Method Article
La purificación de todo el genoma de ADN circular Extracromosómica a partir de células eucariotas
En este artículo
Resumen
This paper presents a sensitive method called Circle-Seq for purifying extrachromosomal circular DNA (eccDNA). The method encompasses column purification, removal of remaining linear chromosomal DNA, rolling-circle amplification and high-throughput sequencing. Circle-Seq is applicable to genome-scale screening of eukaryotic eccDNA and studying genome instability and copy-number variation.
Resumen
ADN circular extracromosómico (eccDNAs) son elementos genéticos comunes en Saccharomyces cerevisiae y se presentan en otros eucariotas también. EccDNAs contribuyen a la variación genética entre las células somáticas en organismos multicelulares y para la evolución de los eucariotas unicelulares. Se necesitan métodos sensibles para detectar eccDNA para aclarar cómo estos elementos afectan a la estabilidad del genoma y cómo los factores ambientales y biológicos inducen su formación en células eucariotas. Este video presenta un método de purificación eccDNA-sensible llamado Círculo-Seq. El método comprende la purificación en columna de ADN circular, la eliminación de ADN lineal restante cromosómica, amplificación círculo rodante de eccDNA, la secuenciación de profundidad, y la cartografía. se requiere tratamiento con exonucleasa extensa para suficiente degradación del ADN cromosómico lineal. La etapa de amplificación por círculo rodanteine-height:. normal "> φ 29 de la polimerasa de ADN enriquecido para circular sobre ADN lineal Validación del método Circle-Seq en tres poblaciones de S. cerevisiae CEN.PK de 10 10 células detecta cientos de perfiles eccDNA en tamaños más grandes de 1 kilobase . descubrimientos repetidos de ASP3-1, COS111, CUP1, RSC30, HXT6, Hxt7 genes en el ADN circular en tanto S288c y CEN.PK sugiere que la circularización del ADN se conserva entre las cepas en estos loci. en resumen, el método Circle-Sec tiene una amplia aplicabilidad para la detección de genoma escala para eccDNA en eucariotas, así como para la detección de tipos específicos eccDNA.
Introducción
La detección de la amplificación cromosómica temprano o transitoria es difícil, ya que requiere la identificación de alteraciones en las moléculas individuales de ADN en grandes poblaciones de células. Las variaciones del número de copias cromosómicas (CNV) generalmente se detectaron bien después de su establecimiento, dejando sólo la estructura final CNV como prueba del mecanismo que genera la variación 1,2. Detección y recuperación de ADN extracromosómico circular (eccDNA) en las primeras etapas de la formación de la CNV podría dilucidar los procesos en curso en reordenamientos genómicos.
Anteriormente, de novo descubrimiento de eccDNA era por micrografías electrónicas 3, Giemsa tinción de cromosomas en metafase 4, o electroforesis en gel de dos dimensiones 5. Estos métodos proporcionan poca o ninguna información acerca de la secuencia del ADN circular. Técnicas específicas tales como transferencia Southern 6,7, 8 PCR inversa, o fluorescencia en situ hybridization 9 proporcionan evidencia sólo de elementos eccDNA específicos. Ninguno de estos métodos proporcionan la secuencia de todos los tipos eccDNA existentes en una población celular.
Divergencia genómico en una piscina de las células se puede caracterizar por la secuenciación del genoma y / o suelo de baldosas arrays 10,11. La detección de una deleción o amplificación por métodos de purificación de ADN convencionales por lo general requiere que un alelo mutado representan al menos 0,1 a 1% de la población de células 12,13. Se espera que acéntrico eccDNAs a ser aún más transitoria en un cultivo celular debido a su falta de centrómeros y el potencial ausencia de síntesis de ADN en replicación. Por lo tanto, ya que la mayoría eccDNAs presumiblemente están en bajas cantidades y sus secuencias se asemejan el genoma, se necesitan métodos de extracción de ADN alternativos para detectar eccDNAs.
Varias técnicas de purificación de ADN circular explotan las diferencias estructurales entre los cromosomas y el ADN circular. Por ejemplo, ultracentrifug de alta velocidadación en gradientes de cloruro de cesio se utiliza para aislar 350-3000 pares de bases (pb) eccDNAs grandes de la línea celular HeLa de cáncer humano 14. Sin embargo, a alta velocidad puede romper o una muesca en la columna vertebral de las estructuras de ADN circular superenrollado, la alteración de la velocidad de sedimentación 15 y el rendimiento eccDNA. Dutta y sus colaboradores desarrollaron un método de novo, la identificación del genoma a gran escala de ADN circular a partir de tejidos de ratón, así como a partir de cultivos de células humanas y de pollo 16,17. Su método es la extracción de núcleos de tejido homogeneizado por ultracentrifugación sacarosa seguido de la purificación del plásmido y varias rondas de reacciones enzimáticas y las extracciones de ADN. Su protocolo identifica principalmente 200-400 pb eccDNAs, llamados microDNAs. Dutta y compañeros de trabajo también intentaron purificación de microDNAs de Saccharomyces cerevisiae, pero no fueron capaces de grabar microDNA de esta especie de levadura 16.
Hemos desarrollado un nuevo método parade novo detección de eccDNA de levadura llamada Circle-Seq. Este método permite estudios de genoma escala de moléculas de ADN circular lo suficientemente grandes para llevar a genes enteros y tan grandes como el ADN 86 kilobases (kb) mitocondrial (ADNmt). El método Circle-Sec se desarrolló a partir de un método de purificación de plásmidos procariotas bien establecida 18,19, optimizado para las células de levadura eucariotas y en combinación con la secuenciación profunda. Utilizando el enfoque Círculo-Sec, 1756 eccDNAs diferentes, todos mayores de 1 kb, fueron detectados desde las diez S. cerevisiae S288c poblaciones 20. Un tamaño de corte fue elegido para centrarse en eccDNA que eran lo suficientemente grandes para transportar genes enteros. Círculo-Sec era muy sensible; se detectó un único eccDNA dentro de miles de células 20. En el estudio actual, Círculo-Sec se utilizó para aislar e identificar 294 eccDNAs de tres réplicas biológicas de otro S. cerevisiae cepa de levadura, CEN.PK. Los datos revelan que eccDNA es un eleme genética comúnnt en S. cepas de Saccharomyces cerevisiae.
Protocolo
NOTA: Una visión general de la purificación de ADN circular y método de secuenciación (Círculo-Sec) se ilustra en la Figura 1.
1. El cultivo, la cosecha de la célula y la membrana plasmática Interrupción
- Inocular células de levadura (por ejemplo Saccharomyces cerevisiae) de un cultivo O / N en 50 ml de medio nutriente completo de dextrosa de levadura de peptona (YPD). Inocular a una densidad celular inicial baja de 1-3 x 10 5 células / ml o una densidad óptica de aproximadamente 0,01 OD 600.
- Se incuban las células a 30 ° C con agitación a 150 revoluciones por minuto (rpm) hasta que las células alcanzan la densidad celular máxima de aproximadamente 1 x 10 10 células, aproximadamente después de 24 a 48 horas o una densidad óptica a OD 600> 10,0.
NOTA: El tiempo de cultivo no es crucial, ya que las concentraciones de células inferiores pueden ser utilizados.
