
Method Article
רשתות מיקרוסקופיות אלקטרוניות של שידור מיקרו-מפזרנינג למיקום תאים ישירים בתהליכי עבודה של טומוגרפיה קריו-אלקטרונים של תאים שלמים
* These authors contributed equally
In This Article
Summary
מטרת פרוטוקול זה היא לכוון הידבקות תאים וצמיחה לאזורים ממוקדים של רשתות למיקרוסקופיה קריו-אלקטרונית. זה מושג על ידי החלת שכבה נגד עכירות כי הוא ablated בדפוסים שצוינו על ידי המשתמש ואחריו תצהיר של חלבוני מטריצה חוץ תאית באזורים בדוגמת לפני זריעת התא.
Abstract
טומוגרפיה של קריו-אלקטרון של תאים שלמים (cryo-ET) היא טכנולוגיה רבת עוצמה המשמשת לייצור מבני רזולוציה ברמת ננומטר של מקרומולקולות הנמצאות בהקשר התאי ונשמרות במצב כמעט מקורי של לחות קפואה. עם זאת, ישנם אתגרים הקשורים פולחן ו /או הדבקת תאים ברשתות TEM באופן המתאים טומוגרפיה תוך שמירה על התאים במצבם הפיזיולוגי. כאן, פרוטוקול מפורט צעד אחר צעד מוצג על השימוש micropatterning כדי לכוון ולקדם צמיחת תאים אאוקריוטיים על רשתות TEM. במהלך micropatterning, צמיחת התא מכוונת על ידי הפקדת חלבוני מטריצה חוץ תאית (ECM) בתוך דפוסים ומיקומים שצוינו על רדיד הכסף של רשת TEM בעוד האזורים האחרים נשארים מצופים בשכבה נגד עכירות. גמישות בבחירת ציפוי פני השטח ועיצוב התבניות הופכת את המיקרו-מפזרינג ליישום רחב עבור מגוון רחב של סוגי תאים. Micropatterning שימושי עבור מחקרים של מבנים בתוך תאים בודדים, כמו גם מערכות ניסיוניות מורכבות יותר כגון אינטראקציות מארח-פתוגן או קהילות רב תאיות מובחנות. Micropatterning עשוי גם להיות משולב בזרימות עבודה רבות במורד הזרם של קריו-ET של תאים מלאים, כולל אור מתאם ומיקרוסקופיה אלקטרונית (קריו-CLEM) וכרסום קרן יונים ממוקדת (cryo-FIB).
Introduction
עם ההתפתחות, ההתרחבות והרב-תכליתיות של מיקרוסקופיה קריו-אלקטרונית (cryo-EM), החוקרים בחנו מגוון רחב של דגימות ביולוגיות במצב כמעט מקומי ממקרומולקולרי (~ 1 ננומטר) ועד רזולוציה גבוהה (~ 2 Å). טכניקות הקפאה-EM ועקיפת אלקטרונים של חלקיקים בודדים מוחלות בצורה הטובה ביותר על מקרומולקולות מטוהרות בתמיסה או במצב גבישי, בהתאמה 1,2. בעוד טומוגרפיה קריו-אלקטרונים (קריו-ET) מתאימה באופן ייחודי למחקרים מבניים ואולטרה-מבניים כמעט מקומיים של עצמים הטרולוגיים גדולים כגון חיידקים, וירוסים פלומורפיים ותאים אוקריוטים3. בהקפאה-ET, מידע תלת מימדי (3D) מתקבל על ידי הטיה פיזית של המדגם על במת המיקרוסקופ ורכישת סדרה של תמונות דרך המדגם בזוויות שונות. תמונות אלה, או סדרת הטיה, מכסות לעתים קרובות טווח של +60/-60 מעלות במרווחים של דרגה אחת עד שלוש מעלות. לאחר מכן ניתן לשחזר את סדרת ההטיה באופן חישובי לאמצעי אחסון תלת-ממדי, הידוע גם בשם טומוגרמה4.
כל טכניקות ההקפאה-EM דורשות שהדגימה תהיה מוטבעת בשכבה דקה של קרח אמורפי, לא גבישי, זגוגי. אחת מטכניקות ההקפאה הנפוצות ביותר היא הקפאת צלילה, שם המדגם מוחל על רשת EM, מוכתם, וצולל במהירות לתוך אתאן נוזלי או תערובת של אתאן נוזלי פרופן. טכניקה זו מספיקה עבור vitrification של דגימות מ 100 ננומטר < ~ 10 מיקרומטר בעובי, כולל תאים אנושיים בתרבית, כגון תאי HeLa5,6. דגימות עבות יותר, כגון מיני אורגנוידים או ביופסיות רקמות, בעובי של עד 200 מיקרומטר, יכולות להיות vitrified על ידי הקפאה בלחץ גבוה7. עם זאת, בשל פיזור אלקטרונים מוגבר של דגימות עבות יותר, מדגם ועובי קרח עבור cryo-ET מוגבל ~ 0.5 - 1 מיקרומטר ב 300 kV שידור מיקרוסקופים אלקטרונים. לכן, קריו-ET של תאים של תאים שלמים של תאים אוקריוטים רבים מוגבל לפריפריה של התא או הרחבות של תאים אלא אם כן נעשה שימוש בשלבי הכנה מדגם נוספים, כגון cryo-sectioning8 או כרסום קרן יון ממוקדת9,10,11.
מגבלה של ניסויים רבים בהדמיית קריו-ET של תאים שלמים היא תפוקת איסוף נתונים12. שלא כמו cryo-EM חלקיק יחיד, שבו אלפי חלקיקים מבודדים לעתים קרובות ניתן לדמיין מתוך ריבוע רשת TEM יחיד, תאים הם גדולים, להתפשט, ויש לגדל בצפיפות נמוכה מספיק כדי לאפשר את התאים להישמר בשכבה דקה של קרח זגוגית. לעתים קרובות אזור העניין מוגבל לתכונה מסוימת או לאזור משנה מסוים של התא. תפוקה מגבילה נוספת היא הנטייה של תאים לגדול באזורים שאינם נוחים להדמיית TEM, כגון בסרגלי רשת TEM או בקרבתם. בשל גורמים בלתי צפויים הקשורים לתרבות התאים ברשתות TEM, יש צורך בפיתוחים טכנולוגיים כדי לשפר את הנגישות והתפוקה לדוגמה לרכישת נתונים.
מיקרו-פסטיבל מצע עם חלבוני מטריצה חוץ-תאית דבקים (ECM) היא טכניקה מבוססת היטב למיקרוסקופיה של אור חי כדי לכוון את צמיחת התאים על משטחים קשיחים, עמידים ושקופים אופטית כגון זכוכית ומצעים אחרים של תרבית רקמות 13,14. Micropatterning בוצע גם על משטחים רכים ו /או תלת מימדיים (3D). טכניקות כאלה לא רק אפשרו מיקום מדויק של תאים; הם גם תמכו ביצירת רשתות רב-תאיות, כגון מעגלי תאים עצביים בדפוס15. הבאת micropatterning כדי cryo-ET לא רק להגדיל את התפוקה, אבל זה יכול גם לפתוח מחקרים חדשים לחקר microenvironments הסלולר מורכב ודינמי.
לאחרונה, מספר קבוצות החלו להשתמש בטכניקות micropatterning ברשתות TEM באמצעות גישות מרובות16,17. כאן, השימוש בטכניקת photopatterning ללא מסכה עבור רשתות TEM מתואר באמצעות מערכת micropatterning Alvéole PRIMO, הכוללת תבניות ברזולוציה גבוהה וללא מגע. עם מערכת micropatterning זו, שכבה נגד עכירות מוחלת על החלק העליון של המצע, ואחריו היישום של photocatalyst אבלציה של שכבת אנטי עכירות בדפוסים המוגדרים על ידי המשתמש עם לייזר UV. לאחר מכן ניתן להוסיף חלבוני ECM לתבניות של תרבית התאים המתאימה. שיטה זו שימשה מספר קבוצות למחקרי קריו-ET של אפיתל הפיגמנט הרשתית-1 (RPE1), מאדין-דארבי כלב כליות-II (MDCKII), פיברובלסט עורלה אנושית (HFF), וקווי תאי אנדותל16,17,18. מערכת micropatterning זו תואמת עם מצעים מרובים נגד עכירות שכבה, כמו גם ריאגנט פוטוקטליסט נוזלי או ג'ל. ניתן לבחור מגוון חלבוני ECM ולהתאים אותם לספציפיות של קו התא, המעניקים רב-תכליתיות למשתמש.
Micropatterning יושם בהצלחה על מספר פרויקטים בתוך המעבדה19. כאן, פרוטוקול micropatterning מוצג, כולל התאמות ספציפיות ללמוד תאי HeLa מתורבתים, וירוס סינסיטיאלי נשימתי (RSV) נגוע BEAS-2B תאים, נוירונים זחל ראשי Drosophila melanogaster20.
Protocol
הפרוטוקול המתואר כאן הוא אוסף של תרבות התאים, micropatterning, ושיטות הדמיה המשמשות את מעבדת רייט ואת מרכז המחקר Cryo-EM באוניברסיטת ויסקונסין, מדיסון. זרימת העבודה מוצגת באיור 1. הדרכה וחומרי הדרכה נוספים זמינים באתרים הבאים: https://cryoem.wisc.edu או https://wrightlab.wisc.edu
1. הכנת רשתות לדפוסי דפוס
- מעבירים את רשתות ה- TEM למגלשת זכוכית נקייה, צד פחמן כלפי מעלה (העובי הסטנדרטי של רדיד הפחמן הוא 12 ננומטר). באמצעות מאייד פחמן, ACE600, לאדות 5-8 ננומטר של פחמן נוסף על הרשתות כדי להגדיל את העמידות הכוללת של סרט פחמן.
הערה: שלב זה אינו נחוץ עבור רשתות SiO2 . צעד זה עשוי להיעשות גם מראש; לאחסן את הרשתות מצופות בסביבה לחות נמוכה כגון ייבוש ואקום. - העבירו את הרשתות למחזיק הכנה לרשת ושחררו בזוהר את הרשתות בצד הפחמן כלפי מעלה. באמצעות מערכת פריקה זוהר, זוהר לפרוק את הרשתות עבור 60 s ב 10 mA עם מרחק עבודה 80 מ"מ ולחץ ואקום של 1.0 x 10-3 mbar. עשה זאת תוך 15-30 דקות מהשלב הבא.
הערה: ניתן לקנות באופן מסחרי או להכין את מחזיקי הכנת הרשת באופן מסחרי או תוצרת בית עם פיסת נייר סינון על צלחת פטרי קטנה.
2. יישום השכבה נגד עכירות
הערה: יש להשתמש בטכניקה סטרילית נכונה בעת טיפול ברשתות, וכל הפתרונות צריכים להיות סטריליים ו/או מסננים מעוקרים.
- מעבירים את הרשתות (צד פחמן למעלה) למגלשת זכוכית נקייה או מכסים עם הפרדה של 1 ס"מ לפחות בין הרשתות. פיפטה 10 μL של 0.05% פולי-L-ליזין (PLL) על כל רשת. לדגור על הרשתות בתא לח, כגון קופסת פלסטיק סגורה עם מגבות נייר לחות, במשך 30 דקות לפחות.
הערה: ניתן להרחיב שלב זה עד הלילה. ודא שרמת הלחות בתא מספיקה כדי למנוע מהרשתות להתייבש. - לשטוף כל רשת שלוש פעמים עם 15 μL של 0.1 M HEPES pH 8.5. עבור כל לשטוף, להסיר את רוב הנוזל מהרשת עם פיפטה מבלי לתת לרשת להתייבש. מוסיפים 15 μL של חוצץ טרי, דגירה עבור לפחות 30 s ולחזור על. השאירו כל רשת ב 15 μL של 0.1 M HEPES לאחר הכביסה הסופית.
הערה: בשלב זה ובשלבים עתידיים, חשוב לשמור על הרשת רטובה ולהימנע ממגע בין הפיפטה לרשת. - הכן 10 μL של 100 מ"ג / מ"ל פוליאתילן גליקול-succinimidyl valerate (PEG-SVA) ב 0.1 M HEPES pH 8.5 עבור כל רשת. PEG-SVA יתמוסס במהירות עם ערבוב עדין וכתוצאה מכך פתרון ברור.
הערה: אין להכין מראש את פתרון PEG-SVA. PEG-SVA יש מחצית חיים של 10 דקות ב pH 8.5. הימנעו מחשיפת מלאי PEG-SVA ללחות מופרזת על ידי אחסונו בסביבה מתייבשת או יבשה בטמפרטורת החדר לפני הפתיחה. - מיד לאחר הכנת פתרון PEG-SVA, להסיר את טיפת 15 μL של HEPES pH 8.5 מכל רשת (דואג לא לייבש את הרשת) ולהוסיף טיפה 10 μL של פתרון PEG-SVA. לדגור על הרשתות בתא לח לפחות 1 שעות.
