
Method Article
شبكات المجهر الإلكتروني للإرسال بالنمط المجهري لتحديد مواقع الخلايا مباشرة ضمن سير عمل التصوير المقطعي بالتبريد والإلكترون للخلية الكاملة
* These authors contributed equally
In This Article
Summary
الهدف من هذا البروتوكول هو توجيه التصاق الخلايا ونموها إلى المناطق المستهدفة من الشبكات للفحص المجهري الإلكتروني المبرد. ويتحقق ذلك من خلال تطبيق طبقة مضادة للقاذورات يتم استئصالها في أنماط محددة من قبل المستخدم متبوعة بترسب بروتينات المصفوفة خارج الخلية في المناطق المنقوشة قبل بذر الخلايا.
Abstract
التصوير المقطعي بالتبريد والإلكترون كامل الخلية (cryo-ET) هو تقنية قوية تستخدم لإنتاج هياكل دقة على مستوى النانومتر للجزيئات الكبيرة الموجودة في السياق الخلوي والمحفوظة في حالة رطبة مجمدة شبه أصلية. ومع ذلك ، هناك تحديات مرتبطة بزراعة و / أو التمسك بالخلايا على شبكات TEM بطريقة مناسبة للتصوير المقطعي مع الاحتفاظ بالخلايا في حالتها الفسيولوجية. هنا ، يتم تقديم بروتوكول مفصل خطوة بخطوة حول استخدام النمط الدقيق لتوجيه وتعزيز نمو الخلايا حقيقية النواة على شبكات TEM. أثناء النمط الدقيق ، يتم توجيه نمو الخلايا عن طريق إيداع بروتينات المصفوفة خارج الخلية (ECM) ضمن أنماط ومواضع محددة على رقائق شبكة TEM بينما تظل المناطق الأخرى مغلفة بطبقة مضادة للقاذورات. المرونة في اختيار طلاء السطح وتصميم النمط تجعل النمط الدقيق قابلا للتطبيق على نطاق واسع لمجموعة واسعة من أنواع الخلايا. يعد النمط الدقيق مفيدا لدراسات الهياكل داخل الخلايا الفردية بالإضافة إلى الأنظمة التجريبية الأكثر تعقيدا مثل التفاعلات بين المضيف والممرض أو المجتمعات متعددة الخلايا المتباينة. يمكن أيضا دمج النمط الدقيق في العديد من مهام سير عمل Cryo-ET للخلايا الكاملة في المصب ، بما في ذلك المجهر الضوئي والإلكتروني المرتبط (cryo-CLEM) وطحن الحزمة الأيونية المركزة (cryo-FIB).
Introduction
مع تطور وتوسع وتنوع المجهر الإلكتروني المبرد (cryo-EM) ، فحص الباحثون مجموعة واسعة من العينات البيولوجية في حالة شبه أصلية من الجزيئات الكبيرة (~ 1 نانومتر) إلى دقة عالية (~ 2 Å). من الأفضل تطبيق تقنيات cryo-EM أحادية الجسيم وحيود الإلكترون على الجزيئات الكبيرة النقية في محلول أو في حالة بلورية ، على التوالي1,2. في حين أن التصوير المقطعي الإلكتروني المبرد (cryo-ET) مناسب بشكل فريد للدراسات الهيكلية وفوق الهيكلية شبه الأصلية للأجسام الكبيرة غير المتجانسة مثل البكتيريا والفيروسات متعددة الأشكال والخلايا حقيقية النواة3. في cryo-ET ، يتم الحصول على معلومات ثلاثية الأبعاد (3D) عن طريق إمالة العينة فعليا على مرحلة المجهر والحصول على سلسلة من الصور من خلال العينة بزوايا مختلفة. غالبا ما تغطي هذه الصور، أو سلسلة الإمالة، نطاقا يتراوح بين +60/-60 درجة بزيادات من درجة إلى ثلاث درجات. يمكن بعد ذلك إعادة بناء سلسلة الإمالة حسابيا إلى حجم 3D ، المعروف أيضا باسم التصوير المقطعي 4.
تتطلب جميع تقنيات التبريد الكهرومغناطيسي أن تكون العينة مدمجة في طبقة رقيقة من الجليد الزجاجي غير المتبلور وغير البلوري. واحدة من تقنيات التثبيت بالتبريد الأكثر استخداما هي التجميد الغطس ، حيث يتم تطبيق العينة على شبكة EM ، وتلطخ ، وتغرق بسرعة في الإيثان السائل أو خليط من الإيثان السائل والبروبان. هذه التقنية كافية لتزجيج العينات من <100 نانومتر إلى ~ 10 ميكرومتر في السمك ، بما في ذلك الخلايا البشرية المستزرعة ، مثل خلايا HeLa5,6. يمكن تزجيج العينات الأكثر سمكا، مثل المواد العضوية الصغيرة أو خزعات الأنسجة، التي يصل سمكها إلى 200 ميكرومتر، عن طريق التجميد عالي الضغط7. ومع ذلك ، نظرا لزيادة تشتت الإلكترونات للعينات الأكثر سمكا ، تقتصر العينة وسماكة الجليد ل cryo-ET على ~ 0.5 - 1 ميكرومتر في المجاهر الإلكترونية الناقلة 300 كيلو فولت. لذلك ، يقتصر التبريد الكامل للخلايا ET للعديد من الخلايا حقيقية النواة على محيط الخلية أو امتدادات الخلايا ما لم يتم استخدام خطوات إضافية لإعداد العينة ، مثل التقسيم بالتبريد8 أو طحن الحزمة الأيونية المركزة9،10،11.
يتمثل أحد القيود المفروضة على العديد من تجارب التصوير بالتبريد ET للخلايا الكاملة في نقل البيانات12. على عكس الجسيم الواحد cryo-EM ، حيث يمكن في كثير من الأحيان تصوير الآلاف من الجسيمات المعزولة من مربع شبكة TEM واحد ، فإن الخلايا كبيرة ومنتشرة ، ويجب زراعتها بكثافة منخفضة بما يكفي للسماح بحفظ الخلايا في طبقة رقيقة من الجليد الزجاجي. غالبا ما تقتصر منطقة الاهتمام على ميزة معينة أو منطقة فرعية معينة من الخلية. ومما يحد من الإنتاجية أيضا ميل الخلايا إلى النمو في المناطق غير القابلة لتصوير TEM ، مثل قضبان شبكة TEM أو بالقرب منها. نظرا للعوامل غير المتوقعة المرتبطة بزراعة الخلايا على شبكات TEM ، هناك حاجة إلى تطورات تكنولوجية لتحسين إمكانية الوصول إلى العينات والإنتاجية للحصول على البيانات.
النمط الدقيق للركيزة مع بروتينات المصفوفة خارج الخلية الملتصقة (ECM) هي تقنية راسخة للفحص المجهري الضوئي للخلايا الحية لتوجيه نمو الخلايا على أسطح صلبة ومتينة وشفافة بصريا مثل الزجاج وركائز زراعة الأنسجة الأخرى13,14. كما تم إجراء النمط الدقيق على الأسطح الناعمة و / أو ثلاثية الأبعاد (3D). ولم تسمح هذه التقنيات بتحديد المواقع بدقة للخلايا فحسب؛ بل سمحت أيضا بتحديد المواقع بدقة. كما دعموا إنشاء شبكات متعددة الخلايا، مثل دوائر الخلايا العصبية المنقوشة15. لن يؤدي جلب الأنماط الدقيقة إلى cryo-ET إلى زيادة الإنتاجية فحسب ، بل يمكن أن يفتح أيضا دراسات جديدة لاستكشاف البيئات الدقيقة الخلوية المعقدة والديناميكية.
في الآونة الأخيرة ، بدأت العديد من المجموعات في استخدام تقنيات النمط المصغر على شبكات TEM من خلال نهج متعددة16,17. هنا ، يتم وصف استخدام تقنية النمط الضوئي بدون قناع لشبكات TEM باستخدام نظام Alvéole PRIMO micropatterning ، والذي يتميز بنقوش عالية الدقة وبدون تلامس. باستخدام نظام الأنماط الدقيقة هذا ، يتم تطبيق طبقة مضادة للقاذورات في الجزء العلوي من الركيزة ، تليها تطبيق محفز ضوئي واستئصال الطبقة المضادة للقاذورات في أنماط يحددها المستخدم باستخدام ليزر الأشعة فوق البنفسجية. يمكن بعد ذلك إضافة بروتينات ECM إلى أنماط زراعة الخلايا المناسبة. تم استخدام هذه الطريقة من قبل عدة مجموعات لدراسات cryo-ET لصبغة الشبكية الظهارية -1 (RPE1) ، والكلى الكلبية Madin-Darby -II (MDCKII) ، والخلايا الليفية القلفة البشرية (HFF) ، وخطوط الخلايا البطانية16،17،18. يتوافق نظام الأنماط الدقيقة هذا مع ركائز طبقة متعددة مضادة للقاذورات بالإضافة إلى كاشف محفز ضوئي سائل أو هلامي. يمكن اختيار مجموعة متنوعة من بروتينات ECM وتكييفها لخصوصية خط الخلية ، مما يمنح براعة للمستخدم.
وقد تم تطبيق النمط المصغر بنجاح على عدد من المشاريع داخل المختبر19. هنا ، يتم تقديم بروتوكول للنمط الدقيق ، بما في ذلك تعديلات محددة لدراسة خلايا HeLa المستزرعة ، وخلايا BEAS-2B المصابة بالفيروس المخلوي التنفسي (RSV) ، والخلايا العصبية اليرقية الأولية Drosophila melanogaster 20.
Protocol
البروتوكول الموصوف هنا هو تجميع لثقافة الخلايا ، والنمط الدقيق ، وطرق التصوير المستخدمة من قبل مختبر رايت ومركز أبحاث Cryo-EM في جامعة ويسكونسن ، ماديسون. يتم عرض سير العمل في الشكل 1. تتوفر مواد تدريبية وتعليمية إضافية في المواقع التالية: https://cryoem.wisc.edu أو https://wrightlab.wisc.edu
1. إعداد الشبكات للتنسيق
- انقل شبكات TEM إلى شريحة زجاجية نظيفة ، جانب الكربون لأعلى (السماكة القياسية لرقائق الكربون هي 12 نانومتر). باستخدام مبخر الكربون ، ACE600 ، تبخر 5-8 نانومتر من الكربون الإضافي على الشبكات لزيادة المتانة الإجمالية لفيلم الكربون.
ملاحظة: هذه الخطوة ليست ضرورية لشبكات SiO2 . ويمكن أيضا القيام بهذه الخطوة مقدما؛ قم بتخزين الشبكات المطلية في بيئة منخفضة الرطوبة مثل مجفف الفراغ. - انقل الشبكات إلى حامل إعداد الشبكة وقم بتفريغ الشبكات بجانب الكربون لأعلى. باستخدام نظام تفريغ التوهج ، قم بتفريغ الشبكات لمدة 60 ثانية عند 10 مللي أمبير مع مسافة عمل 80 مم وضغط فراغ 1.0 × 10-3 مللي بار. قم بذلك في غضون 15-30 دقيقة من الخطوة التالية.
ملاحظة: يمكن شراء حاملات إعداد الشبكة تجاريا أو محلية الصنع باستخدام قطعة من ورق الترشيح على طبق بتري صغير.
2. تطبيق طبقة مضادة للقاذورات
ملاحظة: يجب استخدام تقنية التعقيم المناسبة عند التعامل مع الشبكات ، ويجب أن تكون جميع المحاليل معقمة و / أو معقمة بالفلتر.
- انقل الشبكات (جانب الكربون لأعلى) إلى شريحة زجاجية نظيفة أو غطاء مع فصل 1 سم على الأقل بين الشبكات. ماصة 10 ميكرولتر من 0.05٪ بولي L-ليسين (PLL) على كل شبكة. احتضن الشبكات في غرفة رطبة ، مثل صندوق بلاستيكي مغلق مع مناشف ورقية رطبة ، لمدة 30 دقيقة على الأقل.
ملاحظة: يمكن تمديد هذه الخطوة إلى بين عشية وضحاها. تأكد من أن مستوى الرطوبة في الغرفة يكفي لمنع الشبكات من الجفاف. - اغسل كل شبكة ثلاث مرات ب 15 ميكرولتر من 0.1 M HEPES الرقم الهيدروجيني 8.5. لكل غسلة ، قم بإزالة معظم السائل من الشبكة باستخدام ماصة دون ترك الشبكة تجف. أضف 15 ميكرولتر من المخزن المؤقت الطازج ، واحتضن لمدة 30 ثانية على الأقل وكرر. اترك كل شبكة في 15 ميكرولتر من 0.1 M HEPES بعد الغسيل النهائي.
ملاحظة: في هذه الخطوة والخطوات المستقبلية ، من المهم الحفاظ على الشبكة رطبة وتجنب الاتصال بين الماصة والشبكة. - تحضير 10 ميكرولتر من 100 ملغم / مل من البولي إيثيلين جلايكول -سكسينيميديل فاليرات (PEG-SVA) في 0.1 M HEPES الرقم الهيدروجيني 8.5 لكل شبكة. سوف يذوب PEG-SVA بسرعة مع الخلط اللطيف مما يؤدي إلى حل واضح.
ملاحظة: لا تقم بإعداد حل PEG-SVA مسبقا. PEG-SVA له عمر نصف يبلغ 10 دقائق عند درجة الحموضة 8.5. تجنب تعريض مخزون PEG-SVA للرطوبة الزائدة عن طريق تخزينه في مجفف أو بيئة جافة عند -20 درجة مئوية والاحترار إلى درجة حرارة الغرفة قبل الفتح. - مباشرة بعد تحضير محلول PEG-SVA ، قم بإزالة قطرة 15 ميكرولتر من HEPES pH 8.5 من كل شبكة (مع الحرص على عدم تجفيف الشبكة) وإضافة قطرة 10 ميكرولتر من محلول PEG-SVA. احتضان الشبكات في غرفة رطبة لمدة 1 ساعة على الأقل.