- Se incuban las células a 30 ° C con agitación a 150 revoluciones por minuto (rpm) hasta que las células alcanzan la densidad celular máxima de aproximadamente 1 x 10 10 células, aproximadamente después de 24 a 48 horas o una densidad óptica a OD 600> 10,0.
- La transferencia de la cultura de crecimiento excesivo a un tubo cónico de 50 ml, sedimentar las células por centrifugación a800 g durante 3 min y descartar el sobrenadante.
- Lavar el precipitado con 25 ml de solución tampón de Tris-Cl 10 mM, EDTA 1 mM, pH 8,0, volver a sedimentar las células por centrifugación a 800 xg durante 3 min y desechar el sobrenadante.
- Resuspender el sedimento celular en 1,2 ml de tampón de resuspensión suministrada desde un kit de plásmido de columna de purificación.
- Paso opcional: Añadir plásmidos altamente diluidas como controles para la purificación de elementos de ADN circular 20.
NOTA: En el conjunto de datos actual, se aplicó una mezcla de plásmidos 7,7 l de cada muestra que contiene 10 10 células. La mezcla de plásmidos social consistió en tres plásmidos en diferentes concentraciones; pBR322 a 38 ng / muestra, pUC19 en 0,5 ng / muestra, y pUG72 en 0,01 ng / muestra. - Transferir la suspensión celular en dos tubos de 2 ml de microcentrífuga, cada uno suplementado con perlas de vidrio de 0,5 mm en una proporción de 1: 3 del volumen de la suspensión total.
- Vortex cada tubo a máxima velocidad durante 10 min para interrumpir de células plasmáticasmembranas. Sedimentar las perlas por centrifugación a 268 xg durante 30 seg y transferir los 1,2 ml de sobrenadante combinado de los dos tubos de microcentrífuga a un nuevo tubo.
NOTA: Alternativa al paso 1,6-1,7, utilice zimoliasa para romper las células en solución tampón 0,6 ml resuspensión. Diez unidades de zimoliasa pueden perturbar 5 x 10 7 células dentro de 1,5 horas a 35 ° C.
2. EccDNA Enriquecimiento mediante cromatografía en columna
- Siga el protocolo de un kit de purificación en columna de plásmidos. En resumen, el tratamiento de cada muestra con una solución alcalina de 1,2 ml, mezclar suavemente y se incuba 3 minutos a temperatura ambiente.
- Añadir 1,2 ml de tampón de neutralización, mezclar suavemente y centrifugar a 9650 xg durante 5 min.
- Cargar la solución en una columna equilibrada con solución 1 ml de equilibrado y permite que el líquido fluya a través de la columna por gravedad.
- Lavar la columna con solución de lavado 4 ml. Cuando la solución ha pasado a través de la resina, añadir con cuidado 0,3 ml de elución por lolución para reemplazar la mayor parte de la columna de 0,35 ml de volumen vacío.
- Eluir el ADN en un nuevo tubo de recogida con una solución de 1 ml de elución y precipitar el ADN mediante la adición de 0,8 ml de mezcla de precipitación. Centrifugar a 9650 xg durante 10 min.
- Lavar el sedimento de ADN con 0,5 ml 70% de etanol, se centrifuga a 9650 xg durante 5 min, secar al aire durante 5 a 15 min y se disuelve el ADN purificado en 25 l de agua estéril.
NOTA: Sólo el almacenamiento a corto plazo del ADN en agua se recomienda. Preferentemente, vaya directamente al paso 3.
3. La digestión del ADN cromosómico lineal restante
- Paso opcional: Para facilitar la digestión específica de ADN lineal por exonucleasa, tratar el ADN purificado con una endonucleasa rara-corte, tal como NotI. Para 5 g de ADN, utilizar 1 unidad NotI, 5 l de 10x tampón de digestión y agua estéril hasta un volumen total de 50 l. Incubar la reacción a 37 ° C durante 16 horas y el calor inactivar la endonucleasa a 80 ° C durante 5 min.
- Añadir 20unidades de exonucleasa (2 l), 4 ATP l (25 mM), 34 l de agua estéril y 10 l de 10x tampón de reacción directamente a la 50 l de ADN endonucleasa escinde para llegar a un volumen de reacción 1x de 100 l, utilizando la exonucleasa dependiente de ATP equipo.
- Realizar la hidrólisis de DNA de cadena simple y de cadena doble lineal a 37 ° C durante 5 días o más. Añadir un adicional de ATP 4 l (25 mM), tampón de reacción 0,6 l de 10x y 20 unidades de exonucleasa cada 24 horas para continuar la digestión del ADN enzimática en un volumen de reacción 1x.
- Después de la eliminación de ADN lineal, muestra de 2 l de la solución tratada exonucleasa para confirmar la eliminación de ADN lineal cromosómico mediante reacción en cadena de la polimerasa cuantitativa (qPCR), utilizando un marcador cromosómico como la actina ACT1 gen 20.
- Cada 20 l de volumen de reacción qPCR contiene 2 l de muestra exonucleasa tratados, 150 nm cebadores 5'-ACT1 TCCGTCTGGATTGGTGGTTCTA-3 'y 5'-TGGACCACTTTCGTCGTATTC-3 ', 2% (volumen / volumen) de sulfóxido de dimetilo, y 10 l verde fluorescente mezcla maestra.
- Utilice las condiciones de reacción; 3 min a 95 ° C, seguido por 45 ciclos de 15 segundos a 95 ° C y 30 segundos a 60 ° C.
NOTA: ACT1 es un marcador especialmente adecuado para ADN lineal ya que las variaciones del número de copias de este gen son perjudiciales 21-23 por lo que no debe llevar a eccDNA ACT1. - Alternativas a análisis de digestión de ADN mediante qPCR son PCR estándar (4,3) o tinción de yoduro de propidio (4,4).
- Utilice 2 l de muestra exonucleasa tratados como molde de PCR con los cebadores 5'-ACT1 TGGATTCTGGTATGTTCTAGC-3 'y 5'-GAACGACGTGAGTAACACC-3'. Como control positivo ACT1, utilice 50-100 ng genómico de S. ADN cerevisiae como molde. condiciones de reacción de PCR; 3 min a 95 ° C, seguido de 35 ciclos de 30 segundos a 95 ° C, 30 seg a 56 ° C y 1 min a 72 ° C.
- Ejecutar PCR reacciones por electrophore gelsis en 1% de agarosa con 0,5 mg / ml de bromuro de etidio. Busque una banda de 0,8 kb ACT1.
- La ausencia o presencia de ADN lineal también pueden ser examinados por tinción con yoduro de propidio antes y después de la amplificación de ADN.
- Mezcle cada muestra de ADN en una relación 1: 1 con un volumen de 1: 1.000 H 2 O-solución diluida de 20 mM de yoduro de propidio de valores. Deje la solución en la oscuridad durante 10 a 20 min a TA y analizar la tinción de ADN por microscopía de fluorescencia a 100 aumentos utilizando un filtro de excitación de fluorescencia roja en 663 a 738 nm y un tiempo de exposición de 5 a 30 seg. Como control DNA-tinción, el uso ø29-amplificó el ADN genómico de levadura y / o plásmido ø29-amplificado.
- Heat inactivar la solución de exonucleasa en 70 ° C durante 30 min.
4. Amplificación de ADN
- Amplificar el purificada y enriquecida eccDNA desde el paso 3.5) con ADN polimerasa de ø29 24-26 acuerdoing con el protocolo del fabricante de la polimerasa.
