
Method Article
Micropatterning Transmissão Electron Microscopy Grids para Posicionamento Direto celular dentro de fluxos de trabalho de tomografia crio-elétron inteira
* Estes autores contribuíram igualmente
Neste Artigo
Resumo
O objetivo deste protocolo é direcionar a adesão celular e o crescimento para áreas direcionadas de redes para microscopia crio-elétron. Isso é conseguido aplicando uma camada anti-incrustada que é ablada em padrões especificados pelo usuário seguido pela deposição de proteínas de matriz extracelular nas áreas padronizadas antes da semeadura celular.
Resumo
A tomografia crio-elétron de células inteiras (crio-ET) é uma tecnologia poderosa que é usada para produzir estruturas de resolução de nível de nanômetros de macromoléculas presentes no contexto celular e preservadas em um estado quase nativo congelado-hidratado. No entanto, há desafios associados à cultura e/ou adesão de células nas grades TEM de uma maneira adequada para a tomografia, mantendo as células em seu estado fisiológico. Aqui, um protocolo passo-a-passo detalhado é apresentado sobre o uso de micropatterning para direcionar e promover o crescimento de células eucarióticas nas redes TEM. Durante a micropattersagem, o crescimento celular é direcionado depositando proteínas de matriz extracelular (ECM) dentro de padrões e posições especificados na folha da grade TEM, enquanto as outras áreas permanecem revestidas com uma camada anti-incrustar. A flexibilidade na escolha do revestimento de superfície e do design de padrões torna a micropatterning amplamente aplicável para uma ampla gama de tipos de células. A micropattersão é útil para estudos de estruturas dentro de células individuais, bem como sistemas experimentais mais complexos, como interações hospedeiro-patógeno ou comunidades multicelulares diferenciadas. A micropattersagem também pode ser integrada em muitos fluxos de trabalho crio-ET de células a jusante, incluindo a microscopia de luz correlativa e elétron (crio-CLEM) e a fresagem de feixe de íons focal (crio-FIB).
Introdução
Com o desenvolvimento, expansão e versatilidade da microscopia crio-elétron (crio-EM), pesquisadores examinaram uma ampla gama de amostras biológicas em um estado quase nativo, de resolução macromolecular (~1 nm) a alta (~2 Å). As técnicas de crio-EM de partículas únicas e difração eletrônica são melhor aplicadas a macromoléculas purificadas em solução ou em estado cristalino, respectivamente 1,2. Considerando que a tomografia crio-elétron (crio-ET) é exclusivamente adequada para estudos estruturais e ultraestruturais quase nativos de objetos grandes e heterologos, como bactérias, vírus pleomórficos e células eucarióticas3. No crio-ET, as informações tridimensionais (3D) são obtidas inclinando fisicamente a amostra no estágio do microscópio e adquirindo uma série de imagens através da amostra em diferentes ângulos. Essas imagens, ou séries de inclinação, geralmente cobrem uma faixa de +60/-60 graus em incrementos de um a três graus. A série de inclinação pode então ser reconstruída computacionalmente em um volume 3D, também conhecido como tomograma4.
Todas as técnicas crio-EM requerem que a amostra seja incorporada em uma fina camada de gelo amorfo, não cristalino e vítreo. Uma das técnicas de criopensa mais utilizadas é o congelamento de mergulho, onde a amostra é aplicada à grade EM, borrada e rapidamente mergulhada em etano líquido ou uma mistura de etano líquido e propano. Esta técnica é suficiente para a vitrificação de amostras de <100 nm a ~10 μm de espessura, incluindo células humanas cultivadas, como células HeLa5,6. Amostras mais grossas, como mini-organóides ou biópsias teciduais, até 200 μm de espessura, podem ser vitrificadas por congelamento de alta pressão7. No entanto, devido ao aumento da dispersão de elétrons de amostras mais espessas, a amostra e a espessura do gelo para crio-ET é limitada a ~0,5 - 1 μm em microscópios eletrônicos de transmissão de 300 kV. Portanto, crio-ET de células inteiras de muitas células eucarióticas está limitado à periferia celular ou extensões de células, a menos que sejam usadas etapas adicionais de preparação de amostras, como crio-seccionamento8 ou moagem de feixe de íons focais9,10,11.
Uma limitação de muitos experimentos de imagem crio-ET de células inteiras é a coleta de dados através de 12. Ao contrário do crio-EM de partículas únicas, onde milhares de partículas isoladas podem muitas vezes ser imagens de um único quadrado de grade TEM, as células são grandes, espalhadas, e devem ser cultivadas em baixa densidade suficiente para permitir que as células sejam preservadas em uma fina camada de gelo vítreo. Muitas vezes a região de interesse é limitada a uma característica particular ou subárea da célula. Limitando ainda mais o throughput é a propensão de as células crescerem em áreas que não são favoráveis à imagem TEM, como em ou perto de barras de grade TEM. Devido a fatores imprevisíveis associados à cultura celular nas redes TEM, os desenvolvimentos tecnológicos são necessários para melhorar a acessibilidade e o throughput da amostra para aquisição de dados.
Micropatterning substrato com proteínas de matriz extracelular (ECM) é uma técnica bem estabelecida para microscopia de luz de células vivas para direcionar o crescimento de células em superfícies rígidas, duráveis e opticamente transparentes, como vidro e outras culturas teciduais substratos13,14. A micropatters também foi realizada em superfícies macias e/ou tridimensionais (3D). Tais técnicas não só permitiram o posicionamento preciso das células; eles também apoiaram a criação de redes multicelulares, como circuitos de células neurais padronizadas15. Trazer micropatterning para crio-ET não só aumentará o rendimento, mas também pode abrir novos estudos para explorar microambientes celulares complexos e dinâmicos.
Recentemente, vários grupos começaram a usar técnicas de micropattering em grades TEM através de múltiplas abordagens16,17. Aqui, o uso de uma técnica de fotopatterning sem máscara para grades TEM é descrito usando o sistema de micropatterning Alvéole PRIMO, que possui padronização de alta resolução e sem contato. Com este sistema de micropatterning, uma camada anti-incrustação é aplicada na parte superior do substrato, seguida pela aplicação de um fotocatalyst e ablação da camada anti-incrustação em padrões definidos pelo usuário com um laser UV. As proteínas ECM podem então ser adicionadas aos padrões para a cultura celular apropriada. Este método tem sido usado por vários grupos para estudos crio-ET de pigmento retiniano epitelial-1 (RPE1), Madin-Darby canino kidney-II (MDCKII), fibroblast de prepúcio humano (HFF) e linhas celulares endoteliais16,17,18. Este sistema de micropatterning é compatível com vários substratos de camada anti-incrustar, bem como um reagente de fotocatalyst líquido ou gel. Uma variedade de proteínas ECM pode ser selecionada e adaptada para a especificidade da linha celular, conferindo versatilidade para o usuário.
A micropattersão foi aplicada com sucesso a uma série de projetos dentro do laboratório19. Aqui, é apresentado um protocolo de micropatterning, incluindo adaptações específicas para estudar células HeLa cultivadas, células BEAS-2B infectadas pelo vírus sincicial respiratório (RSV) e neurônios mamóquios 20 larval primários.
Protocolo
O protocolo descrito aqui é uma compilação da cultura celular, micropatterning e métodos de imagem usados pelo laboratório Wright e pelo Centro de Pesquisa Cryo-EM da Universidade de Wisconsin, Madison. O fluxo de trabalho é apresentado na Figura 1. Treinamento adicional e materiais instrutivos estão disponíveis nos seguintes locais: https://cryoem.wisc.edu ou https://wrightlab.wisc.edu
1. Preparação de grades para padronização
- Transfira as grades TEM para um escorregador de vidro limpo, lado carbono para cima (a espessura padrão da folha de carbono é de 12 nm). Usando um evaporador de carbono, ACE600, evaporar 5-8 nm de carbono adicional nas redes para aumentar a durabilidade geral da película de carbono.
NOTA: Esta etapa não é necessária para grades SiO2 . Esta etapa também pode ser feita com antecedência; armazene as grades revestidas em um ambiente de baixa umidade, como um dessecador de vácuo. - Transfira as grades para um suporte de preparação de grade e exacar as grades do lado carbono para cima. Utilizando um sistema de descarga de brilho, o brilho descarrega as grades para 60 s a 10 mA com uma distância de trabalho de 80 mm e pressão de vácuo de 1,0 x 10-3 mbar. Faça isso dentro de 15-30 minutos do próximo passo.
NOTA: Os suportes de preparação da grade podem ser comprados comercialmente ou caseiros com um pedaço de papel filtro em uma pequena placa de Petri.
2. Aplicação da camada anti-incrusto
NOTA: Deve ser utilizada uma técnica estéril adequada ao manusear as grades, e todas as soluções devem ser estéreis e/ou filtradas.
- Transfira as grades (lado carbono para cima) para um deslizamento de vidro limpo ou deslizamento de tampas com pelo menos 1 cm de separação entre as grades. Pipeta 10 μL de 0,05% poli-L-lysine (PLL) em cada grade. Incubar as grades em uma câmara úmida, como uma caixa de plástico fechada com toalhas de papel úmidas, por pelo menos 30 minutos.
NOTA: Esta etapa pode ser estendida para durante a noite. Certifique-se de que o nível de umidade na câmara é suficiente para evitar que as grades sequem. - Lave cada grade três vezes com 15 μL de 0,1 M HEPES pH 8.5. Para cada lavagem, remova a maior parte do líquido da rede com uma pipeta sem deixar a rede secar. Adicione 15 μL de tampão fresco, incubar por pelo menos 30 s e repita. Deixe cada grade em 15 μL de 0,1 M HEPES após a lavagem final.
NOTA: Nesta etapa e etapas futuras, é importante manter a grade molhada e evitar o contato entre a pipeta e a grade. - Prepare 10 μL de 100 mg/mL de polietileno glicol-succinimidyl valerate (PEG-SVA) em 0,1 M HEPES pH 8,5 para cada grade. O PEG-SVA dissolverá-se rapidamente com uma mistura suave resultando em uma solução clara.
NOTA: Não prepare a solução PEG-SVA com antecedência. PEG-SVA tem uma meia-vida de 10 min em pH 8,5. Evite expor o estoque PEG-SVA à umidade excessiva armazenando-o em um ambiente desiccador ou seco a -20 °C e aquecendo-se à temperatura ambiente antes de abrir. - Imediatamente após a preparação da solução PEG-SVA, remova a gota de 15 μL de HEPES pH 8,5 de cada grade (tomando cuidado para não secar a grade) e adicione uma gota de 10 μL da solução PEG-SVA. Incubar as grades em uma câmara úmida por pelo menos 1 h.
NOTA: Esta etapa pode ser estendida para durante a noite. Certifique-se de que a umidade na câmara é suficiente para evitar que as grades sequem. - Lave cada grade três vezes com 15 μL de água estéril. Para cada lavagem, retire a maior parte do líquido da rede com uma pipeta sem deixar a rede secar, adicione 15 μL de água doce, incubar por pelo menos 30 s e repetir. Deixe cada grade em 15 μL de água após a lavagem final.
3. Aplicação do gel PLPP
- Prepare uma tampa de microscópio limpo para cada grade. Complete as etapas a seguir para cada grade, uma grade de cada vez, para minimizar a chance de a grade secar.
