Method Article
تنقية الجينوم على نطاق خارج الصبغي التعميم الحمض النووي من خلايا حقيقية النواة
In This Article
Summary
This paper presents a sensitive method called Circle-Seq for purifying extrachromosomal circular DNA (eccDNA). The method encompasses column purification, removal of remaining linear chromosomal DNA, rolling-circle amplification and high-throughput sequencing. Circle-Seq is applicable to genome-scale screening of eukaryotic eccDNA and studying genome instability and copy-number variation.
Abstract
السلطات الوطنية المعينة دائرية خارج الصبغي (eccDNAs) هي عناصر جينية مشتركة في خميرة الخباز وذكرت في حقيقيات النوى الأخرى كذلك. المساهمة EccDNAs إلى الاختلاف الجيني بين الخلايا الجسدية في الكائنات متعددة الخلايا وتطور حقيقيات النوى وحيدة الخلية. وهناك حاجة إلى أساليب حساسة للكشف عن eccDNA لتوضيح كيفية تأثير هذه العناصر استقرار الجينوم وكيف العوامل البيئية والبيولوجية تحفز تكوين في الخلايا حقيقية النواة. يعرض هذا الفيديو طريقة eccDNA تنقية حساسة تسمى دائرة تسلسل. يشمل طريقة تنقية عمود من الحمض النووي الدائري، وإزالة ما تبقى من الحمض النووي الخطي الكروموسومات، الدائرة المتداول التضخيم من eccDNA، تسلسل عميق، ورسم الخرائط. كان مطلوبا العلاج نوكلياز خارجية واسعة لكافية الخطي تدهور الكروموسومات الحمض النووي. الخطوة المتداول دائرة التضخيم من قبلالمعهد الوطني للإحصاء الطول: عادي؛ "> φ 29 البلمرة المخصب لالحمض النووي DNA دائري فوق الخطي التحقق من طريقة الدائرة بعدها على ثلاثة سكان البيرة س CEN.PK من 10 10 خلايا الكشف عن مئات التشكيلات eccDNA في أحجام أكبر من 1 كيلو قاعدة . النتائج المتكررة من ASP3-1، COS111، CUP1، RSC30، HXT6، HXT7 الجينات في الحمض النووي التعميم في كل S288c وتقترح CEN.PK أن التعميم الحمض النووي وحفظها بين السلالات في هذه المكاني. وباختصار، فإن طريقة الدائرة يليها ديه واسعة تطبيق لفحص الجينوم على نطاق ولeccDNA في حقيقيات النوى وكذلك للكشف عن أنواع eccDNA محددة.
Introduction
كشف المبكر أو عابرة التضخيم الكروموسومات من الصعب لأنه يتطلب تحديد التعديلات في جزيئات الحمض النووي احدة في أعداد كبيرة من الخلايا. تم الكشف عن الكروموسومات الاختلافات نسخ رقم (التنوعات) بشكل جيد عموما بعد إنشائها، ولم يتبق سوى هيكل CNV النهائي كدليل على الآلية التي ولدت الاختلاف 1،2. كشف واستعادة الحمض النووي خارج الصبغية دائري (eccDNA) في المراحل السابقة من تشكيل CNV قد توضيح العمليات الجارية في إعادة ترتيب الجينوم.
سابقا، كان دي نوفو اكتشاف eccDNA بواسطة الميكروسكوب الإلكتروني 3، تلطيخ بالغيمزا من الكروموسومات الطورية 4، أو ثنائي الأبعاد هلام الكهربائي 5. وتوفر هذه الأساليب معلومات قليلة أو معدومة عن تسلسل الحمض النووي دائري. تقنيات المستهدفة مثل جنوب النشاف 6،7، معكوس PCR 8، أو مضان في hybridizatio الموقعن 9 تقديم أدلة فقط عن عناصر eccDNA محددة. أيا من هذه الأساليب توفير سلسلة من جميع أنواع eccDNA الموجودة في السكان الخلية.
الاختلاف الجيني في مجموعة من الخلايا يمكن أن تتسم تسلسل الجينوم و / أو صفائف تبليط 10،11. الكشف عن الحذف أو التضخيم من قبل وسائل تنقية الحمض النووي التقليدية عادة ما يتطلب أن أليل تحور تمثل 0،1-1٪ على الأقل من سكان الخلية 12،13. ومن المتوقع أن تكون أكثر عابرة في زراعة الخلايا لامركزي eccDNAs بسبب افتقارها إلى القسيم وغياب محتمل لتوليف الحمض النووي في النسخ المتماثل. وهكذا، منذ أكثر eccDNAs ويفترض في كميات قليلة وتسلسل بهم تشبه الجينوم، وهناك حاجة إلى طرق استخراج الحمض النووي بديلة للكشف عن eccDNAs.
تقنيات عدة لتنقية الحمض النووي دائرية استغلال الاختلافات البنيوية بين الكروموسومات والحمض النووي دائري. على سبيل المثال، عالية السرعة ultracentrifugيستخدم أوجه في التدرجات السيزيوم كلوريد لعزل 350-3000 basepairs (بي بي) eccDNAs كبير من البشرية السرطان هيلا خط الخلية 14. ومع ذلك، يمكن عالية السرعة كسر أو شق العمود الفقري للبنية الحمض النووي دائرية supercoiled، تغيير سرعة الترسيب (15) والعائد eccDNA. وضعت دوتا وزملاء العمل وسيلة لمن جديد، وتحديد الجينوم على نطاق من الحمض النووي دائري من الأنسجة الماوس وكذلك من ثقافات الدجاج والخلايا البشرية 16،17. منهجهم هو استخراج النواة من النسيج المتجانس من قبل تنبيذ فائق السكروز تليها تنقية البلازميد وعدة جولات من التفاعلات الإنزيمية والاستخراج الحمض النووي. بروتوكول بهم ويحدد في المقام الأول 200-400 eccDNAs بي بي، ودعا microDNAs. كما حاول دوتا وزملاء العمل تنقية microDNAs من خميرة الخباز ولكن لم يتمكنوا من تسجيل microDNA من هذا أنواع من الخميرة 16.
لقد قمنا بتطوير طريقة جديدة لدي نوفو الكشف عن eccDNA من الخميرة تسمى دائرة تسلسل. تمكن هذه الطريقة استطلاعات الجينوم على نطاق ولجزيئات الحمض النووي دائرية كبيرة بما يكفي لتحمل جينات كاملة وكبيرة مثل 86 كيلو قاعدة (كيلوبايت) الحمض النووي (و mtDNA). تم تطوير طريقة الدائرة تسلسل من البلازميد في بدائيات النواة راسخة طريقة تنقية 18،19 الأمثل لخلايا الخميرة حقيقية النواة وجنبا إلى جنب مع تسلسل عميق. باستخدام نهج الدائرة وما يليها، 1756 eccDNAs مختلفة، كل كيلوبايت أكبر من 1، تم الكشف عن عشر س. الخباز S288c ظل السكان 20. وقد تم اختيار حجم قطع للتركيز على eccDNA التي كانت كبيرة بما يكفي لتحمل جينات بأكملها. كانت دائرة تسلسل حساسة للغاية. ذلك الكشف عن eccDNA احد ضمن آلاف الخلايا 20. في الدراسة الحالية، استخدمت الدائرة تسلسل لعزل وتحديد 294 eccDNAs من ثلاثة مكررات البيولوجية للS. آخر الخباز الخميرة سلالة، CEN.PK. يكشف عن البيانات التي eccDNA هو إيليمي الجيني المشتركالإقليم الشمالي في س. سلالات الخباز.
