
Method Article
Tüm Hücreli Kriyo-Elektron Tomografi İş Akışlarında Doğrudan Hücre Konumlandırmasına Mikropatterning İletim Elektron Mikroskopi Izgaraları
* Bu yazarlar eşit katkıda bulunmuştur
Bu Makalede
Özet
Bu protokolün amacı, hücre yapıştırma ve büyümesini kriyo-elektron mikroskopisi için hedeflenen ızgara alanlarına yönlendirmektir. Bu, kullanıcı tarafından belirtilen desenlerde ablated bir anti-kirlenme tabakasının uygulanması ve ardından hücre tohumlamadan önce desenli alanlarda hücre dışı matris proteinlerinin birikmesi ile elde edilir.
Özet
Tüm hücreli kriyo-elektron tomografisi (kriyo-ET), hücresel bağlamda bulunan makromoleküllerin nanometre düzeyinde çözünürlük yapılarını üretmek için kullanılan ve neredeyse yerli donmuş-hidratlı bir durumda korunan güçlü bir teknolojidir. Bununla birlikte, hücreleri fizyolojik durumlarında tutarken tomografiye uygun bir şekilde TEM ızgaralarına hücrelerin kültleme ve/veya yapışması ile ilgili zorluklar vardır. Burada, TEM şebekelerinde ökaryotik hücre büyümesini yönlendirmek ve teşvik etmek için mikropatterning kullanımı hakkında ayrıntılı bir adım adım protokol sunulmaktadır. Mikropatterning sırasında hücre büyümesi, ekstra hücresel matris (ECM) proteinlerinin TEM ızgarasının folyo üzerinde belirtilen desen ve konumlar içinde birikmesiyle yönlendirilirken, diğer alanlar kirlenme önleyici bir tabaka ile kaplanmıştır. Yüzey kaplaması ve desen tasarımı seçimlerindeki esneklik, mikropatterning'i çok çeşitli hücre tipleri için geniş ölçüde uygulanabilir hale getirir. Mikropatterning, bireysel hücrelerdeki yapıların yanı sıra konak-patojen etkileşimleri veya farklılaştırılmış çok hücreli topluluklar gibi daha karmaşık deneysel sistemlerin çalışmaları için yararlıdır. Mikropatterning ayrıca korelatif ışık ve elektron mikroskopisi (kriyo-CLEM) ve odaklanmış iyon ışın frezeleme (kriyo-FIB) dahil olmak üzere birçok aşağı akış tüm hücreli kriyo-ET iş akışına entegre edilebilir.
Giriş
Kriyo-elektron mikroskopisinin (kriyo-EM) geliştirilmesi, genişlemesi ve çok yönlülüğü ile araştırmacılar, makromoleküler (~1 nm) ila yüksek (~2 Å) çözünürlüğe kadar neredeyse yerli bir durumda çok çeşitli biyolojik örnekleri incelediler. Tek parçacıklı kriyo-EM ve elektron kırınım teknikleri en iyi çözeltideki veya kristal durumdaki saflaştırılmış makromoleküllere uygulanır, sırasıyla1,2. Kriyo-elektron tomografisi (kriyo-ET), bakteriler, pleomorfik virüsler ve ökaryotik hücreler gibi büyük, heterolog nesnelerin neredeyse yerli yapısal ve ultrayapısal çalışmaları için benzersizdir3. Cryo-ET'de, numunenin mikroskop aşamasında fiziksel olarak eğilmesi ve farklı açılarda numune üzerinden bir dizi görüntü elde ederek üç boyutlu (3D) bilgiler elde edilir. Bu görüntüler veya eğim serileri genellikle bir ila üç derecelik artışlarla +60/-60 derecelik bir aralığı kapsar. Eğim serisi daha sonra hesaplamalı olarak tomogram4 olarak da bilinen bir 3D birime yeniden yapılandırılabilir.
Tüm kriyo-EM teknikleri, numunenin ince bir amorf, kristal olmayan, vitreus buz tabakasına gömülmesini gerektirir. En sık kullanılan kriyo fiksasyon tekniklerinden biri, numunenin EM ızgarasına uygulandığı, şiştiği ve sıvı etan veya sıvı etan ve propan karışımına hızla daldırıldığı dalma dondurmadır. Bu teknik, HeLa hücreleri5,6 gibi kültürlü insan hücreleri de dahil olmak üzere kalınlıkta 100 nm ila ~10 μm < örneklerin vitrifikasyonu için yeterlidir. Kalınlığı 200 μm'ye kadar olan mini organoidler veya doku biyopsileri gibi daha kalın örnekler, yüksek basınçlı dondurma ile vitrifiye edilebilir7. Bununla birlikte, daha kalın numunelerin artan elektron saçılımı nedeniyle, kriyo-ET için numune ve buz kalınlığı 300 kV iletim elektron mikroskoplarında ~ 0.5 - 1 μm ile sınırlıdır. Bu nedenle, kriyo-kesit8 veya odaklanmış iyon ışın frezeleme9,10,11 gibi ek numune hazırlama adımları kullanılmadığı sürece, birçok ökaryotik hücrenin tüm hücre kriyo-ET'si hücre çevresi veya hücrelerin uzantıları ile sınırlıdır.
Birçok tüm hücreli kriyo-ET görüntüleme deneyinin bir sınırlaması veri toplama aktarım hızıdır12. Binlerce izole parçacığın genellikle tek bir TEM ızgara karesinden görüntülenebildiği tek parçacıklı kriyo-EM'nin aksine, hücreler büyüktür, yayılmıştır ve hücrelerin ince bir vitreus buz tabakasında korunmasına izin vermek için yeterince düşük yoğunlukta yetiştirilmelidir. Genellikle ilgi alanı hücrenin belirli bir özelliği veya alt alanı ile sınırlıdır. Verimi daha da sınırlayan, hücrelerin TEM ızgara çubukları üzerinde veya yakınında olduğu gibi TEM görüntülemeye uygun olmayan alanlarda büyüme eğilimidir. TEM şebekelerindeki hücre kültürüyle ilişkili öngörülemeyen faktörler nedeniyle, veri toplama için örnek erişilebilirliği ve veri verimini iyileştirmek için teknolojik gelişmelere ihtiyaç vardır.
Yapışan ekstra hücresel matris (ECM) proteinleri ile substrat mikropatterning, cam ve diğer doku kültürü substratları gibi sert, dayanıklı ve optik olarak şeffaf yüzeylerde hücrelerin büyümesini yönlendirmek için canlı hücreli ışık mikroskopisi için iyi kurulmuş bir tekniktir13,14. Mikropatterning yumuşak ve/veya üç boyutlu (3D) yüzeylerde de gerçekleştirilmiştir. Bu tür teknikler sadece hücrelerin hassas bir şekilde konumlandırılmasına izin vermedi; ayrıca desenli sinir hücresi devreleri15 gibi çok hücreli ağların oluşturulmasını da desteklemişlerdir. Mikropatterning'i kriyo-ET'ye getirmek sadece verimi artırmakla kalmayacak, aynı zamanda karmaşık ve dinamik hücresel mikroçevranlıkları keşfetmek için yeni çalışmalar da açabilir.
Son zamanlarda, çeşitli gruplar birden fazla yaklaşımla TEM şebekelerinde mikropatterning tekniklerini kullanmaya başladı16,17. Burada, TEM ızgaraları için maskesiz bir fotopatterning tekniğinin kullanımı, yüksek çözünürlüklü ve temassız desenleme özelliğine sahip Alvéole PRIMO mikropatterning sistemi kullanılarak açıklanmaktadır. Bu mikropatterning sistemi ile, alt tabakanın üstüne bir kirlenme önleyici tabaka uygulanır, ardından bir fotokatalistin uygulanması ve UV lazer ile kullanıcı tanımlı desenlerde kirlenme önleyici tabakanın ablasyonu. ECM proteinleri daha sonra uygun hücre kültürü için desenlere eklenebilir. Bu yöntem retina pigment epitel-1 (RPE1), Madin-Darby köpek böbrek-II (MDCKII), insan foreskin fibroblast (HFF) ve endotel hücre hatları kriyo-ET çalışmaları için çeşitli gruplar tarafından kullanılmıştır16,17,18. Bu mikropatterning sistemi, birden fazla kirlenme önleyici tabaka substratın yanı sıra bir sıvı veya jel fotokatalist reaktif ile uyumludur. Çeşitli ECM proteinleri hücre hattının özgüllüğünden seçilebilir ve uyarlanabilir ve kullanıcı için çok yönlülük sağlar.
Mikropatterning laboratuvar içerisinde bir dizi projeye başarıyla uygulanmıştır19. Burada, kültürlü HeLa hücrelerini, solunum sinsitial virüsünü (RSV) enfekte BEAS-2B hücrelerini ve birincil larva Drosophila melanogaster nöronlarını incelemek için spesifik adaptasyonlar dahil olmak üzere bir mikropatterning protokolü sunulmaktadır20.
Protokol
Burada açıklanan protokol, Wright laboratuvarı ve Madison Wisconsin Üniversitesi Cryo-EM Araştırma Merkezi tarafından kullanılan hücre kültürü, mikropatterning ve görüntüleme yöntemlerinin bir derlemesidir. İş akışı Şekil 1'de sunulmaktadır. Ek eğitim ve öğretim materyalleri aşağıdaki sitelerde mevcuttur: https://cryoem.wisc.edu veya https://wrightlab.wisc.edu
1. Desenleme için ızgaraların hazırlanması
- TEM ızgaralarını temiz bir cam kaydırağa, karbon tarafı yukarıya aktarın (karbon folyonun standart kalınlığı 12 nm'dir). ACE600 adlı bir karbon evaporatör kullanarak, genel karbon filmi dayanıklılığını artırmak için ızgaralara 5-8 nm ek karbon buharlaştırın.
NOT: Bu adım SiO2 ızgaraları için gerekli değildir. Bu adım önceden de yapılabilir; kaplamalı ızgaraları vakum kurutucu gibi düşük nem ortamında saklayın. - Izgaraları bir ızgara hazırlık tutucusuna aktarın ve ızgaraları karbon tarafı yukarı doğru deşarj edin. Işıma deşarj sistemi kullanarak, 80 mm çalışma mesafesi ve 1,0 x 10-3 mbar vakum basıncı ile 10 mA'da 60 s için ızgaraları boşaltın. Bunu bir sonraki adımdan 15-30 dakika içinde yapın.
NOT: Izgara hazırlık sahipleri, küçük bir Petri kabında bir filtre kağıdı parçası ile ticari olarak satın alabilir veya ev yapımı olabilir.
2. Kirlenme önleyici tabakanın uygulanması
NOT: Izgaraları kullanırken uygun steril teknik kullanılmalı ve tüm çözeltiler steril olmalı ve/veya filtre sterilize edilmelidir.
- Izgaraları (karbon tarafı yukarı) ızgaralar arasında en az 1 cm ayırma ile temiz bir cam kaydırağa veya kapak ucuna aktarın. Pipet 10 μL% 0.05 poli-L-lizin (PLL) her ızgaraya. Izgaraları nemli kağıt havlulu kapalı plastik bir kutu gibi nemli bir odada en az 30 dakika kuluçkaya yatırın.
NOT: Bu adım bir geceye uzatılabilir. Izgaraların kurumasını önlemek için haznedeki nem seviyesinin yeterli olduğundan emin olun. - Her ızgarayı 15 μL 0,1 M HEPES pH 8,5 ile üç kez yıkayın. Her yıkama için, ızgaranın kurumasına izin vermeden sıvının çoğunu bir pipetle ızgaradan çıkarın. 15 μL taze tampon ekleyin, en az 30 sn kuluçkaya yatırın ve tekrarlayın. Son yıkamadan sonra her ızgarayı 15 μL 0,1 M HEPES'te bırakın.
NOT: Bu adımda ve gelecekteki adımlarda, ızgarayı ıslak tutmak ve pipet ile ızgara arasında temastan kaçınmak önemlidir. - Her ızgara için 0,1 M HEPES pH 8,5'te 10 μL 100 mg/mL polietilen glikol-süksinimimidyl valerate (PEG-SVA) hazırlayın. PEG-SVA, yumuşak karıştırma ile hızlı bir şekilde çözünür ve net bir çözüm elde olur.
NOT: PEG-SVA çözümünü önceden hazırlamayın. PEG-SVA, pH 8.5'te 10 dakikalık bir yarı ömrağa sahiptir. PEG-SVA stoğunu -20 °C'de kuru bir ortamda saklayarak ve açılmadan önce oda sıcaklığına ısınarak aşırı neme maruz kalmaktan kaçının. - PEG-SVA çözeltisini hazırladıktan hemen sonra, hepes pH 8.5'in 15 μL'lik damlasını her ızgaradan çıkarın (ızgarayı kurutmamaya dikkat edin) ve PEG-SVA çözeltisinin 10 μL'lik bir damlasını ekleyin. Izgaraları nemli bir odada en az 1 saat kuluçkaya yatırın.