- Coloque uma gota de 1,0 μL de água no centro do deslizamento de tampas para ajudar a colocar a grade na tampa e manter a grade molhada. Transfira cuidadosamente a rede da gota de água de 15 μL para a gota de água de 1,0 μL na mancha de cobertura. Certifique-se de colocar o lado de carbono da rede para cima.
- Coloque cuidadosamente um estêncil de polidimetilatilaxitano (PDMS) sobre a grade, tomando o cuidado de manter a rede centrada e minimizar o contato de estêncil com a folha de carbono da rede.
- Adicione 1,0 μL de 4-benzoilbenzyl-trimethylammonium cloreto (PLPP) na grade. Pipeta suavemente para misturar (não toque na grade com a ponta da pipeta).
- Mova a tampa com a grade para um local escuro para secar. O gel secará em aproximadamente 15-30 min.
4. Calibração e design do micropattern
- Colorir um lado de uma tampa de vidro com um marcador. Adicione linhas pretas de um marcador permanente de ponta fina para facilitar o foco. Coloque a mancha no microscópio de tal forma que o lado colorido enfrente a lente objetiva. Usando o modo brightfield, concentre-se no marcador.
- Certifique-se de que o microscópio e o sistema de micropatterning estão ligados e o caminho correto da luz está definido. Abra o Micromanager e o software Leonardo (Plugins > Leonardo) no computador do microscópio.
- Selecione calibrar e siga as instruções na tela. Ajuste o foco do microscópio para que a imagem projetada no slide esteja em foco. O tempo de exposição pode precisar ser diminuído. Após a calibração, selecione Padrão Agora.
- Registo micrometro/pixel (μm/px) relatados sob dados de calibração na janela superior esquerda do programa (Figura 2, área 1). Use essa proporção para determinar o número de pixels a serem usados por micrômetro ao projetar um padrão.
- Após a calibração, certifique-se de que o software está agora aberto com uma visão brightfield ao vivo do microscópio. Carregue uma grade preparada em um deslizamento (seção 3) no palco com a grade voltada para a lente objetiva. Posicione o estágio e ajuste o foco para que a grade fique visível na janela do software.
- Meça o tamanho dos quadrados da grade e barras de grade em micrômetros. O software inclui uma régua ativada pelo botão perto do canto inferior esquerdo para medir a grade (Figura 2, área 2). Por exemplo, os padrões usados aqui para uma grade de malha de 200 correspondem a ~87 × quadrados de grade de 87 μm e barras de grade de ~36 μm.
NOTA: O software oferece flexibilidade na redimensionamento de padrões em tempo real, de modo que pequenas imprecisões na medição podem ser toleradas. - Com base nas medições e razões acima, crie padrões com qualquer software de criação de imagem. O tamanho mínimo do recurso com um objetivo de 20 × é de 1,2 μm. Os padrões devem ser salvos como arquivos .tiff de 8 bits não comprimidos.
- Certifique-se de que o software não redimensione as imagens para um tamanho de pixel diferente ao salvar. O padrão deve caber dentro de uma caixa de 800 × 800 pixels, suficiente para cobrir quatro quadrados de grade.
NOTA: Pixels com valor de 255 (branco) serão padronizados na maior intensidade (dose total do laser) e pixels com valor de zero (preto) não serão padronizados. Quaisquer pixels com um valor intermediário serão padronizados com uma dose de aproximadamente (X/255)*dose total. Na Figura 3A, foram utilizados valores de pixels de 255 e 129 para os padrões de escala de cinza. Uma vez que o padrão é projetado, ele pode ser salvo e reutilizado sem modificação.
- Certifique-se de que o software não redimensione as imagens para um tamanho de pixel diferente ao salvar. O padrão deve caber dentro de uma caixa de 800 × 800 pixels, suficiente para cobrir quatro quadrados de grade.
5. Micropatterning
- Após a calibração, certifique-se de que o software está agora aberto com uma visão brightfield ao vivo do microscópio. Carregue uma grade preparada em um deslizamento (seção 3) no palco com a grade voltada para a lente objetiva. Posicione o estágio e ajuste o foco para ver a grade no software.
- Para uma execução inicial, projete um novo modelo. No software, selecione Adicionar ROI (não mostrado, na localização da área 3 da Figura 2 ) e escolha um círculo de 3.000 μm. Posicione o ROI do círculo sobre a grade usando a imagem de campo brilhante na tela como um guia. Pressione a trava para fixar o ROI.
- Após bloquear o ROI no lugar, selecione Adicionar padrão (não mostrado, na localização da figura 2 área 3). Escolha o padrão projetado na seção 4. Divida a rede em seis regiões para permitir que o foco e o posicionamento independentes em cada região considerem grades irregulares. Uma região quadrada de 8 × 8 grades para cada canto da grade e uma região quadrada de 2 × 8 grades em cada lado do centro do centro, deixando o centro quatro quadrados de grade sem ser forreado (Figura 2, imagem central).
- Use as opções de replicação (Figura 2, área 4) para gerar cópias do padrão inicial para atingir o número desejado de cópias totais do padrão. Ajuste o espaçamento entre cópias para combinar o espaçamento entre os quadrados da grade, se necessário.
- Defina a Dose Total para o padrão. 30 mJ/mm2 é um bom ponto de partida. Consulte a seção de discussão para mais detalhes.
- Em Opções especializadas (Figura 2, área 4) ajuste o ângulo da região para combinar com o dos quadrados da grade. As regiões podem ser reposicionadas usando o mouse. A razão (tamanho) dos padrões também pode ser ajustada. Iterar através do ajuste do ângulo, posição, espaço entre e proporção do padrão até que os padrões se alinhem com os quadrados da grade. Mova o estágio do microscópio para alterar a região da grade no visor de campo brilhante ao vivo.
- Pressione lock para salvar as alterações feitas na região.
- Para copiar uma região, clique no botão Duplicata (Figura 2, área 5, duas folhas de ícone de papel) ao lado de seu nome no painel Ações à esquerda. Para reposicionar, renomear ou editar a cópia, clique em seu nome no painel Ação.
- Repita as etapas 5.4-5.9 conforme necessário para preencher todas as regiões desejadas.
- Uma vez que o modelo completo seja projetado e posicionado, salve o arquivo de modelo dentro do software (Figura 2, área 6, barra com ícone de seta para cima na barra de ferramentas superior).
- Ao carregar um modelo previamente salvo (barra com ícone de seta para baixo) centralizar o ROI sobre a grade e pressionar bloquear. Clique em cada região no Painel de Ação para alterar o ângulo, posição, dose e/ou arquivo padrão.
- Uma vez que o modelo e os padrões estejam posicionados, desmarque todas, exceto uma das regiões do Painel de Ação no software.
- Use o estágio do microscópio para navegar até aquela região e se concentrar na folha de carbono. Clicar no ícone Eyeball no painel Ação (Figura 2, área 5) alternará a exibição da sobreposição de padrão sobre a sobreposição ou desligada.
- Uma vez que a grade esteja em foco, feche o obturador brightfield e pressione o ícone Play no canto inferior direito do software para iniciar o processo de padronização, que pode ser monitorado ao vivo.
- No painel de ação, selecione a caixa para a próxima região. Abra o obturador brightfield para que a grade seja visível e centralizar essa região usando o estágio do microscópio. Repita as etapas 5.13-5.14 para cada região no Painel de Ação.
- Remova a mancha de cobertura com a grade do microscópio e, imediatamente, pipeta 10 μL de soro fisiológico estéril tamponado de fosfato (PBS) na grade.
- Depois de 10 min, retire o estêncil com pinças e depois lave a grade 3x com 15 μL de PBS. Após a lavagem final, coloque cada grade em 15 μL de PBS e mova as grades para um local escuro.
6. Deposição de proteínas ECM
- Para células cultivadas, siga os passos 6.2-6.5; para neurônios primários de Drosophila , siga os passos 6.6-6.10.
- Prepare pelo menos 15 μL de ECM para cada grade. Para células BEAS-2B, prepare uma concentração final de fibronectina bovina de 0,01 mg/mL e 0,01 mg/mL de fibrinogênio conjugado por fluoródono em PBS estéril. Para células HeLa, prepare 0,01 mg/mL de colágeno bovino I e 0,1 mg/mL de fibrinogênio conjugado fluorophore em PBS estéril.
- Remova a maior parte do PBS de cada grade e aplique 15 μL do ECM. Incubar a rede em uma câmara úmida à temperatura ambiente por pelo menos 1 h.
NOTA: Esta etapa pode ser estendida para durante a noite a 4 °C. - Após a incubação em ECM, lave cada grade 5x com PBS estéril. Para cada lavagem, retire a maior parte do líquido com uma pipeta sem deixar a rede secar, adicione 15 μL de PBS fresco, incubar por pelo menos 30 s e repita. Deixe cada grade na PBS após a lavagem final.
NOTA: As grades podem ser armazenadas por até uma semana em PBS a 4 °C sem deterioração observada na qualidade. - Use um microscópio de fluorescência para detectar o fluoróforo no ECM para confirmar a padronização e que a folha de carbono permaneceu intacta. Alguns quadrados quebrados são geralmente toleráveis.
- Para neurônios primários de Drosophila , mova as grades padronizadas para uma antena de fundo de vidro de 30 mm contendo PBS estéril.
- Aspire o PBS do prato e aplique 2 mL de concanavalina conjugada a fluorophore a. Incubar durante a noite a 25 °C em um ambiente estéril.
- Retire a concanavalina Uma solução do prato (sem secar as grades) e lave as grades 3x com PBS. Para cada lavagem, adicione e retire 2 mL PBS do prato.
- Use um microscópio de fluorescência para detectar o fluoróforo no ECM para confirmar a padronização e que a folha de carbono permaneceu intacta. Alguns quadrados quebrados são geralmente toleráveis.
- Após a lavagem final, retire o PBS do prato de fundo de vidro e adicione 2 mL de recém-preparado, filtros estéreis A Drosophila media21 de Schneider, contendo 20% de soro bovino fetal inativado por calor (FBS), 5 μg/mL de insulina, 100 μg/mL penicilina, 100 μg/mL estreptomicina e 10 μg/mL tetraciclina. Incubar a 25 °C em um ambiente estéril até que os neurônios estejam prontos para serem banhados.
7. Preparação de células primárias de Drosophila antes da semeadura
- Esterilize um prato de dissecção de 55 mm com 70% de EtOH e, em seguida, adicione ao prato 2-3 mL de soro fisiológico de dissecção filtrada 1× (9,9 mM HEPES pH 7.5, 137 mM NaCl, 5,4 mM KCl, 0,17 mM NaH2PO4, 0,22 mM KH2PO4, 3,3 mM de glicose, 43,8 mM de sacarose)21.
- Escolha 30-40 3ª larva instar suavemente da comida usando um par de pinças.
- Coloque as larvas no tubo com 1× PBS, depois transfira-as para o segundo tubo com 1× PBS para lavar as larvas.
- Transfira as larvas para o tubo com 70% de EtOH para lavar o PBS e, em seguida, transfira-as para o segundo tubo com 70% de EtOH. Deixe as larvas no segundo tubo por 2-3 minutos para esterilizar as larvas.
