
Method Article
Optik tabanlı bir sistem kullanarak hidrojel gömülü bozulmamış fare kas liflerinin yüksek verimli kasılma ölçümleri
* Bu yazarlar eşit katkıda bulunmuştur
Bu Makalede
Özet
İskelet kası fonksiyonu, geleneksel olarak zahmetli, düşük verimli yaklaşımlar kullanılarak, izole kas liflerinin kontraktilitesini ölçerek değerlendirilebilir. Burada, hidrojel gömülü kas liflerinin kontraktilitesini ölçmek için optik tabanlı, yüksek verimli bir yöntem tarif ediyoruz. Bu yaklaşımın ilaç taraması ve terapötik gelişim için uygulamaları vardır.
Özet
İn vitro hücre kültürü, hücresel süreçleri değerlendirmek ve terapötik stratejileri test etmek için güçlü bir araçtır. İskelet kası için en yaygın yaklaşımlar, miyojenik progenitör hücrelerin olgunlaşmamış miyotüplere ayırt edilmesini veya izole edilmiş bireysel kas liflerinin kısa süreli ex vivo kültürünü içerir. Ex vivo kültürün in vitro üzerindeki önemli bir yararı, karmaşık hücresel mimarinin ve kontraktil özelliklerin korunmasıdır. Burada, bozulmamış fleksör digitorum brevis kas liflerinin farelerden izolasyonu ve ardından ex vivo kültürü için deneysel bir protokolü detaylandırıyoruz. Bu protokolde, kas lifleri, lifleri hareketsiz hale getirmek ve kasılma işlevlerini sürdürmek için fibrin bazlı ve bazal membran matris hidrojel içine gömülür. Daha sonra optik tabanlı, yüksek verimli bir kontraktilite sistemi kullanarak kas lifi kasılma fonksiyonunu değerlendirmek için yöntemler açıklıyoruz. Gömülü kas lifleri, kasılmaları indüklemek için elektriksel olarak uyarılır, daha sonra sarkomer kısalması ve kasılma hızı gibi fonksiyonel özellikleri, optik bazlı niceleme kullanılarak değerlendirilir. Kas lifi kültürünün bu sistemle birleştirilmesi, farmakolojik ajanların kasılma fonksiyonu üzerindeki etkilerinin yüksek verimli test edilmesine ve genetik kas bozukluklarının ex vivo çalışmalarına izin verir. Son olarak, bu protokol canlı hücre mikroskobu kullanarak kas liflerindeki dinamik hücresel süreçleri incelemek için de uyarlanabilir.
Giriş
İn vitro hücre kültürü tekniklerindeki ilerlemeler, doku rejeneratif yeteneklerini, patofizyolojik hücresel mekanizmaları ve müteakip terapötik stratejileri incelemek için yeni olanaklar ortaya çıkarmıştır ve bunların hepsi memeli dokularını iyi kontrol edilen koşullar altında kullanmaktadır 1,2,3. İn vitro kültür sistemlerinin kullanımı kas araştırma alanında iyi bir şekilde kurulmuştur 4,5. Genel olarak, miyojenik progenitör hücrelerden in vitro diferansiye olgunlaşmamış miyotüpler 2,6,7,8 kullanılır. Daha olgun kas lifleri üretmek için farklılaşma protokolünde ilerleme kaydedilmesine rağmen9, bunların olgunlaşmamışlığı hala bulguların in vivo ayar 1,10'a çevrilmesini sınırlamaktadır. Kas biyolojisi alanındaki merkezi bir konu, in vitro diferansiye miyotüplerin, doğal kas dokusunda gözlenen karmaşık hücre içi yapıları, hücre sinyal süreçlerini ve hücre dışı etkileşimleri tam olarak özetleyememesi ve daha da önemlisi, kas lifleri tarafından üretilen kasılma kuvvetlerini özetleyememesidir 1,2,10,11,12 . Ek olarak, farklılaşma işlemi sırasında miyotüplerin koordinasyonsuz kasılması genellikle kültür yemeklerinden kendiliğinden ayrılma ile sonuçlanır, bu da in vitro farklılaştırılmış miyotüplerin kontraktil değerlendirmesini zorlaştırır ve kalitatif veya yarı kantitatif değerlendirme ile sınırlı kılar 8,11,12,13. Bu sınırlamalar genellikle hayvanlarla düzenli in vivo deneyler yapılmasını gerektirir, özellikle de kas kontraktilitesi birincildeneysel sonuçsa1.
İn vitro diferansiye miyotüplerin kültürlenmesine bir alternatif, izole olgun kas liflerinin ex vivo kültürüdür 1,14. Ex vivo kültür sırasında, gelişimsel olarak olgun kas dokusu vücuttan eksize edilir, ardından laboratuvar koşullarında yetiştirme için tek hücreli izolasyon yapılır 1,14. İzole olgun kas lifleri, doğal doku14,15 içinde gözlenen kompleks hücresel yapılarını korur ve bu yöntem, iyi tanımlanmış ve kontrol edilebilir bir kültür ortamında genetik manipülasyon ve ilaç taraması gibi doğrudan müdahaleler için olasılık açar. İskelet kası lif izolasyonu ve ex vivo kültürü ile ilgili ilk raporlardan biri 1930'lara kadar uzanmaktadır; Bununla birlikte, bu protokolden elde edilebilir liflerin verimi düşüktü16. İzolasyon prosedürünün ve kültür koşullarının sürekli optimizasyonu ile, canlı ve fonksiyonel kas liflerinin miktarında önemli bir iyileşme artık mümkündür 14,15,17,18,19. Kültür koşullarındaki böyle bir gelişme, izole kas liflerinin kültür kabına yapışmasını teşvik etmek için kültür yemeklerinin hücre dışı matris proteinleri ile kaplanmasını içerir15,18,20. Genellikle, laminin kaplama kullanılır, çünkü laminin, kasların hücre dışı matrisi içinde en bol bulunan elementlerden biridir20,21. İzolasyon prosedürünün optimizasyonu, kültür yemeklerinin kaplanması ile birleştiğinde, kas araştırma alanının, kültürde bozulmamış hücresel mimariye ve kasılma işlevselliğine sahip izole edilebilir canlı kas liflerini kısa süreliğine tutmasını sağlamıştır 1,15,18,22.
Kas alanında kuvvet ve kasılma yeteneklerini ölçmek için kullanılan en geleneksel yaklaşım, bireysel kas liflerini bir uzunluk sürücü motoru ile bir kuvvet dönüştürücü23,24 arasına monte etmektir. Genel olarak, bu motor tahrikli kurulumlar için kullanılan kas lifleri, ya dondurulmuş ya da taze dokudan diseke edilir, ardından kas lifi kasılmasını indüklemek için değişen kalsiyum konsantrasyonlarının kullanıldığı harici kalsiyum aktivasyonuna izin veren geçirgenlik veya "derileme" yapılır24. Bu yöntem kas lifi kasılma ölçümleri için altın standart olsa da, bir seferde sadece tek bir kas lifi ölçülebilir, bu da bu tekniği zahmetli ve zaman alıcı bir prosedür haline getirir25. Ayrıca, kas liflerinin izolasyonu ve derilendirme prosedürü, uyarılma-kasılma eşleşmesinde (yani, kalsiyum salınımı ve ardından sarkoplazmik retikuluma geri alımı) yer alan çeşitli yapıları bozar, böylece gevşeme kinetiğinin ve bu süreci etkileyebilecek herhangi bir hastalığın incelenmesine izin vermez26,27. Derili lif preparatına bir alternatif, elektriksel aktivasyona yanıt olarak kasılma kuvvetlerinin ölçülebildiği sağlam kas liflerini izole etmek için mekanik diseksiyon kullanmaktır28; Bununla birlikte, bu yaklaşım teknik olarak zorlu ve çok zaman alıcıdır, bu da düşük verimli ölçümlerle sonuçlanır. Son olarak, hem derili hem de sağlam preparatlarda, kas hücreleri kasılma ölçümleri sırasında hücre dışı ortamdan tamamen çıkarılır 24, bu da hücre dışı matriks kompozisyonunun / sertliğinin kas lifi kasılması üzerindeki etkisinin araştırılmasını imkansız kılar24. Sonuç olarak, izole edilmiş sağlam kas liflerinin kas lifi kontraktilitesi ölçümlerini yüksek verimli bir şekilde sağlamak ve kas lifleri ile hücre dışı matris arasındaki bağlantıyı yeniden oluşturmak için alternatif yöntemlerin geliştirilmesine ihtiyaç vardır.