NOT: Bu adım bir geceye uzatılabilir. Haznedeki nemin ızgaraların kurumasını önlemek için yeterli olduğundan emin olun. - Her ızgarayı 15 μL steril su ile üç kez yıkayın. Her yıkama için, ızgaranın kurumasına izin vermeden sıvının çoğunu bir pipetle ızgaradan çıkarın, 15 μL tatlı su ekleyin, en az 30 sn kuluçkaya yatırın ve tekrarlayın. Son yıkamadan sonra her ızgarayı 15 μL suda bırakın.
3. PLPP jelin uygulanması
- Her ızgara için temiz bir mikroskop kapak kapağı hazırlayın. Şebekenin kuruma olasılığını en aza indirmek için her ızgara için her ızgara için aynı anda bir ızgara olmak üzere aşağıdaki adımları tamamlayın.
- Şebekeyi kapak kapağına yerleştirmeye ve ızgarayı ıslak tutmaya yardımcı olmak için kapak çizgisinin ortasına 1,0 μL'lik bir damla su yerleştirin. Şebekeyi 15 μL su damlasından kapak üzerindeki 1,0 μL su damlasına dikkatlice aktarın. Izgara karbon tarafını yukarı yerleştirdiniz.
- Izgarayı ortalanmış tutmaya ve ızgaranın karbon folyosuyla kalıp temasını en aza indirmeye dikkat ederek ızgaranın üzerine bir polidimetilsiloksan (PDMS) kalıbı dikkatlice yerleştirin.
- Izgaranın üzerine 1,0 μL 4 benzoylbenzyl-trimetilamimonyum klorür (PLPP) jel ekleyin. Pipet hafifçe karıştırmak için (pipet ucu ile ızgaraya dokunmayın).
- Izgara ile kapak kapağını kurumak için karanlık bir konuma taşıyın. Jel yaklaşık 15-30 dakika içinde kurur.
4. Mikropattern kalibrasyonu ve tasarımı
- Cam kapakçığın bir tarafını vurgulayıcı ile renklendirin. Odaklanmayı kolaylaştırmak için ince uçlu kalıcı bir işaretleyiciden siyah çizgiler ekleyin. Kapak altlığı mikroskop üzerine renkli tarafı objektif lense baka olacak şekilde yerleştirin. Parlak alan modunu kullanarak vurgulayıcıya odaklanın.
- Mikroskobun ve mikropatterning sisteminin açık olduğundan ve doğru ışık yolunun ayarlı olduğundan emin olun. Mikroskop bilgisayarında Micromanager ve Leonardo yazılımını (Plugins > Leonardo) açın.
- Kalibre et'i seçin ve ekrandaki yönergeleri izleyin. Mikroskop odağını, slayda yansıtılan görüntünün odakta olması için ayarlayın. Maruz kalma süresinin azaltılması gerekebilir. Kalibrasyondan sonra Şimdi Desenle'yi seçin.
- Kalibrasyon verileri altında bildirilen mikrometre/piksel (μm/px) oranını programın sol üst penceresinde kaydedin (Şekil 2, alan 1). Bir desen tasarlarken mikrometre başına kullanılacak piksel sayısını belirlemek için bu oranı kullanın.
- Kalibrasyondan sonra, yazılımın mikroskoptan canlı bir parlak alan görünümüyle açık olduğundan emin olun. Hazırlanan bir ızgarayı bir kapak kapağına (bölüm 3) ızgara objektif lense bakacak şekilde sahne alanına yükleyin. Sahneyi konumlandırın ve odağı ızgaranın yazılım penceresinde görülebilecek şekilde ayarlayın.
- Izgara karelerinin ve ızgara çubuklarının boyutunu mikrometreler halinde ölçün. Yazılım, ızgarayı ölçmek için sol alt köşenin yanındaki düğme tarafından etkinleştirilen bir cetvel içerir (Şekil 2, alan 2). Örneğin, burada 200 mesh ızgara için kullanılan desenler ~87 × 87 μm ızgara karelerine ve ~36 μm ızgara çubuklarına karşılık gelir.
NOT: Yazılım, desenleri anında yeniden boyutlandırmada esneklik sunar, bu nedenle ölçümdeki küçük yanlışlıklar tolere edilebilir. - Yukarıdaki ölçümlere ve oranlara bağlı olarak, herhangi bir görüntü oluşturma yazılımıyla desenler oluşturun. 20 × hedefli minimum özellik boyutu 1,2 μm'dir. Desenler sıkıştırılmamış 8 bit .tiff dosyaları olarak kaydedilmelidir.
- Kaydederken yazılımın görüntüleri farklı bir piksel boyutuna yeniden ölçeklendirmediğine emin olun. Desen, dört ızgara karesini kapsayacak şekilde yeterli olan 800 × 800 piksellik bir kutuya sığmalıdır.
NOT: Değeri 255 (beyaz) olan pikseller en yüksek yoğunlukta (lazerin toplam dozu) desenlenecek ve sıfır (siyah) değerine sahip pikseller desenlendirilmeyecektir. Ara değere sahip pikseller yaklaşık (X/255)*toplam dozda bir dozla desenlenecektir. Şekil 3A'da gri tonlamalı desenler için 255 ve 129 piksel değerleri kullanılmıştır. Desen tasarlandıktan sonra, değiştirilmeden kaydedilebilir ve yeniden kullanılabilir.
- Kaydederken yazılımın görüntüleri farklı bir piksel boyutuna yeniden ölçeklendirmediğine emin olun. Desen, dört ızgara karesini kapsayacak şekilde yeterli olan 800 × 800 piksellik bir kutuya sığmalıdır.
5. Mikropatterning
- Kalibrasyondan sonra, yazılımın mikroskoptan canlı bir parlak alan görünümüyle açık olduğundan emin olun. Hazırlanan bir ızgarayı bir kapak kapağına (bölüm 3) ızgara objektif lense bakacak şekilde sahne alanına yükleyin. Sahneyi konumlandırın ve odağı yazılımdaki ızgarayı görecek şekilde ayarlayın.
- İlk çalıştırma için yeni bir şablon tasarlayin. Yazılımda Yatırım Getirisi Ekle'yi ( Şekil 2 alan 3 konumunda gösterilmez) seçin ve 3.000 μm daire seçin. Ekrandaki parlak alan görüntüsünü kılavuz olarak kullanarak daire yatırım getirisini ızgaranın üzerine yerleştirin. Yatırım getirisinin güvenliğini sağlamak için kilit tuşuna basın.
- Yatırım getirisini yerinde kilitledikten sonra Desen Ekle'yi seçin ( Şekil 2 alan 3'ün konumunda gösterilmez). Bölüm 4'te tasarlanan deseni seçin. Her bölgede bağımsız odaklama ve konumlandırmanın eşit olmayan ızgaraları hesaba katmasına izin vermek için ızgarayı altı bölgeye bölün. Izgaranın her köşesi için 8 × 8 ızgara kare bölgesi ve merkezin her iki tarafında 2 × 8 ızgara kare bölgesi, orta dört ızgara karesini boş bırakır (Şekil 2, orta görüntü).
- Desenin istenen toplam kopya sayısına ulaşmak için başlangıç deseninin kopyalarını oluşturmak için çoğaltma seçeneklerini (Şekil 2, alan 4) kullanın. Gerekirse, kopyalar arasındaki aralığı kılavuz kareleri arasındaki aralıkla eşleşecek şekilde ayarlayın.
- Desen için Toplam Doz'ı ayarlayın. 30 mJ/mm2 iyi bir başlangıç noktasıdır. Daha fazla ayrıntı için tartışma bölümüne bakın.
- Uzman Seçenekleri (Şekil 2, alan 4) altında, bölgenin açısını ızgara karelerinin açısıyla eşleşecek şekilde ayarlayın. Bölgeler fare kullanılarak yeniden konumlandırılabilir. Desenlerin oranı (boyutu) da ayarlanabilir. Desenler ızgara kareleriyle hizalanana kadar desenin açısını, konumunu, arasını ve oranını ayarlayarak yinelenin. Canlı parlak alan ekranında ızgaranın bölgesini değiştirmek için mikroskop aşamasını hareket ettinin.
- Bölgede yapılan değişiklikleri kaydetmek için Kilitle'ye basın.
- Bir bölgeyi kopyalamak için, soldaki Eylemler panelinde adının yanındaki Çoğalt düğmesini (Şekil 2, alan 5, iki sayfa kağıt simgesi) tıklatın. Kopyayı yeniden konumlandırmak, yeniden adlandırmak veya düzenlemek için Eylem panelinde adını tıklatın.
- İstediğiniz tüm bölgeleri doldurmak için 5.4-5.9 adımlarını gerektiği gibi yineleyin.
- Şablonun tamamı tasarlanıp konumlandıktan sonra, Şablon dosyasını yazılım içinde kaydedin (Şekil 2, alan 6, üst araç çubuğunda yukarı ok simgesi bulunan çubuk).
- Daha önce kaydedilmiş bir şablonu yüklerken (aşağı ok simgesine sahip çubuk) yatırım getirisini ızgaranın üzerine ortalayın ve Kilitle'ye basın. Açı, konum, doz ve/veya desen dosyasını değiştirmek için Eylem Paneli'ndeki her bölgeye tıklayın.
- Şablon ve desenler konumlandıktan sonra, yazılımdaki Eylem Paneli'ndeki bölgelerden biri hariç tüm bölgelerin işaretini kaldırın.
- Mikroskop aşamasını kullanarak o bölgeye gidin ve karbon folyoya odaklanın. Eylem panelinde (Şekil 2, alan 5) Göz küresi simgesine tıklayıp desen kaplamasının görüntülenmesini açar veya kapatır.
- Izgara netleme işlemine başladıktan sonra, parlak alan deklanşörünü kapatın ve yazılımın sağ alt köşesindeki Oynat simgesine basarak canlı olarak izlenebilen desenleme işlemine başlayın.
- Eylem panelinde, bir sonraki bölgenin kutusunu seçin. Parlak alan deklanşörini açın, böylece ızgara görünür hale gelir ve mikroskop aşamasını kullanarak o bölgeyi ortalar. Eylem Paneli'nde her bölge için 5.13-5.14 arası adımları yineleyin.
- Şebeke ile kapak kapağını mikroskoptan çıkarın ve hemen ızgaraya 10 μL steril fosfat tamponlu salin (PBS) pipetlayın.
- 10 dakika sonra, kalıbı cımbızla çıkarın, ardından ızgarayı 15 μL PBS ile 3x yıkayın. Son yıkamadan sonra, her ızgarayı 15 μL PBS'ye yerleştirin ve ızgaraları karanlık bir konuma taşıyın.
6. ECM proteinlerinin birikmesi
- Kültürlü hücreler için 6.2-6.5 adımlarını izleyin; primer Drosophila nöronları için 6.6-6.10 adımlarını izleyin.
- Her ızgara için en az 15 μL ECM hazırlayın. BEAS-2B hücreleri için steril PBS'de 0.01 mg/mL sığır fibronektin ve 0.01 mg/mL florofor konjuge fibrinojen son konsantrasyonu hazırlayın. HeLa hücreleri için steril PBS'de 0.01 mg/mL sığır kollajeni I ve 0.1 mg/mL florofor-konjuge fibrinojen hazırlayın.
- PBS'nin çoğunu her ızgaradan çıkarın ve 15 μL ECM uygulayın. Izgarayı oda sıcaklığında nemli bir odada en az 1 saat kuluçkaya yatırın.
NOT: Bu adım 4 °C'de geceleme süresine kadar uzatılabilir. - ECM'de inkübasyondan sonra, her ızgarayı steril PBS ile 5x yıkayın. Her yıkama için, ızgaranın kurumasına izin vermeden sıvının çoğunu bir pipetle çıkarın, 15 μL taze PBS ekleyin, en az 30 sn kuluçkaya yatırın ve tekrarlayın. Son yıkamadan sonra her ızgarayı PBS'de bırakın.
NOT: Izgaralar PBS'de 4 °C'de bir haftaya kadar saklanabilir ve kalitede herhangi bir bozulma gözlenmez. - Deseni ve karbon folyonun bozulmadan kaldığını doğrulamak için ECM'deki floroforu tespit etmek için bir floresan mikroskobu kullanın. Birkaç kırık kare genellikle tolere edilebilir.
- Birincil Drosophila nöronları için, desenli ızgaraları steril PBS içeren 30 mm'lik bir cam alt tabağa taşıyın.
- PBS'yi tabaktan epire edin ve steril bir ortamda 25 °C'de bir gecede 0,5 mg/mL florofor konjuge konanavalin A. Incubate uygulayın.
- Concanavalin A çözeltisini tabaktan çıkarın (ızgaraları kurutmadan) ve ızgaraları PBS ile 3x yıkayın. Her yıkama için, çanaktan 2 mL PBS ekleyin ve çıkarın.