- Transfira as larvas para um tubo com soro fisiológico de dissecção de 1× e, em seguida, transfira-as imediatamente para o segundo tubo com soro fisiológico de dissecção de 1×.
- Transfira larvas individuais para o prato de dissecação contendo salina de dissecção de 1×. Com um par de fórceps e um microscópio de dissecção, rasgue rapidamente cada larva para extrair o cérebro e transferi-lo para o terceiro tubo com 1× dissecção salina. Repita até que todos os cérebros sejam extraídos.
- Centrifugar o tubo contendo o cérebro a 300 x g por 1 min.
- Descarte o supernasal e lave com 1 mL de 1× dissecção salina e centrífugas do tubo a 300 x g por 1 min. Repita este passo mais uma vez.
- Descarte o supernasciente até que 200-250 μL seja deixado no tubo e adicione 20 μL de 2,5 mg/mL Liberase em 1x dissecção salina.
- Gire o tubo em um rotador por 1h à temperatura ambiente; durante esta hora, pipeta a solução 25-30 vezes a cada 10 min. No final, a solução deve ser um pouco opaca.
- Centrifugar as células a 300 × g por 5 min.
- Descarte o supernatante, depois adicione 1 mL de mídia de Schneider complementada. Pipeta a solução 30 vezes para misturar.
- Centrifugar as células a 300 × g por 5 min.
- Descarte o supernatante e lave a pelota celular adicionando 1 mL de mídia de Schneider suplementada. Pipeta a solução 30 vezes para misturar.
- Centrifugar as células a 300 × g por 5 min.
- Descarte o supernatante e, em seguida, resuspenque a pelota celular com 300 μL de mídia de Schneider suplementada. Pipeta a solução 30-40 vezes para misturar.
8. Infecção cultural e RSV de células BEAS-2B e HeLa
- Mantenha as células HeLa e AS-2B em frascos T75 a 37 °C e 5 % de CO2. Células de passagem a cada 3-4 dias, uma vez que atingem aproximadamente 80% de confluência. Manter células HeLa em DMEM + 10% FBS + 1× Antibiótico-Antimycotico. Manter o BEAS-2B em RPMI + 10% FBS + 1× Antibiótico-Antimycótico6,22,23.
- Para semeadura de células não infectadas, pule para a seção 9. As células BEAS-2B e HeLa são suscetíveis à infecção por RSV; As células BEAS-2B foram utilizadas para todos os experimentos envolvendo RSV mostrados aqui.
NOTA: Realize todas as etapas do BSL-2 em conformidade com os protocolos institucionais utilizando um gabinete de biossegurança (BSC) e equipamentos de proteção individual (EPI) apropriados. - Antes da infecção por RSV de células, a passagem 5 × 104 células por poço em uma placa de 6 poços (área de superfície ~9,6 cm2) com 2 mL de mídia de crescimento e incubar durante a noite.
- Tentepsinizar e contar um poço de células. Para tentarpsinizar, aspire a mídia de um poço e lave com 2 mL de PBS estéril sem Mg2+ e Ca2+ para remover a mídia residual. Adicione 500 μL de solução de trippsina de 0,25%. Incubar a 37 °C por 5-10 min. Verifique periodicamente as células para ver se elas são liberadas da superfície. Uma vez que as células são liberadas, adicione 1,5 mL de mídia cultural.
- Misture 100 μL de células experimentadas com 100 μL de azul trypan. Pipeta 10 μL de mistura de células diluídas em um hemócito. Conte as células e calcule o número de células por poço. Use este número para calcular MOI abaixo.
- Prepare uma diluição do RSV-A2mK+24 em mídia de crescimento para alcançar um MOI de 10 por poço em 750 μL de mídia. O MOI de RSV-A2mK+ pode ser calculado a partir de títulos fluorescentes de unidades focais fluorescentes (FFU) do estoque (Por exemplo: para 1,0 × 105 células por poço e um estoque de RSV de 1,0 × 108 FFU/mL, diluir o estoque viral 1:75 a 1 × 106 FFU/750 μL ou 1,33 × 106 FFU/mL).
- Aspire a mídia das células no prato de 6 poços e adicione 750 μL da solução viral de cima para cada poço.
- Balance a placa em temperatura ambiente por 1h.
- Depois de 1h, leve o volume total por poço até 2 mL com mídia de crescimento pré-aquecida para 37 °C e coloque a placa em uma incubadora definida para 37 °C com 5% de CO2 por 6h.
- Tentesinizar as células para soltá-las e proceder à semeadura conforme descrito abaixo. Após a semeadura, incubar as grades por mais 18 h antes de mergulhar no congelamento (para um total de 24 h pós-infecção).
9. Semeadura de células em grades micropatteradas
- Para células cultivadas, siga os passos 9.2-9.8; para neurônios primários de Drosophila , siga 9.9-9.11.
- Tentepsinizar as células para soltá-las (ver passo 4 na seção 8 acima). Para reduzir a agregação celular, tentepsinizar as células a 60 % ou menos confluência.
- Misture 100 μL de células experimentadas com 100 μL de azul trypan. Pipeta 10 μL da célula diluída se mistura em um hemótmetro e conte as células.
- Diluir as células na mídia para 2 × 104 células/mL.
- Adicione 1 μL de mídia ao centro de uma antena de fundo de vidro de 30 mm para ajudar na colocação da grade e evitar que ela seque. Transfira a grade do PBS no deslizamento de tampas para o centro do prato de fundo de vidro. Adicione 10 μL de solução celular à grade.
- Usando um microscópio de campo brilhante, observe a adesão celular à rede após 5 minutos. Se a maioria dos padrões permanecer desocupada, adicione uma gota adicional de 10 μL da solução celular. Mantenha as grades e a solução celular a 37 °C durante as incubações.
- Repita o passo 9.6 até que a maioria dos padrões sejam ocupados ou muitos padrões ocupados comecem a ter múltiplas células. Incubar a rede por 2h na incubadora (37 °C, 5% CO2).
- Inunde o prato com 2 mL de mídia pré-aquecida e incubar durante a noite (37 °C, 5% de CO2).
- Para neurônios primários de Drosophila , remova a mídia do prato contendo grade e coloque as células sobre o prato.
- Espere 30-60 min para as células anexarem, em seguida, adicione 2 mL de mídia de Schneider complementada.
- Cultue os neurônios em uma incubadora de 25 °C por um mínimo de 2-3 dias antes do congelamento do mergulho.
10. Imagem e vitrificação de grades padronizadas
- Coloque o prato de fundo de vidro contendo a grade padronizada e células cultivadas no microscópio de fluorescência.
- Adquira imagens da grade usando brightfield e os canais fluorescentes apropriados para detectar o padrão e qualquer outra rotulagem nas células. Certifique-se de que a densidade e o posicionamento das células são adequados para imagens e análises a jusante.
NOTA: Imagens brilhantes e fluorescentes foram processadas no pacote de software FIJI25. - Prepare um freezer crio-mergulho; o tipo de dispositivo de congelamento dependerá da disponibilidade, custo e recursos mais adequados para a amostra.
NOTA: Os neurônios primários de Drosophila foram preparados em um congelador automatizado, e as células BEAS-2B foram preparadas utilizando um congelador semi-automatizado. - Aplique fiduciais de ouro nas amostras para o alinhamento adequado da série de inclinação. Amostras de manchas para remover o excesso de mídia, em seguida, congelar as amostras em um criogeno, como o etano líquido resfriado por nitrogênio líquido. Para neurônios primários de Drosophila , blot para 4 s do traseiro. Para as células HeLa e BEAS-2B, blot de ambos os lados para 4-6 s. As redes congeladas podem então ser armazenadas em nitrogênio líquido até que sejam mais utilizados.
- Células vitrificadas de imagem em um microscópio crio-elétron, operado a 300 kV com uma câmera detector de elétrons direta. Configure a coleção de séries de inclinação para cada região de interesse com softwares como SerialEM26 para coleta de dados crio-EM/cryo-ET.
NOTA: A série de inclinação de neurônios Drosophila primários foi coletada em um detector de elétrons direto de -60° a 60° bidirecionalmente a incrementos de 2° a -8 μm defoco com um tamanho de pixel de 4.628 Å para uma dose total de 70-75 e-/Å2. A série de inclinação do BEAS-2B infectado por RSV foi coletada em um detector de elétrons direto com um filtro de energia (20 eV fenda) a -5 μm defoco com um tamanho de pixel de 4.603 Å e uma dose total de ~80 e-/Å2. - Processe a série de inclinação para reconstruir os tomogramas.
NOTA: Os tomogramas aqui apresentados foram reconstruídos utilizando o pacote IMOD27; a filtragem lowpass foi feita usando o pacote de software EMAN2228.
Resultados
Este procedimento foi usado para padronizar grades EM para experimentos crio-ET de células inteiras. Todo o fluxo de trabalho apresentado neste estudo, incluindo preparações iniciais de cultura celular, micropatterning (Figura 1) e imagem, abrange de 3 a 7 dias. Um procedimento de duas etapas foi utilizado para gerar a camada anti-incrustamento, aplicando PLL à grade e, posteriormente, vinculando PEG por adição do PEG-SVA reativo. A camada anti-incrustion também pode ser aplicada em um único passo adicionando PLL-g-PEG em uma incubação. O gel PLPP é um catalisador para a micropatterning UV, que também está disponível como um líquido menos concentrado. O gel permite a padronização em uma dose significativamente reduzida em comparação com o líquido, o que resulta em uma padronização muito mais rápida. Com este sistema, o tempo real de padronização de uma grade TEM completa foi de ~2 minutos. O fluxo de trabalho de micropatterning sozinho geralmente abrange de 5 a 6 horas e permite que um indivíduo padrone oito grades para cultura celular padrão em grades TEM.
Algumas etapas durante o processo de micropatterning requerem longos tempos de incubação (ver etapas 2.1, 2.3, 6.4). Convenientemente, algumas dessas etapas, como a passivação PLL (2.1) ou a passivação PEG-SVA (2.3), podem ser estendidas para uma incubação noturna. Além disso, as grades podem ser padronizadas com antecedência e armazenadas em uma solução da proteína ECM ou PBS para uso posterior. Em nosso estudo, essas opções foram valiosas em casos em que o tempo de preparação e semeadura celular é crítico, como neurônios primários de Drosophila e infecção por RSV de células BEAS-2B.
As grades foram preparadas em um ambiente geral de laboratório de nível de biossegurança 2 (BSL-2) utilizando ferramentas limpas, soluções estéreis e incluiu antibióticos/antimípticos nas mídias de crescimento6,22,29,30. Para amostras particularmente sensíveis à contaminação microbiana, a camada anti-incrustante e o ECM podem ser aplicados em uma capa de cultura tecidual ou outro ambiente estéril. Além disso, a rede poderia ser lavada em etanol entre a padronização e a aplicação do ECM. Se trabalhar com agentes infecciosos, é importante adaptar o procedimento para cumprir os protocolos adequados de biossegurança.
Este fluxo de trabalho e os procedimentos apresentados (Figura 1) permitiram que as células HeLa (Figura 4), células BEAS-2B infectadas por RSV (Figura 3, Figura 5) e neurônios larvais de Drosophila primários (Figura 6, Figura 7) fossem semeadas em grades EM padronizadas para uma coleta ideal de dados crio-ET.