Son zamanlarda, yüksek verimli kas lifi kontraktilitesi ölçümleri için optik tabanlı yeni bir yaklaşım geliştirilmiştir29. Bu optik tabanlı sistem, yüksek hızlı görüntüleme kullanarak kasılma sırasında sarkomer uzunluğunu değerlendirmek için sarkomerlerin periyodikliğini ölçer. Bu sistemde, optikler hareket ettirilirken hücreler kültür kabında yerinde kalır, böylece birden fazla hücrenin ölçümleri arasında gereken süre en aza indirilir29. Bu yüksek verimli, optik tabanlı yaklaşımı kullanmanın en büyük avantajı, doğal dokununkine benzer kültür koşullarının geliştirilmesine izin vermesidir. Doğal in vivo koşulları taklit etmek için kullanılan bir yaklaşım, hücreleri hidrojellere gömmektir30. Tipik olarak, bir hidrojel, hacmini ve şeklini koruyabilen viskoelastik bir malzemedir ve hidrojeller hem katı hem de sıvı malzemelerin özelliklerine sahiptir31. Katı kısım, birbirine çapraz bağlanmış polimer zincirlerinden oluşur ve net30,31'e benzeyen bir yapı oluşturur. Hidrojellerin malzeme özellikleri, kasların matris birikimini taklit edecek şekilde ayarlanabilir30,31. Bu nedenle, yüksek verimli, optik tabanlı bir sistemin hidrojellere gömülü hücrelerle kombinasyonu, hücre dışı matris kompozisyonunun ve mekanik özelliklerin kas lifi işlevselliği üzerindeki etkilerini değerlendirmek için yeni olanaklar açmaktadır.
Bu makalenin genel amacı, 1) doğal doku ortamını taklit eden koşullarda kas liflerinin enzimatik izolasyonu ve ex vivo kültürü için metodolojiyi tanımlamak ve 2) yüksek verimli bir yaklaşım kullanarak kas lifi kontraktilitesini değerlendirmektir. Enzimatik sindirim kullanarak fleksör digitorum brevis (FDB) kasından çok sayıda tek kas lifini kolayca izole etmek için ayrıntılı bir metodoloji tarif ediyoruz. Ek olarak, kasların doğal ortamını taklit etmek ve kas lifi canlılığını ve kontraktilitesini geliştirmek için izole edilmiş kas liflerini fibrin bazlı bir hidrojele gömmek için bir teknik tarif ediyoruz. Daha sonra, yakın zamanda geliştirilen bu sistemi kullanarak canlı kas lifi kasılmalarını in vitro olarak ölçmek için yüksek verimli bir yöntemin ana hatlarını çiziyoruz. Bu gömme prosedürünün ek bir avantajı, büzülme sırasında liflerin hareketsiz hale getirilmesidir ve bu da bu ölçümlerin sinyal-gürültü oranını artırabilir. Bu jel gömme yöntemi, hem tek polimer hem de kompozit jel kapsülleme prosedürleri için uygulanabilir ve hücre dışı matris bileşiminin kas lifi kontraktilitesi üzerindeki etkilerinin değerlendirilmesini kolaylaştırır.
Protokol
Ex vivo kasılma çalışmaları için, Hollanda Hayvan Araştırmaları Kanunu'nun izniyle Avrupa Konseyi Direktifi (2010/63/EU) uyarınca VU Üniversitesi'nin diğer onaylanmış araştırma projeleri ve / veya üreme fazlası için kurban edilen hayvanlardan ölüm sonrası doku elde edilmiştir.
1. Malzeme hazırlama
- Pipetleri tritürasyon için hazırlayın (kullanıma kadar bir akış kabininde %70 etanol içinde saklayın). Farklı delik boyutları oluşturmak için iki P1.000 ucun uçlarını kesin; Deliğin, kasın pipeti bloke etmeden geçebileceği kadar büyük olduğundan emin olun. Kesim uçlarını bir alevden geçirin, böylece pürüzsüz hale gelirler.
- Sylgard yemeklerini32. başka bir yerde açıklandığı gibi hazırlayın ve izolasyon sırasında pençe ve kasın sabitlenmesinde kullanılmak üzere önceden% 70 etanol ile sterilize edin.
NOT: Sylgard bulaşıkları, deiyonize su ve% 70 etanol ile iyice temizlendikten sonra tekrar kullanılabilir. - İzolasyon prosedüründen önce aşağıdaki çözeltileri hazırlayın: diseksiyon ortamı, fibrin kültürü ortamı ve kas sindirim ortamı (bkz. Tablo 1).
NOT: Kas sindirim ortamı, 0,22 μm'lik bir filtre kullanılarak filtreyle sterilize edilmelidir. Hazırlanan tüm çözeltiler, kullanımdan önce en az 30 dakika boyunca 37 ° C'de standart bir hücre kültürü inkübatöründe ve% 5 CO2'de dengelenmelidir. - Kas sindiriminden sonra, hidrojeli dökmek için aşağıdaki çözeltileri hazırlayın: hücre karışımı ve matris karışımı (bkz. Tablo 2). Hücre karışımını ve matris karışımını, 2.5 mg / mL'lik nihai fibrin konsantrasyonu için 1: 1 oranında birleştirin.
NOT: Matrisi buz üzerinde hazırlayın ve bazal membran matrisinin erken polimerizasyonunu önlemek için önceden soğutulmuş pipet uçları kullanın. Burada kullanılan trombin:fibrinojen oranı 1:10 (mg fibrinojen başına birim) 30 dakikalık bir polimerizasyon süresi için optimize edilmiştir. Jel 30 dakika içinde polimerize olursa, daha düşük bir trombin konsantrasyonu kullanılmalıdır. Daha fazla bilgi için tartışmaya bakın.
2. FDB kas diseksiyonu
- Servikal çıkık ile fareyi ötenazi yapın. Arka bacağınızı% 70 alkolle dezenfekte edin.
- Alt arka bacağı ayak bileğinin üstünden kesin. Ayağın sırt tarafındaki cildi ayak parmaklarına doğru kesin.
NOT: Daha sonra ihtiyaç duyulursa alt ekstremite kaslarına ve tibiaya zarar gelmesini önlemek için ayak bileği eklemindeki alt arka bacağı kesin. - Cildi ayak parmaklarına doğru dikkatlice soyun. Kaslara zarar vermemeye dikkat edin. FDB, ayağın ventral tarafındaki en yüzeysel kastır.
- Disseke edilmiş ayağı, 37 ° C'de 10 mL önceden ısıtılmış diseksiyon ortamına sahip bir Sylgard kabına yerleştirin. Ayağı hala ayak parmaklarına bağlı olan deriden geçirin ve alt bacağı ayak bileğinin ötesine sabitleyin.
- Kasın üstündeki bağ dokusunu dikkatlice çıkarın. Tendonu topuktan kesin ve kası tendonu ile yukarı kaldırın.
- Bağ dokusu boyunca kasın yanında ve altında kesin. Ayak parmağı tendonları açığa çıkana kadar kesmeye devam edin.
- Üç tendonun uzunluğunun yarısı açığa çıktığında, tendonları kesin ve kası ayaktan serbest bırakın. İSTEĞE BAĞLI: Dördüncü lateral tendonu ve kas liflerini kesin.
- Bağ dokusunu kastan temizleyin ve önceden ısıtılmış diseksiyon ortamı içeren bir tüpe aktarın.
3. FDB kas sindirimi
- Kas sindirim ortamını Tablo 1'e uygun olarak hazırlayın.
- FDB kaslarını serolojik bir pipet kullanarak kas sindirim ortamına aktarın. Bir doku kültürü inkübatöründe 37 ° C'de ve% 5 CO2'de 80 dakika boyunca inkübe edin.
NOT: Bu süre, her kollajenaz partisi için optimize edilmelidir. Kas yıpranmaya başladığında ve genişlemiş göründüğünde sindirim tamamlanır. Sindirim süresinin optimizasyonu için tartışmaya bakın. - Sindirimden sonra, kası 3 mL diseksiyon ortamı içeren 15 mL'lik bir tüpe aktarın ve tritürasyondan önce 30 dakika boyunca inkübe edin.