- Deseni ve karbon folyonun bozulmadan kaldığını doğrulamak için ECM'deki floroforu tespit etmek için bir floresan mikroskobu kullanın. Birkaç kırık kare genellikle tolere edilebilir.
- Son yıkamadan sonra, PBS'yi cam alt tabaktan çıkarın ve 2 mL taze hazırlanmış ekleyin, steril filtreli takviye schneider's Drosophila media21, %20 ısı inaktive fetal sığır serumu (FBS), 5 μg/mL insülin, 100 μg/mL penisilin, 100 μg/mL streptomisin ve 10 μg/mL tetrasiklin içerir. Nöronlar kaplanmaya hazır olana kadar steril bir ortamda 25 °C'de kuluçkaya yatırın.
7. Tohumlamadan önce birincil Drosophila hücrelerinin hazırlanması
- 55 mm'lik bir diseksiyon kabını % 70 EtOH ile sterilize edin ve ardından yemeğe 2-3 mL steril filtreli 1× diseksiyon salin ekleyin (9,9 mM HEPES pH 7,5, 137 mM NaCl, 5.4 mM KCl, 0.17 mM NaH2PO4, 0.22 mM KH2PO4, 3.3 mM glikoz, 43.8 mM sakkaroz)21.
- Bir çift cımbız kullanarak yiyeceklerden 30-40 3.
- Larvaları 1× PBS ile tüpe yerleştirin, ardından larvaları yıkamak için 1× PBS ile ikinci tüpe aktarın.
- PBS'yi yıkamak için larvaları% 70 EtOH ile tüpe aktarın, ardından% 70 EtOH ile ikinci tüpe aktarın. Larvaları sterilize etmek için larvaları ikinci tüpte 2-3 dakika bırakın.
- Larvaları 1× diseksiyon salinli bir tüpe aktarın, ardından hemen 1× diseksiyon salin ile ikinci tüpe aktarın.
- Bireysel larvaları 1× diseksiyon salin içeren diseksiyon kabına aktarın. Bir çift tokmak ve bir diseksiyon mikroskobu ile, beyni çıkarmak için her larvayı hızla yırtın ve 1× diseksiyon salin ile üçüncü tüpe aktarın. Tüm beyinler çıkarılana kadar tekrarlayın.
- Beyinleri içeren tüpü 1 dakika boyunca 300 x g'da santrifüj edin.
- Süpernatantı atın ve 1 mL 1× diseksiyon salin ile yıkayın ve tüpü 1 dakika boyunca 300 x g'da santrifüj edin. Bu adımı bir kez daha yineleyin.
- Tüpte 200-250 μL kalana kadar süpernatant atın ve 1x diseksiyon salin içine 2,5 mg/mL Liberase 20 μL ekleyin.
- Tüpü oda sıcaklığında 1 saat boyunca bir rotatör üzerinde döndürün; Bu saat boyunca, çözeltiyi her 10 dakikada bir 25-30 kez pipetlayın. Sonunda, çözelti biraz opak olmalıdır.
- Hücreleri 5 dakika boyunca 300 × g'da santrifüj edin.
- Süpernatantı atın, ardından 1 mL takviye schneider medyası ekleyin. Pipet çözeltisini karıştırmak için 30 kez.
- Hücreleri 5 dakika boyunca 300 × g'da santrifüj edin.
- Süpernatantı atın ve 1 mL takviye schneider medyası ekleyerek hücre peletini yıkayın. Pipet çözeltisini karıştırmak için 30 kez.
- Hücreleri 5 dakika boyunca 300 × g'da santrifüj edin.
- Süpernatantı atın, ardından hücre peletini 300 μL takviye schneider'ın ortamıyla yeniden atın. Pipet çözeltisini karıştırmak için 30-40 kez.
8. BEAS-2B ve HeLa hücrelerinin kültür ve RSV enfeksiyonu
- HeLa hücrelerini ve BEAS-2B hücrelerini T75 şişelerinde 37 °C ve % 5 CO2'de sakla. Pasaj hücreleri her 3-4 günde bir yaklaşık% 80 izdiah eder. HeLa hücrelerini DMEM + %10 FBS + 1× Antibiyotik-Antimykotik olarak koruyun. BEAS-2B'yi RPMI + %10 FBS + 1× Antibiyotik-Antimycotic6,22,23'te koruyun.
- Enfekte olmayan hücrelerin tohumlaması için bölüm 9'a atlayın. BEAS-2B ve HeLa hücreleri RSV enfeksiyonuna karşı hassastır; Beas-2B hücreleri burada gösterilen RSV ile ilgili tüm deneyler için kullanılmıştır.
NOT: Tüm BSL-2 adımlarını uygun bir biyogüvenlik kabini (BSC) ve kişisel koruyucu ekipman (KKD) kullanarak kurumsal protokollere uygun olarak gerçekleştirin. - Hücrelerin RSV enfeksiyonundan önce, kuyu başına 5 × 104 hücreyi 2 mL büyüme ortamına sahip 6 kuyu plakasına (yüzey alanı ~9,6 cm2) geçirin ve bir gecede kuluçkaya yatırın.
- Trypsinize ve hücrelerin bir kuyu saymak. Trypsinize için, bir kuyudan medya aspire ve artık medya kaldırmak için Mg2 + ve Ca2 + olmadan steril PBS 2 mL ile yıkayın. 500 μL% 0.25 tripsin çözeltisi ekleyin. 5-10 dakika boyunca 37 °C'de kuluçkaya yaslanın. Yüzeyden salınıp salınmadıklarını görmek için hücreleri periyodik olarak kontrol edin. Hücreler serbest bırakıldıktan sonra, 1,5 mL kültür ortamı ekleyin.
- 100 μL tripzil hücreyi 100 μL trippan mavisi ile karıştırın. Pipet 10 μL seyreltilmiş hücre karışımı bir hemositometre içine. Hücreleri sayın ve kuyu başına hücre sayısını hesaplayın. Aşağıdaki MOI'yi hesaplamak için bu numarayı kullanın.
- 750 μL medyada kuyu başına 10 MOI elde etmek için büyüme ortamlarında RSV-A2mK +24 seyreltme hazırlayın. RSV-A2mK+'nın MOI'si, stokun floresan odak birimlerinden (FFU) titrelerinden hesaplanabilir (Örneğin: kuyu başına 1.0 × 105 hücre ve 1.0 × 108 FFU/mL RSV stoku için, viral stoku 1:75 ila 1 × 106 FFU/750 μL veya 1,33 × 106 FFU/mL) seyreltin.
- Medyayı 6 kuyulu tabaktaki hücrelerden epire edin ve her kuyuya yukarıdan 750 μL viral çözelti ekleyin.
- Plakayı oda sıcaklığında 1 saat sallayın.
- 1 saat sonra, 37 °C'ye kadar önceden ısıtılmış büyüme ortamı ile kuyu başına toplam hacmi 2 mL'ye kadar getirin ve plakayı 6 saat boyunca% 5 CO2 ile 37 °C'ye ayarlanmış bir inkübatöre yerleştirin.
- Hücreleri serbest bırakmak için deneyin ve aşağıda açıklandığı gibi tohumlamaya devam edin. Tohumlamadan sonra, ızgaraları donmadan önce 18 saat daha kuluçkaya yatırın (enfeksiyon sonrası toplam 24 saat için).
9. Mikropatterned ızgaralara hücre tohumlama
- Kültürlü hücreler için 9.2-9.8 adımlarını izleyin; primer Drosophila nöronları için 9.9-9.11'i takip edin.
- Hücreleri serbest bırakmak için trypsinize (yukarıdaki bölüm 8'deki 4. adıma bakın). Hücre toplamayı azaltmak için hücreleri %60 veya daha az izdiahta deneyin.
- 100 μL tripzil hücreyi 100 μL trippan mavisi ile karıştırın. Seyreltilmiş hücrenin pipeti 10 μL bir hemositometreye karışır ve hücreleri sayar.
- Ortamdaki hücreleri 2 × 104 hücre/mL'ye seyreltin.
- Izgaranın yerleştirilmesine yardımcı olmak ve kurumasını önlemek için 30 mm'lik cam alt kabın ortasına 1 μL ortam ekleyin. Izgarayı kapak üzerindeki PBS'den cam alt kabın ortasına aktarın. Izgale 10 μL hücre çözeltisi ekleyin.
- Parlak alan mikroskobu kullanarak, 5 dakika sonra ızgaraya hücre yapışmasını gözlemleyin. Desenlerin çoğu boş kalırsa, hücre çözeltisinin ek 10 μL damlasını ekleyin. Kuluçka sırasında ızgaraları ve hücre sokalımını 37 °C'de tutun.
- Çoğu desen işgal edilene veya birçok meşgul desen birden çok hücreye sahip olmaya başlayana kadar 9.6 adımını yineleyin. Izgarayı inkübatörde 2 saat kuluçkaya yatırın (37 °C, %5 CO2).
- Yemeği önceden ısıtılmış 2 mL ortamla doldurun ve bir gecede kuluçkaya yatırın (37 °C, %5 CO2).
- Birincil Drosophila nöronları için, ortamı ızgara içeren tabaktan çıkarın ve hücreleri yemeğe yerleştirin.
- Hücrelerin bağlanması için 30-60 dakika bekleyin, ardından 2 mL takviye schneider medyası ekleyin.
- Nöronları dalma donmadan önce en az 2-3 gün boyunca 25 °C inkübatörde kültüre edin.
10. Desenli ızgaraların görüntülenmesi ve vitrifikasyonu
- Desenli ızgarayı ve kültürlü hücreleri içeren cam alt kabı floresan mikroskobuna yerleştirin.
- Hücrelerdeki deseni ve diğer etiketlemeleri algılamak için parlak alan ve uygun floresan kanalları kullanarak ızgaranın görüntülerini alın. Hücre yoğunluğunun ve konumlandırmasının aşağı akış görüntüleme ve analizi için uygun olduğundan emin olun.
NOT: Brightfield ve floresan görüntüler FIJI yazılım paketinde işlendi25. - Kriyo-dalma dondurucu hazırlayın; dondurma cihazının türü, numune için en uygun kullanılabilirliğe, maliyete ve özelliklere bağlı olacaktır.
NOT: Primer Drosophila nöronları otomatik bir dalma-dondurucuda, BEAS-2B hücreleri ise yarı otomatik dalma-dondurucu kullanılarak hazırlanmıştır. - Eğim serisinin uygun hizalanması için numunelere altın fiducials uygulayın. Fazla ortamı çıkarmak için leke örnekleri, daha sonra numuneleri sıvı nitrojenle soğutulan sıvı etan gibi bir kriyojene daldırma-dondurma. Birincil Drosophila nöronları için, arkadan 4 sn leke. HeLa ve BEAS-2B hücreleri için, her iki taraftan 4-6 sn leke. Donmuş ızgaralar daha sonra daha fazla kullanıma kadar sıvı nitrojende saklanabilir.
- Kriyo-elektron mikroskoptaki görüntü vitrifiye hücreler, doğrudan elektron dedektörü kamerasıyla 300 kV'da çalıştırılır. Cryo-EM/cryo-ET veri toplama için SerialEM26 gibi yazılımlarla her ilgi alanı için eğim serisi koleksiyonu ayarlayın.
NOT: Birincil Drosophila nöronlarının eğim serisi, toplam 70-75 e-/Å2 doz için 4.628 şpiksel boyutuna sahip -8 μm defocus'ta -8 μm defoksta 2° artışlarla -60° ila 60 ° çift yönlü olarak doğrudan bir elektron dedektörü üzerinde toplanarak toplandı. RSV ile enfekte beas-2B tilt serisi, 4.603 şpiksel boyutuna ve toplam ~80 e-/Å2 doza sahip -5 μm defocus'ta bir enerji filtresi (20 eV yarık) ile doğrudan bir elektron dedektörü üzerinde toplandı. - Tomogramları yeniden oluşturmak için eğim serisini işleyin.
NOT: Burada sunulan tomogramlar IMOD paketi27 kullanılarak yeniden inşa edilmiştir; lowpass filtreleme EMAN2 yazılım paketi28 kullanılarak yapıldı.
Sonuçlar
Bu prosedür, tüm hücre kriyo-ET deneyleri için EM ızgaralarını desenlamak için kullanıldı. İlk hücre kültürü preparatları, mikropatterning (Şekil 1) ve görüntüleme dahil olmak üzere bu çalışmada sunulan tüm iş akışı 3-7 günü kapsamaktadır. PLL'yi ızgaraya uygulayarak ve daha sonra reaktif PEG-SVA'yı ekleerek PEG'i bağlayarak kirlenme önleyici tabakayı oluşturmak için iki aşamalı bir prosedür kullanılmıştır. Kirlenme önleyici tabaka, bir inkübasyonda PLL-g-PEG eklenerek tek bir adımda da uygulanabilir. PLPP jeli, daha az konsantre bir sıvı olarak da kullanılabilen UV mikropatterning için bir katalizördür. Jel, sıvıya kıyasla önemli ölçüde azaltılmış bir dozda desenleme sağlar, bu da çok daha hızlı desenleme ile sonuçlanır. Bu sistemle, tam bir TEM ızgarasının gerçek desenleme süresi ~ 2 dakika idi. Mikropatterning iş akışı tek başına genellikle 5-6 saate yayılır ve bireyin TEM ızgaralarında standart hücre kültürü için sekiz ızgara desenlemesine izin verir.