Protocol
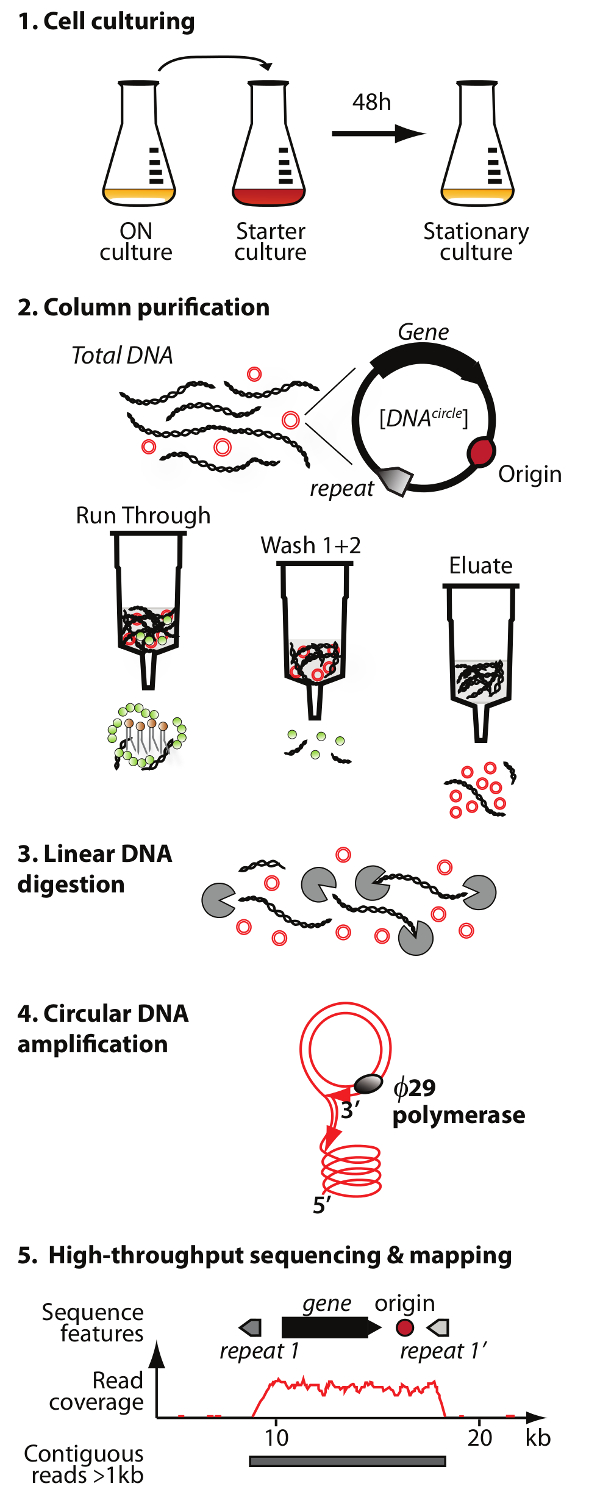
ملاحظة: لمحة عامة عن تنقية الحمض النووي دائرية وطريقة التسلسل (دائرة تسلسل) هو موضح في الشكل 1.
1. زراعة، خلية الحصاد والبلازما غشاء تعطيل
- تطعيم خلايا الخميرة (على سبيل المثال خميرة الخباز) من O / N ثقافة إلى 50 مل وسط غذائي كامل من سكر العنب الخميرة ببتون (YPD). تطعيم في انخفاض كثافة الخلايا الأولية من 1-3 × 10 5 خلية / مل أو الكثافة البصرية ما يقرب من 0.01 OD 600.
- احتضان الخلايا عند 30 درجة مئوية مع الإثارة في 150 طلقة في الدقيقة (دورة في الدقيقة) حتى خلايا وصول إلى أقصى كثافة الخلية حوالي 1 × 10 10 خلايا، بعد حوالي 24 إلى 48 ساعة أو الكثافة البصرية في OD 600> 10.0.
ملاحظة: الساعة زراعة ليس حاسما كما تركيزات الخلية السفلية يمكن استخدامها.
- احتضان الخلايا عند 30 درجة مئوية مع الإثارة في 150 طلقة في الدقيقة (دورة في الدقيقة) حتى خلايا وصول إلى أقصى كثافة الخلية حوالي 1 × 10 10 خلايا، بعد حوالي 24 إلى 48 ساعة أو الكثافة البصرية في OD 600> 10.0.
- نقل ثقافة تجاوزت إلى أنبوب مخروطي 50 مل، بيليه الخلايا بواسطة الطرد المركزي في800 x ج لمدة 3 دقائق والتخلص من طاف.
- غسل بيليه مع 25 مل من محلول عازلة من 10 ملي تريس الكلورين، 1 ملم EDTA، ودرجة الحموضة 8.0، وإعادة بيليه الخلايا بواسطة الطرد المركزي في 800 x ج لمدة 3 دقائق والتخلص من طاف.
- resuspend الكرية خلية في 1.2 مل العازلة إعادة تعليق الموردة من مجموعة البلازميد العمود تنقية.
- خطوة اختيارية: قم بإضافة البلازميدات مخففة للغاية والضوابط لتنقية عناصر الحمض النووي دائرية 20.
ملاحظة: في ورقة العمل الحالي، تم تطبيق 7.7 ميكرولتر البلازميد خليط لكل عينة تحتوي على 10 10 الخلايا. خليط الأسهم البلازميد يتألف من ثلاثة البلازميدات في تركيزات مختلفة؛ pBR322 في 38 نانوغرام / عينة، pUC19 عند 0.5 نانوغرام / عينة، وpUG72 عند 0.01 نانوغرام / عينة. - نقل تعليق الخلية إلى قسمين 2 مل أنابيب الطرد المركزي الصغيرة، كل تستكمل مع الخرز الزجاجي 0.5 ملم في نسبة 1: 3 من إجمالي حجم تعليق.
- دوامة كل أنبوب في أقصى سرعة لمدة 10 دقيقة إلى تعطيل خلية بلازماالأغشية. بيليه الخرز بواسطة الطرد المركزي في 268 x ج لمدة 30 ثانية، ونقل 1.2 مل طاف جنبا إلى جنب من أنابيب microcentrifuge اثنين إلى أنبوب جديد.
ملاحظة: البديل إلى الخطوة 1،6-1،7، استخدم zymolyase لتعطيل الخلايا في 0.6 مل إعادة تعليق الحل العازلة. عشر وحدات من zymolyase يمكن أن يحدث خللا 5 × 10 7 خلايا خلال 1.5 ساعة عند 35 درجة مئوية.
2. EccDNA التخصيب بواسطة اللوني العمود
- متابعة بروتوكول من مجموعة لتنقية عمود من البلازميدات. وباختصار، علاج كل عينة مع 1.2 مل من محلول قلوي، مزيج بلطف واحتضان 3 دقائق على RT.
- إضافة 1.2 مل العازلة تحييد المزيج بلطف وأجهزة الطرد المركزي في 9650 x ج لمدة 5 دقائق.
- تحميل الحل على عمود معايرتها مع حل 1 مل موازنة وتسمح للسائل تتدفق من خلال العمود عن طريق الجاذبية.
- غسل العمود مع حل غسل 4 مل. عندما اجتاز حل من خلال الراتنج، إضافة بعناية 0.3 مل شطف ذلكlution ليحل محل معظم 0.35 مل العمود حجم باطلة.
- أزل الحمض النووي في أنبوب مجموعة جديدة مع الحل 1 مل شطف وترسيب الحمض النووي عن طريق إضافة 0.8 مل هطول الخليط. أجهزة الطرد المركزي في 9650 x ج لمدة 10 دقيقة.
- غسل بيليه الحمض النووي مع 0.5 مل 70٪ من الإيثانول، الطرد المركزي في 9650 x ج لمدة 5 دقائق، والهواء الجاف لمدة 5 إلى 15 دقيقة وحل الحمض النووي المنقى في 25 ميكرولتر الماء المعقم.
ملاحظة: فقط تخزين على المدى القصير من الحمض النووي في الماء الموصى بها. تفضيلي، الانتقال مباشرة إلى الخطوة 3.
3. هضم باقي الخطي الصبغية الحمض النووي
- خطوة اختيارية: تيسير الهضم معين من الحمض النووي الخطي نوكلياز خارجية، علاج الحمض النووي المنقى مع نوكلياز داخلية قطع نادرة مثل نوتي. لمدة 5 ميكروغرام الحمض النووي، واستخدام 1 وحدة نوتي، 5 ميكرولتر العازلة 10X الهضم والماء المعقم إلى وحدة تخزين ما مجموعه 50 ميكرولتر. احتضان رد الفعل عند 37 درجة مئوية لمدة 16 ساعة والحرارة تعطيل للنوكلياز داخلية في 80 درجة مئوية لمدة 5 دقائق.