4. FDB kas tritürasyonu ve yerçekimi sedimantasyonu
- Daha önce hazırlanmış tritürasyon uçlarını (adım 1.1) kullanarak kası pipetleyerek en büyükten en küçük boyuta geçerek kası tritüre edin. Bu adım 5 dakikadan uzun sürerse, dinlenmesine izin vermek için kası 5 dakika boyunca inkübatöre yerleştirin.
- Kas lifleri çoğunlukla tendondan çıkana kadar triturat yapın ve tendon bir P200 ucundan geçebilir. Tendonları çıkarın.
- Ayrışmış FDB liflerini, 10 mL diseksiyon ortamı içeren 15 mL'lik bir tüpe ekleyin ve liflerin inkübatöre 20 dakika boyunca yerleşmesine izin verin. Oluşan pelet gözlemleyin.
- İsteğe bağlı: Üstten 10 mL ortamı çıkarın ve 4.3 adımını tekrarlayın. Bu adım, fazla kalıntıların ve ilişkili monoçekirdekli hücrelerin çıkarılmasını kolaylaştırır.
5. FDB fiber gömme
- Tüm ortamı fiber peletin üstünden dikkatlice çıkarın. FDB kası başına (buz üzerinde) 875 μL hücre karışımındaki hücreleri yeniden askıya alın.
NOT: Bir FDB kası, 24 delikli bir plakanın yedi kuyucuğu için yeterli lif verir. Bu yoğunluk, deneyin ihtiyaçlarına göre ayarlanabilir. Aşağıdaki jel hacmi (250 μL) 24 delikli bir format için optimize edilmiştir, ancak diğer formatlar için buna göre ölçeklendirilebilir. - Hücrenin Aliquot 125 μL'si tek mikrosantrifüj tüplerine karışır.
- Her seferinde bir kuyucuk, bir hücre süspansiyonu aliquot'a 125 μL matris karışımı ekleyin ve kabarcıkların oluşumunu önleyerek dikkatlice yukarı ve aşağı pipetleyerek karıştırın.
- Son karışımı hemen bir kuyuya aktarın.
NOT: Karışımı kuyunun ortasına boruladığınızdan emin olun. Her kuyucuk için adım 5.3 ve adım 5.4'ü yineleyin. - Jelleri bir inkübatörde 30-45 dakika boyunca katılaştırın. Katılaşmadan sonra, kültür ortamını dikkatlice kuyucuklara ekleyin.
NOT: Hızlı pipetleme, hidrojeli kuyudan ayırabilir. Bu noktadan itibaren, lifler uyarılabilir ve ölçülebilir. Bununla birlikte, deneyimlerimize göre, liflerin 24 saat boyunca kültür koşullarına uyum sağlamasına izin vermek, lif kontraktilitesini artırabilir. - Daha uzun bir kültür için, kültür ortamını her 2 günde bir yarı değişiklik yaparak doldurun. Bunu yapmak için, kültür ortamının yarısını çıkarın ve eşit miktarda taze ortam ile değiştirin.
6. Optik tabanlı kontraktil ölçümler
- Optik tabanlı kontraktil ölçüm sistemini (Malzeme Tablosuna bakın), floresan lambayı, elektrik hücresi pacer'ı ve bilgisayarı açın. İzole kas liflerini uyarmak için elektrik uyarıcısını 1,0 Hz, 10,0 V ve 5,00 ms nabız süresine ayarlayın.
- Plakayı ölçüm sistemine yerleştirin. Pacer'ı pacing ekine bağlayın ve kültür plakasına yerleştirin.
- IonWizard programını açın ve Dosya'ya tıklayarak yeni bir dosya açın (ekranın sol üst köşesinde) | Yeni.
- Programın doğru deneyde olup olmadığını kontrol edin, İskelet Sarkomer, Yeni | Deney toplayın. Denemeyi değiştirmek için istediğiniz denemeyi tıklayın ve ekle'ye basın. İskelet sarkomer deneyi için aşağıdaki ayarları uygulayın: Sarc 20x, Ortalama çizgiler, tek FFT, 250 Hz örnekleme hızı ve 10 sn edinme süresi.
NOT: Deney ayarları deneyden önce hazırlanmalıdır. Ayarlar, denemenin ihtiyaçlarına göre ayarlanabilir. - Ölçüm sisteminin sıcaklığını 25 °C'ye ayarlayın. Araç çubuğunun altındaki hücre bulucuyu aç'a tıklayın ve yeni bir ekranın açılmasını bekleyin. Bu ekranın sağ üst köşesinde, plaka tipini ve aktif kuyucukları seçin.
NOT: Ölçümler, hızlı seğiren FDB kas liflerinin kasılma hızını azaltmak için 25 ° C'de gerçekleştirilir, böylece kasılma olayının yetersiz örneklenmesini önler. - Netleme kaydırma çubuğunu ayarlayarak fiberleri netlemeye getirin. Alternatif olarak, bu odaklama işlevi için W tuşunu ve S tuşunu kullanın.
- Elektriksel stimülasyonu başlatmak için tempoyu etkinleştirin. Liflerin şimdi seğirmeye başladığını gözlemleyin.
NOT: Hiçbir lif seğirmiyorsa, tüm tellerin bağlı olduğundan ve pacer'ın tamamen suya batırıldığından emin olun. Bundan sonra hala hareket yoksa, lifler aşırı sindirime veya tritürasyon sırasında aşırı hasara neden olabilir. - Ölçüm alanını, sarkomerler dikey olarak çalışacak şekilde bir fiberin ucuna odaklayın. Sarkomerlerin odaklandığından emin olun. Sarkomerler odaktaysa, araç çubuğunda tek bir tepe noktası görünür. Kasılma sırasında, bu tepe sarkomer kısaldıkça sağa doğru hareket edecektir.
NOT: Mor ölçüm alanı, deneye başlamadan önce ayarlanabilir. Doğru bir ölçüm sağlamak için, ~ 20 sarkomer ekleyin. Bu tepe noktası kasılma sırasında şekil değiştirirse, bu, kasılma sırasında sarkomerin gizlendiğini veya odak dışında olduğunu gösterebilir, bu da gürültüye neden olur. - Araç çubuğunda başlat'ı tıklayarak denemeyi başlatın. Bir ölçüm başlatmak için Q tuşuna basın ve programın 10 kasılma geçicisini ölçmesini bekleyin. Dörtten fazla geçici ses gürültüsüz görünüyorsa, Z tuşuna basarak ölçümü kabul edin. Geçicilerin çok fazla gürültüsü varsa, X tuşuna basarak ölçümü reddedin.
- Koşul başına 10 lif ile 20 lif arasında ölçüm yapın. Daha önce ölçülen liflerin yerleri korunur.
- İsteğe bağlı: Bileşikler ekleyin, daha sonra lifler bu noktada yeniden ölçülebilir.
- Deneme tamamlandığında, hücre bulucu penceresini kapatmak için alt araç çubuğundaki durdur düğmesine basın. Dosyayı kaydedin ve yeni bir dosya başlatın.
- Verileri analiz etmek için "Cytosolver desktop" programını açın. İçe aktar'a tıklayın ve analiz edilecek dosyaları seçin.
- Program analizi bitirdikten sonra mavi, kırmızı ve gri tepeleri arayın. Mavi zirveler, program tarafından kabul edilen geçicilerdir. Kırmızı tepe noktaları program tarafından reddedilen geçici, gri tepe noktaları ise kullanıcı tarafından reddedilen geçici tepe noktalarıdır.
NOT: Rejeksiyon kriterleri analiz yazılımında ayarlanabilir. Genel olarak, değerler maksimum türev limitini aşarsa reddedilir ve geçiciler <0,95'lik eğri uyumuR2 değerlerine göre reddedilir. - Dışa aktar'a tıklayın. Aşağıdaki kutuları işaretleyin: Geçici verilerin ortalaması alındı ve excel'e aktarıldı.
- İşiniz bittiğinde, tüm makineleri kapatın. Kültür tabağını çıkarın ve atın. Pacer elektrotlarını deiyonize su ve% 70 etanol ile temizleyin.
