Method Article
A purificação de todo o genoma de DNA circular Extracromossômica de células eucarióticas
Neste Artigo
Resumo
This paper presents a sensitive method called Circle-Seq for purifying extrachromosomal circular DNA (eccDNA). The method encompasses column purification, removal of remaining linear chromosomal DNA, rolling-circle amplification and high-throughput sequencing. Circle-Seq is applicable to genome-scale screening of eukaryotic eccDNA and studying genome instability and copy-number variation.
Resumo
DNAs circulares extracromossômicos (eccDNAs) são elementos genéticos comuns em Saccharomyces cerevisiae e são relatados em outros eucariotas também. EccDNAs contribuir para a variação genética entre células somáticas em organismos multicelulares e evolução das eucariotas unicelulares. métodos sensíveis para detectar eccDNA são necessários para esclarecer como esses elementos afetam a estabilidade do genoma e como fatores ambientais e biológicos induzir a sua formação em células eucarióticas. Este vídeo apresenta um método eccDNA-purificação sensível, chamado Circle-Seq. O método envolve a purificação de DNA circular, a remoção do restante DNA linear cromossômica, amplificação rolling-círculo de eccDNA, sequenciamento de profundidade, e mapeamento de coluna. tratamento exonuclease extensa foi necessária para a degradação do DNA cromossômico linear suficiente. O passo de amplificação rolling-círculoine-height:. normal; "> φ 29 polimerase enriquecido para DNA circular sobre o DNA linear de validação do método Circle-Seq em três populações de S. cerevisiae CEN.PK de 10 a 10 células detectadas centenas de perfis eccDNA em tamanhos maiores do que 1 quilobases . resultados repetidos de ASP3-1, COS111, CUP1, RSC30, HXT6, HXT7 genes no DNA circular em ambos S288c e CEN.PK sugere que circularização DNA é conservado entre as estirpes nestes loci. em suma, o método Circle-Seq tem ampla aplicabilidade para rastreio de escala genoma para eccDNA em eucariotas, bem como para a detecção de tipos específicos eccDNA.
Introdução
Detecção de amplificação cromossómico cedo ou transiente é difícil porque requer a identificação de alterações nas moléculas de ADN em grandes populações de células. Cromossómicas variações no número de cópias (CNVs) são geralmente bem detectado após o seu estabelecimento, deixando apenas a estrutura final CNV como prova de que o mecanismo de variação gerado a 1,2. Detecção e recuperação DNA extrachromosomal circular (eccDNA) nos estágios iniciais de formação de CNV poderia elucidar os processos em curso no rearranjos genômicos.
Anteriormente, de novo descoberta de eccDNA foi de micrografias eletrônicas 3, Giemsa de cromossomos em metáfase 4 ou eletroforese bidimensional 5. Estes métodos oferecem pouca ou nenhuma informação sobre a sequência do DNA circular. Técnicas visados como o Southern blotting 6,7, inverse PCR 8, ou de fluorescência em hybridizatio situn 9 fornecer provas apenas cerca elementos eccDNA específicos. Nenhum destes métodos proporcionam a sequência de todos os tipos eccDNA existentes numa população de células.
Divergência genómico em uma associação de células pode ser caracterizado por sequenciação do genoma e / ou matrizes ladrilhos 10,11. Detecção de uma deleção ou amplificação por métodos de purificação de ADN convencionais geralmente requer que um alelo mutado representam, pelo menos, 0,1-1% da população de células 12,13. Acentric eccDNAs se espera que sejam ainda mais transiente em uma cultura de células, devido à sua falta de centrómeros e potencial ausência da síntese de ADN em replicação. Assim, desde que a maioria eccDNAs presumivelmente estão em quantidades baixas e as suas sequências de se parecer com o genoma, métodos de extracção de ADN alternativos são necessários para detectar eccDNAs.
Várias técnicas de purificação de ADN circular explorar as diferenças estruturais entre os cromossomas e ADN circular. Por exemplo, ultracentrifug de alta velocidadeção em gradientes de césio-cloreto é usado para isolar 350-3000 pares de bases (pb) a partir de grandes eccDNAs a linha celular de cancro humano HeLa 14. No entanto, a alta velocidade pode quebrar ou entalhe da espinha dorsal das estruturas circulares de DNA super-enrolados, alterando a velocidade de sedimentação 15 e no rendimento eccDNA. Dutta e colaboradores desenvolveram um método para de novo, a identificação escala genoma de ADN circular a partir de tecidos de ratinho, bem como a partir de culturas de células humanas e de galinha 16,17. Seu método é a extração de núcleos de tecido homogeneizado por ultracentrifugação sacarose seguida de purificação de plasmídeos e várias rodadas de reações enzimáticas e extrações de DNA. Seu protocolo identifica principalmente 200-400 eccDNAs pb, chamados microDNAs. Dutta e colegas de trabalho também tentou a purificação de microDNAs de Saccharomyces cerevisiae, mas não foram capazes de gravar microDNA desta espécie de levedura 16.
Desenvolvemos um novo método para adetecção de novo de eccDNA de fungo chamado Círculo-Seq. Este método permite pesquisas escala genoma para moléculas circulares de DNA, grandes o suficiente para transportar genes inteiros e tão grande como o DNA 86 quilobases (kb) mitocondrial (mtDNA). O método de círculo-Seq foi desenvolvido a partir de um método de purificação de plasmídeo procarióticos bem estabelecida 18,19, optimizado para as células de levedura e eucariotas combinado com sequenciação profunda. Utilizando a abordagem Círculo-Seq, 1756 eccDNAs diferentes, todo maior do que 1 kb, foram detectados a partir de dez S. cerevisiae S288c populações 20. Um tamanho de corte foi escolhido para se concentrar em eccDNA que eram grandes o suficiente para transportar genes inteiros. Circle-Seq foi altamente sensível; ele detectou um único eccDNA dentro de milhares de células 20. No estudo atual, Circle-Seq foi usado para isolar e identificar 294 eccDNAs de três repetições biológicas de outro S. cerevisiae cepa de levedura, CEN.PK. Os dados revelam que eccDNA é um eleme genética comumnt em S. cerevisiae.