ملاحظة: يمكن تمديد هذه الخطوة إلى بين عشية وضحاها. تأكد من أن الرطوبة في الغرفة كافية لمنع الشبكات من الجفاف. - اغسل كل شبكة ثلاث مرات ب 15 ميكرولتر من الماء المعقم. لكل غسلة ، قم بإزالة معظم السائل من الشبكة باستخدام ماصة دون ترك الشبكة تجف ، وأضف 15 ميكرولتر من الماء العذب ، واحتضن لمدة 30 ثانية على الأقل وكرر. اترك كل شبكة في 15 ميكرولتر من الماء بعد الغسيل النهائي.
3. تطبيق هلام PLPP
- قم بإعداد غطاء مجهر نظيف لكل شبكة. أكمل الخطوات التالية لكل شبكة، شبكة واحدة في كل مرة، لتقليل فرصة جفاف الشبكة.
- ضع قطرة ماء سعة 1.0 ميكرولتر في وسط الغطاء للمساعدة في وضع الشبكة على الغطاء والحفاظ على الشبكة رطبة. انقل الشبكة بعناية من قطرة الماء 15 ميكرولتر إلى قطرة الماء 1.0 ميكرولتر على الغطاء. تأكد من وضع جانب الكربون الشبكي لأعلى.
- ضع استنسل بولي ديميثيل سيلوكسان (PDMS) بعناية فوق الشبكة ، مع الحرص على إبقاء الشبكة متمركزة وتقليل ملامسة الاستنسل لرقائق الكربون في الشبكة.
- أضف 1.0 ميكرولتر من هلام كلوريد 4-بنزويل بنزيل ثلاثي ميثيل الأمونيوم (PLPP) إلى الشبكة. ماصة بلطف للخلط (لا تلمس الشبكة بطرف الماصة).
- حرك زلة الغطاء مع الشبكة إلى مكان مظلم لتجف. سوف يجف الجل في حوالي 15-30 دقيقة.
4. معايرة وتصميم النمط الدقيق
- لون جانبا واحدا من غطاء زجاجي باستخدام قلم تمييز. أضف خطوطا سوداء من علامة دائمة ذات رؤوس دقيقة لتسهيل التركيز البؤري. ضع الغطاء على المجهر بحيث يواجه الجانب الملون العدسة الموضوعية. باستخدام وضع brightfield، ركز على أداة التمييز.
- تأكد من تشغيل المجهر ونظام النمط الدقيق ، ويتم تعيين مسار الضوء الصحيح. افتح Micromanager وبرنامج Leonardo (Plugins > Leonardo) على الكمبيوتر المجهري.
- حدد معايرة واتبع التعليمات التي تظهر على الشاشة. اضبط تركيز المجهر بحيث تكون الصورة المعروضة على الشريحة في بؤرة التركيز. قد يلزم تقليل وقت التعرض. بعد المعايرة، حدد النمط الآن.
- سجل نسبة الميكرومتر/البكسل (μm/px) المبلغ عنها تحت بيانات المعايرة في النافذة العلوية اليسرى من البرنامج (الشكل 2، المنطقة 1). استخدم هذه النسبة لتحديد عدد وحدات البكسل التي يجب استخدامها لكل ميكرومتر عند تصميم نمط.
- بعد المعايرة، تأكد من أن البرنامج مفتوح الآن من خلال عرض مباشر ل brightfield من المجهر. قم بتحميل شبكة معدة على شفة غطاء (القسم 3) على المسرح مع الشبكة التي تواجه العدسة الموضوعية. ضع المرحلة واضبط التركيز بحيث تكون الشبكة مرئية في نافذة البرنامج.
- قم بقياس حجم مربعات الشبكة وقضبان الشبكة بالميكرومتر. يتضمن البرنامج مسطرة يتم تنشيطها بواسطة الزر بالقرب من الزاوية اليسرى السفلى لقياس الشبكة (الشكل 2 ، المنطقة 2). على سبيل المثال ، تتوافق الأنماط المستخدمة هنا لشبكة شبكية 200 مع ~ 87 × مربعات شبكة 87 ميكرومتر و ~ 36 ميكرومتر قضبان الشبكة.
ملاحظة: يوفر البرنامج مرونة في تغيير حجم الأنماط أثناء التنقل ، لذلك يمكن تحمل عدم الدقة الطفيفة في القياس. - استنادا إلى القياسات والنسب المذكورة أعلاه ، قم بإنشاء نمط (أنماط) باستخدام أي برنامج لإنشاء الصور. الحد الأدنى لحجم الميزة مع هدف 20× هو 1.2 ميكرومتر. يجب حفظ الأنماط كملفات .tiff 8 بت غير مضغوطة.
- تأكد من أن البرنامج لا يعيد قياس الصور إلى حجم بكسل مختلف عند الحفظ. يجب أن يتناسب النمط مع مربع 800 × 800 بكسل ، وهو ما يكفي لتغطية أربعة مربعات شبكة.
ملاحظة: سيتم نقش وحدات البكسل ذات القيمة 255 (أبيض) بأعلى كثافة (الجرعة الإجمالية لليزر) ولن يتم نقش وحدات البكسل ذات القيمة صفر (أسود). سيتم نقش أي وحدات بكسل ذات قيمة وسيطة بجرعة تبلغ تقريبا (X/255)*الجرعة الإجمالية. في الشكل 3A ، تم استخدام قيم البكسل 255 و 129 لأنماط التدرج الرمادي. بمجرد تصميم النمط ، يمكن حفظه وإعادة استخدامه دون تعديل.
- تأكد من أن البرنامج لا يعيد قياس الصور إلى حجم بكسل مختلف عند الحفظ. يجب أن يتناسب النمط مع مربع 800 × 800 بكسل ، وهو ما يكفي لتغطية أربعة مربعات شبكة.
5. النمط المصغر
- بعد المعايرة، تأكد من أن البرنامج مفتوح الآن من خلال عرض مباشر ل brightfield من المجهر. قم بتحميل شبكة معدة على شفة غطاء (القسم 3) على المسرح مع الشبكة التي تواجه العدسة الموضوعية. ضع المسرح واضبط التركيز لرؤية الشبكة في البرنامج.
- للتشغيل الأولي، قم بتصميم قالب جديد. في البرنامج، حدد إضافة عائد استثمار (غير معروض، في موقع منطقة الشكل 2 3) واختر دائرة 3000 ميكرومتر. ضع عائد استثمار الدائرة فوق الشبكة باستخدام صورة brightfield على الشاشة كدليل. اضغط على القفل لتأمين عائد الاستثمار.
- بعد قفل عائد الاستثمار في مكانه، حدد إضافة نمط (غير معروض، في موقع منطقة الشكل 2 3). اختر النمط المصمم في القسم 4. قسم الشبكة إلى ست مناطق للسماح بالتركيز المستقل وتحديد المواقع في كل منطقة لمراعاة الشبكات غير المستوية. منطقة مربعة شبكة 8 × 8 لكل ركن من أركان الشبكة ومنطقة مربعة 2 × 8 شبكة على كل جانب من المركز ، تاركة المركز أربعة مربعات شبكة غير مرتبة (الشكل 2 ، الصورة المركزية).
- استخدم خيارات النسخ المتماثل (الشكل 2، المنطقة 4) لإنشاء نسخ من النمط الأولي للوصول إلى العدد المطلوب من إجمالي نسخ النمط. اضبط التباعد بين النسخ لمطابقة التباعد بين مربعات الشبكة إذا لزم الأمر.
- اضبط الجرعة الإجمالية للنمط. 30 mJ / mm2 هي نقطة انطلاق جيدة. راجع قسم المناقشة لمزيد من التفاصيل.
- ضمن خيارات الخبراء (الشكل 2، المساحة 4)، اضبط زاوية المنطقة لتتناسب مع زاوية مربعات الشبكة. يمكن تغيير موضع المناطق باستخدام الماوس. يمكن أيضا ضبط نسبة (حجم) الأنماط. كرر من خلال ضبط زاوية النمط وموضعه والمسافة بينه ونسبته حتى تصطف الأنماط مع مربعات الشبكة. حرك مرحلة المجهر لتغيير منطقة الشبكة في شاشة brightfield الحية.
- اضغط على Lock لحفظ التغييرات التي تم إجراؤها على المنطقة.
- لنسخ منطقة، انقر زر تكرار (الشكل 2، المنطقة 5، أيقونة ورقتين) بجوار اسمها في لوحة الإجراءات على اليمين. لتغيير موضع النسخة أو إعادة تسميتها أو تحريرها، انقر على اسمها في لوحة الإجراءات.
- كرر الخطوات من 5.4 إلى 5.9 حسب الضرورة لملء كافة المناطق المطلوبة.
- بمجرد تصميم القالب الكامل ووضعه، احفظ ملف القالب داخل البرنامج (الشكل 2، المنطقة 6، شريط مع رمز السهم لأعلى في شريط الأدوات العلوي).
- عند تحميل قالب محفوظ مسبقا (شريط مع رمز السهم لأسفل) قم بتوسيط عائد الاستثمار فوق الشبكة واضغط على Lock. انقر فوق كل منطقة في لوحة الإجراءات لتغيير ملف الزاوية والموضع والجرعة و/أو النمط.
- بمجرد وضع القالب والأنماط، قم بإلغاء تحديد جميع المناطق الموجودة في لوحة الإجراءات في البرنامج باستثناء منطقة واحدة.
- استخدم مرحلة المجهر للانتقال إلى تلك المنطقة والتركيز على رقائق الكربون. سيؤدي النقر فوق أيقونة مقلة العين في لوحة الإجراءات (الشكل 2، المنطقة 5) إلى تبديل عرض تراكب النقش أو إيقاف تشغيله.
- بمجرد أن تصبح الشبكة في بؤرة التركيز ، أغلق مصراع brightfield واضغط على أيقونة Play في الزاوية اليمنى السفلى من البرنامج لبدء عملية النقش ، والتي يمكن مراقبتها مباشرة.
- في لوحة الإجراءات، حدد المربع للمنطقة التالية. افتح مصراع brightfield بحيث تكون الشبكة مرئية ومركز تلك المنطقة باستخدام مرحلة المجهر. كرر الخطوات من 5.13 إلى 5.14 لكل منطقة في مجموعة العمل.
- قم بإزالة الغطاء مع الشبكة من المجهر ، وقم على الفور بإخراج 10 ميكرولتر من محلول ملحي معقم بالفوسفات المخزن مؤقتا (PBS) على الشبكة.
- بعد 10 دقائق ، قم بإزالة الاستنسل باستخدام ملاقط ، ثم اغسل الشبكة 3x مع 15 ميكرولتر من PBS. بعد الغسيل النهائي ، ضع كل شبكة في 15 ميكرولتر من PBS وانقل الشبكات إلى مكان مظلم.
6. ترسب بروتينات ECM
- بالنسبة للخلايا المستزرعة، اتبع الخطوات من 6.2 إلى 6.5؛ بالنسبة للخلايا العصبية الأولية لذبابة الفاكهة ، اتبع الخطوات 6.6-6.10.
- قم بإعداد ما لا يقل عن 15 ميكرولتر من ECM لكل شبكة. بالنسبة لخلايا BEAS-2B ، قم بإعداد تركيز نهائي قدره 0.01 ملغم / مل من الفيبرونيكتين البقري و 0.01 ملغ / مل من الفيبرينوجين المقترن بالفلوروفور في PBS المعقم. بالنسبة لخلايا HeLa ، قم بإعداد 0.01 ملغ / مل من الكولاجين البقري I و 0.1 ملغ / مل من الفيبرينوجين المترافق مع الفلوروفور في PBS المعقم.
- قم بإزالة معظم PBS من كل شبكة وتطبيق 15 ميكرولتر من ECM. احتضان الشبكة في غرفة رطبة في درجة حرارة الغرفة لمدة 1 ساعة على الأقل.
ملاحظة: قد يتم تمديد هذه الخطوة إلى بين عشية وضحاها عند 4 درجات مئوية. - بعد الحضانة في ECM ، اغسل كل شبكة 5x مع PBS معقمة. لكل غسلة ، قم بإزالة معظم السائل باستخدام ماصة دون ترك الشبكة تجف ، وأضف 15 ميكرولتر من PBS الطازج ، واحتضن لمدة 30 ثانية على الأقل ، وكرر. اترك كل شبكة في PBS بعد الغسيل النهائي.
ملاحظة: يمكن تخزين الشبكات لمدة تصل إلى أسبوع في PBS عند 4 درجات مئوية دون أي تدهور ملحوظ في الجودة. - استخدم المجهر الفلوري للكشف عن الفلوروفور في ECM لتأكيد النقش وأن رقائق الكربون ظلت سليمة. بعض المربعات المكسورة يمكن تحملها بشكل عام.
- بالنسبة للخلايا العصبية الأولية لذبابة الفاكهة ، انقل الشبكات المنقوشة إلى طبق سفلي زجاجي 30 مم يحتوي على PBS معقم.
- شفط PBS من الطبق وتطبيق 2 مل من 0.5 ملغ / مل concanavalin A. احتضان الفلوروفور المترافق بين عشية وضحاها عند 25 درجة مئوية في بيئة معقمة.
- قم بإزالة محلول concanavalin A من الطبق (دون تجفيف الشبكات) واغسل الشبكات 3x باستخدام PBS. لكل غسلة ، أضف وأزل 2 مل PBS من الطبق.