Mikropatterning işlemi sırasındaki adımların bir kısmı uzun kuluçka süreleri gerektirir (bkz. adımlar 2.1, 2.3, 6.4). PLL passivasyonu (2.1) veya PEG-SVA pasivasyonu (2.3) gibi bu adımlardan bazıları bir gecede inkübasyona kadar uzatılabilir. Ek olarak, ızgaralar önceden desenlenebilir ve daha sonra kullanılmak üzere ECM proteini veya PBS çözeltisinde saklanabilir. Çalışmamızda, birincil Drosophila nöronları ve BEAS-2B hücrelerinin RSV enfeksiyonu gibi hücre hazırlama ve tohumlama zamanlamasının kritik olduğu durumlarda bu seçenekler değerliydi.
Izgaralar genel biyogüvenlik düzeyi 2 (BSL-2) laboratuvar ortamında temiz aletler, steril çözeltiler kullanılarak hazırlandı ve büyüme ortamına antibiyotik/antimycotics dahil edildi6,22,29,30. Özellikle mikrobiyal kontaminasyona duyarlı numuneler için, kirlenme önleyici tabaka ve ECM bir doku kültürü davlumbazında veya başka steril bir ortamda uygulanabilir. Ek olarak, ızgara desenleme ve ECM uygulaması arasında etanolde yıkanabilir. Enfeksiyöz ajanlarla çalışıyorsanız, prosedürü uygun biyogüvenlik protokollerine uyacak şekilde uyarlamak önemlidir.
Bu iş akışı ve sunulan prosedürler (Şekil 1), HeLa hücrelerinin (Şekil 4), RSV ile enfekte beas-2B hücrelerinin (Şekil 3, Şekil 5) ve birincil Drosophila larva nöronlarının (Şekil 6, Şekil 7) optimal kriyo-ET veri toplama için desenli EM ızgaralarına tohumlanmasını sağladı.
Mikropatterned TEM ızgaralarına tohumlanan HeLa hücreleri, kalsein-AM ve ethidyum homodimer-1 bazlı hücre canlılığı tahlilleri kullanılarak floresan boyama ile belirlendiği gibi canlı kalır (Şekil 4A,B). Kollajen ve fibrinojenden oluşan karışık bir ECM kullanan HeLa hücreleri, ızgaradaki desenlere kolayca yapışır (Şekil 4A,C). Desen boyunca genişleyen hücrelerin genel morfolojisi, bölünmemiş ızgaralarda yetişen hücrelerinkine benzer (Şekil 4C,D). HeLa hücreleri durumunda, toplam hücre kalınlığı ~ < 10 μm ve önemli ölçüde daha ince alanlar ~ hücre çevresinin yakınında 1 μm kalınlığında < kalır (Şekil 4E, F).
RSV çalışmaları için, kenarlarında düşük dozda maruz kalma ve merkeze doğru daha yüksek doz deseni ile bir degrade kullanarak tüm ızgara karelerini desenledik (Şekil 3A). Degrade desenleri, hücrelerin çevresinin yakınında bulunan serbest bırakılmış virüsleri ararken daha iyi sonuçlar verdi. Bu desenlerle, hücrelerin tercihen daha yüksek ECM konsantrasyonuna yapıştıkları, ancak aynı zamanda daha düşük ECM konsantrasyonlarına yapışabildikleri ve büyüyebildikleri bulunmuştur. Birden fazla doz gerektiren desenler kullanırken alanlar arasındaki göreli dozun optimize edilmesi gerekecektir. Dozlar ve dolayısıyla ECM konsantrasyonları birbirine çok benzer veya çok farklıysa, birden fazla doz kullanmanın etkisi kaybolacaktır.
Şekil 3'te, bir TEM ızgarası desenlendi ve daha sonra RSV ile enfekte BEAS-2B hücreleri ile tohumlandı ve kriyo-EM veri toplama için kullanıldı. Şekil 4A, gradyan deseni kullanılarak BIR TEM ızgarasına desenli ECM'nin floresan görüntüsüdür. Desenin orta bölgesi boyunca hücre yapışma ve büyümesi Şekil 3B'de tohumlamadan 18 saat sonra hücrelerin parlak bir görüntüsü olarak görülebilir. Şekil 3C'de, RSV-A2mK+'nın replikasyonundan gelen floresan sinyal (kırmızı), ECM'den gelen sinyalle kaplanır. Enfekte hücrelerin çoğunluğu gradyan deseninin daha yüksek yoğunluklu merkezi bölgesi boyunca konumlandırılmıştır. Şebeke sonrası kriyo fiksasyonunun düşük maglı bir TEM haritası, ızgara karelerinin merkezine yakın karbon folyoya yerleştirilmiş RSV ile enfekte olmuş hücreler de dahil olmak üzere bir dizi hücreyi ortaya koymaktadır. Standart TEM ızgaralarında yetişen hücreler için daha önce gösterildiği gibi22, tilt serisi, mikropatterned ızgaralarda yetişen enfekte BEAS-2B hücrelerinin çevresine yakın bir yerde RSV virionlarının bulunduğu ve toplandığı (Şekil 5A,B). RSV yapısal proteinlerinin çoğu, nükleocapsid (N) ve viral füzyon proteini (F) (Sırasıyla Şekil 5C, mavi ve kırmızı oklar) dahil olmak üzere tomogramlarda tanımlanabilir.
Birincil Drosophila nöron çalışmaları için, yazılım tarafından sunulan çözünürlük sınırına yakın olan dar desenin (desenin kalınlığının 2 μm olduğu) bir ila birkaç hücrenin bir ızgara karesi içinde izole edilmesine izin verdiği bulunmuştur (Şekil 6). Nöronal soma, nötrallerini desen içinde birkaç günlük bir süre boyunca genişletebildi. Bu, neurites'in, yazılmamış ızgaralarda kültürlenen nöronlara kıyasla kolay tanımlanmasına ve eğim serisi edinimine izin verdi (Şekil 7). Ayrıca in vitro Drosophila nöronal kültürleri20,21 için ECM olarak kullanılan bir kürsü olan floresan etiketli concanavalin A'nın desenleme için uygun olduğu bulunmuştur.
Üçüncü instar larvalardan drosophila nöronları daha önce yayınlanan protokollere göre izole edildi20,21,31. Nöronal preparatlar, hücre yerleşimini, yayılmasını ve organizasyonuni düzenlemek için concanavalin A'nın desen üzerine biriktiği mikropatterned kriyo-EM ızgaralarına uygulandı. Desenli veya girilmemiş ızgaralardaki nöronların en az 48-72 saat kuluçkaya yatmasına izin verildi ve ızgaralar daha sonra donduruldu. Şekil 6A'da, desenli bölgelere dağılmış birkaç Drosophila nöronlu mikropatterned EM ızgarasının temsili bir görüntüsü gösterilmiştir. Membranda pan-nöronal GFP ekspresyasyonu olan transgenik bir sinek suşundan türetilen bu nöronlar, sadece floresan etiketlemesi nedeniyle değil, aynı zamanda mikropatternler içindeki konumu nedeniyle de ışık mikroskopisi ile kolayca takip edilebilir. Kırılmamış ızgaralarda kültürlenen nöronlar GFP sinyali ile ışık mikroskopisi (Şekil 7A, sarı daire) ile de izlenebiliyorken, hücresel döküntülerin varlığı ve medyadan kaynaklanan kirlenme nedeniyle kriyo-EM'de yer almak önemli ölçüde zorlaştı (Şekil 7B, sarı daire). Bu tür bir varlık, muhtemelen hücre kalıntılarını yaparak geri püskürten desenli olmayan bölgelerin kirlenme önleyici tabakasındaki PEG nedeniyle, desenli ızgaralardaki nöronlar için azaltıldı. Nöron hücre gövdesinin boyutları ve genişletilmiş neurites (Şekil 6A,B, sarı daire) nedeniyle, hücrelerin daha ince bölgeleri boyunca kriyo-ET eğim serisi toplanmıştır (Şekil 6C,D, kırmızı daire). Nöronal hücre zarı, mitokondrion (siyan), mikrotübüller (mor), aktisin filamentler (mavi), veziküler yapılar (turuncu ve yeşil) ve ribozomlar (kırmızı) gibi makromoleküller, 3D tomogramdan daha yüksek büyütme görüntü montajlarında ve dilimlerinde iyi çözüldü (Şekil 6E). Benzer alt hücresel özellikler, 3D tomogramlardan (Şekil 7E) görülebilirken, veri toplama için uygulanabilir hücresel hedefleri bulmadaki zorluk önemli ölçüde azaldı.
Şekil 8'de, bu sorunlardan bazılarını içeren ızgaralardan gelen temsili görüntüler, tanımlama ve sorun gidermelerine yardımcı olmak için bir araya getirildi. En uygun koşullar belirlendikten sonra, mikropatterning, hücrelerin kriyo-TEM için ızgaralar üzerinde konumlandırılması için güvenilir ve tekrarlanabilir bir yöntemdir.

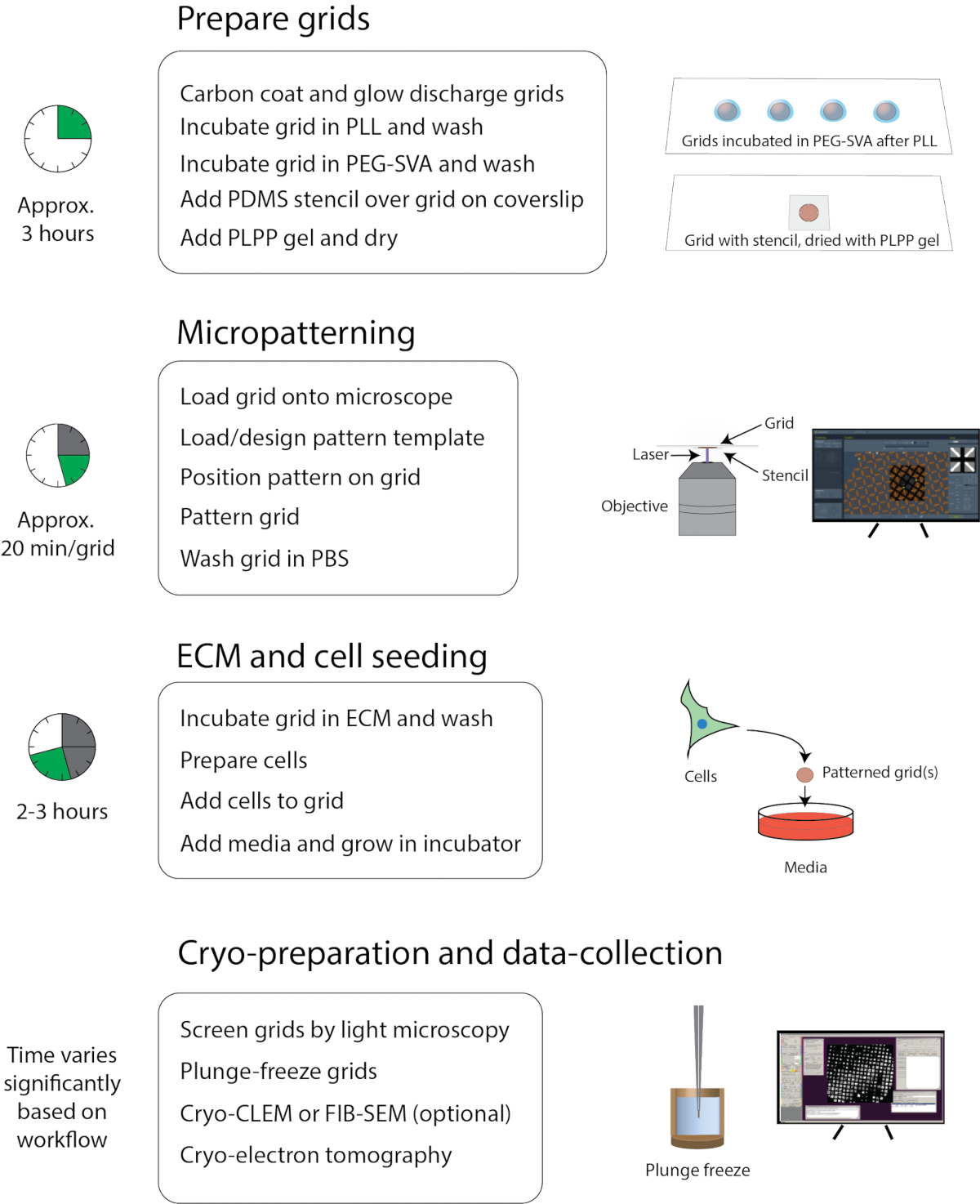
Şekil 1: Kriyo-EM için mikropatterning genel iş akışı. İş akışı kabaca dört bölüme ayrılabilir: Izgara hazırlama, mikropatterning, ECM ve hücre tohumlama ve kriyo hazırlama ve veri toplama. Her bölümün ana adımları başlıkların altında listelenir ve her bölümün tamamlanması için yaklaşık süre sola gösterilir. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.