- En resumen, mezclar 5 l enriquecidos eccDNA con tampón de desnaturalización 5 l.
- Después de 3 min a RT, añadir tampón de neutralización 10 l. Mezclar suavemente y añadir 30 l de mezcla maestra que contiene tampón de reacción de 29 l y 1 l de ADN polimerasa de ø29. Incubar la reacción a 30 ° C durante 16 horas o más (hasta 72 h). Heat inactivar la ADN polimerasa ø29 a 65 ° C durante 3 min.
5. Secuenciación y Análisis de Datos
- Cizallar la eccDNA amplificado con un ultrasonicador enfocada a un tamaño medio de pico objetivo de 300 pb. Utilice los siguientes parámetros para una muestra de ADN l 130: potencia pico de intensidad de 450 W, 60 seg tratamiento, factor de trabajo del 30%, a 200 ciclos por ráfaga, la temperatura de 7 ° C.
- Añadir etiquetas de códigos de barras y los adaptadores de índice a la fragmentada lecturas para la síntesis de bibliotecas para la secuenciación, utilizando un método adecuado para la preparación de la biblioteca.
- Ejecutar la secuenciación de profundidad, por ejemplo, como 141-de un solo nucleótido de extremo lee en una plataforma de secuenciación de alto rendimiento.
- Mapa lee a la referencia del genoma de la levadura bajo investigación y permitir que lee para asignar a múltiples regiones. Por ejemplo, utilizar un sistema de flujo de trabajo de libre acceso y de 27,28 a corto lectura software de mapas alineador 29.
- Identificar lecturas de las regiones de eccDNAs putativos utilizando contigua lee, por ejemplo, más de siete contigua lee (> 1 kb) sin huecos 20.
NOTA: El software está disponible para explorar 27,28 asignada lee en regiones genómicas de interés.
Resultados
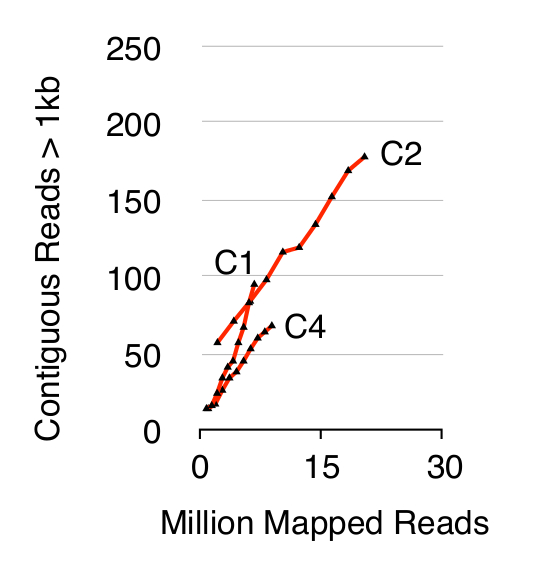
Para validar el método Circle-Sec, tres S. poblaciones de células cerevisiae CEN.PK 1 x 10 10 fueron seleccionados después de las células se cultivaron por separado en YPD durante diez generaciones. Eliminación de ADN lineal cromosómico se confirmó por la ausencia de una señal de qPCR ACT1 como se describió anteriormente 20 (datos no presentados). Purificada y enriquecida eccDNA fue secuenciado hasta 68 millones lee (141-de un solo nucleótido de extremo lee) y se asigna a la referencia del genoma CEN.PK113-7D (la versión 19 junio 2012). Grabaciones de eccDNAs putativos de las tres muestras con nombre C1, C2 y C4 fueron asignados a regiones genómicas mapeadas por contigua lee más de 1 kb. Sobre la base de 10.000 simulaciones de Monte Carlo, el significado de cada región mapeada por contigua lee más largo que se estimó 1 kb. A partir de este 79, 159 y 56 regiones fueron anotados como probables secuencias eccDNA (p <0,1, Conjunto de datos 1). El número de contiguo grabadanos lee> 1 kb aumentó como una función de la profundidad de la secuencia que sugiere que habrían sido registrados aún más elementos eccDNA si las muestras habían sido secuenciados aún más (Figura 2). Como era de esperar, el método Circle-Seq extrajo numerosos lee de un número de elementos conocidos circulares de ADN que incluye el plásmido 2μ, ADN mitocondrial, genes de ARN ribosómico en el cromosoma XII, y los tres plásmidos de control interno pBR322, pUC19 y pUG72 que fueron añadidos en muestras justo antes de la purificación en columna (Figura 3).
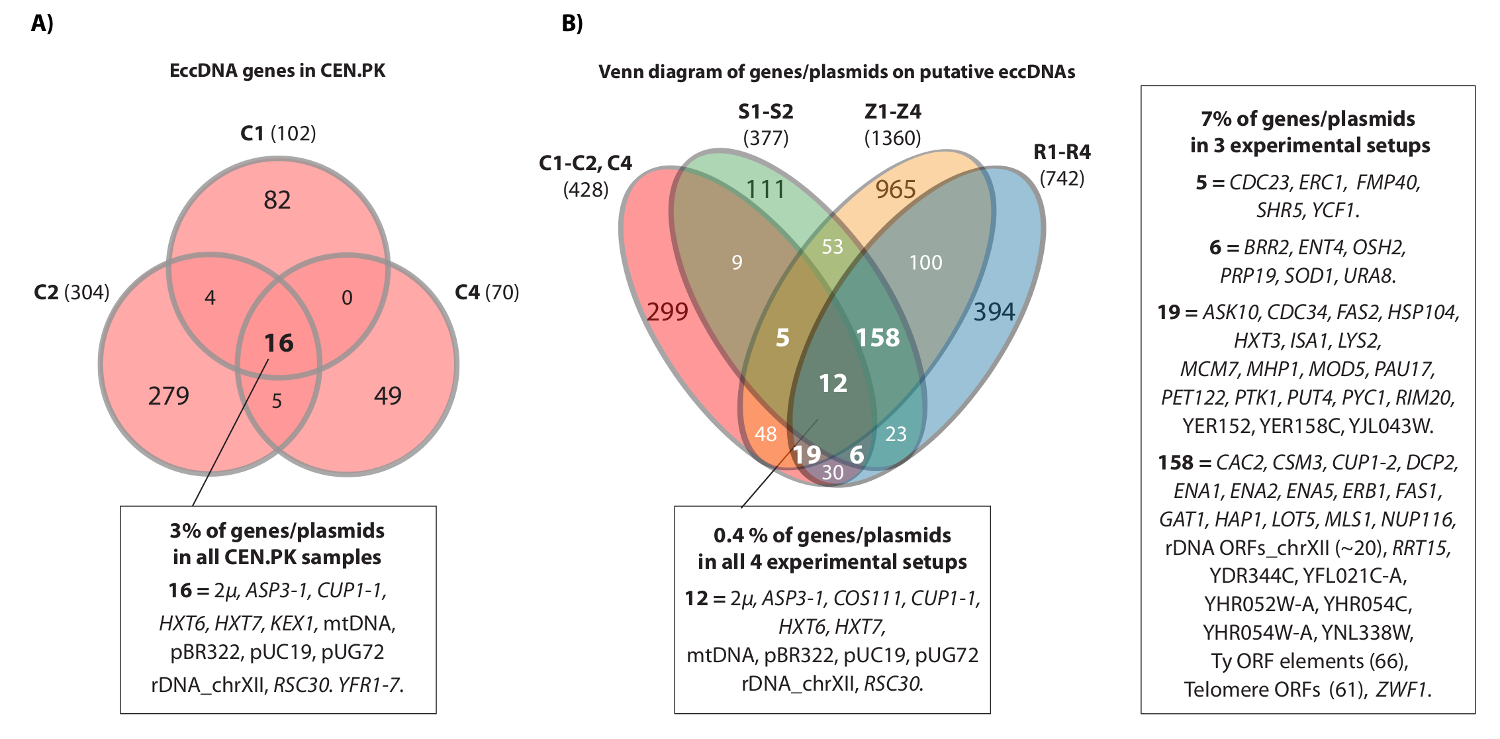
El video muestra un ejemplo de mapeado que lee contigua al lugar HXT7 _ARS432_ HXT6 en el cromosoma IV. Anteriormente, el [HXT6 / 7 círculo] fue detectado por Circle-Seq en diez poblaciones S288C (cada uno con células 1 x 10 10) y la estructura del ADN circular se confirmó por análisis de PCR inversa 20. El ci [HXT6 / 7rcle] también fue grabado en cada una de las tres poblaciones CEN.PK (Figura 4A). Además, la mayoría de los genes eccDNA comunes entre muestras repetidas de CEN.PK se traslapan con genes eccDNA de los conjuntos de datos S288c (Figura 4B).