Sonuçlar
Bu protokolü kullanarak, tek FDB kas lifleri izole edildi ve hidrojel içine gömüldü. Kas diseksiyonu prosedürüne genel bir bakış Şekil 1'de gösterilmiştir. FDB kası sağlam tendonlarla açığa çıkar ve fasyadan gevşek kesilir. Kasların tendonlarını fiksasyon noktaları olarak tutmak, izolasyon prosedürü sırasında kas liflerine potansiyel hasarı en aza indirir. Fazla bağ dokusu, kalıntıları ve ikincil hücre tiplerinin büyümesini azaltmak için kesilebilir. Kas eksize edildikten ve yeterince temizlendikten sonra, kas kollajenaz kullanılarak enzimatik olarak sindirilir ve izole edilmiş hücreleri hidrojellere gömmeden önce tritürasyon yoluyla tek kas lifleri salınır. FDB kas liflerinin izolasyonu, kolayca manipüle edilme avantajına sahip nispeten kısa kas lifleri sağlar. Boyutları nedeniyle, FDB kas lifleri dolaşmaya bağlı hasar olmadan güvenli bir şekilde pipetlenebilir ve kolayca bir hidrojel içine gömülebilir. Tek kas lifleri kültür plakalarına çok iyi yapışmadığından, liflerin hidrojel içine gömülmesi, liflerin hücre kültürü ve kasılma ölçümleri sırasında yerinde kalmasını sağlar. Ayrıca, fibrin jeline bir bazal membran matrisinin eklenmesi, kas lifleri ile matris arasındaki etkileşimlere izin verir ve doğal in vivo ortamı taklit eder. İzole edilmiş tek kas lifleri, izolasyondan sonra birkaç gün boyunca kültürde manipüle edilebilir ve korunabilir. Şekil 2'de, bir hidrojel matrisine gömülü izole edilmiş FDB liflerinin bir örneği gösterilmiştir. Sağlıklı lifler görünür sarkomerlere sahiptir ve düz bir şekilde uzatılır (mavi ok), kavisli lifler ise tipik olarak hasar görür veya dayanamaz (sarı ok) ve ölçümlerden çıkarılmalıdır. Hiper büzülmüş lifler, matriste koyu renkli toplanmış nesneler olarak görünür (kırmızı ok). İzolasyon prosedürü başarılı olsaydı, sağlıklı liflerin oranı ~% 75 olmalıdır. Hiperkontrakte liflerin daha büyük bir kısmı genellikle izolasyon prosedürü sırasında hasarı gösterir. Kas liflerinin zarı, kasın aşırı sindirimi veya tritürasyon sırasında liflerin zarar görmesi nedeniyle zarar görebilir. Tritürasyon hasarı esas olarak kas az sindirilmişse ve bu nedenle kolayca parçalanmazsa ortaya çıkar.
Lif işlevselliği ve canlılığı, optik tabanlı, yüksek verimli bir kontraktil ölçüm sistemi kullanılarak kas liflerinin kasılma ölçümleri ile değerlendirildi. Sistem, sarkomer uzunluğu, sarkomer kısalma yüzdesi, kasılma hızı ve gevşeme hızı gibi çeşitli parametreler çıkarır. Kontraktil ölçüm sistemi kullanılarak, kas lifi başına sarkomer kasılması ölçülebilir. Şekil 3 , ölçüm sistemi kullanılarak ölçülen kas lifi kasılmalarının örneklerini göstermektedir. Tek bir kasılma geçicisinden aşağıdaki parametreler elde edilir: başlangıçta sarkomer uzunluğu, kasılma süresi, maksimum kasılmada sarkomer uzunluğu ve gevşeme süresi (Şekil 3A). Bu parametreler sarkomer kısalma, kasılma ve gevşeme hızlarının yüzdesini hesaplamak için kullanılır. Ortalama hız değerleri, gerekirse süre ve mutlak kısaltma değerlerinden de hesaplanabilir. Geçerli bir büzülme ölçümünde düz bir taban çizgisi, ardından zirveye bir düşüş ve taban çizgisine bir dönüş bulunur (Şekil 3A). Gürültü, odaklanmamış sarkomerler veya fiberin anormal hareketi ölçümü geçici olarak etkileyebilir (Şekil 3B, C) ve bu ölçümler manuel olarak atılabilir veya analiz programı tarafından reddedilir. Bu yaklaşımda, sarkomerlerin net bir şekilde görselleştirilmesi, kasılmaların ölçülmesi için önemlidir; Bu nedenle, sarkomerlerin görünürlüğünü azaltan herhangi bir şey gürültüye neden olabilir. Bu, kasılma sırasında sarkomerin odak düzleminin dışında hareketi varsa ortaya çıkabilir. Bir dizi kasılmanın hız veya derinlik bakımından farklılık gösterdiği ölçümler de veri kümesinden çıkarılmalıdır.
Bu sistemle elde edilen tek kas liflerinin kasılma verileri, farklı kültür koşullarını karşılaştırmak için kullanılabilir. Sistemin etkinliği Şekil 4'te gösterilmiştir. Burada, FDB kas liflerinin kasılmalarını hem 2D (laminin kaplı kültür plakaları) hem de 3D (fibrin hidrojel) kültür formatlarında ölçtük. Liflerin jele gömülmesi, yanal hareketin ve diğer hareket artefaktlarının ölçümü etkilemesini önlediğinden, 3D'de kullanılabilir ölçümlerin daha yüksek bir yüzdesi elde edildi (Şekil 4A). Liflerin gömülmesinin, 2D kültürlü liflere kıyasla sarkomer kısalması veya maksimum büzülme hızı değerleri üzerinde anlamlı bir etkisi yoktu (Şekil 4B). Farklı matrislerin FDB kas liflerinin kasılmasını nasıl etkilediğini göstermek için, bu fibrin hidrojelini saf bir bazal membran matrisi (4 mg / mL) ile karşılaştırdık (Şekil 4C). Saf bazal membran matrisinde gözlenen azalmış kontraktilite, muhtemelen jelin sertliğinden veya artan hücre-matris etkileşimlerinden kaynaklanıyordu. 7 mg / mL'ye kadar fibrin konsantrasyonları da test edilmiştir, kasılma hızı ve kısalması üzerinde önemli bir etkisi yoktur (yayınlanmamış veriler). Bu fibrin bazlı hidrojelin kullanımı, kontraktil parametrelerle minimum parazit sağlar.

Şekil 1: FDB kas diseksiyonu prosedürüne genel bakış. (A) Arka bacak ayak bileğinin üzerinde kesilir (kırmızı kesikli çizgi) ve (B) mavi kesikli çizgi boyunca ayağın üst kısmı boyunca kesilerek cilt çıkarılır. (C) Ayak ayak bileğinden ve ayak parmaklarındaki deriden sabitlenir. (D) Maruz kalan kasın ve çevresindeki bağ dokusunun mikroskobik görünümü. Fasya, vaskülatürün içinden geçtiği beyaz opak bir tabaka olarak görülebilir. (E) Fasya kastan çıkarılır ve tendon kırmızı kesikli çizgi boyunca kesilir. (F) FDB kası, kasın altında ve yanında kaldırılarak ve kesilerek altta yatan dokudan ayrılır. (G) Ayak parmaklarının tendonları açıkça görülebildiğinde, FDB kırmızı kesikli çizgi boyunca gevşek kesilir. (H) FDB tendonlar tarafından sabitlenir ve dördüncü lateral tendon ve lifleri kırmızı kesikli çizgi boyunca kesilerek çıkarılabilir. Fazla fasya artık kesilebilir. (I) Temizlendikten sonra, FDB kası kollajenaz çözeltisine aktarılır. Ölçek çubukları = (D-I) 2 mm. Kısaltma: FDB = flexor digitorum brevis. Bu şeklin daha büyük bir versiyonunu görmek için lütfen buraya tıklayın.