- استخدم المجهر الفلوري للكشف عن الفلوروفور في ECM لتأكيد النقش وأن رقائق الكربون ظلت سليمة. بعض المربعات المكسورة يمكن تحملها بشكل عام.
- بعد الغسيل النهائي، قم بإزالة PBS من الطبق السفلي الزجاجي وأضف 2 مل من وسائط ذبابة الفاكهة المكملة المعقمة والمعقمة من شنايدر21، والتي تحتوي على 20٪ من مصل الجنين البقري المعطل للحرارة (FBS)، و5 ميكروغرام/مل من الأنسولين، و100 ميكروغرام/مل من البنسلين، و100 ميكروغرام/مل من الستربتومايسين، و10 ميكروغرام/مل من التتراسيكلين. احتضن عند 25 درجة مئوية في بيئة معقمة حتى تصبح الخلايا العصبية جاهزة للطلاء.
7. إعداد خلايا ذبابة الفاكهة الأولية قبل البذر
- عقم طبق تشريح 55 مم مع 70٪ EtOH ، ثم أضف إلى الطبق 2-3 مل من محلول ملحي 1× مفلتر معقم (9.9 ملليمتر HEPES درجة الحموضة 7.5 ، 137 ملليمتر كلوريد الصوديوم ، 5.4 ملليمتر كلوريد الصوديوم ، 0.17 ملليمتر NaH2PO4 ، 0.22 ملليمتر KH2PO4 ، 3.3 ملليمتر جلوكوز ، 43.8 ملليمتر سكروز)21.
- اختر 30-40 يرقات 3rd instar بلطف من الطعام باستخدام زوج من الملقط.
- ضع اليرقات في الأنبوب مع 1× PBS ، ثم انقلها إلى الأنبوب الثاني مع 1× PBS لغسل اليرقات.
- انقل اليرقات إلى الأنبوب بنسبة 70٪ EtOH لغسل PBS ، ثم انقلها إلى الأنبوب الثاني بنسبة 70٪ EtOH. اترك اليرقات في الأنبوب الثاني لمدة 2-3 دقائق لتعقيم اليرقات.
- انقل اليرقات إلى أنبوب به محلول ملحي 1× تشريح ، ثم انقلها على الفور إلى الأنبوب الثاني مع 1× محلول ملحي للتشريح.
- انقل اليرقات الفردية إلى طبق التشريح الذي يحتوي على 1× محلول ملحي للتشريح. باستخدام زوج من الملقط ومجهر التشريح ، قم بتمزيق كل يرقات بسرعة لاستخراج الدماغ ونقله إلى الأنبوب الثالث بمحلول ملحي للتشريح × 1. كرر حتى يتم استخراج جميع الأدمغة.
- جهاز الطرد المركزي الأنبوب الذي يحتوي على الأدمغة عند 300 × جم لمدة 1 دقيقة.
- تخلص من المادة الفائقة ، واغسلها ب 1 مل من محلول ملحي للتشريح × وقم بطرد الأنبوب عند 300 × جم لمدة دقيقة واحدة. كرر هذه الخطوة مرة أخرى.
- تخلص من المادة الفائقة حتى يتم ترك 200-250 ميكرولتر في الأنبوب وأضف 20 ميكرولتر من 2.5 ملغم / مل Liberase في محلول ملحي 1x تشريح.
- تدوير الأنبوب على الدوار لمدة 1 ساعة في درجة حرارة الغرفة ؛ خلال هذه الساعة ، ماصة الحل 25-30 مرة كل 10 دقائق. في النهاية ، يجب أن يكون الحل غير شفاف قليلا.
- جهاز طرد مركزي الخلايا في 300 × غرام لمدة 5 دقائق.
- تخلص من المادة الفائقة، ثم أضف 1 مل من وسائط شنايدر المكملة. ماصة الحل 30 مرة للخلط.
- جهاز طرد مركزي للخلايا عند 300 × جم لمدة 5 دقائق.
- تخلص من المادة الفائقة واغسل حبيبات الخلايا بإضافة 1 مل من وسائط شنايدر المكملة. ماصة الحل 30 مرة للخلط.
- الطرد المركزي للخلايا في 300 × غرام لمدة 5 دقائق.
- تخلص من المادة الفائقة ، ثم أعد تعليق حبيبات الخلية مع 300 ميكرولتر من وسائط شنايدر المكملة. ماصة الحل 30-40 مرة للخلط.
8. ثقافة وعدوى RSV من خلايا BEAS-2B و HeLa
- الحفاظ على خلايا HeLa وخلايا BEAS-2B في قوارير T75 عند 37 درجة مئوية و 5٪ CO2. خلايا المرور كل 3-4 أيام بمجرد الوصول إلى ما يقرب من 80 ٪ التقاء. الحفاظ على خلايا HeLa في DMEM + 10٪ FBS + 1× مضاد حيوي مضاد للفطريات. الحفاظ على BEAS-2B في RPMI + 10٪ FBS + 1× مضاد حيوي مضاد للفطريات6،22،23.
- لبذر الخلايا غير المصابة ، انتقل إلى القسم 9. خلايا BEAS-2B و HeLa عرضة للإصابة بعدوى RSV ؛ تم استخدام خلايا BEAS-2B لجميع التجارب التي تنطوي على RSV الموضحة هنا.
ملاحظة: تنفيذ جميع خطوات BSL-2 وفقا للبروتوكولات المؤسسية باستخدام خزانة مناسبة للسلامة الأحيائية (BSC) ومعدات الحماية الشخصية (PPE). - قبل عدوى RSV للخلايا ، يمر 5 × 104 خلايا لكل بئر في صفيحة من 6 آبار (مساحة السطح ~ 9.6 سم 2) مع 2 مل من وسائط النمو ويحضن بين عشية وضحاها.
- Trypsinize وعد بئر واحد من الخلايا. للتريبسين ، قم بشفط الوسائط من بئر واحد واغسلها ب 2 مل من PBS المعقم بدون Mg2 + و Ca2 + لإزالة الوسائط المتبقية. أضف 500 ميكرولتر من محلول التربسين بنسبة 0.25٪. الحضانة عند 37 درجة مئوية لمدة 5-10 دقائق. تحقق بشكل دوري من الخلايا لمعرفة ما إذا كانت قد تم إطلاقها من السطح. بمجرد تحرير الخلايا ، أضف 1.5 مل من وسائط الثقافة.
- امزج 100 ميكرولتر من الخلايا التربسينية مع 100 ميكرولتر من أزرق المثقبية. ماصة 10 ميكرولتر من مزيج الخلايا المخففة في مقياس للخلايا الدموية. عد الخلايا واحسب عدد الخلايا لكل بئر. استخدم هذا الرقم لحساب وزارة الداخلية أدناه.
- قم بإعداد تخفيف RSV-A2mK + 24 في وسائط النمو لتحقيق MOI من 10 لكل بئر في 750 ميكرولتر من الوسائط. يمكن حساب MOI من RSV-A2mK + من عيار وحدات التركيز الفلوري (FFU) من المخزون (على سبيل المثال: ل 1.0 × 105 خلايا لكل بئر ومخزون RSV من 1.0 × 108 FFU / مل ، تمييع المخزون الفيروسي 1:75 إلى 1 × 106 FFU / 750 ميكرولتر أو 1.33 × 106 FFU / مل).
- شفط الوسائط من الخلايا الموجودة في طبق 6 آبار وأضف 750 ميكرولتر من المحلول الفيروسي من الأعلى إلى كل بئر.
- هز اللوحة في درجة حرارة الغرفة لمدة 1 ساعة.
- بعد 1 ساعة ، اجعل الحجم الإجمالي لكل بئر يصل إلى 2 مل مع تسخين وسائط النمو مسبقا إلى 37 درجة مئوية ووضع اللوحة في حاضنة مضبوطة على 37 درجة مئوية مع 5٪ CO2 لمدة 6 ساعات.
- Trypsinize الخلايا لإطلاقها والمضي قدما في البذر كما هو موضح أدناه. بعد البذر ، احتضن الشبكات لمدة 18 ساعة إضافية قبل التجميد الغارق (لما مجموعه 24 ساعة بعد الإصابة).
9. بذر الخلايا على شبكات ذات أنماط دقيقة
- بالنسبة للخلايا المستزرعة، اتبع الخطوات من 9.2 إلى 9.8؛ بالنسبة للخلايا العصبية الأولية لذبابة الفاكهة ، اتبع 9.9-9.11.
- جرب الخلايا لتحريرها (انظر الخطوة 4 في القسم 8 أعلاه). لتقليل تجميع الخلايا، جرب الخلايا عند التقاء 60٪ أو أقل.
- امزج 100 ميكرولتر من الخلايا التربسينية مع 100 ميكرولتر من أزرق المثقبية. ماصة 10 ميكرولتر من الخلية المخففة مزيج في مقياس للخلايا الدموية وعد الخلايا.
- تمييع الخلايا في وسائل الإعلام إلى 2 × 104 خلية / مل.
- أضف 1 ميكرولتر من الوسائط إلى وسط طبق سفلي زجاجي 30 مم للمساعدة في وضع الشبكة ومنعها من الجفاف. انقل الشبكة من PBS على الغطاء إلى وسط الطبق السفلي الزجاجي. أضف 10 ميكرولتر من محلول الخلية إلى الشبكة.
- باستخدام مجهر برايتفيلد ، راقب التصاق الخلية بالشبكة بعد 5 دقائق. إذا ظلت غالبية الأنماط غير مشغولة ، فأضف قطرة إضافية تبلغ 10 ميكرولتر من محلول الخلية. حافظ على الشبكات ومحلول الخلايا عند 37 درجة مئوية أثناء الحضانة.
- كرر الخطوة 9.6 حتى يتم شغل معظم الأنماط أو تبدأ العديد من الأنماط المشغولة في الحصول على خلايا متعددة. احتضان الشبكة لمدة 2 ساعة في الحاضنة (37 درجة مئوية ، 5 ٪ CO2).
- غمر الطبق ب 2 مل من الوسائط المسخنة مسبقا واحتضنها بين عشية وضحاها (37 درجة مئوية ، 5٪ CO2).
- بالنسبة للخلايا العصبية الأولية لذبابة الفاكهة ، قم بإزالة الوسائط من الطبق المحتوي على الشبكة وقم بوضع الخلايا على الطبق.
- انتظر 30-60 دقيقة حتى تلتصق الخلايا، ثم أضف 2 مل من وسائط شنايدر المكملة.
- قم بزراعة الخلايا العصبية في حاضنة 25 درجة مئوية لمدة لا تقل عن 2-3 أيام قبل الغطس والتجميد.
10. تصوير وتزجيج الشبكات المنقوشة
- ضع الطبق السفلي الزجاجي الذي يحتوي على الشبكة المنقوشة والخلايا المستزرعة على المجهر الفلوري.
- احصل على صور للشبكة باستخدام برايتفيلد وقنوات الفلورسنت المناسبة للكشف عن النمط وأي ملصقات أخرى في الخلايا. تأكد من أن كثافة الخلايا وتحديد مواقعها مناسبان للتصوير والتحليل في المراحل النهائية.
ملاحظة: تمت معالجة صور برايتفيلد والفلورسنت في حزمة برامج فيجي25. - إعداد الفريزر الغطس بالتبريد. يعتمد نوع جهاز التجميد على التوافر والتكلفة والميزات الأكثر ملاءمة للعينة.
ملاحظة: تم تحضير الخلايا العصبية الأولية لذبابة الفاكهة على ثلاجة غطس آلية ، وتم إعداد خلايا BEAS-2B باستخدام ثلاجة غطس شبه آلية. - ضع مدخلات الذهب على العينات من أجل المحاذاة المناسبة لسلسلة الإمالة. قم بمسح العينات لإزالة الوسائط الزائدة ، ثم قم بتجميد العينات في مادة مبردة ، مثل الإيثان السائل المبرد بالنيتروجين السائل. بالنسبة للخلايا العصبية الأولية لذبابة الفاكهة ، لطخة لمدة 4 ثوان من الجانب الخلفي. بالنسبة لخلايا HeLa و BEAS-2B ، لطخة من كلا الجانبين لمدة 4-6 ثانية. يمكن بعد ذلك تخزين الشبكات المجمدة في النيتروجين السائل حتى مزيد من الاستخدام.
- صورة الخلايا المزججة في المجهر الإلكتروني المبرد ، تعمل عند 300 كيلو فولت باستخدام كاميرا كاشف الإلكترون المباشر. قم بإعداد مجموعة سلسلة الإمالة لكل منطقة ذات أهمية باستخدام برنامج مثل SerialEM26 لجمع بيانات cryo-EM/cryo-ET.
ملاحظة: تم جمع سلسلة الميل من الخلايا العصبية الأولية لذبابة الفاكهة على كاشف إلكترون مباشر من -60 درجة إلى 60 درجة ثنائي الاتجاه بزيادات 2 درجة عند إلغاء التركيز البؤري -8 ميكرومتر مع حجم بكسل 4.628 Å لجرعة إجمالية قدرها 70-75 e-/Å2. تم جمع سلسلة الإمالة من BEAS-2B المصاب بفيروس RSV على كاشف إلكترون مباشر مع مرشح طاقة (شق 20 eV) عند إلغاء التركيز البؤري -5 ميكرومتر بحجم بكسل 4.603 Å وجرعة إجمالية ~ 80 e-/Å2. - قم بمعالجة سلسلة الإمالة لإعادة بناء التصوير المقطعي.
ملاحظة: أعيد بناء التصوير المقطعي المعروض هنا باستخدام حزمة IMOD27؛ تم إجراء تصفية lowpass باستخدام حزمة برامج EMAN228.