Protocolo
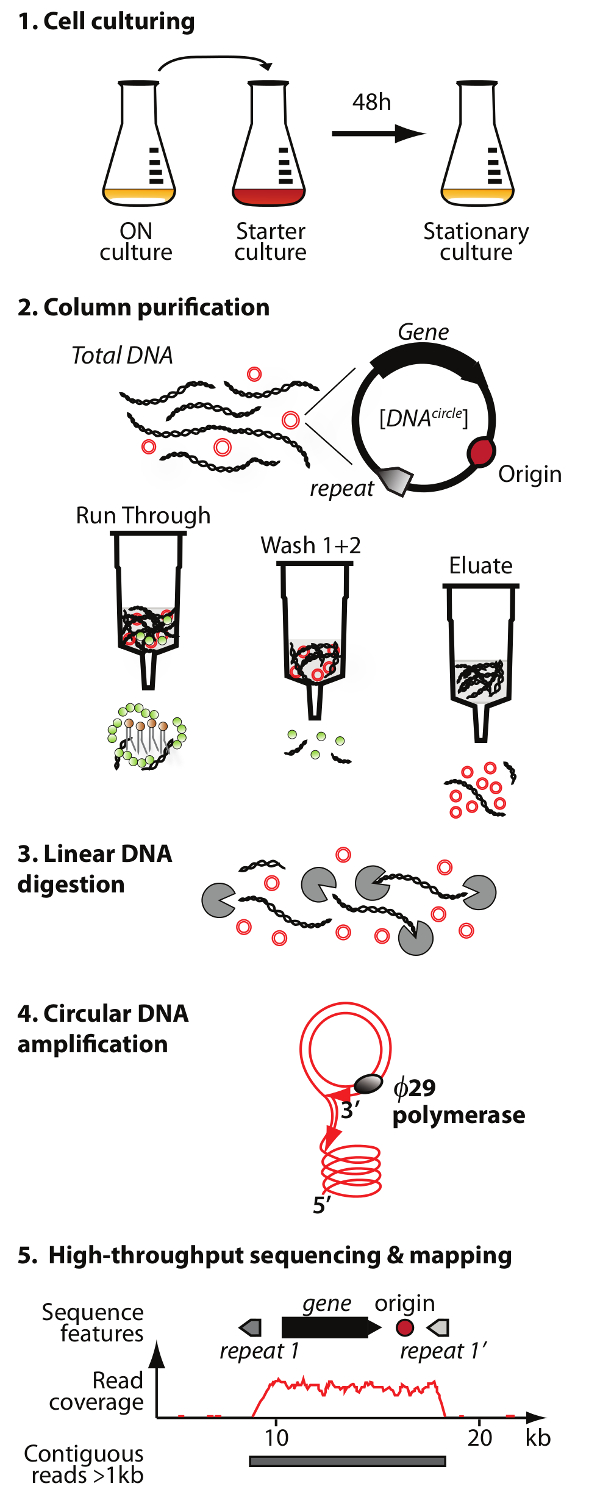
NOTA: Uma visão geral da purificação DNA circular e método de sequenciação (Circle-Seq) é ilustrada na Figura 1.
1. A cultura, celular e de colheita de plasma de membrana Perturbação
- Inoculação das células de levedura (por exemplo Saccharomyces cerevisiae) a partir de uma cultura D / N em 50 ml de meio nutriente completa de dextrose peptona de levedura (YPD). Inocular a uma densidade celular inicial baixo de 1-3 x 10 5 células / ml ou uma densidade óptica de aproximadamente 0,01 OD 600.
- Incubar as células a 30 ° C com agitação a 150 voltas por minuto (rpm) até que as células atingir a densidade máxima de células de cerca de 1 x 10 10 células, aproximadamente após 24 a 48 horas ou uma densidade óptica a 600 OD> 10.0.
NOTA: O tempo de cultura não é crucial como concentrações mais baixas de células pode ser utilizado.
- Incubar as células a 30 ° C com agitação a 150 voltas por minuto (rpm) até que as células atingir a densidade máxima de células de cerca de 1 x 10 10 células, aproximadamente após 24 a 48 horas ou uma densidade óptica a 600 OD> 10.0.
- Transferir a cultura superada para um tubo cónico de 50 ml, sedimentar as células por centrifugação a800 xg durante 3 minutos e descartar o sobrenadante.
- Lavar o sedimento com 25 ml de solução tampão de Tris-Cl 10 mM, EDTA 1 mM, pH 8,0, re-sedimentar as células por centrifugação a 800 xg durante 3 minutos e descartar o sobrenadante.
- Ressuspender o sedimento de células em 1,2 ml de tampão de ressuspensão fornecido a partir de um kit de plasmídeo coluna de purificação.
- Etapa opcional: Adicionar plasmídeos altamente diluídas como controles para a purificação de elementos circulares de DNA, 20.
NOTA: No conjunto de dados atual, uma mistura do plasmídeo 7.7 l foi aplicado para cada amostra contendo 10 a 10 células. A mistura da plasmídeo consistiu de três plasmídeos em diferentes concentrações; pBR322, a 38 ng / amostra, pUC19 a 0,5 ng / amostra, e pUG72 a 0,01 ng / amostra. - Transferir a suspensão celular em dois tubos de 2 ml de micro-centrífuga, cada um complementado com contas de vidro de 0,5 mm a uma razão de 1: 3 do volume total de suspensão.
- Vortex cada tubo à velocidade máxima por 10 min para perturbar células plasmáticasmembranas. Sedimentar as pérolas por centrifugação a 268 xg durante 30 seg e transferir os 1,2 ml de sobrenadante combinado a partir dos dois tubos de microcentrífuga para um novo tubo.
NOTA: Alternativa para o passo 1,6-1,7, usar zimoliase para romper as células em solução tampão 0,6 ml de ressuspensão. Dez unidades de zimoliase pode perturbar 5 x 10 7 células dentro de 1,5 h a 35 ° C.
2. EccDNA Enriquecimento por cromatografia em coluna
- Seguir o protocolo de um kit de purificação de plasmídeos coluna. Em resumo, o tratamento de cada amostra com uma solução de 1,2 ml alcalina, misture delicadamente e incubar 3 min a RT.
- Adicionar 1,2 ml de tampão de neutralização, misturar suavemente e centrifugar a 9650 xg durante 5 min.
- Carregar a solução numa coluna equilibrada com solução 1 ml de equilíbrio e permitir que o líquido a fluir através da coluna por gravidade.
- Lava-se a coluna com uma solução de lavagem de 4 ml. Quando a solução foi passada através da resina, adicionar cuidadosamente 0,3 ml de eluição assimlução para substituir a maior parte do volume de vazio de 0,35 ml de coluna.
- Elui-se o ADN para um novo tubo de recolha com uma solução de 1 ml de eluição e precipitar o ADN pela adição de 0,8 ml de mistura de precipitação. Centrifuga-se a 9650 xg durante 10 min.
- Lava-se a pelete de ADN com 0,5 ml de etanol a 70%, centrifugar a 9650 xg durante 5 min, de ar seco durante 5 a 15 minutos e dissolve-se o ADN purificado em 25 uL de água estéril.
NOTA: Apenas armazenamento a curto prazo de DNA em água é recomendado. Preferencialmente, seguir diretamente para a etapa 3.
3. A digestão do ADN cromossómico restante linear
- Passo opcional: Para facilitar a digestão específica de ADN linear pela exonuclease, tratar o ADN purificado com uma endonuclease rara de corte, tais como Notl. Para 5 ug de ADN, usar uma unidade de Notl, 5 ul de tampão de digestão de 10x e água estéril para um volume total de 50 ul. Incubar a reacção a 37 ° C durante 16 h e inactivar pelo calor a endonuclease a 80 ° C durante 5 min.
- Adicionar 20unidades de exonuclease (2 ul), 4 ul de ATP (25 mM), 34 ul de água estéril e 10 ul de 10x tampão de reacção directa com o ADN clivado-endonuclease de 50 ul para chegar a um volume de reacção de 1x de 100 ul, usando a exonuclease dependente de ATP kit.
- Realizar hidrólise de ADN de cadeia simples e de cadeia dupla linear, a 37 ° C durante 5 dias ou mais. Adicionar um ATP adicional de 4 mL (25 mM), 0,6 ul de 10x tampão de reacção e 20 unidades de exonuclease cada 24 horas para prosseguir a digestão enzimática do ADN a um volume de reacção 1x.
