Method Article
טיהור הגנום כולו של DNA המעגל extrachromosomal מתאי איקריוטיים
In This Article
Summary
This paper presents a sensitive method called Circle-Seq for purifying extrachromosomal circular DNA (eccDNA). The method encompasses column purification, removal of remaining linear chromosomal DNA, rolling-circle amplification and high-throughput sequencing. Circle-Seq is applicable to genome-scale screening of eukaryotic eccDNA and studying genome instability and copy-number variation.
Abstract
DNAs העגול extrachromosomal (eccDNAs) הם מרכיבים גנטיים נפוצים שמר אפייה ומדווח אאוקריוטים אחרים גם כן. EccDNAs לתרום השונות הגנטית בין תאים סומטיים יצורים רב-תאיים ו לאבולוציה של אאוקריוטים unicellular. רגישות שיטות לאיתור eccDNA נדרשות להבהיר כיצד אלמנטים אלה משפיעים על יציבות הגנום וכיצד סביבתי וביולוגי גורם לגרום להיווצרות שלהם בתאים איקריוטיים. וידאו זה מציג שיטת eccDNA טיהור רגישה שנקראה המעגל-Seq. השיטה כוללת טיהור עמודה של DNA מעגלי, הסרת הנותרים דנ"א כרומוזומלי ליניארי, הגברה מתגלגל מעגל eccDNA, רצף עמוק, ומיפוי. טיפול exonuclease נרחב נדרש שפלת DNA כרומוזומלי ליניארי מספיק. שלב ההגברה מתגלגל מעגל על ידיine-גובה:. נורמלי; "> φ 29 פולימראז מועשר עבור DNA מעגלי מעל DNA ליניארי אימות של שיטת מעגל-Seq על שלוש אוכלוסיות cerevisiae ס CEN.PK 10 10 תאים זוהה מאות פרופילים eccDNA בגדלים גדולים מ -1 kilobase . ממצאים חוזרים של ASP3-1, COS111, CUP1, RSC30, HXT6, HXT7 גנים על DNA המעגלי בשני S288c ו CEN.PK עולה כי circularization DNA נשמר בין זנים ב לוקוסים אלה. לסיכום, השיטה-Seq המעגל יש רחבה תחולה להקרנה בקנה מידה הגנום עבור eccDNA אאוקריוטים וכן לאיתור סוגי eccDNA ספציפיים.
Introduction
זיהוי הגברת כרומוזומליות מוקדם או חולפת קשה מכיוון שהיא דורשת זיהוי שינויים במולקולות דנ"א יחידים באוכלוסיות גדולות של תאים. וריאציות עותק-מספר כרומוזומלית (CNVs) מזוהות היטב בדרך כלל לאחר הקמתם, ומשאירות רק את מבנה CNV הסופי כעדות המנגנון שהפיק את הווריאציה 1,2. איתור מחל DNA המעגל extrachromosomal (eccDNA) בשלבים מוקדמים של היווצרות CNV עשוי להבהיר תהליכים מתמשכים rearrangements גנומית.
בעבר, דה נובו גילוי eccDNA היה על ידי micrographs אלקטרונים 3, מכתים Giemsa של הכרומוזומים metaphase 4, או 5 ג'ל אלקטרופורזה דו מימדי. שיטות אלה מספקים מידע מועט, אם בכלל על הרצף של DNA המעגלי. טכניקות ממוקדות כגון הדרום סופג 6,7, ההופכי 8 PCR, או קרינת hybridizatio באתרוn 9 לספק ראיות רק על אלמנטים eccDNA ספציפיים. אף אחת מהשיטות הללו מספקים את הרצף מכל סוגי eccDNA קיימים באוכלוסיית תא.
הבדלי הגנום בתוך שלולית של תאים יכולים להיות מאופיינים על ידי רצף הגנום ו / או מערכי ריצוף 10,11. זיהוי מחיקה או הגברה בשיטות טיהור DNA קונבנציונליות בדרך כלל דורש כי אלל מוטנטי לייצג לפחות 0.1-1% של תא האוכלוסייה 12,13. Acentric eccDNAs צפויים להיות עוד יותר חולף בתוך תרבית תאים עקב חוסר צנטרומר והיעדרות הפוטנציאל שלהם של סינתזת ה- DNA על שכפול. לפיכך, מאחר שרוב eccDNAs הנראה הנם בסכומים נמוכים הרצפים שלהם מזכירים את הגנום, שיטות מיצוי DNA חלופה נדרשות לזהות eccDNAs.
כמה טכניקות טיהור עגולות DNA לנצל את ההבדלים המבניים בין כרומוזומים דנ"א מעגלי. למשל, במהירות גבוהה ultracentrifugation הדרגתי-צזיום כלוריד ממשמש לבודד 350-3000 basepairs (נ"ב) eccDNAs הגדול מקו התאים הסרטני האנושי הלה 14. עם זאת, מהירות גבוהה יכולה לשבור או ניק עמוד השדרה של מבני DNA המעגלים supercoiled, לשנות את מהירות שקיעת 15 ותשואת eccDNA. דוטה ועמיתים לעבודה פיתחו שיטה דה נובו, זיהוי מידה הגנום של ה- DNA מעגלי מרקמות העכבר כמו גם מתרבויות של עוף ותאי אדם 16,17. השיטה שלהם היא מיצוי של גרעינים מרקמות הומוגני ידי ultracentrifugation סוכרוז ואחריו טיהור פלסמיד וכמה סיבובים של תגובות אנזימטיות ו עקירות DNA. פרוטוקול שלהם בעיקר מזהה 200-400 eccDNAs נ"ב, microDNAs שנקרא. גם דוטה ועמיתים לעבודה טיהור ניסיון microDNAs מ שמר האפייה, אבל לא הצליחו להקליט microDNA ממינים שמרים זה 16.
פיתחנו שיטה חדשהזיהוי דה נובו של eccDNA משמרים שנקרא מעגל-Seq. שיטה זו מאפשרת סקרים בקנה מידה הגנום עבור מולקולות DNA מעגליות גדולות מספיק כדי לשאת גנים שלמים וגדולות כמו 86 kilobase (kb) DNA המיטוכונדריאלי (mtDNA). השיטה מעגל-Seq פותחה מתוך שיטת טיהור פלסמיד פרוקריוטים ומבוססת 18,19, מותאם במיוחד בתאי שמרים האיקריוטים בשילוב עם רצף עמוק. שימוש בגישה מעגל-Seq, eccDNAs שונים 1756, כל הגדולים מ -1 kb, אותרו מעשר ס cerevisiae S288c אוכלוסיות 20. גודל חתוכים נבחר להתמקד eccDNA שהיו גדולים מספיק כדי לשאת גנים כולו. מעגל-Seq היה מאוד רגיש; זה זיהה יחיד eccDNA בתוך אלפי תאים 20. במחקר הנוכחי, המעגל-Seq שמש לבודד ולזהות 294 eccDNAs משלושה ביולוגים משכפלים של אחר S. זן שמרים cerevisiae, CEN.PK. הנתונים מגלים כי eccDNA הוא eleme גנטי משותףNT ב ס זני cerevisiae.