النتائج
تم استخدام هذا الإجراء لنمط شبكات EM لتجارب cryo-ET الخلوية الكاملة. يشمل سير العمل بأكمله المقدم في هذه الدراسة ، بما في ذلك الاستعدادات الأولية لزراعة الخلايا ، والنمط الدقيق (الشكل 1) ، والتصوير ، 3-7 أيام. تم استخدام إجراء من خطوتين لتوليد الطبقة المضادة للقاذورات عن طريق تطبيق PLL على الشبكة ومن ثم ربط PEG عن طريق إضافة PEG-SVA التفاعلي. يمكن أيضا تطبيق الطبقة المضادة للقاذورات في خطوة واحدة عن طريق إضافة PLL-g-PEG في حضانة واحدة. هلام PLPP هو محفز للنمط الدقيق للأشعة فوق البنفسجية ، والذي يتوفر أيضا كسائل أقل تركيزا. يسمح الجل بالنقش بجرعة مخفضة بشكل كبير مقارنة بالسائل ، مما يؤدي إلى نقش أسرع بكثير. مع هذا النظام ، كان وقت النقش الفعلي لشبكة TEM الكاملة ~ 2 دقيقة. يمتد سير عمل النمط المصغر وحده بشكل عام من 5 إلى 6 ساعات ويسمح للفرد بنمط ثماني شبكات لزراعة الخلايا القياسية على شبكات TEM.
يتطلب عدد من الخطوات أثناء عملية النمط المصغر أوقات حضانة طويلة (انظر الخطوات 2.1 و 2.3 و 6.4). بشكل مريح ، يمكن تمديد بعض هذه الخطوات ، مثل تخميل PLL (2.1) أو تخميل PEG-SVA (2.3) ، إلى حضانة بين عشية وضحاها. بالإضافة إلى ذلك ، قد يتم تصميم الشبكات مسبقا وتخزينها في محلول من بروتين ECM أو PBS لاستخدامها لاحقا. في دراستنا ، كانت هذه الخيارات قيمة في الحالات التي يكون فيها توقيت إعداد الخلايا والبذر أمرا بالغ الأهمية ، مثل الخلايا العصبية الأولية لذبابة الفاكهة وعدوى RSV لخلايا BEAS-2B.
تم إعداد الشبكات في إعداد مختبر عام على مستوى السلامة الأحيائية 2 (BSL-2) باستخدام أدوات نظيفة ومحاليل معقمة وشملت المضادات الحيوية / مضادات الفطريات في وسائط النمو 6،22،29،30. بالنسبة للعينات الحساسة بشكل خاص للتلوث الميكروبي ، يمكن تطبيق الطبقة المضادة للقذورات و ECM في غطاء زراعة الأنسجة أو بيئة معقمة أخرى. بالإضافة إلى ذلك ، يمكن غسل الشبكة في الإيثانول بين النقش وتطبيق ECM. وفي حالة العمل مع العوامل المعدية، من المهم تكييف الإجراء للامتثال لبروتوكولات السلامة الأحيائية المناسبة.
سمح سير العمل هذا والإجراءات المقدمة (الشكل 1) بزرع خلايا HeLa (الشكل 4) ، وخلايا BEAS-2B المصابة بفيروس RSV (الشكل 3 ، الشكل 5) ، والخلايا العصبية اليرقية الأولية لذبابة الفاكهة (الشكل 6 ، الشكل 7) على شبكات EM منقوشة لجمع بيانات cryo-ET على النحو الأمثل.
تظل خلايا HeLa المزروعة على شبكات TEM ذات الأنماط الدقيقة قابلة للحياة على النحو الذي يحدده تلطيخ الفلورسنت باستخدام فحص قابلية صلاحية الخلايا القائمة على calcein-AM و ethidium homodimer-1 (الشكل 4A ، B). باستخدام ECM مختلط من الكولاجين والفيبرينوجين ، تلتصق خلايا HeLa بسهولة بالأنماط عبر الشبكة (الشكل 4A ، C). التشكل العام للخلايا التي تتوسع على طول النمط مشابه لمورفولوجيا الخلايا المزروعة على شبكات غير نمطية (الشكل 4C، D). في حالة خلايا HeLa ، يبقى سمك الخلية الكلي ~ < 10 ميكرومتر مع مناطق أرق بكثير ~ < سمكها 1 ميكرومتر بالقرب من محيط الخلية (الشكل 4E ، F).
بالنسبة لدراسات RSV ، قمنا بنقش مربعات الشبكة بأكملها باستخدام تدرج ، مع التعرض لجرعة منخفضة على الحواف ونمط جرعة أعلى نحو المركز (الشكل 3A). أسفرت أنماط التدرج عن نتائج أفضل عند البحث عن الفيروسات المطلقة الموجودة بالقرب من محيط الخلايا. مع هذه الأنماط ، وجد أن الخلايا تلتزم بشكل تفضيلي بتركيز ECM الأعلى ، ولكنها قادرة أيضا على الالتصاق بتركيزات ECM المنخفضة والنمو عليها. ستحتاج الجرعة النسبية بين المناطق إلى التحسين عند استخدام الأنماط التي تتطلب جرعات متعددة. إذا كانت الجرعات وبالتالي تركيزات ECM متشابهة جدا أو متباينة جدا مع بعضها البعض ، فقدان تأثير استخدام جرعات متعددة.
في الشكل 3 ، تم تصميم شبكة TEM ثم زرعت بخلايا BEAS-2B المصابة بفيروس RSV واستخدمت لجمع بيانات cryo-EM. الشكل 4A هو صورة فلورية ل ECM منقوشة على شبكة TEM باستخدام نمط تدرج. يمكن رؤية التصاق الخلايا ونموها على طول المنطقة الوسطى من النمط في الشكل 3B كصورة برايتفيلد للخلايا بعد 18 ساعة من البذر. في الشكل 3C ، يتم تراكب إشارة الفلورسنت (الحمراء) من تكرار RSV-A2mK + مع إشارة من ECM. يتم وضع غالبية الخلايا المصابة على طول المنطقة الوسطى ذات الكثافة العالية من نمط التدرج. تكشف خريطة TEM منخفضة الصهارة لتثبيت الشبكة بعد التبريد عن عدد من الخلايا ، بما في ذلك الخلايا المصابة بفيروس RSV ، الموضوعة على رقائق الكربون بالقرب من مركز مربعات الشبكة. وكما هو مبين سابقا بالنسبة للخلايا المزروعة على شبكات TEM القياسية22، تم تحديد موقع سلسلة الإمالة وجمعها من فيروسات RSV على مقربة من محيط خلايا BEAS-2B المصابة المزروعة على شبكات ذات أنماط دقيقة (الشكل 5A، B). يمكن تحديد العديد من البروتينات الهيكلية RSV داخل التصوير المقطعي بما في ذلك nucleocapsid (N) وبروتين الاندماج الفيروسي (F) (الشكل 5C ، الأسهم الزرقاء والحمراء على التوالي).
بالنسبة للدراسات الأولية للخلايا العصبية لذبابة الفاكهة ، وجد أن النمط الضيق ، بالقرب من حد الدقة الذي يوفره البرنامج (حيث كان سمك النمط 2 ميكرومتر) ، يسمح بعزله من خلية واحدة إلى بضع خلايا داخل مربع شبكة (الشكل 6). كانت السوما العصبية قادرة على تمديد عصبياتها على مدى عدة أيام ضمن النمط. وقد سمح ذلك بسهولة التعرف على الخلايا العصبية المكتسبة من الخلايا العصبية المزروعة على شبكات غير منقوشة والحصول على سلسلة إمالة لها مقارنة بالخلايا العصبية المزروعة على شبكات غير نمطية (الشكل 7). ووجد أيضا أن concanavalin A المسمى بالفلورسنت ، وهو الليكتين الذي تم استخدامه كإدارة ECM للثقافات العصبية Drosophila في المختبر20,21 ، قابل للتنسيق.
تم عزل الخلايا العصبية ذبابة الفاكهة من يرقات النجم الثالث وفقا للبروتوكولات المنشورة سابقا20،21،31. تم تطبيق المستحضرات العصبية على شبكات cryo-EM ذات النمط الدقيق حيث تم إيداع concanavalin A على النمط لتنظيم وضع الخلايا وانتشارها وتنظيمها. سمح للخلايا العصبية الموجودة على الشبكات المنقوشة أو غير المنقوشة بالاحتضان لمدة لا تقل عن 48-72 ساعة ، ثم تم تجميد الشبكات. يظهر الشكل 6A صورة تمثيلية لشبكة EM ذات نمط مصغر مع العديد من الخلايا العصبية لذبابة الفاكهة الموزعة عبر المناطق المنقوشة. هذه الخلايا العصبية ، المشتقة من سلالة ذبابة معدلة وراثيا لها تعبير GFP عصبي في الغشاء ، يمكن تتبعها بسهولة عن طريق المجهر الضوئي ليس فقط بسبب وضع العلامات الفلورية ، ولكن أيضا بسبب موقعها داخل الأنماط الدقيقة. في حين يمكن أيضا تتبع الخلايا العصبية المستزرعة على شبكات غير نمطية من خلال إشارات GFP الخاصة بها بواسطة المجهر الضوئي (الشكل 7A ، الدائرة الصفراء) ، أصبح تحديد موقعها في cryo-EM أكثر صعوبة بكثير بسبب وجود الحطام الخلوي والتلوث من الوسائط (الشكل 7B ، الدائرة الصفراء). تم تقليل هذا الوجود للخلايا العصبية على الشبكات المنقوشة ، على الأرجح بسبب PEG في الطبقة المضادة للقاذورات في المناطق غير المنقوشة التي تصد حطام الخلية من الالتصاق. نظرا لأبعاد جسم الخلية العصبية والعصبيات الممتدة (الشكل 6A ، B ، الدائرة الصفراء) ، تم جمع سلسلة إمالة cryo-ET على طول المناطق الأرق من الخلايا (الشكل 6C ، D ، الدائرة الحمراء). تم حل غشاء الخلية العصبية ، والميتوكوندريون (السماوي) ، والأنابيب الدقيقة (الأرجواني) ، وخيوط الأكتين (الأزرق) ، والهياكل الحويصلية (البرتقالي والأخضر) ، والجزيئات الكبيرة مثل الريبوسومات (الحمراء) بشكل جيد في مونتاج الصور الأعلى تكبيرا وشرائح من خلال التصوير المقطعي ثلاثي الأبعاد (الشكل 6E). في حين يمكن رؤية ميزات مماثلة تحت الخلوية من التصوير المقطعي ثلاثي الأبعاد للخلايا العصبية غير المتناثرة (الشكل 7E) ، فإن صعوبة تحديد الأهداف الخلوية القابلة للتطبيق لجمع البيانات قللت الإنتاجية بشكل كبير.
في الشكل 8 ، تم تجميع صور تمثيلية من الشبكات مع بعض هذه المشكلات للمساعدة في تحديدها واستكشاف الأخطاء وإصلاحها. بمجرد تحديد الظروف المثلى ، يعد النمط الدقيق طريقة موثوقة وقابلة للتكرار لوضع الخلايا على شبكات cryo-TEM.

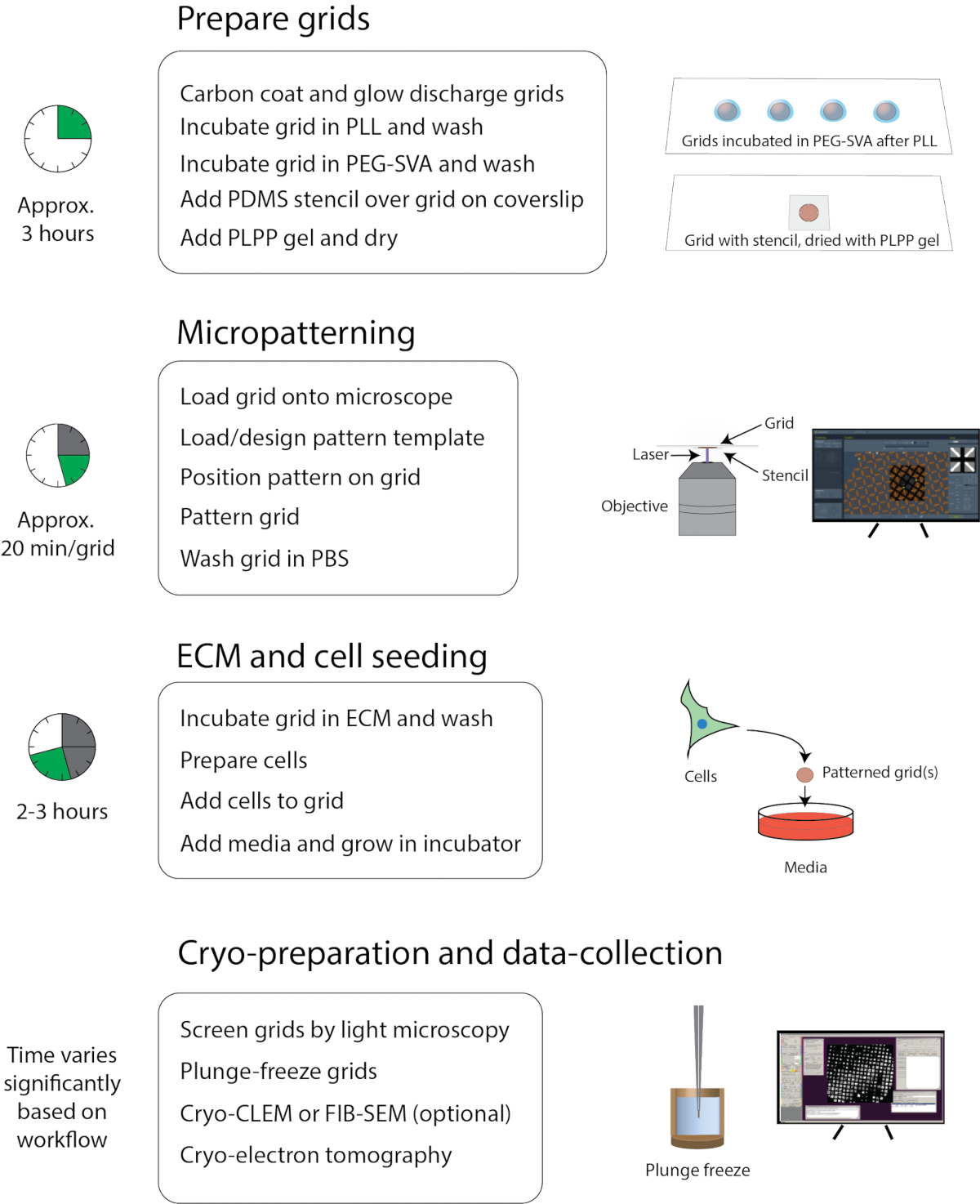
الشكل 1: سير العمل العام للنمط المصغر ل cryo-EM. يمكن تقسيم سير العمل تقريبا إلى أربعة أجزاء: إعداد الشبكة ، والنمط المصغر ، و ECM وبذر الخلايا ، وإعداد التبريد وجمع البيانات. يتم سرد الخطوات الرئيسية لكل قسم أسفل العناوين ويظهر الوقت التقريبي لإكمال كل قسم على اليسار. يرجى النقر هنا لعرض نسخة أكبر من هذا الشكل.
