
Method Article
Gelişimsel ve Hastalık Modelleme Uygulamaları için hiPSC Kaynaklı Bağırsak Organoidlerinin Üretilmesi
Bu Makalede
Özet
Bu protokol, insan pluripotent hücrelerinin bağırsak organoidlerine farklılaşmasına izin verir. Protokol, hücreleri kesin endoderm, arka bağırsak endodermi ve ardından bağırsak epiteli popülasyonuna farklılaştırarak normal insan gelişimini taklit eder. Bu, protokolü hem bağırsak gelişimini hem de hastalık modelleme uygulamalarını incelemek için uygun hale getirir.
Özet
hiPSC'den türetilen bağırsak organoidleri, farklılaşmış hücrelerden karmaşık 3D yapılara kendi kendine birleşen, insan bağırsak epitelini temsil eden ve kript/villus benzeri yapılar sergiledikleri epitel yapılarıdır. Burada, hiPSC'lerin kesin endoderme kademeli olarak farklılaşmasıyla hiPSC'den türetilmiş bağırsak organoidlerinin üretimini, daha sonra 3D kültür koşullarına aktarılmadan önce arka bağırsak epiteli oluşturmak üzere posteriorize edildiğini açıklıyoruz. 3D kültür ortamı, SB202190, A83-01, Gastrin, Noggin, EGF, R-spondin-1 ve CHIR99021 ile desteklenmiş hücre dışı matristen (ECM) (örneğin, Matrigel veya diğer uyumlu ECM) oluşur. Organoidler her 7 günde bir pasaja uğrarlar, burada taze hücre dışı matrise aktarılmadan önce mekanik olarak bozulurlar ve genişlemelerine izin verilir. QPCR ve immünositokimya, hiPSC'den türetilen bağırsak organoidlerinin goblet hücreleri, Paneth hücreleri ve enterositler dahil olmak üzere olgun bağırsak epitel hücre tiplerini içerdiğini doğrulamaktadır. Ek olarak, organoidler, epitel hücrelerinin apikal yüzeyinde lokalize villin ekspresyonu ile polarizasyon kanıtı gösterir.
Elde edilen organoidler, insan bağırsak gelişiminin yanı sıra inflamatuar bağırsak hastalığı da dahil olmak üzere çok sayıda insan bağırsak hastalığını modellemek için kullanılabilir. Bağırsak iltihabını modellemek için organoidler, TNF-α, TGF-β ve bakteriyel LPS gibi enflamatuar mediatörlere maruz bırakılabilir. Proinflamatuar sitokinlere maruz kalan organoidler, yanıt olarak inflamatuar ve fibrotik bir fenotip gösterir. IBD'li hastalardan elde edilen sağlıklı ve hiPSC'lerin eşleştirilmesi, IBD'yi yönlendiren mekanizmaların anlaşılmasında yararlı olabilir. Bu, erken hastalık teşhisine yardımcı olmak için yeni terapötik hedefleri ve yeni biyobelirteçleri ortaya çıkarabilir.
Giriş
Pluripotent kök hücrelerin (PSC'ler) kendini yenileme ve insan vücudunun herhangi bir hücre tipine farklılaşma yeteneği gibi özellikleri, onları gelişim, hastalık patolojisi ve ilaç testi çalışmalarında değerli araçlar haline getirir1. İnsan kaynaklı pluripotent kök hücreler (hiPSC), hastalardan elde edilebildikleri ve hastalık fenotip2,3'ten sorumlu genomu doğrudan yakalayabildikleri için hastalık modelleme çalışmaları için özellikle yararlıdır. Bu tür hiPSC'ler, genetik defektten etkilenen hücre tipine göre ayırt edilebilir ve hastalığın moleküler mekanizmasının dikkatli bir şekilde incelenmesine olanak tanır4.
İnsan PSC'leri için farklılaşma protokolleri, soy bağlılığını ve spesifikasyonunu yöneten spesifik sinyal yollarının aktivasyonu veya inhibisyonu yoluyla hücrelerin ana gelişim aşamaları yoluyla farklılaşmasını yönlendirmeyi amaçlar. HiPSC'nin pluripotent bir durumda tutulması, orta düzeyde Aktivin A (Act-A) sinyali gerektirirken, 3 gün boyunca yüksek dozda Act-A, hiPSC'yi kesin bir endoderm (DE) kaderinebağlar 5,6. Act-A ve Wnt yolakları DE'nin anterior-posterior özdeşliğini yönlendirir. Act-A ile sinyalleme, CDX2 gibi arka bağırsak (HG) genlerinin ekspresyonunu bloke ederken, HHEX, HNF4α ve GATA4 gibi ön bağırsak (FG) belirteçlerini indükler. Wnt sinyali, daha sonra bir HG gen ekspresyon profilini 7,8 benimseyen DE'nin posteriorizasyonunu indükler. HG hücre kimliği oluşturulduktan sonra, farklılaşma 2D'den 3D'ye taşınabilir ve bağırsak organoidlerinin oluşumuna yönlendirilebilir.
Bağırsak organoidleri tipik olarak laminin, kollajen IV ve entaktinden oluşan ve EGF, FGF, PDGF ve IGF-1 gibi büyüme faktörleri ile zenginleştirilmiş bir 3D hücre dışı matris (örneğin, Matrigel veya diğer uyumlu ECM'ler) tabanlı bir kültür sistemi9'da kültürlenir. Organoidler, uzun süreli kültür sırasında bağırsak kök hücre büyümesini ve çoğalmasını uyarmak ve desteklemek için Gastrin, Noggin ve CHIR içeren tanımlanmış bir ortamda kültürlenir.
Bağırsak epitel hücreleri hücre dışı matrise gömüldükten sonra, bağırsak kriptleri oluşmaya ve genişlemeye başlar ve sonunda sferoidler oluşturur. Bunlar, bağırsak epitelinin fizyolojik işleyişini taklit eden organoid yapılara olgunlaşır. Organoidler tipik olarak, fonksiyonel ve gen ekspresyon profillerinde önemli bir kayıp olmadan 1 yıldan fazla kültürlenebilir. Organoidlerin enzimatik sindirimi kullanılarak haftalık olarak pasaj gereklidir, bunlar daha sonra tam organoidler halinde kendi kendine yeniden birleşir.
Yerleşik organoid çizgiler, Crohn Hastalığı, Ülseratif Kolit ve kolorektal kanserlerdahil olmak üzere bağırsakla bağlantılı çok sayıda bozukluğun güvenilir bir modeli olarak kullanılabilir 10,11,12,13. Bu, bu bozukluklarla bağlantılı insan genlerini ifade ettikleri ve insan dokusunda meydana gelene daha çok benzeyen dış uyaranlara daha fazla yanıt verdikleri için hayvan hücrelerine tercih edilen bir modeldir.