- Após a remoção do DNA linear, amostra de 2 ul da solução tratada com exonuclease para confirmar a eliminação do ADN cromossómico linear por reacção em cadeia da polimerase quantitativa (qPCR), usando um marcador cromossómico, tais como o gene da actina ACT1 20.
- Cada 20 ul qPCR volume reaccional contém 2 mL de amostra tratados com exonuclease, a 150 nm, os iniciadores 5'-ACT1 TCCGTCTGGATTGGTGGTTCTA-3 'e 5'-TGGACCACTTTCGTCGTATTC-3 ', 2% (volume / volume) de dimetil sulfóxido, e fluorescente mistura principal 10 ul verde.
- Utilizar a condição de reacção; 3 min a 95 ° C, seguido de 45 ciclos de 15 seg a 95 ° C e 30 segundos a 60 ° C.
NOTA: ACT1 é um marcador particularmente adequado para DNA linear desde variações no número de cópias deste gene são deletérios 21-23 tão eccDNA não deve levar ACT1. - Alternativas a análise da digestão de DNA por qPCR são PCR padrão (4.3) ou coloração com iodeto de propídio (4.4).
- Use 2 jil de amostra tratada com exonuclease como molde de PCR com os iniciadores 5'-ACT1 TGGATTCTGGTATGTTCTAGC-3 'e 5'-GAACGACGTGAGTAACACC-3'. Como controlo positivo ACT1, usar 50-100 ng S. genómico ADN cerevisiae como molde. condições de reacção de PCR; 3 min a 95 ° C, seguido de 35 ciclos de 30 seg a 95 ° C, 30 seg a 56 ° C e 1 min a 72 ° C.
- Executar reacções de PCR por gel electrophoreSIS em 1% de agarose com 0,5 ug / ml de brometo de etídio. Olhe para uma banda ACT1 0,8 kb.
- A ausência ou a presença de DNA linear também pode ser examinado por coloração com iodeto de propidio antes e depois de amplificação de DNA.
- Misturar cada amostra de ADN numa mistura 1: 1 de volume com uma mistura 1: H 2 O 1,000 solução diluída de estoque 20 mM de iodeto de propídio. Deixar solução no escuro durante 10-20 min à temperatura ambiente e analisar a coloração de ADN por microscopia de fluorescência de uma ampliação de 100X, utilizando um filtro de excitação de fluorescência vermelha em 663-738 nm e um tempo de exposição de 5 a 30 seg. Como controlo ADN-coloração, uso O29-ADN genómico amplificado a partir de levedura e / ou plasmídeo amplificado-O29.
- Inactivar pelo calor a solução da exonuclease a 70 ° C durante 30 min.
4. DNA Amplification
- Amplificar o purificado e enriquecido eccDNA a partir do passo 3.5) com DNA polimerase de O29 24-26 acordoção com o protocolo do fabricante polimerase.
- Em resumo, misture 5 ul enriquecidos eccDNA com tampão de desnaturação 5 mL.
- Depois de 3 minutos à temperatura ambiente, adicionar 10 ul de tampão de neutralização. Misturar suavemente e adicionar 30 ul de mistura mestre contendo tampão de reacção de 29 ul e 1 ul de polimerase de ADN O29. Incubar a reacção a 30 ° C durante 16 horas ou mais (até 72 h). Calor inactivar a polimerase de ADN de O29 a 65 ° C durante 3 min.
5. Seqüenciamento e Análise de Dados
- Tosquiarás o eccDNA amplificado com um ultrasonicator focada para um tamanho médio de pico alvo de 300 pb. Utilize as seguintes definições para uma amostra de DNA ul 130: poder 450W pico de intensidade, 60 seg tratamento, factor de utilização de 30%, a 200 ciclos por explosão, temperatura de 7 ° C.
- Adicione etiquetas de índice de código de barras e adaptadores para o fragmentado lê para a síntese de bibliotecas para sequenciação, utilizando um método adequado para a preparação da biblioteca.
- São profundas sequenciação, por exemplo, como 141-single nucleotide-fim lê numa plataforma de sequenciação de alto rendimento.
- Mapa lê o genoma de referência de levedura sob investigação e permitir que lê para mapear para várias regiões. Por exemplo, usar um sistema de fluxo de trabalho disponíveis gratuitamente 27,28 e curto leitura software de mapeamento alinhador 29.
- Identificar lê a partir de regiões de putativas eccDNAs contígua utilizando leituras, por exemplo, mais do que sete contígua lê (> 1 kb), sem lacunas 20.
NOTA: O software está disponível 27,28 para explorar mapeados lê em regiões genômicas de interesse.
Resultados
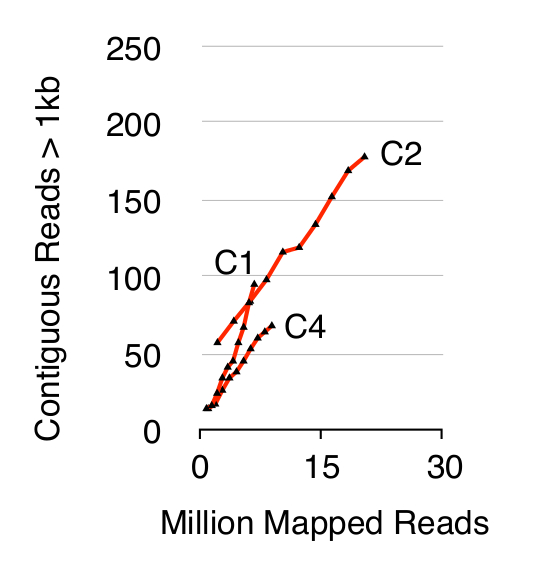
Para validar o método Circle-Seq, três S. populações cerevisiae CEN.PK de 1 x 10 10 células foram testadas depois as células foram cultivadas separadamente em meio YPD durante dez gerações. Eliminação cromossómica de ADN linear foi confirmada pela ausência de um sinal de qPCR ACT1 como descrito anteriormente 20 (dados não mostrados). eccDNA purificado e enriquecido foi sequenciado até 68 milhões lê (141 single-nucleotide-end lê) e mapeado para o genoma de referência CEN.PK113-7D (versão 19 de junho de 2012). As gravações de eccDNAs putativos das três amostras chamados C1, C2 e C4 foram atribuídos a regiões genómicas mapeados por contígua lê mais do que 1 kb. Com base em 10.000 simulações de Monte Carlo, o significado de cada região contígua lê mapeado por mais do que uma kb foi estimada. A partir deste regiões 79, 159 e 56 foram anotados como sequências eccDNA prováveis (p <0,1, Dataset 1). O número de contiguo gravadanos lê> 1 kb aumentou em função da profundidade de sequência, sugerindo que, mesmo mais elementos eccDNA teria sido registada se as amostras tinham sido sequenciados mais (Figura 2). Como esperado, o método de círculo-Seq extraída numerosos lê a partir de um número de elementos conhecidos circulares de DNA incluindo o plasmídeo 2μ, DNA mitocondrial, genes de ARN ribossomal no cromossoma XII, e os três plasmídeos de controlo interno pBR322, pUC19 e pUG72 que foram misturados em amostras apenas antes da purificação em coluna (Figura 3).