{kind=link}

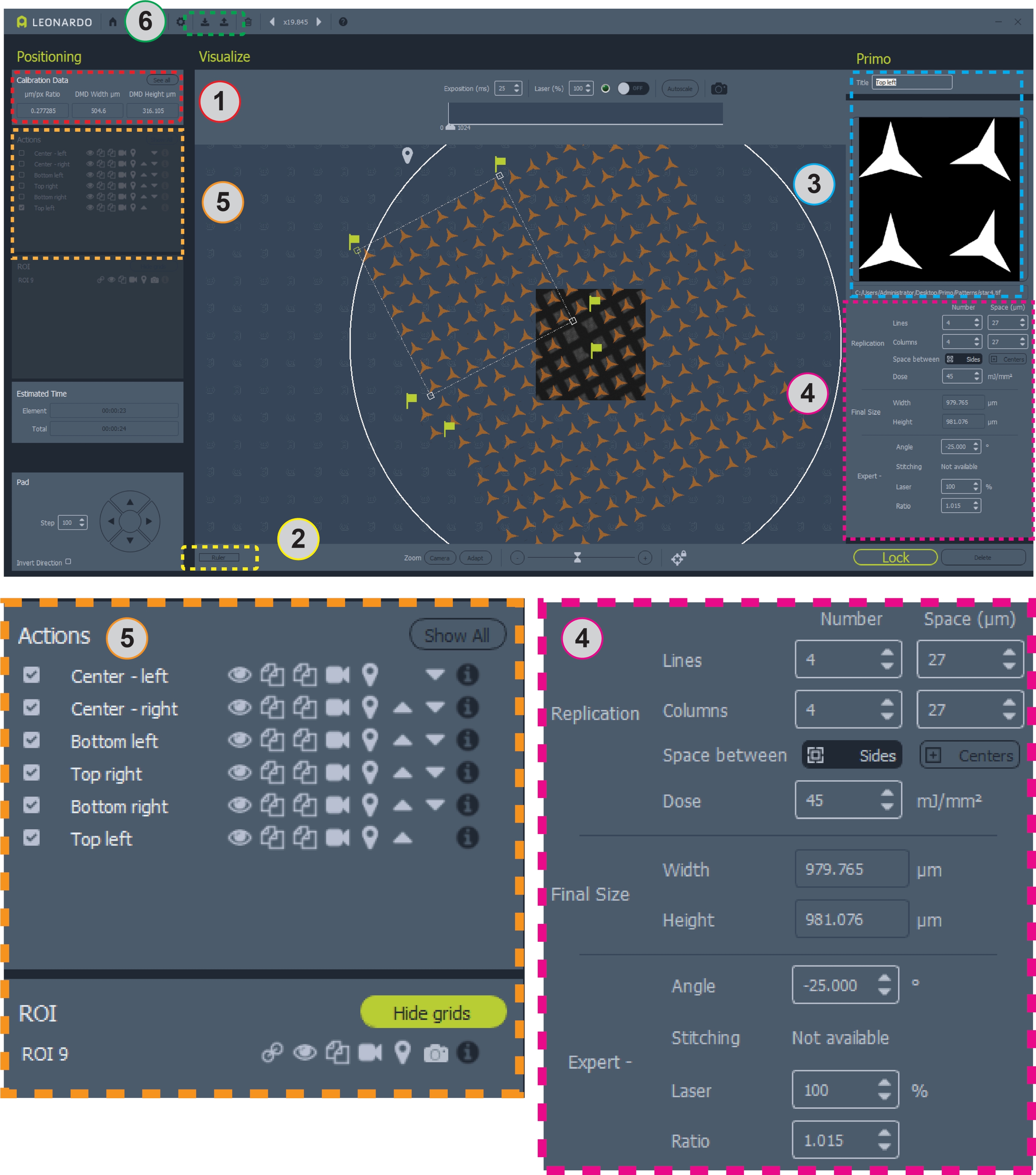
Şekil 2: Yazılımın ızgaraya yerleştirilmiş desenli ekran görüntüsü. Alan 1, desen tasarımı için μm/pix oranını içerir. Alan 2, bir ızgarayı ölçmek için cetveldir. Alan 3, desenlerin ve ROI'lerin ekleneceği veya değiştirildiği yerdir. Alan 4, desen konumlandırma ve doz için tüm bilgileri içerir. Alan 5, kaplamaları değiştir, desenleri kopyalama veya silme ve mikropatterning için desenleri seçme gibi desenler için seçenekler içerir. Alan 6, şablonların kaydedilebileceği ve yüklenebileceği yerdir. Netlik için 4 ve 5 alanlarının daha geniş görünümleri aşağıda gösterilmiştir. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.
{kind=link}

Şekil 3: Desenli kriyo-TEM ızgarasında RSV ile enfekte beas-2B hücreleri. (A) Floresan etiketli ECM ilave edildikten sonra desenli ızgaranın floresan görüntüsü. Giriş deseni sol alt köşede gösterilir. (B) A. (C) Izgara üzerinde yetişen BEAS-2B hücrelerinin brightfield görüntüsü, görüntünün A (siyan) ve B (gri) olarak, dalma dondurmadan hemen önce RSV ile enfekte olmuş hücrelerin (kırmızı) floresan görüntüsüyle birleştirilmesi; enfekte hücreler mKate-2'ye ifade eder. Ölçek çubukları 500 μm'dir. (D) Dalma dondurduktan sonra B'deki ızgaranın düşük büyütmeli kriyo-TEM haritasıdır. Floresan görüntüler sahte renklerdedir. Ölçek çubukları 500 μm'dir. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.
{kind=link}

Şekil 4: Desenli ve düzenlenmemiş hücrelerin canlı/ölü lekelenme. (A) Desenli bir ızgara üzerinde yetişen ve kalcein-AM (canlı hücre lekesi, yeşil) ve ethidyum homodimer-1 (ölü hücre lekesi, kırmızı) ile boyanmış HeLa hücrelerinin floresan görüntüsü. (B) HeLa hücreleri, 0,01 mg/mL kollajen ve fibrinojen 647 ECM (kırmızı) ile desenli quantifoil R2/2 ızgarası üzerinde bir HeLa hücresinin konfokal z-yığınlarının A. (C) Projeksiyonu'nda olduğu gibi boyanmış ve lekelenmiştir. Hücre calcein-AM (yeşil) ve Hoechst-33342 (mavi) ile boyandı. (D) 0.01 mg/mL kollajen ve fibrinojen 647 ECM ile inkübe edilmiş, calcein-AM ve Hoecsht-33342 ile inkübe edilmiş ve lekelenmiş, bölünmemiş ızgaradaki HeLa hücreleri. Floresan görüntüler iletilen ışık (gri tonlama) ile birleştirildi. (E) C.'nin X,Z projeksiyonu (F) X,Z D. Görüntülerin projeksiyonu sahtedir. (A) ve (B) ölçek çubukları 500 μm'dir; (C) - (F) ölçek çubukları 10 μm'dir . Bu şeklin daha büyük bir versiyonunu görüntülemek için lütfen buraya tıklayın.
{kind=link}

Şekil 5: Desenli kriyo-TEM ızgarasında RSV ile enfekte beas-2B hücresinin cryo-ET'si. (A) RSV enfekte BEAS-2B hücresinin kriyo-EM ızgara kare haritası. Yaklaşık hücre sınırı kesikli yeşil çizgi ile gösterilir. (B) (A) kırmızı kutulu alanın daha yüksek çözünürlüklü görüntüsü. Yaklaşık hücre sınırı kesikli yeşil çizgi ile gösterilir. RSV virionları hücre çevresinin yakınında görülebilir (beyaz ok ve sarı kutu). (C) (B) bölgesindeki sarı kutunun alanında toplanan tomogramdan tek z dilimi. Kırmızı oklar RSV F füzyon proteinini, mavi oklar ribonikleoprotein (RNP) kompleksini işaret eder. (A)-(C) ölçü çubukları görüntüye gömülür. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.
{kind=link}

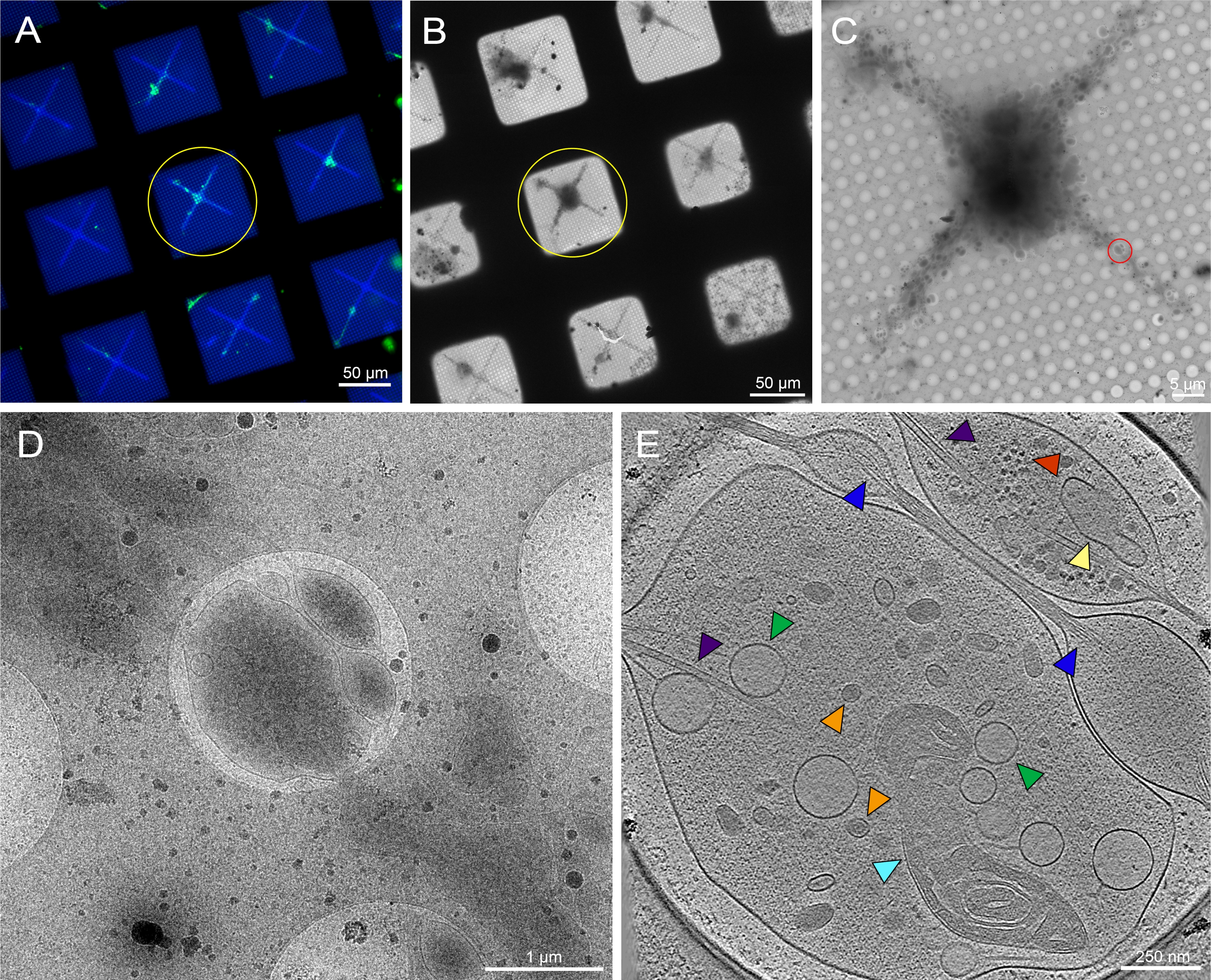
Şekil 6: Desenli kriyo-TEM ızgarasındaki 3. instar Drosophila melanogaster larvalarının beyinlerinden elde edilen birincil nöronlar. (A) 0,5 mg/mL floresan concanavalin A. Green ile desenli ızgara karelerinde membran hedefli GFP'yi ifade eden Drosophila nöronlarının canlı hücreli floresan mikroskopi ızgara montajı: Drosophila nöronları. Mavi: Fotopattern. (B) Kriyo muhafazası sonrası (A) içinde ızgaranın kriyo-EM görüntü montajı. Sarı daire, (A) ile aynı ızgara karesini notlar. (C) (A) ve (B) sarı daire ile vurgulanan karenin kriyo-EM görüntü montajı. (D) Hücrenin neurites üzerinde bir eğim serisi toplanan (C) kırmızı daire ile sınırlanmış alanın daha yüksek büyütme görüntüsü. E. 25 nm kalınlığında tomogram dilimi, (C) içinde kırmızı daireden elde edilen eğim serisinden yeniden inşa edildi. Bu tomogramda mitokondri (siyan), mikrotübüller (mor), yoğun çekirdek veziklinler (turuncu), açık veziklinler (yeşil), endoplazmik retikülü (sarı) ve aktin (mavi) gibi çeşitli organeller görülebilir. Ribozomlar (kırmızı) gibi makromoleküller sağ üst köşede de görülebilir. Floresan görüntüler sahte renklerdedir. (A)-(E) ölçü çubukları görüntüye gömülür. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.