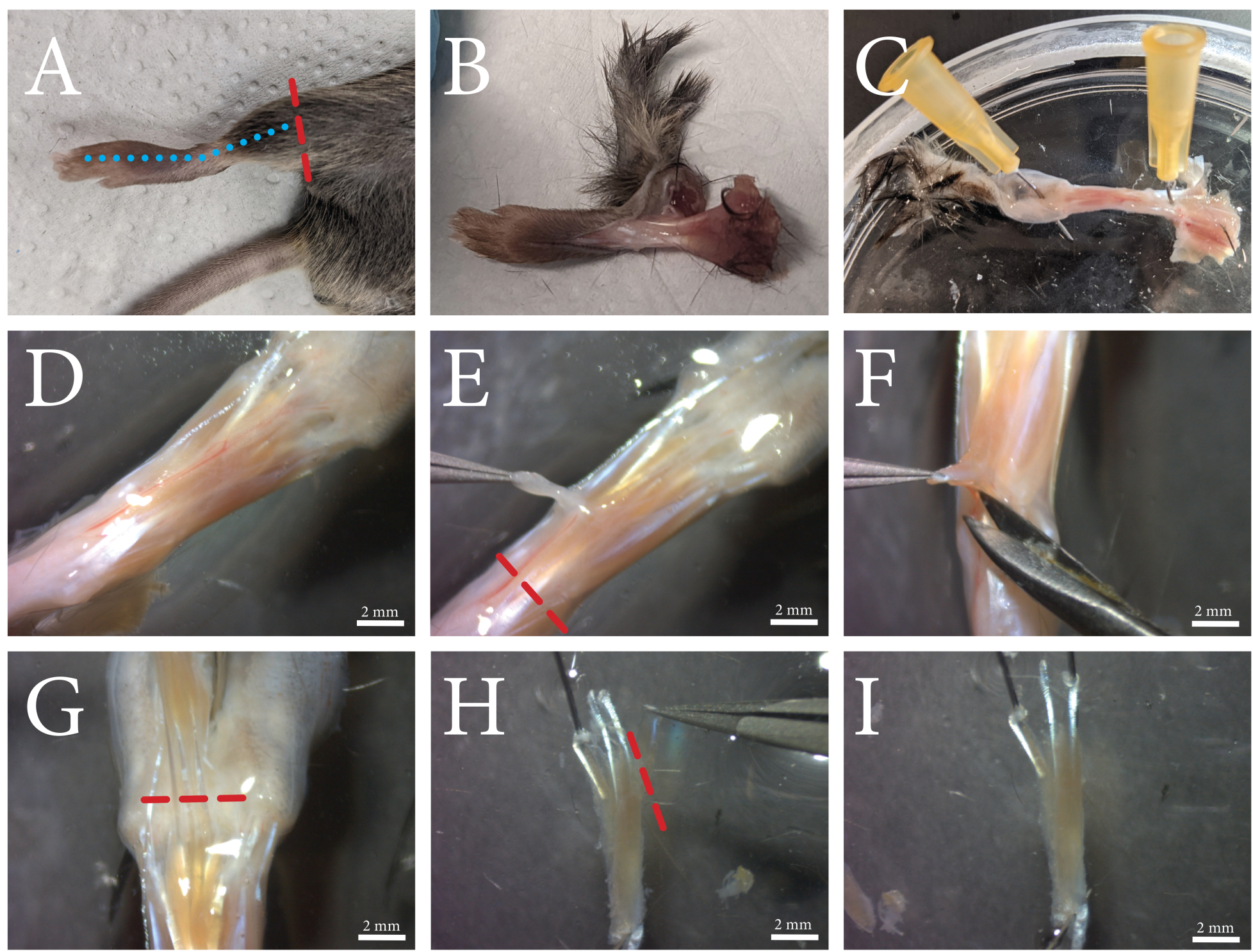{kind=link}

Şekil 2: 24 saatlik kültürden sonra hidrojel içine gömülü FDB kas liflerinin mikroskobik görüntüsü. Mavi ok: Canlı bir FDB kas lifi örneği. Sarı ok: Bükülmüş FDB kas lifi örneği. Bükülmüş kas lifleri canlılığı azaltmış ve kasılmaları bozmuş olabilir ve bu nedenle ölçümlerden çıkarılmalıdır. Kırmızı ok: Hiperkontrasyonlu FDB kas lifi örneği. Aşırı hiperkontraksiyon, tritürasyon çok kuvvetli bir şekilde yapıldığında ortaya çıkar veya kollajenaz, aşırı sindirim, dengesiz kültür ortamının kullanılmasından kaynaklanabilir. Ölçek çubuğu = 100 μm. Kısaltma: FDB = flexor digitorum brevis. Bu şeklin daha büyük bir versiyonunu görmek için lütfen buraya tıklayın.
{kind=link}

Şekil 3: Optik tabanlı, yüksek verimli kontraktil sistem kullanılarak ölçülen fiber büzülme geçicilerine örnek . (A) Normal bir kasılma geçici örneği. Bu geçici, alt araç çubuğunda gösterilen mor kare ile çevrili sarkomerlerden elde edilir. Geçici aşağıdaki bileşenlerden oluşur: başlangıçta sarkomer uzunluğu (yeşil), kasılma süresi (mavi), maksimum kasılmada sarkomer uzunluğu (mor) ve gevşeme süresi (kırmızı). Hız ve büzülme yüzdesi gibi parametreler bu değerlerden hesaplanır. (B) Geçici yetersiz kasılma örneği. Bu ölçümler, gürültü nedeniyle sarkomer sinyali alınmadığında meydana gelir (bkz. kırmızı oklar). (C) Hareket artefaktı olan geçici bir kasılma örneği (bkz. yeşil oklar). Hareket artefaktları, kas lifi kasılma sırasında odağın dışına çıktığında ortaya çıkar. Bu şeklin daha büyük bir versiyonunu görmek için lütfen buraya tıklayın.
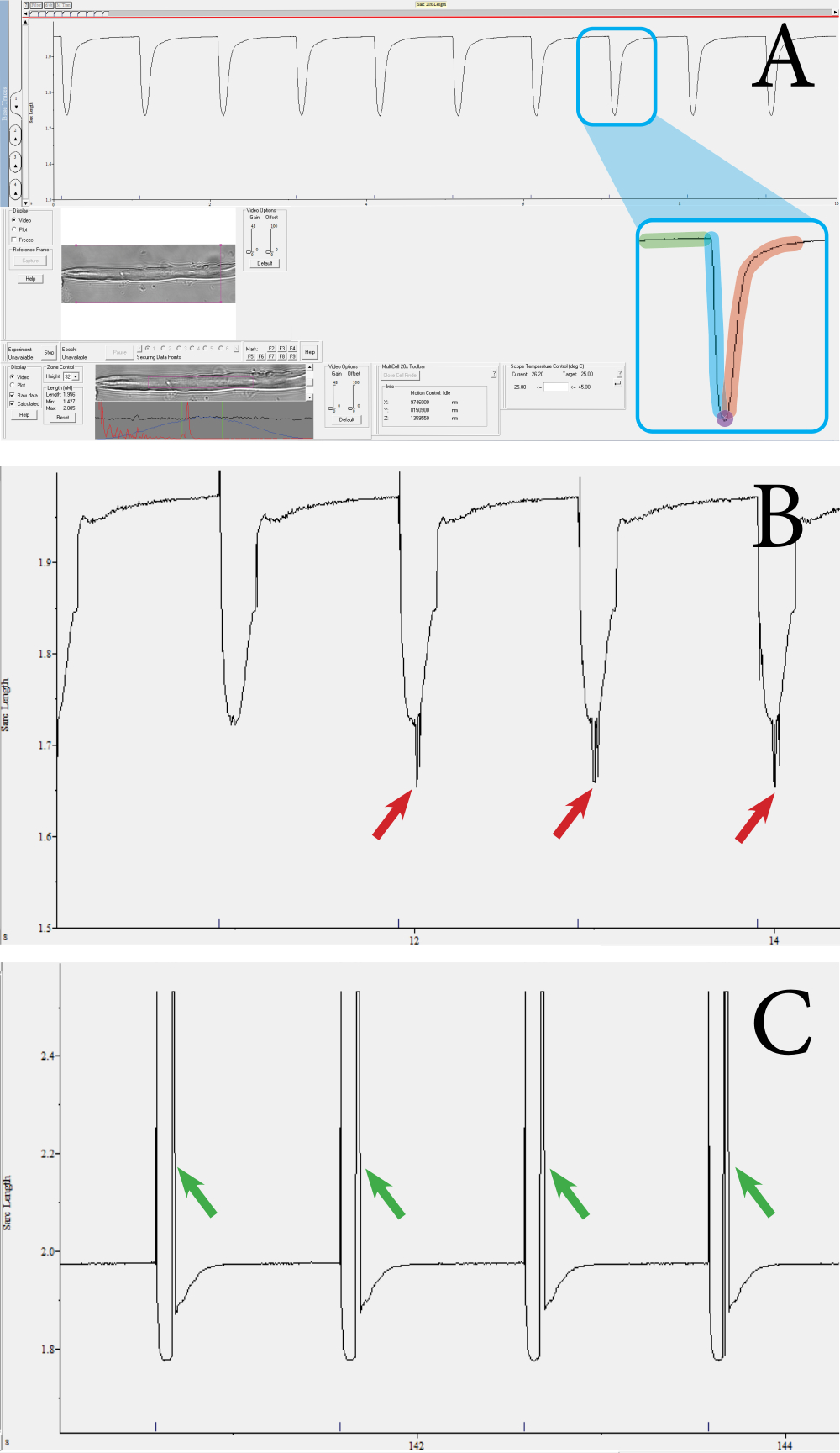{kind=link}

Şekil 4: 2D ile 3D koşulları ve farklı hidrojelleri karşılaştırırken optik tabanlı yüksek verimli kontraktil sistem kullanılarak elde edilen temsili veriler . (A) Veri analiz programı tarafından 2D kültürde ve 30 ölçüme dayanan 3D kültürde bulunan kabul edilen ve reddedilen kasılma ölçümlerinin yüzdesi. (B) 24 saatlik kültürden sonra üç fareden izole edilen 2D kültürlü ve 3D kültürlü kas liflerindeki maksimum kasılma hızının karşılaştırılması. (C) 24 saatlik kültürden sonra üç fareden izole edilen 2D kültürlü ve 3D kültürlü kas liflerinde sarkomer kısalmasının karşılaştırılması. (D) 24 saatlik kültürden sonra üç fareden izole edilen saf bazal membran matrisi (Matrigel) gömülü ve fibrin hidrojel gömülü kas liflerinin maksimum kasılma hızının karşılaştırılması. (E) 24 saatlik kültürden sonra üç fareden izole edilen saf bazal membran matrisi (Matrigel) gömülü ve fibrin hidrojel gömülü kas liflerinin sarkomer kısalmasının karşılaştırılması. Veriler bir Student'ın t-testi kullanılarak analiz edilmiştir ve ortalama ± SD olarak gösterilmiştir. Her veri noktası bir kas lifidir. Bu şeklin daha büyük bir versiyonunu görmek için lütfen buraya tıklayın.