הערה: ניתן להרחיב שלב זה עד הלילה. ודא שהלחות בתא מספיקה כדי למנוע מהרשתות להתייבש. - לשטוף כל רשת שלוש פעמים עם 15 μL של מים סטריליים. עבור כל לשטוף, להסיר את רוב הנוזל מהרשת עם פיפטה מבלי לתת לרשת להתייבש, להוסיף 15 μL של מים מתוקים, לדגור על לפחות 30 s ולחזור על. השאר כל רשת ב 15 μL של מים לאחר הכביסה הסופית.
3. מריחת ג'ל PLPP
- הכן כיסוי מיקרוסקופ נקי עבור כל רשת. השלם את השלבים הבאים עבור כל רשת, רשת אחת בכל פעם, כדי למזער את הסיכוי שהרשת תתייבש.
- מניחים טיפת מים של 1.0 μL במרכז כיסוי כדי לעזור עם הצבת הרשת על כיסוי ושמירה על הרשת רטובה. בזהירות להעביר את הרשת מן טיפת מים 15 μL ל טיפת מים 1.0 μL על כיסוי. הקפד למקם את צד הפחמן של הרשת למעלה.
- בזהירות למקם סטנסיל polydimethylsiloxane (PDMS) על גבי הרשת, הקפדה לשמור על הרשת ממורכזת כדי למזער מגע סטנסיל עם רדיד הפחמן של הרשת.
- הוסף 1.0 μL של 4-בנזואילבנזיל-trimethylammonium כלוריד (PLPP) ג'ל על הרשת. פיפטה בעדינות לערבב (לא לגעת ברשת עם קצה פיפטה).
- הזז את כיסוי הכיסוי עם הרשת למיקום חשוך לייבוש. הג'ל יתייבש בעוד כ-15-30 דקות.
4. כיול ועיצוב המיקרו-פאטרן
- צבע צד אחד של כיסוי זכוכית עם עט סימון. הוסף קווים שחורים מסמן קבוע עם קצה דק כדי להקל על המיקוד. מניחים את כיסוי הכיסוי על המיקרוסקופ כך שהצד הצבעוני פונה לעדשת המטרה. באמצעות מצב Brightfield, התמקדו בעט הסימון.
- ודא שהמיקרוסקופ ומערכת המיקרו-פצירה מופעלים, ונתיב האור הנכון מוגדר. פתח את Micromanager ואת תוכנת לאונרדו (תוספים > לאונרדו) במחשב המיקרוסקופ.
- בחר כיול ובצע את ההוראות המופיעות על המסך. התאם את מוקד המיקרוסקופ כך שהתמונה המוקרנת על השקופית תהיה בפוקוס. ייתכן שיהיה צורך להפחית את זמן החשיפה. לאחר הכיול, בחרו 'תבנית עכשיו'.
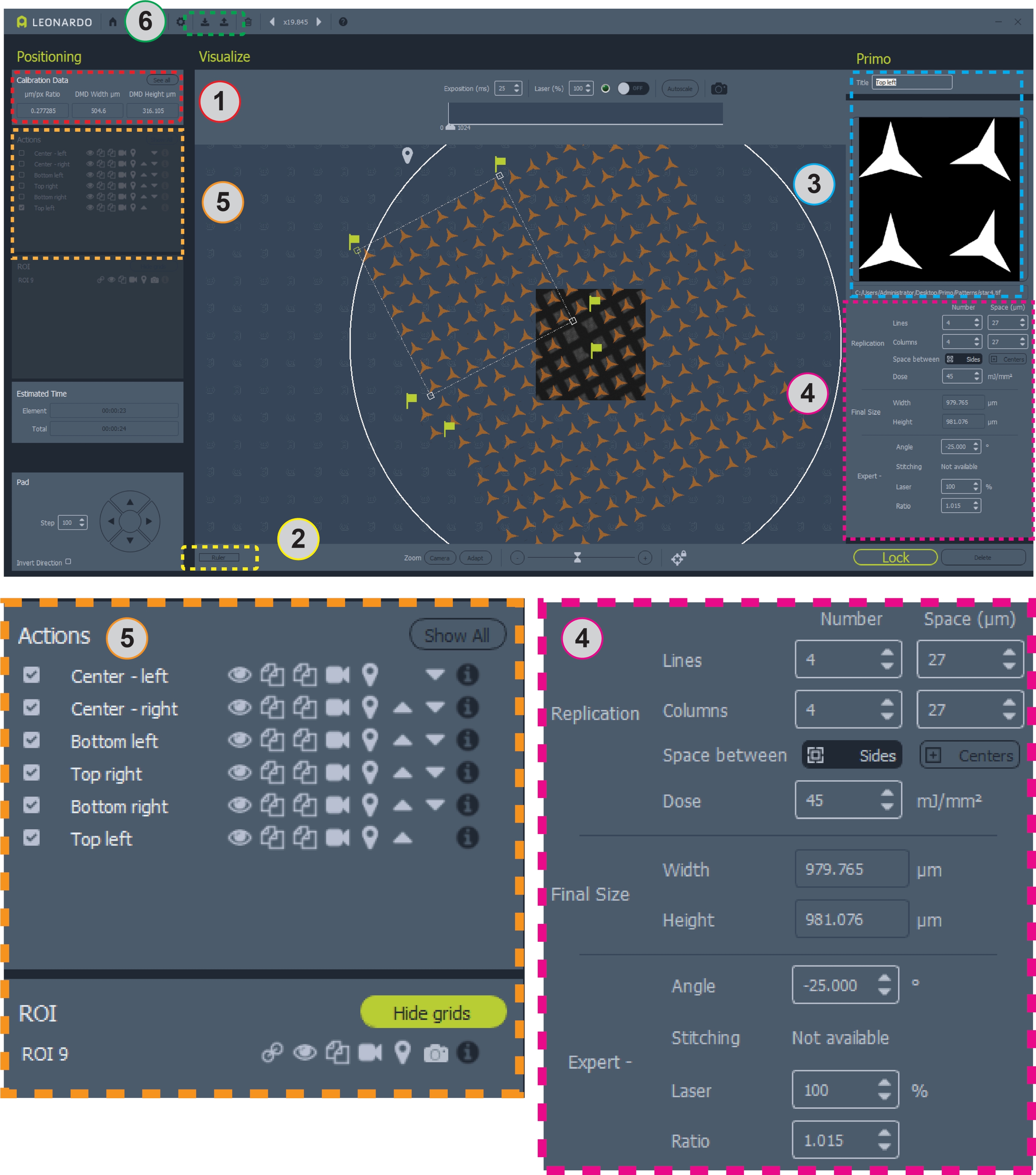
- הקלט את יחס המיקרומטר/פיקסל (מיקרומטר/פיקסל) המדווח תחת נתוני כיול בחלון הימני העליון של התוכנית (איור 2, אזור 1). השתמש ביחס זה כדי לקבוע את מספר הפיקסלים שיש להשתמש בהם לכל מיקרומטר בעת עיצוב תבנית.
- לאחר הכיול, ודא שהתוכנה פתוחה כעת עם תצוגה חיה של Brightfield מהמיקרוסקופ. טען רשת מוכנה על כיסוי (סעיף 3) על הבמה כאשר הרשת פונה לעדשת המטרה. מקם את השלב והתאם את המוקד כך שהרשת תהיה גלויה בחלון התוכנה.
- מדוד את הגודל של ריבועי הרשת ופסי הרשת במיקרומטרים. התוכנה כוללת סרגל המופעל על ידי הלחצן ליד הפינה השמאלית התחתונה כדי למדוד את הרשת (איור 2, אזור 2). לדוגמה, התבניות המשמשות כאן עבור רשת שינוי של 200 תואמות ל- ~ 87 × ריבועי רשת של 87 מיקרומטר ופסי רשת ~ 36 מיקרומטר.
הערה: התוכנה מציעה גמישות בשינוי גודל דפוסים תוך כדי תנועה, כך שניתן לסבול אי דיוקים קלים במדידה. - בהתבסס על המדידות והיחסים לעיל, צור תבניות עם כל תוכנה ליצירת תמונות. גודל התכונה המינימלי עם יעד של 20× הוא 1.2 מיקרומטר. יש לשמור תבניות כקבצי .tiff של 8 סיביות שאינם דחוסים.
- ודא שהתוכנה אינה משנה את גודל התמונות לגודל פיקסל שונה בעת השמירה. התבנית אמורה להתאים לתיבה של 800 ×800 פיקסלים, מספיק כדי לכסות ארבעה ריבועי רשת.
הערה: פיקסלים עם ערך של 255 (לבן) יעוצבו בעוצמה הגבוהה ביותר (מינון כולל של הלייזר) ופיקסלים עם ערך של אפס (שחור) לא יעוצבו. כל פיקסל עם ערך ביניים יהיה בדוגמת מנה של כ (X/255)*מינון כולל. באיור 3A, ערכי פיקסלים של 255 ו-129 שימשו לתבניות בגווני אפור. לאחר העיצוב של התבנית ניתן לשמור אותו ולהשתמש בו שוב ללא שינוי.
- ודא שהתוכנה אינה משנה את גודל התמונות לגודל פיקסל שונה בעת השמירה. התבנית אמורה להתאים לתיבה של 800 ×800 פיקסלים, מספיק כדי לכסות ארבעה ריבועי רשת.
5. מיקרו-פאטרנינג
- לאחר הכיול, ודא שהתוכנה פתוחה כעת עם תצוגה חיה של Brightfield מהמיקרוסקופ. טען רשת מוכנה על כיסוי (סעיף 3) על הבמה כאשר הרשת פונה לעדשת המטרה. מקם את הבמה והתאם את המוקד כדי לראות את הרשת בתוכנה.
- עבור הפעלה ראשונית, עצב תבנית חדשה. בתוכנה, בחרו ' הוסף החזר השקעה' (לא מוצג, במיקום של איור 2 אזור 3) ובחרו עיגול של 3,000 מיקרומטר. מקם את ההחזר על ההשקעה במעגל מעל הרשת באמצעות תמונת brightfield על המסך כמדריך עזר. לחץ על מנעול כדי לאבטח את ההחזר על ההשקעה.
- לאחר נעילת ההחזר על ההשקעה במקום, בחרו ' הוסף דוגמת מילוי' (לא מוצגת, במיקום של איור 2 אזור 3). בחר את התבנית המעוצבת בסעיף 4. חלק את הרשת לשישה אזורים כדי לאפשר מיקוד ומיקום עצמאיים בכל אזור כדי לקחת בחשבון רשתות לא אחידות. 8 × 8 אזור ריבועי רשת עבור כל פינה ברשת ו-2 × 8 אזור ריבועי רשת בכל צד של המרכז, מה שמשאיר את ארבעת ריבועי הרשת המרכזיים לא מרוסנים (איור 2, תמונת מרכז).
- השתמש באפשרויות השכפול (איור 2, אזור 4) כדי ליצור עותקים של התבנית ההתחלתית כדי להגיע למספר הרצוי של העותקים הכוללים של התבנית. התאם את המרווח בין עותקים כך שיתאים למרווח בין ריבועי רשת במידת הצורך.
- הגדר את המינון הכולל עבור התבנית. 30 mJ / mm2 היא נקודת התחלה טובה. עיין בסעיף הדיון לקבלת פרטים נוספים.
- תחת אפשרויות מומחה (איור 2, אזור 4) התאם את זווית האזור כך שתתאים לזו של ריבועי הרשת. ניתן למקם מחדש אזורים באמצעות העכבר. ניתן גם להתאים את היחס (גודל) של התבניות. חזרו על הפעולה באמצעות התאמת הזווית, המיקום, הרווח בין התבנית והיחס אליה עד שהתבניות יתיישרו עם ריבועי הרשת. הזז את שלב המיקרוסקופ כדי לשנות את אזור הרשת בתצוגת הבהירות החיה.
- הקש Lock כדי לשמור את השינויים שבוצעו באזור.
- להעתקת אזור, לחצו על הלחצן 'שכפל ' (איור 2, אזור 5, סמל שני גליונות נייר) לצד שמו בחלונית 'פעולות ' מימין. למיקום מחדש, שינוי שם או עריכה של העותק, לחצו על שמו בחלונית 'פעולה'.
- חזור על שלבים 5.4-5.9 לפי הצורך כדי למלא את כל האזורים הרצויים.
- לאחר שהתבנית המלאה מעוצבת וממוקמת, שמור את קובץ התבנית בתוך התוכנה (איור 2, אזור 6, עמודה עם סמל חץ למעלה בסרגל הכלים העליון).
- בעת טעינת תבנית שנשמרה בעבר (מייצג פעילות עם סמל חץ למטה) מרכז את ההחזר על ההשקעה מעל הרשת והקש Lock. לחץ על כל אזור בלוח הפעולות כדי לשנות את הזווית, המיקום, המינון ו/או קובץ התבנית.
- לאחר מיקום התבנית והתבניות, בטל את הסימון של כל האזורים פרט לאחד בלוח הפעולות בתוכנה.
- השתמש בשלב המיקרוסקופ כדי לנווט לאזור זה ולהתמקד בנייר הפחמן. לחיצה על סמל גלגל העין בחלונית 'פעולה' (איור 2, אזור 5) תעביר את תצוגת שכבת-העל של דוגמת המילוי להפעלה או לביטול.