Protocol
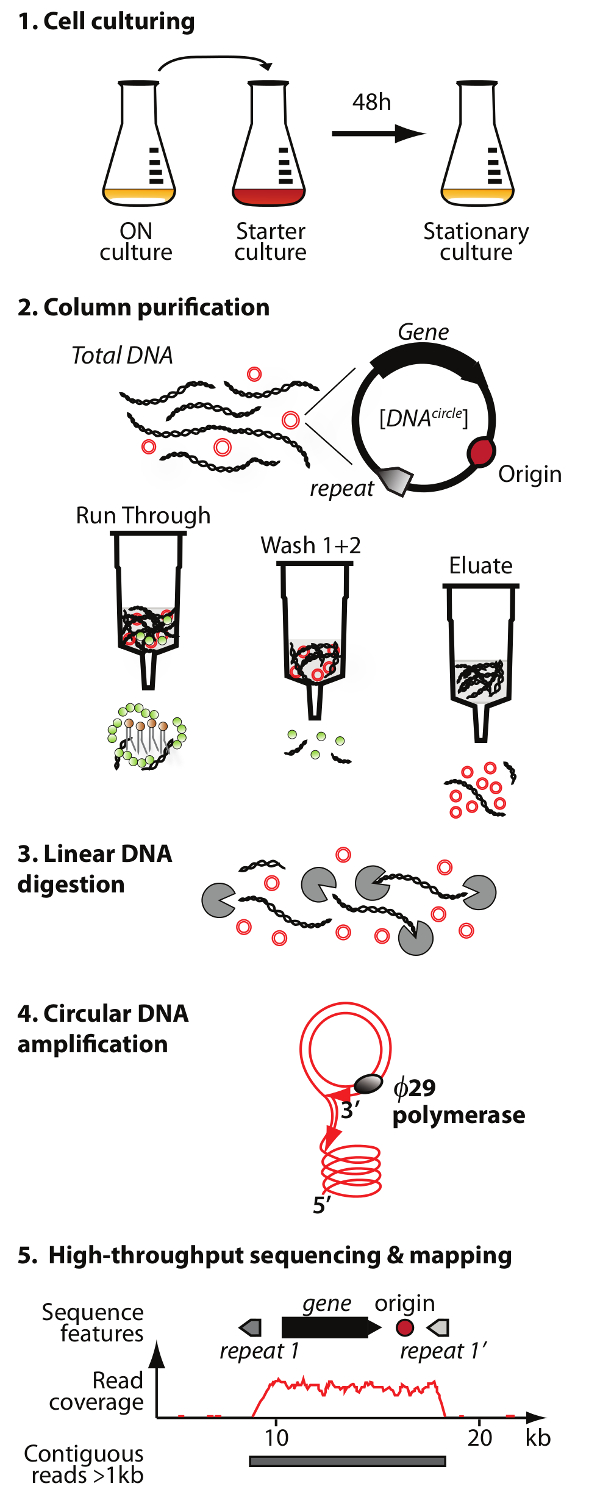
הערה: סקירה של טיהור DNA מעגלי שיטת סידור (מעגל-Seq) מתוארת באיור 1.
1. טיפוח, תא קציר פלזמה שיבוש ממברנה
- לחסן תאי שמרים (למשל שמר האפייה) מתרבות O / N לתוך 50 מ"ל מזין בינוני מלאה של דקסטרוז שמרים peptone (YPD). לחסן בצפיפות תא ראשונית נמוכה של 1-3 x 10 5 תאים / מ"ל או צפיפות אופטית של כ 0.01 OD 600.
- דגירת התאים ב 30 מעלות צלזיוס עם תסיסה ב 150 סיבובים לכל (סל"ד) דקות עד תאים להגיע צפיפות תא מרבית של כ 1 x 10 10 תאים, כ לאחר 24 עד 48 שעות או צפיפות אופטית ב OD 600> 10.0.
שים לב: בפעם הטיפוח היא לא קריטית, מאחר שניתן להשתמש בם ריכוזי תא תחתונים.
- דגירת התאים ב 30 מעלות צלזיוס עם תסיסה ב 150 סיבובים לכל (סל"ד) דקות עד תאים להגיע צפיפות תא מרבית של כ 1 x 10 10 תאים, כ לאחר 24 עד 48 שעות או צפיפות אופטית ב OD 600> 10.0.
- מעבירים את התרבות הקטן מדי כדי צינור חרוטי 50 מ"ל, גלולה התאים על ידי צנטריפוגה ב800 XG במשך 3 דקות וזורקים supernatant.
- שטוף את הכדור עם פתרון חיץ 25 מיליליטר של 10 המ"מימ טריס-Cl, 1 mM EDTA, pH 8.0, מחדש גלולת התאים על ידי צנטריפוגה ב 800 XG במשך 3 דקות וזורקי supernatant.
- Resuspend התא גלולה במאגר 1.2 מ"ל resuspension שסופקו ערכת פלסמיד עמודה הטיהור.
- שלב אופציונאלי: להוסיף פלסמידים מדוללים מאוד כפקדים לטיהור של רצפי דנ"א מעגליים 20.
הערה: בסיס הנתונים הנוכחיים, תערובת פלסמיד 7.7 μl יושמה עבור כל מדגם המכיל 10 10 תאים. תערובת מניית פלסמיד מורכבת משלושה פלסמידים בריכוזים שונים; pBR322 ב 38 ng / המדגם, pUC19 ברמה של 0.5 ng / המדגם, pUG72 0.01 ng / המדגם. - מעבירים את ההשעיה התא לתוך צינורות מיקרו צנטריפוגות שני 2 מ"ל, כל השלימו עם חרוזי זכוכית 0.5 מ"מ יחס של 1: 3 מנפח השעיה מוחלטת.
- וורטקס כל צינור במהירות מקסימלית במשך 10 דקות כדי לשבש תא פלזמהממברנות. גלולה חרוזים על ידי צנטריפוגה ב 268 XG במשך 30 שניות ולהעביר את supernatant 1.2 מ"ל בשילוב משני צינורות microcentrifuge צינור חדש.
הערה: חלופה לשלב 1.6-1.7, השתמש zymolyase לשבש תאים בתמיסה חיץ 0.6 מ"ל resuspension. עשר יחידות של zymolyase יכול לשבש 5 x 10 7 תאים בתוך 1.5 שעות ב 35 מעלות צלזיוס.
2. עשרת EccDNA ידי כרומטוגרפיה טור
- פעל על פי הפרוטוקול מתוך ערכה לטיהור פלסמידים עמודה. בקיצור, לטפל כל דגימה עם פתרון 1.2 מ"ל אלקליין, מערבבים בעדינות דגירה 3 דקות ב RT.
- להוסיף 1.2 מ"ל חיץ הניטרול, מערבבים בעדינות צנטריפוגות ב 9650 XG במשך 5 דקות.
- טען את הפתרון על עמודה equilibrated עם פתרון איזון 1 מ"ל ולאפשר לנוזל לזרום דרך העמודה על ידי כוח הכבידה.
- לשטוף את העמודה עם פתרון כביסה 4 מ"ל. כאשר הפתרון עבר דרך השרף, להוסיף בזהירות 0.3 מיליליטר elution כךlution להחליף את רוב נפח חלל 0.35 מ"ל העמודה.
- Elute DNA לתוך צינור אוסף חדש עם פתרון elution 1 מיליליטר ו להאיץ את ה- DNA על ידי הוספת תערובת ממטרים 0.8 מיליליטר. צנטריפוגה ב 9650 XG במשך 10 דקות.
- שטפו את הכדור DNA עם אתנול 0.5 מ"ל 70%, צנטריפוגה XG ב 9650 למשך 5 דקות, אוויר יבש במשך 5 עד 15 דקות ו לפזר את ה- DNA מטוהרים 25 μl מים סטריליים.
הערה: אחסון לטווח קצר בלבד של DNA במים מומלץ. באופן מועדף, להמשיך ישירות לשלב 3.
3. עיכול של הנשארים DNA כרומוזומלית לינארי
- שלב אופציונלי: כדי להקל על העיכול ספציפי של DNA ליניארי על ידי exonuclease, לטפל DNA מטוהרים עם endonuclease נדיר חיתוך כגון NotI. במשך 5 DNA מיקרוגרם, השתמש יחידה 1 NotI, 5 μl 10x חיץ העיכול מים סטריליים כדי בהיקף כולל של 50 μl. דגירת התגובה על 37 מעלות צלזיוס למשך 16 שעות וחום להשבית את endonuclease על 80 מעלות צלזיוס למשך 5 דקות.