- إضافة 20وحدات نوكلياز خارجية (2 ميكرولتر)، 4 ميكرولتر ATP (25 ملم)، 34 ميكرولتر الماء المعقم و 10 ميكرولتر 10X العازلة رد فعل مباشرة إلى 50 ميكرولتر الحمض النووي المشقوق نوكلياز داخلية للوصول إلى حجم رد الفعل 1X 100 ميكرولتر، وذلك باستخدام نوكلياز خارجية ATP التي تعتمد على عدة.
- أداء التحلل من الحمض النووي المفرد الذين تقطعت بهم السبل وتقطعت بهم السبل المزدوج الخطي عند 37 درجة مئوية لمدة 5 أيام أو أكثر. إضافة إضافي 4 ميكرولتر ATP (25 ملم)، و 0.6 ميكرولتر 10X العازلة رد فعل و20 وحدة نوكلياز خارجية كل 24 ساعة لمواصلة عملية الهضم الأنزيمي الحمض النووي في حجم رد الفعل 1X.
- بعد إزالة الحمض النووي الخطي، عينة 2 ميكرولتر من محلول نوكلياز خارجية المعالجة للتأكد من القضاء على الكروموسومات الحمض النووي الخطي الكمي تفاعل البلمرة المتسلسل (QPCR)، وذلك باستخدام علامة الكروموسومات مثل الأكتين الجينات ACT1 20.
- يحتوي كل 20 ميكرولتر QPCR حجم رد الفعل 2 ميكرولتر المعالجة نوكلياز خارجية عينة، 150 نانومتر الاشعال ACT1 5'-TCCGTCTGGATTGGTGGTTCTA-3 "و 5'-TGGACCACTTTCGTCGTATTC-3، 2٪ (حجم / حجم) ثنائي ميثيل سلفوكسيد، و 10 ميكرولتر الأخضر مزيج الرئيسي الفلورسنت.
- استخدام حالة التفاعل؛ 3 دقائق عند درجة حرارة 95 درجة مئوية، تليها 45 دورات من 15 ثانية في 95 درجة مئوية و 30 ثانية في 60 درجة مئوية.
ملاحظة: ACT1 هو علامة مناسبة خاصة لالحمض النووي الخطي منذ الاختلافات في عدد النسخ في هذا الجين هي ضارة 21-23 لذلك ينبغي eccDNA لا تحمل ACT1. - بدائل لتحليل الحمض النووي عن طريق الهضم QPCR هي PCR القياسية (4.3) أو تلطيخ يوديد propidium (4.4).
- استخدام 2 ميكرولتر عينة المعالجة نوكلياز خارجية كقالب PCR مع الاشعال ACT1 5'-TGGATTCTGGTATGTTCTAGC-3 "و 5'-GAACGACGTGAGTAACACC-3". كما تحكم ACT1 إيجابية، استخدم 50-100 نانوغرام الجيني س. الحمض النووي الخباز كقالب. ظروف التفاعل PCR. 3 دقائق عند درجة حرارة 95 درجة مئوية، تليها 35 دورات من 30 ثانية في 95 درجة مئوية، و 30 ثانية في 56 درجة مئوية و 1 دقيقة عند 72 درجة مئوية.
- ردود الفعل تشغيل PCR بواسطة electrophore هلامجهاز الأمن والمخابرات على 1٪ الاغاروز مع 0.5 ميكروغرام / مل إيثيديوم بروميد. ابحث عن الفرقة ACT1 0.8 كيلوبايت.
- غياب أو وجود الحمض النووي الخطي يمكن أيضا أن يفحصه يوديد propidium تلطيخ قبل وبعد التضخيم الحمض النووي.
- خلط كل عينة من الحمض النووي في 1: حجم 1 مع 1: 1000 H 2 O حل المخفف من 20 ملي الأسهم يوديد propidium. ترك الحل في الظلام لمدة 10-20 دقيقة في RT وتحليل الحمض النووي تلطيخ بواسطة المجهر مضان في التكبير 100X باستخدام الإثارة مرشح مضان أحمر في 663-738 نانومتر والوقت تعرض 5-30 ثانية. كما تحكم الحمض النووي تلطيخ، واستخدام ø29 تضخيم الحمض النووي الجيني من الخميرة و / أو البلازميد تضخيم ø29.
- الحرارة تعطيل الحل نوكلياز خارجية عند 70 درجة مئوية لمدة 30 دقيقة.
4. الحمض النووي التضخيم
- تضخيم تنقيته وإثراء eccDNA من الخطوة 3.5) مع ø29 البلمرة DNA 24-26 اتفاقجي لبروتوكول الشركة المصنعة البلمرة.
- وباختصار، مزيج 5 ميكرولتر المخصب eccDNA مع العازلة تمسخ 5 ميكرولتر.
- بعد 3 دقائق على RT، إضافة العازلة تحييد 10 ميكرولتر. المزيج بلطف وإضافة مزيج الرئيسي 30 ميكرولتر تحتوي على 29 ميكرولتر رد فعل العازلة و 1 ميكرولتر ø29 البلمرة DNA. احتضان رد الفعل عند 30 درجة مئوية لمدة 16 ساعة أو أكثر (تصل إلى 72 ساعة). الحرارة تعطيل بوليميريز ø29 الحمض النووي عند 65 درجة مئوية لمدة 3 دقائق.
5. تسلسل وتحليل البيانات
- قص eccDNA تضخيم مع ultrasonicator تركيزا على متوسط ذروة الهدف حجم 300 نقطة أساس. استخدام الإعدادات التالية لعينة الحمض النووي ميكرولتر 130: قوة 450W ذروة كثافة، 60 ثانية العلاج، و 30٪ عامل واجب، 200 دورة في انفجار، ودرجة الحرارة 7 ° C.
- إضافة الباركود تسميات مؤشر ومحولات ليقرأ مجزأة لتوليف مكتبات لتسلسل، وذلك باستخدام طريقة مناسبة لإعداد مكتبة.
- تشغيل تسلسل عميق، على سبيل المثال كما يقرأ من 141 النووية المنفردة نهاية على منصة التسلسل الإنتاجية العالية.
- خريطة يقرأ في الجينوم الخميرة إشارة قيد التحقيق والسماح يقرأ لتعيين مناطق متعددة. على سبيل المثال، استخدام نظام سير العمل متاحة بحرية 27،28 وقصيرة للقراءة برنامج رسم الخرائط اليجنر 29.
- تحديد يقرأ من مناطق eccDNAs المفترضة باستخدام متجاورة يقرأ، على سبيل المثال، يقرأ أكثر من سبعة متجاورة (> 1 كيلوبايت) دون ثغرات 20.
ملاحظة: البرنامج هو متاح 27،28 لاستكشاف معين يقرأ في مناطق الجينوم من الفائدة.