Protokol
NOT: Aşağıda ayrıntıları verilen tüm doku kültürü çalışmaları, Sınıf II laminer akış başlığında yapılmalıdır.
1. İnsan kaynaklı pluripotent kök hücre (hiPSC'ler) hindgut endodermine farklılaşması
- Bir hücre kültürü şişesini hücre dışı matrisle kaplamak için, önce gereken matris miktarını hesaplayın. İşte Matrigel ile kaplanmış 12 oyuklu bir plakanın bir örneği. 12 oyuklu bir plakayı kaplamak için gerekli hücre dışı matris miktarını hesaplayın. Aşağıdaki formülü kullanın:

NOT: Hücre kültürü kaplarını tercih edilen hücre dışı matris ile kaplayın. Matrigel kullanıyorsanız, farklılaşma için tohumlamadan 24 saat önce 0.035 mg /cm2'lik bir konsantrasyon kullanın. Ürün sayfasındaki hücre dışı matrisin parti konsantrasyonunu kontrol edin ve üreticinin talimatlarına göre deneylerden önce daha küçük alikotlar hazırlayın. - Bir p1000 pipeti kullanarak gerekli miktarda hücre dışı matrisi soğuk DMEM ortamında seyreltin.
- 5 mL'lik bir şerit kullanarak iyice karıştırın.
- Her bir oyuklu plakaya 500μL seyreltilmiş matris ekleyin. Seyreltilmiş hücre dışı matrisi eşit olarak dağıtmak için plakayı hafifçe sallayın ve 37 ° C'de en az 12 saat inkübe edin.
- Hücreleri plakaya tohumlamadan önce, fazla hücre dışı matrisi çıkarmak için her bir kuyuyu 500 μL PBS ile yıkayın. Bu, farklılaşma sırasında hücre ayrılmasını önleyecektir.
- Hücre dışı matrisin kurumasını önlemek için tohumlamaya hazır olana kadar son yıkamayı kuyularda bırakın.
- Kesin endoderm (DE) veya hindgut'a (HG) doğru hiPSC'lerin farklılaşması için tohum atmak için, tercih edilen bir bakım ortamında (örneğin: temel 8 ortam) kültürlenmiş bir hiPSC şişesi alın ve ortamı aspire edin. (Burada, hücreleri 25.000 hücre / cm2'de tohumluyoruz).
NOT: Tohumlama yoğunluğu, başarılı bir farklılaşma için çok önemlidir ve her bir hiPSC hücre hattı için optimize edilmesi gerekir. - Şişedeki hücreleri 5 mL PBS ile yıkayın. PBS'yi aspire edin.
- 2.5 mL hücre ayrışma çözeltisi ekleyin (ör., TrypLE) ve RT'de 4 dakika bekletin. Hücre ayrışma solüsyonunu aspire edin ve hücreleri ayırmak için şişeye hafifçe vurun.
- Şişeyi 37 ° C'ye ısıtılmış 5 mL DMEM ortamı ile yıkayın ve tüm hücreleri 15 mL'lik bir tüpte toplayın.
- 10 μL resüspanse hücre çözeltisi alın ve hücre yoğunluğunu bir hemositometre ile ölçün.
- 1.05 x 106 hücre toplamak için yeterli miktarda hücre süspansiyonu alın ve 15 mL'lik bir tüpe yerleştirin.
- 3 dakika boyunca 160 x g'da döndürün. Hücreler santrifüjlenirken, hücre dışı matris kaplı 12 oyuklu plakadan PBS'yi aspire edin.
- Santrifüjlemeyi takiben, süpernatanı aspire edin ve hücre peletini ROCK inhibitörü (10 μM) ile 12 mL bakım ortamında yeniden süspanse edin.
- Bir p1000 pipeti kullanarak, 12 oyuklu plakanın her bir oyuğuna 1 mL hücre süspansiyonu ekleyin. Kuyucuklar arasında eşit dağılımı sağlamak için hücreleri iyice süspanse edin.
- Hücreleri plakanın oyuklarına dağıtmak için plakayı hafifçe sallayın, ancak hücreleri kuyucukların ortasında yoğunlaştıracağından ortamın dönmesini önleyin.
- 37 °C, %5CO2 inkübatöre yerleştirin.
- Tohumlamadan 24 saat sonra medyayı yalnızca bakım medyasına (örn. temel 8 medya) değiştirin (ROCK inhibitörü yok).
- DE'ye farklılaşmaya başlamak için, endoderm bazal ortamını Tablo 1'e göre hazırlayın.
NOT: Farklılaşmanın başlangıcında hücreler %60-80 arasında birleşim şeklinde olmalıdır. - Endoderm bazal ortama Activin A (100 ng/mL) ve Wnt3 (50 ng/mL) ekleyerek 12 mL DE ortamı hazırlayın.
- Ortamı 37 °C'ye ısıtın.
- Doku kültürü plakasının veya şişesinin her bir oyuğundan ortamı aspire edin.
- 12 oyuklu plakanın her bir oyuğuna 1 mL DE ortamı ekleyerek DE farklılaşmasını başlatın.
- DE diferansiyasyonunu (D1 DE) başlattıktan 24 saat sonra, yeni DE ortamı hazırlayın ve ortam değişimi gerçekleştirin.
- Adım 1.24'ü 48 saatte tekrarlayın (D2 DE). Bu aşamada çok fazla hücre ölümü varsa, ortam değiştirmeden önce tüm kuyucukları 500 μL PBS ile yıkayın.
NOT: Başarılı endoderm farklılaşmasını doğrulamak için, SOX17 ekspresyonunu değerlendirmek için akış sitometrisi yapın. Normalde hücrelerin %>80'inin D3 DE ile SOX17 pozitif olmasını bekleriz. SOX17'nin ekspresyonu yetersiz ise, hücre tohumlama yoğunluğu ve Act-A konsantrasyonları optimize edilmelidir. - HG farklılaşmasını bu noktada, yani DE farklılaşmasının başlamasından 72 saat sonra başlatın. Endoderm bazal ortama CHIR99021 (3 μM) ve RA (1 μM) ekleyerek 12 mL HG ortamı hazırlayın (Tablo 1).
NOT: D3 DE'de hücre tek tip bir tek tabaka oluşturmuş olmalıdır. Optimal DE farklılaşması, farklılaşmanın sonraki aşamaları için çok önemlidir. - DE ortamını aspire edin.
- 12 oyuklu plakanın her bir oyuğuna 1 mL HG ortamı ekleyerek HG farklılaşmasını başlatın.
