
Method Article
Génération d’organoïdes intestinaux dérivés de hiPSC pour des applications de modélisation du développement et des maladies
Dans cet article
Résumé
Ce protocole permet de différencier les cellules pluripotentes humaines en organoïdes intestinaux. Le protocole imite le développement humain normal en différenciant les cellules en une population d’endoderme définitif, d’endoderme de l’intestin postérieur, puis d’épithélium intestinal. Cela rend le protocole adapté à la fois à l’étude du développement intestinal et aux applications de modélisation des maladies.
Résumé
Les organoïdes intestinaux dérivés de hiPSC sont des structures épithéliales qui s’auto-assemblent à partir de cellules différenciées en structures 3D complexes, représentatives de l’épithélium intestinal humain, dans lesquelles ils présentent des structures ressemblant à des cryptes/villosités. Ici, nous décrivons la génération d’organoïdes intestinaux dérivés des hiPSC par la différenciation progressive des hiPSC en endoderme définitif, qui est ensuite postériorisé pour former un épithélium de l’intestin postérieur avant d’être transféré dans des conditions de culture 3D. L’environnement de culture 3D est constitué d’une matrice extracellulaire (MEC) (p. ex., Matrigel ou autre MEC compatible) complétée par du SB202190, de l’A83-01, de la gastrine, de la noggine, de l’EGF, de la R-spondine-1 et du CHIR99021. Les organoïdes subissent un passage tous les 7 jours, où ils sont mécaniquement perturbés avant d’être transférés dans une matrice extracellulaire fraîche et de se développer. La QPCR et l’immunocytochimie confirment que les organoïdes intestinaux dérivés de hiPSC contiennent des types de cellules épithéliales intestinales matures, notamment des cellules caliciformes, des cellules de Paneth et des entérocytes. De plus, les organoïdes montrent des signes de polarisation par l’expression de la ville, localisée sur la surface apicale des cellules épithéliales.
Les organoïdes qui en résultent peuvent être utilisés pour modéliser le développement intestinal humain ainsi que de nombreuses maladies intestinales humaines, y compris les maladies inflammatoires de l’intestin. Pour modéliser l’inflammation intestinale, les organoïdes peuvent être exposés à des médiateurs inflammatoires tels que le TNF-α, le TGF-β et le LPS bactérien. Les organoïdes exposés à des cytokines pro-inflammatoires présentent un phénotype inflammatoire et fibreux en réponse. L’appariement des CSP saines et hiPS dérivées de patients atteints de MII peut être utile pour comprendre les mécanismes à l’origine des MII. Cela pourrait révéler de nouvelles cibles thérapeutiques et de nouveaux biomarqueurs pour aider au diagnostic précoce de la maladie.
Introduction
Les propriétés des cellules souches pluripotentes (CSP), telles que l’auto-renouvellement et la capacité de se différencier en n’importe quel type de cellule du corps humain, en font des outils précieux dans l’étude du développement, de la pathologie des maladies et des tests de médicaments1. Les cellules souches pluripotentes induites humaines (hiPSC) sont particulièrement utiles pour les études de modélisation de la maladie car elles peuvent être dérivées de patients, capturant directement le génome responsable du phénotype 2,3 de la maladie. Ces hiPSC peuvent être différenciées en fonction du type de cellule affecté par l’anomalie génétique, ce qui permet un examen minutieux du mécanisme moléculaire de la maladie4.
Les protocoles de différenciation pour les CSP humaines visent à diriger la différenciation des cellules à travers les principaux stades de développement par l’activation ou l’inhibition de voies de signalisation spécifiques régissant l’engagement et la spécification de la lignée. Le maintien de l’hiPSC dans un état pluripotent nécessite des niveaux modérés de signalisation de l’activine A (Act-A), tandis qu’une dose élevée d’Act-A pendant 3 jours engage l’hiPSC dans un destin définitif de l’endoderme (DE) 5,6. Les voies Act-A et Wnt dirigent l’identité antéro-postérieure de l’ED. La signalisation par Act-A induit des marqueurs de l’intestin antérieur (FG) tels que HHEX, HNF4α et GATA4, tout en bloquant l’expression des gènes de l’intestin postérieur (HG) tels que CDX2. La signalisation Wnt induit la postériorisation de l’ED, qui adopte alors un profil d’expression du gène HG 7,8. Une fois l’identité des cellules HG établie, la différenciation peut être déplacée de la 2D à la 3D et dirigée vers la formation d’organoïdes intestinaux.
Les organoïdes intestinaux sont généralement cultivés dansun système de culture basé sur la matrice extracellulaire 3D (par exemple, Matrigel ou d’autres ECM compatibles) 9, composé de laminine, de collagène IV et d’entactine et enrichi de facteurs de croissance tels que l’EGF, FGF, PDGF et IGF-1 pour contribuer à la survie et à la prolifération. Les organoïdes sont cultivés dans un milieu défini contenant de la gastrine, de la noggine et du CHIR pour stimuler et soutenir la croissance et la prolifération des cellules souches intestinales pendant la culture à long terme.
Une fois que les cellules épithéliales intestinales sont intégrées dans la matrice extracellulaire, les cryptes intestinales commencent à se former et à se développer, formant finalement des sphéroïdes. Ceux-ci mûrissent en structures organoïdes qui imitent le fonctionnement physiologique de l’épithélium intestinal. Les organoïdes peuvent généralement être cultivés pendant plus d’un an sans perte significative des profils fonctionnels et d’expression génique. Le passage est nécessaire sur une base hebdomadaire en utilisant la digestion enzymatique des organoïdes en fragments plus petits qui s’auto-assemblent ensuite en organoïdes complets.
Les lignées organoïdes établies peuvent être utilisées comme modèle fiable de nombreux troubles liés à l’intestin, notamment la maladie de Crohn, la colite ulcéreuse et les cancers colorectaux 10,11,12,13. Il s’agit d’un modèle privilégié pour les cellules animales car elles expriment des gènes humains liés à ces troubles et répondent à des stimuli externes plus proches de ceux qui se produisent dans les tissus humains in vivo.
Protocole
REMARQUE : Tous les travaux de culture tissulaire détaillés ci-dessous doivent être effectués dans une hotte à flux laminaire de classe II.