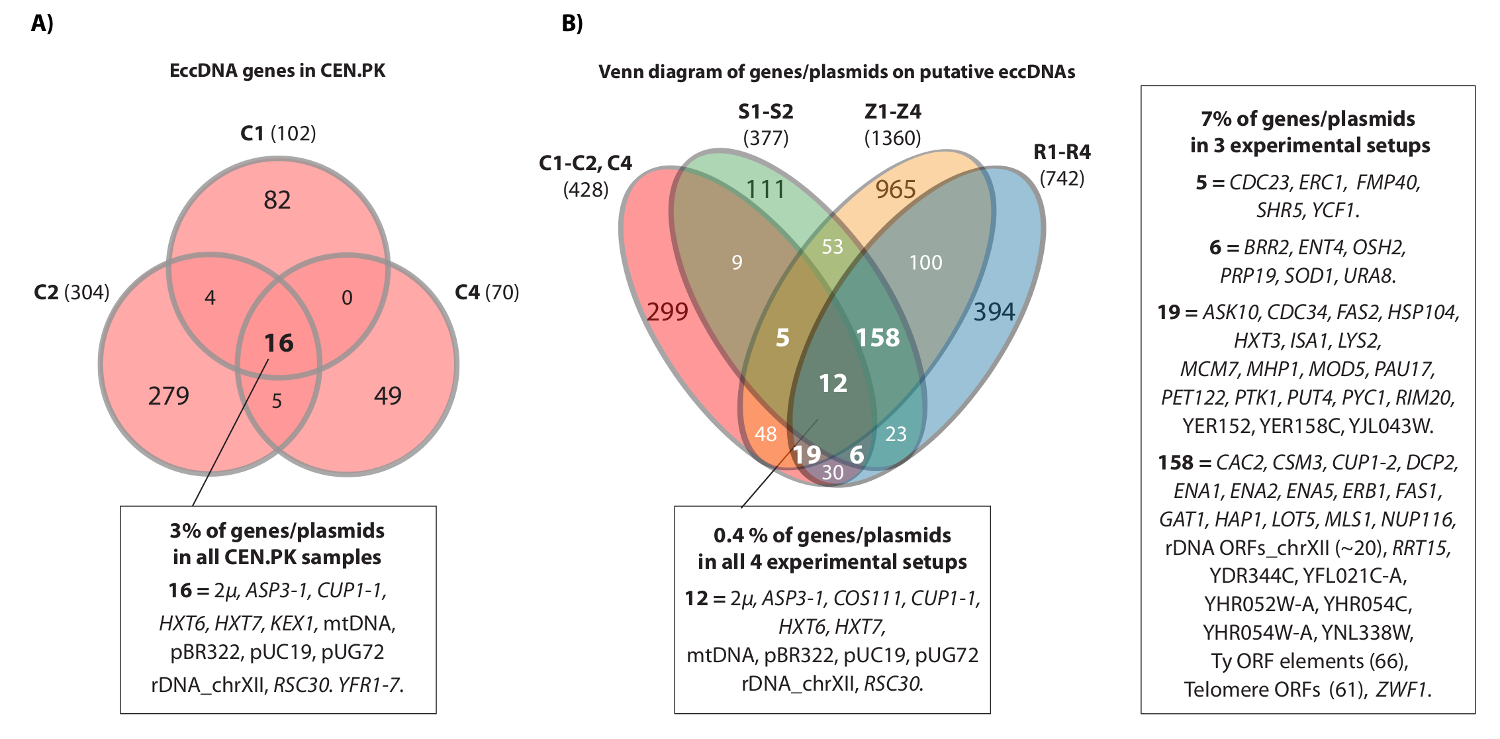
O vídeo mostra um exemplo de contígua lê que mapeada para o locus de HXT7 _ARS432_ HXT6 no cromossoma IV. Anteriormente, o [HXT6 / 7 círculo] foi detectada por Círculo-Seq em dez populações S288C (cada um com 1 x 10 10 células) e a estrutura de ADN circular foi confirmada por análise de PCR inverso 20. A ci [HXT6 / 7rcle] também foi registada em cada uma das três populações CEN.PK (Figura 4A). Além disso, a maioria dos genes comuns eccDNA entre as amostras replicadas de CEN.PK sobreposto eccDNA genes a partir de conjuntos de dados S288C (Figura 4B).
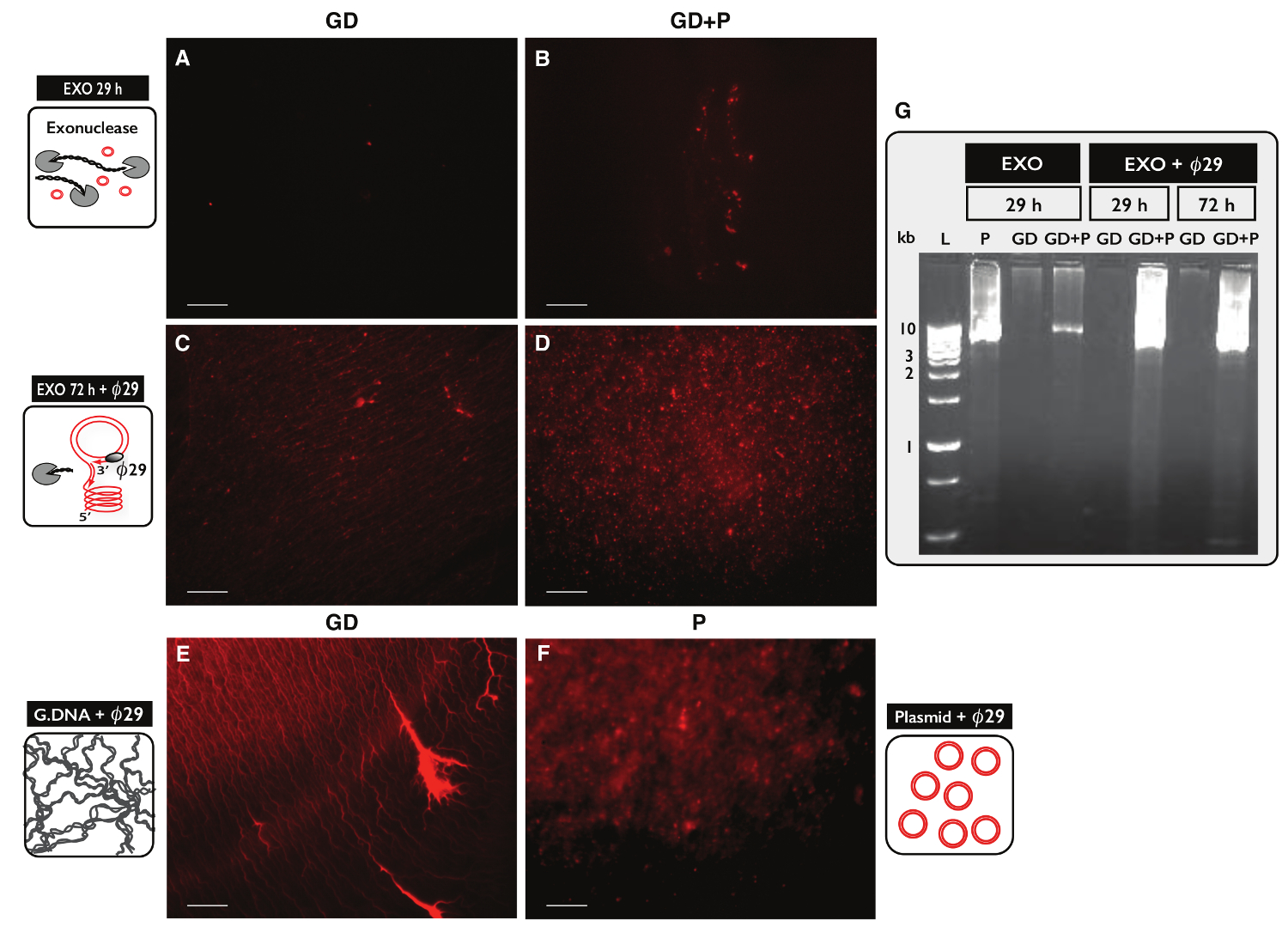
Para testar a especificidade do protocolo Círculo-Seq para a purificação de ADN circular, de duas amostras, cada uma com o ADN genómico de 30 ug, foram testados. Uma amostra foi suplementado com 100 ng de DNA plasmídeo e eccDNA de ambas as amostras foram purificadas pelo protocolo Círculo-Seq. Após separação em coluna, o rendimento de ADN foi de 1,27% (380 ng) para a amostra sem plasmídeo (GD) e 1,60% (480 ng) para a amostra com o plasmídeo (GD + P). A eficiência do tratamento de exonuclease foi testado para o teor de ADN linear após 29 h e 72 h utilizando PCR contra ACT1. Sem amostras continham amplificado ACT1 (dados não mostrados). Uma fracção de cada amostra tratada com exonuclease foi ainda umamplified pelo O29 polimerase e os produtos de reacções enzimáticas foram analisadas por coloração com iodeto de propídio (Figura 5A-F) e electroforese em gel de agarose (Figura 5G). As amostras após o tratamento de exonuclease mostrou mínima de propídio iodo-mancha (Figura 5A-B). O O29 - amostra amplificada com apenas ADN genómico revelou estruturas fio (Figura 5C) semelhantes à amostra controle (Figura 5E). O O29 - amostra amplificada que tinha plasmídeo acrescentou revelou focos (Figura 5D) assemelham-se o controle de plasmídeo (Figura 5F). As imagens indicou que O29 polimerase de ADN enriquecido para circular sobre o ADN linear. A maioria do ADN cromossómico linear foi removido a partir de amostras após o tratamento 29 horas exonuclease (Figura 5A-B, G). No entanto, a extensa tratamento exonuclease por mais de 100 horas e usando mais de 100 unidades foi necessária para remover todo o ADN cromossómico linear, como O29 - amostras amplificadas ainda mostrou um fundo de estruturas em forma de fio após 72 horas de tratamento com exonuclease (Figura 5C-D).

. Figura 1. Esquema do método de círculo-Seq O protocolo possui 5 etapas: 1) cultura das células, 2) purificação e do enriquecimento de eccDNA por cromatografia em coluna, 3) digestão de ADN cromossómico restante linear da fracção de eluato, 4) de amplificação ADN por O29 a polimerase de ADN, e 5) a sequenciação de eccDNA altamente enriquecido e mapeamento das leituras para a S. genoma de referência cerevisiae. Por favor clique aqui para ver uma versão maior desta figura.
{kind=link}
p_upload / 54239 / 54239fig2.jpg "/>
