp-nanosensor geneticamente codificado ATeam1.03YEMK (14) em neurônios e astrócitos de fatias de cérebro de rato que podem ser mantidas na cultura celular por várias semanas15. Um procedimento de como remover a cicatriz glial que cobre fatias cultivadas do tecido é descrito, que melhora a acessibilidade ótica e a imagem latente das pilhas nas camadas organotypic do tecido embaixo. Finalmente, mostramos como o ATeam1.03YEMK pode ser usado para realizar imagens baseadas em FRET de alterações nos níveis de ATP celular nesta preparação. Este método hospeda as principais vantagens de que não requer procedimentos cerebrais cirúrgicos, fornece altos níveis de expressão do sensor e especificidade do tipo celular em fatias cerebrais cultivadas, reduzindo a invasividade ou estresse nas células em comparação com outros métodos, como a transfecção por eletroporação ou transdução com outros vetores virais10,16,17. Além disso, este protocolo pode ser aplicado a outros nanosensores baseados em FRET, entre eles variantes do ATeam1.03 que fornecem menor afinidade de ligação para ATP14.
Protocolo
O presente estudo foi realizado em estrita conformidade com as orientações institucionais da Universidade Heinrich Heine de Düsseldorf, bem como com a Diretiva do Conselho Comunitário Europeu (2010/63/UE). Todos os experimentos com culturas organotípicas de fatias cerebrais foram comunicados e aprovados pelo Escritório de Bem-Estar Animal no Animal Care and Use Facility da Universidade Heinrich Heine Düsseldorf (número do ato institucional: O50/05). De acordo com as recomendações da Comissão Europeia18,animais com até 10 dias de idade foram mortos por decapitação.
1. Preparação de culturas organotípicas da fatia do cérebro (OTCs)
- No dia anterior ou pelo menos 30 min antes do procedimento
- Prepare a placa de Petri (em condições estéreis). Retire a tampa da placa de 6 poços e coloque 800-850 μL de meio OTC em cada poço. Mantenha a placa na incubadora (37 °C, 5% CO2/95% O2) até que seja necessário.
- Prepare a lavagem de placas de Petri (em condições estéreis). Adicione 3 mL de HBSS a cada 30 mm de placa de Petri. Um total de 5 pratos por procedimento é necessário. Coloque-os na incubadora (37 °C, 5% CO2/95% O2) por pelo menos 30 min - durante a noite até o necessário.
- Prepare o ACSF(Tabela 1). Mantenha o soline sem glicose a 4 °C até o dia seguinte.
| Salines e Mídia - Formulação | ||||
| Nome | Abreviação | Composição | Concentração [mM] | Comentários |
| Solução artificial de líquido cefalorraquidiano | ACSF ACSF | Nacl | 125 | Borbulhado com 5% CO2/95% O2, pH 7,4 |
| Kcl | 2.5 | Adicione sempre a glicose direita antes do uso. | ||
| CaCl 2 CaCl2 | 2 | Não guarde por mais de um dia com glicose | ||
| MgCl 2 MgCl2 | 1 | ~310 mOsm/L ~310 mOsm/L | ||
| Nah 2 PO 4 Nah2PO4 | 1.25 | |||
| NaHCO 3 NahCO3 | 26 | |||
| Glicose | 20 | |||
| Experimental ACSF | E-ACSF E-ACSF | Nacl | 136 | Borbulhado com 5% CO2/95% O2, pH 7,4 |
| Kcl | 3 | Adicione sempre a glicose direita antes do uso. | ||
| CaCl 2 CaCl2 | 2 | Não guarde por mais de um dia com glicose | ||
| MgCl 2 MgCl2 | 1 | ~320 mOsm/L ~320 mOsm/L | ||
| Nah 2 PO 4 Nah2PO4 | 1.25 | |||
| NaHCO 3 NahCO3 | 24 | |||
| Glicose | 5 | |||
| Lactato | 1 | |||
| Solução química da isquemia | Cis | Nacl | 136 | Borbulhado com 5% CO2/95% O2, pH 7,4 |
| Kcl | 3 | ~318 mOsm/L ~318 mOsm/L | ||
| CaCl 2 CaCl2 | 2 | |||
| MgCl 2 MgCl2 | 1 | |||
| Nah 2 PO 4 Nah2PO4 | 1.25 | |||
| NaHCO 3 NahCO3 | 24 | |||
| 2, 2-Deoxiglicose | 2 | |||
| Nan 3 Nan3 | 5 | |||
| 8 mM acsf potássio | 8 mM K+ ACSF | Nacl | 128 | Borbulhado com 5% CO2/95% O2, pH 7,4 |
| Kcl | 8 | ~320 mOsm/L ~320 mOsm/L | ||
| CaCl 2 CaCl2 | 2 | |||
| MgCl 2 MgCl2 | 1 | |||
| Nah 2 PO 4 Nah2PO4 | 1.25 | |||
| NaHCO 3 NahCO3 | 24 | |||
| Glicose | 5 | |||
| Lactato | 1 | |||
| ACSF com tampão de hepes | H-ACSF H-ACSF | Nacl | 125 | Ajustado ao pH 7.4 com NaOH |
| Kcl | 3 | Sempre adicione glicose logo antes de usar | ||
| CaCl 2 CaCl2 | 2 | Não guarde por mais de um dia com glicose | ||
| MgSO 4 MgSO4 | 2 | ~310 mOsm/L (ajustado com sacarose) | ||
| Nah 2 PO 4 Nah2PO4 | 125 | |||
| Hepes Hepes | 25 | |||
| Glicose | 10 | |||
| Solução de sal equilibrado de Hanks | HBSS HBSS | Sigma (catálogo número H9394). | ||
| Sinológico tampão de fosfato de Dulbecco | DPBS DPBS | Gibco (catálogo número 14287-080) | ||
| Meio de Cultura Organotípica | OTC médio | soro de cavalo inativado de calor | 20% | 34°C, 5% CO2,pH 7,4, condições de cultivo |
| Mem | 79% | ~320 mOsm/L ~320 mOsm/L | ||
| L-glutamina | 1 | |||
| Insulina | 0,01 mg/mL | |||
| Nacl | 14.5 | |||
| MgSO 4 MgSO4 | 2 | |||
| CaCl 2 CaCl2 | 1.44 | |||
| Ácido ascórbico | 0.00125 % | |||
| D-glicose | 13 |
Tabela 1: Composição da solução.
- No dia da preparação, adicione a glicose ao ACSF, coloque-a no gelo e comece a borbulhar com 95% O2/5% CO2 por pelo menos 30 min para resultar em um pH de 7,4.
- Dissecação e corte
- Sacrifique o mouse (BalbC, ambos os sexos) nos dias pós-natal 6 a 8 por decapitação rápida e coloque a cabeça em uma placa de Petri de vidro contendo ACSF gelada.
- Exponha o crânio cortando a pele da parte de trás até a ponta posterior do osso nasal. Em seguida, corte cuidadosamente o crânio do foramen magnum usando uma tesoura cirúrgica e expor o cérebro.
Nota: Verifique se o procedimento está de acordo com as diretrizes da instituição. - Retire o cérebro e coloque-o em uma membrana de filtro em uma placa de Petri gelada cheia de ACSF.
- Separe os hemisférios e realize um corte parasagitivo em um ângulo de 45°. Corrija um hemisfério no estágio de tecido vibratome com supercola. Transfira imediatamente o bloco de tecido para o banho vibratome contendo ACSF gelado (borbulhado com 5% co2/95% O2). Finalmente alinhar o tecido. Mantenha o segundo hemisfério em ACSF gelado até cortar.
- Ajuste o vibratome para cortar fatias em 250-400 μm. Cortar a 250 μm renderá aproximadamente 12 fatias por animal (400 μm: ~7 fatias).