- Günlük medya değişiklikleri ile 4 gün boyunca farklılaşmaya devam edin.
NOT: DE posteriorizasyonunun başarısını belirlemek için, CDX2 ekspresyonunu değerlendirmek için akış sitometrisi yapın. HG farklılaşmasının D4'ü ile normalde hücrelerin %>80'inin CDX2 pozitif olmasını bekleriz. Posteriorizasyon yetersiz ise, CHIR99021 konsantrasyonu optimize edilmelidir. - RNA ekstraksiyonu için bir numune toplamak için, farklılaşma ortamını aspire edin ve kuyuyu 500 ul PBS ile yıkayın.
- PBS'yi aspire edin ve bir RNA ekstraksiyon kitinden uygun hacimde hücre lizis tamponu ekleyin.
- Bir p1000 pipeti kullanarak, tüm hücrelerin parçalanmasını sağlamak için kuyunun dibini kazıyın.
- Hücre lizatını aspire edin ve temiz bir tüpe koyun.
- RNA ekstraksiyonuna devam edin veya lizatı RNA ekstraksiyonu için hazır olana kadar -20 °C'de dondurun.
- İmmün boyama için, farklılaşma ortamını aspire edin ve kuyucukları 500 μL PBS ile yıkayın.
- PBS'yi aspire edin ve 500 μL% 4 PFA ekleyin.
NOT: PFA toksiktir. Uygun KKD kullanın ve PFA imhası için yerel laboratuvar prosedürlerini izleyin. - 4 °C'de 20 dakika inkübe edin.
- PFA'yı çıkarın ve kuyuları üç kez 500 μL PBS ile yıkayın.
- İmmün boyama yapmaya hazır olana kadar hücreler üzerindeki son PBS yıkamasını bırakın.
2. Bağırsak organoidlerinin pasajı
- Bağırsak bazal ortamını Tablo 2'ye göre hazırlayın.
- 2D'den 3D hücre kültürüne aktarmak için, hiPSCS'den DE hücreleri oluşturmak için 6 oyuklu bir plaka kullanın. Hücrelerin tek tabakasını ayırın, 5 mL'lik bir şerit kullanarak 6 hücreli plakayı oluşturun.
- Bir hücre peleti üretmek için 1 dakika boyunca 400 x g'da santrifüjlemeden önce hücreleri 15 mL'lik bir santrifüj tüpünde toplayın.
- Büyüme faktörleri içeren bağırsak büyüme ortamında hücre peletini yeniden süspanse edin: SB202190 (10 μM), A83-01 (500 nM), Gastrin (10 nM), Noggin (100 ng/μL), EGF (500 ng/μL), R-Spondin1 (100 ng/mL), CHIR99021 (6 μM), ROCK inhibitörü (10 μM).
- Kaplanan kuyucuk sayısına bağlı olarak uygun hacimde hücre dışı matris ekleyin. Bu, aşağıdaki denklemler kullanılarak hesaplanabilir.
[1] Hücre dışı matriks/ortamın toplam hacmi (μL) = 30 μL * 48 oyuklu plakanın kuyucuk sayısı
[2] Gerekli hücre dışı matriks hacmi (μL) = [1] * 2/3'ten gelen cevap
[3] Gerekli ortam hacmi (μL) = [1] * 1/3'ten yanıt - 48 oyuklu bir plakanın her bir oyuğunun ortasına 30 μL hücre süspansiyonu ekleyin.
- Hücre dışı matrisin sertleşmesine izin vermek için plakayı 37 ° C'lik bir inkübatörde en az 5 dakika inkübe edin.
- Hücre dışı matriks kubbeleri ayarlandıktan sonra, her bir oyuğa tüm büyüme faktörlerini içeren 300 μL bağırsak büyüme ortamı ekleyin.
- Plakayı %5 CO2 içeren 37 °C'lik bir inkübatöre geri koyun.
NOT: 48 saat sonra hiçbir organoid gözlenemezse ve/veya önemli hücre ölümü varsa, ROCKi ve NOGGIN konsantrasyonlarının optimize edilmesi gerekebilir. - Organoidleri geçmek için, organoidlerin yoğunluğunu ve boyutunu değerlendirmek ve geçişe ihtiyaç duyup duymadıklarını belirlemek için hücreleri ışık mikroskobu altında gözlemleyin.
- Hücre kültürü ortamını buz gibi soğuk PBS ile değiştirin.
NOT: Organoid kültürlerin yaklaşık 5-7 günde bir bölünmesi gerekecektir. Organoid lümen içinde belirgin bir hücre kalıntısı birikimi ve çevredeki bağırsak büyüme ortamının sararması durumunda bağırsak organoid kültürlerinin geçirilmesi gerekir. - 5 mL'lik bir şerit ile kazıma hareketi kullanarak organoidleri ve hücre dışı matris küresini plakadan mekanik olarak ayırın.
- Organoidleri her bir kuyucuktan 15 mL'lik bir santrifüj tüpüne toplayın.
- Organoidleri peletlemek için organoid süspansiyonu 400 x g'da 1 dakika santrifüjleyin. Süpernatanı, hücre peletinin tepesine ulaşana kadar aspire edin, görünür hücre dışı matris tabakasına ulaştığında dikkatli olun.
- Peletleri 15 mL buz gibi soğuk PBS'de yeniden süspanse edin.
NOT: Bu adım, ilk dönüşte çıkarılmayan kalan hücre dışı matrisi yıkamaya yarar. - 400 x g'da 1 dakika santrifüjleyin. Organoidleri içeren pelete kadar ortamı aspire edin.
- 1 mL buz gibi soğuk PBS'de tekrar süspanse edin. Bir p200 pipeti kullanarak, birkaç kez yukarı ve aşağı pipetleyerek sağlam organoidleri manuel olarak parçalayın.
NOT: Daha fazla ayrışmaları gerekip gerekmediğini belirlemek için ışık mikroskobu kullanarak organoidlerin boyutunu gözlemleyin. - Büyüme faktörleri olmadan 9 mL ortam ekleyin.
- 400 x g'da 1 dakika santrifüjleyin. Organoidler peletine kadar süpernatanı aspire edin.
- Aşağıdaki denklemleri kullanarak gerekli hücre dışı matris ve ortam miktarını hesaplayın:
[1] Hücre dışı matriks/ortamın toplam hacmi (μL) = 30 μL * 48 oyuklu plakanın kuyucuk sayısı
[2] Gerekli hücre dışı matriks hacmi (μL) = [1] * 2/3'ten gelen cevap
[3] Gerekli ortam hacmi (μL) = [1] * 1/3'ten yanıt - Organoid peleti, büyüme faktörleri (Noggin ve ROCK inhibitörü dahil) ile hesaplanan bağırsak ortamı hacminde yeniden süspanse edin.