As células HeLa semeadas em grades TEM micropatteradas permanecem viáveis conforme determinado pela coloração fluorescente usando um ensaio de viabilidade celular baseado em calceínio-AM e ethidium-1 (Figura 4A,B). Usando um ECM misto de colágeno e fibrinogênio, as células HeLa facilmente aderem a padrões através da grade (Figura 4A,C). A morfologia geral das células que se expandem ao longo do padrão é semelhante à das células cultivadas em grades não padronizadas (Figura 4C,D). No caso das células HeLa, a espessura total da célula permanece ~< 10 μm com áreas significativamente mais finas ~< 1 μm de espessura perto da periferia celular (Figura 4E,F).
Para estudos de RSV, padronizamos quadrados de grade inteiros usando um gradiente, com uma exposição de baixa dose nas bordas e um padrão de dose maior em direção ao centro (Figura 3A). Os padrões de gradiente produziram melhores resultados na busca por vírus liberados presentes perto da periferia das células. Com esses padrões, as células foram encontradas preferencialmente aderindo à maior concentração de ECM, mas também são capazes de aderir e crescer nas concentrações de ECM mais baixas. A dose relativa entre as áreas precisará ser otimizada ao usar padrões que requerem múltiplas doses. Se as doses e, portanto, as concentrações de ECM forem muito semelhantes ou muito díspares umas às outras, o efeito do uso de múltiplas doses será perdido.
Na Figura 3, uma grade TEM foi padronizada e posteriormente semeada com células BEAS-2B infectadas pelo RSV e utilizada para coleta de dados crio-EM. Figura 4A é uma imagem fluorescente de ECM padronizada em uma grade TEM usando um padrão de gradiente. A adesão celular e o crescimento ao longo da região central do padrão podem ser vistos na Figura 3B como uma imagem brilhante das células 18 horas após a semeadura. Na Figura 3C, o sinal fluorescente (vermelho) da replicação do RSV-A2mK+ é sobreposto com sinal do ECM. A maioria das células infectadas estão posicionadas ao longo da região central de maior densidade do padrão gradiente. Um mapa TEM de baixa mag da grade pós crio-fixação revela uma série de células, incluindo células infectadas por RSV, posicionadas na folha de carbono perto do centro dos quadrados da grade. Como mostrado anteriormente para células cultivadas nas grades tem padrão22, séries de inclinação foram localizadas e coletadas de virions RSV nas proximidades da periferia de células BEAS-2B infectadas cultivadas em grades micropadrificadas (Figura 5A,B). Muitas das proteínas estruturais RSV podem ser identificadas dentro dos tomogramas, incluindo nucleocapsídeo (N) e a proteína de fusão viral (F) (Figura 5C, setas azuis e vermelhas, respectivamente).
Para estudos primários de neurônios de Drosophila, verificou-se que o padrão estreito, próximo ao limite de resolução oferecido pelo software (onde a espessura do padrão era de 2 μm), permitia que de uma a poucas células fosse isolada dentro de um quadrado de grade (Figura 6). A soma neuronal foi capaz de estender seus neurites durante um período de vários dias dentro do padrão. Isso permitiu fácil identificação e aquisição de séries de inclinação dos neurites em comparação com neurônios cultivados em grades não respingadas (Figura 7). Verificou-se também que a concanavalina fluorescente A, uma lectina que tem sido usada como ECM para culturas neuronais in vitro Drosophila20,21, é favorável à padronização.
Os neurônios de drosophila da terceira larva instar foram isolados de acordo com protocolos publicados anteriormente20,21,31. As preparações neuronais foram aplicadas em grades crio-EM micropatteradas onde a concanavalina A foi depositada no padrão para regular a colocação, a disseminação e a organização das células. Os neurônios em grades padronizadas ou não padronizadas foram autorizados a incubar por um mínimo de 48-72 horas, e as grades foram então mergulhadas congeladas. Uma imagem representativa de uma rede EM micropatterada com vários neurônios de Drosophila distribuídos pelas regiões padronizadas é mostrada na Figura 6A. Esses neurônios, derivados de uma cepa de mosca transgênica que tem expressão de GFP pan-neuronal na membrana, podem ser facilmente rastreados por microscopia leve não apenas devido à sua rotulagem fluorescente, mas também por causa de sua localização dentro das micropatterns. Enquanto neurônios cultivados em redes não respingadas também podem ser rastreados através de sua sinalização GFP por microscopia leve (Figura 7A, círculo amarelo), localizá-los em crio-EM tornou-se substancialmente mais difícil devido à presença de detritos celulares e contaminação da mídia (Figura 7B, círculo amarelo). Tal presença foi diminuída para neurônios em grades padronizadas, provavelmente devido ao PEG na camada anti-incrustante das regiões não padronizadas que repeliram os detritos celulares da adesão. Devido às dimensões do corpo celular neurônio e aos neurites estendidos (Figura 6A,B, círculo amarelo), as séries de inclinação crio-ET foram coletadas ao longo de regiões mais finas das células (Figura 6C,D, círculo vermelho). A membrana celular neuronal, uma mitocôndria (ciano), microtúbulos (roxo), filamentos de actina (azul), estruturas vesiculares (laranja e verde) e macromoléculas como ribossomos (vermelho) foram bem resolvidas em montagens de imagem de maior ampliação e fatias através do tomograma 3D (Figura 6E). Embora características subcelulares semelhantes possam ser vistas a partir de tomogramas 3D de neurônios não padrões (Figura 7E), a dificuldade em localizar alvos celulares viáveis para coleta de dados diminuiu substancialmente o rendimento.
Na Figura 8, imagens representativas de grades com algumas dessas questões foram reunidas para auxiliar na identificação e solução de problemas. Uma vez determinadas condições ideais, a micropattering é um método confiável e reprodutível para o posicionamento de células em grades para crio-TEM.

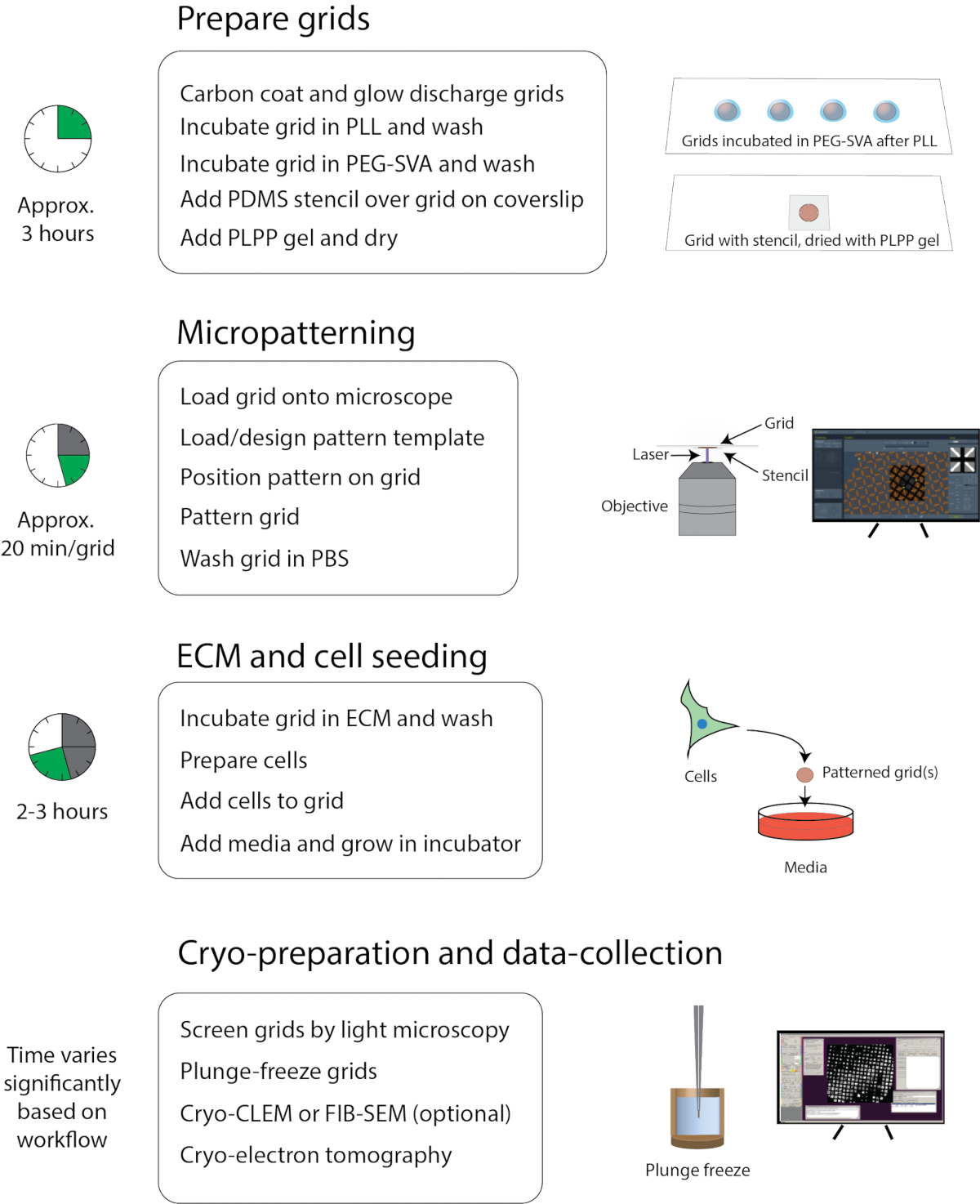
Figura 1: Fluxo de trabalho geral de micropatterning para crio-EM. O fluxo de trabalho pode ser aproximadamente dividido em quatro partes: preparação da grade, micropatterning, ECM e semeadura celular, e crio-preparação e coleta de dados. As principais etapas de cada seção estão listadas abaixo dos títulos e o tempo aproximado para completar cada seção é mostrado à esquerda. Clique aqui para ver uma versão maior desta figura.