1. Différenciation des cellules souches pluripotentes induites humaines (hiPSCs) vers l’endoderme de l’intestin postérieur
- Pour enrober un flacon de culture cellulaire de matrice extracellulaire, calculez d’abord la quantité de matrice nécessaire. Voici un exemple d’une plaque à 12 puits recouverte de Matrigel. Calculez la quantité requise de matrice extracellulaire pour recouvrir une plaque à 12 puits. Utilisez la formule suivante :

REMARQUE : Enrober les boîtes de culture cellulaire avec la matrice extracellulaire de votre choix. Si vous utilisez Matrigel, utiliser une concentration de 0,035 mg/cm2 24 h avant le semis pour la différenciation. Vérifiez la concentration du lot de matrice extracellulaire sur la fiche produit et préparez des aliquotes plus petites avant les expériences selon les instructions du fabricant. - Diluer la quantité requise de matrice extracellulaire dans un milieu DMEM froid à l’aide d’une pipette p1000.
- Bien mélanger à l’aide d’une stripette de 5 ml.
- Ajouter 500 μL de matrice diluée à chaque plaque de puits. Agiter doucement la plaque pour répartir uniformément la matrice extracellulaire diluée et incuber pendant au moins 12 h à 37 °C.
- Avant d’ensemencer les cellules dans la plaque, laver chaque puits avec 500 μL de PBS pour éliminer l’excès de matrice extracellulaire. Cela empêchera le détachement des cellules pendant la différenciation.
- Laissez le dernier lavage dans les puits jusqu’au moment de semer pour éviter le dessèchement de la matrice extracellulaire.
- Pour amorcer la différenciation des hiPSCs vers l’endoderme définitif (DE) ou l’intestin postérieur (HG), prélever une fiole de hiPSCs cultivée dans un milieu d’entretien de votre choix (ex. : milieu essentiel 8) et aspirer le milieu. (Ici, nous ensemenceons les cellules à 25 000 cellules/cm2).
REMARQUE : La densité d’ensemencement est cruciale pour une différenciation réussie et doit être optimisée pour chaque lignée cellulaire hiPSC individuelle. - Laver les cellules dans le flacon avec 5 mL de PBS. Aspirez le PBS.
- Ajouter 2,5 mL de solution de dissociation cellulaire (p. ex., TrypLE) et laisser reposer 4 min. Aspirez la solution de dissociation cellulaire et tapotez doucement le flacon pour détacher les cellules.
- Lavez le flacon avec 5 mL de média DMEM chauffé à 37 °C et collectez toutes les cellules dans un tube de 15 mL.
- Prélever 10 μL de solution cellulaire remise en suspension et mesurer la densité cellulaire à l’aide d’un hémocytomètre.
- Prélevez suffisamment de suspension cellulaire pour recueillir 1,05 x 106 cellules et placez-la dans un tube de 15 ml.
- Essorer à 160 x g pendant 3 min. Pendant que les cellules sont centrifugées, aspirez le PBS de la plaque à 12 puits recouverte d’une matrice extracellulaire.
- Après centrifugation, aspirer le surnageant et remettre en suspension la pastille cellulaire dans 12 mL de milieu d’entretien avec inhibiteur ROCK (10 μM).
- À l’aide d’une pipette p1000, ajouter 1 mL de suspension cellulaire dans chaque puits de la plaque à 12 puits. Bien remettre les cellules en suspension pour assurer une répartition égale entre les puits.
- Secouez doucement la plaque pour répartir les cellules à l’intérieur des puits de la plaque, mais évitez de faire tourbillonner le milieu car cela concentrera les cellules au milieu des puits.
- Placer dans un incubateur à 37 °C, 5 % de CO2 .
- Remplacer le milieu par un milieu d’entretien (p. ex., milieu essentiel 8) seulement (pas d’inhibiteur de ROCK) 24 h après l’ensemencement.
- Pour commencer la différenciation en DE, préparer le milieu basal de l’endoderme conformément au tableau 1.
REMARQUE : Au début de la différenciation, les cellules doivent être à une confluence comprise entre 60 et 80%. - Préparer 12 mL de milieu DE en ajoutant de l’activine A (100 ng/mL) et du Wnt3 (50 ng/mL) au milieu basal de l’endoderme.
- Réchauffez le support à 37 °C.
- Aspirer le milieu de chaque puits de la plaque ou du flacon de culture tissulaire.
- Commencez la différenciation de l’ED en ajoutant 1 mL de média DE dans chaque puits de la plaque à 12 puits.
- À 24 h après le début de la différenciation DE (D1 DE), préparer un nouveau milieu DE et effectuer un changement de milieu.
- Répétez l’étape 1.24 à 48 h (D2 DE). S’il y a beaucoup de mort cellulaire à ce stade, laver tous les puits avec 500 μL de PBS avant de changer de milieu.
REMARQUE : Pour confirmer la réussite de la différenciation de l’endoderme, effectuer une cytométrie en flux pour évaluer l’expression de SOX17. Nous nous attendons normalement à ce que >80 % des cellules soient positives pour le SOX17 à J3 DE. Si l’expression de SOX17 est sous-optimale, la densité d’ensemencement cellulaire doit être optimisée, ainsi que les concentrations d’Act-A. - Commencer la différenciation HG à ce stade, c’est-à-dire 72 h après le début de la différenciation DE. Préparer 12 mL de milieu HG en ajoutant CHIR99021 (3 μM) et RA (1 μM) au milieu basal de l’endoderme (tableau 1).
REMARQUE : À D3 DE, la cellule doit avoir formé une monocouche uniforme. Une différenciation optimale de l’EDD est cruciale pour les étapes ultérieures de la différenciation. - Aspirer le milieu DE.
- Commencez la différenciation HG en ajoutant 1 mL de milieu HG à chaque puits de la plaque à 12 puits.
- Continuez la différenciation pendant 4 jours avec des changements de supports quotidiens.
REMARQUE : Pour déterminer le succès de la postériorisation de l’ED, effectuer une cytométrie en flux pour évaluer l’expression de CDX2. À D4 de la différenciation HG, nous nous attendons normalement à ce que >80% des cellules soient CDX2 positives. Si la postériorisation n’est pas optimale, la concentration de CHIR99021 doit être optimisée. - Pour prélever un échantillon pour l’extraction de l’ARN, aspirer le milieu de différenciation et laver le puits avec 500 ul de PBS.