- לאחר שהרשת נמצאת בפוקוס, סגור את תריס brightfield ולחץ על סמל הפעל בפינה השמאלית התחתונה של התוכנה כדי להתחיל בתהליך התבנית, שניתן לפקח עליו בשידור חי.
- בחלונית הפעולה, בחרו בתיבה של האזור הבא. פתח את תריס brightfield כך שהרשת תהיה גלויה ומרכזת אזור זה באמצעות שלב המיקרוסקופ. חזור על שלבים 5.13-5.14 עבור כל אזור בלוח הפעולות.
- הסר את כיסוי כיסוי עם הרשת מן המיקרוסקופ, ומיד פיפטה 10 μL של תמיסת מלח פוספט-חוצץ סטרילי (PBS) על הרשת.
- לאחר 10 דקות, להסיר את הסטנסיל עם פינצטה, ולאחר מכן לשטוף את הרשת 3x עם 15 μL של PBS. לאחר השטיפה הסופית, מניחים כל רשת ב-15 μL של PBS ומעבירים את הרשתות למיקום חשוך.
6. התצהיר של חלבוני ECM
- עבור תאים בתרבית, בצע את השלבים 6.2-6.5; עבור נוירונים Drosophila ראשוני, בצע את השלבים 6.6-6.10.
- הכן לפחות 15 μL של ECM עבור כל רשת. עבור תאי BEAS-2B, הכן ריכוז סופי של 0.01 מ"ג / מ"ל פיברונקטין בקר ו 0.01 מ"ג / מ"ל פלואורופור-מצומד פיברינוגן PBS סטרילי. עבור תאי HeLa, הכן 0.01 מ"ג / מ"ל קולגן בקר I ו 0.1 מ"ג / מ"ל פלואורופור-מצומד פיברינוגן ב- PBS סטרילי.
- הסר את רוב ה- PBS מכל רשת והחיל 15 μL של ECM. לדגור על הרשת בתא לח בטמפרטורת החדר לפחות 1 שעות.
הערה: שלב זה עשוי להיות מורחב ללילה ב 4 °C (65 °F). - לאחר הדגירה ב- ECM, לשטוף כל רשת 5x עם PBS סטרילי. עבור כל לשטוף, להסיר את רוב הנוזל עם פיפטה מבלי לתת לרשת להתייבש, להוסיף 15 μL של PBS טרי, לדגור על לפחות 30 s, ולחזור על. השאר כל רשת ב- PBS לאחר הכביסה הסופית.
הערה: ניתן לאחסן רשתות עד שבוע ב- PBS בטמפרטורה של 4 °C (65 °F) ללא הידרדרות נצפתה באיכות. - השתמש במיקרוסקופ פלואורסצנטי כדי לזהות את הפלואורופור ב- ECM כדי לאשר דפוסים וכי רדיד הפחמן נשאר שלם. כמה ריבועים שבורים הם בדרך כלל נסבלים.
- עבור נוירונים Drosophila העיקריים, להעביר את הרשתות בדוגמת צלחת תחתונה זכוכית 30 מ"מ המכיל PBS סטרילי.
- לשאוף את PBS מן המנה ולהחיל 2 מ"ל של 0.5 מ"ג / מ"ל פלואורופור מצומד concanavalin A. דגירה לילה ב 25 °C (55 °F) בסביבה סטרילית.
- הסר את concanavalin פתרון מהצלחת (מבלי לייבש את הרשתות) ולשטוף את הרשתות 3x עם PBS. לכל שטיפה, מוסיפים ומסירים 2 מ"ל PBS מהמנה.
- השתמש במיקרוסקופ פלואורסצנטי כדי לזהות את הפלואורופור ב- ECM כדי לאשר דפוסים וכי רדיד הפחמן נשאר שלם. כמה ריבועים שבורים הם בדרך כלל נסבלים.
- לאחר השטיפה הסופית, הסירו את ה-PBS מהצלחת התחתונה של הזכוכית והוסיפו 2 מ"ל של דרוזופילה media21 של שניידר, המכיל 20% סרום בקר עוברי מושבת בחום (FBS), 5 מיקרוגרם/מ"ל אינסולין, 100 מיקרוגרם/מ"ל פניצילין, 100 מיקרוגרם/מ"ל סטרפטומיצין ו-10 מיקרוגרם/מ"ל טטרציקלין. דגירה בטמפרטורה של 25 מעלות צלזיוס בסביבה סטרילית עד שהנוירונים יהיו מוכנים לציפוי.
7. הכנת תאי דרוזופילה ראשוניים לפני הזריעה
- לחטא צלחת ביתור 55 מ"מ עם 70% EtOH, ולאחר מכן להוסיף למנה 2-3 מ"ל של סטרילי מסונן 1× לחתוך מלוחים (9.9 מ"מ HEPES pH 7.5, 137 מ"מ NaCl, 5.4 mM KCl, 0.17 mM NaH2PO4, 0.22 mM KH2PO4, 3.3 mM גלוקוז, 43.8 mM סוכרוז)21.
- בחרו 30-40 זחלי אינסטאר 3 בעדינות מהאוכל באמצעות זוג פינצטה.
- מניחים את הזחלים לתוך הצינור עם 1× PBS, ולאחר מכן להעביר אותם לתוך הצינור השני עם 1× PBS לשטוף את הזחלים.
- העבר את הזחלים לתוך הצינור עם 70% EtOH לשטוף את PBS, ולאחר מכן להעביר אותם לתוך הצינור השני עם 70% EtOH. השאירו את הזחלים בצינור השני במשך 2-3 דקות כדי לעקר את הזחלים.
- מעבירים את הזחלים לצינור עם תמיסת מלח 1×, ואז מעבירים אותם מיד לצינור השני עם תמיסת מלח 1×.
- מעבירים זחלים בודדים לצלחת הניתוח המכילה תמיסת מלח 1×. עם זוג מלקחיים ומיקרוסקופ ניתוח, לקרוע במהירות כל זחל כדי לחלץ את המוח ולהעביר אותו לצינור השלישי עם 1× תמיסת מלח. חזור על הפעולה עד שכל המוחות יחולצו.
- צנטריפוגה הצינור המכיל את המוח ב 300 x g במשך 1 דקה.
- להשליך את supernatant, ולשטוף עם 1 מ"ל של 1× לחתוך תמיסת מלח וצנטריפוגה הצינור ב 300 x g במשך 1 דקה. חזור על שלב זה פעם נוספת.
- להשליך את supernatant עד 200-250 μL נשאר בצינור ולהוסיף 20 μL של 2.5 מ"ג / מ"ל ליבראז ב 1x תמיסת מלח.
- לסובב את הצינור על סיבוב במשך 1 שעה בטמפרטורת החדר; במהלך שעה זו, פיפטה הפתרון 25-30 פעמים כל 10 דקות. בסוף, הפתרון צריך להיות מעט אטום.
- צנטריפוגה התאים ב 300 × גרם במשך 5 דקות.
- להשליך את supernatant, ולאחר מכן להוסיף 1 מ"ל של מדיה נוספת שניידר. פיפטה הפתרון 30 פעמים לערבב.
- צנטריפוגה התאים ב 300 × גרם במשך 5 דקות.
- להשליך את supernatant ולשטוף את גלולה התא על ידי הוספת 1 מ"ל של מדיה נוספת שניידר. פיפטה הפתרון 30 פעמים לערבב.
- צנטריפוגה התאים ב 300 × גרם במשך 5 דקות.
- להשליך את supernatant, ולאחר מכן resuspend את גלולה התא עם 300 μL של מדיה נוספת של שניידר. פיפטה הפתרון 30-40 פעמים לערבב.
8. תרבות וזיהום RSV של תאי BEAS-2B ו- HeLa
- שמור על תאי HeLa ותאי BEAS-2B בבקבוקונים T75 ב 37 °C (5 ° C ) ו 5 % CO2. תאי מעבר כל 3-4 ימים מגיעים פעם אחת לכ-80% מפגש. שמור על תאי HeLa ב- DMEM + 10% FBS + 1× אנטיביוטית-אנטי-אנטיביוטית. שמור על BEAS-2B ב- RPMI + 10% FBS + 1× אנטיביוטיקה-אנטי-אנטי-מיקרוטית6,22,23.
- לזריעה של תאים לא נגועים, דלג לסעיף 9. תאי BEAS-2B ו- HeLa רגישים לזיהום RSV; תאי BEAS-2B שימשו לכל הניסויים מעורבים RSV המוצגים כאן.
הערה: בצע את כל שלבי BSL-2 בהתאם לפרוטוקולים מוסדיים באמצעות ארון בטיחות ביולוגית (BSC) מתאים וציוד מגן אישי (PPE). - לפני זיהום RSV של תאים, מעבר 5 × 104 תאים לבאר לתוך צלחת 6-well (שטח הפנים ~ 9.6 cm2) עם 2 מ"ל של מדיה צמיחה ודירה בן לילה.
- בצע טריפסין וספור באר אחת של תאים. כדי לנסות לנסות, שאפו מדיה מבאר אחת ושטפו עם 2 מ"ל של PBS סטרילי ללא Mg2+ ו- Ca2+ כדי להסיר את שאריות המדיה. הוסף 500 μL של 0.25% פתרון טריפסין. דגירה ב 37 °C (5-10 דקות). בדוק מעת לעת את התאים כדי לראות אם הם משתחררים מפני השטח. לאחר שחרור התאים, הוסף 1.5 מ"ל של מדיה תרבותית.
- מערבבים 100 μL של תאים טריפסיניים עם 100 μL של כחול טריפאן. פיפטה 10 μL של תערובת תאים מדוללים לתוך hemocytometer. ספור את התאים וחשב את מספר התאים לבאר. השתמש במספר זה כדי לחשב את MOI להלן.
- הכן דילול של RSV-A2mK +24 במדיית צמיחה כדי להשיג MOI של 10 לבאר ב 750 μL של מדיה. MOI של RSV-A2mK + ניתן לחשב מתוך יחידות מיקוד פלואורסצנטיות (FFU) titers של המניה (לדוגמה: עבור 1.0 × 105 תאים לבאר ומלאי RSV של 1.0 × 108 FFU / mL, לדלל את המלאי הנגיפי 1:75 ל 1 × 106 FFU / 750 μL או 1.33 × 106 FFU / mL).
- שאפו את המדיה מהתאים בצלחת 6-well ולהוסיף 750 μL של הפתרון הנגיפי מלמעלה לכל באר.
- מנענעים את הצלחת בטמפרטורת החדר למשך שעה אחת.
- לאחר 1 שעות, להביא את הנפח הכולל לבאר עד 2 מ"ל עם מדיה צמיחה מחומם מראש ל 37 °C (6 °F) ומניחים את הצלחת בחממה להגדיר 37 °C (5° C) עם 5% CO2 עבור 6 שעות.
- נסה את התאים כדי לשחרר אותם ולהמשיך לזרוע כמתואר להלן. לאחר הזריעה, לדגור על הרשתות עבור 18 שעות נוספות לפני הקפאת צלילה (עבור סך של 24 שעות לאחר ההדבקה).
9. זריעת תאים לרשתות מיקרו-מרופדות
- עבור תאים בתרבית, בצע את שלבים 9.2-9.8; עבור נוירונים Drosophila ראשוני, לעקוב אחר 9.9-9.11.
- בצע טריפסינציה של התאים כדי לשחרר אותם (ראה שלב 4 בסעיף 8 לעיל). כדי להפחית את צבירת התאים, בצע טריפסין לתאים ב- 60 % או פחות.
- מערבבים 100 μL של תאים טריפסיניים עם 100 μL של כחול טריפאן. פיפטה 10 μL של התא המדולל לערבב לתוך hemocytometer ולספור את התאים.
- לדלל את התאים במדיה ל 2 × 104 תאים / מ"ל.
- הוסף 1 μL של מדיה למרכז צלחת תחתונה זכוכית 30 מ"מ כדי לסייע בהצבת הרשת ולמנוע ממנו להתייבש. מעבירים את הרשת מה-PBS על כיסוי הכיסוי למרכז צלחת הזכוכית התחתונה. הוסף 10 μL של פתרון התא לרשת.
- באמצעות מיקרוסקופ ברייטפילד, שימו לב להידבקות התא לרשת לאחר 5 דקות. אם רוב התבניות נשארות פנויות, הוסף שחרור נוסף של 10 μL של פתרון התא. שמור על הרשתות ואת פתרון התא ב 37 °C (55 °F) במהלך הדגירה.
- חזור על שלב 9.6 עד שרוב התבניות תפוסות או שתבניות תפוסות רבות מתחילות לכלול תאים מרובים. לדגור על הרשת במשך 2 שעות באינקובטור (37 °C (37 °C (5° פרנהייט, 5% CO2).
- מציפים את המנה עם 2 מ"ל של מדיה מחוממת מראש ודגור לילה (37 °C (37 °C (5% CO2).
- עבור נוירונים Drosophila העיקריים, להסיר את המדיה מן המנה המכילה את הרשת צלחת צלחת התאים על המנה.