- הוסף 20יחידות exonuclease (2 μl), 4 ATP μl (25 מ"מ), 34 μl מים סטריליים 10 μl 10x חיץ התגובה ישירות ל- DNA 50 μl-ביקע endonuclease להגיע נפח התגובה 1x של 100 μl, באמצעות exonuclease תלוי ATP קִיט.
- בצע הידרוליזה של DNA ליניארי חד גדילי ו פעמיים תקועים על 37 מעלות צלזיוס למשך 5 ימים או יותר. הוסף ATP נוסף 4 μl (25 מ"מ), 0.6 μl חיץ התגובה 10x ו -20 יחידות exonuclease כל 24 שעות על מנת להמשיך את מערכת העיכול DNA האנזימטית בווליום התגובה 1x.
- לאחר הסרת DNA ליניארי, μl מדגם 2 מפתרון מטופלים exonuclease לאשר חיסול DNA ליניארי כרומוזומליות ידי תגובת שרשרת פולימרז כמותי (qPCR), באמצעות סמן כרומוזומליות כגון ACT1 הגן אקטין 20.
- כל כרך תגובה 20 μl qPCR מכיל 2 μl מדגם שטופלו exonuclease, 150 פריימרים ACT1 ננומטר 5'-TCCGTCTGGATTGGTGGTTCTA-3 '5'-TGGACCACTTTCGTCGTATTC-3 ", 2% (נפח / נפח) sulfoxide דימתיל, ו -10 μl ירוק תערובת אמן ניאון.
- השתמש במצב התגובה; 3 דקות ב 95 מעלות צלזיוס, ואחריו 45 מחזורים של 15 שניות על 95 מעלות צלזיוס ו -30 שניות על 60 מעלות צלזיוס.
הערה: ACT1 הוא סמן מתאים במיוחד DNA ליניארי מאז וריאציות מספר עותק בגן הזה הן מזיקות 21-23 כך eccDNA לא צריך לשאת ACT1. - תחליפים ניתוח של מערכת העיכול DNA על ידי qPCR הם PCR סטנדרטי (4.3) או מכתים יודיד propidium (4.4).
- השתמש 2 μl מדגם שטופלו exonuclease כתבנית PCR עם פריימרים ACT1 5'-TGGATTCTGGTATGTTCTAGC-3 '5'-GAACGACGTGAGTAACACC-3 ". כפי בקרה ACT1 חיובית, להשתמש 50-100 ng ס גנומית DNA cerevisiae כתבנית. תנאי תגובת PCR; 3 דקות ב 95 מעלות צלזיוס, ואחריו 35 מחזורים של 30 שניות על 95 מעלות צלזיוס, 30 שניות ב 56 מעלות צלזיוס ו 1 דקות ב 72 מעלות צלזיוס.
- הפעלת PCR תגובות על ידי ג'ל electrophoresis על agarose 1% עם 0.5 מיקרוגרם / מ"ל ethidium ברומיד. חפש להקה 0.8 kb ACT1.
- העדר או נוכחות של DNA ליניארי ניתן לבדוק גם על ידי מכתים יודיד propidium לפני ואחרי הגברה DNA.
- מערבבים כל דגימת DNA ביחס של 1: 1 נפח עם 1: המניה יודיד 1,000 H 2 O-מדולל פתרון של 20 מ"מ propidium. השאר פתרון בחושך במשך 10-20 דקות ב RT ולנתח מכתים DNA על ידי מיקרוסקופ פלואורסצנטי בהגדלה 100x באמצעות מסנן קרינת עירור אדום 663-738 ננומטר וחשיפה פעמית של 5 עד 30 שניות. כפי בקרה DNA מכתים, שימוש ø29 מוגבר הדנ"א הגנומי של שמרים ו / או פלסמיד מוגבר ø29.
- מחממים להשבית הפתרון exonuclease ב 70 מעלות צלזיוס למשך 30 דקות.
4. DNA הגברה
- הגבר את מטוהרים מועשר eccDNA משלב 3.5) עם DNA פולימרז ø29 24-26 בהסכמהing לפרוטוקול של היצרן פולימראז.
- בקיצור, לערבב 5 μl מועשר eccDNA עם חיץ denaturation 5 μl.
- לאחר 3 דקות ב RT, להוסיף למאגר נטרול 10 μl. מערבבים בעדינות ומוסיפים 30 μl תערובת מאסטר המכיל 29 μl חיץ התגובה 1 DNA פולימרז ø29 μl. דגירת התגובה על 30 מעלות צלזיוס למשך 16 שעות או יותר (עד 72 שעות). מחממים להשבית DNA פולימרז ø29 על 65 מעלות צלזיוס במשך 3 דקות.
רצף 5. וניתוח נתונים
- גזירת eccDNA המוגבר עם ultrasonicator ממוקד לגודל שיא היעד ממוצע של 300 נ"ב. השתמש בהגדרות הבאות עבור דגימת DNA 130 μl: כוח עוצמת שיא 450W, 60 טיפול שני, 30% מקדמים העומס, 200 מחזורי פרץ, טמפרטורה 7 מעלות צלזיוס.
- להוסיף תוויות מדד ברקוד ומתאמים אל המקוטעת קוראה לסינתזה של ספריות עבור סידור, באמצעות שיטה מתאימה כנה ספרייה.
- הפעל רצף עמוק, למשל כמו-הסוף יחיד 141 נוקליאוטידים קורא על פלטפורמת רצף תפוקה גבוהה.
- מפת מקריא בגנום שמרים הפניה תחת חקירה ולאפשר קורא למפה למספר אזורים. לדוגמא, להשתמש במערכת עבודה זמינה באופן חופשי 27,28 ו תוכנת מיפוי aligner קצרה לקריאת 29.
- זהה קורא מאזורים של eccDNAs המשוערת באמצעות רציף קורא, למשל, יותר משבע רציף קורא (> 1 kb) ללא פערי 20.
הערה: התוכנה זמינה 27,28 לחקר ממופה קורא לעבר אזורים הגנומי של עניין.
תוצאות
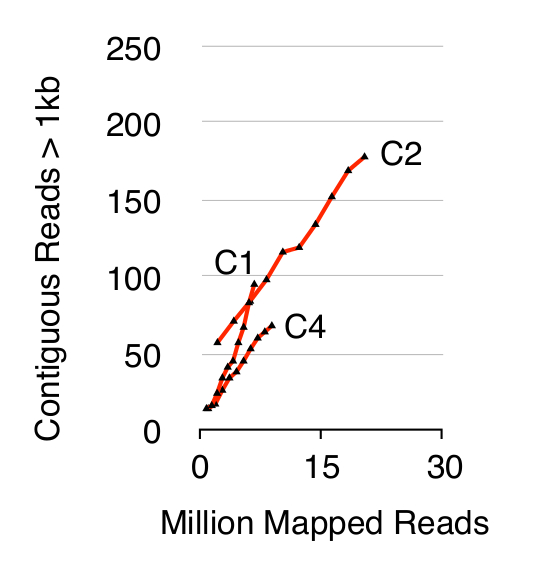
כדי לאמת את שיטת המעגל-Seq, שלוש S. cerevisiae CEN.PK אוכלוסיות של 1 x 10 10 תאים הוקרנו לאחר התאים גדלו בנפרד YPD במשך עשרה דורות. חיסול כרומוזומלית DNA ליניארי אושר על ידי בהעדר אות qPCR ACT1 כפי שתואר לעיל 20 (מידע לא מוצג). eccDNA מטוהרים מועשר היה רצף עד 68 מיליון קורא (141 נוקליאוטידים לקצה יחיד כניסות), ומיפו את הגנום התייחסות CEN.PK113-7D (גרסה 19 יוני 2012). הקלטות של eccDNAs המשוערת מהמדגמים השלושה בשם C1, C2 ו- C4 חולקו לאזורים גנומית ממופים על ידי רציף קורא יותר מ -1 kb. בהתבסס על 10,000 סימולציות מונטה קרלו, המשמעות של כל אזור ממופה על ידי רציף קורא יותר מ -1 kb נאמד. מתוך 79 זה, 159 ו -56 אזורים היו מפורשים כמו רצפי eccDNA סבירים (p <0.1, מערך נתונים 1). מספר contiguo המוקלטתלנו קורא> 1 kb גדל כפונקציה של עומק רצף המצביעה על כך אפילו יותר אלמנטי eccDNA היו נרשמים אם דגימות היו רצף נוסף (איור 2). כצפוי, השיטה מעגל-Seq חילוץ רבים קורא ממספר רצפי דנ"א מעגלית ידוע כולל הפלסמיד 2μ, הדנ"א המיטוכונדרי, גנים RNA ריבוזומלי בכרומוזום XII, ושלושת פלסמידים הבקרה הפנימית pBR322, pUC19 ו pUG72 שהיו זינק לתוך דגימות רק לפני טיהור עמודה (איור 3).