النتائج
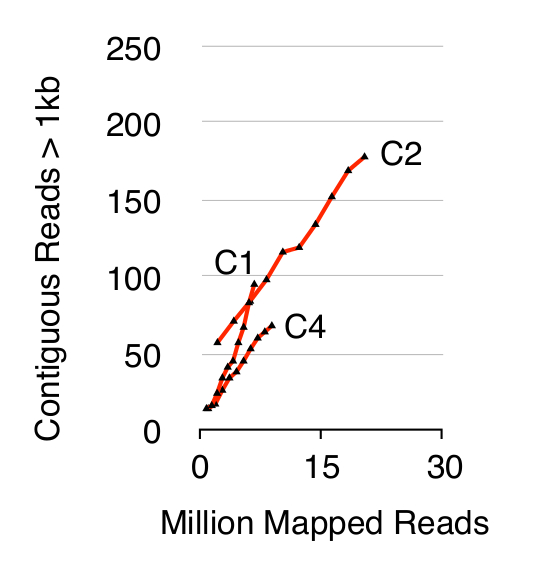
للتحقق من صحة طريقة الدائرة تسلسل، ثلاثة س. تم فحص السكان الخباز CEN.PK من 1 × 10 10 الخلايا بعد ما نمت الخلايا على حدة في YPD لعشرة أجيال. وأكد الكروموسومات القضاء الحمض النووي الخطي بعدم وجود إشارة QPCR ACT1 كما هو موضح سابقا 20 (لا تظهر البيانات). كان التسلسل تنقيته وإثراء eccDNA ما يصل الى 68 مليون يقرأ (141 النوكليوتيدات واحدة في نهاية يقرأ) وتعيينها إلى الجينوم إشارة CEN.PK113-7D (الإصدار 19 يونيو 2012). تم تعيين تسجيلات eccDNAs المفترضة من العينات الثلاث التي ذكرت C1، C2 و C4 إلى مناطق الجينوم التي رسمها متجاورة يقرأ أطول من 1 كيلو بايت. واستنادا إلى 10،000 محاكاة مونت كارلو، وأهمية كل منطقة تعيينها من قبل متجاورة يقرأ أطول من قدرت 1 كيلو بايت. من كان المشروح هذا 79 و 159 و 56 مناطق ما يحتمل تسلسل eccDNA (ص <0.1، الإدراجات 1). عدد contiguo المسجلةلنا يقرأ ارتفعت> 1 كيلوبايت بوصفها وظيفة من تسلسل عمق مما يشير إلى أن المزيد من العناصر eccDNA قد تم تسجيل إذا كان التسلسل عينات أخرى (الشكل 2). كما هو متوقع، استخراج طريقة الدائرة تسلسل العديد يقرأ من عدد من العناصر المعروفة دائرية الحمض النووي بما في ذلك البلازميد 2μ، الحمض النووي والجينات الحمض النووي الريبي الريباسي على الكروموسوم الثاني عشر، والبلازميدات الرقابة الداخلية ثلاثة pBR322، pUC19 وpUG72 التي ارتفعت إلى عينات قبل تنقية عمود (الشكل 3).
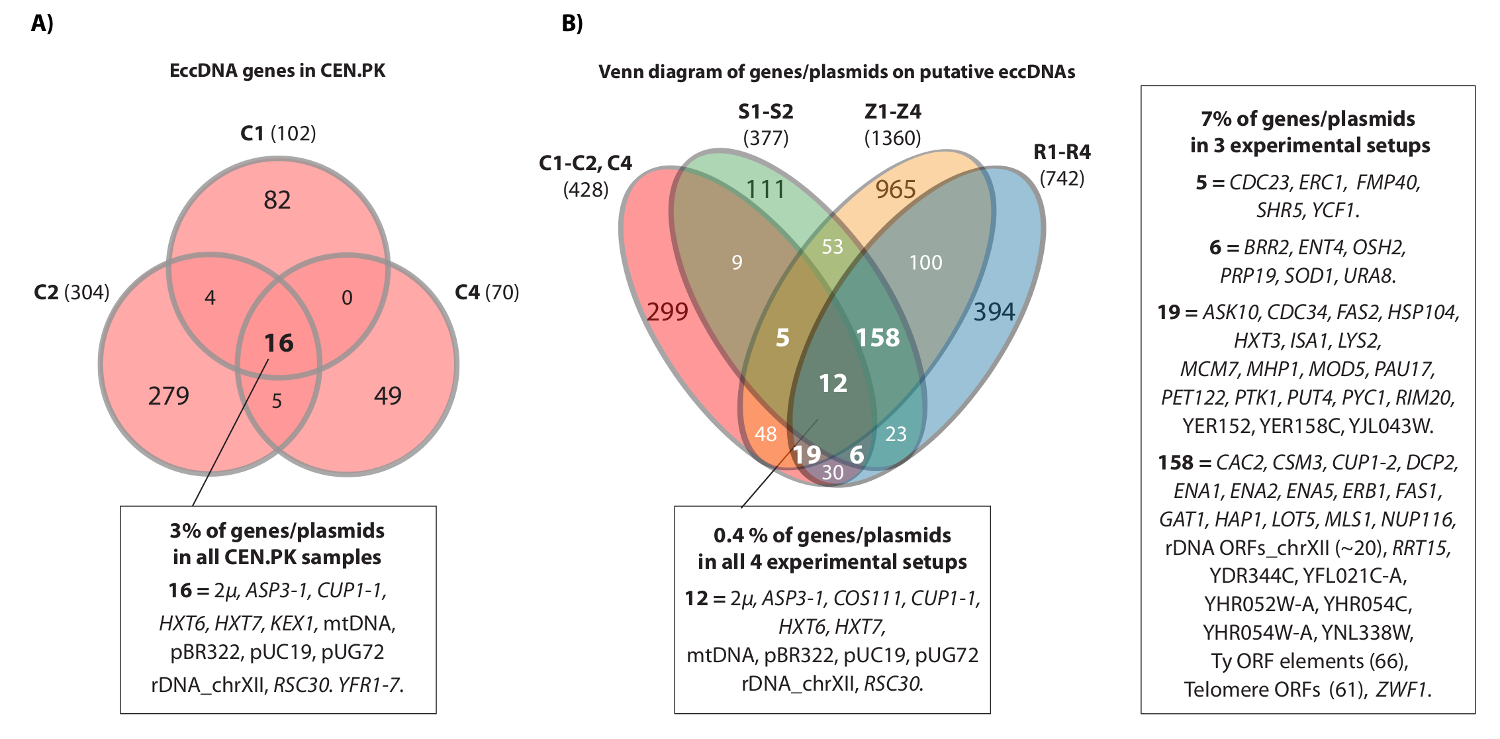
ويظهر شريط الفيديو مثال متجاورة يقرأ أن تعيينها إلى موضع HXT7 _ARS432_ HXT6 على الكروموسوم الرابع. سابقا، تم اكتشاف [HXT6 / 7 دائرة] من قبل دائرة تسلسل في عشرة السكان S288c (كل مع 1 × 10 10 خلايا) وبنية الحمض النووي التعميم ما أكده تحليل PCR معكوس 20. و[HXT6 / 7 كاريتاس الدوليةrcle] كما تم تسجيل في كل من الثلاثة السكان CEN.PK الشكل (4A). وعلاوة على ذلك، فإن معظم الجينات eccDNA مشتركة بين العينات تكرار من CEN.PK تداخل الجينات eccDNA من مجموعات البيانات S288c (الشكل 4B).