- Bu hücre süspansiyonuna gerekli hacimde hücre dışı matris ekleyin ve organoidlerin eşit dağılımını sağlamak için yeniden süspanse edin.
- Bu süspansiyonun 30 μL'sini 48 oyuklu bir plakanın (tercihen 37 °C'lik bir inkübatörde önceden ısıtılmış) her bir oyuğunun ortasına pipetleyin.
- Hücre dışı matris ayarlanana kadar plakayı 37 ° C'lik bir inkübatöre 5 dakika geri koyun.
- Bağırsak ortamını büyüme faktörleri (+ ROCK inhibitörü) ile hazırlayın (48 oyuklu plaka başına yaklaşık 17 mL).
- Her kuyucuğa 300 μL ekleyin.
- 37 °C'de %5 CO2'de inkübe edin.
- Geçişten sonra, bağırsak organoidleri için ortamı aspire edin ve her 2-4 günde bir büyüme faktörleri (Noggin ve ROCK inhibitörü olmadan) içeren taze bağırsak ortamı ile değiştirin.
NOT: 48 saat sonra herhangi bir organoid gözlenemezse ve/veya önemli hücre ölümü varsa, ROCKi ve NOGGIN konsantrasyonlarının optimize edilmesi gerekebilir.
NOT: Bağırsak organoidleri, in vivo olarak bir dizi inflamatuar mediatöre yanıt verir. TNFa, çeşitli enflamatuar süreçlerde yer alan bir hücre sinyal proteinidir. - Bağırsak organoidlerinde inflamatuar bir yanıtı tetiklemek için, TNFa'yı sadece bazal ortamda 40 ng / mL konsantrasyonda hazırlayın.
- 48 oyuklu plakadan aspire ortamı.
- 40 ng/mL TNFa içeren 300 μL hazırlanmış bazal ortam ekleyin.
- Pro-inflamatuar bir ortamı çoğaltmak için plakayı 37 ° C'de 48 saat inkübe edin.
NOT: Bağırsak organoidleri, in vivo olarak bir dizi inflamatuar mediatöre yanıt verir. TNFa, çeşitli enflamatuar süreçlerde yer alan bir hücre sinyal proteinidir. - Kalan hücreleri% 5 Trigene içeren bir vakum tuzağına aspirasyon yoluyla atın.
Sonuçlar
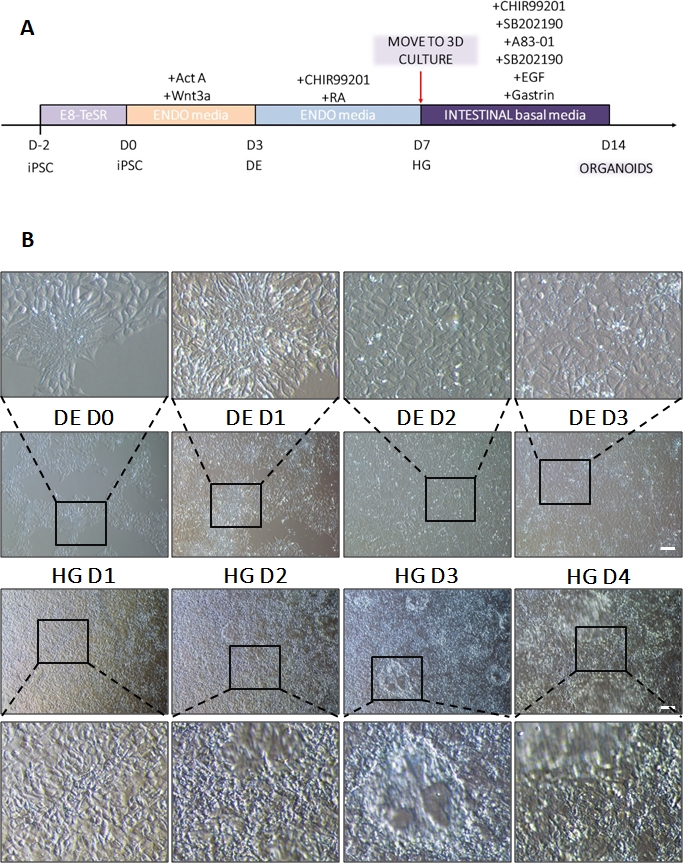
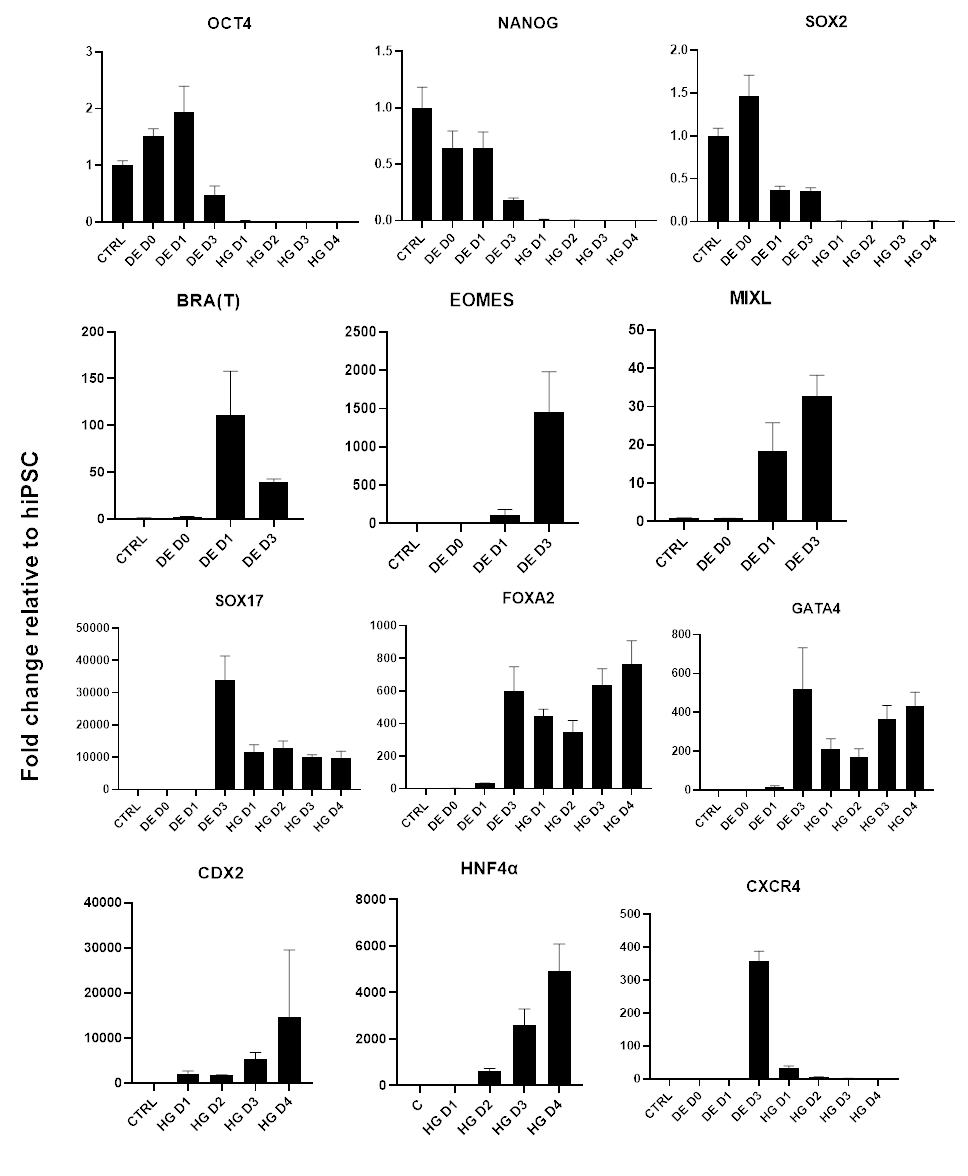
Farklılaşma protokolünün bir şeması Şekil 1A'da gösterilmiştir. Protokolün 1. gününde, hiPSC'ler kompakt olmalı ve toplam birleşme yaklaşık% 50-60 olan küçük koloniler oluşturmalıdır. DE'ye doğru farklılaşmanın indüksiyonunu takiben 24 saat sonra hücreler, tek bir hücre tabakası oluşturmak için kök hücre kolonilerinden