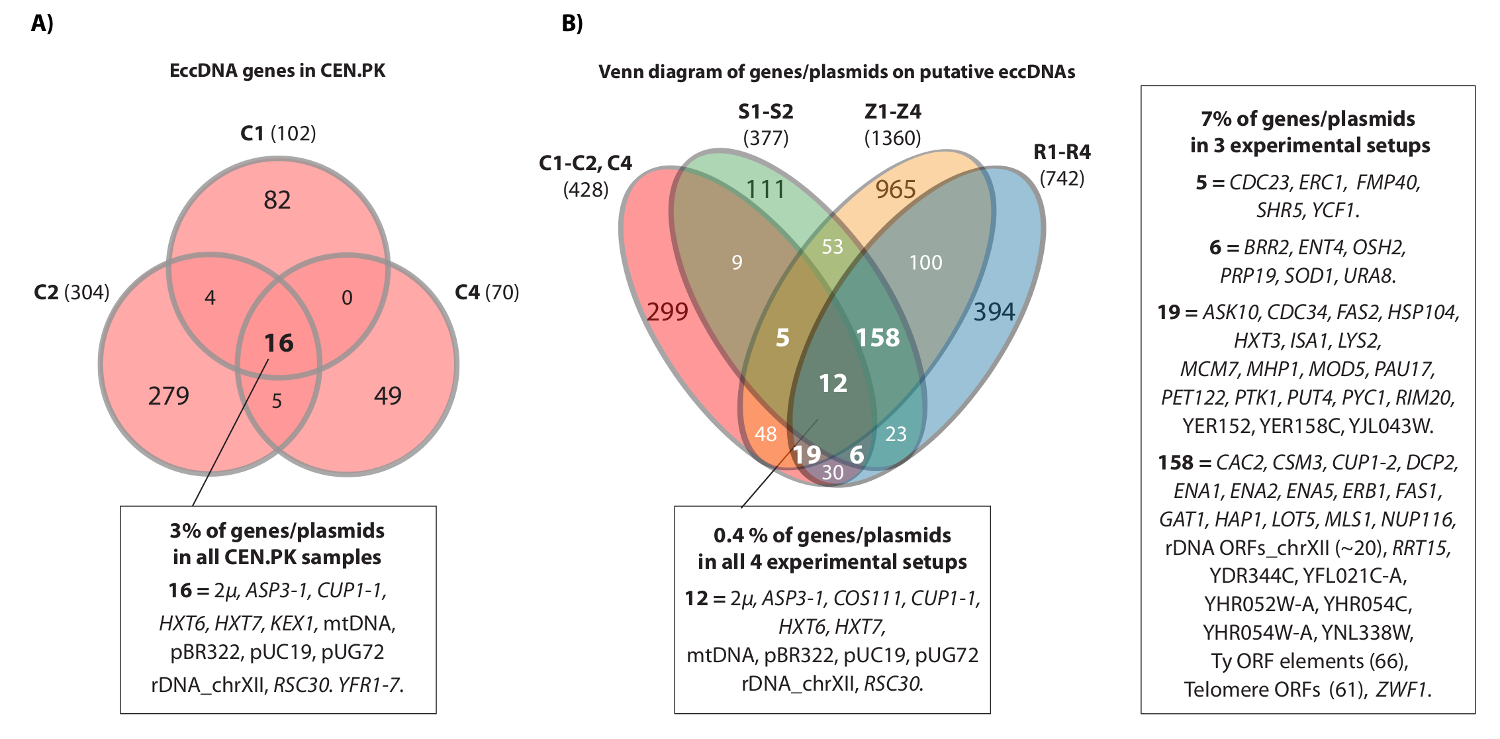
הוידאו מציג דוגמה רציף קורא כי ממופה אל מוקד HXT7 _ARS432_ HXT6 על IV כרומוזום. בעבר, [מעגל HXT6 / 7] זוהה על ידי מעגל-Seq בעשר אוכלוסיות S288c (כל אחד עם 1 x 10 10 תאים) ואת מבנה ה- DNA מעגלי אושרה על ידי ניתוח הפוך PCR 20. ה [HXT6 / 7 CIrcle] גם נרשמה כל אחת (איור 4 א) שלוש אוכלוסיות CEN.PK. יתר על כן, רוב גני eccDNA הנפוצים בקרב דגימות לשכפל של CEN.PK חפפו גני eccDNA מן מערכי נתוני S288c (האיור 4B).
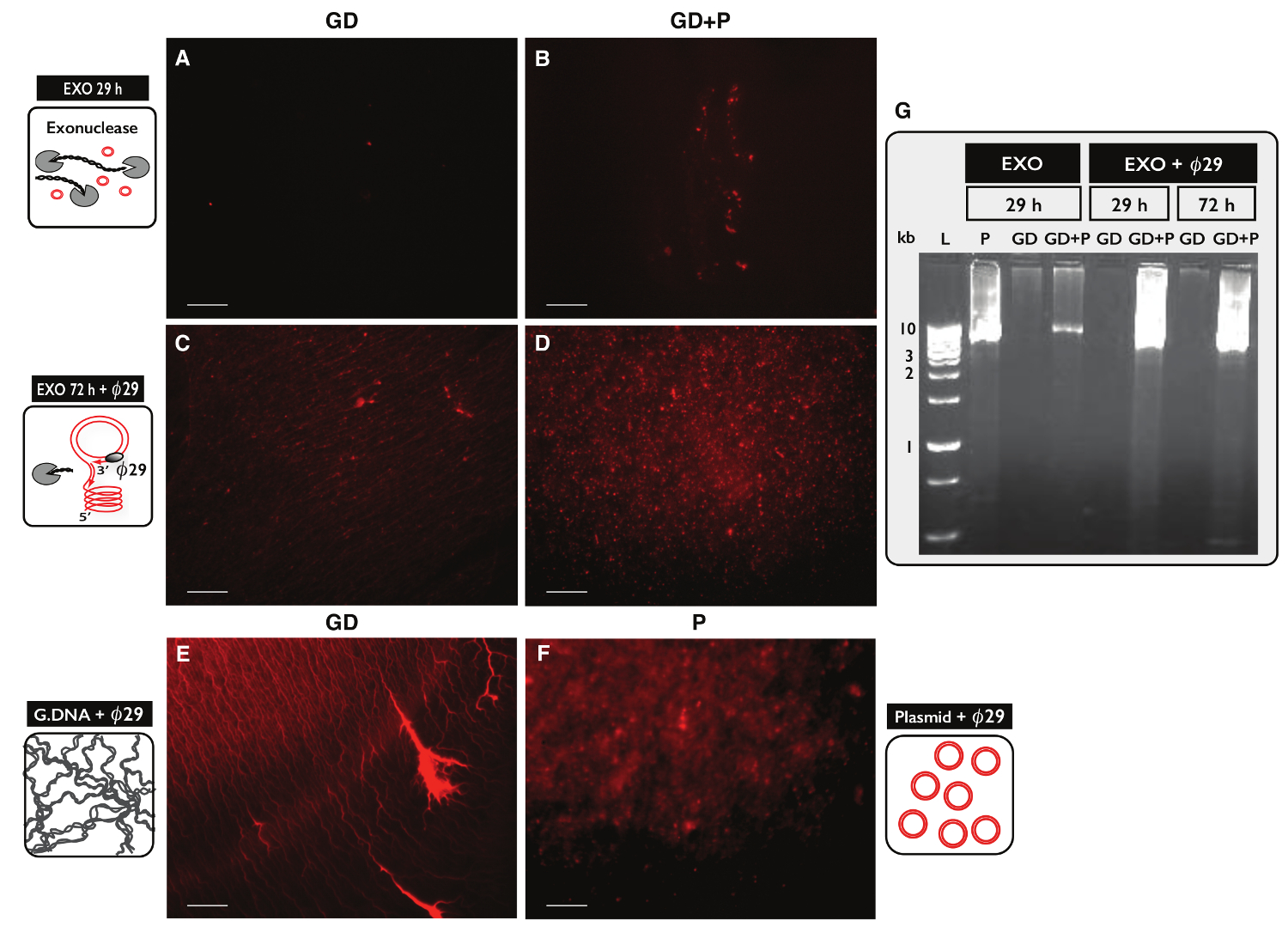
כדי לבדוק את הספציפיות של הפרוטוקול-Seq המעגל עבור טיהור DNA מעגלית, שתי דגימות, כל אחד עם 30 מיקרוגרם DNA גנומי, נבדקו. דגימה אחת נוספה עם 100 ננוגרם פלסמיד דנ"א eccDNA משני דגימות היו מטוהרים ידי פרוטוקול מעגל-Seq. לאחר ההפרדה עמודה, התשואה DNA היה 1.27% (380 ng) למדגם ללא פלסמיד (GD) ו 1.60% (480 ng) למדגם עם פלסמיד (GD + P). היעילות של טיפול exonuclease נבדק עבור תוכן DNA ליניארי לאחר 29 שעות ו -72 שעות באמצעות PCR נגד ACT1. אין דגימות הכילו מוגבר ACT1 (מידע לא מוצג). שבריר של כל דגימה שטופל exonuclease היה עוד יותר אתmplified ידי ø29 פולימראז ואת המוצרים של תגובות אנזימטיות נותחו על ידי מכתים יודיד propidium (איור 5 א-F) ג'ל אלקטרופורזה agarose (איור 5G). דוגמאות לאחר טיפול exonuclease הראו יוד-כתם propidium מינימאלי (איור 5 א-ב). Ø29 - מדגם מוגבר עם הדנ"א הגנומי רק חשף מבנים דמויי חוט (איור 5 ג), בדומה למה מדגם הביקורת (איור 5E). Ø29 - מדגם מוגבר כי היה פלסמיד הוסיף חשף מוקדים (איור 5D) הדומה השליטה פלסמיד (איור 5F). התמונות עולה כי פולימראז ø29 מועשר עבור DNA מעגלי מעל DNA ליניארי. רוב ה- DNA כרומוזומלי ליניארי הוסר ממדגמים לאחר 29 טיפול hr exonuclease (איור 5 א-ב, ז). עם זאת, טיפול exonuclease נרחב עבור יותר מ -100 שעות ושימוש יותר מ -100 יחידות היו צריך להסיר את כל ה- DNA ליניארי כרומוזומליות, כמו ø29 - דגימות מוגברות עדיין הראו רקע של מבנים דמויי חוט לאחר טיפול 72 שעות exonuclease (איור 5 ג-ד).

. באיור 1. תמציתי של שיטת מעגל-Seq הפרוטוקול יש 5 שלבים: 1) culturing תא, 2) טיהור והעשרה של eccDNA ידי כרומטוגרפיה טור, 3) העיכול של הנותרים דנ"א כרומוזומלי ליניארי בשבריר eluate, 4) הגברה של ה- DNA על ידי DNA פולימרז ø29, ו -5) רצף של eccDNA מועשר ומיפוי קורא אל ס הגנום התייחסות cerevisiae. אנא לחץ כאן כדי לצפות בגרסה גדולה יותר של דמות זו.
{kind=link}
p_upload / 54,239 / 54239fig2.jpg "/>