{kind=link}

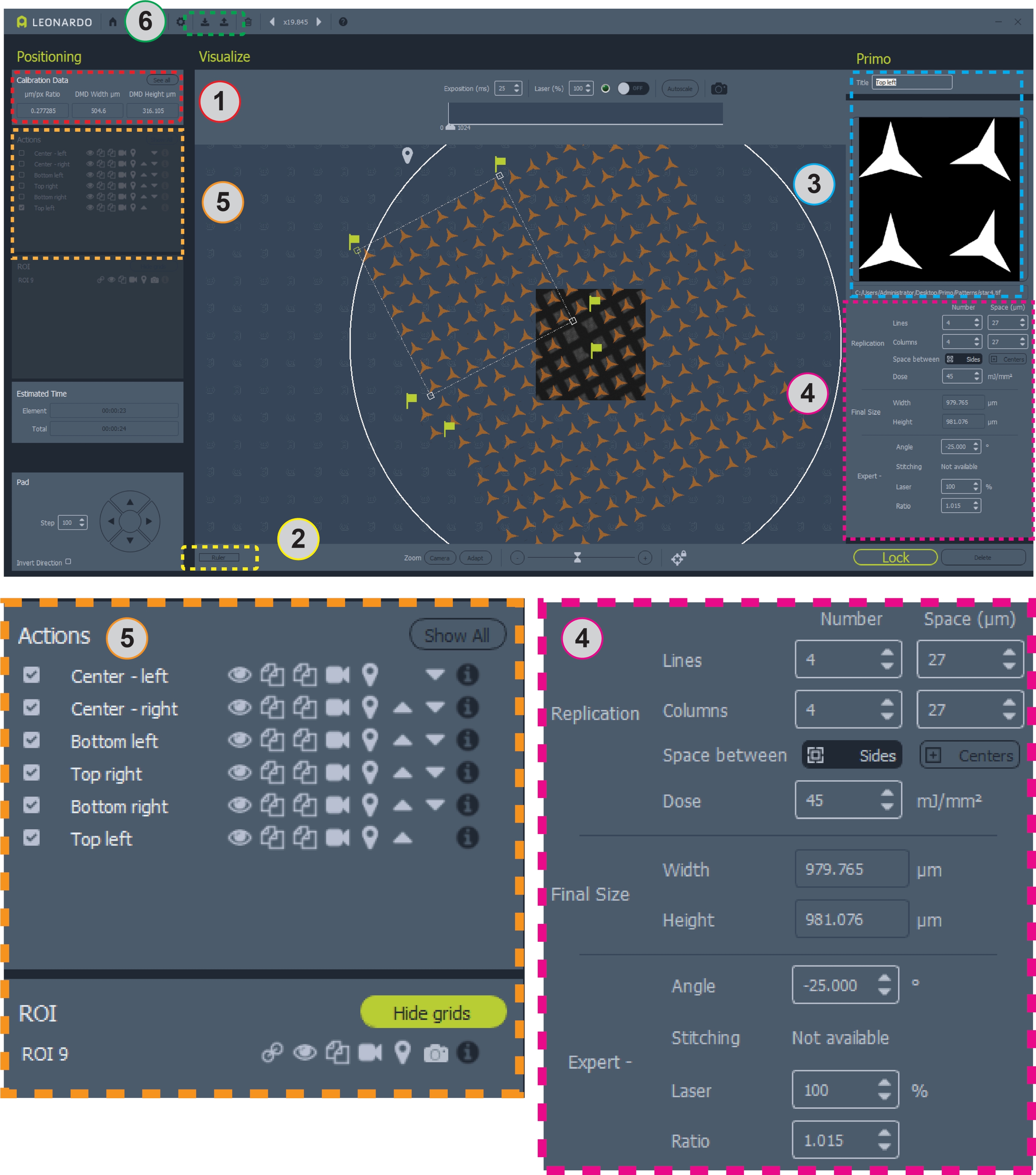
الشكل 2: لقطة شاشة للبرنامج مع وضع نمط على الشبكة. تحتوي المنطقة 1 على نسبة ميكرومتر/ بكسل لتصميم النمط. المساحة 2 هي المسطرة لقياس الشبكة. المنطقة 3 هي المكان الذي يمكن فيه إضافة أو تغيير الأنماط وعائد الاستثمار. تحتوي المنطقة 4 على جميع المعلومات الخاصة بتحديد موضع النمط والجرعة. تحتوي المنطقة 5 على خيارات للأنماط، بما في ذلك تبديل التراكبات ونسخ الأنماط أو حذفها وتحديد أنماط للأنماط الدقيقة. المنطقة 6 هي المكان الذي يمكن فيه حفظ القوالب وتحميلها. وترد أدناه مشاهدات أكبر للمناطق 4 و5 من أجل الوضوح. يرجى النقر هنا لعرض نسخة أكبر من هذا الشكل.
{kind=link}

الشكل 3: خلايا BEAS-2B المصابة بفيروس RSV على شبكة cryo-TEM المنقوشة. (A) صورة الفلورسنت للشبكة المنقوشة بعد إضافة ECM المسمى بالفلورسنت. يظهر نمط الإدخال في الزاوية السفلية اليسرى. (ب) صورة برايتفيلد لخلايا BEAS-2B المزروعة على الشبكة في A. (C) دمج الصورة في A (سماوي) و B (رمادي) مع صورة فلورسنت للخلايا المصابة بفيروس RSV (الأحمر) مباشرة قبل التجميد الغارق ؛ الخلايا المصابة تعبر عن mKate-2. أشرطة المقياس هي 500 ميكرومتر. (D) خريطة التبريد TEM منخفضة التكبير للشبكة في B بعد التجمد الغطس. صور الفلورسنت زائفة اللون. قضبان المقياس هي 500 ميكرومتر. يرجى النقر هنا لعرض نسخة أكبر من هذا الشكل.
{kind=link}

الشكل 4: تلطيخ الخلايا الحية/الميتة للخلايا المنقوشة وغير النمطية. (أ) صورة فلورسنت لخلايا HeLa المزروعة على شبكة منقوشة وملطخة ب calcein-AM (بقعة الخلية الحية ، الخضراء) و ethidium homodimer-1 (بقعة الخلايا الميتة ، حمراء). (ب) خلايا HeLa نمت على شبكة غير منقوشة وملطخة كما هو الحال في A. (C) إسقاط أكوام z البؤرية لخلية HeLa على شبكة Quantifoil R2/2 منقوشة مع 0.01 mg / mL الكولاجين والفيبرينوجين 647 ECM (أحمر). كانت الخلية ملطخة بالكالسين-AM (الأخضر) و Hoechst-33342 (الأزرق). (د) خلايا HeLa على شبكة غير منقوشة محتضنة ب 0.01 ملغم / مل من الكولاجين والفيبرينوجين 647 ECM ، محتضنة وملطخة ب calcein-AM و Hoecsht-33342. تم دمج الصور الفلورية مع الضوء المرسل (تدرج رمادي). (ه) إسقاط X,Z لإسقاط C. (F) X,Z ل D. الصور زائفة اللون. أشرطة المقياس في (A) و (B) هي 500 ميكرومتر ؛ أشرطة المقياس في (C) - (F) هي 10 ميكرومتر. يرجى النقر هنا لعرض نسخة أكبر من هذا الشكل.
{kind=link}

الشكل 5: Cryo-ET لخلية BEAS-2B المصابة بفيروس RSV على شبكة CRYO-TEM المنقوشة. (A) خريطة مربعة لشبكة Cryo-EM لخلية BEAS-2B المصابة بفيروس RSV. يشار إلى حدود الخلية التقريبية بالخط الأخضر المتقطع. (ب) صورة عالية الدقة للمساحة المربعة باللون الأحمر في (أ). يشار إلى حدود الخلية التقريبية بخط أخضر متقطع. يمكن رؤية فيروسات RSV بالقرب من محيط الخلية (السهم الأبيض والصندوق الأصفر). (ج) شريحة Z واحدة من التصوير المقطعي تم جمعها في منطقة الصندوق الأصفر في (B). تشير الأسهم الحمراء إلى بروتين الاندماج RSV F ، وتشير الأسهم الزرقاء إلى مركب البروتين النووي الريبي (RNP). يتم تضمين أشرطة المقياس في (A)-(C) في الصورة. يرجى النقر هنا لعرض نسخة أكبر من هذا الشكل.
{kind=link}

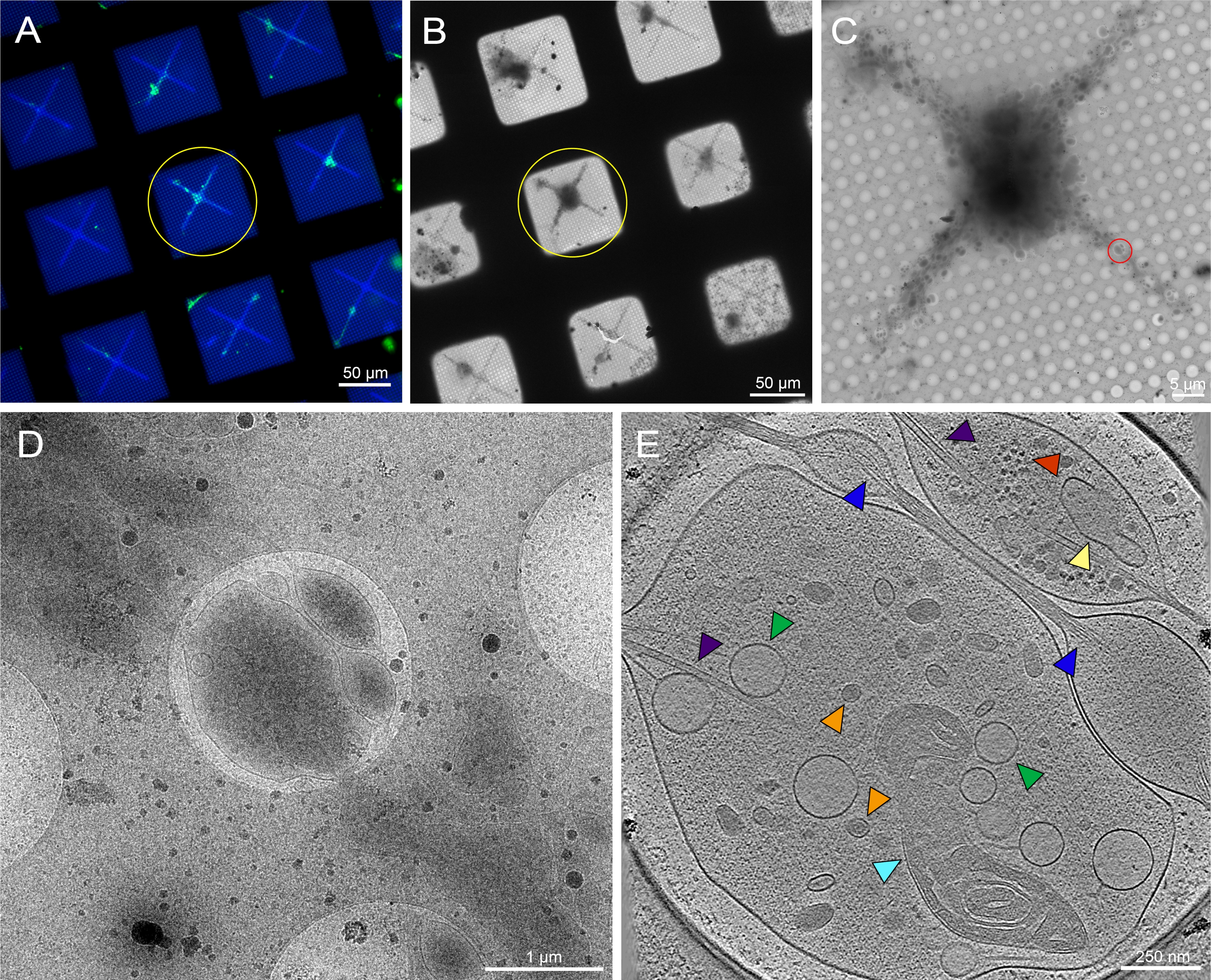
الشكل 6: الخلايا العصبية الأولية المشتقة من أدمغة يرقات ذبابة الفاكهة 3rd instar على شبكة cryo-TEM المنقوشة. (أ) مونتاج شبكة الفحص المجهري الفلوري للخلايا الحية المتراكبة للخلايا العصبية ذبابة الفاكهة التي تعبر عن GFP المستهدف بالغشاء على مربعات الشبكة المنقوشة مع 0.5 ملغم / مل من الكونكانافالين الفلوري A. الأخضر: الخلايا العصبية ذبابة الفاكهة . الأزرق: نمط ضوئي. (ب) مونتاج صورة Cryo-EM للشبكة في (A) بعد الحفظ بالتبريد. تشير الدائرة الصفراء إلى نفس مربع الشبكة كما في (A). (ج) مونتاج الصور Cryo-EM للمربع الذي تبرزه الدائرة الصفراء في (A) و (B). (د) صورة تكبير أعلى للمنطقة التي تحدها الدائرة الحمراء في (C)، حيث تم جمع سلسلة إمالة على عصبيات الخلية. E. شريحة بسمك 25 نانومتر من التصوير المقطعي المعاد بناؤها من سلسلة الإمالة التي تم الحصول عليها من الدائرة الحمراء في (C). يمكن رؤية عضيات مختلفة في هذا التصوير المقطعي ، مثل الميتوكوندريا (سماوي) ، والأنابيب الدقيقة (الأرجواني) ، والحويصلات الأساسية الكثيفة (البرتقالية) ، والحويصلات الخفيفة (الخضراء) ، والشبكة الإندوبلازمية (الأصفر) ، والأكتين (الأزرق). يمكن أيضا رؤية الجزيئات الكبيرة ، مثل الريبوسومات (الحمراء) ، في الزاوية اليمنى العليا. صور الفلورسنت زائفة اللون. يتم تضمين أشرطة المقياس في (A)-(E) في الصورة. يرجى النقر هنا لعرض نسخة أكبر من هذا الشكل.