uzaklaşmaya başlar. Bu, takip eden 3 gün boyunca devam eder ve DE farklılaşmasının D3'ünde tam bir tek tabaka oluşturmalıdır (Şekil 1). Gen ekspresyonu, D0'da yüksek oranda eksprese edilen ve DE farklılaşması sırasında hızla aşağı regüle edilen pluripotens belirteçleri (OCT4, NANOG, SOX2) ile farklılaşma boyunca izlenmelidir. DE farklılaşması sırasında T ekspresyonu D1'de zirve yapmalı, ardından D2'de EOMES ve MIXL gelmelidir. D2'de DE genleri (SOX17, FOXA2, GATA4, CXCR4) eksprese edilmeye başlamalı ve D3'te zirve yapmalıdır (Şekil 2 ve Şekil 3). Hücreler DE D3 tarafından tek tabakalı olmalıdır ve daha sonra arka bağırsak endodermine posteriorize edilebilir. Posteriorizasyon olayı sırasında 3D yapılar D2 kadar erken oluşmaya başlayacaktır. Ancak, bazen sadece D4'te görünmeye başlarlar veya hiç görünmezler; bu her zaman hücrelerin ilerleyip ilerlemeyeceğinin ve bağırsak organoidleri oluşturup oluşturmayacağının göstergesi değildir (Şekil 1). HG spesifikasyonu sırasında CDX2 ve HNF4a ekspresyonu indüklenmeli ve zamanla artmalıdır (Şekil 3).
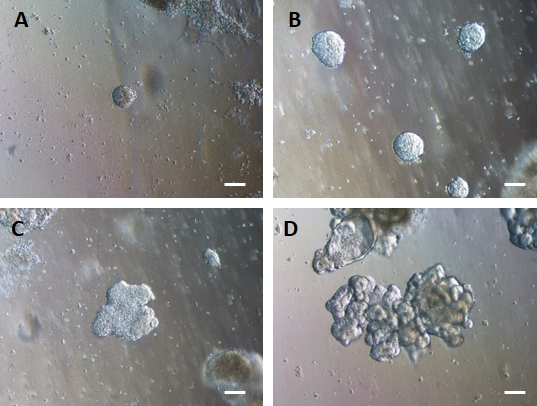
2D hücre tabakalarının hücre dışı matrise aktarılmasını takiben, ilk 24 saat boyunca 2D hücre kümeleri gözlemlenecektir. 48 saat sonra, hücre tabakaları, başlangıçta küçük olan (Şekil 4A), daha sonra 7-10 günlük kültür boyunca boyut ve karmaşıklık açısından kademeli olarak artan (Şekil 4B ve Şekil 4C) daha sıkıştırılmış 3D küresel yapılar halinde otomatik olarak organize olmaya başlamalıdır. Organoidler, lümen organoid/sferoidin merkezine bakacak şekilde belirgin epitel ile berrak bir organoid/sferoid morfolojisi elde edene kadar geçilmemelidir (Şekil 4D). Bu aşamada immünositokimya, villin ve CDX2 gibi bağırsak belirteçlerinin ekspresyonunu doğrulamak için kullanılabilir (Şekil 5). Tüm 2D hücre kümeleri organoidlere dönüşmeyecek ve hücre dışı matris içinde bazı kirletici ölü hücreler olacaktır. Bu ölü hücre tabakaları, hayatta kalan hücreler büyük organoidler oluşturana ve geçmeye hazır olana kadar göz ardı edilmelidir.