Figura 2. Adjacente lê> 1 kb como função da profundidade sequência. EccDNA a partir de 1 x 10 10 células aumentam em função da profundidade de sequência (em milhões de mapeado lê). Mostrado: triplicados biológicos de haplóides CEN.PK S. cerevisiae populações (C1, C2, C4) separados por 10 a 10 divisões celulares. Por favor clique aqui para ver uma versão maior desta figura.
{kind=link}

Figura 3. Detecção de elementos de DNA circular conhecidos. (AB) Espalhe parcelas de cobertura de leitura (ler densidade) em por cento para plasmídeos em CEN.PK biológica replica C1, C2 e C4. (A) lê mapeada para os plasmídeos de levedura endógenos foram: 2μ; [Círculo rDNA] (genes de ARN ribossómico de cromossoma XII); e mtDNA (o ADN mitocondrial). (B) original lê mapeado para controlar plasmídeos. plasmídeos de controlo foram doseadas em amostras antes da purificação em coluna. rácios de plasmídeo por célula foram: pBR322 (mais sinais) 1: 1, pUC19 (círculos) 1:50, e pUG72 (triângulos) 1: 2500.

Figura 4. Elementos eccDNA comum em CEN.PK e S288c. (A) diagrama de Venn exibindo sobreposição entre os 476 genes em 294 eccDNA elementos nas três amostras CEN.PK (C1, C2, C4). Os 16 sobrepostas genes eccDNA / plasmídeos comuns são anotados (todos os nomes de genes estão em Dataset 1). (B) diagrama de Venn de todos os genes registados em eccDNAs putativos das três amostras CEN.PK (C1, C2, C4), em comparação com todos os genes registados em eccDNAs putativas a partir de 10 amostras de S288C: S1-S2, R1-R4, Z1-Z4 (ver referência 20). São mostrados 13 repetições biológicas (S1-S2, R1-R4, Z1-Z4, C1-C3) com genes / plasmídeos e regiões eccDNA putativos que se sobrepunham um mínimo de 2 estirpe origens e 3 ou mais experimentais configurações. As amostras C, CEN.PK; amostras R e Z, S288c BY4741; Amostras S, S288c M3750. Por favor clique aqui para ver uma versão maior desta figura.
{kind=link}

Figura 5. A visualização das amostras de ADN depois de exonuclease e O 2 9 de tratamento. (AF) de iodeto de propídio a coloração do ADN. Barra de escala, 10 | im. (A, C e E) com amostras de ADN genómico (GD); (B e D) GD amostras com mais de plasmídeo (GD + P). ( ong> AB) Após o tratamento exonuclease 29 hr (EXO 29 h); (CD) após o tratamento com exonuclease 72 h seguido por O29 a amplificação com polimerase (exo 72 H +) O29. (E) o controle DNA genômico depois e: a amplificação da polimerase O29; (F) Controlo de plasmídeo (5,5 kb) depois amplificação 29 polimerase; (Ø G) agarose gel-eletroforese. A partir da esquerda: L, 1 marcadores kb; P, o controlo de plasmídeo (5,5 kb) depois EXO 29 h; GD, após EXO 29 h (amostra, tal como em A); GD + P, depois de EXO 29 hr (amostra como B); GD e GD + P, depois de EXO 29 hr + O29; GD e GD + P, depois de EXO 72 hr + O29 (amostra como no CD). Veja a Tabela S1 para fornecer mais detalhes. Por favor clique aqui para ver uma versão maior desta figura.
{kind=link}
Aset 1 "src =" / files / ftp_upload / 54239 / 54239dataset1.jpg "/>
Conjunto de Dados 1. regiões DNA circularização potenciais em CEN.PK. Por favor clique aqui para baixar esse arquivo.