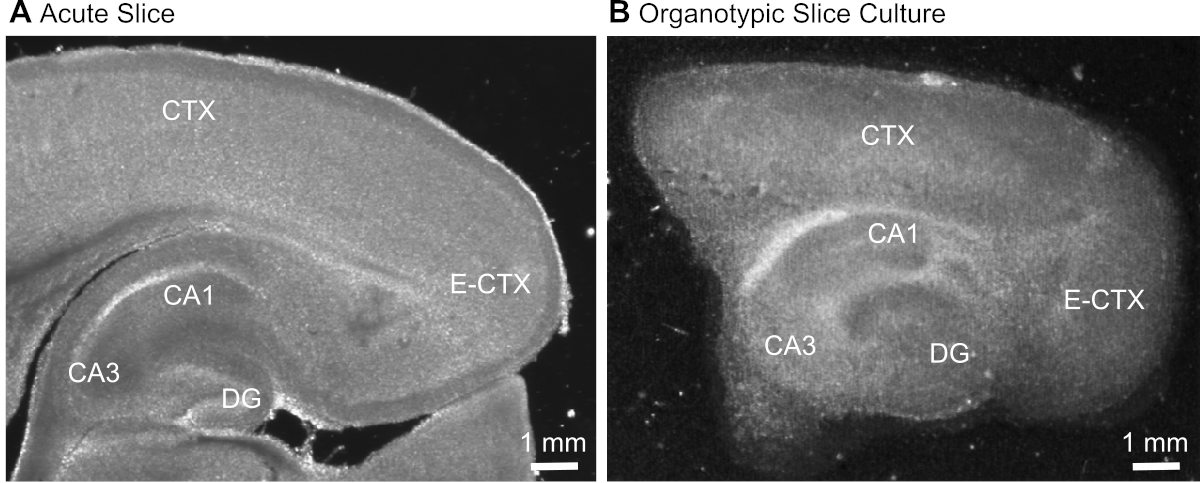
- Depois de cortar a fatia (Figura 1A),identifique a formação hipocampal com base em sua aparência morfológica típica (Figura 1)e isolá-la usando agulhas hipodérmicas (calibre 23, 1), mantendo a parte do córtex cerebral adjacente ao hipocampo.
Nota: Preservar o neocórtex enquanto se aventura ajuda a preservar a integridade do hipocampo. No entanto, o hipocampo pode ser isolado e cultivado sem córtex, se necessário. - Coloque a fatia em uma malha em aquecido, ACSF (34 °C, borbulhado com 5% CO2/95% O2) até que todas as fatias são coletadas.
- Transfira as fatias para o armário de fluxo laminar para continuar condições estéreis.

Figura 1: Imagens representativas da transmissão de preparações agudas e organotípicas da fatia do cérebro. Comparação de uma fatia do cérebro parasagital agudamente isolada (A)e uma fatia do cérebro organotypic do parasagittal mantida na cultura por 12 dias(B)usando a microscopia wide-field da translumination. DG = giro dentado; CA1/3 = CA1/CA3-região do hipocampo; E-CTX = Córtex entorrinal; CTX = (neo-) córtex. Clique aqui para ver uma versão maior deste número.
{kind=link}
2. Culturing as fatias
- Transfira delicadamente as fatias do ACSF em uma das placas de Petri pré-aquecidas enchidas com a solução de sal de Hank estéril usando uma pipeta estéril invertida do pasteur do vidro.
Nota: A esterilização do tecido é alcançada pela diluição (passo 3.2). Transfira o menor ACSF possível para a placa de Petri. - Mude a pipeta e transfira as fatias para a segunda placa de Petri. Repita o processo 5 vezes no geral. Transfira o mínimo possível de HBSS para as seguintes placas de Petri.
- Coloque delicadamente uma fatia de cada vez no topo da inserção da cultura. Repita o processo para cada fatia. Evite turbulências na pipeta e espere até que a fatia desça até a ponta da pipeta Pasteur. Pode-se colocar até 4 fatias em uma única membrana.
- Retire cuidadosamente qualquer solução de Hank em excesso da parte superior da inserção usando uma ponta fina.
- Mantenha as culturas(Figura 2)em uma incubadora na interface entre gás (carbogênio, 95% O2 /5% CO2)e o líquido a 37 °C até o dia do experimento. Substitua o meio a cada 2-3 dias.

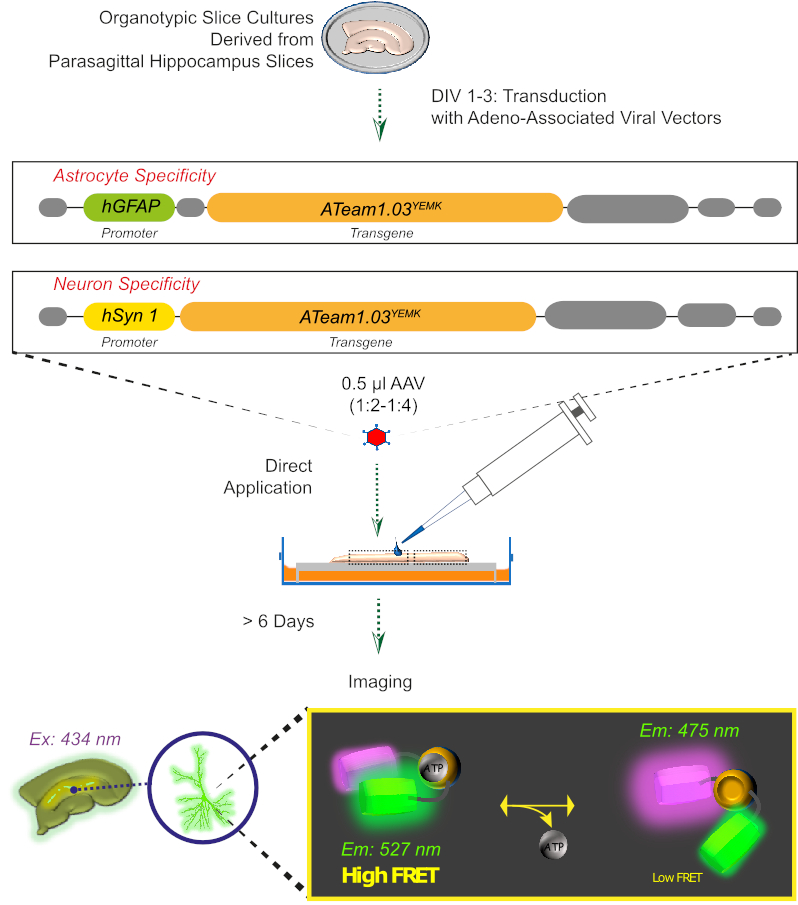
Figura 2: Princípio da imagem ATP baseada em FRET em fatias de cérebro organotipato cultivadas usando o sensor geneticamente codificado ATeam1.03YEMK. Representação esquemática do protocolo apresentado neste trabalho. Brevemente, fatias organotípicas parasagitais, cultivadas por 1-3 dias, são transinduzidas com um vetor viral associado ao adeno contendo qualquer um, o hGFAP específico do astrocito ou o hSyn-promotor específico do neurônio e a seqüência para a expressão de ATeam1.03YEMK. Alíquotas diluídas desses vetores (1:2-1:4) são aplicadas diretamente no topo de uma fatia, que é mantida em condições de cultivo por pelo menos mais 6 dias. As alterações nos níveis intracelulares de ATP podem então ser visualizadas nas células que expressam o sensor empolgá-lo a 434 nm e adquirindo emissão de fluorescência simultaneamente em 527 (aceitador) e 475 (doador) nm. Clique aqui para ver uma versão maior deste número.
{kind=link}
3. Expressão de sensores ATP com um Vetor Viral associado a Aadeno(Figura 2)
Nota: Certifique-se de atender a todos os requisitos para o manuseio de organismos geneticamente modificados!
- Para lidar com o vetor viral, aliquot adeno associados vetores virais (AAV2/5) em 1-2 μL para evitar congelamento repetido e descongelamento. Guarde os alíquos a -80 °C.
- Coloque um frasco contendo 10% de lixívia no banco estéril para descartar todo o material residual usado que estava em contato com o vetor.
- Para a transdução, prepare uma diluição de 1 μL do vetor com 2-3 μL de DPBS. As soluções de ações normalmente exibem um díter físico na magnitude de 1012 genomas virais por mL (vg/mL).
- Transfira uma inserção contendo uma fatia cultivada no capô estéril.
- Sem tocar no tecido, aplique 0,5 μL do vetor diluído diretamente na parte superior de cada fatia.
Nota: Melhor expressão nas camadas mais profundas do tecido é obtida transduzindo fatias cultivadas em 1-3 dias in vitro (DIV). A transdução de culturas mais antigas pode resultar em uma expressão predominante de células na cicatriz glial circundante ou uma baixa expressão nos neurônios, respectivamente. - Finalmente, coloque as fatias de volta para a incubadora e mantê-los por pelo menos mais 6 dias. Não altere o meio no dia da transdução.
4. Remoção da Cicatriz Glial (Figura 3)
- Pouco antes de iniciar um experimento, transfira uma inserção contendo fatias cultivadas no capô estéril e coloque-a em um prato de 30 mm, contendo 1 mL de meio OCT ou MEM.
- Coloque o prato o estereoscópio e concentre-se na superfície da fatia.