{kind=link}

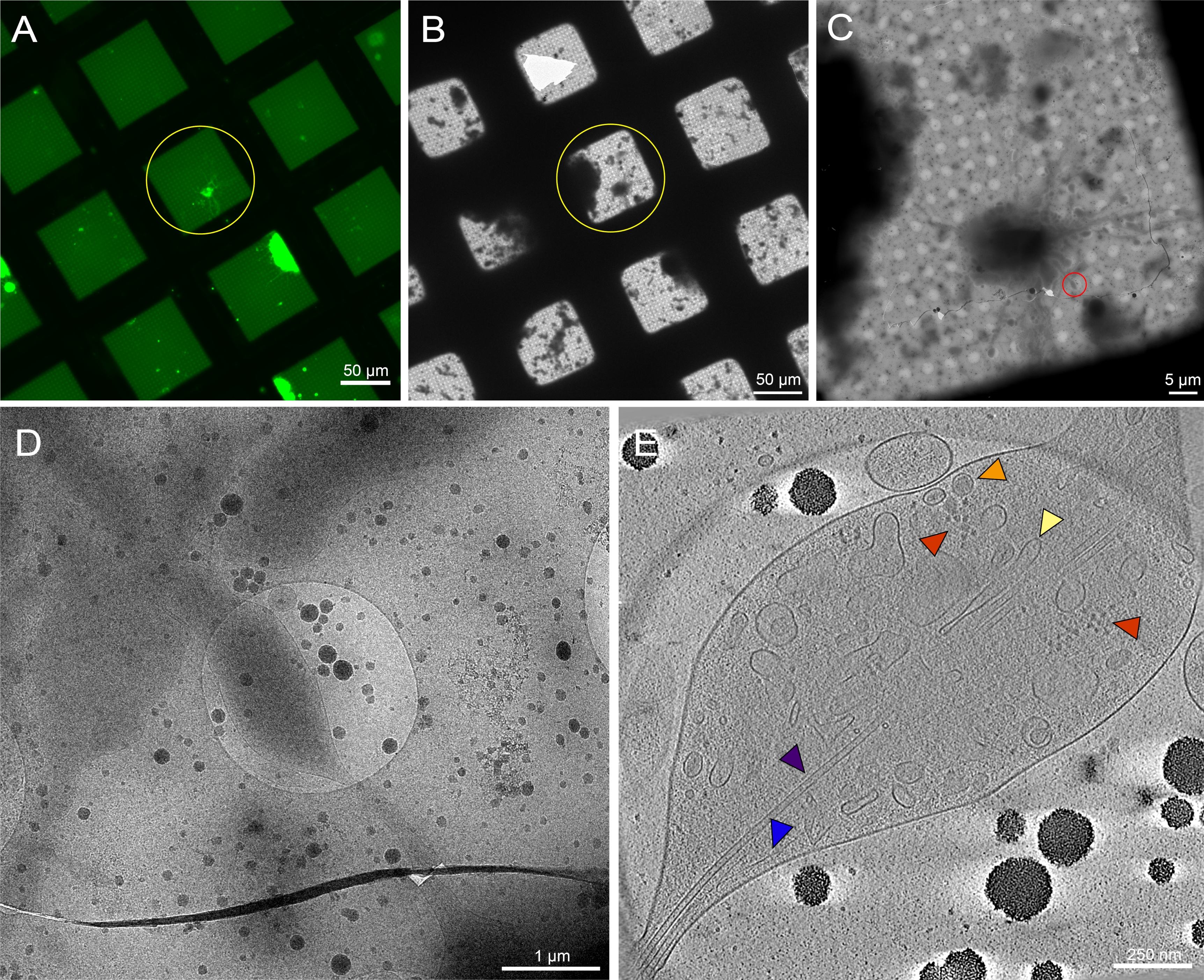
Şekil 7: 3. instar Drosophila melanogaster larvalarının beyinlerinden elde edilen primer nöronlar, unpatterned ızgaralarda. (A) 0,5 mg/mL concanavalin A. Green ile ızgara karelerinde membran hedefli GFP'yi ifade eden Drosophila nöronlarının canlı hücreli floresan mikroskopi ızgara montajı: Drosophila nöronları. (B) Dalma dondurduktan sonra (A) içinde aynı ızgaranın kriyo-EM ızgara montajı. Sarı daire (A) ile aynı ızgara karesini gösterir. Desenli ızgaralara kıyasla hedef tanımlamayı zorlaştıran hücresel enkaz ve ortam kirlenmesinin varlığına dikkat edin. (C) (A) ve (B) haritalarındaki sarı daireler tarafından vurgulanan karenin kriyo-EM görüntü montajı. (D) Hücrenin neurites üzerinde bir eğim serisi toplanan (C) kırmızı daire ile sınırlanmış alanın daha yüksek büyütme görüntüsü. (E) (C) ve (D) eğim serisinden yeniden yapılandırılmış tomogramın 25 nm kalınlığında dilimi. Bu tomogramda mikrotübüller (mor), aktin (mavi), endoplazmik retikül (sarı) ve yoğun çekirdek veziklinler (turuncu) gibi bir dizi organel görülebilir. Ribozomlar (kırmızı) gibi makromoleküller de görülebilir. Floresan görüntüler sahte renklerdedir. (A)-(E) ölçü çubukları görüntüye gömülür. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.
{kind=link}

Şekil 8: Desenleme ile ilgili olası sorunlara örnekler. Mikropatterned ızgaralara yatırılan etiketli ECM'nin floresan görüntüleri. (A) PLPP jelinin düzensiz dağılımı nedeniyle ızgara boyunca düzensiz desenleme. (B) ECM, desenleme sırasında PDMS kalıbını kaplayan alanlara yapışamaz. (C) Doymuş degrade deseni (sağ taraf) veya ters desen (sol) çok yüksek toplam dozda desenli bir ızgara üzerinde. (D) ECM, desenleme sırasında UV lazerin yansımaları nedeniyle ızgara çubuklarındaki alanların yanı sıra desenli alana da bağlı kalmaktadır. Görüntüler sahte renklendirilir; giriş deseni sol altta gösterilir; ölçek çubukları 100 μm'dir . Bu şeklin daha büyük bir versiyonunu görüntülemek için lütfen buraya tıklayın.
{kind=link}
| Sorun | Olası nedenler | Sorun giderme |
| Mikropatterning | ||
| PRIMO lazerden aydınlatma görülemiyor | • Işık yolu doğru kurulmama | • Mikroskop ışık yolunun düzgün ayarlı olduğundan emin olun |
| • PRIMO lazer açık değildir veya lazer birbirine kenetlenmiştir | ||
| Birçok kırık ızgara karesi | • Kullanım yaparken ızgara folyosuna cımbız veya pipetle dokunmak | • Izgaraları özenle ele alın |
| • Kuluçka veya yıkama sırasında ızgara kurudu | • Yıkamalar ve kuluçkalar sırasında ızgaranın kurumasına izin vermeyin | |
| Büyük, unpatterned alanlar | • Yetersiz jel kapsama alanı | • Ekleme yaparken jelin ızgaraya eşit şekilde yayıldığını sağlayın |
| • Desenleme sırasında odak dışı ızgara folyo | • Ek bir jel mikroliter ekleyin | |
| •Şablonla kaplı alan | • Her bölgeyi desenlemeden önce odağı kontrol edin | |
| • Izgarayı kalıpta dikkatlice ortala | ||
| Doymuş veya ters desen | • Yanlış doz | • Desen için bir dizi toplam doz deneyin |
| • Yetersiz jel kapsama alanı | • Izgaranın jel ile eşit şekilde kaplandığını sağlayın | |
| • Gri tonlamalı desenler için farklı değerler deneyin | ||
| Bulanık desen | • Desenleme sırasında zayıf odaklama | • PRIMO kalibrasyonunu numuneyle aynı yükseklikte tekrarlayın |
| • Yanlış kalibrasyon | • Desenlemeden önce ızgara folyosuna odaklanın | |
| • Deseni desenleme için ek bölgelere bölün | ||
| Desenin dışına yapışan ECM | • Jel veya tozdan yansımalar | • Desenlemeden önce jelin kuru olduğundan emin olun |
| • Kapak ve objektif lensin temiz olduğundan emin olun | ||
| Desenlemeden sonra ECM görünmüyor | • Fotoğraf ağartma | • Görüntülemeden önce ECM'ye ışık maruziyeti en aza indirin |
| • Desenleme sırasında yanlış doz | • Desen için bir dizi toplam doz değeri deneyin | |
| • Yetersiz ECM kuluçka süresi | • ECM için kuluçka süresini artırın | |
| Hücre tohumlama | ||
| Hücreler topaklama | • Aşırı sindirim | • Yapışan hücrelerin salınması için daha düşük tripsin yüzdesi veya zaman kullanın |
| • Yüksek hücre yoğunluğu | • Daha düşük izdiahta hücrelerin geçişi ve/veya sindirilmesi | |
| • Serbest bırakma sırasında hücreleri ajite etmeyin | ||
| • Hücre sokunu hafifçe pipetle veya hücre süzgeçlerini kullanın | ||
| Desenli alanlara bağlı olmayan hücreler | • ECM hücre tipi için uygun değildir | • Farklı ECM konsantrasyonlarını ve bileşimini deneyin |
| • Tohumlamadan önce hücrelerin canlılığı azalır | • Hücre kültürünün ve hücre salınım koşullarının hücrelere zarar vermediklarından emin olun | |
| Yapışıklık sonrası hücreler genişlemiyor | • Hücre tipi için uygun olmayan ECM veya desen | • Farklı desenler ve ECM deneyin |
| • Bazı durumlarda daha sürekli bir folyo (R1.2/20 vs R2/1) hücre genişlemesini teşvik edebilir |
Tablo 1: Mikropatterning sırasında olası sorunlar. Bu tabloda, bir kullanıcının mikropatterning veya hücre tohumlama sırasında karşılaşabileceği bazı sorunlar açıklanmaktadır. Her sorun için olası nedenler ve sorun giderme sağlanır. Şekil 8'de bazı sorunların temsili görüntüleri görülebilir.
Tartışmalar
Modern, gelişmiş elektron mikroskopları ve yazılım paketleri artık yüzlerce ila binlerce pozisyonun birkaç gün içinde hedeflenebildiği ve görüntülenebildiği kolaylaştırılmış otomatik kriyo-EM ve kriyo-ET veri toplamayı destekliyor32,33,34,35. Tüm hücreli kriyo-ET iş akışları için önemli bir sınırlayıcı faktör, ızgara başına yeterli sayıda toplanabilir hedef elde etmektir. Son zamanlarda, bir dizi grup kriyo-EM için mikropatterning ızgaraları için protokoller geliştirdi ve bir avantajı gelişmiş veri toplama verimliliği16,17,18 oldu. Burada, birincil Drosophila nöronlarının ve kültürlü insan hücre hatlarının (enfekte olmayan veya RSV enfekte) kriyo-ET çalışmaları için TEM ızgaralarını mikropattern etmek için piyasada bulunan bir mikropatterning sistemi kullanmak için bir protokol sunulmaktadır. Bu mikropatterning sistemi çok yönlüdür ve birçok adım belirli deneysel hedeflere uyacak şekilde optimize edilebilir ve uyarlanabilir. TEM ve floresan mikroskopi deneyimine sahip bir kullanıcı, şebeke hazırlama ve mikropatterning konusunda hızla yetenekli hale gelebilir. Dikkatli bir uygulama ile, birkaç yinelemeden sonra iyi sonuçlar elde edilmelidir. Aşağıda, mevcut seçeneklerden bazıları, kullanıcı konuları, potansiyel faydalar ve kriyo-EM için mikropatterning'in gelecekteki uygulamaları tartışılmıştır.