איור 2. רציף קורא> 1 kb כפונקציה של עומק רצף. EccDNA מ 1 x 10 10 תאים להגדיל כפונקציה של עומק רצף (במיליוני ממופה כניסות). מוצג: triplicates ביולוגי בין הפלואידים CEN.PK ס אוכלוסיות cerevisiae (C1, C2, C4) מופרדים על ידי 10 10 חלוקות תא. אנא לחץ כאן כדי לצפות בגרסה גדולה יותר של דמות זו.
{kind=link}

איור 3. איתור של רצפי דנ"א מעגלית ידוע. (AB) מפזרים חלקות כיסוי קריאה (לקרוא צפיפות) באחוזים עבור פלסמידים CEN.PK ביולוגי משכפל C1, C2 ו- C4. (א) ממופים מקריא פלסמידים שמרים אנדוגני היו: 2μ; [מעגל rDNA] (גנים RNA ריבוזומלי מ XII כרומוזום); ו mtDNA (בדנ"א המיטוכונדריאלי). (ב) הייחודי קורא ממופה לשלוט פלסמידים. פלסמידים הבקרה היו זינק לתוך דגימות לפני טיהור עמודה. יחסי פלסמיד לכל תא היו: pBR322 (סימני פלוס) 1: 1, pUC19 (עיגולים) 1:50, ו pUG72 (משולשים) 1: 2,500.

איור 4. אלמנטי eccDNA הנפוץ CEN.PK ו S288c. (א) דיאגרמת ון מוצגת חפיפה בין 476 גנים על 294 אלמנטים eccDNA בשלוש דגימות CEN.PK (C1, C2, C4). 16 הגנים המשותפים החופפים eccDNA / פלסמידים הם מבוארים (כל שמות הגן נמצאים בסיס נתוני 1). (ב) ון דיאגרמה של כל הגנים רשמו על eccDNAs משוערת משלוש דגימות CEN.PK (C1, C2, C4), לעומת כל הגנים רשמו על eccDNAs המשוערת מ -10 דגימות S288c: S1-S2, R1-R4, Z1-Z4 (ראה התייחסות 20). מוצגים הם 13 משכפל ביולוגי (S1-S2, R1-R4, Z1-Z4, C1-C3) עם גנים / פלסמידים ואזורי eccDNA משוערים אשר חפפו מינימום של 2 רקע מתח ואו 3 או יותר setups הניסיון. דגימות C, CEN.PK; דגימות R ו- Z, S288c BY4741; דגימות S, S288c M3750. אנא לחץ כאן כדי לצפות בגרסה גדולה יותר של דמות זו.
{kind=link}

איור 5. ויזואליזציה של דגימות DNA לאחר exonuclease וטיפול O 2 9. (AF) Propidium יודיד מכתים של DNA. סרגל קנה מידה, 10 מיקרומטר. (A, C ו- E) דוגמאות עם הדנ"א הגנומי (GD); (B ו- D) דגימות עם GD בתוספת פלסמיד (GD + P). ( אונג> א.ב.) לאחר טיפול exonuclease 29 שעות (EXO 29 ח); (CD) לאחר 72 שעות טיפול exonuclease ואחריו הגברה פולימראז ø29 (EXO 72 שעות + ø29). (ה) שליטת הדנ"א הגנומי לאחר דואר: הגברת פולימראז ø29; (F) שליטה פלסמיד (5.5 kb) לאחר 29 פולימראז הגברה; (O G) ג'ל-eletrophoresis agarose. משמאל: L, 1 סמנים kb; P, בקרת פלסמיד (5.5 kb) אחרי שעה 29 EXO; GD, לאחר EXO 29 שע '(מדגם כמו א); GD + P, לאחר EXO 29 שע '(מדגם כמו B); GD ו- GD + P, לאחר EXO 29 שעות + ø29; GD ו- GD + P, לאחר EXO 72 שעות + ø29 (מדגם כמו CD). ראה טבלה S1 לפרטים נוספים. אנא לחץ כאן כדי לצפות בגרסה גדולה יותר של דמות זו.