- המתן 30-60 דקות עד שהתאים יצורפו, ולאחר מכן הוסף 2 מ"ל של מדיה נוספת של שניידר.
- תרבות הנוירונים בחממה של 25 °C (25 °C) לפחות 2-3 ימים לפני הקפאת הצלילה.
10. הדמיה ו vitrification של רשתות דפוס
- מניחים את צלחת הזכוכית התחתונה המכילה את הרשת המעוצבת ואת התאים בתרבית על מיקרוסקופ הפלואורסצנטיות.
- השג תמונות של הרשת באמצעות brightfield ואת ערוצי פלורסנט המתאימים כדי לזהות את התבנית וכל תיוג אחר בתאים. ודא שצפיפות התאים והמיצוב מתאימים להדמיה וניתוח במורד הזרם.
הערה: תמונות ברייטפילד ופלואורסצנטיות עובדו בחבילת התוכנה של FIJI25. - מכינים מקפיא קפוא; סוג התקן ההקפאה יהיה תלוי בזמינות, בעלות ובתכונות המתאימות ביותר לדגימה.
הערה: נוירונים דרוזופילה ראשוניים הוכנו על מקפיא עמוק אוטומטי, ותאי BEAS-2B הוכנו באמצעות מקפיא עמוק חצי אוטומטי. - החל fiducials זהב על הדגימות ליישור תקין של סדרת ההטיה. כתמים דגימות כדי להסיר מדיה עודפת, ולאחר מכן לצלול-להקפיא את הדגימות לתוך קריוגן, כגון אתאן נוזלי מקורר על ידי חנקן נוזלי. עבור נוירונים Drosophila העיקרי, כתם עבור 4 s מן הישבן. עבור תאי HeLa ו- BEAS-2B, כתם משני הצדדים עבור 4-6 s. לאחר מכן ניתן לאחסן את הרשתות הקפואות בחנקן נוזלי עד לשימוש נוסף.
- תמונה vitrified תאים במיקרוסקופ קריו-אלקטרון, מופעל ב 300 kV עם מצלמת גלאי אלקטרונים ישירה. הגדר איסוף סדרות הטיה עבור כל אזור מעניין באמצעות תוכנה כגון SerialEM26 לאיסוף נתונים קריו-EM/cryo-ET.
הערה: סדרת הטיה של נוירונים Drosophila העיקריים נאספו על גלאי אלקטרונים ישיר מ -60° עד 60° באופן דו-כיווני במרווחים של 2° ב -8 מיקרומטר דפוקוס עם גודל פיקסל של 4.628 Å עבור מנה כוללת של 70-75 e-/Å2. סדרת הטיה של BEAS-2B נגוע RSV נאספו על גלאי אלקטרונים ישיר עם מסנן אנרגיה (20 חריץ eV) ב -5 מיקרומטר defocus עם גודל פיקסל של 4.603 Å ומינון כולל של ~ 80 e/Å2. - עבד את סדרת ההטיה כדי לשחזר טומוגרמה.
הערה: טומוגרמה שהוצגה כאן שוחזרה באמצעות חבילת IMOD27; סינון lowpass נעשה באמצעות חבילת התוכנה EMAN228.
תוצאות
הליך זה שימש לתבנית רשתות EM לניסויי קריו-ET של תאים שלמים. זרימת העבודה כולה המוצגת במחקר זה, כולל הכנות ראשוניות לתרבות התאים, מיקרו-פצפוצינג (איור 1) והדמיה, כוללת 3-7 ימים. הליך דו-שלבי שימש ליצירת השכבה למניעת עכירות על-ידי החלת PLL על הרשת ולאחר מכן קישור PEG על-ידי הוספת PEG-SVA תגובתי. ניתן להחיל את השכבה נגד עכירות גם בשלב אחד על ידי הוספת PLL-g-PEG בדגרה אחת. ג'ל PLPP הוא זרז עבור micropatterning UV, אשר זמין גם כנוזל מרוכז פחות. הג'ל מאפשר דפוס במינון מופחת באופן משמעותי בהשוואה לנוזל, מה שגורם דפוסים הרבה יותר מהר. עם מערכת זו, זמן התבנית בפועל של רשת TEM מלאה היה ~ 2 דקות. זרימת העבודה של המיקרו-תיקון לבדה משתרעת בדרך כלל על פני 5-6 שעות ומאפשרת לאדם לעצב שמונה רשתות עבור תרבית תאים סטנדרטית ברשתות TEM.
מספר שלבים במהלך תהליך micropatterning דורשים זמני דגירה ארוכים (ראה שלבים 2.1, 2.3, 6.4). בנוחות, חלק משלבים אלה, כגון פסיביזציה PLL (2.1) או PEG-SVA פסיבינציה (2.3), עשויים להיות מורחבים לדגירה לילית. בנוסף, רשתות עשויות להיות מעוצבות מראש ומאוחסנות בתמיסה של חלבון ECM או PBS לשימוש מאוחר יותר. במחקר שלנו, אפשרויות אלה היו יקרות ערך במקרים שבהם התזמון של הכנת תאים וזריעה הוא קריטי, כגון נוירונים Drosophila ראשוני ו RSV-זיהום של תאי BEAS-2B.
הרשתות הוכנו במסגרת מעבדה כללית ברמת בטיחות ביולוגית 2 (BSL-2) תוך שימוש בכלים נקיים, פתרונות סטריליים, וכללו אנטיביוטיקה/אנטי-מיקוטיקה במדיית הצמיחה6,22,29,30. עבור דגימות רגישות במיוחד לזיהום מיקרוביאלי, ניתן ליישם את השכבה נגד עכירות ו- ECM במכסה המנוע של תרבית הרקמות או בסביבה סטרילית אחרת. בנוסף, ניתן לשטוף את הרשת באתנול בין דפוס ויישום ECM. אם עובדים עם גורמים זיהומיים, חשוב להתאים את ההליך כדי לעמוד בפרוטוקולים המתאימים של בטיחות ביולוגית.
זרימת עבודה זו וההליכים שהוצגו (איור 1) אפשרו לתאי HeLa (איור 4), לתאי BEAS-2B נגועים ב-RSV (איור 3, איור 5) ולנוירונים זחליים ראשיים של Drosophila (איור 6, איור 7) לזרוע על גבי רשתות EM בדוגמת דפוס לאיסוף נתוני קריו-ET אופטימלי.
תאי HeLa הנזרעים על רשתות TEM מיקרו-מרופדות נשארים ברי קיימא כפי שנקבעו על ידי כתמים פלואורסצנטיים באמצעות בדיקת כדאיות תאים מבוססת קלצין-AM ואתידיום הומודימר-1 (איור 4A,B). באמצעות ECM מעורב של קולגן ופיברינוגן, תאי HeLa נצמדים בקלות לדפוסים ברחבי הרשת (איור 4A, C). המורפולוגיה הכוללת של תאים המתרחבים לאורך התבנית דומה לזו של תאים הגדלים ברשתות לא מרוסנות (איור 4C, D). במקרה של תאי HeLa, עובי התא הכולל נשאר ~ < 10 מיקרומטר עם אזורים דקים משמעותית ~ < 1 מיקרומטר בעובי ליד פריפריית התא (איור 4E, F).
במחקרי RSV, עיצבנו ריבועי רשת שלמים באמצעות מעבר צבע, עם חשיפה במינון נמוך בקצוות ותבנית מינון גבוהה יותר לכיוון המרכז (איור 3A). דפוסי מעבר צבע הניבו תוצאות טובות יותר בעת חיפוש וירוסים שפורסמו הנמצאים ליד הפריפריה של תאים. עם דפוסים אלה, תאים נמצאו לדבוק באופן מועדף ריכוז ECM גבוה יותר, אבל הם גם מסוגלים לדבוק ולגדול על ריכוזי ECM נמוכים יותר. המינון היחסי בין אזורים יצטרך להיות ממוטב בעת שימוש בדפוסים הדורשים מינונים מרובים. אם המינונים ולכן ריכוזי ECM דומים מדי או שונים מדי זה מזה, ההשפעה של שימוש במינונים מרובים תאבד.
באיור 3, רשת TEM עוצבה ולאחר מכן נזרעה עם תאי BEAS-2B נגועים RSV ושימשה לאיסוף נתוני קריו-EM. איור 4A הוא תמונה פלואורסצנטית של ECM המעוצבת ברשת TEM באמצעות תבנית הדרגתית. הידבקות תאים וצמיחה לאורך האזור המרכזי של התבנית ניתן לראות באיור 3B כתמונת שדה בהיר של התאים 18 שעות לאחר הזריעה. באיור 3C, אות פלואורסצנטי (אדום) משכפול של RSV-A2mK+ מכוסה באות מה-ECM. רוב התאים הנגועים ממוקמים לאורך האזור המרכזי בצפיפות גבוהה יותר של תבנית השיפוע. מפת TEM במג נמוכה של הרשת לאחר ההקפאה-קיבעון חושפת מספר תאים, כולל תאים נגועים RSV, ממוקם על רדיד הפחמן ליד מרכז ריבועי הרשת. כפי שניתן לראות בעבר עבור תאים הגדלים ברשתות TEM סטנדרטיות22, סדרות הטיה אותרו ונאספו של נגיפי RSV בסמיכות לפריפריה של תאי BEAS-2B נגועים הגדלים על רשתות מיקרו-מרופדות (איור 5A,B). רבים מהחלבונים המבניים של RSV ניתנים לזיהוי בתוך הטומוגרמות, כולל נוקלאוקפפסיד (N) וחלבון ההיתוך הנגיפי (F) (איור 5C, חצים כחולים ואדומים בהתאמה).
במחקרים ראשוניים של נוירונים Drosophila, נמצא כי התבנית הצרה, בסמוך למגבלת הרזולוציה שמציעה התוכנה (שבה עובי התבנית היה 2 מיקרומטר), אפשרה לבודד בין תא אחד לכמה תאים בתוך ריבוע רשת (איור 6). הסומה העצבית הצליחה להאריך את הנוריטים שלה על פני תקופה של מספר ימים בתוך התבנית. הדבר איפשר זיהוי קל ורכישת סדרות הטיה של הניוריטים בהשוואה לנוירונים בתרבית ברשתות לא מרוסנות (איור 7). כמו כן נמצא כי פלואורסצנטית שכותרתו concanavalin A, לקטין ששימש ECM עבור תרבויות עצביות Drosophila במבחנה20,21, הוא מקובל עבור דפוס.
נוירונים Drosophila מזחלי instar השלישי היו מבודדים על פי פרוטוקולים שפורסמו בעבר20,21,31. ההכנות העצביות הוחלו על רשתות קריו-EM micropatterned שבו concanavalin A הופקד על התבנית כדי לווסת את מיקום התא, התפשטות, וארגון. הנוירונים ברשתות מעוצבות או לא מרופדות הורשו לדגור למשך 48-72 שעות לפחות, והרשתות היו אז קפואות. תמונה מייצגת של רשת EM מיקרו-מרופדת עם מספר נוירונים Drosophila המופצים על פני האזורים המעוצבים מוצגת באיור 6A. נוירונים אלה, נגזר זן זבוב מהונדס כי יש ביטוי GFP pan-עצבי בממברנה, ניתן לעקוב בקלות על ידי מיקרוסקופיה אור לא רק בשל תיוג פלואורסצנטי שלה, אלא גם בגלל המיקום שלה בתוך micropatterns. בעוד שניתן לעקוב אחר תאי עצב בתרבית ברשתות לא מרוסנות גם באמצעות איתות GFP באמצעות מיקרוסקופיה קלה (איור 7A, עיגול צהוב), איתורם בהקפאה-EM הפך לקשה יותר באופן משמעותי בשל נוכחותם של פסולת תאית וזיהום מהתקשורת (איור 7B, עיגול צהוב). נוכחות כזו הופחתה עבור נוירונים על רשתות דפוס, ככל הנראה בשל PEG בשכבה נגד עכירות של האזורים שאינם בדוגמת הדוחה את פסולת התא מהיצמדות. בשל הממדים של גוף תאי העצב והנוריטים המורחבים (איור 6A, B, עיגול צהוב), נאספו סדרות הטיית קריו-ET לאורך אזורים דקים יותר של התאים (איור 6C,D, עיגול אדום). קרום התא העצבי, המיטוכונדריון (ציאן), מיקרוטובולים (סגול), חוטי אקטין (כחול), מבני שלפוחית (כתום וירוק) ומקרומולקולות כגון ריבוזומים (אדום) נפתרו היטב במונטאז'ים ופרוסות תמונה בהגדלה גבוהה יותר באמצעות טומוגרמה תלת-ממדית (איור 6E). בעוד שניתן לראות תכונות תת-תאיות דומות מטומוגרמות תלת-ממדיות של נוירונים לא מרוסנים (איור 7E), הקושי באיתור יעדים תאיים מעשיים לאיסוף נתונים ירד משמעותית בתפוקה.