{kind=link}
Tablo 1: Kasın diseksiyonunda kullanılan diseksiyon ortamının, izole kas liflerinin kültürlenmesinde kullanılan fibrin kültür ortamının ve izole kasları enzimatik olarak sindirmek için kullanılan kas sindirim ortamının bileşimi. Bu Tabloyu indirmek için lütfen tıklayınız.
Tablo 2: Hidrojelleri dökmek için kullanılan hücre karışımının bileşimi ve hidrojelleri dökmek için kullanılan matris karışımı. Bu Tabloyu indirmek için lütfen tıklayınız.
Tartışmalar
Burada, FDB kas liflerinin enzimatik izolasyonunu ve kültürünü 3D kültür formatında gerçekleştirmek için bir protokolü detaylandırıyoruz, ardından optik tabanlı bir kontraktil ölçüm sistemi kullanarak kontraktil ölçümler yapıyoruz. Bu protokolün, 1) birçok sağlam kas lifinin tek bir kastan düz ve zamanında izole edilmesi; 2) kas liflerinin ayarlanabilir bir hidrojel matrisine gömülmesi; 3) optik tabanlı sistemi kullanarak yüksek verimli kontraktilite ölçümlerinin performansı; ve 4) bir müdahaleyi takiben aynı kas liflerinin tekrarlanan ölçümlerini yapma yeteneği. Tek canlı kas liflerinin izolasyonu, kasılma fonksiyonlarını koruyan olgun kas hücreleri sağlar. Fare FDB'sinden elde edilen kas lifleri nispeten küçük olduğundan, izolasyon sırasında kolayca manipüle edilir ve düz şekillerini koruyarak aşağı akış kasılma ölçümlerine izin verir. Sistem öncelikle kardiyomiyosit kasılmasını incelemek için geliştirilmiş olmasına rağmen, iskelet kası lifleri kolayca ayırt edilebilen sarkomer desenli aynı kasılma makinesini içerir ve bu nedenle bu sistem29 kullanılarak da ölçülebilir. Tek hücreli kontraktil ölçümlerin canlı ex vivo kas lifi kültürü ile birleştirilmesi, olgun kas lifi sağlığını değerlendirmek ve elektriksel aktivasyona yanıt olarak işlev görmek için güçlü bir araçtır.
Ayrışmış kas liflerinin kullanılmasının bir sınırlaması, liflere uygulanan dış kuvvetlerin (yani pasif gerilme) eksikliğidir, bu da in vivo bulunanlara kıyasla daha düşük dinlenme sarkomer uzunluklarına neden olur. Her ne kadar 2.4-2.5 μm'lik bir sarkomer uzunluğu optimum kuvvet üretimi sağlasa da, dinlenme sarkomer uzunluğu büyük ölçüdedeğişebilir 33. FDB'nin in vivo dinlenme sarkomer uzunluğu henüz tanımlanmamış olsa da, kendi yayınlanmamış verilerimiz ortalama 2.2 μm uzunluğunda olduğunu göstermektedir. Mevcut sonuçlar, kültürde 24 saat sonra yüksüz FDB liflerinde ortalama dinlenme sarkomer uzunluğunun ~ 1.95 μm olduğunu göstermektedir (Şekil 3). Bu düşük dinlenme sarkomer uzunluğu daha düşük kuvvet üretimine neden olsa da, ~ 1.95 μm'lik bir uzunluk hala maksimum kuvvet34'ün% >90'ını üretmelidir. Bu nedenle, bu sarkomer uzunlukları, farklı genetik modeller arasındaki lif fonksiyonundaki farklılıkları belirlemek veya ilaç tedavilerini takip etmek için yeterli olmalıdır. Ek olarak, liflerin bir hidrojel içine gömülmesi, serbest yüzen 2D kültürlü liflere kıyasla yapışma için birçok ek nokta sağlar ve bu da zamanla daha fazla sarkomer kısalmasını sınırlar.
Bu kas lifi izolasyon protokolünün bir avantajı, ekstansör digitorum longus (EDL) gibi diğer kaslara kıyasla nispeten küçük kas liflerinden oluşan, kolayca disseke edilebilen hızlı seğirme kasının kullanılmasıdır. Daha küçük boyutları, kas izolasyonunu tritürasyon bazlı ayırma için daha uygun hale getirir, böylece kas liflerinde pipet veya dolaşmaya bağlı hasar olasılığını azaltır. FDB kaslarının hücre dışı matrisi, kollajenaz ile enzimatik olarak kolayca sindirilebilir ve kısa sürede yüzlerce kas lifinin izolasyonuna izin verir. Bununla birlikte, aşırı sindirim kas liflerine zarar verebilir. Kas liflerinin aşırı sindirimi, kas tritüre edilirken kas neredeyse anında parçalandığında veya hücre tohumlama prosedürü sırasında hücre hacminin büyük bir kısmı hiperkontraksiyona girdiğinde tanınabilir. Kasın aşırı sindirimini önlemek için, sindirim süresinin her kollajenaz partisi için optimize edilmesi gerekir. Bunu test etmek için, iki FDB kası, aralarında 5 dakikalık kademeli bir sindirim süresine paralel olarak sindirilmelidir. Canlı kas liflerinin en yüksek verimine sahip sindirim süresi seçilmelidir. Bu optimizasyon daha sonra ikinci kez, yine sindirim süresinde 5 dakikalık ayırma ile yapılmalıdır. En yüksek canlı kas liflerini veren sindirim süresi, mevcut kollajenaz partisi için en uygun sindirim süresi olarak kullanılmalıdır. Kollajenazın partiden partiye değişkenliğini sınırlamanın bir başka yolu, stok çözeltisinin mililitresi başına aktivite birimlerini doğrudan hesaplamak ve daha sonra sonraki kollajenaz partilerini aynı miktarda yeniden oluşturmaktır. Son olarak, sindirim sürelerinin farklı fare suşları arasında optimize edilmesi gerekebilir, örneğin, artan hücre dışı matris birikimi sergileyen yaşlı veya hastalıklı hayvanları inceliyorsanız35,36.