{kind=link}

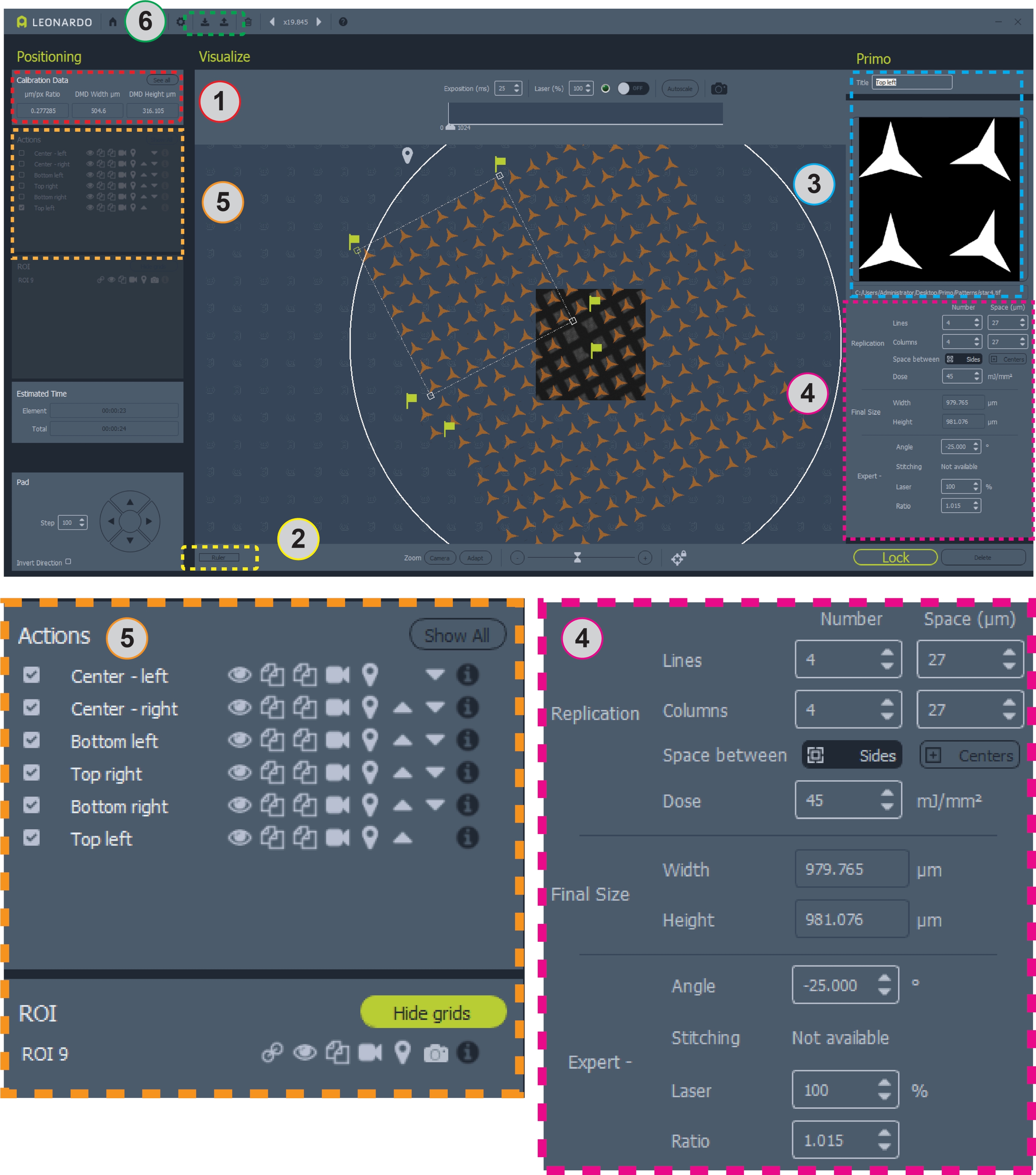
Figura 2: Captura de tela do software com padrão posicionado na grade. A área 1 contém a razão μm/pix para o design do padrão. A Área 2 é a régua para medir uma rede. A Área 3 é onde adicionar ou alterar padrões e ROIs. A Área 4 contém todas as informações para posicionamento padrão e dose. A Área 5 contém opções para padrões, incluindo sobreposições de toggling, copiar ou excluir padrões e selecionar padrões para micropatterning. A área 6 é onde os modelos podem ser salvos e carregados. Vistas maiores das áreas 4 e 5 são mostradas abaixo para clareza. Clique aqui para ver uma versão maior desta figura.
{kind=link}

Figura 3: Células BEAS-2B infectadas por RSV na grade crio-TEM padronizada. (A) Imagem fluorescente da grade padronizada após a adição de ECM rotulado fluorescentemente. O padrão de entrada é mostrado no canto inferior esquerdo. (B) Imagem de campo brilhante das células BEAS-2B cultivadas na grade em A. (C) Mesclagem da imagem em A (ciano) e B (cinza) com imagem fluorescente de células infectadas por RSV (vermelho) imediatamente antes do congelamento do mergulho; células infectadas expressam mKate-2. As barras de escala são 500 μm. (D) Mapa crio-TEM de baixa ampliação da grade em B após o congelamento de mergulho. Imagens fluorescentes são pseudocoloridas. As barras de escala são de 500 μm. Clique aqui para ver uma versão maior desta figura.
{kind=link}

Figura 4: Coloração viva/morta de células padronizadas e não padronizadas. (A) Imagem fluorescente de células HeLa cultivadas em uma grade padronizada e manchadas com calceíno-AM (mancha de célula viva, verde) e homodimer-1 (mancha de célula morta, vermelho). (B) Células HeLa cultivadas em uma grade não padronizada e manchadas como em A. (C) Projeção de pilhas z confocal de uma célula HeLa em uma grade Padrão Quantifoil R2/2 com colágeno de 0,01 mg/mL e fibrinogen 647 ECM (vermelho). A célula estava manchada com calcein-AM (verde) e Hoechst-33342 (azul). (D) Células HeLa em grade sem valor incubadas com colágeno de 0,01 mg/mL e fibrinogeno 647 ECM, incubadas e manchadas com calcein-AM e Hoecsht-33342. As imagens fluorescentes foram fundidas com luz transmitida (escala de cinza). (E) X, Z projeção de C. (F) X, Z projeção de D. As imagens são pseudocoloridas. As barras de escala em (A) e (B) são de 500 μm; barras de escala em (C) - (F) são 10 μm. Por favor clique aqui para ver uma versão maior desta figura.
{kind=link}

Figura 5: Cryo-ET da célula BEAS-2B infectada por RSV na grade crio-TEM padronizada. (A) Mapa quadrado da grade Cryo-EM da célula BEAS-2B infectada por RSV. O limite aproximado da célula é indicado pela linha verde tracejada. (B) Imagem de maior resolução da área encaixotado em vermelho em (A). O limite aproximado da célula é indicado pela linha verde tracejada. Virions RSV podem ser vistos perto da periferia celular (seta branca e caixa amarela). (C) Única fatia z do tomograma coletada na área da caixa amarela em (B). Setas vermelhas apontam para a proteína de fusão RSV F, setas azuis apontam para o complexo de ribonucleoproteína (RNP). As barras de escala em (A)-(C) estão embutidas na imagem. Clique aqui para ver uma versão maior desta figura.
{kind=link}

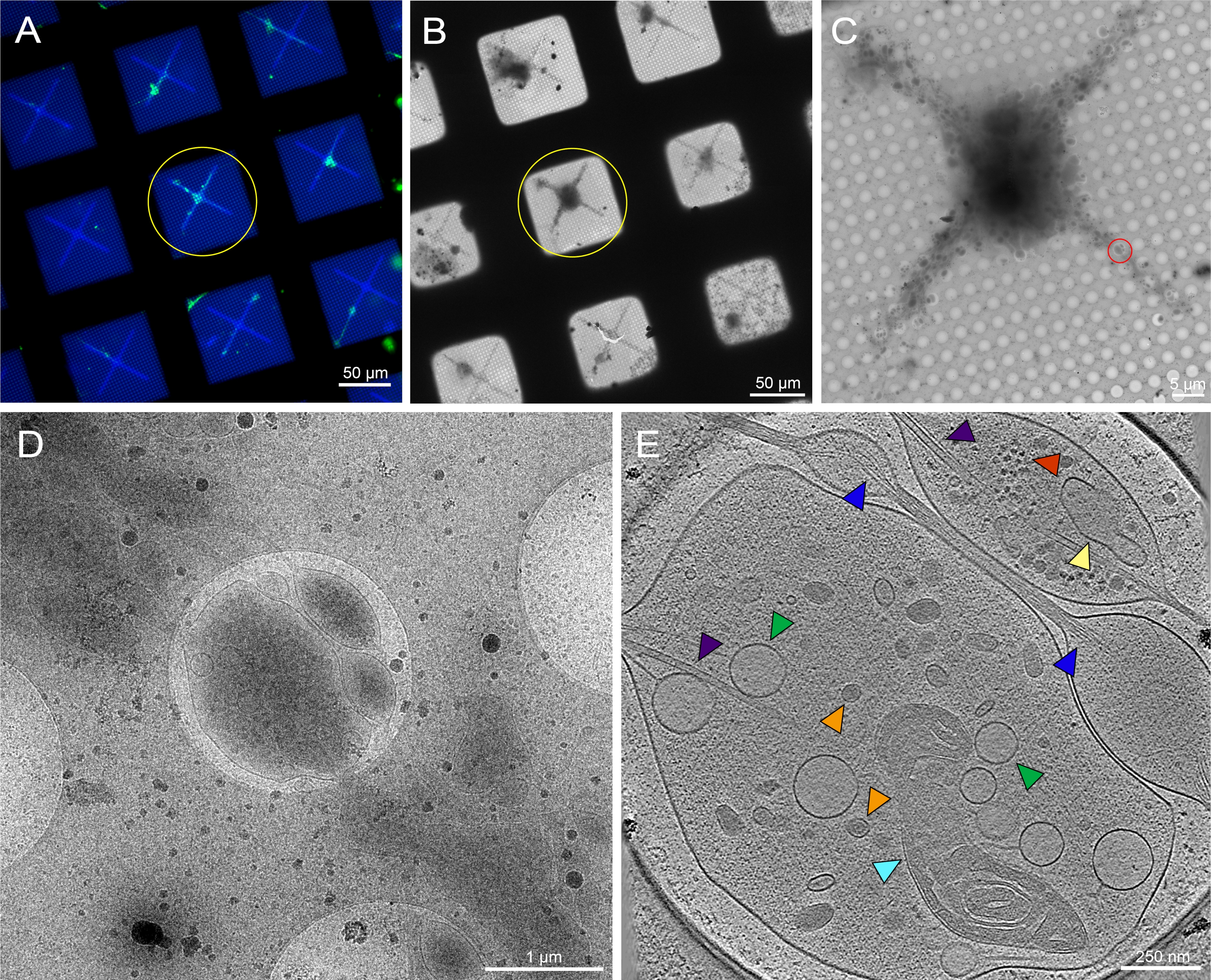
Figura 6: Neurônios primários derivados do cérebro da 3ª instar Drosophila melanogaster larvas na grade crio-TEM padronizada. (A) Montagem da rede de microscopia de fluorescência de células vivas sobrepostas de neurônios Drosophila expressando GFP com direção de membrana em quadrados de grade padronizados com concanavalina fluorescente de 0,5 mg/mL A. Verde: neurônios de Drosophila . Azul: Photopattern. (B) montagem de imagem cryo-EM da grade em (A) após criopreservação. O círculo amarelo observa o mesmo quadrado de grade que em (A). (C) montagem de imagem cryo-EM do quadrado destacada pelo círculo amarelo em (A) e (B). (D) Imagem de ampliação mais elevada da área delimitada pelo círculo vermelho em (C), onde uma série de inclinação foi coletada nos neurites da célula. E. 25 nm de espessura fatia de um tomograma reconstruído a partir da série de inclinação que foi adquirida do círculo vermelho em (C). Várias organelas podem ser vistas neste tomograma, como as mitocôndrias (ciano), microtúbulos (roxo), vesículas de núcleo densa (laranja), vesículas leves (verde), o réticulo endoplasmático (amarelo) e actina (azul). Macromoléculas, como ribossomos (vermelho), também podem ser vistas no canto superior direito. Imagens fluorescentes são pseudocoloridas. As barras de escala em (A)-(E) estão embutidas na imagem. Clique aqui para ver uma versão maior desta figura.