- Aspirez le PBS et ajoutez un volume approprié de tampon de lyse cellulaire à partir d’un kit d’extraction d’ARN.
- À l’aide d’une pipette p1000, grattez le fond du puits pour assurer la lyse de toutes les cellules.
- Aspirez le lysat cellulaire et placez-le dans un tube propre.
- Procéder à l’extraction de l’ARN ou congeler le lysat à -20 °C jusqu’à ce qu’il soit prêt pour l’extraction de l’ARN.
- Pour l’immunomarquage, aspirer les milieux de différenciation et laver les puits avec 500 μL de PBS.
- Aspirer le PBS et ajouter 500 μL de PFA à 4 %.
REMARQUE : Le PFA est toxique. Utilisez l’EPI approprié et suivez les procédures de laboratoire locales pour l’élimination des PFA. - Incuber à 4 °C pendant 20 min.
- Retirez les PFA et lavez les puits avec 500 μL de PBS trois fois.
- Laissez le dernier lavage PBS sur les cellules jusqu’à ce que vous soyez prêt à effectuer l’immunomarquage.
2. Passage d’organoïdes intestinaux
- Préparer le milieu basal intestinal conformément au tableau 2.
- Pour passer d’une culture cellulaire 2D à une culture cellulaire 3D, utilisez une plaque à 6 puits pour générer des cellules DE formant des hiPSCS. Détacher la monocouche de cellules de la plaque à 6 cellules à l’aide d’une stripette de 5 mL.
- Recueillir les cellules dans un tube à centrifuger de 15 mL, avant de centrifuger à 400 x g pendant 1 min pour produire une pastille de cellule.
- Remettre en suspension la pastille cellulaire dans un milieu de croissance intestinal contenant des facteurs de croissance : SB202190 (10 μM), A83-01 (500 nM), Gastrin (10 nM), Noggin (100 ng/μL), EGF (500 ng/μL), R-Spondin1 (100 ng/mL), CHIR99021 (6 μM), inhibiteur de ROCK (10 μM).
- Ajouter le volume approprié de matrice extracellulaire en fonction du nombre de puits plaqués. Cela peut être calculé à l’aide des équations ci-dessous.
[1] Volume total de la matrice/du milieu extracellulaire (μL) = 30 μL * nombre de puits de la plaque à 48 puits
[2] Volume requis de matrice extracellulaire (μL) = Réponse de [1] * 2/3
[3] Volume requis de média (μL) = Réponse de [1] * 1/3 - Ajouter 30 μL de suspension cellulaire au centre de chaque puits d’une plaque à 48 puits.
- Incuber la plaque dans un incubateur à 37 °C pendant au moins 5 min pour permettre à la matrice extracellulaire de se fixer.
- Une fois que les dômes de la matrice extracellulaire ont pris, ajoutez 300 μL de milieu de croissance intestinal contenant tous les facteurs de croissance dans chaque puits.
- Remettez la plaque dans un incubateur à 37 °C avec 5 % de CO2.
REMARQUE : Si après 48 h, aucun organoïde ne peut être observé et/ou s’il y a une mort cellulaire importante, il peut être nécessaire d’optimiser les concentrations de ROCKi et de NOGGIN. - Pour faire passer les organoïdes, observez les cellules au microscope optique afin d’évaluer la densité et la taille des organoïdes et de déterminer s’ils doivent être transportés.
- Remplacez les milieux de culture cellulaire par du PBS glacé.
REMARQUE : Les cultures organoïdes devront être divisées environ tous les 5 à 7 jours. Le passage de cultures d’organoïdes intestinaux est nécessaire une fois qu’il y a une accumulation de débris cellulaires évidente dans la lumière organoïde et un jaunissement du milieu de croissance intestinal environnant. - Détacher mécaniquement les organoïdes et la sphère de la matrice extracellulaire de la plaque à l’aide d’un mouvement de raclage à l’aide d’une bande de 5 mL.
- Recueillir les organoïdes de chaque puits dans un tube à centrifuger de 15 ml.
- Centrifuger la suspension d’organoïdes à 400 x g pendant 1 min pour granuler les organoïdes. Aspirer le surnageant jusqu’à ce qu’il atteigne le sommet de la pastille cellulaire, en faisant attention lorsqu’il a atteint la couche visible de la matrice extracellulaire.
- Remettre la pastille en suspension dans 15 mL de PBS glacé.
REMARQUE : Cette étape sert à laver toute matrice extracellulaire restante qui n’a pas été retirée lors de l’essorage initial. - Centrifuger à 400 x g pendant 1 min. Aspirer le milieu jusqu’à la pastille contenant les organoïdes.
- Remettre en suspension dans 1 mL de PBS glacé. À l’aide d’une pipette p200, perturbez manuellement les organoïdes intacts en pipetant plusieurs fois de haut en bas.
REMARQUE : Observez la taille des organoïdes à l’aide d’un microscope optique pour déterminer s’ils doivent être dissociés davantage. - Ajouter 9 mL de milieu sans facteurs de croissance.
- Centrifuger à 400 x g pendant 1 min. Aspirer le surnageant jusqu’à la pastille d’organoïdes.
- Calculez la quantité requise de matrice extracellulaire et de milieux à l’aide des équations ci-dessous :
[1] Volume total de la matrice/du milieu extracellulaire (μL) = 30 μL * nombre de puits de la plaque à 48 puits
[2] Volume requis de matrice extracellulaire (μL) = Réponse de [1] * 2/3
[3] Volume requis de média (μL) = Réponse de [1] * 1/3 - Remettre en suspension la pastille organoïde dans le volume calculé du milieu intestinal avec des facteurs de croissance (y compris l’inhibiteur de Noggin & ROCK).
- Ajoutez le volume requis de matrice extracellulaire dans cette suspension cellulaire et remettez-la en suspension pour assurer une distribution uniforme des organoïdes.
- Pipeter 30 μL de cette suspension au centre de chaque puits d’une plaque à 48 puits (de préférence préchauffée dans un incubateur à 37 °C).
- Remettez la plaque dans un incubateur à 37 °C pendant 5 min jusqu’à ce que la matrice extracellulaire ait pris.