{kind=link}

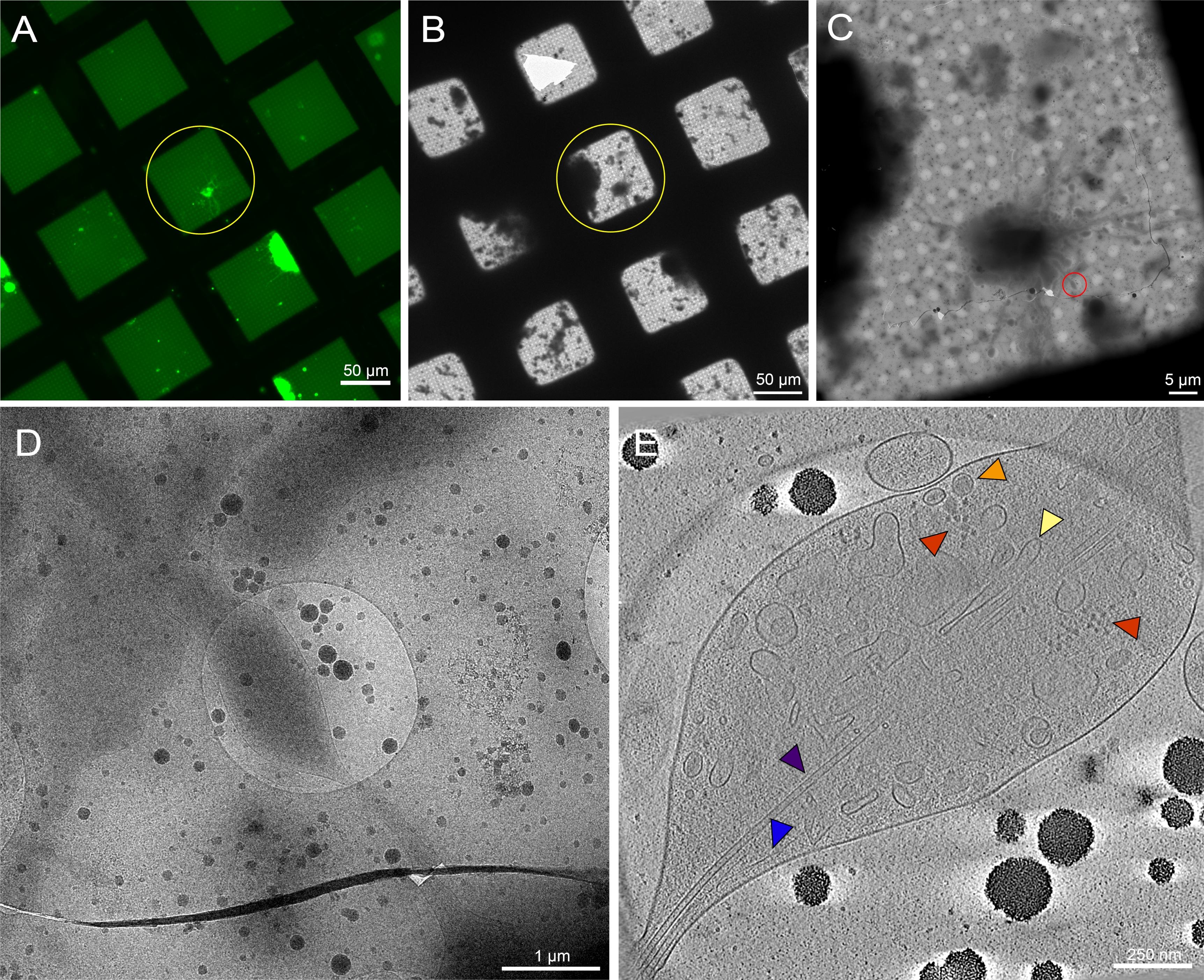
الشكل 7: الخلايا العصبية الأولية المشتقة من أدمغة يرقات ذبابة الفاكهة 3rd instar على شبكات غير نمطية. (أ) مونتاج شبكة الفحص المجهري الفلوري للخلايا العصبية لذبابة الفاكهة التي تعبر عن GFP المستهدف بالغشاء على مربعات الشبكة مع 0.5 ملغ / مل concanavalin A. الأخضر: الخلايا العصبية ذبابة الفاكهة . (ب) مونتاج شبكة Cryo-EM لنفس الشبكة في (A) بعد التجميد الغطس. تظهر الدائرة الصفراء نفس مربع الشبكة كما في (A). لاحظ وجود حطام خلوي وتلوث وسائط، مما جعل تحديد الهدف صعبا مقارنة بالشبكات المنقوشة. (ج) مونتاج الصور Cryo-EM للمربع الذي أبرزته الدوائر الصفراء في الخرائط (A) و (B). (د) صورة تكبير أعلى للمنطقة التي تحدها الدائرة الحمراء في (C)، حيث تم جمع سلسلة إمالة على عصبيات الخلية. (هاء) شريحة بسمك 25 نانومتر من التصوير المقطعي المعاد بناؤه من سلسلة الإمالة من (C) و (D). يظهر عدد من العضيات في هذا التصوير المقطعي ، مثل الأنابيب الدقيقة (الأرجواني) ، والأكتين (الأزرق) ، والشبكة الإندوبلازمية (الصفراء) ، والحويصلات الأساسية الكثيفة (البرتقالي). يمكن أيضا رؤية الجزيئات الكبيرة ، مثل الريبوسومات (الحمراء). صور الفلورسنت زائفة اللون. يتم تضمين أشرطة المقياس في (A)-(E) في الصورة. يرجى النقر هنا لعرض نسخة أكبر من هذا الشكل.
{kind=link}

الشكل 8: أمثلة على المشاكل المحتملة مع النقش. صور الفلورسنت ل ECM الموسومة المودعة على شبكات ذات أنماط دقيقة. (أ) النقش غير المتكافئ عبر الشبكة بسبب التوزيع غير المتكافئ لهلام PLPP. (ب) لا يمكن لإدارة المحتوى المؤسسي الالتصاق بالمناطق التي يغطيها استنسل PDMS أثناء النقش. (ج) نمط التدرج المشبع (الجانب الأيمن) أو النمط المقلوب (يسار) على شبكة منقوشة بجرعة إجمالية عالية جدا. (د) ECM هو التمسك المناطق على قضبان الشبكة وكذلك المنطقة المنقوشة بسبب انعكاسات الليزر فوق البنفسجي أثناء النقش. الصور زائفة اللون. يظهر نمط الإدخال في أسفل اليسار ؛ أشرطة المقياس هي 100 ميكرومتر. يرجى النقر هنا لعرض نسخة أكبر من هذا الشكل.
{kind=link}
| أصدر | السبب (الأسباب) المحتملة | استكشاف الاخطاء |
| النمط المصغر | ||
| لا يمكن رؤية الإضاءة من ليزر PRIMO | • لم يتم إعداد مسار الضوء بشكل صحيح | • تحقق من إعداد مسار ضوء المجهر بشكل صحيح |
| • ليزر PRIMO غير قيد التشغيل أو الليزر متشابك | ||
| العديد من مربعات الشبكة المكسورة | • لمس احباط الشبكة مع ملاقط أو ماصة أثناء المناولة | • مقبض على الشبكات بعناية |
| • جفت الشبكة أثناء الحضانة أو الغسيل | • لا تسمح للشبكة بأن تجف أثناء الغسيل والحضانات | |
| مناطق كبيرة غير منقوشة | • تغطية غير كافية للجل | • تأكدي من توزيع الجل بالتساوي على الشبكة أثناء الإضافة |
| • رقائق شبكية خارج نطاق التركيز البؤري أثناء النقش | • أضيفي ميكرولتر إضافي من الجل | |
| • المساحة المغطاة بالاستنسل | • تحقق من التركيز البؤري قبل نقش كل منطقة | |
| • شبكة في المنتصف بعناية في الاستنسل | ||
| نمط مشبع أو مقلوب | • جرعة غير صحيحة | • جرب مجموعة من الجرعات الإجمالية للنمط |
| • تغطية غير كافية للجل | • تأكد من تغطية الشبكة بالتساوي بالجل | |
| • جرب قيما مختلفة لأنماط التدرج الرمادي | ||
| نمط ضبابي | • ضعف التركيز أثناء النقش | • تكرار معايرة PRIMO على نفس ارتفاع العينة |
| • معايرة غير صحيحة | • ركزي على رقائق الشبكة قبل النقش | |
| • تقسيم النمط إلى مناطق إضافية للتنسيق | ||
| ECM التمسك خارج النمط | • انعكاسات من الجل أو الغبار | • تأكدي من جفاف الجل قبل النقش |
| • تأكد من نظافة الغطاء والعدسة الموضوعية | ||
| ECM غير مرئي بعد النقش | • تبييض الصور | • تقليل التعرض للضوء إلى ECM قبل التصوير |
| • جرعة غير صحيحة أثناء النقش | • جرب مجموعة من قيم الجرعة الإجمالية للنمط | |
| • عدم كفاية وقت حضانة ECM | • زيادة وقت حضانة ECM | |
| بذر الخلايا | ||
| تكتل الخلايا | • الإفراط في الهضم | • استخدام نسبة أقل من التربسين أو الوقت لإطلاق الخلايا الملتصقة |
| • كثافة خلايا عالية | • مرور و / أو هضم الخلايا عند التقاء أقل | |
| • لا تهيج الخلايا أثناء التحرير | ||
| • محلول خلية ماصة بلطف أو استخدم مصافي الخلايا | ||
| الخلايا التي لا تلتصق بالمناطق المنقوشة | • ECM غير مناسب لنوع الخلية | • جرب تركيزات وتكوين ECM مختلفة |
| • يتم تقليل صلاحية الخلايا قبل البذر | • التأكد من أن زراعة الخلايا وظروف إطلاق الخلايا لا تضر بالخلايا | |
| الخلايا لا تتوسع بعد الالتصاق | • ECM أو نمط غير مناسب لنوع الخلية | • جرب أنماطا مختلفة و ECM |
| • في بعض الحالات ، قد تعزز رقائق أكثر استمرارية (R1.2/20 مقابل R2/1) توسع الخلايا |
الجدول 1: القضايا المحتملة أثناء النمط المصغر. يصف هذا الجدول بعض المشكلات التي قد يواجهها المستخدم أثناء الأنماط الدقيقة أو زرع الخلايا. يتم توفير الأسباب المحتملة واستكشاف الأخطاء وإصلاحها لكل مشكلة. يمكن رؤية الصور التمثيلية لبعض المشاكل في الشكل 8.
Discussion
تدعم المجاهر الإلكترونية الحديثة والمتقدمة وحزم البرامج الآن جمع بيانات cryo-EM و cryo-ET المؤتمتة حيث يمكن استهداف مئات إلى آلاف المواقع وتصويرها في غضون أيام قليلة32,33,34,35. أحد العوامل المحددة الهامة لسير عمل cryo-ET كامل الخلية هو الحصول على أعداد كافية من الأهداف القابلة للتحصيل لكل شبكة. في الآونة الأخيرة ، طور عدد من المجموعات بروتوكولات لشبكات الأنماط الدقيقة ل cryo-EM ، مع ميزة واحدة تتمثل في تحسين كفاءة جمع البيانات 16،17،18. هنا يتم تقديم بروتوكول لاستخدام نظام النمط الدقيق المتاح تجاريا لشبكات TEM micropattern لإجراء دراسات cryo-ET للخلايا العصبية الأولية لذبابة الفاكهة وخطوط الخلايا البشرية المستزرعة (غير المصابة أو المصابة بفيروس RSV). نظام الأنماط الدقيقة هذا متعدد الاستخدامات ويمكن تحسين العديد من الخطوات وتخصيصها لتناسب أهدافا تجريبية محددة. يمكن للمستخدم الذي يتمتع بخبرة TEM والفحص المجهري الفلوري أن يصبح ماهرا بسرعة في إعداد الشبكة والنمط الدقيق. مع الممارسة الدقيقة ، يجب أن تكون النتائج الجيدة قابلة للتحقيق بعد بضع تكرارات. فيما يلي ، تتم مناقشة بعض الخيارات المتاحة ، واعتبارات المستخدم ، والفوائد المحتملة ، والتطبيقات المستقبلية للنمط المصغر ل cryo-EM.