São mostrados os dados de sequências e análises para 348 regiões. As colunas são AD, mapeamento eccDNA. A (primeira coluna a partir da esquerda), a amostra a partir do qual foi identificado eccDNA putativo; B, cromossoma; CD, início e fim coordenadas eccDNAs putativos. EH, o conteúdo eccDNA. E, sequência de replicação autónoma (ARS) na região; F, gene completo da região; L, parte do gene incluído na região; H, BLASTN do gene identificado. IO, a cobertura EccDNA e valores de p. I, região mais longo com uma sequência anotada exclusivamente no pb; J, número de todos mapeados lê; K, a cobertura de todos mapeados lê por fragmentos por kb a partir de um milhão mapeado lê (FPKM); L, p-valor para o putativo eccDNA comparado a ocorrência por acaso a partir de simulações de Monte Carlo; M, número de uniquely mapeados lê; NÃO; como K e L utilizando apenas mapeados exclusivamente lê (UFPKM). Parâmetros para mapeamento de leituras e simulações de Monte Carlo foram descrito 20.
Discussão
O método Circle-Seq permite a detecção escala genoma de eccDNA a partir de células de levedura com resolução de nível sequência. O método é uma purificação eccDNA leve que não requer vórtice intensiva ou pipetagem e usa a separação da coluna pela força da gravidade para limitar eccDNA ruptura que levaria a digestão de exonuclease no passo subsequente. Estas características do método pode ser crucial para a detecção de grandes eccDNAs que contêm sequências de genes. Circle-Seq detectado numerosas eccDNAs incluindo genes completos (Conjunto de Dados 1). Ele também detectou a 86 kb levedura DNA mitocondrial. Assim, esse protocolo facilita a purificação de grandes elementos circulares de DNA. Mantendo o número de passos de extracção de ADN para um mínimo reduz o risco de perda eccDNA e maximiza o rendimento. Com base nos resultados de controlo, cravado-em plasmídeos, Círculo-Seq é altamente sensível, a detecção de um único ADN circular de 2.500 células. Além disso, a remoção de plasmídeos endógenos abundantes tais como 2μ; plasmídeo ou ADN mitocondrial pode melhorar significativamente a sensibilidade. Cura de 2μ a partir de culturas de levedura foi descrito 30. Alternativamente, 2μ e remoção do DNA mitocondrial pode ser alcançada com uma endonuclease rara de corte, tais como SwaI. No entanto, o passo enzima de restrição poderia ter como alvo outras eccDNAs de interesse e limitar o rendimento total eccDNA.
As etapas críticas para a detecção eccDNA foram remoção de ADN linear (passo 3) e sequenciação de DNA (passo 5) a uma profundidade adequada. Para gravar a maioria dos eccDNAs a partir de uma população de células, sequenciamento de profundidade pode ser necessária 20. sequenciamento-end emparelhado deve fornecer uma confiança ainda maior de detecção eccDNA, como se espera junções circulares de DNA para produzir emparelhado-end lê esse mapa discordantemente. Estas discrepâncias suporta a descoberta de estruturas de ADN circular e pode, potencialmente, ser utilizado como um filtro de detecção de eccDNA adicional.
O Círculo-Semétodo q foi validado utilizando três S. independente populações cerevisiae CEN.PK. Sequências detectadas incluído anteriormente relatado eccDNAs, plasmídeos endógenos e cravado-in plasmídeos e centenas de eccDNAs putativos (Conjunto de Dados 1). Estes resultados suportam conjuntos de dados Círculo-Seq anteriores de S. cerevisiae S288c 20. A descoberta de vários eccDNAs comuns para as populações CEN.PK S288C e indica que estes loci têm uma propensão para existir como elementos circulares (Figura 4). Temos anteriormente demonstrado que o [círculo GAP1] é enriquecido em condições limitadas de nitrogênio no fundo CEN.PK 8, embora não tenha sido encontrada evidência de [círculo GAP1] em outra estirpe fundos. Encontrar de eccDNA do CUP1-1 RSC30, ASP3-1, COS111 e HXT6 HXT7 loci tanto S288c e CEN.PK sugere que uma predisposição para a circularização do DNA é conservido entre as estirpes de levedura. Ele continua a ser mostrado se [HXT6 / 7 círculo], [círculo ASP3-1], [círculo COS111] e [círculo CUP1-1 RSC30] conferem vantagens selectivas para células ou se a sua existência é meramente um efeito de altas taxas de circularização do ADN.
Tomados em conjunto, os resultados indicam que o círculo-Seq é bem adequado para detectar eccDNAs quilobases de tamanho e tem vantagens para a identificação de genes completos com eccDNAs. Circle-Seq é um método altamente sensível que permite telas inteiras escala genoma de eccDNAs de levedura. O método Circle-Seq poderia abrir um novo campo de investigação destinada a elucidar o papel de eccDNA na geração de deleções de genes e amplificações. Dado que a arquitetura e estrutura do DNA são em grande parte conservada desde levedura eucariótica para eucariotas superiores, o método Circle-Seq deve, em princípio, ser applicablE para todas as células eucarióticas, com ligeiras modificações. Actualmente, o método não parece ter quaisquer limitações, embora a sua capacidade de purificar eccDNAs tamanho Megabase-se ainda a ser mostrado. Além disso, a utilização de O29 ADN-polimerase, que utiliza um método de amplificação de rolamento-círculo 31, cria uma tendência para as pequenas eccDNAs eccDNA tornando mais difícil a quantificação. Circle-Seq detecta eccDNAs grandes o suficiente para transportar genes completos, tornando-o adequado para estudos de DNA minutos circular dupla a partir de células somáticas humanas. Minutos duplas podem contribuir para o câncer quando proto-oncogenes são amplificados sobre esses elementos 32-37. Estudos de eccDNAs em células germinativas poderia ser usado para medir as taxas de mutação germinativa e avaliar a qualidade do esperma, por exemplo, na pecuária. Assim, Círculo-Seq tem o potencial para produzir insights sobre a taxa a que a variação genética surge sob a forma de variação do número de cópias, e levam a uma nova compreensão das doenças que envolvem contra cópia do genevariação do número 38-40.
Divulgações
The authors declare that they have no competing financial interests.
Agradecimentos
Thanks to Kenn D. Møller and Claus Sternberg (DTU) for technical assistance and to Tue S. Jørgensen for quantitative PCR analysis.