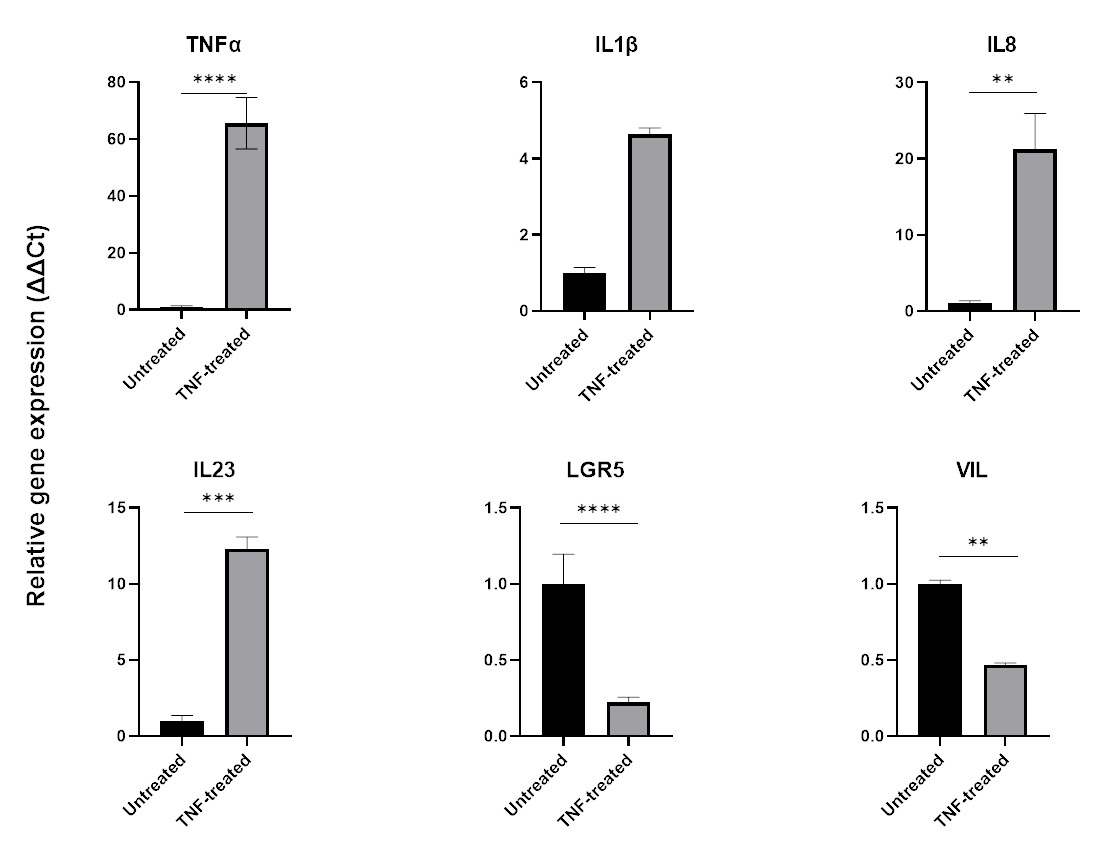
Enflamasyonu modellemek için, doku kültürü ortamına 24-48 saat boyunca TNFa eklenebilir. Proinflamatuar moleküllerle inkübasyonun ardından, organoidler, izolasyonları ve pasajları için kullanılan aynı teknik kullanılarak toplanır ve daha sonra QPCR veya western blotting gibi hücre tamponu uyumlu bir uygulama kullanılarak parçalanır. Daha kısa maruziyetler gerekiyorsa, organoidler önce hücre dışı matristen çıkarılmalı ve 1.5 mL'lik bir tüp kullanılarak süspansiyon halinde TNFa'ya maruz bırakılmalıdır. Bağırsak organoidlerinin 48 saat boyunca TNFa ile tedavisi tipik olarak proinflamatuar belirteçlerin (TNFa, IL1B, IL8, IL23) ekspresyonunu indüklerken, bağırsak epitel belirteçlerinin (LGR5, VIL) ekspresyonunu olumsuz etkiler (Şekil 6).
| Endoderm bazal media | 50 mL | |
| DEV/DAK 1640 DEV/DAK | 48,5 mL | |
| B27 Takviyesi | 1 mL | |
| %1 NEAA | 0,5 mL | |
Tablo 1: Endoderm farklılaşması için endodermal bazal ortamın bileşimi.
| Bağırsak Bazal Medya | 50 mL | |
| Gelişmiş DMEM/F12 | 46,5 mL | |
| HEPES Tamponu | 0,5 mL | |
| GlutaMAX | 0,5 mL | |
| Nikotinamid | 0,5 mL | |
| N2 Takviyesi | 0,5 mL | |
| B27 Takviyesi | 1.0 mL | |
| Kalem/Strep | 0,5 mL | |
Tablo 2: Bağırsak organoidlerinin kültürü için bağırsak bazal ortamının bileşimi

Şekil 1: HiPSC'nin kesin endoderm yoluyla arka bağırsak soyuna farklılaşması sırasında morfolojik değişiklikler.
(A) Bağırsak farklılaşma protokolüne şematik genel bakış. HiPSC'nin bu hücre hattı, yüksek çekirdek-sitoplazma oranına sahip küçük hücrelerin gevşek kolonilerini oluşturur. Farklılaşma ilerledikçe, hücreler epitelyalden mezenkimal fenotipe geçişle tutarlı değişikliklere uğrar ve DE D3 ile tek tip bir tek tabaka oluşturur. Uygun sinyaller iletildikten sonra, DE hücreleri uzar ve HD D3 olur olmaz ortaya çıkan 3D sferoidlerle daha yoğun bir şekilde paketlenmiş bir tek tabaka oluşturur, ancak bu kullanılan hücre hattına bağlıdır ve 3D kültüre (B) geçiş için bir gereklilik değildir. Ölçek çubuğu: 100 μm. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.
{kind=link}

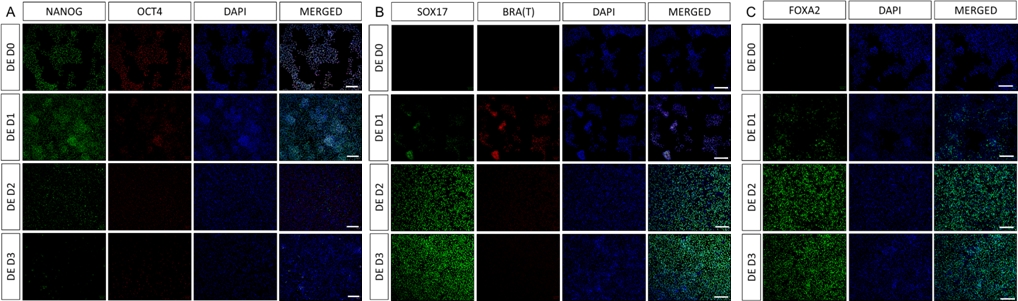
Şekil 2: HiPSC'lerin HG endodermine farklılaşması, endodermal genlerin ekspresyonunu indükler.
Kesin endodermden farklılaşan hiPSC'nin immün boyaması, protein düzeyinde TF'lerin ekspresyonunda değişiklikler gösterir. Pluripotens belirteçleri (NANOG ve OCT4) DE D3 (A) tarafından aşağı regüle edilir. Mezendoderm belirteci BRA (T) ekspresyonu protokolün (B) D1'inde bulunur ve DE'ye özgü TF'ler SOX17 ve FOXA2 D2'de (B ve C) görülür. Ölçek çubuğu: 200 mm. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.
{kind=link}

Şekil 3: Hindgut endoderm'e (HG) hiPSC farklılaşması sırasında qPCR ile gen ekspresyonu değişiklikleri.
Pluripotens ile ilişkili genler aşağı regüle edilir (OCT4, NANOG, SOX2), ardından mezendoderm genlerinin geçici ekspresyonu (T, EOMES, MIXL1) ve son olarak DE genlerinin ekspresyonu (SOX17, FOXA2, CXCR4) ve arka bağırsak genleri (CDX2, GATA4, HNF4a). Veriler ortalama ±SD olarak sunulmuştur. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.