{kind=link}
1 Aset "src =" / files / ftp_upload / 54,239 / 54239dataset1.jpg "/>
מערך נתוני 1. אזורי circularization DNA פוטנציאלי CEN.PK. אנא לחץ כאן כדי להוריד את הקובץ.
מוצגים הם נתונים רצף ומנתח עבור 348 אזורים. עמודות הן לספירה, מיפוי eccDNA. A (טור ראשון משמאל), מדגם שממנו המשוערת eccDNA זוהה; B, כרומוזום; CD, התחלה וסיום הקואורדינטות של eccDNAs המשוערת. EH, תוכן eccDNA. E, משכפלים רצף באופן אוטונומי (ARS) באזור; F, גן מלא באזור; G, חלק הגן ייכלל באזור; H, BLASTN-מזוהה הגן. IO, כיסוי ו- p-values EccDNA. אני, באזור הארוך עם רצף מבואר ייחודי נ"ב; J, מספר כל ממופה קורא; K, כיסוי של כל ממופה קורא מרסיסים לכל kb מתוך מיליון ממופה קורא (FPKM); L, p-value עבור המשוערת eccDNA לעומת התרחשות במקרה ממונטה קרלו סימולציות; M, מספר בלתיiquely ממופה קורא; לא; כמו K ו- L באמצעות רק ממופה באופן ייחודי קורא (UFPKM). פרמטרים למיפוי של קורא מונטה קרלו היו כמתואר 20.
Discussion
השיטה-Seq המעגל מאפשרת זיהוי בקנה המידה הגנום של eccDNA מתאי שמרים עם רזולוצית רצף ברמה. השיטה היא היטהרות eccDNA קלה שאינה דורשת נמרץ מערבולת או pipetting ומשתמש ההפרדה עמודה על ידי כוח הכבידה להגביל שבירה eccDNA שיוביל העיכול exonuclease ב לשלב הבא. תכונות אלה של השיטה עשויות להיות מכריעות לאיתור eccDNAs גדולה המכילות רצפי גנים. המעגל-Seq זוהה eccDNAs רב, כוללים גנים מלאים (מערך נתונים 1). כמו כן זיהה את ה- DNA שמרים המיטוכונדריה 86-kb. לפיכך, פרוטוקול זה הופך לפשוט טיהור של רצפי דנ"א עגולים גדולים. שמירה על מספר צעדי מיצוי DNA למינימום מפחיתה את הסיכון eccDNA אובדן למקסימום תשואה. בהתבסס על תוצאות לבקרה, ממוסמר-ב פלסמידים, מעגל-Seq היא רגישה מאוד, מזהה DNA מעגלי יחיד מ -2,500 תאים. יתר על כן, הסרת פלסמידים אנדוגני בשפע כגון 2μ; פלסמיד או הדנ"א המיטוכונדרי עשוי לשפר את הרגישות באופן משמעותי. אשפרה של 2μ מתרבויות שמרים תוארה 30. לחלופין, 2μ והסרה הדנ"א המיטוכונדרי עלול להיות מושגת באמצעות endonuclease נדיר חיתוך, כגון SwaI. עם זאת, צעד אנזים ההגבלה יכול למקד eccDNAs האחר בעלי עניין ולהגביל את תשואת eccDNA הכולל.
צעדים קריטיים לגילוי eccDNA היו הסרת DNA ליניארי (שלב 3) ו רצפי DNA (שלב 5) עד לעומק נכון. כדי להקליט את רוב eccDNAs מאוכלוסיית תא, רצף עמוק ייתכן שיידרש 20. רצף מותאם-הסוף צריך לספק ביטחון עוד יותר של זיהוי eccDNA, כמו צמתי DNA מעגליות צפויות להניב-סוף לזווג קוראות המפה discordantly. פערים אלה תומכים גילוי מבני DNA המעגליים וניתן להשתמש כמכילי מסנן eccDNA-זיהוי נוסף.
החוג-Seשיטת q הייתה תוקף באמצעות שלושה ס עצמאי אוכלוסיות CEN.PK cerevisiae. רצפים זוהו כלולים שדווחו בעבר eccDNAs, פלסמידים אנדוגני ומשובצים-ב פלסמידים והמאה eccDNAs המשוערת (מערך נתונים 1). ממצאים אלה תומכים מערכי נתונים מעגלים-Seq קודמים מתוך ס cerevisiae S288c 20. התגלית של כמה eccDNAs המשותף לאוכלוסיות CEN.PK ו S288c עולה כי לוקוסים אלה יש נטייה להתקיים אלמנטים עגולים כמו (איור 4). הראינו בעבר כי [מעגל GAP1] מועשר בתנאי מגבילי חנקן ברקע CEN.PK 8, אם כי עדות [מעגל GAP1] ברקע זן אחר לא נמצאה. מציאת של eccDNA מן CUP1-1 RSC30, ASP3-1, COS111, ו HXT6 HXT7 לוקוסים בשני S288c ו CEN.PK עולה כי ולנטייה circularization DNA הוא conשמש בין זני שמרים. נותר רק לראות אם [HXT6 / 7 מעגל], [מעגל ASP3-1], [מעגל COS111], ו [מעגל CUP1-1 RSC30] להעניק יתרונות סלקטיבית לתאים או אם קיומם הוא רק השפעה של שיעור גבוה של DNA circularization.
יחדיו, התוצאות מצביעות על כך המעגל-Seq הוא גם מתאים לאיתור eccDNAs בגודל kilobase ויש לו יתרונות לזיהוי eccDNAs עם גנים שלמים. המעגל-Seq היא שיטה רגישה מאוד המאפשרת מסכים בקנה מידת הגנום כולו של eccDNAs משמר. השיטה מעגל-Seq יכול לפתוח תחום חדש של מחקר שמטרתה הבהרת תפקיד eccDNA ביצירת מחיקות גנים amplifications. בהתחשב בכך ארכיטקטורה ומבנה הדנ"א שמורים בעיקר משמרים איקריוטיים כדי אאוקריוטים גבוהים, שיטת המעגל-Seq צריכה, באופן עקרוני, להיות applicablדואר לכל תאים איקריוטיים, עם שינויים קלים. כיום, השיטה לא נראה לי שום מגבלות, למרות יכולתו לטהר eccDNAs megabase בגודל טרם לראות. בנוסף, השימוש ø29 DNA פולימרז, אשר משתמש בשיטת הגברה מתגלגל מעגל 31, יוצר הטיה כלפי eccDNAs הקטן עושה כימות eccDNA קשה יותר. המעגל-Seq מזהה eccDNAs הגדול מספיק כדי לשאת גנים מלאים, מה שהופך אותו מתאים מחקרים על הדנ"א כפול דקות עגולות מן תאי הסומטיים אדם. דקות זוגיות יכולות לגרום לסרטן כאשר-אונקוגנים פרוטו הם מוגברים על האלמנטים האלה 32-37. מחקרים של eccDNAs בתאים germline יכול לשמש כדי למדוד את שיעורי מוטציה בתאי מין וגם להעריך את איכות הזרע, למשל בעלי חיים. לפיכך, מעגל-Seq יש פוטנציאל להניב תובנות לתוך הקצב שבו וריאציה גנטית מתעוררת בצורת וריאציה מספר עותק, ולהוביל להבנה הרומן של מחלות שקשורות הגן copy-מספר וריאציה 38-40.