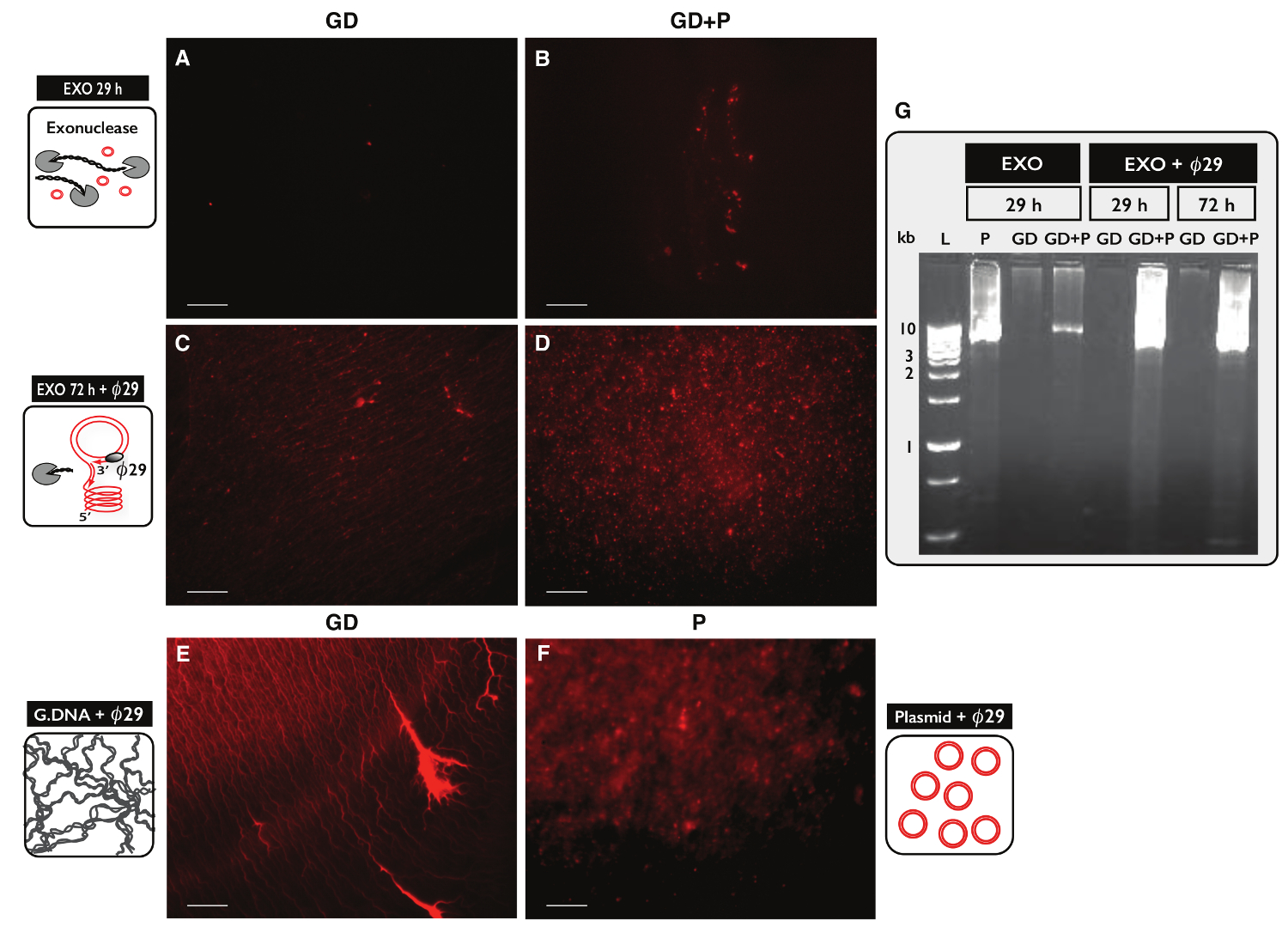
Para probar la especificidad del protocolo Circle-Seq para la purificación de ADN circular, dos muestras, cada una con ADN genómico 30 g, fueron probados. Una muestra se complementó con el plásmido de ADN 100 ng y eccDNA de ambas muestras fueron purificados por el protocolo Circle-Seq. Después de la separación en columna, el rendimiento de ADN fue de 1,27% (380 ng) para la muestra sin plásmido (GD) y 1,60% (480 ng) para la muestra con el plásmido (GD + P). La eficiencia del tratamiento de exonucleasa se ensayó para determinar el contenido de ADN lineal después de 29 hr y 72 hr utilizando PCR contra ACT1. No hay muestras contenían amplificó ACT1 (datos no mostrados). Una fracción de cada muestra de exonucleasa-tratado fue además unamplified por el ø29 polimerasa y los productos de las reacciones enzimáticas se analizaron por tinción con yoduro de propidio (Figura 5A-F) y electroforesis en gel de agarosa (Figura 5G). Las muestras después del tratamiento con exonucleasa mostraron mínima de propidio yodo-mancha (Figura 5A-B). El ø29 - muestra amplificada con sólo el ADN genómico reveló estructuras a modo de hilo (Figura 5C) similares a la muestra de control (Figura 5E). El ø29 - muestra amplificada que tenía plásmido añadido reveló focos (Figura 5D) se asemejan al control de plásmido (Figura 5F). Las imágenes indicaron que ø29 polimerasa enriquecido para el ADN circular sobre DNA lineal. La mayor parte del ADN cromosómico lineal fue retirado de las muestras después del tratamiento 29 hr exonucleasa (Figura 5A-B, G). Sin embargo, un tratamiento extenso exonucleasa durante más de 100 horas y se utilizan más de 100 unidades fue sea necesario para eliminar todo el ADN cromosómico lineal, como O29 - muestras amplificadas todavía mostraron un fondo de estructuras filiformes después de 72 horas de tratamiento con exonucleasa (Figura 5C-D).