Materiais
| Name | Company | Catalog Number | Comments |
| Bacto peptone | BD Difco | 211677 | Alternative product can be used. |
| Brilliant III SYBR Green PCR Master Mix | Agilent Technologies | 600882 | For qPCR analysis. Alternative product can be used. |
| Dextrose (D-glucose) | Carl Roth | HN06.4 | Alternative product can be used. |
| Disruptor Beads, 0.5 mm | Scientific Industries, Inc. | SI-BG05 | Glass beads to disrupt plasma cell membranes. Alternative product can be used. |
| Ethidium bromide | Carl Roth | 2218.2 | Agarose gel stain for detecting DNA/RNA. |
| GeneJet plasmid miniprep kit | Thermo Fisher | K0502 | Plasmid purifcation from bacteria. Alternative product can be used |
| NotI, FastDigest | Life Technologies - Thermo Fisher Scientific, USA | FD0594 | Endonuclease. Alternative product can be used. |
| Plasmid Mini AX kit | A&A Biotechnology, Poland | 010-50 | Plasmid purifcation kit used to purify eccDNA. |
| Plasmid-Safe ATP-dependent DNase kit | Epicentre, USA | E3105K | ATP-dependent exonuclease kit. Alternative product can be used. |
| Propidium iodide | Sigma-Aldrich, USA | 81845 | Alternative product can be used. |
| pUG6 plasmid | EUROSCARF, Germany | P30114 | Marker gene: loxP-PAgTEF1-kanMX-TAgTEF1-loxP. Plasmid requests: Please contact Dr. Peter Philippsen@unibas.ch |
| QIAGEN genomic-tip 100/G | Qiagen, USA | 13343 | Genomic DNA purifcation from yeast. Alternative product can be used. |
| REPLI-g Mini Kit protocol | Qiagen, USA | 150023 | Amplification of eccDNA by the phi29 polymerase |
| Yeast extract | BD Difco | 210929 | Alternative product can be used. |
| Zymolyase 100T (Lyticase, Yeast Lytic Enzyme) | Nordic BioSite, Sweden | Z1004-3 | Alternative product can be used. |
| Data access to sequence files | European Nucleotide Archive | EccDNA dataset from Saccharomyces cerevisiae CEN.PK113-7D. Study accession number PRJEB9684. 2nd accession number is ERP010820. Locus tag prefix is BN2032. | |
| Strains | |||
| Saccharomyces cerevisiae CEN.PK113-7D | Genotype MATa MAL2-8c SUC2 | ||
| Saccharomyces cerevisiae yeast deletion library pool | EUROSCARF, Germany | S288c BY4741 pool of 4400 viable single-gene deletion mutants disrubted by KanMX module. Genotypes MATa his3∆1 leu2∆0 met15∆0 ura3∆0 genexxx::KanMX. | |
| Equipments | |||
| DNA Spectrophotometer | NanoDrop 1000 Spectrophotometer, Thermo Fisher | Measuring DNA concentration. Alternative product can be used. | |
| Fluorescence microscopy | Nikon Optronics Magnafire. Red excitation fluorescence filter, 663-738 nm. | Alternative product can be used. | |
| Robotic library-build system | Apollo 324, IntegenX Inc. | DNA library preparation. Alternative product can be used. | |
| Sequencing platform | Illumina HiSeq 2000 platform, Illumina Inc. | DNA sequencing. Alternative product can be used. | |
| Ultrasonicator | Covaris LE220, microTUBE AFA Fiber tubes | Alternative product can be used. | |
| Methods | |||
| 2% YPD media | Mix 10 g Dextrose, 10 g Yeast extract, 20 g Bacto peptone and add H2O to a total volume of 1000 ml and autoclave. | ||
| Circle-Seq test on genomic DNA | Genomic DNA was purified (Qiagen) from a pool of the yeast deletion library (Euroscarf). The DNA concentration was measured by nanodrop and 30 µg genomic DNA was pipetted into two micro centrifuge tubes. One micro centrifuge tube was supplemented with 100 nanogram plasmid (pUG6). The DNA samples were purified by Circle-Seq, omitting the protocol steps 1.1-1.3 and 1.5-1.7. The eluted DNA concentrations were measured by nanodrop and the entire DNA yield from sample GD and GD+P was treated with exonuclease for a period of 29 hours. A 10% fraction was collected for phi29-amplification and PCR analysis, while the remaining DNA was subjected to 72 hour exonuclease treatment. The samples were analyzed for linear DNA content by PCR, using the ACT1 gene as chromosomal marker. A 5% fraction of each of the exonuclease treated samples was amplified by the phi29 DNA polymerase for 16 hours (Qiagen). The presence of DNA in each sample was examined by loading an equal amount (7 µl) in wells on an 0.5 µg/ml ethidium-bromide 0.9% agarose gel after running gel-electrophoresis. | ||
| Mapping software | Bowtie2 aligner, John Hopkins University | Ultrafast short read alignment. Reference: Langmead B, Salzberg S. Fast gapped-read alignment with Bowtie 2. Nature Methods. 2012, 9:357-359. | |
| Propidium iodide stain | Images of propidium iodine stained DNA were captured by fluorescence microscopy at 100x magnification (100x/1.30 oil, Nikon) in the RFP channel (red excitation fluorescence filter, 663-738 nm) using identical exposition time (5 seconds). | ||
| Workflow bioinformatic system | Galaxy, Open source. | A free web-based platform for data intensive biomedical research. References: Goecks, J, Nekrutenko, A, Taylor, J and The Galaxy Team. Galaxy: a comprehensive approach for supporting accessible, reproducible, and transparent computational research in the life sciences. Genome Biol. 2010 Aug 25;11(8):R86. Blankenberg D, Von Kuster G, Coraor N, Ananda G, Lazarus R, Mangan M, Nekrutenko A, Taylor J. "Galaxy: a web-based genome analysis tool for experimentalists". Current Protocols in Molecular Biology. 2010 Jan; Chapter 19:Unit 19.10.1-21. Giardine B, Riemer C, Hardison RC, Burhans R, Elnitski L, Shah P, Zhang Y, Blankenberg D, Albert I, Taylor J, Miller W, Kent WJ, Nekrutenko A. "Galaxy: a platform for interactive large-scale genome analysis." Genome Research. 2005 Oct; 15(10):1451-5. | |
Referências
- Kugelberg, E., Kofoid, E., et al. The Tandem Inversion Duplication in Salmonella enterica.: Selection Drives Unstable Precursors to Final Mutation Types. Genetics. 185 (1), 65-80 (2010).
- Reams, A. B., Kofoid, E., Savageau, M., Roth, J. R. Duplication Frequency in a Population of Salmonella enterica. Rapidly Approaches Steady State With or Without Recombination. Genetics. 184 (4), 1077-1094 (2010).
- Smith, C. A., Vinograd, J. Small polydisperse circular DNA of HeLa cells. Journal of Molecular Biology. 69 (2), 163-178 (1972).
- Carroll, S. M., DeRose, M. L., et al. Double Minute Chromosomes Can Be Produced from Precursors Derived from a Chromosomal Deletion. Molecular and cellular biology. 8 (4), 1525-1533 (1988).
- Cohen, S., Yacobi, K., Segal, D. Extrachromosomal Circular DNA of Tandemly Repeated Genomic Sequences in Drosophila. Genome research. 13 (6A), 1133-1145 (2003).
- Horowitz, H., Haber, J. E. Identification of Autonomously Replicating Circular Subtelomeric Y' Elements in Saccharomyces cerevisiae. Molecular and cellular biology. 5 (9), 2369-2380 (1985).
- Moore, I. K., Martin, M. P., Dorsey, M. J., Paquin, C. E. Formation of Circular Amplifications in Saccharomyces cerevisiae by a Breakage-Fusion-Bridge Mechanism. Environmental and molecular mutagenesis. 36 (2), 113-120 (2000).
- Gresham, D., Usaite, R., Germann, S. M., Lisby, M., Botstein, D., Regenberg, B. Adaptation to diverse nitrogen-limited environments by deletion or extrachromosomal element formation of the GAP1 locus. Proceedings of the National Academy of Sciences of the United States of America. 107 (43), 18551-18556 (2010).
- Windle, B., Draper, B. W., Yin, Y. X., O'Gorman, S., Wahl, G. M. A central role for chromosome breakage in gene amplification, deletion formation, and amplicon integration. Genes & development. 5 (2), 160-174 (1991).