- Préparer le milieu intestinal avec des facteurs de croissance (+ inhibiteur de ROCK) (environ 17 mL par plaque de 48 puits).
- Ajouter 300 μL dans chaque puits.
- Incuber à 37 °C à 5 % de CO2.
- Après le passage, aspirez le milieu pour les organoïdes intestinaux et remplacez-le par un milieu intestinal frais avec des facteurs de croissance (sans inhibiteur de Noggin & ROCK) tous les 2 à 4 jours.
REMARQUE : Si après 48 heures, aucun organoïde ne peut être observé et/ou s’il y a une mort cellulaire importante, il peut être nécessaire d’optimiser les concentrations de ROCKi et de NOGGIN.
REMARQUE : Les organoïdes intestinaux répondent à une gamme de médiateurs inflammatoires in vivo. Le TNFα est une protéine de signalisation cellulaire impliquée dans plusieurs processus inflammatoires. - Pour déclencher une réponse inflammatoire dans les organoïdes intestinaux, préparer le TNFα à une concentration de 40 ng/mL dans un milieu basal uniquement.
- Aspirer le milieu à partir d’une plaque à 48 puits.
- Ajouter 300 μL de milieu basal préparé contenant 40 ng/mL de TNFα.
- Incuber la plaque à 37 °C pendant 48 heures pour reproduire un environnement pro-inflammatoire.
REMARQUE : Les organoïdes intestinaux répondent à une gamme de médiateurs inflammatoires in vivo. Le TNFα est une protéine de signalisation cellulaire impliquée dans plusieurs processus inflammatoires. - Éliminer les cellules restantes par aspiration dans un piège à vide contenant 5 % de trigène.
Résultats
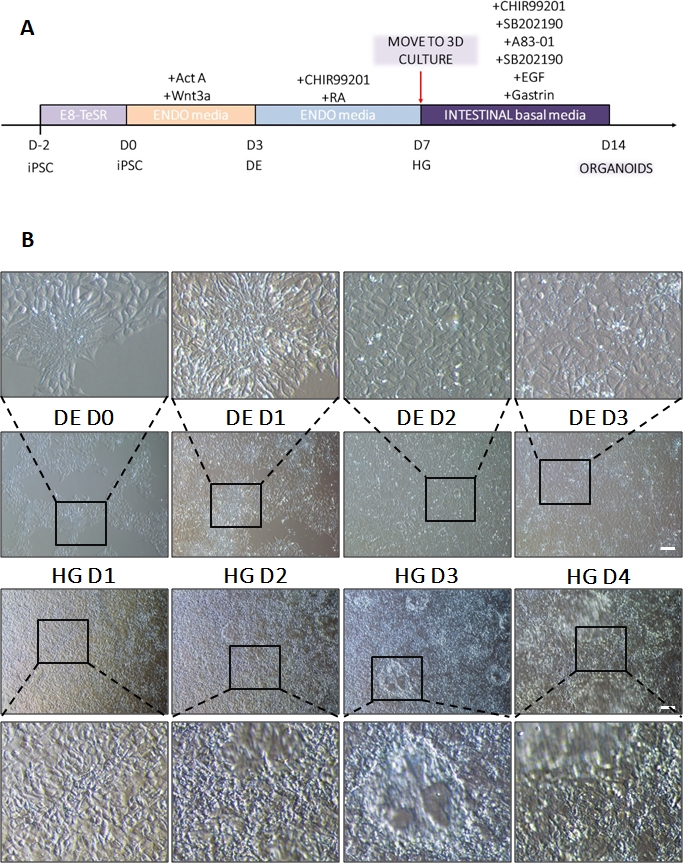
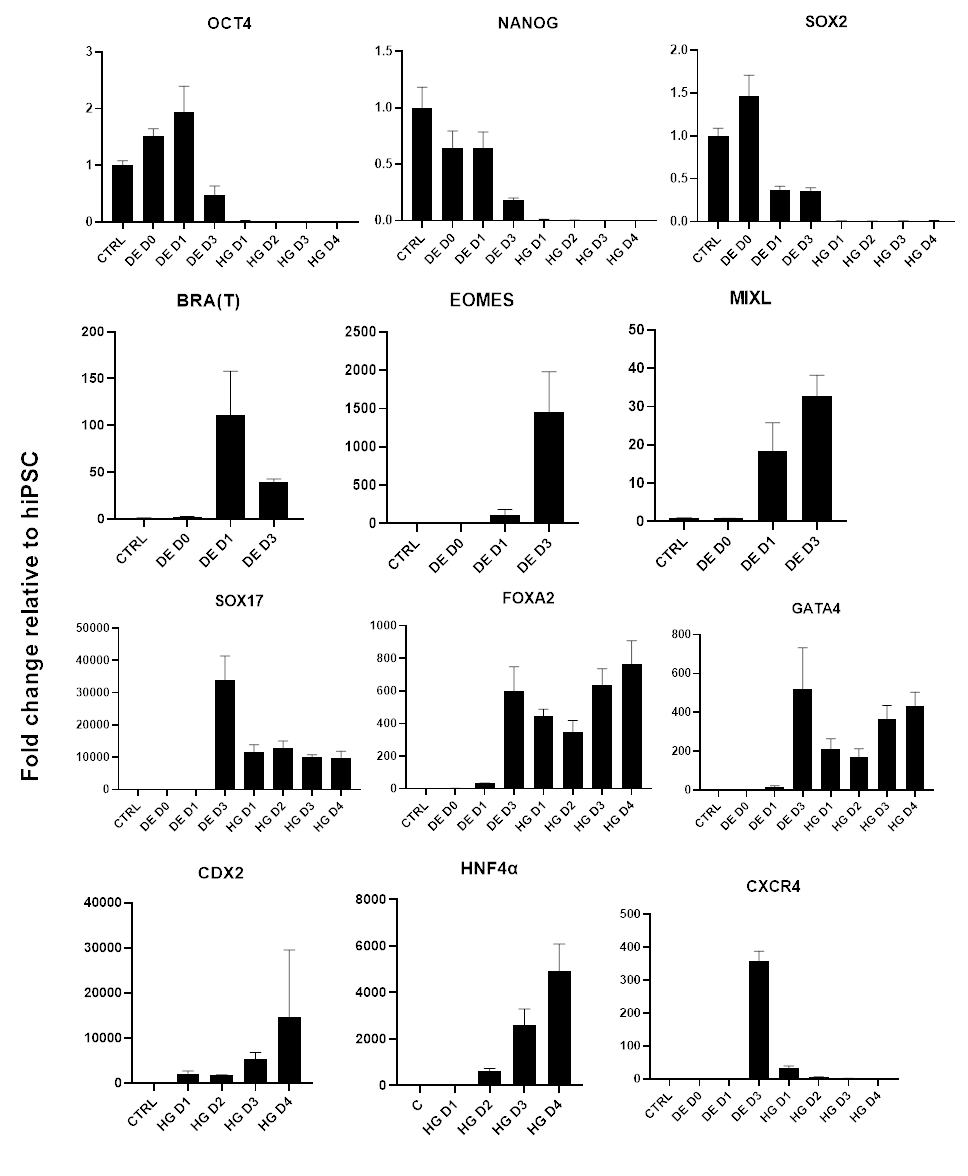
Un schéma du protocole de différenciation est présenté à la figure 1A. Au jour 1 du protocole, les hiPSC doivent être compacts et former de petites colonies avec une confluence totale d’environ 50 à 60 %. 24 heures après l’induction de la différenciation vers l’ED, les cellules commencent à migrer loin des colonies de cellules souches pour former une monocouche de cellules. Cela se poursuit pendant les 3 jours suivants et devrait former une monocouche complète sur D3 de différenciation DE (Figure 1). L’expression des gènes doit être