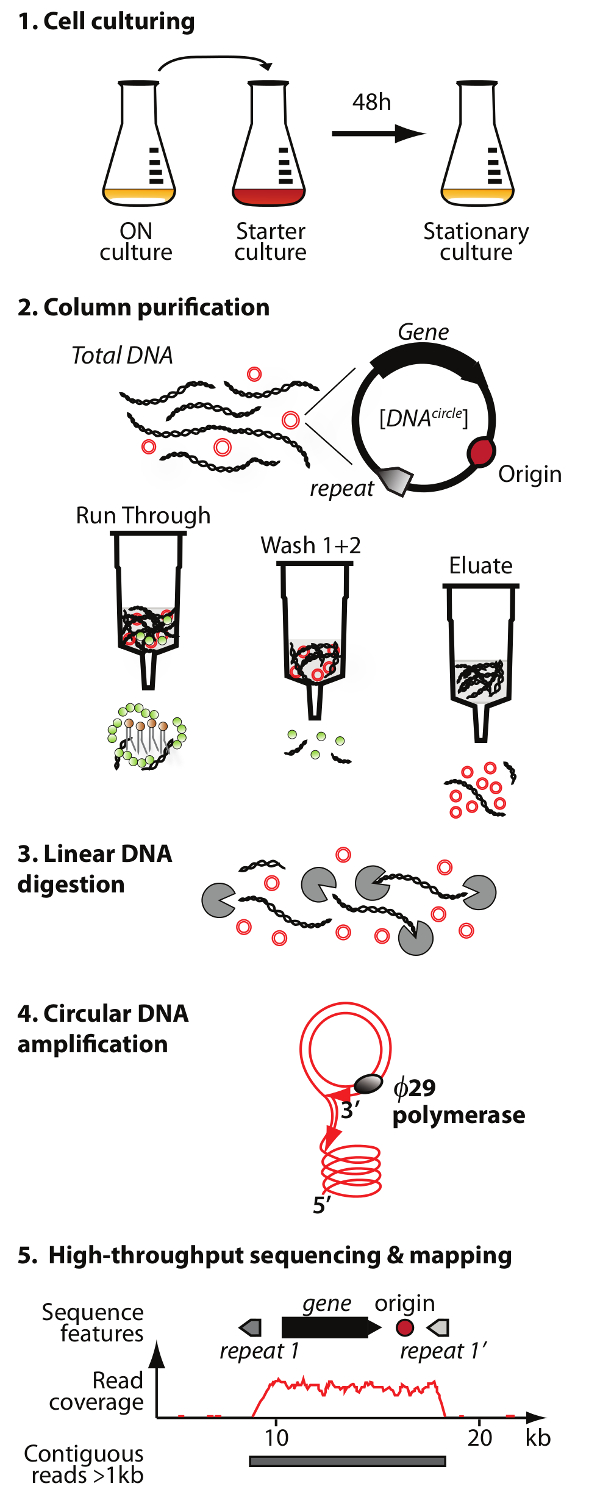
. Figura 1. Esquema del método Circle-Seq El protocolo tiene 5 pasos: 1) el cultivo de células, 2) purificación y enriquecimiento de eccDNA por cromatografía en columna, 3) la digestión de permanecer ADN cromosómico lineal en la fracción del eluido, 4) de amplificación de ADN por ø29 ADN polimerasa, y 5) secuenciación de eccDNA altamente enriquecido y el mapeo de lee a la S. cerevisiae genoma de referencia. Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}
p_upload / 54239 / 54239fig2.jpg "/>
Figura 2. Contiguo lee> 1 kb como función de la profundidad de secuencia. EccDNA de 1 x 10 10 células aumentan como una función de la profundidad de secuencia (en millones de mapeado lecturas). Se muestra: triplicados biológicos de haploides CEN.PK S. poblaciones cerevisiae (C1, C2, C4) separadas por 10 10 divisiones celulares. Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}

Figura 3. Detección de elementos de ADN circular conocidos. (AB) Dispersión parcelas de la cobertura de lectura (lectura de densidad) en el porcentaje de plásmidos en CEN.PK biológica repeticiones C1, C2 y C4. (A) asignada lee a los plásmidos de levaduras endógenas fueron: 2μ; [Círculo ADNr] (ARN ribosomal genes del cromosoma XII); y ADNmt (ADN mitocondrial). (B) lee único asignado a controlar plásmidos. plásmidos de control fueron añadidos en las muestras antes de la purificación en columna. relaciones de plásmido por célula fueron: pBR322 (signos más) 1: 1, pUC19 (círculos) 01:50, y pUG72 (triángulos) 1: 2500.

Figura 4. Elementos comunes en eccDNA CEN.PK y S288c. (A) Diagrama de Venn que muestra la superposición entre los 476 genes en 294 elementos eccDNA en las tres muestras CEN.PK (C1, C2, C4). Los 16 genes superpuestos eccDNA / plásmidos comunes son anotados (todos los nombres de genes son en conjunto de datos 1). (B) diagrama de Venn de todos los genes grabados en eccDNAs putativos de las tres muestras CEN.PK (C1, C2, C4), en comparación con todos los genes grabados en eccDNAs putativo de 10 muestras S288C: S1-S2, R1-R4, Z1-Z4 (véase la referencia 20). Se demuestran las 13 réplicas biológicas (S1-S2, R1-R4, Z1-Z4, C1-C3) con genes / plásmidos y regiones eccDNA putativos que se superponen un mínimo de 2 fondos de deformación y de 3 o más configuraciones experimentales. Ejemplos de C, CEN.PK; R muestras y Z, S288c BY4741; Muestras S, S288c M3750. Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}

Figura 5. La visualización de muestras de ADN después de exonucleasa y O 2 9 tratamiento. (AF) de propidio tinción con yoduro de ADN. Barra de escala, 10 micras. Las muestras con ADN genómico (A, C y E) (GD); (B y D) muestras con GD plus plásmido (GD + P). ( ong> AB) Después del tratamiento con exonucleasa 29 hr (EXO 29 h); (CD) después del tratamiento con exonucleasa de 72 horas seguido de la amplificación de la polimerasa de ø29 (EXO 72 h + O29). (E) de control de ADN genómico después de correo: la amplificación de la polimerasa de ø29; (F) el control plásmido (5,5 kb) después amplificación 29 de la polimerasa; (Ø G) electroforesis en gel de agarosa. De izquierda a derecha: L, 1 kb marcadores; P, control plásmido (5,5 kb) después de EXO 29 hr; GD, después de EXO 29 hr (muestra como en A); GD + P, después de EXO 29 horas (muestra como B); GD y GD + P, después de EXO 29 hr + ø29; GD y GD + P, después de EXO 72 hr + ø29 (muestra como en CD). Véase la Tabla S1 para detalles adicionales. Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}
aset 1 "src =" / files / ftp_upload / 54239 / 54239dataset1.jpg "/>
Del conjunto de datos 1. Las regiones circularización del ADN potenciales en CEN.PK. Haga clic aquí para descargar este archivo.