- Gresham, D., Ruderfer, D. M., et al. Genome-Wide Detection of Polymorphisms at Nucleotide Resolution with a Single DNA Microarray. Science. 311 (5769), 1932-1936 (2006).
- Kidd, J. M., Cooper, G. M., et al. Mapping and sequencing of structural variation from eight human genomes. Nature. 453 (7191), 56-64 (2008).
- Gresham, D., Desai, M. M., Botstein, D., Dunham, M. J. The Repertoire and Dynamics of Evolutionary Adaptations to Controlled Nutrient-Limited Environments in Yeast. PLoS Genetics. 4 (12), 1-19 (2008).
- Lang, G. I., Botstein, D., Desai, M. M. Genetic Variation and the Fate of Beneficial Mutations in Asexual Populations. Genetics. 188 (3), 647-661 (2011).
- van Loon, N., Miller, D., Murnane, J. P. Formation of extrachromosomal circular DNA in HeLa cells by nonhomologous recombination. Nucleic Acids Research. 22 (13), 2447-2452 (1994).
- Vinograd, J., Lebowitz, J. Physical and Topological Properties of Circular Dna. Journal of General Physiology. 49 (6P2), 103 (1966).
- Shibata, Y., Kumar, P., et al. Extrachromosomal MicroDNAs and Chromosomal Microdeletions in Normal Tissues. Science. 336 (6077), 82-86 (2012).
- Dillon, L. W., Kumar, P., et al. Production of Extrachromosomal MicroDNAs Is Linked to Mismatch Repair Pathways and Transcriptional Activity. Cell Reports. 11 (11), 1749-1759 (2015).
- Li, L. L., Norman, A., Hansen, L. H., Sørensen, S. J. Metamobilomics - our knowledge on the pool of plasmid encoded traits in natural environments using high-throughput sequencing. Clinical microbiology and infection : the official publication of the European Society of Clinical Microbiology and Infectious Diseases. 18, 5-7 (2012).
- Brown Kav, A., Sasson, G., Jami, E., Doron-Faigenboim, A., Benhar, I., Mizrahi, I. Insights into the bovine rumen plasmidome. Proceedings of the National Academy of Sciences of the United States of America. 109 (14), 5452-5457 (2012).
- Møller, H. D., Parsons, L., Jørgensen, T. S., Botstein, D., Regenberg, B. Extrachromosomal circular DNA is common in yeast. Proceedings of the National Academy of Sciences of the United States of America. , 201508825 (2015).
- Drubin, D. G., Miller, K. G., Botstein, D. Yeast Actin-Binding Proteins - Evidence for a Role in Morphogenesis. The Journal of cell biology. 107 (6), 2551-2561 (1988).
- Magdolen, V., Drubin, D. G., Mages, G., Bandlow, W. High levels of profilin suppress the lethality caused by overproduction of actin in yeast cells. FEBS letters. 316 (1), 41-47 (1993).
- Sandrock, T. M., Brower, S. M., Toenjes, K. A., Adams, A. Suppressor analysis of fimbrin (Sac6p) overexpression in yeast. Genetics. 151 (4), 1287-1297 (1999).
- Blanco, L., Bernad, A., Lázaro, J. M., Martìn, G., Garmendia, C., Salas, M. Highly Efficient DNA Synthesis by the Phage ø29 DNA Polymerase. The Journal of biological chemistry. 264 (15), 8935-8940 (1989).
- Dean, F. B. Rapid Amplification of Plasmid and Phage DNA Using Phi29 DNA Polymerase and Multiply-Primed Rolling Circle Amplification. Genome research. 11 (6), 1095-1099 (2001).
- Hutchison, C. A., Smith, H. O., Pfannkoch, C., Venter, J. C. Cell-free cloning using ø29 DNA polymerase. Proceedings of the National Academy of Sciences of the United States of America. 102 (48), 17332-17336 (2005).
- Goecks, J., Nekrutenko, A., Taylor, J., Galaxy Team, T. Galaxy: a comprehensive approach for supporting accessible, reproducible, and transparent computational research in the life sciences. Genome Biology. 11 (8), 86 (2010).
- Giardine, B., Riemer, C., et al. Galaxy: A platform for interactive large-scale genome analysis. Genome research. 15 (10), 1451-1455 (2005).
- Langmead, B., Salzberg, S. L. Fast gapped-read alignment with Bowtie 2. Nature Methods. 9 (4), 357 (2012).
- Tsalik, E. L., Gartenberg, M. R. Curing Saccharomyces cerevisiae of the 2 micron plasmid by targeted DNA damage. Yeast. 14 (9), 847-852 (1998).
- Norman, A., Riber, L., Luo, W., Li, L. L., Hansen, L. H., Sørensen, S. J. An Improved Method for Including Upper Size Range Plasmids in Metamobilomes. PLoS ONE. 9 (8), e104405 (2014).
- Storlazzi, C. T., Lonoce, A., et al. Gene amplification as double minutes or homogeneously staining regions in solid tumors: Origin and structure. Genome research. 20 (9), 1198-1206 (2010).
- Von Hoff, D. D., Needham-VanDevanter, D. R., Yucel, J., Windle, B. E., Wahl, G. M. Amplified human MYC localized to replicating submicroscopic circular DNA molecules. Proceedings of the National Academy of Sciences of the United States of America. 85 (13), 4804-4808 (1988).
- Raymond, E., Faivre, S., et al. Effects of hydroxyurea on extrachromosomal DNA in patients with advanced ovarian carcinomas. Clinical cancer research : an official journal of the American Association for Cancer Research. 7 (5), 1171-1180 (2001).
- Shimizu, N. Extrachromosomal Double Minutes and Chromosomal Homogeneously Staining Regions as Probes for Chromosome Research. Cytogenetic and genome research. 124 (3-4), 3-4 (2009).
- Eckhardt, S. G., Dai, A., Davidson, K. K., Forseth, B. J., Wahl, G. M., Von Hoff, D. D. Induction of differentiation in HL60 cells by the reduction of extrachromosomally amplified c-myc. Proceedings of the National Academy of Sciences of the United States of America. 91 (14), 6674-6678 (1994).
- Vogt, N., Lefèvre, S. -. H., et al. Molecular structure of double-minute chromosomes bearing amplified copies of the epidermal growth factor receptor gene in gliomas. Proceedings of the National Academy of Sciences of the United States of America. 101 (31), 11368-11373 (2004).
- Ahn, K., Gotay, N., et al. High rate of disease-related copy number variations in childhood onset schizophrenia. Molecular psychiatry. 19 (5), 568-572 (2013).
- Girirajan, S., Johnson, R. L., et al. Global increases in both common and rare copy number load associated with autism. Human molecular genetics. 22 (14), 2870-2880 (2013).
- Vogt, N., Gibaud, A., Lemoine, F., de la Grange, P., Debatisse, M., Malfoy, B. Amplicon rearrangements during the extrachromosomal and intrachromosomal amplification process in a glioma. Nucleic Acids Research. , (2014).
Reimpressões e Permissões
Solicitar permissão para reutilizar o texto ou figuras deste artigo JoVE
Solicitar PermissãoThis article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. Todos os direitos reservados