באיור 8, תמונות מייצגות מרשתות עם חלק מהבעיות הללו הורכבו כדי לסייע בזיהוי ובפתרון הבעיות שלהן. לאחר קביעת התנאים האופטימליים, micropatterning היא שיטה אמינה וניתנת לשחזור למיקום תאים ברשתות עבור cryo-TEM.

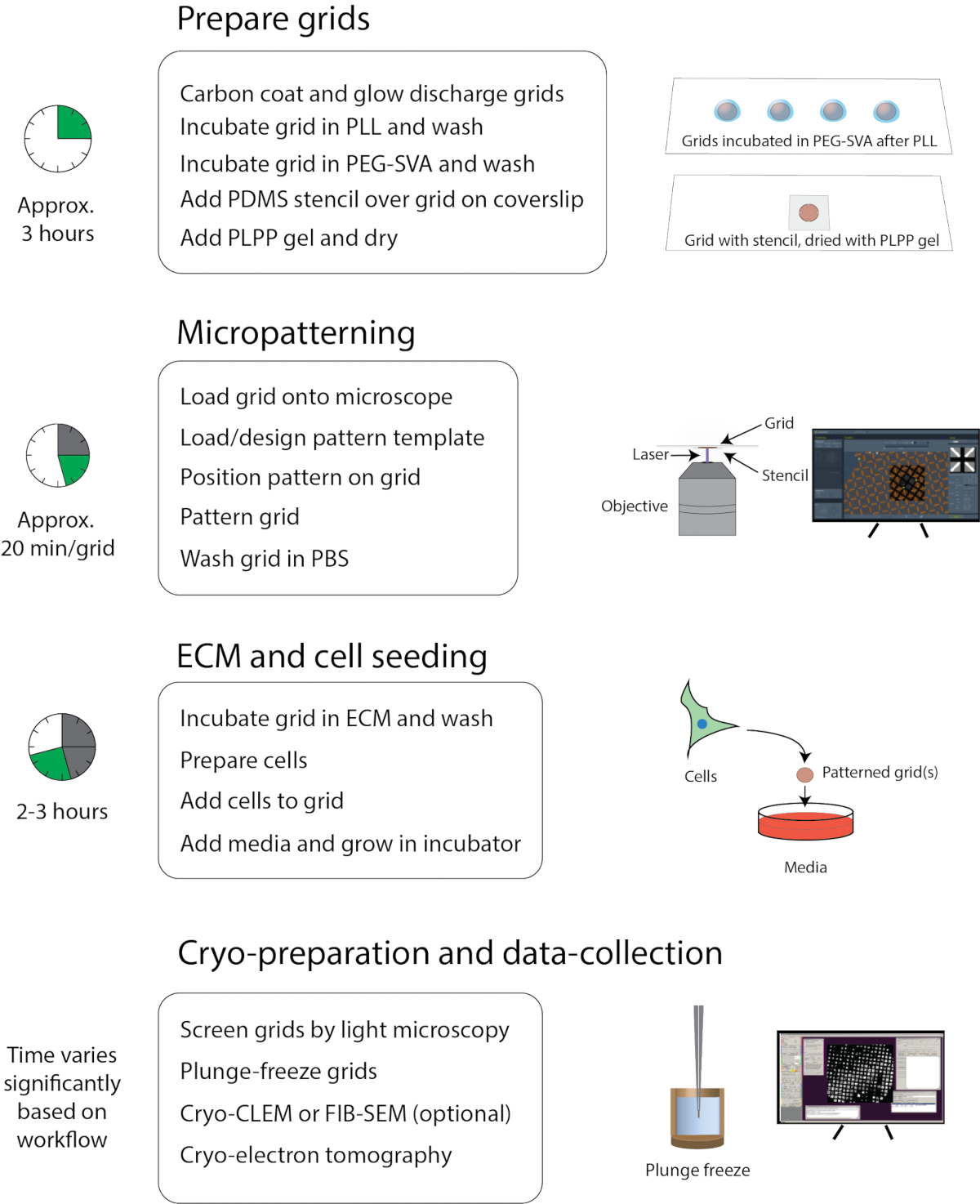
איור 1: זרימת עבודה כללית של מיקרו-פצפוצים להקפאה-EM. זרימת העבודה יכולה להיות מחולקת בערך ארבעה חלקים: הכנת רשת, micropatterning, ECM וזריעת תאים, והכנת קריו ואיסוף נתונים. השלבים העיקריים של כל מקטע מפורטים מתחת לכותרות והזמן המשוער להשלמת כל מקטע מוצג משמאל. לחץ כאן כדי להציג גירסה גדולה יותר של איור זה.
{kind=link}

איור 2: צילום מסך של התוכנה עם תבנית הממוקמת ברשת. אזור 1 מכיל את יחס המיקרומטר/פיקס לעיצוב תבנית. אזור 2 הוא הסרגל למדידת רשת. אזור 3 הוא המקום שבו יש להוסיף או לשנות תבניות ו- ROIs. אזור 4 מכיל את כל המידע עבור מיקום דפוס ומינון. אזור 5 מכיל אפשרויות עבור תבניות, כולל החלפת שכבות-על, העתקה או מחיקה של דוגמאות מילוי ובחירת תבניות למיקרו-חלוקה. אזור 6 הוא המקום שבו ניתן לשמור ולטעון תבניות. תצוגות גדולות יותר של אזורים 4 ו- 5 מוצגות להלן לקבלת בהירות. לחץ כאן כדי להציג גירסה גדולה יותר של איור זה.
{kind=link}

איור 3: תאי BEAS-2B נגועים ב-RSV ברשת ההקפאה-TEM בדוגמת התבנית. (A) תמונה פלואורסצנטית של הרשת בעלת התבנית לאחר הוספה של ECM בעל תווית פלואורסצנטית. תבנית הקלט מוצגת בפינה השמאלית התחתונה. (B) תמונת ברייטפילד של תאי BEAS-2B הגדלים ברשת ב- A. (C) מיזוג התמונה ב- A (ציאן) ו- B (אפור) עם תמונה פלואורסצנטית של תאים נגועים ב- RSV (אדום) מיד לפני הקפאת הצלילה; תאים נגועים מבטאים mKate-2. סרגלי קנה מידה הם 500 מיקרומטר. (D) מפת קריו-TEM בהגדלה נמוכה של הרשת ב- B לאחר הקפאת צלילה. תמונות פלואורסצנטיות צבועות בצבעים פסאודו-צבעיים. סרגלי קנה מידה הם 500 מיקרומטר. לחץ כאן כדי להציג גירסה גדולה יותר של איור זה.
{kind=link}

איור 4: כתמים חיים/מתים של תאים בדוגמת דפוס ולא מרוסנים. (A) תמונה פלואורסצנטית של תאי HeLa הגדלים על רשת בדוגמת ומוכתמים בקלצין-AM (כתם תא חי, ירוק) ואתידיום הומודימר-1 (כתם תא מת, אדום). (B) תאי HeLa הגדלים על רשת לא מרוסנת ומוכתמים כמו ב- A. (C) הקרנה של ערימות z קונפוקליות של תא HeLa על רשת Quantifoil R2/2 בדוגמת עם 0.01 מ"ג / מ"ל קולגן ופיברינוגן 647 ECM (אדום). התא היה מוכתם בקלצין-AM (ירוק) ובהוכסט-33342 (כחול). (D) תאי HeLa על רשת לא מרוסנת דגירה עם קולגן 0.01 מ"ג / מ"ל ופיברינוגן 647 ECM, דגירה ומוכתמת עם calcein-AM ו Hoecsht-33342. התמונות הפלואורסצנטיות מוזגו עם אור משודר (גווני אפור). (ה) X,Z הקרנה של C. (F) X, Z הקרנה של D. תמונות הן פסאודוקולריות. סרגלי קנה מידה ב (A) ו-(B) הם 500 מיקרומטר; סרגלי קנה מידה ב- (C) - (F) הם 10 מיקרומטר. אנא לחץ כאן כדי להציג גירסה גדולה יותר של איור זה.
{kind=link}

איור 5: Cryo-ET של תא BEAS-2B נגוע RSV ברשת קריו-TEM בדוגמת. (A) מפת ריבוע רשת Cryo-EM של תא BEAS-2B נגוע RSV. גבול התא המשוער מצוין על-ידי הקו הירוק המקווקו. (B) תמונה ברזולוציה גבוהה יותר של אזור בקופסה באדום ב- (A). גבול תא משוער מצוין על-ידי קו ירוק מקווקו. נגיפי RSV ניתן לראות ליד פריפריית התא (חץ לבן וקופסה צהובה). (ג) פרוסת z בודדת מטומוגרמה שנאספה באזור הקופסה הצהובה ב- (B). חצים אדומים מצביעים על חלבון היתוך RSV F, חצים כחולים מצביעים על קומפלקס ריבונוקלאופרוטאין (RNP). סרגלי קנה המידה ב- (A)-(C) מוטבעים בתמונה. לחץ כאן כדי להציג גירסה גדולה יותר של איור זה.
{kind=link}

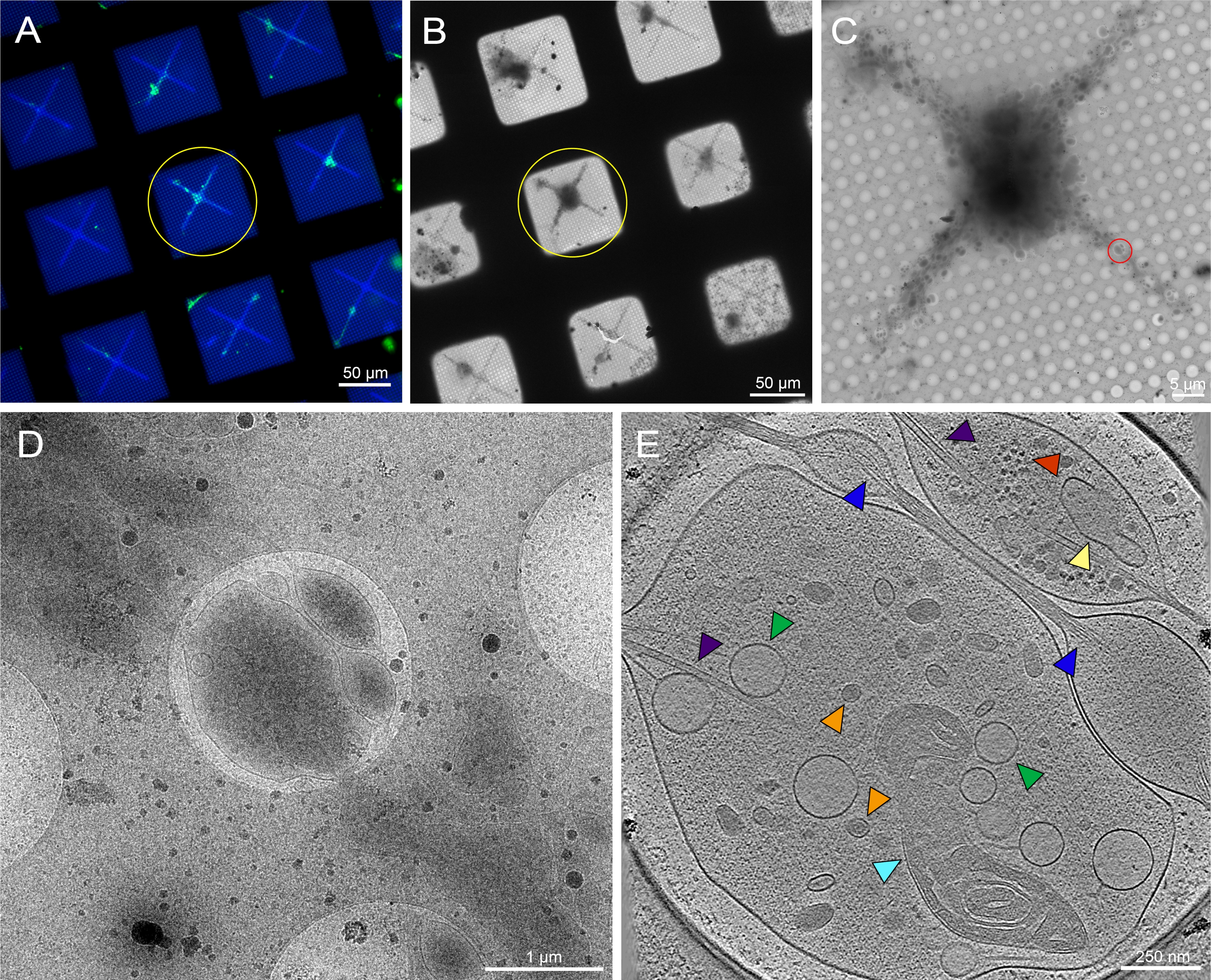
איור 6: נוירונים ראשוניים שמקורם במוחם של זחלי ה-Instar השלישי Drosophila melanogaster על רשת ההקפאה-TEM בדוגמתה. (A) שכבת רשת מיקרואורסצנטיות של פלואורסצנטיות של תאי הפלואורסצנטיים של נוירונים Drosophila המבטאים GFP ממוקד ממברנה בריבועי רשת בדוגמת ריבועי רשת עם 0.5 מ"ג / מ"ל פלואורסצנטית קונבוולין A. ירוק: נוירונים Drosophila. כחול: פוטופטרן. (B) מונטאז' תמונה Cryo-EM של הרשת ב(A) לאחר שימור קריו. עיגול צהוב מציין את אותו ריבוע רשת כמו ב- (A). (C) מונטאז' התמונה Cryo-EM של הריבוע המודגש על-ידי העיגול הצהוב ב- (A) ו- (B). (D) תמונת הגדלה גבוהה יותר של האזור התחום על-ידי העיגול האדום ב- (C), שבו נאספה סדרת הטיה על הנוריטים של התא. ה. פרוסה בעובי 25 ננומטר של טומוגרמה ששוחזרה מסדרת ההטיה שנרכשה מהעיגול האדום ב- (C). אברונים שונים ניתן לראות טומוגרמה זו, כגון המיטוכונדריה (ציאן), microtubules (סגול), שלפוחיות ליבה צפופות (כתום), שלפוחיות אור (ירוק), רשתית אנדופלסמית (צהוב), ו actin (כחול). מקרומולקולות, כגון ריבוזומים (אדום), ניתן לראות גם בפינה הימנית העליונה. תמונות פלואורסצנטיות צבועות בצבעים פסאודו-צבעיים. סרגלי קנה המידה ב- (A)-(E) מוטבעים בתמונה. לחץ כאן כדי להציג גירסה גדולה יותר של איור זה.