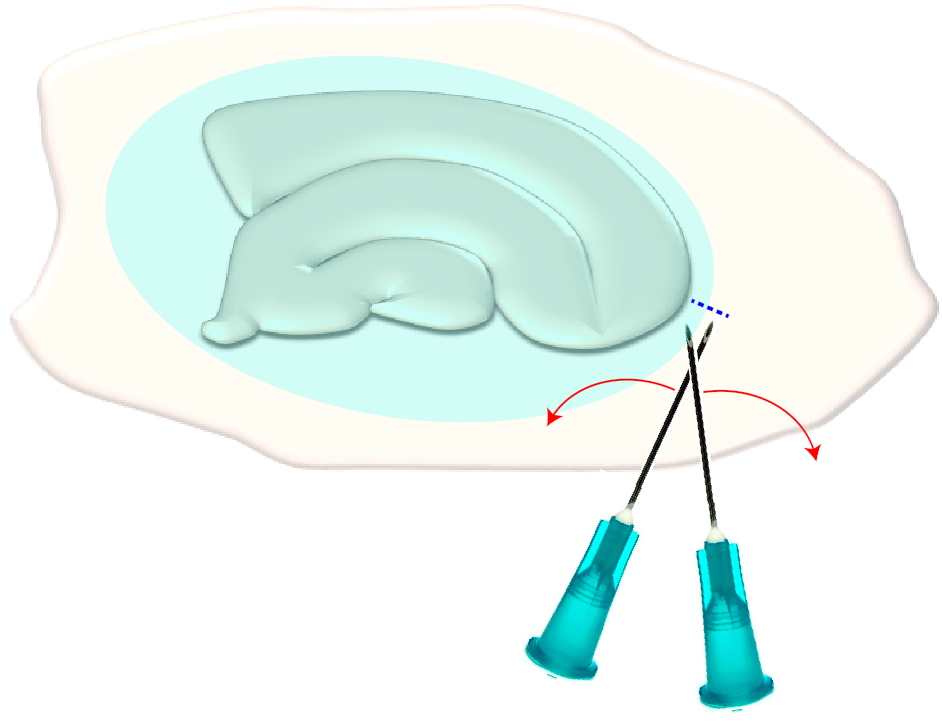
- Use duas agulhas hipodérmicas estéreis (23 G, 1") para fazer um curto corte cruzado bem nas bordas estreitas de uma fatia escolhida (Figura 3). Este procedimento liberará a tensão na superfície criada pela cicatriz glial que faz com que retraia, expondo desse modo as camadas subjacentes (veja a figura 5).
Nota: A primeira camada de tecido (cicatriz glial) é formada principalmente por astrócitos reativos. Em imagens de campo largo, esta camada de tecido densa resultará em dispersão adicional da luz, resultando em imagens borradas. Remover a cicatriz é, portanto, vantajoso para obter uma melhor visibilidade das camadas mais profundas, que contêm o tecido organotypic adequado. Assim, tenha o cuidado de realizar este corte exclusivamente na borda e na camada superior da preparação da fatia apenas e não danificar o tecido embaixo. Não observamos diferenças entre os dados obtidos em OTCs com cicatriz glial com os de OTCs sem cicatrizes (dados não mostrados). - Retire a fatia preparada da inserção. Para este fim, use um bisturi estéril e extirpar-lo, fazendo cortes paralelos em linha reta para a membrana, formando um quadrado ou um triângulo com a fatia no centro, mantendo as bordas da membrana com pinças. Se a inserção hospeda fatias adicionais, transferi-lo de volta para a placa original e para a incubadora. A tensão superficial do meio impedirá seu vazamento na superfície da membrana.

Figura 3: Ilustração esquemática da remoção mecânica da cicatriz glial. A figura mostra uma cultura de fatia hipocampal que é coberta por uma cicatriz glial (ellipsoid azulado). Por um tempo tosquiando as pontas de duas agulhas de seringa no menor pólo da cultura e na borda da cicatriz glial (linha azul tracejada), a cicatriz vai virar de lado. Clique aqui para ver uma versão maior deste número.
{kind=link}
5. FRET-based ATP Imaging (Figura 4)
- Antes do experimento, prepare o E-ACSF e borbulhacom 95% O2/5% CO2 por pelo menos 30 min para obter um pH de 7,4. Ligue a fonte de luz fluorescente (lâmpada Xenon) do monochromator (Figura 4). Comece a perfusão pouco antes de tirar a fatia da incubadora.
Nota: Mantenha o soline borbulhado com 95% O2 e 5% CO2 durante todo o experimento. - Transfira a fatia para uma câmara experimental que é constantemente perfundida com E-ACSF recém-carbogenado usando uma bomba peristaltic (Figura 4). Em seguida, conserte a fatia com uma grade. Coloque a câmara no estágio do microscópio e conecte o sistema de perfusão. A tubulação de laboratório à prova de gás é recomendada para perfusão.
Nota: Experimentos podem ser realizados à temperatura ambiente ou perto da temperatura fisiológica, dependendo do projeto experimental. Verifique a estabilidade e confiabilidade do fluxo de perfusão para evitar mudanças de foco induzidas pelo movimento do tecido e/ou alterações no estresse de cisalhamento. As velocidades padrão da perfusão para o trabalho da fatia, usada por nós e por muitos outros laboratórios, são 1.5-2.5 mL/min.

Figura 4: Configuração da configuração de imagem fret. (A)Ilustração esquemática dos diferentes componentes e seu arranjo espacial necessário para a configuração de imagem FRET. O arranjo consiste em um monochromator com uma lâmpada de xenônio como fonte de luz, um microscópio de estágio fixo vertical (1), um sistema de divisor de imagem (2), uma câmera CCD ou CMOS digital para gravação de lapso de tempo (3) e um banho experimental adaptado para perfusão constante estável (4). A perfusão de banho é realizada por uma bomba peristaltic com taxa de fluxo ajustável. Imagemdo espaço de trabalho experimental. A configuração de imagem FRET é montada em uma mesa com vibração úmida carregando um estágio x/y-translational, no qual o banho experimental é incorporado. Números: ver (A). C) Visão esquemática do caminho de luz do monocromrador para a câmera digital. Indicado é a posição dos diferentes filtros e do espelho dicroico. Números: ver (A). Clique aqui para ver uma versão maior deste número.
{kind=link}
- Traga a fatia cultivada no foco usando a luz da transmissão. Identifique a área onde os experimentos serão realizados (exemplo: região CA1 do hipocampo). Antes de iniciar experimentos de imagem, espere pelo menos 15 min para permitir que as fatias se adaptem às condições shosinas. Para a configuração da configuração experimental, consulte a Figura 4.
- Ligue a câmera e o software de imagem. Em seguida, selecione o cubo de filtro adequado.
- Excite a proteína fluorescente doadora (eCFP) em 435/17 nm (~435 nm). Defina o tempo de exposição entre 40 e 90 ms.
Nota: Forte exposição de fatias à luz fluorescente pode resultar em efeitos fototóxicos. - A excitação em 435 nm resulta em emissão em 475 nm (eCFP; doador) e 527 nm (Vênus; aceitador). Divida a emissão de fluorescência a 500 nm com um divisor de imagem de emissão e empregue filtros de passe de banda em 483/32 e 542/27 para isolar ainda mais a fluorescência de doadores e aceitadores. A expressão forte pode resultar na saturação dos detectores. Neste caso, você pode usar um filtro de densidade neutra para reduzir a intensidade da excitação.
- Selecione uma região de interesse (ROI) aparentemente desprovida de fluorescência celular para subtração em segundo plano. Em seguida, crie ROIs delineando corpos celulares.
- Defina a frequência de aquisição de imagem e o tempo de gravação geral. Para experimentos longos (>30 min), uma frequência de aquisição de 0,2-0,5 Hz é recomendada para prevenir a fototoxicidade.
- Comece a gravação. Recomenda-se registrar pelo menos 5 minutos em condições de base para garantir a estabilidade da preparação.
Nota: Ajuste o foco da célula durante a gravação, se necessário. - Para induzir alterações no ATP intracelular, transfira o tubo de perfusão do ACSF padrão para um soro fisiológico contendo inibidores metabólicos (por exemplo, CIS, consulte a Tabela 1 e abaixo). Alternativamente, use um salino com concentração elevada de potássio para imitar a liberação de potássio de neurônios ativos.
Nota: A aplicação por perfusão de banho é um processo relativamente lento, que globalmente atua em toda a preparação. Tome nota do momento em que a nova solução realmente começou a entrar no banho experimental. Dependendo da distância entre a câmara e o reservatório do salino, bem como da velocidade da perfusão, um tempo de atraso deve ser considerado.
6. Documentação de alta resolução de fluorescência celular ateam
- Logo após as gravações, transfira a câmara de gravação contendo a cultura de fatia para o microscópio de varredura a laser confocal.