surveillée au cours de la différenciation avec des marqueurs de pluripotence (OCT4, NANOG, SOX2) fortement exprimés sur D0 et rapidement régulés à la baisse lors de la différenciation DE. Au cours de la différenciation DE, l’expression de T devrait culminer à D1, suivie d’EOMES et de MIXL à D2. Sur D2, les gènes DE (SOX17, FOXA2, GATA4, CXCR4) devraient commencer à s’exprimer et culminer à D3 (Figure 2 et Figure 3). Les cellules doivent être monocouches par DE D3 et peuvent ensuite être postériorisées dans l’endoderme de l’intestin postérieur. Au cours de l’événement de postériorisation, des structures 3D commenceront à se former dès J2. Cependant, il arrive qu’ils ne commencent à apparaître qu’à J4 ou pas du tout ; cela n’indique pas toujours si les cellules vont ou non se transformer en organoïdes intestinaux (figure 1). Au cours de la spécification HG, l’expression de CDX2 et HNF4a doit être induite et augmenter avec le temps (Figure 3).
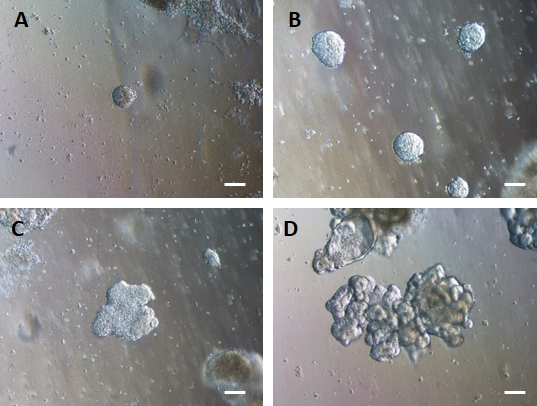
Après le transfert de feuillets de cellules 2D dans la matrice extracellulaire, des amas de cellules 2D seront observés pendant les 24 premières heures. Après 48 h, les feuilles de cellules devraient commencer à s’organiser automatiquement en structures sphéroïdes 3D plus compactes qui sont initialement petites (Figure 4A), puis augmentent progressivement en taille et en complexité au cours des 7 à 10 jours de culture (Figure 4B et Figure 4C). Les organoïdes ne doivent pas être passés tant qu’ils n’ont pas atteint une morphologie organoïde/sphéroïde claire avec un épithélium évident avec la lumière orientée vers le centre de l’organoïde/sphéroïde (Figure 4D). À ce stade, l’immunocytochimie peut être utilisée pour confirmer l’expression de marqueurs intestinaux tels que la villine et le CDX2 (Figure 5). Tous les amas de cellules 2D ne se développeront pas en organoïdes et il y aura des cellules mortes contaminantes dans la matrice extracellulaire. Ces feuilles mortes de cellules doivent être ignorées jusqu’à ce que les cellules survivantes aient formé de gros organoïdes et soient prêtes à passer.