{kind=link}

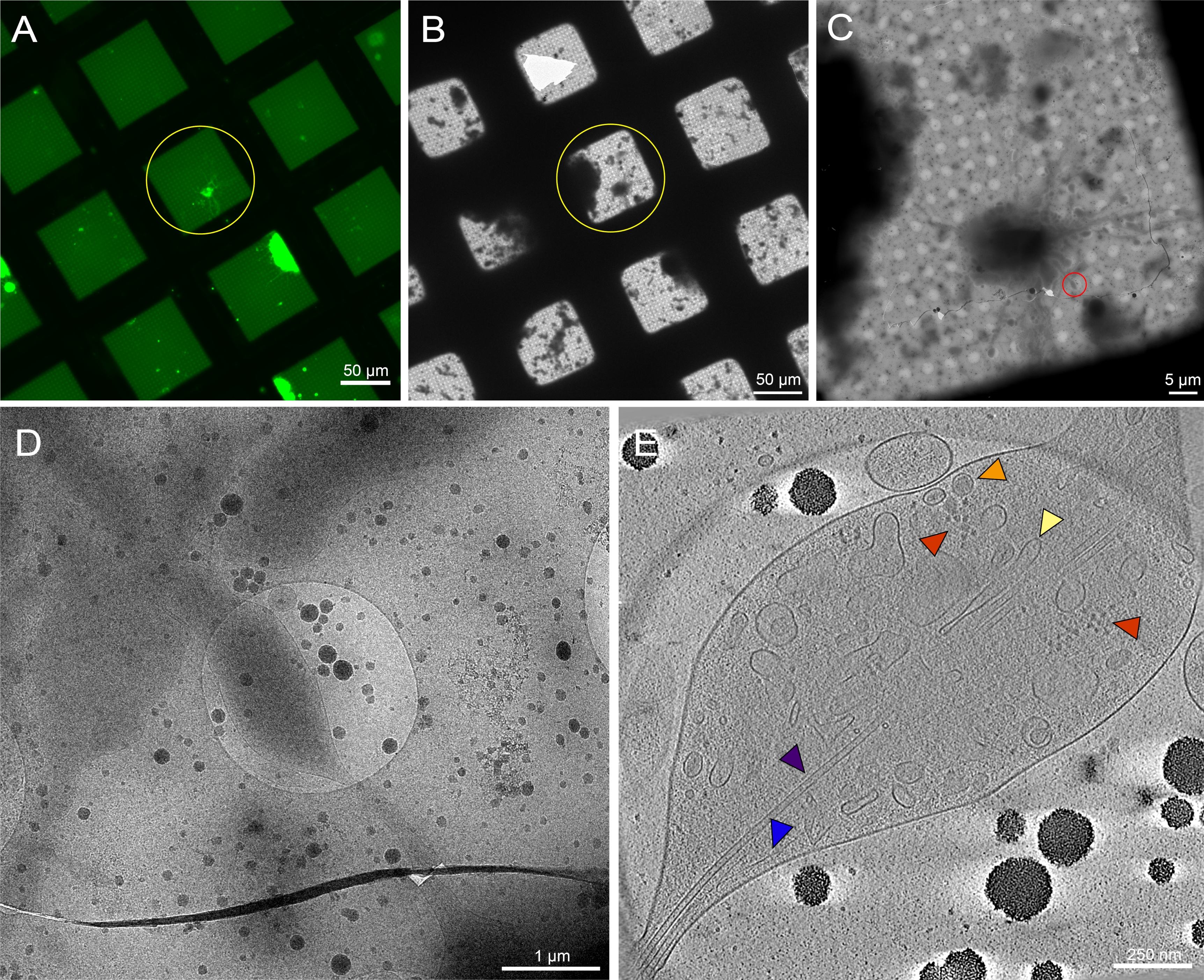
Figura 7: Neurônios primários derivados do cérebro da 3ª instar Drosophila melanogaster larvas em redes não respingadas. (A) Montagem da rede de microscopia de fluorescência de células vivas de neurônios Drosophila expressando GFP direcionado à membrana em quadrados de grade com concanavalina de 0,5 mg/mL A. Verde: neurônios de drosophila . (B) montagem da grade Cryo-EM da mesma grade em (A) após o congelamento do mergulho. O círculo amarelo mostra a mesma grade quadrada que em (A). Observe a presença de detritos celulares e contaminação da mídia, o que dificultou a identificação do alvo em comparação com as grades padronizadas. (C) montagem de imagem cryo-EM da praça destacada pelos círculos amarelos nos mapas (A) e (B). (D) Imagem de ampliação mais elevada da área delimitada pelo círculo vermelho em (C), onde uma série de inclinação foi coletada nos neurites da célula. (E) 25 nm de espessura da fatia reconstruída do tomograma reconstruído da série de inclinação de (C) e (D). Uma série de organelas são visíveis neste tomograma, como microtúbulos (roxo), actina (azul), réticulo endoplasmático (amarelo) e vesículas de núcleo densa (laranja). Macromoléculas, como ribossomos (vermelho), também podem ser vistas. Imagens fluorescentes são pseudocoloridas. As barras de escala em (A)-(E) estão embutidas na imagem. Clique aqui para ver uma versão maior desta figura.
{kind=link}

Figura 8: Exemplos de possíveis problemas com a padronização. Imagens fluorescentes de ECM rotulado depositados em grades micropatteradas. (A) Padronização desigual em toda a rede devido à distribuição desigual do gel PLPP. (B) O ECM não pode aderir às áreas abrangidas pelo estêncil PDMS durante a padronização. (C) Padrão de gradiente saturado (lado direito) ou padrão invertido (esquerda) em uma grade padronizada com dose total muito alta. (D) O ECM está aderindo a áreas nas barras de grade, bem como área padronizada devido a reflexos do laser UV durante a padronização. As imagens são pseudocoloridas; padrão de entrada é mostrado no inferior esquerdo; barras de escala são 100 μm. Por favor clique aqui para ver uma versão maior desta figura.
{kind=link}
| Questão | Causa potencial(s) | Solucionando problemas |
| Micropatterning | ||
| Não é possível ver a iluminação do laser PRIMO | • O caminho da luz não está configurado corretamente | • Verifique se o caminho da luz do microscópio está configurado corretamente |
| • O laser PRIMO não está ligado ou o laser está entrelaçado | ||
| Muitos quadrados de grade quebrados | • Tocar a folha da grade com pinças ou tubulações durante o manuseio | • Manuseie grades com cuidado |
| • Grade seca durante incubações ou lavagem | • Não permita que a grade seque durante lavagens e incubações | |
| Grandes áreas não respingadas | • Cobertura de gel insuficiente | • Certifique-se de que o gel se espalhe uniformemente sobre a grade ao adicionar |
| • Folha da grade fora de foco durante a padronização | • Adicione um microliter adicional de gel | |
| • Área coberta por estêncil | • Verifique o foco antes de padronizar cada região | |
| • Rede cuidadosamente centralizada em estêncil | ||
| Padrão saturado ou invertido | • Dose incorreta | • Experimente uma série de doses totais para o padrão |
| • Cobertura de gel insuficiente | • Certifique-se de que a grade esteja coberta uniformemente com gel | |
| • Experimente valores diferentes para padrões em escala de cinza | ||
| Padrão embaçado | • Mau foco durante a padronização | • Repita a calibração primo na mesma altura da amostra |
| • Calibração incorreta | • Foco na folha de grade antes da padronização | |
| • Dividir o padrão em regiões adicionais para padronização | ||
| ECM aderindo fora do padrão | • Reflexos de gel ou poeira | • Certifique-se de que o gel está seco antes da padronização |
| • Certifique-se de que a tampa e as lentes objetivas estejam limpas | ||
| ECM não visível após a padronização | • Branqueamento de fotos | • Minimizar a exposição à luz ao ECM antes da imagem |
| • Dose incorreta durante a padronização | • Experimente uma gama de valores totais de dose para padrão | |
| • Tempo insuficiente de incubação do ECM | • Aumentar o tempo de incubação do ECM | |
| Semeadura celular | ||
| Células desajeitadas | • Sobre a digestão | • Use menor percentual de trippsina ou tempo para liberação de células aderentes |
| • Alta densidade celular | • Passagem e/ou digestão de células com menor confluência | |
| • Não agitar células durante a liberação | ||
| • Solução de células de tubulação suave ou usar filtros de células | ||
| Células que não aderiram a áreas padronizadas | • O ECM não é adequado para o tipo celular | • Experimente diferentes concentrações e composição de ECM |
| • A viabilidade das células é diminuída antes da semeadura | • Garantir que a cultura celular e as condições de liberação celular não estejam danificando as células | |
| Células não se expandem após a adesão | • ECM ou padrão inadequado para tipo de célula | • Experimente padrões diferentes e ECM |
| • Em alguns casos, uma folha mais contínua (R1.2/20 vs R2/1) pode promover a expansão celular |
Tabela 1: Problemas potenciais durante a micropatterning. Esta tabela descreve alguns problemas que um usuário pode experimentar durante a micropatterning ou semeadura de células. Possíveis causas e solução de problemas são fornecidas para cada problema. Imagens representativas de alguns problemas podem ser vistas na Figura 8.
Discussão
Microscópios eletrônicos modernos e avançados e pacotes de software agora suportam a coleta automatizada de dados crio-EM e crio-ET, onde centenas a milhares de posições podem ser direcionadas e imagens dentro de alguns dias32,33,34,35. Um fator limitante significativo para fluxos de trabalho crio-ET de células inteiras tem obtido um número suficiente de metas colecionáveis por rede. Recentemente, vários grupos desenvolveram protocolos para redes de micropatterning para crio-EM, com uma vantagem sendo a eficiência da coleta de dados 16,17,18. Aqui, um protocolo é apresentado para o uso de um sistema de micropatterning comercialmente disponível para micropattern redes TEM para estudos crio-ET de neurônios primários de Drosophila e linhas de células humanas cultivadas (não infectadas ou infectadas pelo RSV). Este sistema de micropatterning é versátil e muitos passos podem ser otimizados e adaptados para atender a objetivos experimentais específicos. Um usuário com experiência em microscopia de tem e fluorescência pode rapidamente se tornar qualificado na preparação da grade e micropatterning. Com uma prática cuidadosa, bons resultados devem ser alcançáveis após algumas iterações. Abaixo, algumas das opções disponíveis, considerações do usuário, benefícios potenciais e aplicações futuras de micropatterning para crio-EM são discutidas.