Canlı izole kas liflerinin pipetlenmesi olasılığı, çeşitli kültür koşullarında FDB kas liflerinin kültürlenmesine olanak tanır. Böyle bir seçenek, doğal doku kültürü ortamını taklit etmek için bu liflerin hidrojellerde kültürlenmesidir. Bu gömme protokolü, kontraktil ölçümler sırasında liflerin yerinde kalmasını sağlar ve jel setlerinden önce liflerin plakanın dibine yerleşmesini sağlamak için optimize edilmiştir. Bununla birlikte, bu protokolün trombin ve fibrinojen stoklarındaki farklılıkları karşılamak için ayarlanması gerekebilir. Trombin aktivitesi çok yüksekse, jel erken yerleşir ve lifler mikroskobun odak düzleminin dışındaki daha yüksek yerlerde asılı kalabilir. Bu olursa, trombin:fibrinojen oranının ayarlanması gerekir. Bu, lifleri giderek daha düşük trombin konsantrasyonlarında kaplayarak ve polimerizasyonun ne kadar sürdüğünü not ederek test edilebilir. Tipik olarak, bu 30 dakikadan daha hızlı gerçekleşmemelidir. Bununla birlikte, çok düşük bir trombin konsantrasyonuna sahip olmak da polimerizasyon işlemini bozabilir. Liflerin doğru odak düzleminde olmasını sağlamak için bir başka yöntem, önce bir 2D protokolü kullanarak tohumlamak ve daha sonra kültür plakasına yapıştıktan sonra liflerin üzerine bir hidrojel tabakası eklemektir. Bununla birlikte, ortamın liflerden çıkarılmasının, kurumaya duyarlı oldukları için hiperkasılmaya neden olabileceğinin farkında olunmalıdır. Hidrojelin kültür plakasına tam olarak yapışıp yapışmayacağı da belirsizdir ve daha kolay gevşeyebilir. Bu nedenle, bu gömme prosedürü, lifleri canlı tutmak ve büzülme ölçümleri için yerinde tutmak için tercih edilir.
Bu protokolün kullanımı, olgun kas liflerinin kasılma dinamiklerinin ex vivo olarak incelenmesini sağlar ve hem sağlıklı farelere hem de kas hastalıkları için genetik mutasyonlar taşıyanlara uygulanabilir. Aynı şekilde, kültür koşullarının veya bileşiklerin eklenmesinin kas lifi fonksiyonunu nasıl etkilediğinin test edilmesini sağlar. Optik tabanlı sistem kullanılarak elde edilen kasılma verileri, canlı tek kas liflerinin kasılma kabiliyetinin bir göstergesidir ve bu yetenekteki değişiklikler lif sağlığı ile ilişkilendirilebilir. Bununla birlikte, bu veriler tek başına, bu değişikliklerin kas kasılmasının aktin-miyozin çapraz köprüleme veya kalsiyum salınım aşamalarında meydana gelip gelmediğini belirlemek için yeterli değildir. Bu protokolde kalsiyum sinyallemesini ölçmek için yöntemler tanımlamasak da, bu kurulum aynı zamanda kas hücrelerinin kasılmasında Fura bazlı kalsiyum geçicilerini de ölçebilir29. Bu sistemin bir dezavantajı, FDB kasının sadece hızlı seğirme tip IIa / IIx kas lifleri içermesidir ve bu boyuttaki yavaş seğirme tip I liflerini içeren kaslar henüz tanımlanmamıştır37. Bu, bu yöntemi kullanarak lif tipine özgü işleyişi inceleme yeteneğini ortadan kaldırır. Burada önerdiğimiz protokol, lif tipi farklılıklarını incelemek için EDL veya soleus gibi diğer kaslar için potansiyel olarak uyarlanabilir. Daha büyük boyutlarından dolayı, bu protokolün bu kaslar için daha da optimize edilmesi gerekecektir. Daha uzun lifler, yerçekimi çökeltme adımı sırasında karışma eğilimindedir ve pipetleme ile manipüle edilirse yırtılır, bu da daha düşük verime yol açar. Bu nedenle, pipetleme ile uyumsuzlukları nedeniyle, daha uzun lifler jel gömme tekniğiyle de daha az uyumludur. Bu liflerin ölçümleri hala 2D kültür formatında yapılabilir, ancak fiberler boyutlarından dolayı büzülme sırasında daha fazla hareket edebilir ve böylece sinyal-gürültü oranını etkileyebilir. Bu sistemin bir diğer sınırlaması, diğer sağlam kas lifi preparatları kullanılarak elde edilebilen kuvvet ölçümleri gibi kasılma ölçümlerinin yanı sıra kuvvet ölçümlerinin de yapılamamasıdır28. Bununla birlikte, bu sınırlama, kas lifi tarafından üretilen kuvveti tahmin ederek aşılabilir. Kas liflerinin üretilen kuvveti, kasılmış ve gevşemiş durumlar sırasında kas lifi şeklinin yanı sıra matrisin Young modülünün25 olarak bilinmesi durumunda tahmin edilebilir. Bununla birlikte, bu optik tabanlı sistem, kas kasılma fonksiyonunu incelemek için kullanımı kolay, yüksek verimli bir yaklaşım sağlar ve genetik kas hastalıklarını ve terapötik müdahaleleri incelemek için bir dizi yeni olasılık açar.
Açıklamalar
Yazarların beyan edecekleri çıkar çatışmaları yoktur.
Teşekkürler
Yazarlar, Sylvia Bogaards, Sanna Luijcx, Valentijn Jansen, Michiel Helmes ve Emmy Manders'e bu protokolün geliştirilmesine yardımcı olan teknik uzmanlıkları için teşekkür eder. Bu çalışma, Kas Distrofisi Derneği (T.J.K'ya MDA603238 Gelişim Ödülü), Hollanda Kardiyovasküler İttifakı (T.J.K'ya Yetenek Bursu) ve Ulusal Sağlık ve Tıbbi Araştırma Konseyi (NHMRC, Avustralya; Burs APP1121651 M.Y.'ye).
Malzemeler
| Name | Company | Catalog Number | Comments |
| Aprotinin, from Bovine Lung | Thermo Scientific | AAJ63039MC | 100 mM stock solution in PBS can be stored at -20 °C. Sterilize stock solution using a 0.22 µm filter. |
| Collagenase type 2 | Worthington | 77336 | 10% (w/v) stock solution can be stored at -20 °C. Weighing collagenase should be done in a safety cabinet as inhalation is dangerous. |
| Fetal Bovine Serum | Thermo Fisher | 10500064 | |
| Fibrinogen from Bovine Plasma | Sigma Aldrich | 50-176-5054 | 20 mg/mL stock solution in PBS can be stored at -80 °C. Sterilize stock solution using a 0.22 µm filter. |
| Geltrex LDEV-Free Reduced Growth Factor Basement Membrane Matrix | Thermo Fisher | A1413201 | 4 mg/mL stock solution is prepared in MEM and stored at -20 °C. |
| Gibco MEM High glucose + pyruvate | Thermo Fisher | 11095080 | |
| Horse serum | Thermo Fisher | H1270 | |
| Matrigel GFR Membrane Matrix | Corning | CB-40230 | 4 mg/mL stock solution is prepared in MEM and stored at -20 °C. |
| Penicillin/Streptomycin | Sigma Aldrich | P4333 | |
| Serum Replacement 2 (50x) | Sigma Aldrich | S9388 | |
| Thrombin, Bovine Plasma | Thermo Scientific | AAJ63383EXP | 125 U/mL stock in PBS can be stored at -20 °C. Sterilize stock solution using a 0.22 µm filter. |
| Tranexamic Acid | Thermo Scientific | AC228042500 | 80 mM stock solution in PBS can be stored at -20 °C. Sterilize stock solution using a 0.22 µm filter. |
| Equipment | |||
| 24-well electrical stimulator | IonOptix | N/a | |
| Dumont #55 Forceps | Fine Science Tools | 11295-51 | |
| Extra Fine Bonn Scissors | Fine Science Tools | 14084-08 | |
| MultiCell Cytocypher | IonOptix | N/a | |
| MyoCam-S3 | IonOptix | N/a | |
| MyoPacer | IonOptix | N/a | |
| SYLGARD 184 silicone elastomer, Base & Curing Agent | Dow corning | N/a | |
| Vannas Spring Scissor - 25 mm Cutting Edge | Fine Science Tools | 15002-08 | |
| Software | |||
| CytoSolver | IonOptix | N/a | |
| IonWizard | IonOptix | N/a |
Referanslar
- Smith, L. R., Meyer, G. A. Skeletal muscle explants: Ex-vivo models to study cellular behavior in a complex tissue environment. Connective Tissue Research. 61 (3-4), 248-261 (2020).