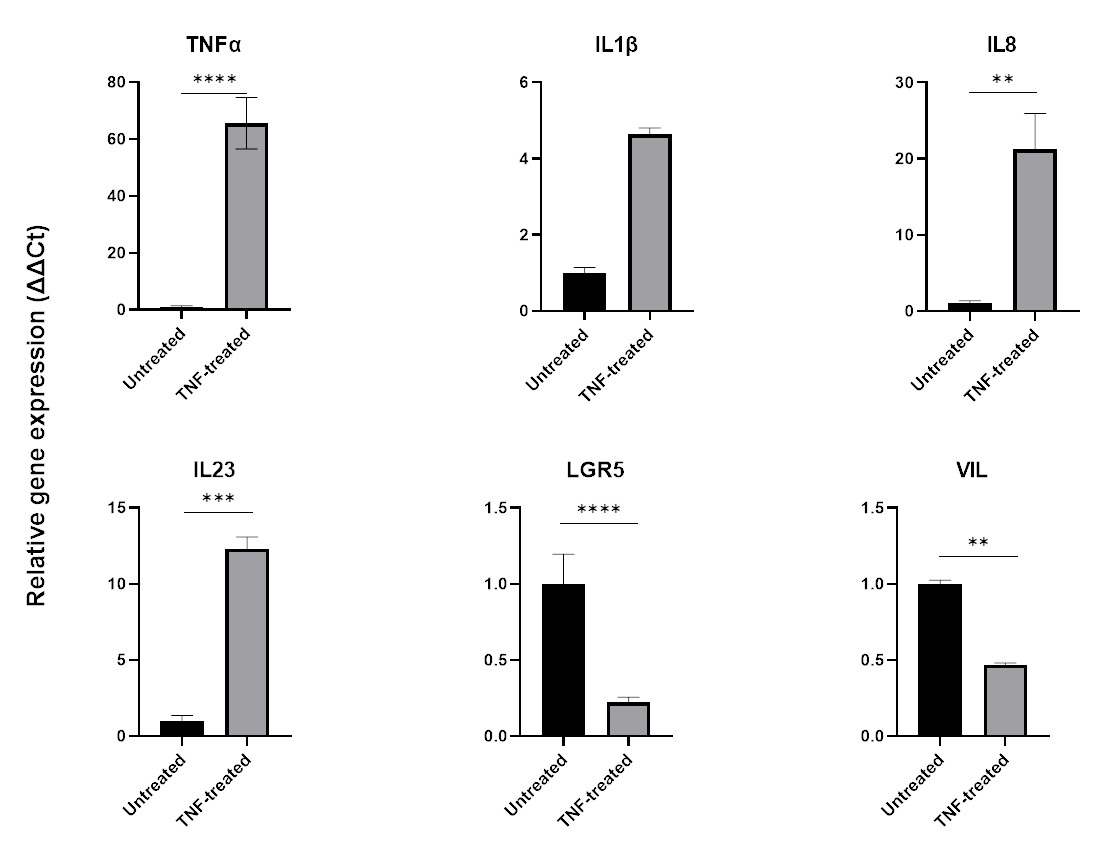
Pour modéliser l’inflammation, le TNFα peut être ajouté au milieu de culture tissulaire pendant 24 à 48 h. Après incubation avec des molécules pro-inflammatoires, les organoïdes sont récoltés en utilisant la même technique que celle utilisée pour leur isolement et leur passage, puis lysés à l’aide d’une application compatible avec les tampons cellulaires telle que la QPCR ou le western blot. Si des expositions plus courtes sont nécessaires, les organoïdes doivent d’abord être retirés de la matrice extracellulaire et exposés au TNFα en suspension à l’aide d’un tube de 1,5 ml. Le traitement des organoïdes intestinaux par TNFα pendant 48 h induit généralement l’expression de marqueurs pro-inflammatoires (TNFα, IL1B, IL8, IL23) tout en affectant négativement l’expression des marqueurs épithéliaux intestinaux (LGR5, VIL) (Figure 6).
| Milieu basal de l’endoderme | 50 ml | |
| RPMI 1640 | 48,5 ml | |
| Supplément B27 | 1 ml | |
| 1 % de la NEAA | 0,5 mL | |
Tableau 1 : Composition du milieu endodermique basal pour la différenciation de l’endoderme.
| Milieux basaux intestinaux | 50 ml | |
| DMEM/F12 avancé | 46,5 mL | |
| Tampon HEPES | 0,5 mL | |
| GlutaMAX | 0,5 mL | |
| Nicotinamide | 0,5 mL | |
| Supplément de N2 | 0,5 mL | |
| Supplément B27 | 1,0 mL | |
| Stylo/streptocoque | 0,5 mL | |
Tableau 2 : Composition des milieux intestinaux basaux pour la culture d’organoïdes intestinaux

Figure 1 : Changements morphologiques au cours de la différenciation de hiPSC via l’endoderme définitif vers la lignée de l’intestin postérieur.
(A) Aperçu schématique du protocole de différenciation intestinale. Cette lignée cellulaire de hiPSC forme des colonies lâches de petites cellules avec un rapport noyau/cytoplasme élevé. Au fur et à mesure de la différenciation, les cellules subissent des changements compatibles avec la transition du phénotype épithélial au phénotype mésenchymateux et forment une monocouche uniforme par DE D3. Une fois que les signaux appropriés sont délivrés, les cellules DE s’allongent et forment une monocouche plus dense avec des sphéroïdes 3D apparaissant dès que HD D3, mais cela dépend de la lignée cellulaire utilisée et n’est pas une exigence pour la transition vers la culture 3D (B). Barre d’échelle : 100 μm. Veuillez cliquer ici pour voir une version agrandie de cette figure.
{kind=link}

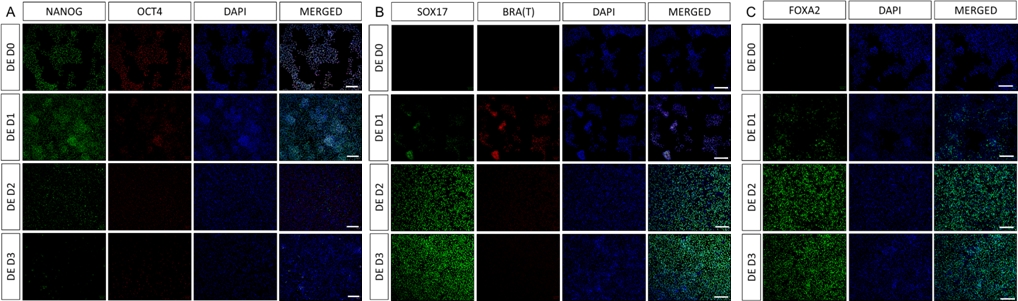
Figure 2 : La différenciation des hiPSCs en endoderme HG induit l’expression des gènes endodermiques.
L’immunomarquage des hiPSC se différenciant en endoderme définitif montre des changements dans l’expression des TF au niveau de la protéine. Les marqueurs de pluripotence (NANOG et OCT4) sont régulés à la baisse par DE D3 (A). L’expression du marqueur mésendodermique BRA(T) est présente à D1 du protocole (B) et les TF spécifiques à l’DE SOX17 et FOXA2 apparaissent à D2 (B et C). Barre d’échelle : 200 mm. Veuillez cliquer ici pour voir une version agrandie de cette figure.
{kind=link}

Figure 3 : Modifications de l’expression génique par qPCR lors de la différenciation des hiPSC vers l’endoderme de l’intestin postérieur (HG).
Les gènes associés à la pluripotence sont régulés à la baisse (OCT4, NANOG, SOX2) suivis de l’expression transitoire des gènes du mésendoderme (T, EOMES, MIXL1), et enfin de l’expression des gènes DE (SOX17, FOXA2, CXCR4) et des gènes de l’intestin postérieur (CDX2, GATA4, HNF4a). Les données sont présentées sous forme de moyenne ±écart-type. Veuillez cliquer ici pour voir une version agrandie de cette figure.