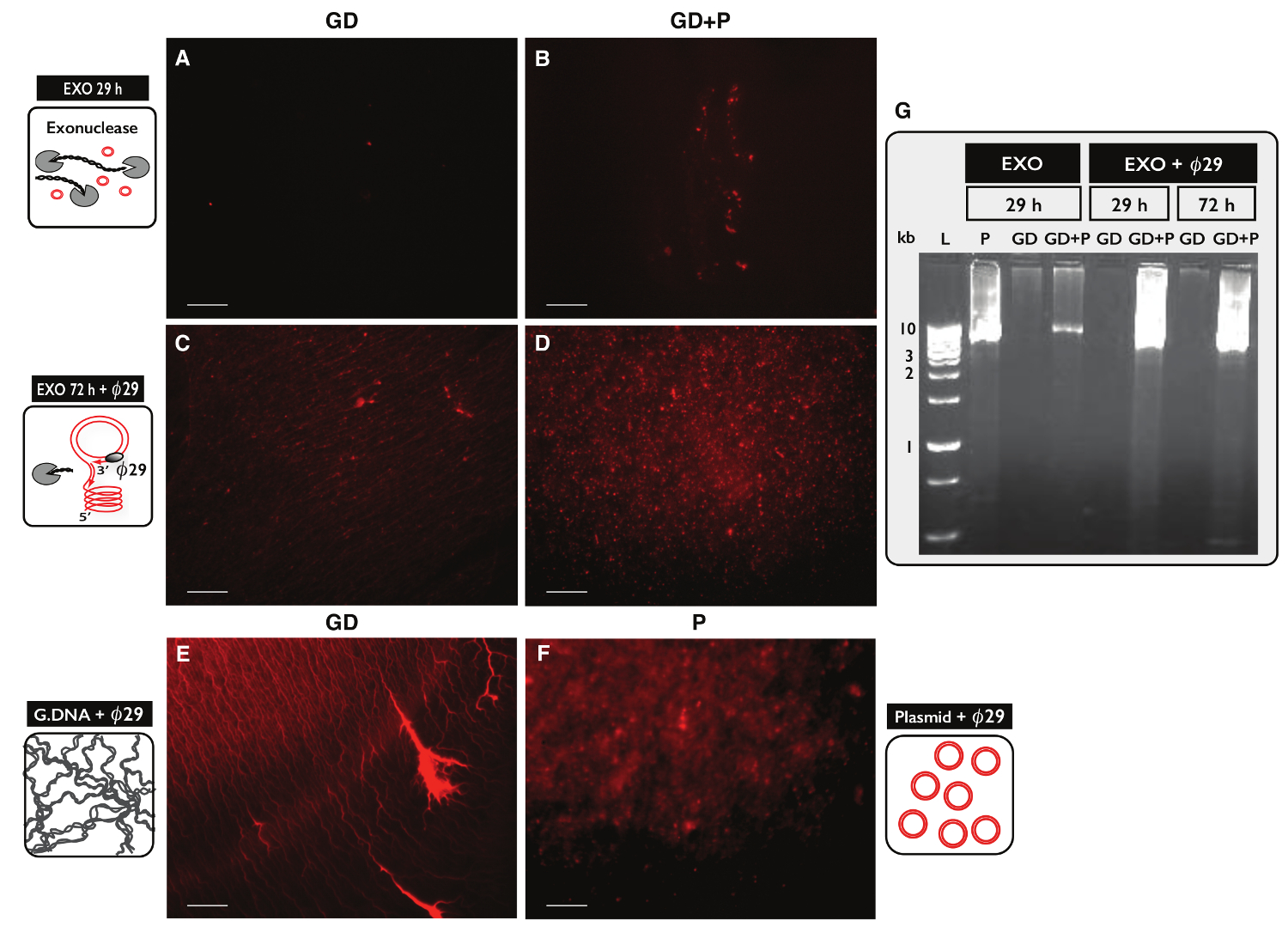
لاختبار خصوصية بروتوكول دائرة يليها لتنقية الحمض النووي دائرية، عينتين، ولكل منها 30 ميكروغرام الحمض النووي الجيني، تم اختبارها. واستكملت عينة واحدة مع 100 نانوغرام DNA البلازميد وeccDNA من وتنقيته من بروتوكول دائرة يليها كل من العينات. بعد فصل العمود، كان العائد DNA 1.27٪ (380 نانوغرام) لعينة من دون البلازميد (GD) و1.60٪ (480 نانوغرام) لعينة مع البلازميد (GD + P). تم اختبار كفاءة العلاج نوكلياز خارجية لمحتوى الحمض النووي الخطي بعد 29 ساعة و 72 ساعة باستخدام PCR ضد ACT1. لا العينات احتوت على ACT1 تضخيم (لا تظهر البيانات). وكان جزء من كل عينة المعالجة نوكلياز خارجية أبعدmplified من قبل ø29 وقد تم تحليل البلمرة والمنتجات من التفاعلات الإنزيمية من تلطيخ يوديد propidium (الشكل 5A-F) والكهربائي للهلام الاغاروز (الشكل 5G). وأظهرت عينات بعد العلاج نوكلياز خارجية الحد الأدنى propidium اليود وصمة عار (الشكل 5A-B). كشفت عينة تضخيم مع الحمض النووي الجيني الوحيد هياكل مثل موضوع (الشكل 5C) مماثلة لعينة السيطرة (الشكل 5E) - وø29. كشفت تضخيم العينة التي كان البلازميد وأضاف البؤر (الشكل 5D) تشبه سيطرة بلازميد (الشكل 5F) - وø29. وأشارت الصور التي ø29 البلمرة المخصب لالحمض النووي DNA دائري فوق خطي. تمت إزالة معظم الحمض النووي الصبغي خطية من العينات بعد العلاج 29 ساعة نوكلياز خارجية (الشكل 5A-B، G). ومع ذلك، فإن العلاج نوكلياز خارجية واسعة لأكثر من 100 ساعة وباستخدام أكثر من 100 وحدة هناك حاجة لإزالة جميع الكروموسومات الحمض النووي الخطي، كما ø29 - عينات تضخيم لا تزال تظهر خلفية هياكل تشبه موضوع بعد 72 ساعة نوكلياز خارجية العلاج (الشكل 5C-D).

. الشكل 1. الخطوط العريضة للطريقة الدائرة يليها البروتوكول قد 5 الخطوات التالية: 1) خلية زراعة، 2) تنقية وإثراء eccDNA بواسطة اللوني العمود، 3) هضم ما تبقى الخطي الحمض النووي الصبغي في جزء الشطافة، 4) من التضخيم الحمض النووي عن طريق ø29 البلمرة DNA، و 5) تسلسل eccDNA التخصيب ورسم الخرائط من يقرأ إلى س. الخباز الجينوم المرجعية. الرجاء انقر هنا لعرض نسخة أكبر من هذا الرقم.
{kind=link}
p_upload / 54239 / 54239fig2.jpg "/>
الشكل 2. ملاصق يقرأ> 1 كيلوبايت وظيفة من تسلسل العمق. EccDNA من 1 × 10 10 خلايا زيادة بوصفها وظيفة من تسلسل عمق (بملايين تعيين قراءة). هو مبين: يثلث البيولوجية من فرداني CEN.PK س. الخباز السكان (C1، C2، C4) مفصولة 10 10 انقسامات الخلية. الرجاء انقر هنا لعرض نسخة أكبر من هذا الرقم.
{kind=link}

الشكل 3. الكشف عن عناصر الحمض النووي الدائري المعروف. (AB) مبعثر المؤامرات التغطية القراءة (قراءة الكثافة) في المئة لالبلازميدات في CEN.PK البيولوجية يعيد C1، C2 و C4. (A) المعينة يقرأ على البلازميدات الخميرة الذاتية هم: 2μ. [دائرة الحمض النووي المؤتلف] (جينات الحمض النووي الريبي الريباسي الثاني عشر من كروموسوم)؛ وو mtDNA (الحمض النووي). (ب) فريد يقرأ معين للسيطرة على البلازميدات. وقد ارتفعت البلازميدات تحكم في عينات قبل تنقية عمود. وكانت نسب البلازميد لكل خلية: pBR322 (زائد علامات) 1: 1، pUC19 (الدوائر) 01:50، وpUG72 (المثلثات) 1: 2500.

الشكل 4. عناصر eccDNA المشتركة في CEN.PK وS288c. (A) فين الرسم البياني عرض التداخل بين 476 الجينات على 294 عناصر eccDNA في عينات CEN.PK ثلاثة (C1، C2، C4). والمشروح 16 مشتركة تداخل الجينات eccDNA / البلازميدات (جميع الأسماء الجين في مجموعة بيانات 1). (ب) فين الرسم البياني لجميع الجينات المسجلة على eccDNAs المفترضة من العينات CEN.PK ثلاثة (C1، C2، C4)، بالمقارنة مع جميع الجينات المسجلة على eccDNAs المفترضة من 10 عينات S288c: S1-S2، R1-R4، Z1-Z4 (أنظر المرجع 20). يظهر عدد 13 مكررات البيولوجية (S1-S2، R1-R4، Z1-Z4، C1-C3) مع الجينات / البلازميدات والمناطق eccDNA المفترضة التي تتداخل ما لا يقل عن 2 الخلفيات سلالة وإما 3 أو أكثر التجريبية الاجهزة. عينات C، CEN.PK. عينات R و Z، S288c BY4741. S عينات، S288c M3750. الرجاء انقر هنا لعرض نسخة أكبر من هذا الرقم.
{kind=link}

الرقم 5. التصور من عينات من الحمض النووي بعد نوكلياز خارجية وø 2 9 العلاج. (AF) يوديد propidium تلطيخ من الحمض النووي. شريط النطاق، 10 ميكرون. (A، C و E) عينات مع الحمض النووي الجيني (GD)؛ (B و D) العينات مع GD بالإضافة إلى البلازميد (GD + P). ( أونج> AB) بعد 29 ساعة العلاج نوكلياز خارجية (EXO 29 ساعة)؛ (CD) بعد العلاج نوكلياز خارجية 72 ساعة تليها ø29 البلمرة التضخيم (EXO 72 ساعة + ø29). (E) الجينومية السيطرة الحمض النووي بعد ه: ø29 البلمرة التضخيم. (F) السيطرة البلازميد (5.5 كيلوبايت) بعد التضخيم 29 البلمرة. (G س) الاغاروز هلام eletrophoresis. من اليسار: L، 1 علامات كيلوبايت. ف، ومراقبة البلازميد (5.5 كيلوبايت) بعد EXO 29 ساعة. GD، بعد EXO 29 ساعة (عينة كما في ألف)؛ GD + P، بعد EXO 29 ساعة (العينة على النحو باء)؛ GD وGD + P، بعد EXO 29 ساعة + ø29. GD وGD + P، بعد EXO 72 ساعة + ø29 (العينة كما هو الحال في CD). انظر الجدول S1 للحصول على تفاصيل إضافية. الرجاء انقر هنا لعرض نسخة أكبر من هذا الرقم.