Nota: Tome cuidado especial. Por causa de potenciais danos fotográficos, executar este passo somente após experimentos. Para fins de documentação, pode-se trocar o E-ACSF com h-ACSF. Portanto, um sistema de perfusão não é necessariamente necessário. - Tome z-pilhas na resolução z mais alta possível na configuração óptica dada.
- Aplique um algoritmo de desconvolução para aumentar a resolução de imagem.
Resultados
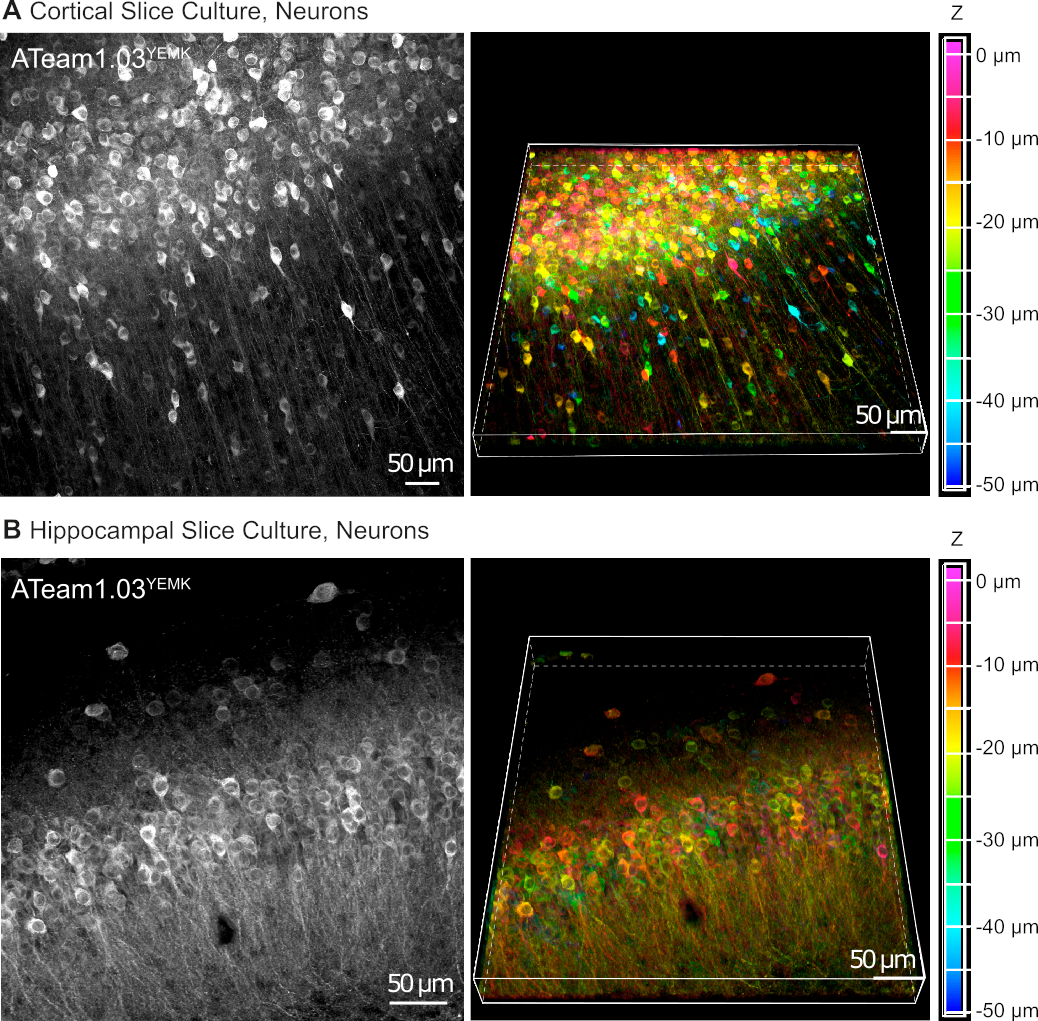
Os vetores de AAV são uma ferramenta confiável para expressar seletivamente genes estranhos em células dentro do tecido vivo16. A aplicação direta de AAVs contendo a fita de seqüência de ATeam1.03YEMK e um promotor específico resulta em uma alta expressão do sensor no tipo de célula escolhida. Na DIV 14 (~10 dias após uma transdução), os neurônios que expressam ATeam o promotor de sinapse humana são encontrados em alta densidade no neocórtex de fatias de tecido cultivado em profundidades de até 50 μm abaixo da superfície da fatia (Figura 5A). Resultados comparáveis podem ser alcançados no hipocampo(Figura 5B).

Figura 5: Visualização de neurônios expressando ATeam1.03YEMK em fatias de cérebro organotipatoorganolípicas parasagitais cultivadas. As imagens à esquerda correspondem a projeções de foco estendida de 43 seções ópticas (1,05 μm cada) de tecido cortical(A)e (B)de 70 seções ópticas (0,6 μm cada) de tecido hipocampal. As imagens à direita representam a visão de volume da mesma projeção. As células são codificadas por cores de acordo com sua profundidade em relação à superfície da fatia, como indicado pela escala de cor à direita. Clique aqui para ver uma versão maior deste número.
{kind=link}
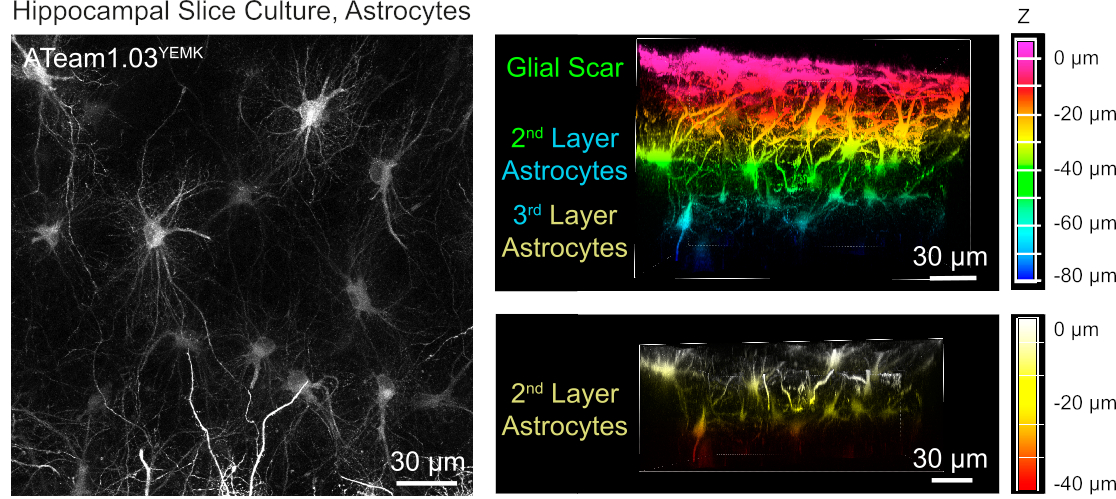
Para a medição dos níveis de ATP em astrócitos, O ATeam1.03YEMK é expresso o controle do promotor de proteína cibrilar ácida glial humana (GFAP). Isso resulta em transdução eficiente de células tanto no neocórtex quanto no hipocampo de fatias de tecido cultivado (Figura 6). Notavelmente, dois fenótipos morfológicos diferentes podem ser distinguidos, dependendo da profundidade em relação à superfície dos preparativos de fatias. Na primeira camada superficial, as células são caracterizadas por processos primários espessos que são predominantemente dispostos em paralelo à superfície. Essas células exibem domínios fortemente sobrepostos, criando uma densa malha de astrócitos aparentemente reativos (Figura 6). Em camadas mais profundas (30-60 μm da superfície), os astrócitos transinduzidos exibem processos celulares finos que formam domínios esféricos em grande parte e sua morfologia se assemelha à de astrócitos in situ, conforme relatado anteriormente19,20,21 ( Figura6). Para obter uma melhor transdução de astrocitos de camada mais profunda, bem como melhor acesso óptico a estas camadas mais profundas, o tecido cicatricial glial pode ser removido como descrito no Passo 4.

Figura 6: Visualização de astrócitos expressando ATeam1.03YEMK em fatias de cérebro organotipato parasagitais cultivadas. A imagem à esquerda corresponde a uma projeção de foco estendida de 191 seções ópticas (0,45 μm cada). Para fins ilustradores, a cicatriz glial foi excluída da projeção de astrócitos. As imagens à direita representam a visão de volume da mesma projeção antes e depois da remoção da cicatriz glial. As células são codificadas por cores de acordo com sua profundidade em relação à superfície da fatia, como indicado pelas escalas de cor à direita. Clique aqui para ver uma versão maior deste número.