أحد الاعتبارات المهمة ل cryo-ET للخلية بأكملها هو اختيار شبكة EM. تتكون شبكات EM من جزأين: إطار شبكي (أو دعم هيكلي) ورقائق معدنية (أو فيلم) ، وهو سطح الفيلم المستمر أو الهولي الذي ستنمو عليه الخلايا. تستخدم الشبكات الشبكية النحاسية بشكل شائع في cryo-EM للبروتينات والمجمعات المعزولة. ومع ذلك ، فهي غير مناسبة ل cryo-ET كامل الخلية بسبب السمية الخلوية للنحاس. بدلا من ذلك ، يتم استخدام شبكة ذهبية بشكل شائع للتصوير المقطعي الخلوي. وتشمل الخيارات الأخرى النيكل أو التيتانيوم، والتي قد توفر فوائد على الذهب مثل زيادة الصلابة16. تتوفر شبكات EM بأبعاد شبكية مختلفة لدعم مجموعة من التطبيقات. توفر أحجام الشبكات الأكبر مساحة أكبر للخلايا للنمو بين قضبان الشبكة والمزيد من المناطق القابلة لجمع سلسلة الإمالة ، على الرغم من ذلك على حساب زيادة هشاشة العينة الإجمالية. الرقائق الأكثر استخداما هي الكربون غير المتبلور المثقب أو الثقب ، مثل Quantifoils أو شبكات C-flat. يمكن تصوير الأهداف البيولوجية إما من خلال الثقوب الموجودة في الكربون أو من خلال الكربون الشفاف للإلكترون. توفر شبكات مثل R 2/1 أو R 2/2 ، حيث يبلغ عرض الثقوب 2 ميكرومتر والمتباعدة 1 و 2 ميكرومتر على التوالي ، عددا كبيرا من الثقوب وبالتالي عددا كبيرا من المناطق المحتملة لجمع البيانات. ومع ذلك ، قد تنمو بعض الخلايا وتتوسع بشكل أفضل على الأسطح الأكثر اتساقا مثل شبكات R 1.2 / 20 أو الكربون المستمر. لمعالجة العينات النهائية عن طريق طحن الحزمة الأيونية المركزة (cryo-FIB) ، تتم إزالة الرقائق من خلال الطحن ، مما يقلل من المخاوف بشأن استمرار وجود الفيلم الأساسي. كما هو الحال مع الشبكة ، تتوفر أيضا رقائق من مواد أخرى ، مع كون بروتوكول النقش المعروض هنا مناسبا بنفس القدر لشبكات SiO2 . تشمل الشبكات شائعة الاستخدام شبكات Quantifoil الذهبية أو الكربون المستمر أو SiO2 film 200-mesh (تباعد 90 ميكرومتر تقريبا بين قضبان الشبكات) للتبريد الخلوي الكامل ET.
هناك عدد من الاعتبارات عند تصميم نمط. تسترشد غالبية هذه القرارات بنوع الخلية والغرض من التجربة. نقطة البداية الجيدة هي اختيار نمط يقترب من شكل وأبعاد الخلايا في الثقافة. أظهرت العديد من الدراسات تأثيرات كبيرة لشكل النمط على نمو الخلايا وترتيب الهيكل الخلوي13,36,37. يجب توخي الحذر بشكل خاص أثناء تصميم النمط إذا كان هذا يمكن أن يغير هدف الاهتمام. تم اختبار عدة أنماط لكل نوع من أنواع الخلايا لتحديد الأنماط التي تعزز الالتصاق الخلوي والنمو. تسمح مرونة نظام الأنماط الدقيقة باختبار أنماط متعددة على شبكة واحدة وتغيير الأنماط للشبكات المختلفة في تجربة واحدة. الأنماط الأكبر (~ 50-90 ميكرومتر) ، مثل تلك المستخدمة هنا ، تزيد من احتمال التصق خلايا متعددة بمنطقة واحدة من النمط وتسمح للخلايا بالتوسع والتمدد بعد الالتصاق. قد تكون الأنماط الأكثر تقييدا (20-30 ميكرومتر) مناسبة في التجارب التي يكون فيها عزل الخلايا أكثر أهمية من توسع الخلية ، مثل تجارب طحن الحزمة الأيونية المركزة (cryo-FIB). بالنسبة لتطبيقات التصوير المقطعي ، قد يحتاج المرء إلى النظر في تأثير محور الميل. إذا تم وضع نمط بحيث تنمو جميع الخلايا بالتوازي مع بعضها البعض في اتجاه واحد ، فمن الممكن أن تكون جميع الخلايا عمودية على محور الميل عند تحميلها على مرحلة المجهر ، مما يؤدي إلى انخفاض جودة البيانات.
على الشبكات غير المتناثرة ، غالبا ما تلتصق الخلايا بشكل تفضيلي بأشرطة الشبكة ، حيث لا يمكن تصويرها بواسطة TEM. حتى على الشبكات المنقوشة ، غالبا ما يلاحظ أن الخلايا توضع في زوايا مربعات الشبكة جزئيا على كل من رقائق الكربون المنقوشة وشريط الشبكة. في الآونة الأخيرة ، تم استخدام النمط المصغر لوضع جزء من الخلية عمدا فوق شريط الشبكة18. يمكن النظر في ذلك في التجارب التي لا يكون فيها من الأهمية بمكان وجود محيط الخلية بالكامل على الرقاقة. يمكن أن يكون هذا مهما بشكل خاص للخلايا التي يمكن أن تنمو أكبر من مربع شبكة واحد ، مثل الخلايا العصبية الأولية التي تنمو على مدار عدة أيام.
هناك العديد من الأدوات التي يمكن استخدامها لتصميم نمط. هنا ، كان النمط مقتصرا على أقل من 800 بكسل في أي بعد بحيث يمكن تدوير النمط إلى أي زاوية ولا يزال مناسبا للمساحة القصوى التي يمكن أن تنقش في إسقاط واحد بواسطة نظام النمط الدقيق هذا. هذا يسمح للمستخدم بتدوير النمط ليكون موجها بشكل صحيح مع الشبكة بغض النظر عن اتجاه الشبكة على المجهر. هنا ، تم تقسيم الشبكة إلى ست مناطق نمط. في المقام الأول ، يسمح هذا بتعديل التركيز البؤري بين مناطق مختلفة من الشبكة. شبكات الذهب ، على وجه الخصوص ، مرنة للغاية وقد لا تستلقي مسطحة تماما على الزجاج. التركيز السليم ضروري للحصول على نتائج نقش نظيفة ومكررة. باستخدام الأنماط المجزأة ، يجب إجراء تعديلات طفيفة فقط على موضع النمط إذا تحولت الشبكة قليلا أثناء عملية النقش ، على الرغم من أن هذه ليست مشكلة عادة عند استخدام هلام PLPP مع استنسل PDMS. وأخيرا ، ظلت مربعات الشبكة الأربعة المركزية للشبكة غير منقوشة. هذا يدعم المستخدم أن يكون قادرا على تحديد مركز الشبكة بوضوح ، وهو أمر مفيد جدا لتجارب التصوير المرتبط.
يحتوي برنامج النقش لنظام الأنماط الدقيقة هذا ، Leonardo ، أيضا على ميزات أكثر تقدما مثل الخياطة والقدرة على استيراد الأنماط كملفات PDF ، والتي تقع خارج نطاق هذا البروتوكول. يتضمن هذا البرنامج أيضا اكتشاف البنية المجهرية وتحديد المواقع التلقائي للنمط الذي يمكن استخدامه على شبكات TEM. تكون هذه الميزة مفيدة للغاية عندما تكون الشبكة مسطحة للغاية ويمكن نقشها دون الحاجة إلى ضبط التركيز البؤري بين المناطق المختلفة.
يمكن أن يكون لاختيار بروتين ECM تأثير كبير على التصاق الخلايا وتوسعها. من المعروف أن بعض الخلايا تخضع لتغيرات فسيولوجية عندما تنمو على ركائز محددة38. تم اختبار بروتينات وتركيزات ECM متعددة لأي نوع جديد من الخلايا بناء على العمل السابق المبلغ عنه في الأدبيات. يستخدم اللامينين والفيبرينوجين والفيبرونيكتين والكولاجين على نطاق واسع للخلايا المستزرعة ويمكن استخدامه كنقطة انطلاق إذا لم تكن البيانات الأخرى متوفرة. ومع ذلك ، يجب أيضا مراعاة بروتينات ECM الأخرى إذا فشلت بروتينات ECM شائعة الاستخدام في منح خصائص الالتزام المناسبة للخلايا. كان هذا صحيحا بشكل خاص بالنسبة للخلايا العصبية الأولية لذبابة الفاكهة ، حيث كان التركيز العالي لنبات اللكتين كونكانافالين A ضروريا للالتصاق الخلوي السليم. يمكن اختبار توافق الالتصاق الخلوي والنمو مع ECM عن طريق النقش على الأطباق الزجاجية أو الشرائح قبل الانتقال إلى شبكات TEM. هذا النهج قبل الفحص فعال من حيث الوقت والتكلفة إذا كان هناك حاجة إلى فحص عدد كبير من المجموعات. يعد تضمين بروتين ECM المترافق مع الفلورسنت أمرا قيما لتقييم نجاح وجودة النقش.
يعد بذر الخلايا أحد أهم الخطوات ل cryo-ET للخلية بأكملها ، سواء مع أو بدون نمط دقيق6,16,39. بالنسبة لذبابة الفاكهة الأولية أو الخلايا العصبية الأخرى ، التي تكون هشة وغير مستقرة في التعليق ، وقد تكون محدودة الكمية ، يتم تفضيل نهج البذر الفردي على بذر الخلايا المتتابعة الخاضعة للمراقبة. تعد خطوة البذر الواحدة بكثافة الخلايا المحسنة ، كما هو موضح في بروتوكول الخلايا العصبية ذبابة الفاكهة ، خيارا قابلا للتطبيق لمعظم أنواع الخلايا. ومع ذلك ، من الممكن أيضا زرع الخلايا على الركيزة بتركيز أولي أقل وإضافة المزيد من الخلايا بطريقة مراقبة كما هو موضح هنا وفي أدبيات أخرى18. يمكن أن يوفر هذا البذر المتسلسل نتائج أكثر اتساقا في بعض الحالات. على غرار زراعة الخلايا القياسية ، يجب توخي الحذر دائما للحفاظ على صلاحية الخلية وتقليل تكتل الخلايا أثناء العزل.
عند البدء لأول مرة بالنمط المصغر ، هناك بعض المزالق المحتملة التي تضر بالنتيجة النهائية. من بين أهم الاعتبارات للنجاح التعامل الدقيق مع الشبكة والتقنية المعقمة ، والتوزيع الموحد لهلام PLPP ، والجرعة المناسبة والتركيز أثناء النقش ، والحفاظ على صلاحية الخلية قبل البذر. وجمعت في الجدول 1 قائمة ببعض المسائل والحلول المحتملة.
يمكن استخدام الشبكات ذات النمط الدقيق للمساعدة في وضع الخلايا لإنشاء كثافة خلايا متسقة عبر الشبكة ووضع المناطق ذات الأهمية في المناطق المناسبة لجمع سلسلة الإمالة16,18. يمكن استخدام موضع الخلايا وتحديد مواقعها كعلامات ائتمانية للارتباط في تجارب cryo-CLEM ، مما يقلل من الحاجة إلى شبكات البحث الهشة وعلامات الفلورسنت الائتمانية. ومع ذلك ، تجدر الإشارة إلى أن هذه العلامات الائتمانية قد لا تزال مفيدة لارتباط دقة الميكرومتر الفرعي29,40. وعلاوة على ذلك، فإن التوزيع المتساوي للخلايا المعزولة مفيد للغاية أيضا لتجارب طحن الحزم الأيونية المركزة (cryo-FIB) لزيادة عدد الخلايا التي يمكن قطع الصفيحة منها16.
وستؤدي إضافة النمط المصغر إلى سير عمل cryo-EM إلى تحسينات قابلة للقياس في إنتاجية البيانات ومن المحتمل أن تمكن من إجراء تجارب جديدة. ومع اعتماد هذه التقنية وتطويرها بشكل أكبر، فإن التطبيقات الأكثر تقدما للمرضى المجهريين بما في ذلك تدرجات ECM، وترسبات ECM المتعددة، وتجميع البنية المجهرية ستزيد من توسيع قدرات cryo-ET لدراسة الأهداف والعمليات البيولوجية في السياق الخلوي الكامل.
Disclosures
وليس لدى صاحبي البلاغ ما يكشفان عنه.