Disclosures
The authors declare that they have no competing financial interests.
Acknowledgements
Thanks to Kenn D. Møller and Claus Sternberg (DTU) for technical assistance and to Tue S. Jørgensen for quantitative PCR analysis.
Materials
| Name | Company | Catalog Number | Comments |
| Bacto peptone | BD Difco | 211677 | Alternative product can be used. |
| Brilliant III SYBR Green PCR Master Mix | Agilent Technologies | 600882 | For qPCR analysis. Alternative product can be used. |
| Dextrose (D-glucose) | Carl Roth | HN06.4 | Alternative product can be used. |
| Disruptor Beads, 0.5 mm | Scientific Industries, Inc. | SI-BG05 | Glass beads to disrupt plasma cell membranes. Alternative product can be used. |
| Ethidium bromide | Carl Roth | 2218.2 | Agarose gel stain for detecting DNA/RNA. |
| GeneJet plasmid miniprep kit | Thermo Fisher | K0502 | Plasmid purifcation from bacteria. Alternative product can be used |
| NotI, FastDigest | Life Technologies - Thermo Fisher Scientific, USA | FD0594 | Endonuclease. Alternative product can be used. |
| Plasmid Mini AX kit | A&A Biotechnology, Poland | 010-50 | Plasmid purifcation kit used to purify eccDNA. |
| Plasmid-Safe ATP-dependent DNase kit | Epicentre, USA | E3105K | ATP-dependent exonuclease kit. Alternative product can be used. |
| Propidium iodide | Sigma-Aldrich, USA | 81845 | Alternative product can be used. |
| pUG6 plasmid | EUROSCARF, Germany | P30114 | Marker gene: loxP-PAgTEF1-kanMX-TAgTEF1-loxP. Plasmid requests: Please contact Dr. Peter Philippsen@unibas.ch |
| QIAGEN genomic-tip 100/G | Qiagen, USA | 13343 | Genomic DNA purifcation from yeast. Alternative product can be used. |
| REPLI-g Mini Kit protocol | Qiagen, USA | 150023 | Amplification of eccDNA by the phi29 polymerase |
| Yeast extract | BD Difco | 210929 | Alternative product can be used. |
| Zymolyase 100T (Lyticase, Yeast Lytic Enzyme) | Nordic BioSite, Sweden | Z1004-3 | Alternative product can be used. |
| Data access to sequence files | European Nucleotide Archive | EccDNA dataset from Saccharomyces cerevisiae CEN.PK113-7D. Study accession number PRJEB9684. 2nd accession number is ERP010820. Locus tag prefix is BN2032. | |
| Strains | |||
| Saccharomyces cerevisiae CEN.PK113-7D | Genotype MATa MAL2-8c SUC2 | ||
| Saccharomyces cerevisiae yeast deletion library pool | EUROSCARF, Germany | S288c BY4741 pool of 4400 viable single-gene deletion mutants disrubted by KanMX module. Genotypes MATa his3∆1 leu2∆0 met15∆0 ura3∆0 genexxx::KanMX. | |
| Equipments | |||
| DNA Spectrophotometer | NanoDrop 1000 Spectrophotometer, Thermo Fisher | Measuring DNA concentration. Alternative product can be used. | |
| Fluorescence microscopy | Nikon Optronics Magnafire. Red excitation fluorescence filter, 663-738 nm. | Alternative product can be used. | |
| Robotic library-build system | Apollo 324, IntegenX Inc. | DNA library preparation. Alternative product can be used. | |
| Sequencing platform | Illumina HiSeq 2000 platform, Illumina Inc. | DNA sequencing. Alternative product can be used. | |
| Ultrasonicator | Covaris LE220, microTUBE AFA Fiber tubes | Alternative product can be used. | |
| Methods | |||
| 2% YPD media | Mix 10 g Dextrose, 10 g Yeast extract, 20 g Bacto peptone and add H2O to a total volume of 1000 ml and autoclave. | ||
| Circle-Seq test on genomic DNA | Genomic DNA was purified (Qiagen) from a pool of the yeast deletion library (Euroscarf). The DNA concentration was measured by nanodrop and 30 µg genomic DNA was pipetted into two micro centrifuge tubes. One micro centrifuge tube was supplemented with 100 nanogram plasmid (pUG6). The DNA samples were purified by Circle-Seq, omitting the protocol steps 1.1-1.3 and 1.5-1.7. The eluted DNA concentrations were measured by nanodrop and the entire DNA yield from sample GD and GD+P was treated with exonuclease for a period of 29 hours. A 10% fraction was collected for phi29-amplification and PCR analysis, while the remaining DNA was subjected to 72 hour exonuclease treatment. The samples were analyzed for linear DNA content by PCR, using the ACT1 gene as chromosomal marker. A 5% fraction of each of the exonuclease treated samples was amplified by the phi29 DNA polymerase for 16 hours (Qiagen). The presence of DNA in each sample was examined by loading an equal amount (7 µl) in wells on an 0.5 µg/ml ethidium-bromide 0.9% agarose gel after running gel-electrophoresis. | ||
| Mapping software | Bowtie2 aligner, John Hopkins University | Ultrafast short read alignment. Reference: Langmead B, Salzberg S. Fast gapped-read alignment with Bowtie 2. Nature Methods. 2012, 9:357-359. | |
| Propidium iodide stain | Images of propidium iodine stained DNA were captured by fluorescence microscopy at 100x magnification (100x/1.30 oil, Nikon) in the RFP channel (red excitation fluorescence filter, 663-738 nm) using identical exposition time (5 seconds). | ||
| Workflow bioinformatic system | Galaxy, Open source. | A free web-based platform for data intensive biomedical research. References: Goecks, J, Nekrutenko, A, Taylor, J and The Galaxy Team. Galaxy: a comprehensive approach for supporting accessible, reproducible, and transparent computational research in the life sciences. Genome Biol. 2010 Aug 25;11(8):R86. Blankenberg D, Von Kuster G, Coraor N, Ananda G, Lazarus R, Mangan M, Nekrutenko A, Taylor J. "Galaxy: a web-based genome analysis tool for experimentalists". Current Protocols in Molecular Biology. 2010 Jan; Chapter 19:Unit 19.10.1-21. Giardine B, Riemer C, Hardison RC, Burhans R, Elnitski L, Shah P, Zhang Y, Blankenberg D, Albert I, Taylor J, Miller W, Kent WJ, Nekrutenko A. "Galaxy: a platform for interactive large-scale genome analysis." Genome Research. 2005 Oct; 15(10):1451-5. | |
References
- Kugelberg, E., Kofoid, E., et al. The Tandem Inversion Duplication in Salmonella enterica.: Selection Drives Unstable Precursors to Final Mutation Types. Genetics. 185 (1), 65-80 (2010).
- Reams, A. B., Kofoid, E., Savageau, M., Roth, J. R. Duplication Frequency in a Population of Salmonella enterica. Rapidly Approaches Steady State With or Without Recombination. Genetics. 184 (4), 1077-1094 (2010).
- Smith, C. A., Vinograd, J. Small polydisperse circular DNA of HeLa cells. Journal of Molecular Biology. 69 (2), 163-178 (1972).
- Carroll, S. M., DeRose, M. L., et al. Double Minute Chromosomes Can Be Produced from Precursors Derived from a Chromosomal Deletion. Molecular and cellular biology. 8 (4), 1525-1533 (1988).
- Cohen, S., Yacobi, K., Segal, D. Extrachromosomal Circular DNA of Tandemly Repeated Genomic Sequences in Drosophila. Genome research. 13 (6A), 1133-1145 (2003).
- Horowitz, H., Haber, J. E. Identification of Autonomously Replicating Circular Subtelomeric Y' Elements in Saccharomyces cerevisiae. Molecular and cellular biology. 5 (9), 2369-2380 (1985).