- Khodabukus, A., Prabhu, N., Wang, J., Bursac, N. In vitro tissue-engineered skeletal muscle models for studying muscle physiology and disease. Advanced Healthcare Materials. 7 (15), 1701498 (2018).
- Fernandez-Costa, J. M., Fernandez-Garibay, X., Velasco-Mallorqui, F., Ramon-Azcon, J. Bioengineered in vitro skeletal muscles as new tools for muscular dystrophies preclinical studies. Journal of Tissue Engineering. 12, 2041731420981339 (2021).
- Romagnoli, C., Iantomasi, T., Brandi, M. L. Available in vitro models for human satellite cells from skeletal muscle. International Journal of Molecular Sciences. 22 (24), 13221 (2021).
- Dessauge, F., Schleder, C., Perruchot, M. -. H., Rouger, K. 3D in vitro models of skeletal muscle: Myopshere, myobundle and bioprinted muscle construct. Veterinary Research. 52 (1), 72 (2021).
- Hosoyama, T., Meyer, M. G., Krakora, D., Suzuki, M. Isolation and in vitro propagation of human skeletal muscle progenitor cells from fetal muscle. Cell Biology International. 37 (2), 191-196 (2013).
- Guo, X., et al. In vitro differentiation of functional human skeletal myotubes in a defined system. Biomaterials Science. 2 (1), 131-138 (2014).
- Denes, L. T., et al. Culturing C2C12 myotubes on micromolded gelatin hydrogels accelerates myotube maturation. Skeletal Muscle. 9 (1), 17 (2019).
- Pimentel, M. R., Falcone, S., Cadot, B., Gomes, E. R. In vitro differentiation of mature myofibers for live imaging. Journal of Visualized Experiments. (119), e55141 (2017).
- Khodabukus, A. Tissue-engineered skeletal muscle models to study muscle function, plasticity, and disease. Frontiers in Physiology. 12, 619710 (2021).
- Engler, A. J., et al. Myotubes differentiate optimally on substrates with tissue-like stiffness: Pathological implications for soft or stiff microenvironments. Journal of Cell Biology. 166 (6), 877-887 (2004).
- Huang, N. F., et al. Myotube assembly on nanofibrous and micropatterned polymers. Nano Letters. 6 (3), 537-542 (2006).
- Earle, A. J., et al. Mutant lamins cause nuclear envelope rupture and DNA damage in skeletal muscle cells. Nature Materials. 19 (4), 464-473 (2020).
- Stange, K., Ahrens, H. E., von Maltzahn, J., Rontgen, M. Isolation and ex vivo cultivation of single myofibers from porcine muscle. In Vitro Cellular and Developmental Biology. Animal. 56 (8), 585-592 (2020).
- Ravenscroft, G., et al. Dissociated flexor digitorum brevis myofiber culture system--A more mature muscle culture system. Cell Motility and the Cytoskeleton. 64 (10), 727-738 (2007).
- Ramsey, R. W., Street, S. F. The isometric length-tension diagram of isolated skeletal muscle fibers of the frog. Journal of Cellular and Comparative Physiology. 15 (1), 11-34 (1940).
- Selvin, D., Hesse, E., Renaud, J. M. Properties of single FDB fibers following a collagenase digestion for studying contractility, fatigue, and pCa-sarcomere shortening relationship. American Journal of Physiology-Regulatory, Integrative and Comparative Physiology. 308 (6), R467-R479 (2015).
- Renzini, A., et al. Culture conditions influence satellite cell activation and survival of single myofibers. European Journal of Translational Myology. 28 (2), 7567 (2018).
- Pasut, A., Jones, A. E., Rudnicki, M. A. Isolation and culture of individual myofibers and their satellite cells from adult skeletal muscle. Journal of Visualized Experiments. (73), e50074 (2013).
- Holmberg, J., Durbeej, M. Laminin-211 in skeletal muscle function. Cell Adhesion and Migration. 7 (1), 111-121 (2013).
- Stuelsatz, P., Keire, P., Yablonka-Reuveni, Z. Isolation, culture, and immunostaining of skeletal muscle myofibers from wildtype and nestin-GFP mice as a means to analyze satellite cell. Methods in Molecular Biology. 1556, 51-102 (2017).
- Alkhateeb, H., Chabowski, A., Bonen, A. Viability of the isolated soleus muscle during long-term incubation. Applied Physiology, Nutrition, and Metabolism. 31 (4), 467-476 (2006).
- Roche, S. M., Gumucio, J. P., Brooks, S. V., Mendias, C. L., Claflin, D. R. Measurement of maximum isometric force generated by permeabilized skeletal muscle fibers. Journal of Visualized Experiments. (100), e52695 (2015).
- Ottenheijm, C. A., et al. Altered myofilament function depresses force generation in patients with nebulin-based nemaline myopathy (NEM2). Journal of Structural Biology. 170 (2), 334-343 (2010).
- Rausch, M., et al. Measurement of skeletal muscle fiber contractility with high-speed traction microscopy. Biophysical Journal. 118 (3), 657-666 (2020).
- de Winter, J. M., et al. KBTBD13 is an actin-binding protein that modulates muscle kinetics. Journal of Clinical Investigation. 130 (2), 754-767 (2020).
- Wijnker, P. J. M., vander Velden, J. Mutation-specific pathology and treatment of hypertrophic cardiomyopathy in patients, mouse models and human engineered heart tissue. Biochimica et Biophysica Acta. Molecular Basis of Disease. 1866 (8), 165774 (2020).
- Cheng, A. J., Westerblad, H. Mechanical isolation, and measurement of force and myoplasmic free [Ca(2+)] in fully intact single skeletal muscle fibers. Nature Protocols. 12 (9), 1763-1776 (2017).
- Cao, L., Manders, E., Helmes, M. Automatic detection of adult cardiomyocyte for high throughput measurements of calcium and contractility. PLoS One. 16 (9), e0256713 (2021).
- Geckil, H., Xu, F., Zhang, X., Moon, S., Demirci, U. Engineering hydrogels as extracellular matrix mimics. Nanomedicine. 5 (3), 469-484 (2010).
- Lin, C. C., Anseth, K. S. PEG hydrogels for the controlled release of biomolecules in regenerative medicine. Pharmaceutical Research. 26 (3), 631-643 (2009).
- Cold Spring Harbor Protocols. Sylgard-coated coverslips and petri dishes. Cold Spring Harbor Protocols. 2022 (8), (2022).
- Moo, E. K., Fortuna, R., Sibole, S. C., Abusara, Z., Herzog, W. In vivo sarcomere lengths and sarcomere elongations are not uniform across an intact muscle. Frontiers in Physiology. 7, 187 (2016).
- Moo, E. K., Leonard, T. R., Herzog, W. The sarcomere force-length relationship in an intact muscle-tendon unit. Journal of Experimental Biology. 223, 215020 (2020).
- Schuler, S. C., et al. Extensive remodeling of the extracellular matrix during aging contributes to age-dependent impairments of muscle stem cell functionality. Cell Reports. 35 (10), 109223 (2021).
- Carberry, S., Zweyer, M., Swandulla, D., Ohlendieck, K. Proteomics reveals drastic increase of extracellular matrix proteins collagen and dermatopontin in the aged mdx diaphragm model of Duchenne muscular dystrophy. International Journal of Molecular Medicine. 30 (2), 229-234 (2012).
- Tarpey, M. D., et al. Characterization and utilization of the flexor digitorum brevis for assessing skeletal muscle function. Skeletal Muscle. 8 (1), 14 (2018).
Yeniden Basımlar ve İzinler
Bu JoVE makalesinin metnini veya resimlerini yeniden kullanma izni talebi
Izin talebiThis article has been published
Video Coming Soon
JoVE Hakkında
Telif Hakkı © 2020 MyJove Corporation. Tüm hakları saklıdır