Se muestran los datos de la secuencia y análisis de 348 regiones. Las columnas son AD, eccDNA mapeo. A (primera columna desde la izquierda), muestra de la que se identificó putativo eccDNA; B, el cromosoma; CD, inicio y final de las coordenadas eccDNAs putativos. EH, el contenido eccDNA. E, secuencia de replicación autónoma (ARS) en la región; F, gen completo de la región; G, parte del gen incluido en la región; H, BLASTN gen-identificado. IO, la cobertura EccDNA y los valores de p. Yo, la región más larga con una secuencia anotada en forma exclusiva pb; J, número de todo planeado lee; K, la cobertura de todos asignada lee por fragmentos por kb de un millón de mapeado lee (FPKM); L, valor de p para putativo eccDNA comparación con ocurrencia por casualidad de simulaciones de Monte Carlo; M, el número de las Naciones Unidasiquely asignada lee; NO; como K y L utilizando sólo asignada de forma única lee (UFPKM). Los parámetros para la asignación de las lecturas y las simulaciones de Monte Carlo fueron como las descritas 20.
Discusión
El método Circle-Sec permite la detección del genoma escala de eccDNA de células de levadura con una resolución a nivel de secuencia. El método es una purificación eccDNA leve que no requiere vórtice intensivo o pipeteo y utiliza la columna de separación por gravedad para limitar eccDNA rotura que conduciría a la digestión de exonucleasa en la etapa subsiguiente. Estas características del método pueden ser cruciales para la detección de grandes eccDNAs que contienen secuencias de genes. Círculo-Sec detectado numerosos eccDNAs incluyendo genes completos (1) del conjunto de datos. También detecta el ADN mitocondrial de levadura 86 kb. Por lo tanto, este protocolo facilita la purificación de grandes elementos de ADN circulares. Manteniendo el número de etapas de extracción de ADN a un mínimo reduce el riesgo de pérdida eccDNA y maximiza el rendimiento. Con base en los resultados para el control, se disparó en plásmidos, Círculo-Sec es muy sensible, detectando un ADN circular a partir de 2.500 células. Además, la eliminación de abundantes plásmidos endógenos tales como 2μ; plásmido o ADN mitocondrial pueden mejorar significativamente la sensibilidad. El curado de 2μ de cultivos de levadura se ha descrito 30. Alternativamente, 2μ y extracción de ADN mitocondrial podrían lograrse con una endonucleasa de corte raro-, como SwaI. Sin embargo, el paso de enzimas de restricción podría dirigirse a otros eccDNAs de interés y limitar el rendimiento total eccDNA.
Los pasos críticos para la detección eccDNA eran eliminación de ADN lineal (paso 3) y la secuenciación del ADN (etapa 5) a una profundidad adecuada. Para grabar la mayoría de eccDNAs a partir de una población de células, la secuenciación profunda podría ser necesario 20. secuenciación de extremo emparejado debe proporcionar incluso una mayor probabilidad de detección eccDNA, pues se espera que los cruces de ADN circulares para producir de gama emparejado lee ese mapa discordante. Estas discrepancias apoyan el descubrimiento de estructuras de ADN circulares y potencialmente se pueden utilizar como un filtro eccDNA de detección adicional.
El Círculo-Seq método fue validado usando tres S. independiente poblaciones cerevisiae CEN.PK. Secuencias detectadas previamente incluidos informó eccDNAs, plásmidos endógenos y se trataron en plásmidos y cientos de eccDNAs putativos (1) del conjunto de datos. Estos hallazgos apoyan los conjuntos de datos anteriores Circle-Sec de S. cerevisiae S288c 20. El descubrimiento de varios eccDNAs comunes a las poblaciones CEN.PK y S288C indica que estos loci tienen una propensión a existir como elementos circulares (Figura 4). Hemos demostrado previamente que el [círculo GAP1] se enriquece en condiciones limitadas de nitrógeno en el fondo CEN.PK 8, aunque no se ha encontrado evidencia de [círculo GAP1] en otras cepa antecedentes. Hallazgo de eccDNA del CUP1-1 RSC30, ASP3-1, COS111, y loci HXT6 HXT7 tanto en S288c y CEN.PK sugiere que una predisposición a la circularización del ADN es estafaservido entre las cepas de levadura. Se queda por demostrar si [HXT6 / 7 círculo], [círculo ASP3-1], [círculo COS111], y [círculo CUP1-1 RSC30] confieren ventajas selectivas a las células o si su existencia no es más que un efecto de las altas tasas de circularización del ADN.
Tomados en conjunto, los resultados indican que el Círculo-Sec es muy adecuado para la detección de eccDNAs-kilobases de tamaño y tiene ventajas para la identificación de eccDNAs con genes completos. Circle-Seq es un método altamente sensible que permite pantallas enteros a escala del genoma de eccDNAs de la levadura. El método Circle-Sec podría abrir un nuevo campo de investigación destinada a esclarecer el papel de eccDNA en la generación de deleciones de genes y amplificaciones. Dado que la arquitectura y la estructura de ADN se conservan en gran medida a partir de levadura eucariota para eucariotas superiores, el método Circle-Seq debe, en principio, ser applicablelectrónico a todas las células eucariotas, con ligeras modificaciones. En la actualidad, el método no parecen tener ningún tipo de limitaciones, aunque su capacidad de purificar eccDNAs tamaño de megabases aún no se ha demostrado. Además, el uso de ø29 ADN polimerasa, que utiliza un método de amplificación de círculo rodante 31, crea un sesgo hacia eccDNAs más pequeños que hacen eccDNA cuantificación más difícil. Círculo-Sec detecta eccDNAs lo suficientemente grandes para transportar genes completos, por lo que es adecuada para los estudios relativos a la doble de ADN circular minutos a partir de células somáticas humanas. Minuto dobles pueden contribuir al cáncer cuando los proto-oncogenes se amplifican en estos elementos 32-37. Los estudios de eccDNAs en las células de la línea germinal se podrían utilizar para medir las tasas de mutación de la línea germinal y evaluar la calidad del esperma, por ejemplo en el ganado. Por lo tanto, Circle-Seq tiene el potencial de ayudar a comprender la velocidad a la que se produce la variación genética en la forma de la variación del número de copias, y dar lugar a una nueva comprensión de las enfermedades que implican de autor genla variación del número 38-40.
Divulgaciones
The authors declare that they have no competing financial interests.
Agradecimientos
Thanks to Kenn D. Møller and Claus Sternberg (DTU) for technical assistance and to Tue S. Jørgensen for quantitative PCR analysis.