Tüm hücre kriyo-ET için önemli hususlardan biri EM ızgara seçimidir. EM ızgaraları iki bölümden oluşur: bir örgü çerçeve (veya yapısal destek) ve hücrelerin büyüyeceği sürekli veya delikli film yüzeyi olan folyo (veya film). Bakır örgü ızgaraları genellikle proteinlerin ve izole komplekslerin kriyo-EM'si için kullanılır. Bununla birlikte, bakırın sitotoksisitesi nedeniyle tüm hücreli kriyo-ET için uygun değildirler. Bunun yerine, hücresel tomografi için genellikle altın bir ağ kullanılır. Diğer seçenekler arasında nikel veya titanyum bulunur, bu da artan sertlik gibi altına göre fayda sağlayabilir16. EM ızgaraları, çeşitli uygulamaları desteklemek için farklı ağ boyutlarıyla mevcuttur. Daha büyük ağ boyutları, hücrelerin ızgara çubukları ve eğim serisi toplanmasına uygun daha fazla alan arasında büyümesi için daha fazla alan sağlar, ancak genel numune kırılganlığının artması pahasına. En sık kullanılan folyo, Quantifoils veya C-flat ızgaralar gibi delikli veya delikli amorf karbondur. Biyolojik hedefler karbondaki deliklerden veya elektron yarı saydam karbondan görüntülenebilir. Deliklerin sırasıyla 1 ve 2 μm aralıklı 2 μm genişliğinde olduğu R 2/1 veya R 2/2 gibi ızgaralar, çok sayıda delik ve dolayısıyla veri toplama için çok sayıda potansiyel alan sağlar. Bununla birlikte, bazı hücreler R 1.2/20 ızgaraları veya sürekli karbon gibi daha düzgün yüzeylerde daha iyi büyüyebilir ve genişleyebilir. Odaklanmış iyon ışın frezeleme (kriyo-FIB) ile aşağı akış numune işleme için folyo frezeleme yoluyla çıkarılır ve alttaki filmin sürekli varlığına ilişkin endişeleri azaltır. Örgüde olduğu gibi, burada sunulan desenleme protokolü SiO2 ızgaraları için eşit derecede uygun olan diğer malzemelerden folyolar da mevcuttur. Yaygın olarak kullanılan ızgaralar arasında altın Quantifoil, sürekli karbon veya sio2 filmi 200 ağ ızgaraları (ızgara çubukları arasında ~90 μm aralık) bulunur.
Bir desen tasarlarken dikkat edilmesi gereken birkaç husus vardır. Bu kararların çoğu, deneyin hücre türüne ve amacına göre yönlendirilir. İyi bir başlangıç noktası, kültürdeki hücrelerin şeklini ve boyutlarını yaklaşık olarak gösteren bir desen seçmektir. Birçok çalışma, desen şeklinin hücre büyümesi ve sitoskeletal düzenleme üzerinde önemli etkileri olduğunu göstermiştir13,36,37. Bu ilgi hedefini değiştirebilecekse, desen tasarımı sırasında özel dikkat gösterilmelidir. Hangi desenlerin hücresel yapıştırma ve büyümeyi teşvik ettiğini belirlemek için her hücre tipi için çeşitli desenler test edilmiştir. Mikropatterning sisteminin esnekliği, tek bir ızgarada birden fazla desenin testine ve tek bir deneyde farklı ızgaralar için kalıpların değiştirilmesine izin sağlar. Burada kullanılanlar gibi daha büyük desenler (~50-90 μm), birden fazla hücrenin desenin tek bir bölgesine yapışma olasılığını arttırır ve hücrelerin yapışma sonrasında genişlemesine ve genişlemesine izin verir. Hücre yalıtımı, odaklanmış iyon ışın frezeleme (kriyo-FIB) deneyleri gibi hücre genişlemesinden daha kritik olduğu deneylerde daha kısıtlı desenler (20-30 μm) uygun olabilir. Tomografi uygulamaları için eğim ekseninin etkisini göz önünde bulundurmanız gerekebilir. Bir desen, tüm hücrelerin birbirine paralel olarak tek bir yönde büyüyecek şekilde konumlandırılırsa, mikroskop aşamasına yüklendiğinde tüm hücrelerin eğim eksenine dik olması mümkündür, bu da daha düşük bir veri kalitesine neden olur.
Bölünmemiş ızgaralarda, hücreler genellikle TERCIHEN TEM tarafından görüntülenemeyecekleri ızgara çubuklarına yapışırlar. Desenli ızgaralarda bile, hücrelerin genellikle ızgara karelerinin köşelerine kısmen hem desenli karbon folyo hem de ızgara çubuğuna yerlendiği gözlenir. Son zamanlarda, hücrenin bir kısmını kasıtlı olarak ızgara çubuğu üzerinde konumlandırmak için mikropatterning kullanıldı18. Bu, tüm hücre çevresinin folyo üzerinde olmasının kritik olmadığı deneyler için düşünülebilir. Bu, özellikle birden fazla gün içinde büyüyen birincil nöronlar gibi tek bir ızgara karesinden daha büyük büyüyebilen hücreler için önemli olabilir.
Bir desen tasarlamak için kullanılabilecek birçok araç vardır. Burada, desen herhangi bir boyutta 800 pikselden daha az bir pikselle sınırlıydı, böylece desen herhangi bir açıya döndürülebilir ve yine de bu mikropatterning sistemi tarafından tek bir projeksiyonda desenlenebilen maksimum alana sığabilir. Bu, kullanıcının mikroskop üzerindeki ızgaranın yönüne bakılmaksızın deseni kılavuzla düzgün bir şekilde yönlendirilecek şekilde döndürmesini sağlar. Burada ızgara altı desen alanına ayrılmıştır. Öncelikle, bu, ızgaranın farklı bölgeleri arasında odak ayarlamasına izin verir. Özellikle altın ızgaraları çok dövülebilir ve camın üzerine tamamen düz uzanmayabilir. Temiz, rafine desenleme sonuçları için uygun odaklama gereklidir. Parçalı desenler kullanılarak, ızgara desenleme işlemi sırasında hafifçe kayarsa, desen konumunda yalnızca küçük ayarlamalar yapılması gerekir, ancak PLPP jelini PDMS kalıplarıyla kullanırken bu genellikle bir sorun değildir. Son olarak, ızgaranın merkezi dört ızgara karesi kayıtsız kaldı. Bu, bir kullanıcının korelatif görüntüleme deneyleri için çok yararlı olan ızgaranın merkezini net bir şekilde tanımlayabilmesini destekler.
Bu mikropatterning sistemi için desenleme yazılımı Leonardo, aynı zamanda dikiş ve bu protokolün kapsamı dışında olan PDF olarak desenleri içe aktarma yeteneği gibi daha gelişmiş özelliklere sahiptir. Bu yazılım ayrıca TEM ızgaralarında kullanılabilecek mikroyapı algılama ve otomatik desen konumlandırma içerir. Bu özellik en çok ızgara çok düz olduğunda kullanışlıdır ve farklı alanlar arasında odağı ayarlamaya gerek kalmadan desenlenebilir.
Bir ECM proteininin seçimi hücre yapıştırma ve genişleme üzerinde önemli bir etkiye sahip olabilir. Bazı hücrelerin belirli substratlarda yetiştirildiğinde fizyolojik değişikliklere uğradığı bilinmektedir38. Literatürde bildirilen önceki çalışmalara dayanarak herhangi bir yeni hücre tipi için birden fazla ECM proteini ve konsantrasyonu test edildi. Laminin, fibrinojen, fibronektin ve kollajen kültürlü hücreler için yaygın olarak kullanılır ve diğer veriler mevcut değilse başlangıç noktası olarak kullanılabilir. Bununla birlikte, yaygın olarak kullanılan ECM proteinleri hücreler için uygun yapışma özelliklerini sağlayamazsa, diğer ECM proteinleri de göz önünde bulundurulmalıdır. Bu özellikle primer Drosophila nöronları için geçerliydi, çünkü uygun hücresel yapışma için A bitki lektin konanavalininin yüksek konsantrasyonu gerekliydi. Hücresel yapışıklık ve büyümenin ECM ile uyumluluğu, TEM ızgaralarına geçmeden önce cam tabaklarda veya kaydıraklarda desenlenerek test edilebilir. Bu ön eleme yaklaşımı, çok sayıda kombinasyonun incelenmesi gerekiyorsa zaman ve uygun maliyetlidir. Floresan konjuge ecm proteininin dahil edilmesi, desenlemenin başarısını ve kalitesini değerlendirmek için değerlidir.
Hücre tohumlama, mikropatterning olsun veya olmasın tüm hücre kriyo-ET için en önemli adımlardan biridir6,16,39. Kırılgan, süspansiyonda kararsız ve miktarı sınırlı olabilen primer Drosophila veya diğer nöronlar için, izlenen, sıralı hücre tohumlama yerine tek tohumlama yaklaşımları tercih edilir. Drosophila nöronları için protokolde açıklandığı gibi, optimize edilmiş bir hücre yoğunluğunda tek bir tohumlama adımı, çoğu hücre tipi için uygun bir seçenektir. Bununla birlikte, hücreleri daha düşük bir başlangıç konsantrasyonunda alt tabakaya tohumlamak ve burada ve diğer literatürde açıklandığı gibi izlenen bir şekilde daha fazla hücre eklemek de mümkündür18. Bu sıralı tohumlama, bazı durumlarda daha tutarlı sonuçlar sağlayabilir. Standart hücre kültürüne benzer şekilde, izolasyon sırasında hücre canlılığını korumaya ve hücre topaklanmasını en aza indirmeye her zaman özen verilmelidir.
Mikropatterning ile ilk başladığımızda, nihai sonuca zararlı olan birkaç potansiyel tuzak vardır. Dikkatli ızgara kullanımı ve steril teknik, PLPP jelinin düzgün dağılımı, desenleme sırasında uygun doz ve odaklama ve tohumlamadan önce hücre canlılığının korunması başarı için en önemli hususlar arasındadır. Olası sorunların ve çözümlerin bir listesi Tablo 1'de toplanmıştır.
Mikropatterned ızgaralar, hücrelerin ızgara boyunca tutarlı bir hücre yoğunluğu oluşturmasına ve eğim serisi toplama için uygun alanlarda ilgi alanlarını konumlandırmasına yardımcı olmak için kullanılabilir16,18. Hücrelerin yerleştirilmesi ve konumlandırılması, kriyo-CLEM deneylerinde korelasyon için fidüsiyal belirteçler olarak kullanılabilir ve kırılgan bulucu ızgaralara ve floresan fidüsiyal belirteçlere olan ihtiyacı azaltır. Bununla birlikte, bu tür fidüsiyal belirteçlerin mikrometre altı doğruluk korelasyonu için hala yararlı olabileceği belirtilmelidir29,40. Ayrıca, izole edilmiş hücrelerin eşit bir dağılımı, lamellerin kesilebileceği hücre sayısını en üst düzeye çıkarmak için odaklanmış iyon ışın frezeleme (kriyo-FIB) deneyleri için de son derece faydalıdır16.
Cryo-EM iş akışlarına mikropatterning eklenmesi, veri veriminde ölçülebilir iyileştirmelere neden olacak ve potansiyel olarak yeni denemelere olanak sağlayacaktır. Teknik daha da benimsendikçe ve geliştirildikçe, ECM gradyanları, çoklu ECM çökeltileri ve mikroyapı montajı dahil olmak üzere daha gelişmiş mikropatterning uygulamaları, kriyo-ET'nin biyolojik hedefleri ve süreçleri tam hücresel bağlamda inceleme yeteneklerini daha da genişletecektir.
Açıklamalar
Yazarların açıklayacak bir şeyi yok.
Teşekkürler
Wisconsin Üniversitesi, Madison Biyokimya Bölümü'nden Dr. Jill Wildonger, Dr. Sihui Z. Yang ve Bayan Josephine W. Mitchell'a elav-Gal4, UAS-CD8::GFP sinek türünü cömertçe paylaştıkları için teşekkür ederiz (Bloomington stok merkezi, #5146). Ayrıca Alvéole'den Dr. Aurélien Duboin, Bay Laurent Siquier ve Bayan Marie-Charlotte Manus'a ve Nanoscale Labs'den Bay Serge Kaddoura'ya bu projedeki cömert destekleri için teşekkür ederiz. Bu çalışma kısmen Wisconsin Üniversitesi tarafından desteklendi. Madison, Wisconsin Üniversitesi Biyokimya Bölümü, Madison ve halk sağlığı servisi R01 GM114561, R01 GM104540, R01 GM104540-03W1 ve U24 GM139168'i E.R.W. ve R01 AI150475'i NIH'den P.W.S.'ye verir. Bu araştırmanın bir kısmı NIH hibe U24 GM129547 tarafından desteklendi ve OHSU'daki PNCC'de gerçekleştirildi ve Biyolojik ve Çevresel Araştırma Ofisi tarafından desteklenen bir DOE Bilim Kullanıcı Tesisi Ofisi olan EMSL (grid.436923.9) aracılığıyla erişildi. Ayrıca Madison Wisconsin Üniversitesi Biyokimya Bölümü'ndeki Cryo-EM Araştırma Merkezi'ndeki tesislerin ve enstrümantasyonun kullanımı için minnettarız.