{kind=link}
يتبعها 1 "SRC =" / ملفات / ftp_upload / 54239 / 54239dataset1.jpg "/>
مجموعة البيانات 1. المحتملة المناطق الحمض النووي التعميم في CEN.PK. الرجاء انقر هنا لتحميل هذا الملف.
أظهرت هي تسلسل البيانات والتحليلات عن 348 المناطق. الأعمدة م، ورسم الخرائط eccDNA. ألف (العمود الأول من اليسار)، عينة من التي تم تحديدها المفترضة eccDNA. B، كروموسوم. مؤتمر نزع السلاح، بداية ونهاية إحداثيات eccDNAs المفترضة. EH، المحتوى eccDNA. E، تكرار مستقل تسلسل (ARS) في المنطقة؛ F، الجينات الكامل في المنطقة؛ G، وهي جزء من الجينات المدرجة في المنطقة؛ H، جينات BLASTN التي تم تحديدها. IO، والتغطية EccDNA والقيم ص. الأول، أطول المنطقة مع تسلسل المشروح فريد في شركة بريتيش بتروليوم. J، عدد كل تعيين يقرأ. K، وتغطية كل تعيين يقرأ بشظايا في كيلوبايت من مليون معين يقرأ (FPKM)؛ L، ف قيمة المفترضة eccDNA مقارنة حدوثها عن طريق الصدفة من مونتي كارلو المحاكاة. M، عدد من الامم المتحدةiquely معين يقرأ. لا؛ كما K و L فقط باستخدام تعيين فريد يقرأ (UFPKM). المعلمات لرسم الخرائط من يقرأ وكانت محاكاة مونت كارلو كما هو موضح 20.
Discussion
على طريقة الدائرة يليها يسمح الكشف عن الجينوم على نطاق من eccDNA من خلايا الخميرة مع قرار على مستوى التسلسل. هذه الطريقة لتنقية eccDNA الخفيفة التي لا تتطلب دوامة المركزة أو pipetting لويستخدم فصل العمود عن طريق الجاذبية للحد من eccDNA الكسر التي من شأنها أن تؤدي إلى الهضم نوكلياز خارجية في خطوة لاحقة. هذه الميزات من الأسلوب قد تكون حاسمة للكشف عن eccDNAs الكبيرة التي تحتوي على تسلسل الجين. الكشف عن دائرة تسلسل العديد eccDNAs بما في ذلك الجينات الكاملة (الإدراجات 1). انها رصدت أيضا 86 كيلوبايت الخميرة الحمض النووي. وهكذا، هذا البروتوكول يسهل تنقية عناصر كبيرة DNA دائرية. الحفاظ على عدد من الخطوات استخراج الحمض النووي إلى أدنى حد ممكن يقلل من مخاطر الخسارة eccDNA ويزيد الغلة. بناء على نتائج للسيطرة، ارتفعت في البلازميدات، دائرة تسلسل حساس للغاية، والكشف عن الحمض النووي دائري واحد من 2500 الخلايا. وعلاوة على ذلك، وإزالة البلازميدات الذاتية وفيرة مثل 2μ. البلازميد أو الحمض النووي قد يعزز إلى حد كبير حساسية. وقد وصفت علاج من 2μ من الثقافات الخميرة 30. بدلا من ذلك، 2μ وإزالة الحمض النووي يمكن أن يتحقق مع نوكلياز داخلية قطع نادرة، مثل صوي. ومع ذلك، يمكن أن الخطوة انزيم التقييد يستهدف eccDNAs الأخرى من الفائدة وتحد من المحصول الكلي eccDNA.
كانت خطوات مهمة لكشف eccDNA إزالة الحمض النووي الخطي (الخطوة 3) وتسلسل الحمض النووي (الخطوة 5) إلى عمق السليم. لتسجيل غالبية eccDNAs من سكان الخلية، قد تكون هناك حاجة تسلسل عميق 20. إقران نهاية التسلسل ينبغي أن توفر الثقة أكبر من الكشف eccDNA، حيث من المتوقع أن تسفر عن إقران نهاية تقاطعات دائرية الحمض النووي يقرأ تلك الخريطة discordantly. هذه التناقضات تدعم اكتشاف هياكل الحمض النووي دائرية ويحتمل أن يتم استخدامها بوصفها عامل تصفية eccDNA الكشف إضافية.
الدائرة سيتم التحقق من صحة طريقة ف باستخدام ثلاثة S. مستقلة السكان الخباز CEN.PK. وتضمنت تسلسل الكشف عنها سابقا eccDNAs، البلازميدات الذاتية وارتفعت في البلازميدات ومئات من eccDNAs المفترضة (الإدراجات 1). هذه النتائج تدعم مجموعات البيانات السابقة، الدائرة وما يليها من س. الخباز S288c 20. اكتشاف العديد eccDNAs مشتركة بين CEN.PK وS288c السكان يشير إلى أن هذه المكاني لديها الميل إلى وجود العناصر كما دائرية (الشكل 4). لقد أظهرنا سابقا أن [دائرة GAP1] تم تخصيب تحت النيتروجين الظروف محدودة في الخلفية CEN.PK 8، رغم أنه لم يتم العثور على دليل على [GAP1 الدائرة] في الخلفيات سلالة أخرى. العثور على eccDNA من CUP1-1 RSC30، ASP3-1، COS111، وHXT6 HXT7 مواضع في كل S288c وCEN.PK تشير إلى أن الاستعداد للالتعميم الحمض النووي هو يخدعخدم بين سلالات الخميرة. يبقى أن يظهر إذا [HXT6 / 7 الدائرة]، [ASP3-1 الدائرة]، [COS111 الدائرة]، و [الدائرة CUP1-1 RSC30] تمنح مزايا انتقائية لخلايا أو إذا وجودها هو مجرد تأثير ارتفاع معدلات التعميم الحمض النووي.
مجتمعة، تشير النتائج إلى أن الدائرة تسلسل مناسب تماما للكشف عن eccDNAs-كيلو قاعدة الحجم ومزاياه لتحديد eccDNAs مع جينات كاملة. دائرة تسلسل هو طريقة حساسة للغاية تمكن شاشات الجينوم على نطاق كامل من eccDNAs من الخميرة. على طريقة الدائرة بعدها يمكن أن يفتح مجالا جديدا من البحوث التي تهدف إلى توضيح دور eccDNA في توليد الحذف الجينات والتكبير. وبالنظر إلى أن العمارة الحمض النووي وهيكل وحفظها إلى حد كبير من الخميرة حقيقية النواة لحقيقيات النوى أعلى، وطريقة الدائرة يليها يجب، من حيث المبدأ، أن يكون applicablالبريد إلى جميع الخلايا حقيقية النواة، مع تعديلات طفيفة. في الوقت الحاضر، لا يظهر طريقة لديك أي قيود، على الرغم من قدرته على تنقية eccDNAs-megabase الحجم لم يتم بعد مبين. وبالإضافة إلى ذلك، فإن استخدام ø29 البلمرة DNA، والذي يستخدم المتداول دائرة أسلوب التضخيم 31، ويخلق وجود تحيز نحو eccDNAs أصغر مما eccDNA تقدير أكثر صعوبة. دائرة تسلسل يكشف eccDNAs كبيرة بما يكفي لتحمل جينات كاملة، مما يجعلها مناسبة للدراسات في الدقيقة دائرية مزدوجة من الحمض النووي من خلايا جسدية الإنسان. يمكن دقيقة مزدوجة تساهم في السرطان عندما يتم تضخيم بروتو الجينات المسرطنة على هذه العناصر 32-37. ويمكن استخدام دراسات eccDNAs في الخلايا الجرثومية لقياس معدلات الطفرة سلالة الجرثومية وتقييم نوعية الحيوانات المنوية، على سبيل المثال في مجال الثروة الحيوانية. وهكذا، دائرة تسلسل لديه القدرة على انتاج الأفكار إلى المعدل الذي ينشأ الاختلاف الجيني في شكل من التباين في عدد النسخ، ويؤدي إلى فهم رواية من الأمراض التي تنطوي على copy- الجينعدد الاختلاف 38-40.