Materiales
| Name | Company | Catalog Number | Comments |
| Bacto peptone | BD Difco | 211677 | Alternative product can be used. |
| Brilliant III SYBR Green PCR Master Mix | Agilent Technologies | 600882 | For qPCR analysis. Alternative product can be used. |
| Dextrose (D-glucose) | Carl Roth | HN06.4 | Alternative product can be used. |
| Disruptor Beads, 0.5 mm | Scientific Industries, Inc. | SI-BG05 | Glass beads to disrupt plasma cell membranes. Alternative product can be used. |
| Ethidium bromide | Carl Roth | 2218.2 | Agarose gel stain for detecting DNA/RNA. |
| GeneJet plasmid miniprep kit | Thermo Fisher | K0502 | Plasmid purifcation from bacteria. Alternative product can be used |
| NotI, FastDigest | Life Technologies - Thermo Fisher Scientific, USA | FD0594 | Endonuclease. Alternative product can be used. |
| Plasmid Mini AX kit | A&A Biotechnology, Poland | 010-50 | Plasmid purifcation kit used to purify eccDNA. |
| Plasmid-Safe ATP-dependent DNase kit | Epicentre, USA | E3105K | ATP-dependent exonuclease kit. Alternative product can be used. |
| Propidium iodide | Sigma-Aldrich, USA | 81845 | Alternative product can be used. |
| pUG6 plasmid | EUROSCARF, Germany | P30114 | Marker gene: loxP-PAgTEF1-kanMX-TAgTEF1-loxP. Plasmid requests: Please contact Dr. Peter Philippsen@unibas.ch |
| QIAGEN genomic-tip 100/G | Qiagen, USA | 13343 | Genomic DNA purifcation from yeast. Alternative product can be used. |
| REPLI-g Mini Kit protocol | Qiagen, USA | 150023 | Amplification of eccDNA by the phi29 polymerase |
| Yeast extract | BD Difco | 210929 | Alternative product can be used. |
| Zymolyase 100T (Lyticase, Yeast Lytic Enzyme) | Nordic BioSite, Sweden | Z1004-3 | Alternative product can be used. |
| Data access to sequence files | European Nucleotide Archive | EccDNA dataset from Saccharomyces cerevisiae CEN.PK113-7D. Study accession number PRJEB9684. 2nd accession number is ERP010820. Locus tag prefix is BN2032. | |
| Strains | |||
| Saccharomyces cerevisiae CEN.PK113-7D | Genotype MATa MAL2-8c SUC2 | ||
| Saccharomyces cerevisiae yeast deletion library pool | EUROSCARF, Germany | S288c BY4741 pool of 4400 viable single-gene deletion mutants disrubted by KanMX module. Genotypes MATa his3∆1 leu2∆0 met15∆0 ura3∆0 genexxx::KanMX. | |
| Equipments | |||
| DNA Spectrophotometer | NanoDrop 1000 Spectrophotometer, Thermo Fisher | Measuring DNA concentration. Alternative product can be used. | |
| Fluorescence microscopy | Nikon Optronics Magnafire. Red excitation fluorescence filter, 663-738 nm. | Alternative product can be used. | |
| Robotic library-build system | Apollo 324, IntegenX Inc. | DNA library preparation. Alternative product can be used. | |
| Sequencing platform | Illumina HiSeq 2000 platform, Illumina Inc. | DNA sequencing. Alternative product can be used. | |
| Ultrasonicator | Covaris LE220, microTUBE AFA Fiber tubes | Alternative product can be used. | |
| Methods | |||
| 2% YPD media | Mix 10 g Dextrose, 10 g Yeast extract, 20 g Bacto peptone and add H2O to a total volume of 1000 ml and autoclave. | ||
| Circle-Seq test on genomic DNA | Genomic DNA was purified (Qiagen) from a pool of the yeast deletion library (Euroscarf). The DNA concentration was measured by nanodrop and 30 µg genomic DNA was pipetted into two micro centrifuge tubes. One micro centrifuge tube was supplemented with 100 nanogram plasmid (pUG6). The DNA samples were purified by Circle-Seq, omitting the protocol steps 1.1-1.3 and 1.5-1.7. The eluted DNA concentrations were measured by nanodrop and the entire DNA yield from sample GD and GD+P was treated with exonuclease for a period of 29 hours. A 10% fraction was collected for phi29-amplification and PCR analysis, while the remaining DNA was subjected to 72 hour exonuclease treatment. The samples were analyzed for linear DNA content by PCR, using the ACT1 gene as chromosomal marker. A 5% fraction of each of the exonuclease treated samples was amplified by the phi29 DNA polymerase for 16 hours (Qiagen). The presence of DNA in each sample was examined by loading an equal amount (7 µl) in wells on an 0.5 µg/ml ethidium-bromide 0.9% agarose gel after running gel-electrophoresis. | ||
| Mapping software | Bowtie2 aligner, John Hopkins University | Ultrafast short read alignment. Reference: Langmead B, Salzberg S. Fast gapped-read alignment with Bowtie 2. Nature Methods. 2012, 9:357-359. | |
| Propidium iodide stain | Images of propidium iodine stained DNA were captured by fluorescence microscopy at 100x magnification (100x/1.30 oil, Nikon) in the RFP channel (red excitation fluorescence filter, 663-738 nm) using identical exposition time (5 seconds). | ||
| Workflow bioinformatic system | Galaxy, Open source. | A free web-based platform for data intensive biomedical research. References: Goecks, J, Nekrutenko, A, Taylor, J and The Galaxy Team. Galaxy: a comprehensive approach for supporting accessible, reproducible, and transparent computational research in the life sciences. Genome Biol. 2010 Aug 25;11(8):R86. Blankenberg D, Von Kuster G, Coraor N, Ananda G, Lazarus R, Mangan M, Nekrutenko A, Taylor J. "Galaxy: a web-based genome analysis tool for experimentalists". Current Protocols in Molecular Biology. 2010 Jan; Chapter 19:Unit 19.10.1-21. Giardine B, Riemer C, Hardison RC, Burhans R, Elnitski L, Shah P, Zhang Y, Blankenberg D, Albert I, Taylor J, Miller W, Kent WJ, Nekrutenko A. "Galaxy: a platform for interactive large-scale genome analysis." Genome Research. 2005 Oct; 15(10):1451-5. | |
Referencias
- Kugelberg, E., Kofoid, E., et al. The Tandem Inversion Duplication in Salmonella enterica.: Selection Drives Unstable Precursors to Final Mutation Types. Genetics. 185 (1), 65-80 (2010).
- Reams, A. B., Kofoid, E., Savageau, M., Roth, J. R. Duplication Frequency in a Population of Salmonella enterica. Rapidly Approaches Steady State With or Without Recombination. Genetics. 184 (4), 1077-1094 (2010).
- Smith, C. A., Vinograd, J. Small polydisperse circular DNA of HeLa cells. Journal of Molecular Biology. 69 (2), 163-178 (1972).
- Carroll, S. M., DeRose, M. L., et al. Double Minute Chromosomes Can Be Produced from Precursors Derived from a Chromosomal Deletion. Molecular and cellular biology. 8 (4), 1525-1533 (1988).
- Cohen, S., Yacobi, K., Segal, D. Extrachromosomal Circular DNA of Tandemly Repeated Genomic Sequences in Drosophila. Genome research. 13 (6A), 1133-1145 (2003).
- Horowitz, H., Haber, J. E. Identification of Autonomously Replicating Circular Subtelomeric Y' Elements in Saccharomyces cerevisiae. Molecular and cellular biology. 5 (9), 2369-2380 (1985).
- Moore, I. K., Martin, M. P., Dorsey, M. J., Paquin, C. E. Formation of Circular Amplifications in Saccharomyces cerevisiae by a Breakage-Fusion-Bridge Mechanism. Environmental and molecular mutagenesis. 36 (2), 113-120 (2000).
- Gresham, D., Usaite, R., Germann, S. M., Lisby, M., Botstein, D., Regenberg, B. Adaptation to diverse nitrogen-limited environments by deletion or extrachromosomal element formation of the GAP1 locus. Proceedings of the National Academy of Sciences of the United States of America. 107 (43), 18551-18556 (2010).
- Windle, B., Draper, B. W., Yin, Y. X., O'Gorman, S., Wahl, G. M. A central role for chromosome breakage in gene amplification, deletion formation, and amplicon integration. Genes & development. 5 (2), 160-174 (1991).