{kind=link}

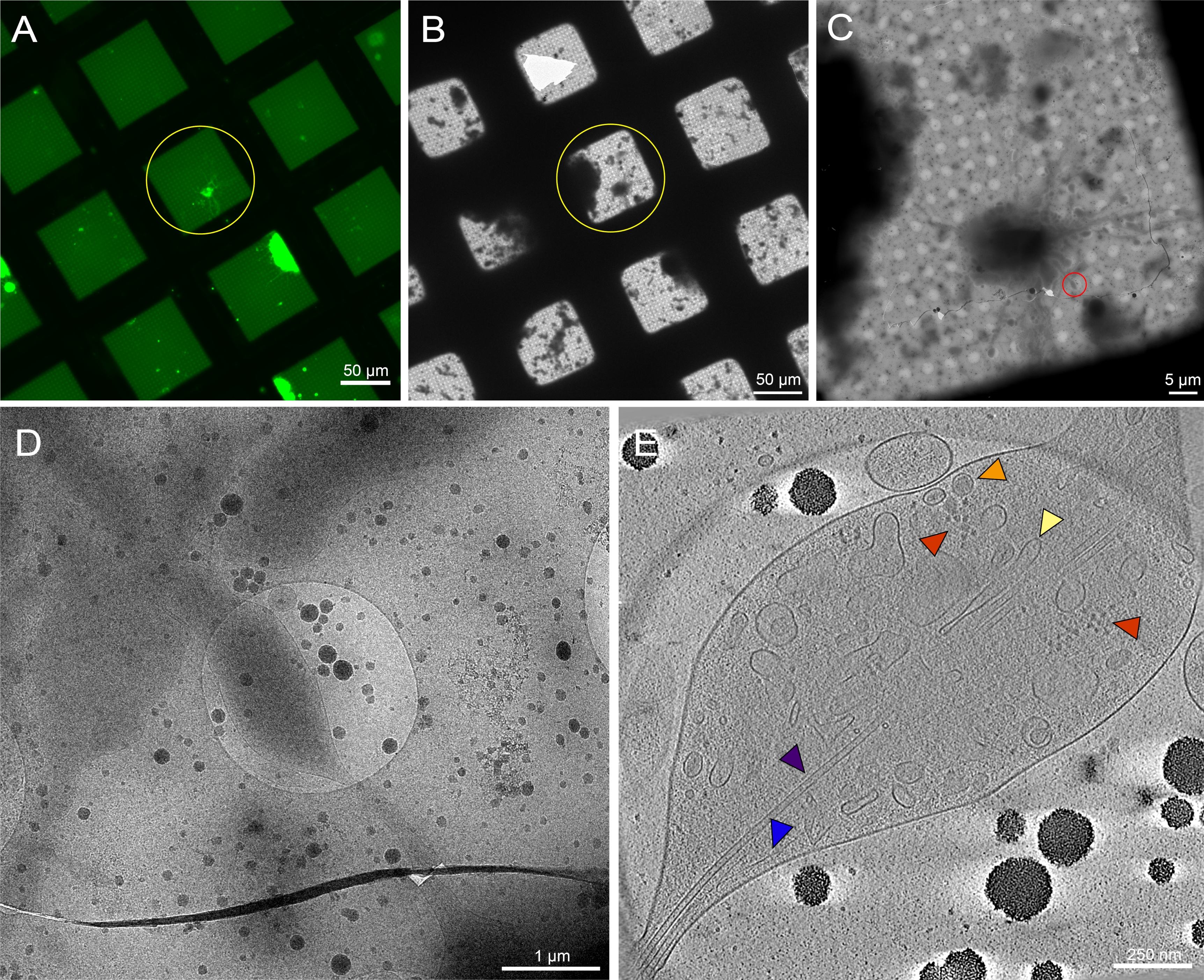
איור 7: נוירונים ראשוניים הנגזרים ממוחו של זחלי הכוכב השלישי דרוזופילה מלנוגסטר על רשתות לא מרוסנות. (A) מיקרואורסצנטיות מיקרואורסצנטיות של רשת פלואורסצנטיות של נוירונים Drosophila המבטאים GFP ממוקד ממברנה בריבועי רשת עם 0.5 מ"ג / מ"ל concanavalin A. ירוק: נוירונים Drosophila . (B) מונטאז' רשת קריו-EM של אותה רשת ב-(A) לאחר הקפאת צלילה. עיגול צהוב מציג את אותו ריבוע רשת כמו ב- (A). שים לב לנוכחות של פסולת סלולרית וזיהום מדיה, מה שהקשה על זיהוי המטרה בהשוואה לרשתות מעוצבות. (ג) מונטאז' התמונה Cryo-EM של הריבוע המודגש על-ידי העיגולים הצהובים במפות (A) ו- (B). (D) תמונת הגדלה גבוהה יותר של האזור התחום על-ידי העיגול האדום ב- (C), שבו נאספה סדרת הטיה על הנוריטים של התא. (E) פרוסה בעובי 25 ננומטר של טומוגרמה משוחזרת מסדרת ההטיה מ- (C) ו- (D). מספר אברונים נראים בטומוגרמה זו, כגון מיקרוטובולים (סגול), אקטין (כחול), הרשתית האנדופלזמית (צהובה) ושלפוחיות ליבה צפופות (כתומות). ניתן לראות גם מקרומולקולות, כגון ריבוזומים (אדום). תמונות פלואורסצנטיות צבועות בצבעים פסאודו-צבעיים. סרגלי קנה המידה ב- (A)-(E) מוטבעים בתמונה. לחץ כאן כדי להציג גירסה גדולה יותר של איור זה.
{kind=link}

איור 8: דוגמאות לבעיות אפשריות בדפוסי דפוס. תמונות פלואורסצנטיות של ECM עם תווית שהופקדו על רשתות מיקרו-מרופדות. (A) דפוסים לא אחידים ברחבי הרשת עקב הפצה לא אחידה של ג'ל PLPP. (B) ל- ECM אין אפשרות לדבוק באזורים המכוסים על-ידי הסטנסיל PDMS במהלך מילוי. (C) תבנית הדרגתית רוויה (צד ימין) או תבנית הפוכה (משמאל) ברשת עם תבנית עם מינון כולל גבוה מדי. (D) ECM דבק באזורים על מוטות הרשת, כמו גם אזור בדוגמת עקב השתקפויות של לייזר UV במהלך דפוס דפוס. התמונות צבועות פסאודו; תבנית קלט מוצגת בפינה הימנית התחתונה; סרגלי קנה מידה הם 100 מיקרומטר. אנא לחץ כאן כדי להציג גרסה גדולה יותר של איור זה.
{kind=link}
| בעיה | גורמים פוטנציאליים | פתרון בעיות |
| Micropatterning | ||
| לא ניתן לראות תאורה מלייזר PRIMO | • נתיב אור אינו מוגדר כראוי | • ודא כי נתיב אור המיקרוסקופ מוגדר כראוי |
| • לייזר PRIMO אינו פועל או שלייזר משולב | ||
| ריבועי רשת שבורים רבים | • נגיעה בנייר כסף עם פינצטה או פיפטה תוך כדי טיפול | • טפלו ברשתות בזהירות |
| • הרשת התייבשה במהלך דגירה או כביסה | • אל תאפשר לרשת להתייבש במהלך שטיפה ודגרה | |
| אזורים גדולים שלא נטענו | • כיסוי ג'ל לא מספיק | • ודאו שג'ל מתפשט באופן שווה על פני הרשת תוך הוספת |
| • רדיד אלומיניום של רשת מחוץ למיקוד במהלך דפוסי דפוס | • הוסיפו מיקרוליטר נוסף של ג'ל | |
| •אזור המכוסה בסטנסיל | • בדוק את המוקד לפני מילוי כל אזור | |
| • רשת מרכזית בקפידה בסטנסיל | ||
| תבנית רוויה או הפוכה | • מינון שגוי | • נסה מגוון של מינונים כוללים עבור דפוס |
| • כיסוי ג'ל לא מספיק | • ודא שהרשת מכוסה באופן שווה בג'ל | |
| • נסה ערכים שונים עבור תבניות בגווני אפור | ||
| תבנית מטושטשת | • מיקוד לקוי במהלך דפוסי דפוס | • כיול PRIMO חוזר באותו גובה כמו מדגם |
| • כיול שגוי | • התמקד בנייר כסף רשת לפני התבנית | |
| • חלוקת תבנית לאזורים נוספים לעיצוב דפוס | ||
| היצמדות ECM מחוץ לתבנית | • השתקפויות מג'ל או אבק | • ודא שהג'ל יבש לפני הדוגמנות |
| • ודא כיסוי ועדשה אובייקטיבית נקיים | ||
| ECM אינו גלוי לאחר דפוס | • הלבנת תמונות | • מזער את החשיפה לאור ל- ECM לפני הדמיה |
| • מינון שגוי במהלך דפוס | • נסה טווח של ערכי מינון כוללים עבור תבנית | |
| • זמן דגירה ECM לא מספיק | • להגדיל את זמן הדגירה עבור ECM | |
| זריעת תאים | ||
| תאים מתגבשים | • מעל העיכול | • השתמש באחוז נמוך יותר של טריפסין או זמן לשחרור של תאים דבקים |
| • צפיפות תאים גבוהה | • מעבר ו/או עיכול תאים במפגש נמוך יותר | |
| • אין להסעיר תאים במהלך השחרור | ||
| • צנרת עדינה של תמיסת תאים או שימוש במסננים של תאים | ||
| תאים שאינם דבקים באזורים עם תבנית | • ECM אינו מתאים לסוג התא | • נסה ריכוזי ECM שונים והרכב |
| • הכדאיות של התאים יורדת לפני הזריעה | • ודא שתנאי תרבית התאים ושחרור התאים אינם פוגעים בתאים | |
| תאים שאינם מתרחבים לאחר הידבקות | • ECM או תבנית שאינם מתאימים לסוג התא | • נסה דפוסים שונים ו- ECM |
| • במקרים מסוימים רדיד אלומיניום רציף יותר (R1.2/20 לעומת R2/1) עשוי לקדם הרחבת תאים |
טבלה 1: בעיות פוטנציאליות במהלך micropatterning. טבלה זו מתארת בעיות מסוימות שמשתמש עלול להיתקל בהן במהלך מיקרו-פצצת תאים או זריעת תאים. סיבות פוטנציאליות ופתרון בעיות מסופקות עבור כל בעיה. תמונות מייצגות של כמה בעיות ניתן לראות באיור 8.
Discussion
מיקרוסקופ אלקטרונים וחבילות תוכנה מודרניות ומתקדמות תומכות כעת באיסוף נתוני קריו-EM והקפאה-ET אוטומטיים יעילים שבהם ניתן לפלח ולדמיין מאות עד אלפי משרות בתוך ימים ספורים32,33,33,34,35. אחד הגורמים המגבילים המשמעותיים לזרימות עבודה של קריו-ET של תאים שלמים היה השגת מספר מספיק של יעדי אספנות לכל רשת. לאחרונה, מספר קבוצות פיתחו פרוטוקולים עבור רשתות micropatterning עבור cryo-EM, עם יתרון אחד להיות יעילות איסוף נתונים משופרת16,17,18. כאן מוצג פרוטוקול לשימוש במערכת micropatterning זמינה מסחרית לרשתות TEM micropattern למחקרי קריו-ET של נוירונים Drosophila ראשוניים וקווי תאים אנושיים מתורבתים (נגועים או נגועים RSV). מערכת micropatterning זו היא רב-תכליתית וניתן למטב שלבים רבים ולהתאים אותם כך שיתאימו למטרות ניסיוניות ספציפיות. משתמש עם ניסיון במיקרוסקופיה של TEM ופלואורסצנטיות יכול להיות מיומן במהירות בהכנת רשת ובמיקרו-פצפוצים. עם תרגול זהיר, תוצאות טובות צריכות להיות ברות השגה לאחר כמה איטרציות. להלן, חלק מהאפשרויות הזמינות, שיקולי משתמש, יתרונות פוטנציאליים, ויישומים עתידיים של micropatterning עבור cryo-EM נדונים.