- Moore, I. K., Martin, M. P., Dorsey, M. J., Paquin, C. E. Formation of Circular Amplifications in Saccharomyces cerevisiae by a Breakage-Fusion-Bridge Mechanism. Environmental and molecular mutagenesis. 36 (2), 113-120 (2000).
- Gresham, D., Usaite, R., Germann, S. M., Lisby, M., Botstein, D., Regenberg, B. Adaptation to diverse nitrogen-limited environments by deletion or extrachromosomal element formation of the GAP1 locus. Proceedings of the National Academy of Sciences of the United States of America. 107 (43), 18551-18556 (2010).
- Windle, B., Draper, B. W., Yin, Y. X., O'Gorman, S., Wahl, G. M. A central role for chromosome breakage in gene amplification, deletion formation, and amplicon integration. Genes & development. 5 (2), 160-174 (1991).
- Gresham, D., Ruderfer, D. M., et al. Genome-Wide Detection of Polymorphisms at Nucleotide Resolution with a Single DNA Microarray. Science. 311 (5769), 1932-1936 (2006).
- Kidd, J. M., Cooper, G. M., et al. Mapping and sequencing of structural variation from eight human genomes. Nature. 453 (7191), 56-64 (2008).
- Gresham, D., Desai, M. M., Botstein, D., Dunham, M. J. The Repertoire and Dynamics of Evolutionary Adaptations to Controlled Nutrient-Limited Environments in Yeast. PLoS Genetics. 4 (12), 1-19 (2008).
- Lang, G. I., Botstein, D., Desai, M. M. Genetic Variation and the Fate of Beneficial Mutations in Asexual Populations. Genetics. 188 (3), 647-661 (2011).
- van Loon, N., Miller, D., Murnane, J. P. Formation of extrachromosomal circular DNA in HeLa cells by nonhomologous recombination. Nucleic Acids Research. 22 (13), 2447-2452 (1994).
- Vinograd, J., Lebowitz, J. Physical and Topological Properties of Circular Dna. Journal of General Physiology. 49 (6P2), 103 (1966).
- Shibata, Y., Kumar, P., et al. Extrachromosomal MicroDNAs and Chromosomal Microdeletions in Normal Tissues. Science. 336 (6077), 82-86 (2012).
- Dillon, L. W., Kumar, P., et al. Production of Extrachromosomal MicroDNAs Is Linked to Mismatch Repair Pathways and Transcriptional Activity. Cell Reports. 11 (11), 1749-1759 (2015).
- Li, L. L., Norman, A., Hansen, L. H., Sørensen, S. J. Metamobilomics - our knowledge on the pool of plasmid encoded traits in natural environments using high-throughput sequencing. Clinical microbiology and infection : the official publication of the European Society of Clinical Microbiology and Infectious Diseases. 18, 5-7 (2012).
- Brown Kav, A., Sasson, G., Jami, E., Doron-Faigenboim, A., Benhar, I., Mizrahi, I. Insights into the bovine rumen plasmidome. Proceedings of the National Academy of Sciences of the United States of America. 109 (14), 5452-5457 (2012).
- Møller, H. D., Parsons, L., Jørgensen, T. S., Botstein, D., Regenberg, B. Extrachromosomal circular DNA is common in yeast. Proceedings of the National Academy of Sciences of the United States of America. , 201508825 (2015).
- Drubin, D. G., Miller, K. G., Botstein, D. Yeast Actin-Binding Proteins - Evidence for a Role in Morphogenesis. The Journal of cell biology. 107 (6), 2551-2561 (1988).
- Magdolen, V., Drubin, D. G., Mages, G., Bandlow, W. High levels of profilin suppress the lethality caused by overproduction of actin in yeast cells. FEBS letters. 316 (1), 41-47 (1993).
- Sandrock, T. M., Brower, S. M., Toenjes, K. A., Adams, A. Suppressor analysis of fimbrin (Sac6p) overexpression in yeast. Genetics. 151 (4), 1287-1297 (1999).
- Blanco, L., Bernad, A., Lázaro, J. M., Martìn, G., Garmendia, C., Salas, M. Highly Efficient DNA Synthesis by the Phage ø29 DNA Polymerase. The Journal of biological chemistry. 264 (15), 8935-8940 (1989).
- Dean, F. B. Rapid Amplification of Plasmid and Phage DNA Using Phi29 DNA Polymerase and Multiply-Primed Rolling Circle Amplification. Genome research. 11 (6), 1095-1099 (2001).
- Hutchison, C. A., Smith, H. O., Pfannkoch, C., Venter, J. C. Cell-free cloning using ø29 DNA polymerase. Proceedings of the National Academy of Sciences of the United States of America. 102 (48), 17332-17336 (2005).
- Goecks, J., Nekrutenko, A., Taylor, J., Galaxy Team, T. Galaxy: a comprehensive approach for supporting accessible, reproducible, and transparent computational research in the life sciences. Genome Biology. 11 (8), 86 (2010).
- Giardine, B., Riemer, C., et al. Galaxy: A platform for interactive large-scale genome analysis. Genome research. 15 (10), 1451-1455 (2005).
- Langmead, B., Salzberg, S. L. Fast gapped-read alignment with Bowtie 2. Nature Methods. 9 (4), 357 (2012).
- Tsalik, E. L., Gartenberg, M. R. Curing Saccharomyces cerevisiae of the 2 micron plasmid by targeted DNA damage. Yeast. 14 (9), 847-852 (1998).
- Norman, A., Riber, L., Luo, W., Li, L. L., Hansen, L. H., Sørensen, S. J. An Improved Method for Including Upper Size Range Plasmids in Metamobilomes. PLoS ONE. 9 (8), e104405 (2014).
- Storlazzi, C. T., Lonoce, A., et al. Gene amplification as double minutes or homogeneously staining regions in solid tumors: Origin and structure. Genome research. 20 (9), 1198-1206 (2010).
- Von Hoff, D. D., Needham-VanDevanter, D. R., Yucel, J., Windle, B. E., Wahl, G. M. Amplified human MYC localized to replicating submicroscopic circular DNA molecules. Proceedings of the National Academy of Sciences of the United States of America. 85 (13), 4804-4808 (1988).
- Raymond, E., Faivre, S., et al. Effects of hydroxyurea on extrachromosomal DNA in patients with advanced ovarian carcinomas. Clinical cancer research : an official journal of the American Association for Cancer Research. 7 (5), 1171-1180 (2001).
- Shimizu, N. Extrachromosomal Double Minutes and Chromosomal Homogeneously Staining Regions as Probes for Chromosome Research. Cytogenetic and genome research. 124 (3-4), 3-4 (2009).
- Eckhardt, S. G., Dai, A., Davidson, K. K., Forseth, B. J., Wahl, G. M., Von Hoff, D. D. Induction of differentiation in HL60 cells by the reduction of extrachromosomally amplified c-myc. Proceedings of the National Academy of Sciences of the United States of America. 91 (14), 6674-6678 (1994).
- Vogt, N., Lefèvre, S. -. H., et al. Molecular structure of double-minute chromosomes bearing amplified copies of the epidermal growth factor receptor gene in gliomas. Proceedings of the National Academy of Sciences of the United States of America. 101 (31), 11368-11373 (2004).
- Ahn, K., Gotay, N., et al. High rate of disease-related copy number variations in childhood onset schizophrenia. Molecular psychiatry. 19 (5), 568-572 (2013).
- Girirajan, S., Johnson, R. L., et al. Global increases in both common and rare copy number load associated with autism. Human molecular genetics. 22 (14), 2870-2880 (2013).
- Vogt, N., Gibaud, A., Lemoine, F., de la Grange, P., Debatisse, M., Malfoy, B. Amplicon rearrangements during the extrachromosomal and intrachromosomal amplification process in a glioma. Nucleic Acids Research. , (2014).
Reprints and Permissions
Request permission to reuse the text or figures of this JoVE article
Request PermissionExplore More Articles
This article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. All rights reserved