Disclosures
The authors declare that they have no competing financial interests.
Acknowledgements
Thanks to Kenn D. Møller and Claus Sternberg (DTU) for technical assistance and to Tue S. Jørgensen for quantitative PCR analysis.
Materials
| Name | Company | Catalog Number | Comments |
| Bacto peptone | BD Difco | 211677 | Alternative product can be used. |
| Brilliant III SYBR Green PCR Master Mix | Agilent Technologies | 600882 | For qPCR analysis. Alternative product can be used. |
| Dextrose (D-glucose) | Carl Roth | HN06.4 | Alternative product can be used. |
| Disruptor Beads, 0.5 mm | Scientific Industries, Inc. | SI-BG05 | Glass beads to disrupt plasma cell membranes. Alternative product can be used. |
| Ethidium bromide | Carl Roth | 2218.2 | Agarose gel stain for detecting DNA/RNA. |
| GeneJet plasmid miniprep kit | Thermo Fisher | K0502 | Plasmid purifcation from bacteria. Alternative product can be used |
| NotI, FastDigest | Life Technologies - Thermo Fisher Scientific, USA | FD0594 | Endonuclease. Alternative product can be used. |
| Plasmid Mini AX kit | A&A Biotechnology, Poland | 010-50 | Plasmid purifcation kit used to purify eccDNA. |
| Plasmid-Safe ATP-dependent DNase kit | Epicentre, USA | E3105K | ATP-dependent exonuclease kit. Alternative product can be used. |
| Propidium iodide | Sigma-Aldrich, USA | 81845 | Alternative product can be used. |
| pUG6 plasmid | EUROSCARF, Germany | P30114 | Marker gene: loxP-PAgTEF1-kanMX-TAgTEF1-loxP. Plasmid requests: Please contact Dr. Peter Philippsen@unibas.ch |
| QIAGEN genomic-tip 100/G | Qiagen, USA | 13343 | Genomic DNA purifcation from yeast. Alternative product can be used. |
| REPLI-g Mini Kit protocol | Qiagen, USA | 150023 | Amplification of eccDNA by the phi29 polymerase |
| Yeast extract | BD Difco | 210929 | Alternative product can be used. |
| Zymolyase 100T (Lyticase, Yeast Lytic Enzyme) | Nordic BioSite, Sweden | Z1004-3 | Alternative product can be used. |
| Data access to sequence files | European Nucleotide Archive | EccDNA dataset from Saccharomyces cerevisiae CEN.PK113-7D. Study accession number PRJEB9684. 2nd accession number is ERP010820. Locus tag prefix is BN2032. | |
| Strains | |||
| Saccharomyces cerevisiae CEN.PK113-7D | Genotype MATa MAL2-8c SUC2 | ||
| Saccharomyces cerevisiae yeast deletion library pool | EUROSCARF, Germany | S288c BY4741 pool of 4400 viable single-gene deletion mutants disrubted by KanMX module. Genotypes MATa his3∆1 leu2∆0 met15∆0 ura3∆0 genexxx::KanMX. | |
| Equipments | |||
| DNA Spectrophotometer | NanoDrop 1000 Spectrophotometer, Thermo Fisher | Measuring DNA concentration. Alternative product can be used. | |
| Fluorescence microscopy | Nikon Optronics Magnafire. Red excitation fluorescence filter, 663-738 nm. | Alternative product can be used. | |
| Robotic library-build system | Apollo 324, IntegenX Inc. | DNA library preparation. Alternative product can be used. | |
| Sequencing platform | Illumina HiSeq 2000 platform, Illumina Inc. | DNA sequencing. Alternative product can be used. | |
| Ultrasonicator | Covaris LE220, microTUBE AFA Fiber tubes | Alternative product can be used. | |
| Methods | |||
| 2% YPD media | Mix 10 g Dextrose, 10 g Yeast extract, 20 g Bacto peptone and add H2O to a total volume of 1000 ml and autoclave. | ||
| Circle-Seq test on genomic DNA | Genomic DNA was purified (Qiagen) from a pool of the yeast deletion library (Euroscarf). The DNA concentration was measured by nanodrop and 30 µg genomic DNA was pipetted into two micro centrifuge tubes. One micro centrifuge tube was supplemented with 100 nanogram plasmid (pUG6). The DNA samples were purified by Circle-Seq, omitting the protocol steps 1.1-1.3 and 1.5-1.7. The eluted DNA concentrations were measured by nanodrop and the entire DNA yield from sample GD and GD+P was treated with exonuclease for a period of 29 hours. A 10% fraction was collected for phi29-amplification and PCR analysis, while the remaining DNA was subjected to 72 hour exonuclease treatment. The samples were analyzed for linear DNA content by PCR, using the ACT1 gene as chromosomal marker. A 5% fraction of each of the exonuclease treated samples was amplified by the phi29 DNA polymerase for 16 hours (Qiagen). The presence of DNA in each sample was examined by loading an equal amount (7 µl) in wells on an 0.5 µg/ml ethidium-bromide 0.9% agarose gel after running gel-electrophoresis. | ||
| Mapping software | Bowtie2 aligner, John Hopkins University | Ultrafast short read alignment. Reference: Langmead B, Salzberg S. Fast gapped-read alignment with Bowtie 2. Nature Methods. 2012, 9:357-359. | |
| Propidium iodide stain | Images of propidium iodine stained DNA were captured by fluorescence microscopy at 100x magnification (100x/1.30 oil, Nikon) in the RFP channel (red excitation fluorescence filter, 663-738 nm) using identical exposition time (5 seconds). | ||
| Workflow bioinformatic system | Galaxy, Open source. | A free web-based platform for data intensive biomedical research. References: Goecks, J, Nekrutenko, A, Taylor, J and The Galaxy Team. Galaxy: a comprehensive approach for supporting accessible, reproducible, and transparent computational research in the life sciences. Genome Biol. 2010 Aug 25;11(8):R86. Blankenberg D, Von Kuster G, Coraor N, Ananda G, Lazarus R, Mangan M, Nekrutenko A, Taylor J. "Galaxy: a web-based genome analysis tool for experimentalists". Current Protocols in Molecular Biology. 2010 Jan; Chapter 19:Unit 19.10.1-21. Giardine B, Riemer C, Hardison RC, Burhans R, Elnitski L, Shah P, Zhang Y, Blankenberg D, Albert I, Taylor J, Miller W, Kent WJ, Nekrutenko A. "Galaxy: a platform for interactive large-scale genome analysis." Genome Research. 2005 Oct; 15(10):1451-5. | |
References
- Kugelberg, E., Kofoid, E., et al. The Tandem Inversion Duplication in Salmonella enterica.: Selection Drives Unstable Precursors to Final Mutation Types. Genetics. 185 (1), 65-80 (2010).
- Reams, A. B., Kofoid, E., Savageau, M., Roth, J. R. Duplication Frequency in a Population of Salmonella enterica. Rapidly Approaches Steady State With or Without Recombination. Genetics. 184 (4), 1077-1094 (2010).
- Smith, C. A., Vinograd, J. Small polydisperse circular DNA of HeLa cells. Journal of Molecular Biology. 69 (2), 163-178 (1972).
- Carroll, S. M., DeRose, M. L., et al. Double Minute Chromosomes Can Be Produced from Precursors Derived from a Chromosomal Deletion. Molecular and cellular biology. 8 (4), 1525-1533 (1988).
- Cohen, S., Yacobi, K., Segal, D. Extrachromosomal Circular DNA of Tandemly Repeated Genomic Sequences in Drosophila. Genome research. 13 (6A), 1133-1145 (2003).