{kind=link}

Şekil 4: HG endodermi, 3D hücre dışı matriks kültüründe 3D bağırsak organoidleri oluşturmak için kendi kendine birleşir.
HG endodermi, uygun bir 3D hücre dışı matris kültürüne aktarılır ve başlangıçta küçük katı hücre kümeleri (A) oluşturur. HG endoderm kümeleri 7-10 günlük kültür (B) boyunca genişler ve daha sonra asimetrik hale gelir ve daha karmaşık bir epitel (C) oluşturmaya başlar, sonunda berrak epitel morfolojisine ve organoidin merkezine (D) bakan bir lümen yüzeyine sahip organoidlere yol açar. Ölçek çubukları = 50 μm Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.
{kind=link}

Şekil 5: Yerleşik hiPSC türevi bağırsak organoidleri bağırsak belirteçlerini eksprese eder.
CDX2 ve Villin ekspresyonunu gösteren immünositokimya. Ölçek çubukları = 100 μm Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.
{kind=link}

Şekil 6: TNFa'nın sağlıklı bağırsak organoidlerinin inflamatuar profili ve bağırsak hücresi ekspresyonu üzerindeki etkisi.
TNFa (40 ng / mL) ile 48 saatlik tedaviyi takiben sağlıklı kolonik organoidlerin inflamatuar profili. Pro-inflamatuar belirteçlerin (TNFa, IL-8 ve IL-23) ekspresyonu, TNFa'ya maruz kaldıktan sonra artarken, aynı zamanda bağırsak epitel belirteçlerinin (LGR5, VIL) ekspresyonu aşağı regüle edilir. İstatistiksel analizler iki taraflı öğrenci t-testi ile yapıldı. Veriler her grubun ortalama ± SD olarak ifade edilmiştir. *P < .01; **P < .001; P < .0001. (n=3). Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.
{kind=link}
Tartışmalar
Burada, insan pluripotent hücrelerinin insan bağırsak organoidlerine farklılaşması için bir protokol açıklıyoruz. Enflamasyonu incelemek için kullanımlarını gösteriyoruz; bununla birlikte, bu çeşitli bağlamlara uygulanabilir ve herhangi bir genetik arka plan üzerinde CRISPR / Cas9 gen düzenleme yaklaşımları14 ile birleştirilebilir. Farklılaştıktan sonra, kesin endoderm, arka bağırsak endodermi ve daha sonra bağırsak epitelinin doğal gelişimsel farklılaşma dizisini takiben, ortaya çıkan organoidler 12 aydan fazla bir süre boyunca sürekli olarak kültürlenebilir ve geçirilebilir.
Bu protokolün kritik bir yönü, endoderm farklılaşmasından önce farklılaşmamış kök hücrelerin ilk kaplama yoğunluğudur. Bu yeterince optimize edilmezse, hücreler muhtemelen ilk DE farklılaşma adımı sırasında ölecek (hücreler çok seyrekse) veya DE farklılaşmasının verimliliğini azaltacaktır (hücreler çok yoğunsa). Kullanılan hücre hattı için doğru başlangıç yoğunluğunun optimize edilmesi gerekir ve doğru yoğunluk DE D3'ün sonunda bir tek katman oluşturmalıdır. DE spesifikasyonunun etkinliğini belirlemek için akış sitometrisi kullanılmalıdır ve tipik olarak SOX17 ve/veya CXCR4 için pozitif hücrelerin %>80'ini görürüz. SOX17 pozitif hücrelerin sayısı %60'tan az olduğunda, HG modellemesinin verimliliği etkilenir, bu da hücre dışı matrise aktarıldığında daha az organoid oluşmasına neden olur. Bu, sonuçta ortaya çıkan organoid kültürlerin başarısız olmasına neden olacaktır. DE'den HG'ye modellemenin başarılı olup olmadığını belirlemek için, akış sitometrisi ile CDX2 pozitif hücrelerin sayısını değerlendiririz ve tipik olarak% >80 pozitif hücre görmeyi bekleriz. Yine, CDX2 pozitif hücrelerin sayısı% 50'nin altına düşerse, bu, 3D hücre dışı matris kültürüne aktarıldığında üretilen bağırsak organoidlerinin sayısı üzerinde olumsuz bir etkiye sahip olacaktır.
2D tek katmanların 3D kültüre aktarılmasını takiben, küçük kompakt küreler transferden 24-48 saat sonra görünmelidir. Kullanılan hücre hattının farklılaşma verimliliğine bağlı olarak büyük ölü hücre tabakaları görünebilir. Bu kalıntıları gidermek için kültürleri hemen geçmek yerine, organoidlerin daha karmaşık, katlanmış yapılarını tam olarak oluşturmalarına ve geliştirmelerine izin veriyoruz. İlk geçişi denemeden önce 7-10 gün beklemek, birçok yeni bağırsak organoidi oluşturmak için yeterli bölünen hücrelerin mevcut olmasını sağlar. Kültürde hala mevcut olan herhangi bir kalıntı, ayrışmış organoid / enkaz karışımlarını organoidleri peletlemek için yeterli hızda bir tüp içinde yavaşça döndürerek, ancak ortamda yüzen hücre tabakasını bırakarak, geçiş işlemi sırasında kolayca çıkarılabilir. Medya ve hücresel kalıntılar daha sonra aspire edilebilir, böylece sadece organoidlerin peleti kalır.
Bu yaklaşımın sınırlaması, hiPSC'den türetilen hücre tiplerinin gen ekspresyonu ve fonksiyonel profiller açısından genellikle tam olarak olgunlaşmamış olmasıdır. HiPSC'den türetilen bağırsak dokusunun spesifik uygulamalar için uygun olup olmadığını belirlemek için organoidler, enterositler (VIL), enteroendokrin hücreler (nörog3), goblet hücreleri (MUC2), geçici amplifiye edici hücreler (CD133), paneth hücreleri (FZD5) ve LGR5+ kök hücreler (LGR5) dahil olmak üzere farklı hücre tipleri için karakterize edilmelidir.