{kind=link}
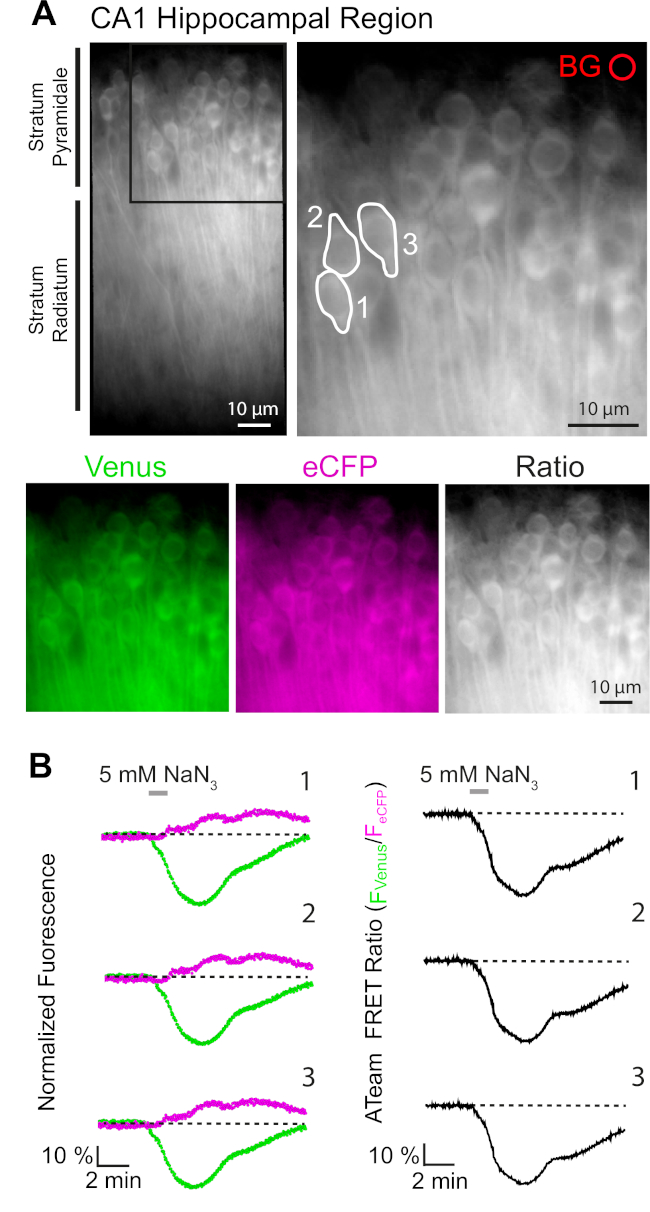
A expressão bem-sucedida do ATeam1.03YEMK permite a medição dinâmica das mudanças nos níveis de ATP em neurônios ou astrócitos, dependendo do promotor usado (veja acima). Experimentos foram realizados em um banho experimental constantemente perfundido com E-ACSF (borbulhado com 95% O2/5% CO2). Em fatias organotípicas expressando ATeam1.03YEMK em neurônios hipocampais, regiões de interesse (ROIs) foram selecionadas antes de iniciar a gravação, representando os somatas das células piramidais (Figura 7A). Além disso, foi escolhida uma região para subtração de fundo(Figura 7A). Emissão de Vênus, bem como fluorescência eCFP foi então coletado para cada um desses ROIs separadamente e descrito como nível de emissão de fluorescência ao longo do tempo (Figura 7B). Depois de registrar fluorescência condições de controle por vários minutos para garantir uma linha de base estável, o metabolismo celular foi inibido por expor a preparação da fatia a uma soro fisiola sem glicose, à qual 5 mM de sódio azide (NaN3)foi adicionado por um minuto (Figura 7B). Essa manipulação induziu mudanças opostas na intensidade de emissão do par de FRET(Figura 7B,painéis esquerdos), com diminuição de Vênus (527 nm) e aumento da emissão de eCFP (475 nm). Calcular a relação FRET dividindo a emissão de fluorescência de Vênus pela eCFP (FVenus/FeCFP)resultou em sinais que refletem as mudanças relativas nos níveis intracelulares de ATP, a chamada "relação ATeam FRET" (Figura 7B, painel direito). Em todos os neurônios registrados (n = 70 células em N = 5 fatias), NaN3 causou uma diminuição reversível na relação ATeam FRET, indicando uma diminuição reversível nos níveis intracelulares de ATP após a inibição do metabolismo celular.

Figura 7: Demonstração de tempo lapso ateam fret ratio imagem. (A)Canto superior esquerdo: Imagem de fluorescência de campo largo da camada piramidal e estrato radiatum da região CA1 de uma fatia hipocampal organotypic cultivada expressando ATeam1.03YEMK em neurônios. Canto superior direito: Visão ampliada da seção encaixotada, conforme indicado à esquerda. Linhas brancas delineam regiões de interesse (ROIs) 1-3 representando corpos celulares de neurônios piramidais CA1 escolhidos para análise em (B). Bg representa o ROI escolhido para correção de fundo. Inferior: Imagens pseudocoloridas que representam a emissão de fluorescência de Vênus (verde), eCFP (roxo) e a proporção de Vênus/eCFP. (B) Gravação de lapso de tempo em ROIs 1-3, representando corpos de células neuronais (ver A). Traços à esquerda mostram emissão normalizada de fluorescência de Vênus (verde) e eCFP (magenta). Os traços à direita mostram a proporção correspondente do ATeam FRET. Note-se que a perfusão com 5 mM NaN3 na ausência de glicose extracelular por 1 minuto (barra cinzenta) induz uma diminuição reversível na relação ATeam FRET, indicando uma diminuição na concentração intracelular de ATP. Clique aqui para ver uma versão maior deste número.
{kind=link}
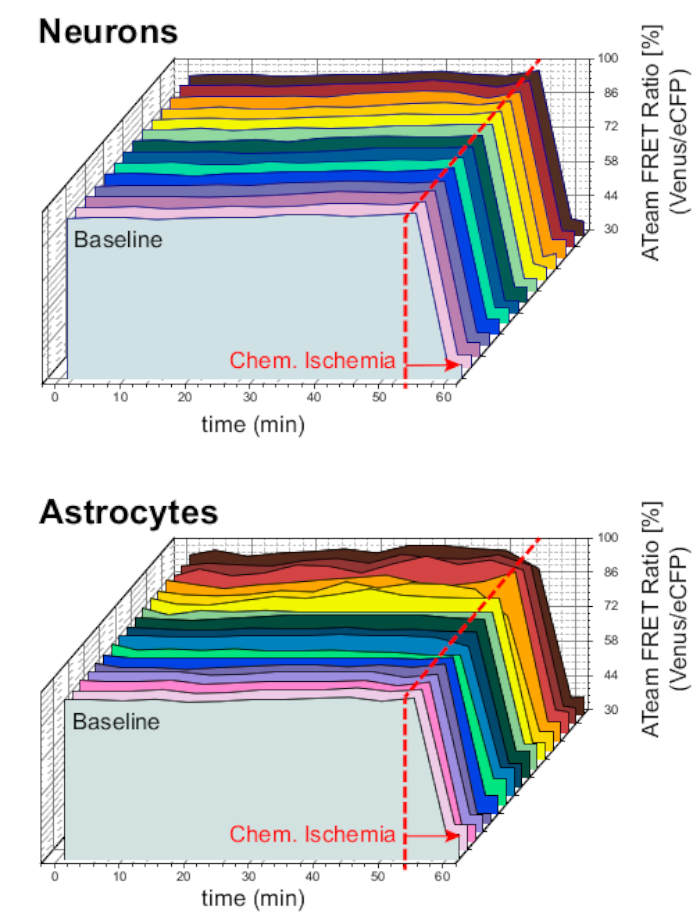
Para garantir a estabilidade da preparação e do sensor em condições de experimento sinuoso de longo prazo, as fatias expressando ATeam em neurônios ou astrócitos eram constantemente perfundidas com ACSF por períodos prolongados (>50 min; n = 12 células cada, N = 3 OTCs de 3 cérebros). Nessas condições, a relação ATeam FRET não mudou(Figura 8). Expor os preparativos à CEI contendo inibidores metabólicos ("Isquemia Química", ver Tabela 1), em contraste, novamente resultou na queda esperada na relação ATeam FRET, conforme observado acima.

Figura 8: Experimentos de base empregando ATeam. Imagens de longo prazo da relação ATeam FRET em 14 células diferentes em condições de base em neurônios (superior) e astrócitos (inferior). Os dados foram dados em condições comparáveis como outros dados experimentais. Ao final de cada medida, a isquemia química foi eliciada pela perfusão com CEI, conforme indicado pela seta. NOTA: Os índices de FRET da ATeam de base estão estáveis ao longo do tempo em condições de base. Clique aqui para ver uma versão maior deste número.