- Horowitz, H., Haber, J. E. Identification of Autonomously Replicating Circular Subtelomeric Y' Elements in Saccharomyces cerevisiae. Molecular and cellular biology. 5 (9), 2369-2380 (1985).
- Moore, I. K., Martin, M. P., Dorsey, M. J., Paquin, C. E. Formation of Circular Amplifications in Saccharomyces cerevisiae by a Breakage-Fusion-Bridge Mechanism. Environmental and molecular mutagenesis. 36 (2), 113-120 (2000).
- Gresham, D., Usaite, R., Germann, S. M., Lisby, M., Botstein, D., Regenberg, B. Adaptation to diverse nitrogen-limited environments by deletion or extrachromosomal element formation of the GAP1 locus. Proceedings of the National Academy of Sciences of the United States of America. 107 (43), 18551-18556 (2010).
- Windle, B., Draper, B. W., Yin, Y. X., O'Gorman, S., Wahl, G. M. A central role for chromosome breakage in gene amplification, deletion formation, and amplicon integration. Genes & development. 5 (2), 160-174 (1991).
- Gresham, D., Ruderfer, D. M., et al. Genome-Wide Detection of Polymorphisms at Nucleotide Resolution with a Single DNA Microarray. Science. 311 (5769), 1932-1936 (2006).
- Kidd, J. M., Cooper, G. M., et al. Mapping and sequencing of structural variation from eight human genomes. Nature. 453 (7191), 56-64 (2008).
- Gresham, D., Desai, M. M., Botstein, D., Dunham, M. J. The Repertoire and Dynamics of Evolutionary Adaptations to Controlled Nutrient-Limited Environments in Yeast. PLoS Genetics. 4 (12), 1-19 (2008).
- Lang, G. I., Botstein, D., Desai, M. M. Genetic Variation and the Fate of Beneficial Mutations in Asexual Populations. Genetics. 188 (3), 647-661 (2011).
- van Loon, N., Miller, D., Murnane, J. P. Formation of extrachromosomal circular DNA in HeLa cells by nonhomologous recombination. Nucleic Acids Research. 22 (13), 2447-2452 (1994).
- Vinograd, J., Lebowitz, J. Physical and Topological Properties of Circular Dna. Journal of General Physiology. 49 (6P2), 103 (1966).
- Shibata, Y., Kumar, P., et al. Extrachromosomal MicroDNAs and Chromosomal Microdeletions in Normal Tissues. Science. 336 (6077), 82-86 (2012).
- Dillon, L. W., Kumar, P., et al. Production of Extrachromosomal MicroDNAs Is Linked to Mismatch Repair Pathways and Transcriptional Activity. Cell Reports. 11 (11), 1749-1759 (2015).
- Li, L. L., Norman, A., Hansen, L. H., Sørensen, S. J. Metamobilomics - our knowledge on the pool of plasmid encoded traits in natural environments using high-throughput sequencing. Clinical microbiology and infection : the official publication of the European Society of Clinical Microbiology and Infectious Diseases. 18, 5-7 (2012).
- Brown Kav, A., Sasson, G., Jami, E., Doron-Faigenboim, A., Benhar, I., Mizrahi, I. Insights into the bovine rumen plasmidome. Proceedings of the National Academy of Sciences of the United States of America. 109 (14), 5452-5457 (2012).
- Møller, H. D., Parsons, L., Jørgensen, T. S., Botstein, D., Regenberg, B. Extrachromosomal circular DNA is common in yeast. Proceedings of the National Academy of Sciences of the United States of America. , 201508825 (2015).
- Drubin, D. G., Miller, K. G., Botstein, D. Yeast Actin-Binding Proteins - Evidence for a Role in Morphogenesis. The Journal of cell biology. 107 (6), 2551-2561 (1988).
- Magdolen, V., Drubin, D. G., Mages, G., Bandlow, W. High levels of profilin suppress the lethality caused by overproduction of actin in yeast cells. FEBS letters. 316 (1), 41-47 (1993).
- Sandrock, T. M., Brower, S. M., Toenjes, K. A., Adams, A. Suppressor analysis of fimbrin (Sac6p) overexpression in yeast. Genetics. 151 (4), 1287-1297 (1999).
- Blanco, L., Bernad, A., Lázaro, J. M., Martìn, G., Garmendia, C., Salas, M. Highly Efficient DNA Synthesis by the Phage ø29 DNA Polymerase. The Journal of biological chemistry. 264 (15), 8935-8940 (1989).
- Dean, F. B. Rapid Amplification of Plasmid and Phage DNA Using Phi29 DNA Polymerase and Multiply-Primed Rolling Circle Amplification. Genome research. 11 (6), 1095-1099 (2001).
- Hutchison, C. A., Smith, H. O., Pfannkoch, C., Venter, J. C. Cell-free cloning using ø29 DNA polymerase. Proceedings of the National Academy of Sciences of the United States of America. 102 (48), 17332-17336 (2005).
- Goecks, J., Nekrutenko, A., Taylor, J., Galaxy Team, T. Galaxy: a comprehensive approach for supporting accessible, reproducible, and transparent computational research in the life sciences. Genome Biology. 11 (8), 86 (2010).
- Giardine, B., Riemer, C., et al. Galaxy: A platform for interactive large-scale genome analysis. Genome research. 15 (10), 1451-1455 (2005).
- Langmead, B., Salzberg, S. L. Fast gapped-read alignment with Bowtie 2. Nature Methods. 9 (4), 357 (2012).
- Tsalik, E. L., Gartenberg, M. R. Curing Saccharomyces cerevisiae of the 2 micron plasmid by targeted DNA damage. Yeast. 14 (9), 847-852 (1998).
- Norman, A., Riber, L., Luo, W., Li, L. L., Hansen, L. H., Sørensen, S. J. An Improved Method for Including Upper Size Range Plasmids in Metamobilomes. PLoS ONE. 9 (8), e104405 (2014).
- Storlazzi, C. T., Lonoce, A., et al. Gene amplification as double minutes or homogeneously staining regions in solid tumors: Origin and structure. Genome research. 20 (9), 1198-1206 (2010).
- Von Hoff, D. D., Needham-VanDevanter, D. R., Yucel, J., Windle, B. E., Wahl, G. M. Amplified human MYC localized to replicating submicroscopic circular DNA molecules. Proceedings of the National Academy of Sciences of the United States of America. 85 (13), 4804-4808 (1988).
- Raymond, E., Faivre, S., et al. Effects of hydroxyurea on extrachromosomal DNA in patients with advanced ovarian carcinomas. Clinical cancer research : an official journal of the American Association for Cancer Research. 7 (5), 1171-1180 (2001).
- Shimizu, N. Extrachromosomal Double Minutes and Chromosomal Homogeneously Staining Regions as Probes for Chromosome Research. Cytogenetic and genome research. 124 (3-4), 3-4 (2009).
- Eckhardt, S. G., Dai, A., Davidson, K. K., Forseth, B. J., Wahl, G. M., Von Hoff, D. D. Induction of differentiation in HL60 cells by the reduction of extrachromosomally amplified c-myc. Proceedings of the National Academy of Sciences of the United States of America. 91 (14), 6674-6678 (1994).
- Vogt, N., Lefèvre, S. -. H., et al. Molecular structure of double-minute chromosomes bearing amplified copies of the epidermal growth factor receptor gene in gliomas. Proceedings of the National Academy of Sciences of the United States of America. 101 (31), 11368-11373 (2004).
- Ahn, K., Gotay, N., et al. High rate of disease-related copy number variations in childhood onset schizophrenia. Molecular psychiatry. 19 (5), 568-572 (2013).
- Girirajan, S., Johnson, R. L., et al. Global increases in both common and rare copy number load associated with autism. Human molecular genetics. 22 (14), 2870-2880 (2013).
- Vogt, N., Gibaud, A., Lemoine, F., de la Grange, P., Debatisse, M., Malfoy, B. Amplicon rearrangements during the extrachromosomal and intrachromosomal amplification process in a glioma. Nucleic Acids Research. , (2014).
Reprints and Permissions
Request permission to reuse the text or figures of this JoVE article
Request PermissionExplore More Articles
This article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. All rights reserved