{kind=link}

Figure 4 : L’endoderme HG s’auto-assemble pour former des organoïdes intestinaux 3D en culture de matrice extracellulaire 3D.
L’endoderme HG est transféré dans une culture de matrice extracellulaire 3D appropriée et forme initialement de petits amas solides de cellules (A). Les amas de l’endoderme HG se dilatent au cours de 7 à 10 jours de culture (B), puis deviennent asymétriques et commencent à former un épithélium plus complexe (C), donnant finalement naissance à des organoïdes avec une morphologie épithéliale claire et une surface luminale orientée vers le centre de l’organoïde (D). Barres d’échelle = 50 μm Veuillez cliquer ici pour voir une version agrandie de cette figure.
{kind=link}

Figure 5 : Les organoïdes intestinaux établis dérivés de hiPSC expriment des marqueurs intestinaux.
Immunocytochimie montrant l’expression de CDX2 et de Villin. Barres d’échelle = 100 μm Veuillez cliquer ici pour voir une version agrandie de cette figure.
{kind=link}

Figure 6 : Effet du TNFα sur le profil inflammatoire et l’expression des cellules intestinales d’organoïdes intestinaux sains.
Profil inflammatoire d’organoïdes du côlon sains après un traitement de 48 heures par TNFα (40 ng/mL). L’expression des marqueurs pro-inflammatoires (TNFα, IL-8 et IL-23) augmente après exposition au TNFα, tandis que dans le même temps, l’expression des marqueurs épithéliaux intestinaux (LGR5, VIL) est régulée à la baisse. Les analyses statistiques ont été effectuées par test t bilatéral de l’étudiant. Les données sont exprimées en moyenne ±écart-type de chaque groupe. *P < .01 ; **P < .001 ; P < .0001. (n = 3). Veuillez cliquer ici pour voir une version agrandie de cette figure.
{kind=link}
Discussion
Nous décrivons ici un protocole de différenciation des cellules pluripotentes humaines en organoïdes intestinaux humains. Nous démontrons leur utilisation pour étudier l’inflammation ; cependant, cela peut être appliqué à divers contextes, et couplé avec les approches d’édition de gènes CRISPR/Cas914, sur n’importe quel fond génétique. Une fois différenciés, suivant la séquence de différenciation développementale naturelle de l’endoderme définitif, de l’endoderme de l’intestin postérieur puis de l’épithélium intestinal, les organoïdes qui en résultent peuvent être cultivés et traités en continu pendant plus de 12 mois.
Un aspect essentiel de ce protocole est la densité initiale de placage des cellules souches indifférenciées avant la différenciation de l’endoderme. Si cela n’est pas suffisamment optimisé, les cellules mourront probablement au cours de l’étape initiale de différenciation de l’ED, ou réduiront l’efficacité de la différenciation de l’ED, (si les cellules sont trop denses). La densité de départ correcte doit être optimisée pour la lignée cellulaire utilisée, et la densité correcte doit générer une monocouche à la fin de DE D3. La cytométrie en flux doit être utilisée pour déterminer l’efficacité de la spécification DE et nous constatons généralement que >80 % des cellules sont positives pour SOX17 et/ou CXCR4. Lorsque le nombre de cellules positives à SOX17 est inférieur à 60 %, l’efficacité de la structuration HG est affectée, ce qui entraîne moins de formation d’organoïdes lors du transfert dans la matrice extracellulaire. Cela finira par entraîner l’échec des cultures organoïdes résultantes. Pour déterminer si la structuration de DE en HG a été couronnée de succès, nous évaluons le nombre de cellules positives CDX2 par cytométrie en flux et nous nous attendons généralement à voir >80 % de cellules positives. Encore une fois, si le nombre de cellules positives CDX2 tombe en dessous de 50%, cela aura un effet négatif sur le nombre d’organoïdes intestinaux générés lors du transfert vers la culture de la matrice extracellulaire 3D.
Après le transfert des monocouches 2D dans la culture 3D, de petites sphères compactes devraient apparaître 24 à 48 h après le transfert. De grandes feuilles de cellules mortes peuvent apparaître en fonction de l’efficacité de différenciation de la lignée cellulaire utilisée. Au lieu de faire passer immédiatement des cultures pour éliminer ces débris, nous permettons aux organoïdes de se former et de développer pleinement leur structure plus complexe et plissée. Attendre 7 à 10 jours avant de tenter le premier passage permet de s’assurer que suffisamment de cellules en division sont présentes pour générer de nombreux nouveaux organoïdes intestinaux. Tous les débris encore présents dans la culture peuvent être facilement éliminés pendant le processus de passage en faisant tourner lentement les mélanges organoïdes/débris dissociés dans un tube avec une vitesse suffisante pour granuler les organoïdes mais laisser une feuille de cellules flottant dans le milieu. Les milieux et les débris cellulaires peuvent ensuite être aspirés de manière à ce qu’il ne reste que la pastille d’organoïdes.
La limite de cette approche est que les types de cellules dérivées de hiPSC ne sont souvent pas complètement matures en termes d’expression génique et de profils fonctionnels. Pour déterminer si le tissu intestinal dérivé de hiPSC convient à des applications spécifiques, les organoïdes doivent être caractérisés pour différents types de cellules, y compris les entérocytes (VIL), les cellules entéroendocrines (neurog3), les cellules caliciformes (MUC2), les cellules amplificatrices transitoires (CD133), les cellules panéthes (FZD5) et les cellules souches LGR5+ (LGR5) afin de déterminer la composition cellulaire des organoïdes.
Dans l’ensemble, l’avantage majeur de ce protocole par rapport à de nombreux autres protocoles de différenciation d’organoïdes est que cette plate-forme de culture est très rentable en raison du remplacement de plusieurs protéines recombinantes et préparations de milieux conditionnés par de petites molécules15,16. La différenciation en HG est très simple et rapide et peut être appliquée à la fois aux cellules souches embryonnaires humaines et aux cellules souches pluripotentes induites avec des résultats identiques. Lorsqu’il est suivi rigoureusement et optimisé pour les lignées cellulaires utilisées, il fournit une plate-forme modèle relativement simple, exempte de cellules mésenchymateuses contaminantes, qui peut ensuite être appliquée à l’étude de l’épithélium intestinal dans divers contextes, notamment l’inflammation, les interactions hôte-pathogène7. La modélisation de la fibrose intestinale pourrait être étudiée en fournissant des stimuli pro-fibrotiques, puis en évaluant l’expression des protéines de la matrice extracellulaire telles que le collagène, la laminine et la fibronectine par QPCR, western blot et ELISA. L’utilisation de techniques d’édition de gènes CRISPR/Cas9 sur des lignées de cellules souches indifférenciées avant la différenciation permet de créer des organoïdes de knock-out génique ou de surexpression de protéines qui pourraient être utilisés pour créer des organoïdes spécifiques à la maladie et des modèles de maladie plus complexes 14,17,18.