- Gresham, D., Ruderfer, D. M., et al. Genome-Wide Detection of Polymorphisms at Nucleotide Resolution with a Single DNA Microarray. Science. 311 (5769), 1932-1936 (2006).
- Kidd, J. M., Cooper, G. M., et al. Mapping and sequencing of structural variation from eight human genomes. Nature. 453 (7191), 56-64 (2008).
- Gresham, D., Desai, M. M., Botstein, D., Dunham, M. J. The Repertoire and Dynamics of Evolutionary Adaptations to Controlled Nutrient-Limited Environments in Yeast. PLoS Genetics. 4 (12), 1-19 (2008).
- Lang, G. I., Botstein, D., Desai, M. M. Genetic Variation and the Fate of Beneficial Mutations in Asexual Populations. Genetics. 188 (3), 647-661 (2011).
- van Loon, N., Miller, D., Murnane, J. P. Formation of extrachromosomal circular DNA in HeLa cells by nonhomologous recombination. Nucleic Acids Research. 22 (13), 2447-2452 (1994).
- Vinograd, J., Lebowitz, J. Physical and Topological Properties of Circular Dna. Journal of General Physiology. 49 (6P2), 103 (1966).
- Shibata, Y., Kumar, P., et al. Extrachromosomal MicroDNAs and Chromosomal Microdeletions in Normal Tissues. Science. 336 (6077), 82-86 (2012).
- Dillon, L. W., Kumar, P., et al. Production of Extrachromosomal MicroDNAs Is Linked to Mismatch Repair Pathways and Transcriptional Activity. Cell Reports. 11 (11), 1749-1759 (2015).
- Li, L. L., Norman, A., Hansen, L. H., Sørensen, S. J. Metamobilomics - our knowledge on the pool of plasmid encoded traits in natural environments using high-throughput sequencing. Clinical microbiology and infection : the official publication of the European Society of Clinical Microbiology and Infectious Diseases. 18, 5-7 (2012).
- Brown Kav, A., Sasson, G., Jami, E., Doron-Faigenboim, A., Benhar, I., Mizrahi, I. Insights into the bovine rumen plasmidome. Proceedings of the National Academy of Sciences of the United States of America. 109 (14), 5452-5457 (2012).
- Møller, H. D., Parsons, L., Jørgensen, T. S., Botstein, D., Regenberg, B. Extrachromosomal circular DNA is common in yeast. Proceedings of the National Academy of Sciences of the United States of America. , 201508825 (2015).
- Drubin, D. G., Miller, K. G., Botstein, D. Yeast Actin-Binding Proteins - Evidence for a Role in Morphogenesis. The Journal of cell biology. 107 (6), 2551-2561 (1988).
- Magdolen, V., Drubin, D. G., Mages, G., Bandlow, W. High levels of profilin suppress the lethality caused by overproduction of actin in yeast cells. FEBS letters. 316 (1), 41-47 (1993).
- Sandrock, T. M., Brower, S. M., Toenjes, K. A., Adams, A. Suppressor analysis of fimbrin (Sac6p) overexpression in yeast. Genetics. 151 (4), 1287-1297 (1999).
- Blanco, L., Bernad, A., Lázaro, J. M., Martìn, G., Garmendia, C., Salas, M. Highly Efficient DNA Synthesis by the Phage ø29 DNA Polymerase. The Journal of biological chemistry. 264 (15), 8935-8940 (1989).
- Dean, F. B. Rapid Amplification of Plasmid and Phage DNA Using Phi29 DNA Polymerase and Multiply-Primed Rolling Circle Amplification. Genome research. 11 (6), 1095-1099 (2001).
- Hutchison, C. A., Smith, H. O., Pfannkoch, C., Venter, J. C. Cell-free cloning using ø29 DNA polymerase. Proceedings of the National Academy of Sciences of the United States of America. 102 (48), 17332-17336 (2005).
- Goecks, J., Nekrutenko, A., Taylor, J., Galaxy Team, T. Galaxy: a comprehensive approach for supporting accessible, reproducible, and transparent computational research in the life sciences. Genome Biology. 11 (8), 86 (2010).
- Giardine, B., Riemer, C., et al. Galaxy: A platform for interactive large-scale genome analysis. Genome research. 15 (10), 1451-1455 (2005).
- Langmead, B., Salzberg, S. L. Fast gapped-read alignment with Bowtie 2. Nature Methods. 9 (4), 357 (2012).
- Tsalik, E. L., Gartenberg, M. R. Curing Saccharomyces cerevisiae of the 2 micron plasmid by targeted DNA damage. Yeast. 14 (9), 847-852 (1998).
- Norman, A., Riber, L., Luo, W., Li, L. L., Hansen, L. H., Sørensen, S. J. An Improved Method for Including Upper Size Range Plasmids in Metamobilomes. PLoS ONE. 9 (8), e104405 (2014).
- Storlazzi, C. T., Lonoce, A., et al. Gene amplification as double minutes or homogeneously staining regions in solid tumors: Origin and structure. Genome research. 20 (9), 1198-1206 (2010).
- Von Hoff, D. D., Needham-VanDevanter, D. R., Yucel, J., Windle, B. E., Wahl, G. M. Amplified human MYC localized to replicating submicroscopic circular DNA molecules. Proceedings of the National Academy of Sciences of the United States of America. 85 (13), 4804-4808 (1988).
- Raymond, E., Faivre, S., et al. Effects of hydroxyurea on extrachromosomal DNA in patients with advanced ovarian carcinomas. Clinical cancer research : an official journal of the American Association for Cancer Research. 7 (5), 1171-1180 (2001).
- Shimizu, N. Extrachromosomal Double Minutes and Chromosomal Homogeneously Staining Regions as Probes for Chromosome Research. Cytogenetic and genome research. 124 (3-4), 3-4 (2009).
- Eckhardt, S. G., Dai, A., Davidson, K. K., Forseth, B. J., Wahl, G. M., Von Hoff, D. D. Induction of differentiation in HL60 cells by the reduction of extrachromosomally amplified c-myc. Proceedings of the National Academy of Sciences of the United States of America. 91 (14), 6674-6678 (1994).
- Vogt, N., Lefèvre, S. -. H., et al. Molecular structure of double-minute chromosomes bearing amplified copies of the epidermal growth factor receptor gene in gliomas. Proceedings of the National Academy of Sciences of the United States of America. 101 (31), 11368-11373 (2004).
- Ahn, K., Gotay, N., et al. High rate of disease-related copy number variations in childhood onset schizophrenia. Molecular psychiatry. 19 (5), 568-572 (2013).
- Girirajan, S., Johnson, R. L., et al. Global increases in both common and rare copy number load associated with autism. Human molecular genetics. 22 (14), 2870-2880 (2013).
- Vogt, N., Gibaud, A., Lemoine, F., de la Grange, P., Debatisse, M., Malfoy, B. Amplicon rearrangements during the extrachromosomal and intrachromosomal amplification process in a glioma. Nucleic Acids Research. , (2014).
Reimpresiones y Permisos
Solicitar permiso para reutilizar el texto o las figuras de este JoVE artículos
Solicitar permisoThis article has been published
Video Coming Soon
ACERCA DE JoVE
Copyright © 2025 MyJoVE Corporation. Todos los derechos reservados