Genel olarak, bu protokolün diğer birçok organoid farklılaşma protokolüne göre en büyük avantajı, bu kültür platformunun, birkaç rekombinant proteinin ve şartlandırılmış ortam preparatlarının küçük moleküllerle değiştirilmesi nedeniyle çok uygun maliyetli olmasıdır15,16. HG'ye farklılaşma çok basit ve hızlıdır ve aynı sonuçlarla hem insan embriyonik hem de indüklenmiş pluripotent kök hücrelere uygulanabilir. Titizlikle takip edildiğinde ve kullanılan hücre hatları için optimize edildiğinde, daha sonra inflamasyon, konakçı patojen etkileşimleri dahil olmak üzere çeşitli bağlamlarda bağırsak epitelini incelemek için uygulanabilen, kontamine mezenkimal hücrelerden arınmış nispeten basit bir model platform sağlar7. İntestinal fibroz modellemesi, profibrotik uyaranlar sağlanarak ve daha sonra kollajen, laminin ve fibronektin gibi hücre dışı matriks proteinlerinin ekspresyonunun QPCR, western blot ve ELISA ile ekspresyonunun değerlendirilmesiyle araştırılabilir. Farklılaşmadan önce farklılaşmamış kök hücre dizilerinde CRISPR/Cas9 gen düzenleme tekniklerinin kullanılması, hastalığa özgü organoidler ve daha karmaşık hastalık modelleri oluşturmak için kullanılabilecek gen nakavt veya protein aşırı ekspresyon organoidlerinin oluşturulmasına izin verir 14,17,18.
Açıklamalar
Yazarların açıklayacak hiçbir şeyi yok.
Teşekkürler
NH, MRC (MR/S009930/1) ve Wellcome Trust (204267/Z/16/Z) tarafından finanse edilmektedir, PD, MRC PHD DTP tarafından finanse edilmektedir, KLF, BBSRC iCASE tarafından finanse edilmektedir.
Malzemeler
| Name | Company | Catalog Number | Comments |
| A83-01 | Tocris | 2939 | |
| Activin A | R&D | 338-AC | |
| Advanced DMEM/F12 (1X) | Life Technologies | 12654-010 | |
| B27 supplement | Gibco | 17504044 | |
| CHIR99021 | Sigma | SML1046-5MG | |
| Epidermal Growth Factor | R&D Systems | 236-EG-01M | |
| Gastrin | Sigma Aldrich | G9145 | |
| GlutaMAX (100X) | Life Technologies | 15630-056 | |
| Growth Factor reduced Matrigel | BD | ||
| HEPES Buffer solution (1M) | Life Technologies | 15630-080 | |
| N2 Supplement (100X) | Gibco | 17502-048 | |
| N-acetyl-cysteine | Sigma Aldrich | A7250 | |
| Nicotinamide | Sigma Aldrich | N0636 | |
| Noggin | R&D Systems | 6057-NG | |
| Non-essential amino acids | Gibco | 11140-050 | |
| Paraformaldehyde | VWR | 9713.5 | |
| Penicillin/Streptomycin | Gibco | 15140122 | |
| Phosphate Buffered Saline | Gibco | 14190-094 | |
| Retinoic Acid | Sigma | 302-79-4 | |
| ROCK inhibitor | Tocris | 1254/1 | |
| ROCK inhibitor Y-27632 | Tocris | 1254 | |
| RPMI | Sigma | R8758-500ml | |
| R-Spondin-1 | Peprotech | 120-38 | |
| SB202190 | Tocris | 1264 | |
| TrypLe Express | Gibco | 12604-021 | |
| Wnt 3a | R&D | 5036-WN |
Referanslar
- Thomson, J. A., et al. Embryonic stem cell lines derived from human blastocysts. Science. 282, 1145-1147 (1998).
- Takahashi, K., Yamanaka, S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell. 126, 663-676 (2006).
- Takahashi, K., et al. Induction of pluripotent stem cells from adult human fibroblasts by defined factors. Cell. 131, 861-872 (2007).
- Rashid, S. T., et al. Modeling inherited metabolic disorders of the liver using human induced pluripotent stem cells. Journal of Clinical Investigation. 120, 3127-3136 (2010).
- Vallier, L., Alexander, M., Pedersen, R. A. Activin/Nodal and FGF pathways cooperate to maintain pluripotency of human embryonic stem cells. Journal of Cell Science. 118, 4495-4509 (2005).
- Jaremko, K. L., Marikawa, Y. Regulation of developmental competence and commitment towards the definitive endoderm lineage in human embryonic stem cells. Stem Cell Research. 10, 489-502 (2013).
- Forbester, J. L., et al. Interaction of Salmonella enterica Serovar Typhimurium with Intestinal Organoids Derived from Human Induced Pluripotent Stem Cells. Infection and Immunity. 83, 2926-2934 (2015).
- Hannan, N. R., et al. Generation of multipotent foregut stem cells from human pluripotent stem cells. Stem Cell Reports. 1, 293-306 (2013).
- Sato, T., et al. Single Lgr5 stem cells build crypt-villus structures in vitro without a mesenchymal niche. Nature. 459, 262-265 (2009).
- de Souza, H. S., Fiocchi, C. Immunopathogenesis of IBD: current state of the art. Nature Reviews Gastroenterology & Hepatology. 13, 13-27 (2016).
- Tripathi, K., Feuerstein, J. D. New developments in ulcerative colitis: latest evidence on management, treatment, and maintenance. Drugs in Context. 8, 212572 (2019).
- Ganesh, K., et al. A rectal cancer organoid platform to study individual responses to chemoradiation. Nature Medicine. 25, 1607-1614 (2019).
- Fair, K. L., Colquhoun, J., Hannan, N. R. F. Intestinal organoids for modelling intestinal development and disease. Philosophical Transactions of the Royal Society B: Biological Sciences. 373, (2018).
- Cuevas-Ocaña, S., Yang, J. Y., Aushev, M., Schlossmacher, G., Bear, C. E., Hannan, N. R. F., Perkins, N. D., Rossant, J., Wong, A. P., Gray, M. A. A Cell- Based Optimised Approach for Rapid and Efficient Gene Editing of Human Pluripotent Stem Cells. Int. J. Mol. Sci. 24, 10266 (2023).
- Spence, J. R., et al. Directed differentiation of human pluripotent stem cells into intestinal tissue in vitro. Nature. 470, 105-109 (2011).
- Sato, T., et al. Long-term Expansion of Epithelial Organoids From Human Colon, Adenoma, Adenocarcinoma, and Barrett's Epithelium. Gastroenterology. 141, 1762-1772 (2011).
- Giacalone, J. C., et al. CRISPR-Cas9-Based Genome Editing of Human Induced Pluripotent Stem Cells. Current Protocols in Stem Cell Biology. 44, 1-22 (2018).
- Bruntraeger, M., Byrne, M., Long, K., Bassett, A. R., Luo, Y. . CRISPR Gene Editing: Methods and Protocols. , 153-183 (2019).
Yeniden Basımlar ve İzinler
Bu JoVE makalesinin metnini veya resimlerini yeniden kullanma izni talebi
Izin talebiThis article has been published
Video Coming Soon
JoVE Hakkında
Telif Hakkı © 2020 MyJove Corporation. Tüm hakları saklıdır