אחד השיקולים החשובים עבור קריו-ET תא שלם הוא בחירת רשת EM. רשתות EM מורכבות משני חלקים: מסגרת רשת שינוי (או תמיכה מבנית) ונייר הכסף (או הסרט), שהוא משטח הסרט הרציף או החורי שעליו יגדלו התאים. רשתות רשת נחושת משמשות בדרך כלל להקפאה-EM של חלבונים ומתחמים מבודדים. עם זאת, הם אינם מתאימים להקפאה-ET של תאים שלמים בשל הציטוטוקסיות של נחושת. במקום זאת, רשת זהב משמשת בדרך כלל לטומוגרפיה תאית. אפשרויות אחרות כוללות ניקל או טיטניום, אשר עשוי לספק יתרונות על פני זהב כגון קשיחות מוגברת16. רשתות EM זמינות עם מידות רשת שינוי שונות כדי לתמוך במגוון יישומים. גדלי רשת גדולים יותר מספקים יותר מקום לתאים לגדול בין סרגלי רשת ואזורים נוספים הניתנים לאיסוף סדרות הטיה, אם כי במחיר של שבריריות דגימה כוללת מוגברת. רדיד הכסף הנפוץ ביותר הוא פחמן אמורפי מחורר או חורי, כגון Quantifoils או רשתות C שטוחות. ניתן לדמיין מטרות ביולוגיות דרך החורים בפחמן או דרך הפחמן שקוף האלקטרונים. רשתות כגון R 2/1 או R 2/2, שבו החורים הם 2 מיקרומטר רוחב כי הם מרווחים 1 ו 2 מיקרומטר זה מזה בהתאמה, לספק מספר רב של חורים ובכך מספר רב של אזורים פוטנציאליים לאיסוף נתונים. עם זאת, תאים מסוימים עשויים לגדול ולהתרחב טוב יותר על משטחים אחידים יותר כגון רשתות R 1.2/20 או פחמן רציף. לעיבוד מדגם במורד הזרם על ידי כרסום קרן יונים ממוקד (קריו-FIB), נייר הכסף מוסר באמצעות כרסום, מה שמפחית את החששות לגבי המשך הנוכחות של הסרט הבסיסי. כמו עם הרשת, רדידים מחומרים אחרים זמינים גם, עם פרוטוקול דפוס המוצג כאן להיות מתאים באותה מידה עבור רשתות SiO2 . רשתות נפוצות כוללות Quantifoil זהב, פחמן רציף, או SiO2 סרט 200-רשת רשתות (~ 90 מיקרומטר מרווח בין סרגלי רשתות) עבור קריו-ET של תאים שלמים.
ישנם מספר שיקולים בעת עיצוב תבנית. רוב ההחלטות הללו מונחות על ידי סוג התא ומטרתו של הניסוי. נקודת התחלה טובה היא לבחור תבנית שמעריכה את הצורה והממדים של התאים בתרבית. מחקרים רבים הראו השפעות משמעותיות של צורת דפוס על צמיחת התאים וסידור ציטו-שלד13,36,37. יש לנקוט בזהירות מיוחדת במהלך עיצוב דפוס אם זה יכול לשנות את היעד של עניין. מספר דפוסים עבור כל סוג תא נבדקו כדי לקבוע אילו דפוסים קידמו הידבקות וצמיחה תאית. הגמישות של מערכת micropatterning מאפשרת בדיקה של דפוסים מרובים על רשת אחת ושינוי דפוסים עבור רשתות שונות בתוך ניסוי אחד. דפוסים גדולים יותר (~ 50-90 מיקרומטר), כגון אלה המשמשים כאן, להגדיל את הסבירות כי תאים מרובים לדבוק באזור אחד של התבנית ולאפשר לתאים להרחיב ולהאריך לאחר הידבקות. דפוסים מוגבלים יותר (20-30 מיקרומטר) עשויים להיות מתאימים בניסויים שבהם בידוד תאים הוא קריטי יותר מאשר הרחבת תאים, כגון לניסויים ממוקדים של כרסום קרן יונים (קריו-FIB). עבור יישומי טומוגרפיה, ייתכן שיהיה צורך לשקול את ההשפעה של ציר ההטיה. אם תבנית ממוקמת כך שכל התאים גדלים במקביל זה לזה בכיוון אחד, ייתכן שכל התאים יהיו מאונכים לציר ההטיה כאשר הם נטענים על במת המיקרוסקופ, וכתוצאה מכך איכות נמוכה יותר של נתונים.
ברשתות לא מרוסנות, תאים לעתים קרובות מעדיפים לדבוק בפסי הרשת, שם לא ניתן לדמיין אותם על-ידי TEM. אפילו על רשתות בדוגמת דפוס, תאים נצפים לעתים קרובות להיות ממוקם בפינות של ריבועי רשת חלקית על רדיד פחמן דפוס וסרגל רשת. לאחרונה, micropatterning שימש בכוונה למקם חלק מהתא מעל סרגל הרשת18. זה יכול להיחשב לניסויים שבהם זה לא קריטי יש פריפריה התא כולו על נייר הכסף. זה יכול להיות חשוב במיוחד עבור תאים שיכולים לגדול גדול יותר מאשר ריבוע רשת יחיד, כגון נוירונים ראשוניים גדל על פני מספר ימים.
ישנם כלים רבים שניתן להשתמש בהם לעיצוב תבנית. כאן, התבנית הוגבלה לפחות מ-800 פיקסלים בכל ממד, כך שניתן לסובב את התבנית לכל זווית ועדיין להתאים אותה לאזור המרבי שיכול להיות מעוצב בהקרנה אחת על ידי מערכת מיקרו-פצפוצים זו. זה מאפשר למשתמש לסובב את התבנית כדי להיות מכוון כראוי עם הרשת ללא קשר לכיוון הרשת במיקרוסקופ. כאן, הרשת חולקה לשישה אזורי דפוס. בראש ובראשונה, הדבר מאפשר התאמת מוקד בין אזורים שונים של הרשת. רשתות זהב, בפרט, הן נזילות מאוד ולא יכול לשכב לגמרי שטוח על הזכוכית. מיקוד נכון חיוני לתוצאות תבניות נקיות ומעודנות. על-ידי שימוש בתבניות מפולחות, יש לבצע רק התאמות קלות למיקום התבנית אם הרשת משתנה מעט במהלך תהליך התבנית, אם כי בדרך כלל זו אינה בעיה בעת שימוש בג'ל PLPP עם סטנסילים PDMS. לבסוף, ארבעת ריבועי הרשת המרכזיים של הרשת נותרו לא מרוסנים. זה תומך משתמש להיות מסוגל לזהות בבירור את מרכז הרשת, וזה מאוד שימושי עבור ניסויי הדמיה מתאם.
תוכנת דפוס עבור מערכת micropatterning זו, לאונרדו, יש גם תכונות מתקדמות יותר כגון תפירה ואת היכולת לייבא דפוסים כמו מסמכי PDF, אשר מעבר להיקף של פרוטוקול זה. תוכנה זו כוללת גם זיהוי מיקרו-מבנים ומיקום דפוס אוטומטי שניתן להשתמש בו ברשתות TEM. תכונה זו שימושית ביותר כאשר הרשת שטוחה מאוד וניתן לעצב אותה ללא צורך להתאים את המוקד בין אזורים שונים.
בחירת חלבון ECM יכולה להשפיע באופן משמעותי על הידבקות והתרחבות התאים. תאים מסוימים ידועים לעבור שינויים פיזיולוגיים כאשר גדל על מצעים ספציפיים38. חלבונים וריכוזים מרובים של ECM נבדקו עבור כל סוג תא חדש בהתבסס על עבודה קודמת שדווחה בספרות. למינין, פיברינוגן, פיברונקטין וקולגן נמצאים בשימוש נרחב עבור תאים בתרבית וניתן להשתמש בהם כנקודת התחלה אם נתונים אחרים אינם זמינים. עם זאת, חלבוני ECM אחרים חייבים להיחשב גם אם חלבוני ECM הנפוצים אינם מצליחים להעניק תכונות דבקות נאותות לתאים. זה היה נכון במיוחד עבור נוירונים Drosophila העיקריים, כמו ריכוז גבוה של הצמח lectin concanavalin A היה הכרחי לדבקות תאית נכונה. התאימות של הידבקות סלולרית וצמיחה עם ECM ניתן לבדוק על ידי דפוס על צלחות זכוכית או שקופיות לפני המעבר לרשתות TEM. גישת טרום הקרנה זו היא זמן וחסכוני אם יש צורך לבחון מספר רב של שילובים. הכללת חלבון ECM מצומד פלואורסצנטי היא בעלת ערך להערכת ההצלחה והאיכות של דפוסי דפוס.
זריעת תאים היא אחד השלבים החשובים ביותר עבור קריו-ET תא שלם, עם או בלי micropatterning6,16,39. עבור Drosophila העיקרי או נוירונים אחרים, אשר שבירים, לא יציבים בהשעיה, ועשויים להיות מוגבלים בכמות, גישות זריעה בודדות עדיפות על פני זריעת תאים מנוטרת ורציפה. שלב זריעה יחיד בצפיפות תאים ממוטבת, כמתואר בפרוטוקול עבור נוירונים Drosophila, הוא אפשרות מעשית עבור רוב סוגי התאים. עם זאת, ניתן גם לזרוע תאים על המצע בריכוז ראשוני נמוך יותר ולהוסיף תאים נוספים בצורה מנוטרת כמתואר כאן ובספרות אחרת18. זריעה רציפה זו יכולה לספק תוצאות עקביות יותר במקרים מסוימים. בדומה לתרבות תאים סטנדרטית, תמיד יש להקפיד לשמור על כדאיות התא ולמזער את גושי התאים במהלך הבידוד.
כאשר מתחילים לראשונה עם micropatterning, יש כמה מלכודות פוטנציאליות כי הם מזיקים לתוצאה הסופית. טיפול זהיר ברשת וטכניקה סטרילית, הפצה אחידה של ג'ל PLPP, מינון ומיקוד נכונים במהלך דפוס, ושמירה על הכדאיות של התא לפני זריעה הם בין השיקולים החשובים ביותר להצלחה. רשימה של חלק מהבעיות הפוטנציאליות וכן פתרונות הורכבו בטבלה 1.
ניתן להשתמש ברשתות micropatterned כדי לסייע במיקום תאים כדי ליצור צפיפות תאים עקבית ברחבי הרשת ולמקם אזורי עניין באזורים המתאימים לאיסוף סדרת הטיה16,18. המיקום והמיצוב של תאים יכול לשמש כסמנים fiducial למתאם בניסויים קריו-CLEM, הפחתת הצורך ברשתות תיווך פתיל שבירות סמנים fiducial פלואורסצנטי. עם זאת, יש לציין כי סמנים fiducial כאלה עדיין עשויים להיות שימושיים עבור מתאם דיוק תת מיקרומטר29,40. יתר על כן, התפלגות אחידה של תאים מבודדים מועילה מאוד גם לניסויי כרסום קרן מיקוד-יון (קריו-FIB) כדי למקסם את מספר התאים שמהם ניתן לחתוך lamella16.
התוספת של מיקרו-מפזרינג לזרימות עבודה של קריו-EM תגרום לשיפורים מדידים בתפוקת הנתונים ועשויה לאפשר ניסויים חדשים. ככל שהטכניקה מאומצת ומפותחת עוד יותר, יישומים מתקדמים יותר של מיקרו-פטרנינג כולל שיפועי ECM, תצהירי ECM מרובים והרכבת מיקרו-מבנה ירחיבו עוד יותר את היכולות של cryo-ET ללמוד מטרות ותהליכים ביולוגיים בהקשר תאי מלא.
Disclosures
למחברים אין מה לחשוף.
Acknowledgements
אנו מודים לד"ר ג'יל ווילדונגר, ד"ר סיהוי ז. יאנג, וגברת ג'וזפין ו. מיטשל במחלקה לביוכימיה, אוניברסיטת ויסקונסין, מדיסון על שיתוף נדיב של אלב-גל4, UAS-CD8::GFP זן זבובים (מרכז מלאי בלומינגטון, #5146). ברצוננו גם להודות לד"ר אורליאן דובוין, מר לורנט סיקייר, וגברת מארי-שרלוט מאנוס מאלבול ומר סרג' קדורה ממעבדות ננומטרי על תמיכתם הנדיבה במהלך פרויקט זה. עבודה זו נתמכה בחלקה על ידי אוניברסיטת ויסקונסין, מדיסון, המחלקה לביוכימיה באוניברסיטת ויסקונסין, מדיסון, ושירות בריאות הציבור מעניק R01 GM1114561, R01 GM104540, R01 GM104540-03W1, ו U24 GM139168 ל- E.R.W. ו R01 AI150475 ל- P.W.S. מה- NIH. חלק ממחקר זה נתמך על ידי מענק NIH U24 GM129547 ובוצע ב- PNCC ב- OHSU ונגיש באמצעות EMSL (רשת.436923.9), משרד DOE של מתקן משתמש מדעי בחסות המשרד למחקר ביולוגי וסביבתי. אנו מודים גם על השימוש במתקנים ומכשורים במרכז המחקר Cryo-EM במחלקה לביוכימיה באוניברסיטת ויסקונסין, מדיסון.