{kind=link}
Em seguida, analisamos as respostas de neurônios e astrócitos expressando ATeam1.03YEMK a um aumento na concentração de potássio extracelular. Depois de estabelecer uma linha de base estável, os neurônios foram perfundidos com um soro salino em que a concentração de potássio foi aumentada de 3 para 8 mM por 3 minutos(Figura 9A). Esta manipulação, no entanto, não resultou em uma mudança detectável na relação ATeam FRET (n = 56 células em N = 5 fatias). Para garantir que o sensor reagisse a uma mudança nos níveis de ATP, as fatias foram novamente expostas a um período sustentado de isquemia química provocada pela substituição do E-ACSF pela CIS. A isquemia química resultou em uma rápida diminuição da relação ATeam FRET para um nível novo e estável, indicando esgotamento nominal do ATP após 2-3 min (Figura 9A).

Figura 9: Experimentos representativos ilustrando mudanças nos níveis de ATP em neurônios e astrócitos. (A,B): Imagens à esquerda mostram fluorescência ATeam de neurônios e astrócitos localizados na região hipocampal CA1 de fatias organotípicas. Traços à direita representam gravações de lapso de tempo da relação ATeam FRET obtidas a partir de um ROI posicionado sobre um único corpo celular. Em ambos os experimentos, as fatias foram submetidas a um aumento na concentração de potássio extracelular por 3 minutos (ver barra), seguida de uma exposição final à isquemia química. Note-se que, enquanto os neurônios não respondem à elevação do potássio extracelular(A),ostrócitos reagem com um aumento no ATP (B). Clique aqui para ver uma versão maior deste número.
{kind=link}
O mesmo protocolo experimental foi realizado com fatias, nas quais o ATeam1.03YEMK foi expresso em astrócitos. Em contraste com o que foi observado nos neurônios, os astrócitos reagiram ao aumento do potássio extracelular por um aumento reversível na relação ATeam FRET, indicando um aumento nos níveis intracelulares de ATP (n = 70 células em N = 5 fatias) (Figura 9B). A exposição subsequente à isquemia química resultou, como esperado, em uma grande queda na relação ATeam FRET, indicativa do esgotamento nominal do ATP intracelular(Figura 9B).
Discussão
Aqui, demonstramos um procedimento para a expressão específica do tipo célula de ATeam1.03YEMK, um nanosensor14geneticamente codificado por FRET, para medição de alterações nos níveis de ATP em astrocitos ou neurônios em culturas organotípicas de fatia de tecido do cérebro do rato15. Em gravações exemplares, mostramos que um aumento na concentração de potássio extracelular não resulta em uma mudança nas concentrações de ATP nos neurônios, enquanto os níveis ascíticos de ATP aumentam em resposta a essa manipulação. Além disso, nossos resultados demonstram que, após a inibição do metabolismo celular, a relação ATeam1.03YEMK FRET cai rapidamente em ambos os tipos de células, indicando uma rápida diminuição do ATP intracelular.
Expressão de ATeam1.03YEMK em culturas organotípicas da fatia exige a manutenção do tecido na cultura circunstâncias controladas no mínimo 7-10 dias. Alternativamente, ATeam1.03YEMK também pode ser empregado para a medição de ATP em fatias de tecido cerebral agudamente isoladas e em nervos ópticos de camundongos13,15. Medidas em tecido suprido agudamente, no entanto, exigem a geração de animais transgênicos ou uma aplicação estereotática de vetores virais no cérebro, envolvendo experimentação animal e protocolos rigorosos de cuidados com animais. A este respeito, a expressão deYEMK ATeam1.03 em culturas organotípicas da fatia do tecido representa uma alternativa útil e valiosa22,23. Há muitos anos, culturas organotípicas de fatias de tecido servem como um sistema modelo estabelecido para estudar propriedades neurais, conectividade e desenvolvimento24,25,26. Eles não só mantêm a arquitetura geral do tecido e laminação (Figura 1), mas também hospedam as propriedades preferenciais das culturas celulares, como acessibilidade superior e controle direto das condições experimentais. As culturas organotípicas da fatia do tecido são empregadas igualmente rotineiramente para expressar genes extrangeiros usando vetores virais27. Vários tipos de vetores virais foram relatados para entregar transgenes no tecido cerebral16,28. Vetores adenovirais induzir alta expressão em células gliais, mas não neurônios hipocampais16, e pode gerar reatividade glial17. Vetores virais associados ao Adeno, como usados aqui, emergem como uma boa alternativa15,e sua eficácia também foi mostrada in vivo29.
Embora seja usado principalmente para o estudo de propriedades neuronais, estudos recentes estabeleceram que culturas organotípicas de fatia sitipade também podem ser empregadas para a análise de astrócitos. As fatias cultivadas são geralmente cobertas por uma camada de astrócitos reativos19,30 (Figura 5), mas os astrócitos exibem uma morfologia e citoarquitetura mais nativas e não reativas em camadas mais profundas19,30 (Figura 5). No presente estudo, descrevemos um procedimento para a remoção mecânica da cicatriz glial externa, o que resulta em uma melhor acessibilidade experimental e óptica de astrócitos nativos dentro das camadas adequadas do tecido organotypic. Além disso, sua remoção melhora a eficácia da expressão em camadas mais profundas das fatias organotípicas; se a cicatriz glial não for removida, a transdução por AAVs pode tender a ser restrita às camadas celulares superficiais.
Vários fatores mecânicos externos precisam ser considerados ao realizar experimentos em fatias de tecido. Uma variação na velocidade da perfusão do banho pode induzir movimentos da preparação inteira e/ou induzir mudanças no foco, tendo por resultado mudanças transientes artificiais do sinal do sensor. Além disso, tanto os astrócitos quanto os neurônios têm sido relatados para responder à deformação mecânica, como imposta s altas taxas de perfusão32,33. Em nossas mãos, usando uma bomba peristaltic de confiança, junto com a manutenção de volumes pequenos e estáveis de salina entre o tecido e o objetivo (menisco) conduz a um FRET-sinal estável circunstâncias da linha de base na velocidade da perfusão usada aqui (1.5-2.5 mL/min; Figura 8).
No presente estudo, também demonstramos que imagens baseadas em FRET com ATeam1.03YEMK podem ser empregadas para monitorar os níveis de ATP em neurônios e astrócitos. Um meio alternativo introduzido anteriormente para a medição do ATP celular é o chamado ensaio luciferin-lúciferase34,35,36,37. Esta abordagem, no entanto, é baseada em imagens de bioluminescência e fornece apenas uma resolução temporal e espacial bastante baixa, em parte devido aos altos níveis de ruído de fundo. Outro método rotineiramente empregado nos últimos anos foi a imagem de mudanças na concentração intracelular de magnésio usando o verde38,39,40. Essa abordagem diz respeito à observação de que o consumo de ATP resulta na liberação de seu cofator magnésio. A imagem latente com verde do magnésio fornece desse modo somente uma estimativa secundária das mudanças em níveis do ATP. Além disso, o verde magnésio também é sensível às mudanças no cálcio intracelular, introduzindo outra dificuldade na interpretação dos resultados obtidos com este método.
O recente desenvolvimento de nanosensores geneticamente codificados para imagens diretas de metabólitos celulares, portanto, representou um grande passo em frente11,12. Vários sensores diferentes foram gerados que podem ser empregados para medição do ATP intracelular36,41,42,43. Entre eles estão o indicador de ATP fluorescente ratiométrica "QUEEN"41, bem como PercevalHR, que detecta a relação ATP:ADP42. Enquanto a última sonda é uma ferramenta valiosa para o estudo do estado energético das células, requer medição simultânea de alterações no pH42.
ATeam é um nanosensor do qual existem várias variantes, que - entre outras - diferem em sua afinidade vinculativa para atp14. In vitro, ATeam1.03YEMK exibe um Kd de 1.2 mM em 37 °C, que está perto dos níveis de ATP celulares determinados em diferentes tipos de células neuronais, variando de hipotálamo e cerebelo34 a hipocampo37,44,45. Nas medições de cuvette, a redução da temperatura em 10 °C resultou em uma diminuição significativa na afinidade vinculativa de ATeam1.03YEMK para ATP, sugerindo que pode não ser ideal para imagens celulares à temperatura ambiente14. Nosso estudo anterior15, no entanto, demonstrou que o comportamento e a resposta do ATeam1.03YEMK expressos em neurônios e astrócitos a diferentes manipulações é semelhante a quase fisiológico e à temperatura ambiente, indicando que o sensor permite determinação confiável dos níveis intracelulares de ATP em ambas as condições. Além disso, nossos experimentos anteriores abordaram a sensibilidade ao pH do ATeam1.03YEMK expressa dentro das células15,mostrando que é insensível às mudanças no pH intracelular em cerca de 0,1-0,2 unidades pH. Se o Kd na faixa de mM baixo é uma preocupação, variantes alternativas ATeam pode ser usado14,entre eles variantes em mudança vermelha de ATeam ("GO-ATeam")43.