Déclarations de divulgation
Les auteurs n’ont rien à divulguer.
Remerciements
NH est financé par le MRC (MR/S009930/1) et le Wellcome Trust (204267/Z/16/Z), le est financé par le MRC PhD DTP, KLF est financé par le BBSRC iCASE.
matériels
| Name | Company | Catalog Number | Comments |
| A83-01 | Tocris | 2939 | |
| Activin A | R&D | 338-AC | |
| Advanced DMEM/F12 (1X) | Life Technologies | 12654-010 | |
| B27 supplement | Gibco | 17504044 | |
| CHIR99021 | Sigma | SML1046-5MG | |
| Epidermal Growth Factor | R&D Systems | 236-EG-01M | |
| Gastrin | Sigma Aldrich | G9145 | |
| GlutaMAX (100X) | Life Technologies | 15630-056 | |
| Growth Factor reduced Matrigel | BD | ||
| HEPES Buffer solution (1M) | Life Technologies | 15630-080 | |
| N2 Supplement (100X) | Gibco | 17502-048 | |
| N-acetyl-cysteine | Sigma Aldrich | A7250 | |
| Nicotinamide | Sigma Aldrich | N0636 | |
| Noggin | R&D Systems | 6057-NG | |
| Non-essential amino acids | Gibco | 11140-050 | |
| Paraformaldehyde | VWR | 9713.5 | |
| Penicillin/Streptomycin | Gibco | 15140122 | |
| Phosphate Buffered Saline | Gibco | 14190-094 | |
| Retinoic Acid | Sigma | 302-79-4 | |
| ROCK inhibitor | Tocris | 1254/1 | |
| ROCK inhibitor Y-27632 | Tocris | 1254 | |
| RPMI | Sigma | R8758-500ml | |
| R-Spondin-1 | Peprotech | 120-38 | |
| SB202190 | Tocris | 1264 | |
| TrypLe Express | Gibco | 12604-021 | |
| Wnt 3a | R&D | 5036-WN |
Références
- Thomson, J. A., et al. Embryonic stem cell lines derived from human blastocysts. Science. 282, 1145-1147 (1998).
- Takahashi, K., Yamanaka, S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell. 126, 663-676 (2006).
- Takahashi, K., et al. Induction of pluripotent stem cells from adult human fibroblasts by defined factors. Cell. 131, 861-872 (2007).
- Rashid, S. T., et al. Modeling inherited metabolic disorders of the liver using human induced pluripotent stem cells. Journal of Clinical Investigation. 120, 3127-3136 (2010).
- Vallier, L., Alexander, M., Pedersen, R. A. Activin/Nodal and FGF pathways cooperate to maintain pluripotency of human embryonic stem cells. Journal of Cell Science. 118, 4495-4509 (2005).
- Jaremko, K. L., Marikawa, Y. Regulation of developmental competence and commitment towards the definitive endoderm lineage in human embryonic stem cells. Stem Cell Research. 10, 489-502 (2013).
- Forbester, J. L., et al. Interaction of Salmonella enterica Serovar Typhimurium with Intestinal Organoids Derived from Human Induced Pluripotent Stem Cells. Infection and Immunity. 83, 2926-2934 (2015).
- Hannan, N. R., et al. Generation of multipotent foregut stem cells from human pluripotent stem cells. Stem Cell Reports. 1, 293-306 (2013).
- Sato, T., et al. Single Lgr5 stem cells build crypt-villus structures in vitro without a mesenchymal niche. Nature. 459, 262-265 (2009).
- de Souza, H. S., Fiocchi, C. Immunopathogenesis of IBD: current state of the art. Nature Reviews Gastroenterology & Hepatology. 13, 13-27 (2016).
- Tripathi, K., Feuerstein, J. D. New developments in ulcerative colitis: latest evidence on management, treatment, and maintenance. Drugs in Context. 8, 212572 (2019).
- Ganesh, K., et al. A rectal cancer organoid platform to study individual responses to chemoradiation. Nature Medicine. 25, 1607-1614 (2019).
- Fair, K. L., Colquhoun, J., Hannan, N. R. F. Intestinal organoids for modelling intestinal development and disease. Philosophical Transactions of the Royal Society B: Biological Sciences. 373, (2018).
- Cuevas-Ocaña, S., Yang, J. Y., Aushev, M., Schlossmacher, G., Bear, C. E., Hannan, N. R. F., Perkins, N. D., Rossant, J., Wong, A. P., Gray, M. A. A Cell- Based Optimised Approach for Rapid and Efficient Gene Editing of Human Pluripotent Stem Cells. Int. J. Mol. Sci. 24, 10266 (2023).
- Spence, J. R., et al. Directed differentiation of human pluripotent stem cells into intestinal tissue in vitro. Nature. 470, 105-109 (2011).
- Sato, T., et al. Long-term Expansion of Epithelial Organoids From Human Colon, Adenoma, Adenocarcinoma, and Barrett's Epithelium. Gastroenterology. 141, 1762-1772 (2011).
- Giacalone, J. C., et al. CRISPR-Cas9-Based Genome Editing of Human Induced Pluripotent Stem Cells. Current Protocols in Stem Cell Biology. 44, 1-22 (2018).
- Bruntraeger, M., Byrne, M., Long, K., Bassett, A. R., Luo, Y. . CRISPR Gene Editing: Methods and Protocols. , 153-183 (2019).
Réimpressions et Autorisations
Demande d’autorisation pour utiliser le texte ou les figures de cet article JoVE
Demande d’autorisationThis article has been published
Video Coming Soon
À PROPOS DE JoVE
Copyright © 2025 MyJoVE Corporation. Tous droits réservés.