Malzemeler
| Name | Company | Catalog Number | Comments |
| 0.1% (w/v) Poly-L-Lysine | Sigma | P8920-100ML | |
| 0.22 µm syringe filters PVDF membrane | Genesee | 25-240 | |
| 22x60-1 Glass cover slip | Fisher | 12545F | |
| 5/15 Tweezers | EMS (Dumont) | 0203-5/15-PO | |
| Antibiotic-Antimycotic (100X) | ThermoFisher (Gibco) | 15240096 | |
| BEAS-2B cells | ATCC | CRL-9609 | |
| Collagen I, bovine | ThermoFisher (Gibco) | A1064401 | |
| Concanavalin A, Alexa Fluor 350 Conjugate | ThermoFisher (Invitrogen) | C11254 | |
| DMEM | Fisher (Lonza) | BW12-604F | |
| EtOH | Fisher (Decon Labs) | 22-032-600 | |
| Fetal Bovine Serum | ATCC | 30-2020 | |
| Fibrinogen From Human Plasma, Alexa Fluor 647 Conjugate | ThermoFisher (Invitrogen) | F35200 | |
| Fibronectin Bovine Protein, Plasma | ThermoFisher (Gibco) | 33010018 | |
| Glass bottom dish | MatTek | P35G-1.5-20-C | |
| Glucose | VWR | 0643-1KG | |
| Grid prep holder | EMS | 71175-01 | |
| HeLa cells | ATCC | CCL-2 | |
| Hemacytometer | Fisher (SKC, Inc.) | 22600100 | |
| HEPES | Fisher (ACROS Organics) | AC172572500 | |
| Hoechst 33342 | ThermoFisher (Invitrogen) | H3570 | |
| Insulin | Fisher (Sigma Aldrich) | NC0520015 | |
| KCl | MP Bio | 194844 | |
| KH2PO4 | Fisher (ACROS Organics) | AC212595000 | |
| Leica-DMi8 | Leica Microsystems | Can be customized with camera, stage, and objective attachments | |
| Leonardo | Alvéole | https://www.alveolelab.com/our-products/leonardo-photopatterning-software/ | |
| Liberase Research Grade | Fisher (Supply Solutions) | 50-100-3280 | |
| LIVE/DEAD Viability/Cytotoxicity Kit | ThermoFisher (Invitrogen) | L3224 | |
| Microscope camera | Hammamatsu | C13440-20CU | |
| Motorized stage | Märzhäuser Wetzlar | 00-24-599-0000 | |
| NaCl | Fisher (Fisher BioReagents) | BP358-1 | |
| NaH2PO4 | Fisher (ACROS Organics) | AC207802500 | |
| NaOH | Fisher (Alfa Aesar) | AAA1603736 | |
| PBS | Corning | 21-040-CV | |
| PDMS stencils | nanoscaleLABS | PDMS_STENCILS_EM | https://www.alveolelab.com/our-products/pdms-stencil-multiwell-plate/ |
| PEG-SVA | nanoscaleLABS | PEG-SVA-1GR | mPEG-Succinimidyl Valerate, MW 5,000 |
| Penicillin | Fisher (Research Products International Corp) | 50-213-641 | |
| pH strips | Fisher (Millipore Sigma) | M1095350001 | pH probe can also be used |
| PLPP gel | nanoscaleLABS | PLPP-GEL-300UL | https://www.alveolelab.com/our-products/plpp-photoactivatable-reagent/ |
| PRIMO | Alvéole | https://www.alveolelab.com/our-products/primo-micropatterning/ | |
| pSynkRSV-I19F (BAC containing RSV A2-mK+ antigenomic cDNA ) | BEI Resources | NR-36460 | https://www.beiresources.org/Catalog/BEIPlasmidVectors/NR-36460.aspx |
| Quantifoil grids | EMS (Quantifoil) | Q2100AR1 | 2 µm holes spaced 1 µm apart, other dimensions are available |
| RPMI | Fisher (Lonza) | BW12-702F | |
| RSV A2-mK+ | see entry for pSynkRSV-19F | - | Described in Hotard et al. [22]. Can be generated from pSynkRSV-ll9F |
| Schneider's Media | ThermoFisher (Gibco) | 21720-024 | |
| SerialEM | SerialEM (https://bio3d.colorado.edu/SerialEM/ ) | https://bio3d.colorado.edu/SerialEM/ | |
| Straight tweezers | EMS (Dumont) | 72812-D | |
| Streptomycin | Fisher (Fisher BioReagents) | BP910-50 | |
| Sucrose | Avantor | 4097-04 | |
| Tetracycline | Sigma | T8032-10MG | |
| Titan Krios electron microscope | ThermoFisher | 300kV, with direct electron detector camera and energy filter | |
| Trypsin | ThermoFisher (Gibco) | 15090046 | |
| Tube Revolver/Rotator | Fisher (Thermo Scientific) | 11676341 | |
| UAS:mcD8:GFP Drosophila fly strain | Bloomington Drosophila Stock Center | 5146 | http://flybase.org/reports/FBtp0002652.html |
Referanslar
- Nogales, E., Scheres, S. H. Cryo-EM: A unique tool for the visualization of macromolecular complexity. Molecular Cell. 58 (4), 677-689 (2015).
- Martynowycz, M. W., Gonen, T. From electron crystallography of 2D crystals to MicroED of 3D crystals. Current Opinion in Colloid and Interface Science. 34, 9-16 (2018).
- Wagner, J., Schaffer, M., Fernandez-Busnadiego, R. Cryo-electron tomography-the cell biology that came in from the cold. FEBS Letters. 591 (17), 2520-2533 (2017).
- Wan, W., Briggs, J. A. Cryo-electron tomography and subtomogram averaging. Methods in Enzymology. 579, 329-367 (2016).
- Bäuerlein, F. J., Pastor-Pareja, J. C., Fernández-Busnadiego, R. Cryo-electron tomography of native Drosophila tissues vitrified by plunge freezing. bioRxiv. , 437159(2021).
- Hampton, C. M., et al. Correlated fluorescence microscopy and cryo-electron tomography of virus-infected or transfected mammalian cells. Nature Protocols. 12 (1), 150-167 (2017).
- Hsieh, C. E., Leith, A., Mannella, C. A., Frank, J., Marko, M. Towards high-resolution three-dimensional imaging of native mammalian tissue: Electron tomography of frozen-hydrated rat liver sections. Journal of Structural Biology. 153 (1), 1-13 (2006).
- Al-Amoudi, A., Norlen, L. P., Dubochet, J. Cryo-electron microscopy of vitreous sections of native biological cells and tissues. Journal of Structural Biolology. 148 (1), 131-135 (2004).
- Rigort, A., et al. Focused ion beam micromachining of eukaryotic cells for cryoelectron tomography. Proceedings of the National Academy of Sciences. 109 (12), 4449-4454 (2012).
- Gorelick, S., et al. PIE-scope, integrated cryo-correlative light and FIB/SEM microscopy. Elife. 8, 45919(2019).
- Wu, G. H., et al. Multi-scale 3D cryo-correlative microscopy for vitrified cells. Structure. 28 (11), 1231-1237 (2020).
- Turk, M., Baumeister, W. The promise and the challenges of cryo-electron tomography. FEBS Letters. 594 (20), 3243-3261 (2020).
- Théry, M. Micropatterning as a tool to decipher cell morphogenesis and functions. Journal of Cell Science. 123 (24), 4201-4213 (2010).
- Tseng, Q., et al. Spatial organization of the extracellular matrix regulates cell-cell junction positioning. Proceedings of the National Academy of Sciences. 109 (5), 1506-1511 (2012).
- Hardelauf, H., et al. Micropatterning neuronal networks. Analyst. 139 (13), 3256-3264 (2014).
- Toro-Nahuelpan, M., et al. Tailoring cryo-electron microscopy grids by photo-micropatterning for in-cell structural studies. Nature Methods. 17 (1), 50-54 (2020).
- Engel, L., et al. Extracellular matrix micropatterning technology for whole cell cryogenic electron microscopy studies. Journal of Micromechanics and Microengineering. 29 (11), (2019).
- Engel, L., et al. Lattice micropatterning for cryo-electron tomography studies of cell-cell contacts. bioRxiv. , 272237(2021).
- Sibert, B. S., Kim, J. Y., Yang, J. E., Wright, E. R. Whole-cell cryo-electron tomography of cultured and primary eukaryotic cells on micropatterned TEM grids. bioRxiv. , 447251(2021).
- Egger, B., van Giesen, L., Moraru, M., Sprecher, S. G. In vitro imaging of primary neural cell culture from Drosophila. Nature Protocols. 8 (5), 958-965 (2013).
- Lu, W., Lakonishok, M., Gelfand, V. I. Kinesin-1-powered microtubule sliding initiates axonal regeneration in Drosophila cultured neurons. Molecular Biology of the Cell. 26 (7), 1296-1307 (2015).
- Ke, Z., et al. The morphology and assembly of respiratory syncytial virus revealed by cryo-electron tomography. Viruses. 10 (8), (2018).
- Stobart, C. C., et al. A live RSV vaccine with engineered thermostability is immunogenic in cotton rats despite high attenuation. Nature Communications. 7, 13916(2016).
- Hotard, A. L., et al. A stabilized respiratory syncytial virus reverse genetics system amenable to recombination-mediated mutagenesis. Virology. 434 (1), 129-136 (2012).
- Schindelin, J., et al. Fiji: an open-source platform for biological-image analysis. Nature Methods. 9 (7), 676-682 (2012).
- Mastronarde, D. N. Automated electron microscope tomography using robust prediction of specimen movements. Journal of Structural Biology. 152 (1), 36-51 (2005).
- Kremer, J. R., Mastronarde, D. N., McIntosh, J. R. Computer visualization of three-dimensional image data using IMOD. Journal of Structural Biology. 116 (1), 71-76 (1996).
- Tang, G., et al. EMAN2: an extensible image processing suite for electron microscopy. Journal of Structural Biology. 157 (1), 38-46 (2007).
- Yang, J. E., Larson, M. R., Sibert, B. S., Shrum, S., Wright, E. R. CorRelator: Interactive software for real-time high precision cryo-correlative light and electron microscopy. Journal of Structural Biology. , 107709(2021).
- Ke, Z., et al. Promotion of virus assembly and organization by the measles virus matrix protein. Nature Communications. 9 (1), 1736(2018).
- Kim, J., Yang, S., Wildonger, J., Wright, E. A new in situ neuronal model for cryo-ET. Microscopy and Microanalysis. 26 (2), 130-132 (2020).
- Bouvette, J., et al. Beam image-shift accelerated data acquisition for near-atomic resolution single-particle cryo-electron tomography. Nature Communications. 12 (1), 1957(2021).
- Schorb, M., Haberbosch, I., Hagen, W. J. H., Schwab, Y., Mastronarde, D. N. Software tools for automated transmission electron microscopy. Nature Methods. 16 (6), 471-477 (2019).
- Weis, F., Hagen, W. J. H., Schorb, M., Mattei, S. Strategies for optimization of cryogenic electron tomography data acquisition. Journal of Visual Experiments. (169), e62383(2021).
- Chreifi, G., Chen, S., Jensen, G. J. Rapid tilt-series method for cryo-electron tomography: Characterizing stage behavior during FISE acquisition. Journal of Structural Biology. 213 (2), 107716(2021).
- Anderson, D. E., Hinds, M. T. Endothelial cell micropatterning: methods, effects, and applications. Annals of Biomedical Engineering. 39 (9), 2329-2345 (2011).
- McWhorter, F. Y., Wang, T., Nguyen, P., Chung, T., Liu, W. F. Modulation of macrophage phenotype by cell shape. Proceedings of the National Academy of Sciences. 110 (43), 17253-17258 (2013).
- Kleinman, H. K., Luckenbill-Edds, L., Cannon, F. W., Sephel, G. C. Use of extracellular matrix components for cell culture. Analytical Biochemistry. 166 (1), 1-13 (1987).
- Fassler, F., Zens, B., Hauschild, R., Schur, F. K. M. 3D printed cell culture grid holders for improved cellular specimen preparation in cryo-electron microscopy. Journal of Structural Biology. 212 (3), 107633(2020).
- Schellenberger, P., et al. High-precision correlative fluorescence and electron cryo microscopy using two independent alignment markers. Ultramicroscopy. 143, 41-51 (2014).
Yeniden Basımlar ve İzinler
Bu JoVE makalesinin metnini veya resimlerini yeniden kullanma izni talebi
Izin talebiThis article has been published
Video Coming Soon
JoVE Hakkında
Telif Hakkı © 2020 MyJove Corporation. Tüm hakları saklıdır