Uma das considerações importantes para o crio-ET de células inteiras é a seleção da grade EM. As grades EM são compostas de duas partes: uma estrutura de malha (ou suporte estrutural) e a folha (ou filme), que é a superfície de filme contínua ou furada na qual as células crescerão. As grades de malha de cobre são comumente usadas para crio-EM de proteínas e complexos isolados. No entanto, eles são inadequados para crio-ET de células inteiras devido à citotoxicidade do cobre. Em vez disso, uma malha de ouro é comumente usada para tomografia celular. Outras opções incluem níquel ou titânio, que pode proporcionar benefícios sobre o ouro, como o aumento da rigidez16. As grades EM estão disponíveis com diferentes dimensões de malha para suportar uma variedade de aplicativos. Tamanhos de malha maiores fornecem mais espaço para as células crescerem entre barras de grade e mais áreas que são favoráveis à coleta de séries de inclinação, embora ao custo do aumento da fragilidade geral dos espécimes. A folha mais usada é carbono amorfo perfurado ou furado, como quantifoils ou grades C-flat. Alvos biológicos podem ser imagens através dos buracos no carbono ou através do carbono translúcido de elétrons. Grades como R 2/1 ou R 2/2, onde os orifícios têm 2 μm de largura que são espaçados 1 e 2 μm de distância, respectivamente, fornecem um grande número de buracos e, portanto, um grande número de áreas potenciais para coleta de dados. No entanto, algumas células podem crescer e expandir melhor em superfícies mais uniformes, como redes R 1,2/20 ou carbono contínuo. Para o processamento da amostra a jusante por moagem de feixe de íons focados (crio-FIB), a folha é removida através da fresagem, reduzindo as preocupações sobre a presença contínua do filme subjacente. Assim como na malha, também estão disponíveis folhas de outros materiais, com o protocolo de padronização aqui apresentado sendo igualmente adequado para grades SiO2 . As grades comumente usadas incluem quantifoil de ouro, carbono contínuo ou grades de 200 malhas de filme SiO2 (~90 μm espaçamento entre grades) para crio-ET de células inteiras.
Há uma série de considerações ao projetar um padrão. A maioria dessas decisões são guiadas pelo tipo celular e propósito do experimento. Um bom ponto de partida é escolher um padrão que se aproxime da forma e dimensões das células na cultura. Muitos estudos demonstraram efeitos significativos da forma padrão no crescimento celular e arranjo citoesquelético13,36,37. Deve-se tomar cuidado especial durante o projeto de padrão se isso pode alterar o alvo de interesse. Vários padrões para cada tipo de célula foram testados para determinar quais padrões promoveram a adesão e o crescimento celular. A flexibilidade do sistema de micropatterning permite o teste de vários padrões em uma única grade e a mudança de padrões para diferentes grades dentro de um único experimento. Padrões maiores (~50-90 μm), como os usados aqui, aumentam a probabilidade de que várias células aderam a uma única região do padrão e permitem que as células se expandam e se estendam após a adesão. Padrões mais restritos (20-30 μm) podem ser apropriados em experimentos onde o isolamento celular é mais crítico do que a expansão celular, como para experimentos de moagem de feixe de íons focados (crio-FIB). Para aplicações de tomografia, pode-se precisar considerar o impacto do eixo de inclinação. Se um padrão for posicionado de tal forma que todas as células cresçam paralelamente umas às outras em uma única direção, é possível que todas as células sejam perpendiculares ao eixo de inclinação quando carregadas no estágio do microscópio, resultando em uma menor qualidade de dados.
Em grades não respingadas, as células geralmente aderem preferencialmente às barras de grade, onde não podem ser imagens pelo TEM. Mesmo em grades padronizadas, as células são frequentemente observadas para serem posicionadas nos cantos dos quadrados de grade parcialmente tanto na folha de carbono padronizada quanto na barra de grade. Recentemente, o micropatterning foi usado para posicionar intencionalmente parte da célula sobre a barra de grade18. Isso pode ser considerado para experimentos onde não é crítico ter toda a periferia celular na folha. Isso pode ser especialmente importante para células que podem crescer maiores que um único quadrado de grade, como neurônios primários crescendo ao longo de vários dias.
Existem muitas ferramentas que podem ser usadas para projetar um padrão. Aqui, o padrão foi limitado a menos de 800 pixels em qualquer dimensão, de modo que o padrão pode ser girado para qualquer ângulo e ainda se encaixar dentro da área máxima que pode padronizar em uma única projeção por este sistema de micropatterning. Isso permite que o usuário gire o padrão para ser devidamente orientado com a grade, independentemente da orientação da grade no microscópio. Aqui, a grade foi dividida em seis áreas de padronização. Em primeiro parte, isso permite o ajuste de foco entre diferentes regiões da rede. As grades de ouro, em particular, são muito maleáveis e podem não se deitar completamente planas no vidro. O foco adequado é essencial para resultados de padronagem limpos e refinados. Ao utilizar padrões segmentados, apenas pequenos ajustes na posição padrão precisam ser feitos se a grade mudar ligeiramente durante o processo de padronização, embora isso geralmente não seja um problema ao usar o gel PLPP com os estêncil PDMS. Finalmente, as quatro praças centrais da grade permaneceram sem ser respingadas. Isso suporta que um usuário seja capaz de identificar claramente o centro da rede, o que é muito útil para experimentos de imagem correlativa.
O software de padronização para este sistema de micropatterning, Leonardo, também tem características mais avançadas, como costura e a capacidade de importar padrões como PDFs, que estão além do escopo deste protocolo. Este software também inclui detecção de microestruturas e posicionamento automatizado de padrões que podem ser usados em grades TEM. Este recurso é mais útil quando a grade é muito plana e pode ser padronizada sem a necessidade de ajustar o foco entre diferentes áreas.
A seleção de uma proteína ECM pode ter um impacto significativo na adesão e expansão celular. Algumas células são conhecidas por sofrer alterações fisiológicas quando cultivadas em substratos específicos38. Várias proteínas e concentrações de ECM foram testadas para qualquer novo tipo de célula com base em trabalhos anteriores relatados na literatura. Laminina, fibrinogênio, fibronectina e colágeno são amplamente utilizados para células cultivadas e podem ser usados como ponto de partida se outros dados não estiverem disponíveis. No entanto, outras proteínas ECM também devem ser consideradas se as proteínas ECM comumente utilizadas não conferem propriedades adequadas de adesão para as células. Isso foi particularmente verdadeiro para os neurônios primários de Drosophila , uma vez que a alta concentração da lectina vegetal concanavalina A era necessária para a adequada adesão celular. A compatibilidade da adesão celular e o crescimento com o ECM podem ser testados por padronização em pratos de vidro ou slides antes da transição para grades TEM. Essa abordagem de pré-triagem é tempo e custo-benefício se um grande número de combinações precisar ser examinada. A inclusão de uma proteína ECM conjugada fluorescente é valiosa para avaliar o sucesso e a qualidade da padronização.
A semeadura celular é um dos passos mais importantes para crio-ET celular inteiro, com ou sem micropatterning6,16,39. Para drosophila primária ou outros neurônios, que são frágeis, instáveis em suspensão, e podem ser limitados em quantidade, abordagens únicas de semeadura são preferidas em vez de sequenciais e monitoradas. Um único passo de semeadura em uma densidade celular otimizada, como descrito no protocolo para neurônios Drosophila, é uma opção viável para a maioria dos tipos de células. No entanto, também é possível semear células no substrato em uma concentração inicial mais baixa e adicionar mais células de forma monitorada como descrito aqui e em outras literaturas18. Esta sequencial sequencial pode proporcionar resultados mais consistentes em alguns casos. Semelhante à cultura celular padrão, deve-se sempre ter cuidado para manter a viabilidade celular e minimizar a aglomeração celular durante o isolamento.
Quando começam com a micropattering, existem algumas armadilhas potenciais que são prejudiciais para o resultado final. O manuseio cuidadoso da grade e a técnica estéril, a distribuição uniforme do gel PLPP, a dose adequada e o foco durante a padronização e a manutenção da viabilidade celular antes da semeadura estão entre as considerações mais importantes para o sucesso. Uma lista de alguns dos problemas potenciais, bem como soluções foram montadas na Tabela 1.
Redes micropatriadas podem ser usadas para ajudar a posicionar as células para estabelecer uma densidade celular consistente em toda a rede e para posicionar regiões de interesse em áreas adequadas para a coleta de séries de inclinação16,18. A colocação e o posicionamento das células podem ser usados como marcadores fiduciais para correlação em experimentos crio-CLEM, reduzindo a necessidade de frágeis grades de localizador e marcadores fiduciais fluorescentes. No entanto, deve-se notar que tais marcadores fiduciários ainda podem ser úteis para a correlação de precisão de subdultímetro29,40. Além disso, uma distribuição uniforme de células isoladas também é altamente benéfica para experimentos de moagem de feixe de íons focados (crio-FIB) para maximizar o número de células das quais a lamella pode ser cortada16.
A adição de micropatterning aos fluxos de trabalho crio-EM resultará em melhorias mensuráveis no throughput de dados e potencialmente permitirá novos experimentos. À medida que a técnica é ainda mais adotada e desenvolvida, aplicações mais avançadas de micropatterning, incluindo gradientes de ECM, deposições múltiplas de ECM e montagem de microestruturas expandirão ainda mais as capacidades do crio-ET para estudar metas e processos biológicos em pleno contexto celular.
Divulgações
Os autores não têm nada a revelar.
Agradecimentos
Agradecemos ao Dr. Jill Wildonger, Dr. Sihui Z. Yang e Sra. Josephine W. Mitchell no Departamento de Bioquímica da Universidade de Wisconsin, Madison por compartilhar generosamente a cepa de mosca elav-Gal4, UAS-CD8::GFP (Bloomington stock center, #5146). Também gostaríamos de agradecer ao Dr. Aurélien Duboin, Ao Sr. Laurent Siquier e à Sra. Marie-Charlotte Manus de Alvéole e ao Sr. Serge Kaddoura da Nanoscale Labs pelo seu generoso apoio durante este projeto. Este trabalho foi apoiado em parte pela Universidade de Wisconsin, Madison, o Departamento de Bioquímica da Universidade de Wisconsin, Madison, e o serviço público de saúde concede R01 GM114561, R01 GM104540, R01 GM104540-03W1 e U24 GM139168 a E.R.W. e R01 AI150475 a P.W.S. do NIH. Parte desta pesquisa foi apoiada pela bolsa NIH U24 GM129547 e realizada na PNCC da OHSU e acessada através da EMSL (grid.436923.9), um DoE Office of Science User Facility patrocinado pelo Escritório de Pesquisa Biológica e Ambiental. Também somos gratos pelo uso de instalações e instrumentação no Cryo-EM Research Center no Departamento de Bioquímica da Universidade de Wisconsin, Madison.