Nossos experimentos usando OYEMK ATeam1.03 demonstram que um aumento na concentração de potássio extracelular em alguns mM apenas (de 3 para 8 mM) resulta em um aumento transitório na relaçãoYEMK ATeam1.03 em astrocitos na cultura de fatia organótípica. Esta observação confirma estudos anteriores15,46 e indica claramente que os astrócitos respondem à liberação de potássio por neurônios ativos com um aumento na sua produção atp, principalmente provável como conseqüência de uma estimulação do Na+/ K+-ATPase e o Na+/ HCO3- cotransportador, respectivamente47,48. Em contraste com isso, os neurônios não mostraram uma resposta, que está em linha com o trabalho anterior também15. Ambos os tipos de células, no entanto, reagiram rapidamente e fortemente à inibição da glicolyse celular e respiração mitocondrial, como mostrado antesde 15. Em condições de isquemia química, as proporções de FRET da ATeam caíram para um novo nível estável, indicando um esgotamento nominal do ATP celular. O último resultado sugere que ambos os neurônios, bem como astrócitos apresentam um consumo relevante de ATP também condições de estado estável, sem estimulação adicional por ativação sináptica ou aplicação de neurotransmissores. Em conjunto, concluímos que a imagem de imagens baseadas em FRET com nanosensores geneticamente codificados, entre eles ATeam1.03YEMK,fornecerá uma abordagem valiosa para elucidar os processos celulares responsáveis por mudanças nos níveis intracelulares de ATP e no consumo de ATP celular em diferentes condições.
Divulgações
Os autores não declaram interesses concorrentes. Os autores receberam apoio financeiro permitindo a publicação de acesso aberto pela Nikon Microscope Solutions, Düsseldorf, Alemanha, que produz instrumentos utilizados no artigo de vídeo. A empresa não estava envolvida na concepção dos experimentos aqui apresentados, nem na execução, nem no manuseio de dados, nem na escrita do manuscrito.
Agradecimentos
Os autores desejam agradecer a Claudia Roderigo e Simone Durry por assistência técnica especializada. Agradecemos ao Dr. Niklas J. Gerkau e ao M.Sc. Joel Nelson pela ajuda na preparação das culturas organotípicas. A pesquisa no laboratório do autor foi financiada pela Associação Alemã de Pesquisa (DFG; PARA 2795: Ro 2327/13-1 e SPP 1757: Ro 2327/8-2 a CRR; e SPP 1757: Financiamento start-up da Glia Young para rl).
Materiais
| Name | Company | Catalog Number | Comments |
| 2-deoxyglucose | Alfa Aesar | L07338 | Non-metabolizable glucose analog |
| 36-IMA-410-019 Argon laser | Melles Griot | 488 nm wavelength argon | |
| Ascorbic acid | Carl Roth | 3525.1 | Antioxidant, Vitamin C |
| band pass filters 483/32 | AHF Analysentechnik AG | Splitter compatible emmision filter | |
| band pass filters 542/27 | AHF Analysentechnik AG | Splitter compatible emmision filter | |
| Beamsplitter T 455 LP | AHF Analysentechnik AG | Excitation dichroic mirror | |
| Beamsplitter T 505 LPXR | AHF Analysentechnik AG | Splitter dichroic | |
| Confocal laser scannig microscope C1 | Nikon Microscope Solutions | Modular confocal microscope system C1 | |
| Data processing Origin Pro 9.0.0 (64-bit) | OriginLab corporation | Scientific graphing and data analysis software | |
| D-glucose monohydrate | Caelo | 2580-1kg | |
| DPBS | GIBCO/Life | 14190250 | Dulbecco's phosphate-buffered saline |
| Eclipse E 600FN upright microscope | Nikon Microscope Solutions | ||
| Eclipse FN1 upright microscope | Nikon Microscope Solutions | ||
| Experimental chamber | custom build | Perfusion chamber for live-cell imaging | |
| EZ-C1 Silver Version 3.91 | Nikon Microscope Solutions | Imaging software for confocal microscope | |
| Hanks' Balanced Salt solution | Sigma-Aldrich | H9394 | With Phenol Red for pH monitoring |
| HERAcell 150 | Thermo Scientific | CO2 incubator HERAcell ® 150 with decontamination routine | |
| HERAsafe KS/KSP | Thermo Scientific | Safety Cabinet | |
| Horse serum | GIBCO/Life | 26050088 | Heat inactivated |
| Huygens Professional | SVI Imaging | Deconvolution software | |
| Image J 1.52i | Wayne Rasban national Institute of Health | Image processing Software available in the public domain | |
| Insulin | Sigma-Aldrich | I6634 | Insulin from bovine pancreas |
| IP serie peristaltic pump | Ismatec | High-precisionmulti-channel pump | |
| Layout software, Illustrator CS6 | Adobe | Vector graphics editor | |
| L-glutamine | GIBCO/Life | 25030024 | |
| Microm HM 650 V | Thermo Scientific | Vibration microtome. Thermo scientific discontinued the production of the device in the meantime. Any other slicer or tissue chopper siutable for slicing living tissue is fine, too. | |
| Microscope stage | custom build | ||
| Microsoft Excel 16 | Microsoft | Spreadsheet software for basic data processing | |
| Millicell culture insert | Merck Millipore | PICM0RG50 | Hydrophilized PTFE, pore size 0.4 μm |
| Minimum Essential Medium Eagle | Sigma-Aldrich | M7278 | Synthetic cell culture media |
| Monochromator Polychrome V | Thermo Scientific/FEI | Ultra fast switching monochromator | |
| NaN3 (Sodium Azide) | Sigma-Aldrich | S-8032 | Mitochondrial inhibitor (complex IV inhibitor). CAUTION: Azide is toxic. Be aware not to accidentally ingest or inhale it, and prevent ist absoption through the skin. |
| Nikon Fluor 40x / 0.80 W DIC M ∞/0 WD 2.0 | Nikon Microscope Solutions | Water Immersion Microscope Objective | |
| NIS Elements 4.50 advanced Research | Nikon Microscope Solutions | Imaging software. Upgraded version for FRET imaging | |
| ORCA-Flash4.0 | Hamamatsu Photonics | Digital CMOS camera | |
| Perfusion tubing | Pro Liquid GmbH | Tygon tubing, 1.52 x 322 mm (Wd: 0.85) | |
| Photoshop CS 6 Version 13.0 | Adobe | Image processing software | |
| Sodium L-lactate | Sigma-Aldrich | 71718-10G | |
| ssAAV-2/2-hSyn1-ATeam1.03YEMK-WPRE-hGHp(A) | ETH Zürich | v244 | Single-stranded AAV vector that induces the expression of ATeam1.03YEMK under the control of the human synapsin 1 promoter fragment hSyn1. |
| ssAAV-5/2-hGFAP-hHBbI/E-ATeam1.03YEMK-WPRE-bGHp(A) | ETH Zürich | v307 | Single-stranded AAV vector that induces the expression of ATeam1.03YEMK under the control of the human glial fibrillary acidic protein promoter fragment ABC1D. |
| WVIEW GEMINI optic system | Hamamatsu Photonics | Emission Image Splitter |
Referências
- Sweadner, K. J. Isozymes of the Na+/K+-ATPase. Biochimica et Biophysica Acta. 988, 185-220 (1989).
- Clapham, D. E. Calcium signaling. Cell. 131, 1047-1058 (2007).
- Cotter, K., Stransky, L., McGuire, C., Forgac, M. Recent Insights into the Structure, Regulation, and Function of the V-ATPases. Trends in Biochemical Sciences. 40, 611-622 (2015).
- Harris, J. J., Jolivet, R., Attwell, D. Synaptic energy use and supply. Neuron. 75, 762-777 (2012).
- Brown, A. M., Ransom, B. R. Astrocyte glycogen and brain energy metabolism. Glia. 55, 1263-1271 (2007).
- Hertz, L., et al. Roles of astrocytic Na+,K+-ATPase and glycogenolysis for K+ homeostasis in mammalian brain. Journal of Neuroscience Research. 93, 1019-1030 (2015).
- Allaman, I., Belanger, M., Magistretti, P. J. Astrocyte-neuron metabolic relationships: For better and for worse. Trends in Neurosciences. 34, 76-87 (2011).