Materials
| Name | Company | Catalog Number | Comments |
| 0.1% (w/v) Poly-L-Lysine | Sigma | P8920-100ML | |
| 0.22 µm syringe filters PVDF membrane | Genesee | 25-240 | |
| 22x60-1 Glass cover slip | Fisher | 12545F | |
| 5/15 Tweezers | EMS (Dumont) | 0203-5/15-PO | |
| Antibiotic-Antimycotic (100X) | ThermoFisher (Gibco) | 15240096 | |
| BEAS-2B cells | ATCC | CRL-9609 | |
| Collagen I, bovine | ThermoFisher (Gibco) | A1064401 | |
| Concanavalin A, Alexa Fluor 350 Conjugate | ThermoFisher (Invitrogen) | C11254 | |
| DMEM | Fisher (Lonza) | BW12-604F | |
| EtOH | Fisher (Decon Labs) | 22-032-600 | |
| Fetal Bovine Serum | ATCC | 30-2020 | |
| Fibrinogen From Human Plasma, Alexa Fluor 647 Conjugate | ThermoFisher (Invitrogen) | F35200 | |
| Fibronectin Bovine Protein, Plasma | ThermoFisher (Gibco) | 33010018 | |
| Glass bottom dish | MatTek | P35G-1.5-20-C | |
| Glucose | VWR | 0643-1KG | |
| Grid prep holder | EMS | 71175-01 | |
| HeLa cells | ATCC | CCL-2 | |
| Hemacytometer | Fisher (SKC, Inc.) | 22600100 | |
| HEPES | Fisher (ACROS Organics) | AC172572500 | |
| Hoechst 33342 | ThermoFisher (Invitrogen) | H3570 | |
| Insulin | Fisher (Sigma Aldrich) | NC0520015 | |
| KCl | MP Bio | 194844 | |
| KH2PO4 | Fisher (ACROS Organics) | AC212595000 | |
| Leica-DMi8 | Leica Microsystems | Can be customized with camera, stage, and objective attachments | |
| Leonardo | Alvéole | https://www.alveolelab.com/our-products/leonardo-photopatterning-software/ | |
| Liberase Research Grade | Fisher (Supply Solutions) | 50-100-3280 | |
| LIVE/DEAD Viability/Cytotoxicity Kit | ThermoFisher (Invitrogen) | L3224 | |
| Microscope camera | Hammamatsu | C13440-20CU | |
| Motorized stage | Märzhäuser Wetzlar | 00-24-599-0000 | |
| NaCl | Fisher (Fisher BioReagents) | BP358-1 | |
| NaH2PO4 | Fisher (ACROS Organics) | AC207802500 | |
| NaOH | Fisher (Alfa Aesar) | AAA1603736 | |
| PBS | Corning | 21-040-CV | |
| PDMS stencils | nanoscaleLABS | PDMS_STENCILS_EM | https://www.alveolelab.com/our-products/pdms-stencil-multiwell-plate/ |
| PEG-SVA | nanoscaleLABS | PEG-SVA-1GR | mPEG-Succinimidyl Valerate, MW 5,000 |
| Penicillin | Fisher (Research Products International Corp) | 50-213-641 | |
| pH strips | Fisher (Millipore Sigma) | M1095350001 | pH probe can also be used |
| PLPP gel | nanoscaleLABS | PLPP-GEL-300UL | https://www.alveolelab.com/our-products/plpp-photoactivatable-reagent/ |
| PRIMO | Alvéole | https://www.alveolelab.com/our-products/primo-micropatterning/ | |
| pSynkRSV-I19F (BAC containing RSV A2-mK+ antigenomic cDNA ) | BEI Resources | NR-36460 | https://www.beiresources.org/Catalog/BEIPlasmidVectors/NR-36460.aspx |
| Quantifoil grids | EMS (Quantifoil) | Q2100AR1 | 2 µm holes spaced 1 µm apart, other dimensions are available |
| RPMI | Fisher (Lonza) | BW12-702F | |
| RSV A2-mK+ | see entry for pSynkRSV-19F | - | Described in Hotard et al. [22]. Can be generated from pSynkRSV-ll9F |
| Schneider's Media | ThermoFisher (Gibco) | 21720-024 | |
| SerialEM | SerialEM (https://bio3d.colorado.edu/SerialEM/ ) | https://bio3d.colorado.edu/SerialEM/ | |
| Straight tweezers | EMS (Dumont) | 72812-D | |
| Streptomycin | Fisher (Fisher BioReagents) | BP910-50 | |
| Sucrose | Avantor | 4097-04 | |
| Tetracycline | Sigma | T8032-10MG | |
| Titan Krios electron microscope | ThermoFisher | 300kV, with direct electron detector camera and energy filter | |
| Trypsin | ThermoFisher (Gibco) | 15090046 | |
| Tube Revolver/Rotator | Fisher (Thermo Scientific) | 11676341 | |
| UAS:mcD8:GFP Drosophila fly strain | Bloomington Drosophila Stock Center | 5146 | http://flybase.org/reports/FBtp0002652.html |
References
- Nogales, E., Scheres, S. H. Cryo-EM: A unique tool for the visualization of macromolecular complexity. Molecular Cell. 58 (4), 677-689 (2015).
- Martynowycz, M. W., Gonen, T. From electron crystallography of 2D crystals to MicroED of 3D crystals. Current Opinion in Colloid and Interface Science. 34, 9-16 (2018).
- Wagner, J., Schaffer, M., Fernandez-Busnadiego, R. Cryo-electron tomography-the cell biology that came in from the cold. FEBS Letters. 591 (17), 2520-2533 (2017).
- Wan, W., Briggs, J. A. Cryo-electron tomography and subtomogram averaging. Methods in Enzymology. 579, 329-367 (2016).
- Bäuerlein, F. J., Pastor-Pareja, J. C., Fernández-Busnadiego, R. Cryo-electron tomography of native Drosophila tissues vitrified by plunge freezing. bioRxiv. , 437159(2021).
- Hampton, C. M., et al. Correlated fluorescence microscopy and cryo-electron tomography of virus-infected or transfected mammalian cells. Nature Protocols. 12 (1), 150-167 (2017).
- Hsieh, C. E., Leith, A., Mannella, C. A., Frank, J., Marko, M. Towards high-resolution three-dimensional imaging of native mammalian tissue: Electron tomography of frozen-hydrated rat liver sections. Journal of Structural Biology. 153 (1), 1-13 (2006).
- Al-Amoudi, A., Norlen, L. P., Dubochet, J. Cryo-electron microscopy of vitreous sections of native biological cells and tissues. Journal of Structural Biolology. 148 (1), 131-135 (2004).
- Rigort, A., et al. Focused ion beam micromachining of eukaryotic cells for cryoelectron tomography. Proceedings of the National Academy of Sciences. 109 (12), 4449-4454 (2012).
- Gorelick, S., et al. PIE-scope, integrated cryo-correlative light and FIB/SEM microscopy. Elife. 8, 45919(2019).
- Wu, G. H., et al. Multi-scale 3D cryo-correlative microscopy for vitrified cells. Structure. 28 (11), 1231-1237 (2020).
- Turk, M., Baumeister, W. The promise and the challenges of cryo-electron tomography. FEBS Letters. 594 (20), 3243-3261 (2020).
- Théry, M. Micropatterning as a tool to decipher cell morphogenesis and functions. Journal of Cell Science. 123 (24), 4201-4213 (2010).
- Tseng, Q., et al. Spatial organization of the extracellular matrix regulates cell-cell junction positioning. Proceedings of the National Academy of Sciences. 109 (5), 1506-1511 (2012).
- Hardelauf, H., et al. Micropatterning neuronal networks. Analyst. 139 (13), 3256-3264 (2014).
- Toro-Nahuelpan, M., et al. Tailoring cryo-electron microscopy grids by photo-micropatterning for in-cell structural studies. Nature Methods. 17 (1), 50-54 (2020).
- Engel, L., et al. Extracellular matrix micropatterning technology for whole cell cryogenic electron microscopy studies. Journal of Micromechanics and Microengineering. 29 (11), (2019).
- Engel, L., et al. Lattice micropatterning for cryo-electron tomography studies of cell-cell contacts. bioRxiv. , 272237(2021).
- Sibert, B. S., Kim, J. Y., Yang, J. E., Wright, E. R. Whole-cell cryo-electron tomography of cultured and primary eukaryotic cells on micropatterned TEM grids. bioRxiv. , 447251(2021).
- Egger, B., van Giesen, L., Moraru, M., Sprecher, S. G. In vitro imaging of primary neural cell culture from Drosophila. Nature Protocols. 8 (5), 958-965 (2013).
- Lu, W., Lakonishok, M., Gelfand, V. I. Kinesin-1-powered microtubule sliding initiates axonal regeneration in Drosophila cultured neurons. Molecular Biology of the Cell. 26 (7), 1296-1307 (2015).
- Ke, Z., et al. The morphology and assembly of respiratory syncytial virus revealed by cryo-electron tomography. Viruses. 10 (8), (2018).
- Stobart, C. C., et al. A live RSV vaccine with engineered thermostability is immunogenic in cotton rats despite high attenuation. Nature Communications. 7, 13916(2016).
- Hotard, A. L., et al. A stabilized respiratory syncytial virus reverse genetics system amenable to recombination-mediated mutagenesis. Virology. 434 (1), 129-136 (2012).
- Schindelin, J., et al. Fiji: an open-source platform for biological-image analysis. Nature Methods. 9 (7), 676-682 (2012).
- Mastronarde, D. N. Automated electron microscope tomography using robust prediction of specimen movements. Journal of Structural Biology. 152 (1), 36-51 (2005).
- Kremer, J. R., Mastronarde, D. N., McIntosh, J. R. Computer visualization of three-dimensional image data using IMOD. Journal of Structural Biology. 116 (1), 71-76 (1996).
- Tang, G., et al. EMAN2: an extensible image processing suite for electron microscopy. Journal of Structural Biology. 157 (1), 38-46 (2007).
- Yang, J. E., Larson, M. R., Sibert, B. S., Shrum, S., Wright, E. R. CorRelator: Interactive software for real-time high precision cryo-correlative light and electron microscopy. Journal of Structural Biology. , 107709(2021).
- Ke, Z., et al. Promotion of virus assembly and organization by the measles virus matrix protein. Nature Communications. 9 (1), 1736(2018).
- Kim, J., Yang, S., Wildonger, J., Wright, E. A new in situ neuronal model for cryo-ET. Microscopy and Microanalysis. 26 (2), 130-132 (2020).
- Bouvette, J., et al. Beam image-shift accelerated data acquisition for near-atomic resolution single-particle cryo-electron tomography. Nature Communications. 12 (1), 1957(2021).
- Schorb, M., Haberbosch, I., Hagen, W. J. H., Schwab, Y., Mastronarde, D. N. Software tools for automated transmission electron microscopy. Nature Methods. 16 (6), 471-477 (2019).
- Weis, F., Hagen, W. J. H., Schorb, M., Mattei, S. Strategies for optimization of cryogenic electron tomography data acquisition. Journal of Visual Experiments. (169), e62383(2021).
- Chreifi, G., Chen, S., Jensen, G. J. Rapid tilt-series method for cryo-electron tomography: Characterizing stage behavior during FISE acquisition. Journal of Structural Biology. 213 (2), 107716(2021).
- Anderson, D. E., Hinds, M. T. Endothelial cell micropatterning: methods, effects, and applications. Annals of Biomedical Engineering. 39 (9), 2329-2345 (2011).
- McWhorter, F. Y., Wang, T., Nguyen, P., Chung, T., Liu, W. F. Modulation of macrophage phenotype by cell shape. Proceedings of the National Academy of Sciences. 110 (43), 17253-17258 (2013).
- Kleinman, H. K., Luckenbill-Edds, L., Cannon, F. W., Sephel, G. C. Use of extracellular matrix components for cell culture. Analytical Biochemistry. 166 (1), 1-13 (1987).
- Fassler, F., Zens, B., Hauschild, R., Schur, F. K. M. 3D printed cell culture grid holders for improved cellular specimen preparation in cryo-electron microscopy. Journal of Structural Biology. 212 (3), 107633(2020).
- Schellenberger, P., et al. High-precision correlative fluorescence and electron cryo microscopy using two independent alignment markers. Ultramicroscopy. 143, 41-51 (2014).
Reprints and Permissions
Request permission to reuse the text or figures of this JoVE article
Request PermissionThis article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. All rights reserved