Materiais
| Name | Company | Catalog Number | Comments |
| 0.1% (w/v) Poly-L-Lysine | Sigma | P8920-100ML | |
| 0.22 µm syringe filters PVDF membrane | Genesee | 25-240 | |
| 22x60-1 Glass cover slip | Fisher | 12545F | |
| 5/15 Tweezers | EMS (Dumont) | 0203-5/15-PO | |
| Antibiotic-Antimycotic (100X) | ThermoFisher (Gibco) | 15240096 | |
| BEAS-2B cells | ATCC | CRL-9609 | |
| Collagen I, bovine | ThermoFisher (Gibco) | A1064401 | |
| Concanavalin A, Alexa Fluor 350 Conjugate | ThermoFisher (Invitrogen) | C11254 | |
| DMEM | Fisher (Lonza) | BW12-604F | |
| EtOH | Fisher (Decon Labs) | 22-032-600 | |
| Fetal Bovine Serum | ATCC | 30-2020 | |
| Fibrinogen From Human Plasma, Alexa Fluor 647 Conjugate | ThermoFisher (Invitrogen) | F35200 | |
| Fibronectin Bovine Protein, Plasma | ThermoFisher (Gibco) | 33010018 | |
| Glass bottom dish | MatTek | P35G-1.5-20-C | |
| Glucose | VWR | 0643-1KG | |
| Grid prep holder | EMS | 71175-01 | |
| HeLa cells | ATCC | CCL-2 | |
| Hemacytometer | Fisher (SKC, Inc.) | 22600100 | |
| HEPES | Fisher (ACROS Organics) | AC172572500 | |
| Hoechst 33342 | ThermoFisher (Invitrogen) | H3570 | |
| Insulin | Fisher (Sigma Aldrich) | NC0520015 | |
| KCl | MP Bio | 194844 | |
| KH2PO4 | Fisher (ACROS Organics) | AC212595000 | |
| Leica-DMi8 | Leica Microsystems | Can be customized with camera, stage, and objective attachments | |
| Leonardo | Alvéole | https://www.alveolelab.com/our-products/leonardo-photopatterning-software/ | |
| Liberase Research Grade | Fisher (Supply Solutions) | 50-100-3280 | |
| LIVE/DEAD Viability/Cytotoxicity Kit | ThermoFisher (Invitrogen) | L3224 | |
| Microscope camera | Hammamatsu | C13440-20CU | |
| Motorized stage | Märzhäuser Wetzlar | 00-24-599-0000 | |
| NaCl | Fisher (Fisher BioReagents) | BP358-1 | |
| NaH2PO4 | Fisher (ACROS Organics) | AC207802500 | |
| NaOH | Fisher (Alfa Aesar) | AAA1603736 | |
| PBS | Corning | 21-040-CV | |
| PDMS stencils | nanoscaleLABS | PDMS_STENCILS_EM | https://www.alveolelab.com/our-products/pdms-stencil-multiwell-plate/ |
| PEG-SVA | nanoscaleLABS | PEG-SVA-1GR | mPEG-Succinimidyl Valerate, MW 5,000 |
| Penicillin | Fisher (Research Products International Corp) | 50-213-641 | |
| pH strips | Fisher (Millipore Sigma) | M1095350001 | pH probe can also be used |
| PLPP gel | nanoscaleLABS | PLPP-GEL-300UL | https://www.alveolelab.com/our-products/plpp-photoactivatable-reagent/ |
| PRIMO | Alvéole | https://www.alveolelab.com/our-products/primo-micropatterning/ | |
| pSynkRSV-I19F (BAC containing RSV A2-mK+ antigenomic cDNA ) | BEI Resources | NR-36460 | https://www.beiresources.org/Catalog/BEIPlasmidVectors/NR-36460.aspx |
| Quantifoil grids | EMS (Quantifoil) | Q2100AR1 | 2 µm holes spaced 1 µm apart, other dimensions are available |
| RPMI | Fisher (Lonza) | BW12-702F | |
| RSV A2-mK+ | see entry for pSynkRSV-19F | - | Described in Hotard et al. [22]. Can be generated from pSynkRSV-ll9F |
| Schneider's Media | ThermoFisher (Gibco) | 21720-024 | |
| SerialEM | SerialEM (https://bio3d.colorado.edu/SerialEM/ ) | https://bio3d.colorado.edu/SerialEM/ | |
| Straight tweezers | EMS (Dumont) | 72812-D | |
| Streptomycin | Fisher (Fisher BioReagents) | BP910-50 | |
| Sucrose | Avantor | 4097-04 | |
| Tetracycline | Sigma | T8032-10MG | |
| Titan Krios electron microscope | ThermoFisher | 300kV, with direct electron detector camera and energy filter | |
| Trypsin | ThermoFisher (Gibco) | 15090046 | |
| Tube Revolver/Rotator | Fisher (Thermo Scientific) | 11676341 | |
| UAS:mcD8:GFP Drosophila fly strain | Bloomington Drosophila Stock Center | 5146 | http://flybase.org/reports/FBtp0002652.html |
Referências
- Nogales, E., Scheres, S. H. Cryo-EM: A unique tool for the visualization of macromolecular complexity. Molecular Cell. 58 (4), 677-689 (2015).
- Martynowycz, M. W., Gonen, T. From electron crystallography of 2D crystals to MicroED of 3D crystals. Current Opinion in Colloid and Interface Science. 34, 9-16 (2018).
- Wagner, J., Schaffer, M., Fernandez-Busnadiego, R. Cryo-electron tomography-the cell biology that came in from the cold. FEBS Letters. 591 (17), 2520-2533 (2017).
- Wan, W., Briggs, J. A. Cryo-electron tomography and subtomogram averaging. Methods in Enzymology. 579, 329-367 (2016).
- Bäuerlein, F. J., Pastor-Pareja, J. C., Fernández-Busnadiego, R. Cryo-electron tomography of native Drosophila tissues vitrified by plunge freezing. bioRxiv. , 437159(2021).
- Hampton, C. M., et al. Correlated fluorescence microscopy and cryo-electron tomography of virus-infected or transfected mammalian cells. Nature Protocols. 12 (1), 150-167 (2017).
- Hsieh, C. E., Leith, A., Mannella, C. A., Frank, J., Marko, M. Towards high-resolution three-dimensional imaging of native mammalian tissue: Electron tomography of frozen-hydrated rat liver sections. Journal of Structural Biology. 153 (1), 1-13 (2006).
- Al-Amoudi, A., Norlen, L. P., Dubochet, J. Cryo-electron microscopy of vitreous sections of native biological cells and tissues. Journal of Structural Biolology. 148 (1), 131-135 (2004).
- Rigort, A., et al. Focused ion beam micromachining of eukaryotic cells for cryoelectron tomography. Proceedings of the National Academy of Sciences. 109 (12), 4449-4454 (2012).
- Gorelick, S., et al. PIE-scope, integrated cryo-correlative light and FIB/SEM microscopy. Elife. 8, 45919(2019).
- Wu, G. H., et al. Multi-scale 3D cryo-correlative microscopy for vitrified cells. Structure. 28 (11), 1231-1237 (2020).
- Turk, M., Baumeister, W. The promise and the challenges of cryo-electron tomography. FEBS Letters. 594 (20), 3243-3261 (2020).
- Théry, M. Micropatterning as a tool to decipher cell morphogenesis and functions. Journal of Cell Science. 123 (24), 4201-4213 (2010).
- Tseng, Q., et al. Spatial organization of the extracellular matrix regulates cell-cell junction positioning. Proceedings of the National Academy of Sciences. 109 (5), 1506-1511 (2012).
- Hardelauf, H., et al. Micropatterning neuronal networks. Analyst. 139 (13), 3256-3264 (2014).
- Toro-Nahuelpan, M., et al. Tailoring cryo-electron microscopy grids by photo-micropatterning for in-cell structural studies. Nature Methods. 17 (1), 50-54 (2020).
- Engel, L., et al. Extracellular matrix micropatterning technology for whole cell cryogenic electron microscopy studies. Journal of Micromechanics and Microengineering. 29 (11), (2019).
- Engel, L., et al. Lattice micropatterning for cryo-electron tomography studies of cell-cell contacts. bioRxiv. , 272237(2021).
- Sibert, B. S., Kim, J. Y., Yang, J. E., Wright, E. R. Whole-cell cryo-electron tomography of cultured and primary eukaryotic cells on micropatterned TEM grids. bioRxiv. , 447251(2021).
- Egger, B., van Giesen, L., Moraru, M., Sprecher, S. G. In vitro imaging of primary neural cell culture from Drosophila. Nature Protocols. 8 (5), 958-965 (2013).
- Lu, W., Lakonishok, M., Gelfand, V. I. Kinesin-1-powered microtubule sliding initiates axonal regeneration in Drosophila cultured neurons. Molecular Biology of the Cell. 26 (7), 1296-1307 (2015).
- Ke, Z., et al. The morphology and assembly of respiratory syncytial virus revealed by cryo-electron tomography. Viruses. 10 (8), (2018).
- Stobart, C. C., et al. A live RSV vaccine with engineered thermostability is immunogenic in cotton rats despite high attenuation. Nature Communications. 7, 13916(2016).
- Hotard, A. L., et al. A stabilized respiratory syncytial virus reverse genetics system amenable to recombination-mediated mutagenesis. Virology. 434 (1), 129-136 (2012).
- Schindelin, J., et al. Fiji: an open-source platform for biological-image analysis. Nature Methods. 9 (7), 676-682 (2012).
- Mastronarde, D. N. Automated electron microscope tomography using robust prediction of specimen movements. Journal of Structural Biology. 152 (1), 36-51 (2005).
- Kremer, J. R., Mastronarde, D. N., McIntosh, J. R. Computer visualization of three-dimensional image data using IMOD. Journal of Structural Biology. 116 (1), 71-76 (1996).
- Tang, G., et al. EMAN2: an extensible image processing suite for electron microscopy. Journal of Structural Biology. 157 (1), 38-46 (2007).
- Yang, J. E., Larson, M. R., Sibert, B. S., Shrum, S., Wright, E. R. CorRelator: Interactive software for real-time high precision cryo-correlative light and electron microscopy. Journal of Structural Biology. , 107709(2021).
- Ke, Z., et al. Promotion of virus assembly and organization by the measles virus matrix protein. Nature Communications. 9 (1), 1736(2018).
- Kim, J., Yang, S., Wildonger, J., Wright, E. A new in situ neuronal model for cryo-ET. Microscopy and Microanalysis. 26 (2), 130-132 (2020).
- Bouvette, J., et al. Beam image-shift accelerated data acquisition for near-atomic resolution single-particle cryo-electron tomography. Nature Communications. 12 (1), 1957(2021).
- Schorb, M., Haberbosch, I., Hagen, W. J. H., Schwab, Y., Mastronarde, D. N. Software tools for automated transmission electron microscopy. Nature Methods. 16 (6), 471-477 (2019).
- Weis, F., Hagen, W. J. H., Schorb, M., Mattei, S. Strategies for optimization of cryogenic electron tomography data acquisition. Journal of Visual Experiments. (169), e62383(2021).
- Chreifi, G., Chen, S., Jensen, G. J. Rapid tilt-series method for cryo-electron tomography: Characterizing stage behavior during FISE acquisition. Journal of Structural Biology. 213 (2), 107716(2021).
- Anderson, D. E., Hinds, M. T. Endothelial cell micropatterning: methods, effects, and applications. Annals of Biomedical Engineering. 39 (9), 2329-2345 (2011).
- McWhorter, F. Y., Wang, T., Nguyen, P., Chung, T., Liu, W. F. Modulation of macrophage phenotype by cell shape. Proceedings of the National Academy of Sciences. 110 (43), 17253-17258 (2013).
- Kleinman, H. K., Luckenbill-Edds, L., Cannon, F. W., Sephel, G. C. Use of extracellular matrix components for cell culture. Analytical Biochemistry. 166 (1), 1-13 (1987).
- Fassler, F., Zens, B., Hauschild, R., Schur, F. K. M. 3D printed cell culture grid holders for improved cellular specimen preparation in cryo-electron microscopy. Journal of Structural Biology. 212 (3), 107633(2020).
- Schellenberger, P., et al. High-precision correlative fluorescence and electron cryo microscopy using two independent alignment markers. Ultramicroscopy. 143, 41-51 (2014).
Reimpressões e Permissões
Solicitar permissão para reutilizar o texto ou figuras deste artigo JoVE
Solicitar PermissãoThis article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. Todos os direitos reservados