Acknowledgements
نشكر الدكتور جيل ويلدونجر والدكتور سيهوي ز. يانغ والسيدة جوزفين دبليو ميتشل في قسم الكيمياء الحيوية بجامعة ويسكونسن في ماديسون على مشاركتهم بسخاء سلالة ذبابة elav-Gal4 و UAS-CD8::GFP (مركز بلومنغتون للأوراق المالية ، # 5146). ونود أيضا أن نشكر الدكتور أوريليان دوبوين، والسيد لوران سيكوييه، والسيدة ماري شارلوت مانوس من ألفيول والسيد سيرج قدورا من مختبرات نانوسكيل على دعمهم السخي خلال هذا المشروع. تم دعم هذا العمل جزئيا من قبل جامعة ويسكونسن ، ماديسون ، قسم الكيمياء الحيوية في جامعة ويسكونسن ، ماديسون ، ومنح خدمات الصحة العامة R01 GM114561 ، R01 GM104540 ، R01 GM104540-03W1 ، و U24 GM139168 إلى E.R.W. و R01 AI150475 إلى P.W.S. من المعاهد الوطنية للصحة. تم دعم جزء من هذا البحث من خلال منحة المعاهد الوطنية للصحة U24 GM129547 وتم إجراؤه في PNCC في OHSU وتم الوصول إليه من خلال EMSL (grid.436923.9) ، وهو مرفق مستخدم تابع لمكتب العلوم التابع لوزارة الطاقة برعاية مكتب البحوث البيولوجية والبيئية. نحن ممتنون أيضا لاستخدام المرافق والأجهزة في مركز أبحاث Cryo-EM في قسم الكيمياء الحيوية في جامعة ويسكونسن ، ماديسون.
Materials
| Name | Company | Catalog Number | Comments |
| 0.1% (w/v) Poly-L-Lysine | Sigma | P8920-100ML | |
| 0.22 µm syringe filters PVDF membrane | Genesee | 25-240 | |
| 22x60-1 Glass cover slip | Fisher | 12545F | |
| 5/15 Tweezers | EMS (Dumont) | 0203-5/15-PO | |
| Antibiotic-Antimycotic (100X) | ThermoFisher (Gibco) | 15240096 | |
| BEAS-2B cells | ATCC | CRL-9609 | |
| Collagen I, bovine | ThermoFisher (Gibco) | A1064401 | |
| Concanavalin A, Alexa Fluor 350 Conjugate | ThermoFisher (Invitrogen) | C11254 | |
| DMEM | Fisher (Lonza) | BW12-604F | |
| EtOH | Fisher (Decon Labs) | 22-032-600 | |
| Fetal Bovine Serum | ATCC | 30-2020 | |
| Fibrinogen From Human Plasma, Alexa Fluor 647 Conjugate | ThermoFisher (Invitrogen) | F35200 | |
| Fibronectin Bovine Protein, Plasma | ThermoFisher (Gibco) | 33010018 | |
| Glass bottom dish | MatTek | P35G-1.5-20-C | |
| Glucose | VWR | 0643-1KG | |
| Grid prep holder | EMS | 71175-01 | |
| HeLa cells | ATCC | CCL-2 | |
| Hemacytometer | Fisher (SKC, Inc.) | 22600100 | |
| HEPES | Fisher (ACROS Organics) | AC172572500 | |
| Hoechst 33342 | ThermoFisher (Invitrogen) | H3570 | |
| Insulin | Fisher (Sigma Aldrich) | NC0520015 | |
| KCl | MP Bio | 194844 | |
| KH2PO4 | Fisher (ACROS Organics) | AC212595000 | |
| Leica-DMi8 | Leica Microsystems | Can be customized with camera, stage, and objective attachments | |
| Leonardo | Alvéole | https://www.alveolelab.com/our-products/leonardo-photopatterning-software/ | |
| Liberase Research Grade | Fisher (Supply Solutions) | 50-100-3280 | |
| LIVE/DEAD Viability/Cytotoxicity Kit | ThermoFisher (Invitrogen) | L3224 | |
| Microscope camera | Hammamatsu | C13440-20CU | |
| Motorized stage | Märzhäuser Wetzlar | 00-24-599-0000 | |
| NaCl | Fisher (Fisher BioReagents) | BP358-1 | |
| NaH2PO4 | Fisher (ACROS Organics) | AC207802500 | |
| NaOH | Fisher (Alfa Aesar) | AAA1603736 | |
| PBS | Corning | 21-040-CV | |
| PDMS stencils | nanoscaleLABS | PDMS_STENCILS_EM | https://www.alveolelab.com/our-products/pdms-stencil-multiwell-plate/ |
| PEG-SVA | nanoscaleLABS | PEG-SVA-1GR | mPEG-Succinimidyl Valerate, MW 5,000 |
| Penicillin | Fisher (Research Products International Corp) | 50-213-641 | |
| pH strips | Fisher (Millipore Sigma) | M1095350001 | pH probe can also be used |
| PLPP gel | nanoscaleLABS | PLPP-GEL-300UL | https://www.alveolelab.com/our-products/plpp-photoactivatable-reagent/ |
| PRIMO | Alvéole | https://www.alveolelab.com/our-products/primo-micropatterning/ | |
| pSynkRSV-I19F (BAC containing RSV A2-mK+ antigenomic cDNA ) | BEI Resources | NR-36460 | https://www.beiresources.org/Catalog/BEIPlasmidVectors/NR-36460.aspx |
| Quantifoil grids | EMS (Quantifoil) | Q2100AR1 | 2 µm holes spaced 1 µm apart, other dimensions are available |
| RPMI | Fisher (Lonza) | BW12-702F | |
| RSV A2-mK+ | see entry for pSynkRSV-19F | - | Described in Hotard et al. [22]. Can be generated from pSynkRSV-ll9F |
| Schneider's Media | ThermoFisher (Gibco) | 21720-024 | |
| SerialEM | SerialEM (https://bio3d.colorado.edu/SerialEM/ ) | https://bio3d.colorado.edu/SerialEM/ | |
| Straight tweezers | EMS (Dumont) | 72812-D | |
| Streptomycin | Fisher (Fisher BioReagents) | BP910-50 | |
| Sucrose | Avantor | 4097-04 | |
| Tetracycline | Sigma | T8032-10MG | |
| Titan Krios electron microscope | ThermoFisher | 300kV, with direct electron detector camera and energy filter | |
| Trypsin | ThermoFisher (Gibco) | 15090046 | |
| Tube Revolver/Rotator | Fisher (Thermo Scientific) | 11676341 | |
| UAS:mcD8:GFP Drosophila fly strain | Bloomington Drosophila Stock Center | 5146 | http://flybase.org/reports/FBtp0002652.html |
References
- Nogales, E., Scheres, S. H. Cryo-EM: A unique tool for the visualization of macromolecular complexity. Molecular Cell. 58 (4), 677-689 (2015).
- Martynowycz, M. W., Gonen, T. From electron crystallography of 2D crystals to MicroED of 3D crystals. Current Opinion in Colloid and Interface Science. 34, 9-16 (2018).
- Wagner, J., Schaffer, M., Fernandez-Busnadiego, R. Cryo-electron tomography-the cell biology that came in from the cold. FEBS Letters. 591 (17), 2520-2533 (2017).
- Wan, W., Briggs, J. A. Cryo-electron tomography and subtomogram averaging. Methods in Enzymology. 579, 329-367 (2016).
- Bäuerlein, F. J., Pastor-Pareja, J. C., Fernández-Busnadiego, R. Cryo-electron tomography of native Drosophila tissues vitrified by plunge freezing. bioRxiv. , 437159(2021).
- Hampton, C. M., et al. Correlated fluorescence microscopy and cryo-electron tomography of virus-infected or transfected mammalian cells. Nature Protocols. 12 (1), 150-167 (2017).
- Hsieh, C. E., Leith, A., Mannella, C. A., Frank, J., Marko, M. Towards high-resolution three-dimensional imaging of native mammalian tissue: Electron tomography of frozen-hydrated rat liver sections. Journal of Structural Biology. 153 (1), 1-13 (2006).
- Al-Amoudi, A., Norlen, L. P., Dubochet, J. Cryo-electron microscopy of vitreous sections of native biological cells and tissues. Journal of Structural Biolology. 148 (1), 131-135 (2004).
- Rigort, A., et al. Focused ion beam micromachining of eukaryotic cells for cryoelectron tomography. Proceedings of the National Academy of Sciences. 109 (12), 4449-4454 (2012).
- Gorelick, S., et al. PIE-scope, integrated cryo-correlative light and FIB/SEM microscopy. Elife. 8, 45919(2019).
- Wu, G. H., et al. Multi-scale 3D cryo-correlative microscopy for vitrified cells. Structure. 28 (11), 1231-1237 (2020).
- Turk, M., Baumeister, W. The promise and the challenges of cryo-electron tomography. FEBS Letters. 594 (20), 3243-3261 (2020).
- Théry, M. Micropatterning as a tool to decipher cell morphogenesis and functions. Journal of Cell Science. 123 (24), 4201-4213 (2010).
- Tseng, Q., et al. Spatial organization of the extracellular matrix regulates cell-cell junction positioning. Proceedings of the National Academy of Sciences. 109 (5), 1506-1511 (2012).
- Hardelauf, H., et al. Micropatterning neuronal networks. Analyst. 139 (13), 3256-3264 (2014).
- Toro-Nahuelpan, M., et al. Tailoring cryo-electron microscopy grids by photo-micropatterning for in-cell structural studies. Nature Methods. 17 (1), 50-54 (2020).
- Engel, L., et al. Extracellular matrix micropatterning technology for whole cell cryogenic electron microscopy studies. Journal of Micromechanics and Microengineering. 29 (11), (2019).
- Engel, L., et al. Lattice micropatterning for cryo-electron tomography studies of cell-cell contacts. bioRxiv. , 272237(2021).
- Sibert, B. S., Kim, J. Y., Yang, J. E., Wright, E. R. Whole-cell cryo-electron tomography of cultured and primary eukaryotic cells on micropatterned TEM grids. bioRxiv. , 447251(2021).
- Egger, B., van Giesen, L., Moraru, M., Sprecher, S. G. In vitro imaging of primary neural cell culture from Drosophila. Nature Protocols. 8 (5), 958-965 (2013).
- Lu, W., Lakonishok, M., Gelfand, V. I. Kinesin-1-powered microtubule sliding initiates axonal regeneration in Drosophila cultured neurons. Molecular Biology of the Cell. 26 (7), 1296-1307 (2015).
- Ke, Z., et al. The morphology and assembly of respiratory syncytial virus revealed by cryo-electron tomography. Viruses. 10 (8), (2018).
- Stobart, C. C., et al. A live RSV vaccine with engineered thermostability is immunogenic in cotton rats despite high attenuation. Nature Communications. 7, 13916(2016).
- Hotard, A. L., et al. A stabilized respiratory syncytial virus reverse genetics system amenable to recombination-mediated mutagenesis. Virology. 434 (1), 129-136 (2012).
- Schindelin, J., et al. Fiji: an open-source platform for biological-image analysis. Nature Methods. 9 (7), 676-682 (2012).
- Mastronarde, D. N. Automated electron microscope tomography using robust prediction of specimen movements. Journal of Structural Biology. 152 (1), 36-51 (2005).
- Kremer, J. R., Mastronarde, D. N., McIntosh, J. R. Computer visualization of three-dimensional image data using IMOD. Journal of Structural Biology. 116 (1), 71-76 (1996).
- Tang, G., et al. EMAN2: an extensible image processing suite for electron microscopy. Journal of Structural Biology. 157 (1), 38-46 (2007).
- Yang, J. E., Larson, M. R., Sibert, B. S., Shrum, S., Wright, E. R. CorRelator: Interactive software for real-time high precision cryo-correlative light and electron microscopy. Journal of Structural Biology. , 107709(2021).
- Ke, Z., et al. Promotion of virus assembly and organization by the measles virus matrix protein. Nature Communications. 9 (1), 1736(2018).
- Kim, J., Yang, S., Wildonger, J., Wright, E. A new in situ neuronal model for cryo-ET. Microscopy and Microanalysis. 26 (2), 130-132 (2020).
- Bouvette, J., et al. Beam image-shift accelerated data acquisition for near-atomic resolution single-particle cryo-electron tomography. Nature Communications. 12 (1), 1957(2021).
- Schorb, M., Haberbosch, I., Hagen, W. J. H., Schwab, Y., Mastronarde, D. N. Software tools for automated transmission electron microscopy. Nature Methods. 16 (6), 471-477 (2019).
- Weis, F., Hagen, W. J. H., Schorb, M., Mattei, S. Strategies for optimization of cryogenic electron tomography data acquisition. Journal of Visual Experiments. (169), e62383(2021).
- Chreifi, G., Chen, S., Jensen, G. J. Rapid tilt-series method for cryo-electron tomography: Characterizing stage behavior during FISE acquisition. Journal of Structural Biology. 213 (2), 107716(2021).
- Anderson, D. E., Hinds, M. T. Endothelial cell micropatterning: methods, effects, and applications. Annals of Biomedical Engineering. 39 (9), 2329-2345 (2011).
- McWhorter, F. Y., Wang, T., Nguyen, P., Chung, T., Liu, W. F. Modulation of macrophage phenotype by cell shape. Proceedings of the National Academy of Sciences. 110 (43), 17253-17258 (2013).
- Kleinman, H. K., Luckenbill-Edds, L., Cannon, F. W., Sephel, G. C. Use of extracellular matrix components for cell culture. Analytical Biochemistry. 166 (1), 1-13 (1987).
- Fassler, F., Zens, B., Hauschild, R., Schur, F. K. M. 3D printed cell culture grid holders for improved cellular specimen preparation in cryo-electron microscopy. Journal of Structural Biology. 212 (3), 107633(2020).
- Schellenberger, P., et al. High-precision correlative fluorescence and electron cryo microscopy using two independent alignment markers. Ultramicroscopy. 143, 41-51 (2014).
Reprints and Permissions
Request permission to reuse the text or figures of this JoVE article
Request PermissionExplore More Articles
This article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. All rights reserved