- Barros, L. F., Deitmer, J. W. Glucose and lactate supply to the synapse. Brain Research Reviews. 63, 149-159 (2010).
- Diaz-Garcia, C. M., et al. Neuronal Stimulation Triggers Neuronal Glycolysis and Not Lactate Uptake. Cell Metabolism. 26, 361-374 (2017).
- Diaz-Garcia, C. M., et al. Quantitative in vivo imaging of neuronal glucose concentrations with a genetically encoded fluorescence lifetime sensor. Journal of Neuroscience Research. 97, 946-960 (2019).
- Barros, L. F., et al. Current technical approaches to brain energy metabolism. Glia. 66, 1138-1159 (2018).
- Tantama, M., Hung, Y. P., Yellen, G. Optogenetic reporters: Fluorescent protein-based genetically encoded indicators of signaling and metabolism in the brain. Progress in Brain Research. 196, 235-263 (2012).
- Trevisiol, A., et al. Monitoring ATP dynamics in electrically active white matter tracts. eLife. 6, e24241(2017).
- Imamura, H., et al. Visualization of ATP levels inside single living cells with fluorescence resonance energy transfer-based genetically encoded indicators. Proceedings of the National Academy of Sciences of the United States of America. 106, 15651-15656 (2009).
- Lerchundi, R., et al. FRET-based imaging of intracellular ATP in organotypic brain slices. Journal of Neuroscience Research. , 1-13 (2018).
- Ehrengruber, M. U., et al. Gene transfer into neurons from hippocampal slices: comparison of recombinant Semliki Forest Virus, adenovirus, adeno-associated virus, lentivirus, and measles virus. Molecular and Cellular Neurosciences. 17, 855-871 (2001).
- Woo, J., et al. Functional Characterization of Resting and Adenovirus-Induced Reactive Astrocytes in Three-Dimensional Culture. Experimental Neurobiology. 26, 158-167 (2017).
- Close, B., et al. Recommendations for euthanasia of experimental animals: Part 2. DGXT of the European Commission. Laboratory Animals. 31, 1-32 (1997).
- Benediktsson, A. M., Schachtele, S. J., Green, S. H., Dailey, M. E. Ballistic labeling and dynamic imaging of astrocytes in organotypic hippocampal slice cultures. Journal of Neuroscience Methods. 141, 41-53 (2005).
- Lanjakornsiripan, D., et al. Layer-specific morphological and molecular differences in neocortical astrocytes and their dependence on neuronal layers. Nature Communications. 9, 1623(2018).
- Bushong, E. A., Martone, M. E., Ellisman, M. H. Examination of the relationship between astrocyte morphology and laminar boundaries in the molecular layer of adult dentate gyrus. The Journal of Comparative Neurology. 462, 241-251 (2003).
- Frotscher, M., Zafirov, S., Heimrich, B. Development of identified neuronal types and of specific synaptic connections in slice cultures of rat hippocampus. Progress in Neurobiology. 45, vii-xxviii (1995).
- Galimberti, I., et al. Long-term rearrangements of hippocampal mossy fiber terminal connectivity in the adult regulated by experience. Neuron. 50, 749-763 (2006).
- Stoppini, L., Buchs, P. A., Muller, D. A simple method for organotypic cultures of nervous tissue. Journal of Neuroscience Methods. 37, 173-182 (1991).
- Forster, E., Zhao, S., Frotscher, M. Laminating the hippocampus. Nature Reviews. Neuroscience. 7, 259-267 (2006).
- Holopainen, I. E. Organotypic Hippocampal Slice Cultures: A Model System to Study Basic Cellular and Molecular Mechanisms of Neuronal Cell Death, Neuroprotection, and Synaptic Plasticity. Neurochemical Research. 30, 1521-1528 (2005).
- Teschemacher, A. G., et al. Targeting specific neuronal populations using adeno- and lentiviral vectors: applications for imaging and studies of cell function. Experimental Physiology. 90, 61-69 (2005).
- Kantor, B., Bailey, R. M., Wimberly, K., Kalburgi, S. N., Gray, S. J. Methods for gene transfer to the central nervous system. Advances in Genetics. 87, 125-197 (2014).
- Mächler, P., et al. In Vivo Evidence for a Lactate Gradient from Astrocytes to Neurons. Cell Metabolism. 23, 94-102 (2016).
- Schreiner, A. E., Berlinger, E., Langer, J., Kafitz, K. W., Rose, C. R. Lesion-Induced Alterations in Astrocyte Glutamate Transporter Expression and Function in the Hippocampus. ISRN Neurology. 2013, 893605(2013).
- Haber, M., Zhou, L., Murai, K. K. Cooperative astrocyte and dendritic spine dynamics at hippocampal excitatory synapses. The Journal of Neuroscience: The Official Journal of the Society for Neuroscience. 26, 8881-8891 (2006).
- Neary, J. T., Kang, Y., Tran, M., Feld, J. Traumatic injury activates protein kinase B/Akt in cultured astrocytes: role of extracellular ATP and P2 purinergic receptors. Journal of Neurotrauma. 22, 491-500 (2005).
- Xia, J., et al. Neurons respond directly to mechanical deformation with pannexin-mediated ATP release and autostimulation of P2X7 receptors. The Journal of Physiology. 590, 2285-2304 (2012).
- Ainscow, E. K., Mirshamsi, S., Tang, T., Ashford, M. L., Rutter, G. A. Dynamic imaging of free cytosolic ATP concentration during fuel sensing by rat hypothalamic neurones: evidence for ATP-independent control of ATP-sensitive K+ channels. The Journal of Physiology. 544, 429-445 (2002).
- Arcuino, G., et al. Intercellular calcium signaling mediated by point-source burst release of ATP. Proceedings of the National Academy of Sciences of the United States of America. 99, 9840-9845 (2002).
- Rajendran, M., Dane, E., Conley, J., Tantama, M. Imaging Adenosine Triphosphate (ATP). The Biological Bulletin. 231, 73-84 (2016).
- Rangaraju, V., Calloway, N., Ryan, T. A. Activity-driven local ATP synthesis is required for synaptic function. Cell. 156, 825-835 (2014).
- Chatton, J. Y., Pellerin, L., Magistretti, P. J. GABA uptake into astrocytes is not associated with significant metabolic cost: implications for brain imaging of inhibitory transmission. Proceedings of the National Academy of Sciences of the United States of America. 100, 12456-12461 (2003).
- Magistretti, P. J., Chatton, J. Y. Relationship between L-glutamate-regulated intracellular Na+ dynamics and ATP hydrolysis in astrocytes. Journal of Neural Transmission (Vienna). 112, 77-85 (2005).
- Langer, J., et al. Rapid sodium signaling couples glutamate uptake to breakdown of ATP in perivascular astrocyte endfeet. Glia. 65, 293-308 (2017).
- Yaginuma, H., et al. Diversity in ATP concentrations in a single bacterial cell population revealed by quantitative single-cell imaging. Scientific Reports. 4, 6522(2014).
- Tantama, M., Martinez-Francois, J. R., Mongeon, R., Yellen, G. Imaging energy status in live cells with a fluorescent biosensor of the intracellular ATP-to-ADP ratio. Nature Communications. 4, 2550(2013).
- Nakano, M., Imamura, H., Nagai, T., Noji, H. Ca2+ Regulation of Mitochondrial ATP Synthesis Visualized at the Single Cell Level. ACS Chemical Biology. 6, 709-715 (2011).
- Mollajew, R., Toloe, J., Mironov, S. L. Single KATP channel opening in response to stimulation of AMPA/kainate receptors is mediated by Na+ accumulation and submembrane ATP and ADP changes. The Journal of Physiology. 591, 2593-2609 (2013).
- Pathak, D., et al. The Role of Mitochondrially Derived ATP in Synaptic Vesicle Recycling. The Journal of Biological Chemistry. 290, 22325-22336 (2015).
- Karus, C., Mondragao, M. A., Ziemens, D., Rose, C. R. Astrocytes restrict discharge duration and neuronal sodium loads during recurrent network activity. Glia. 63, 936-957 (2015).
- Larsen, B. R., Stoica, A., MacAulay, N. Managing Brain Extracellular K+ during Neuronal Activity: The Physiological Role of the Na+/K+-ATPase Subunit Isoforms. Frontiers in Physiology. 7, 141(2016).
- Ruminot, I., et al. NBCe1 mediates the acute stimulation of astrocytic glycolysis by extracellular K+. The Journal of Neuroscience : The Official Journal of the Society for Neuroscience. 31, 14264-14271 (2011).
Reimpressões e Permissões
Solicitar permissão para reutilizar o texto ou figuras deste artigo JoVE
Solicitar PermissãoThis article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. Todos os direitos reservados