
Method Article
전세포 극저온-전자 단층 촬영 워크플로우 내의 직접 세포 위치지정에 마이크로패턴 전송 전자 현미경 검사기 그리드
* 이 저자들은 동등하게 기여했습니다
요약
이 프로토콜의 목표는 극저온 전자 현미경 검사법을 위한 그리드의 표적 영역에 세포 접착 및 성장을 지시하는 것입니다. 이는 세포 종자 전에 패턴화된 영역에서 세포외 매트릭스 단백질의 증착에 따라 사용자 지정 패턴에서 절제되는 항 파울링 층을 적용함으로써 달성된다.
초록
전세포 극저온 전자 단층 촬영(cryo-ET)은 세포 컨텍스트에 존재하고 거의 네이티브 냉동 수화 상태로 보존된 거대 분자의 나노미터 수준의 해상도 구조를 생성하는 데 사용되는 강력한 기술입니다. 그러나, 그들의 생리적인 상태에 있는 세포를 유지하면서 단층 촬영에 적합한 방식으로 TEM 격자에 세포를 배양 및/또는 준수와 관련되었던 도전이 있습니다. 여기서, TEM 그리드에서 진핵 세포 성장을 직접 및 촉진하기 위해 마이크로패터닝을 사용하는 방법에 대한 상세한 단계별 프로토콜이 제시된다. 마이크로패터닝 하는 동안, 세포 성장은 다른 지역이 안티 폴링 층으로 코팅된 채로 남아있는 동안 TEM 그리드의 호일에 지정된 패턴 및 위치 내에서 세포 매트릭스(ECM) 단백질을 증착하여 지시된다. 표면 코팅 및 패턴 설계의 선택의 유연성은 마이크로 패터닝을 광범위한 세포 유형에 광범위하게 적용할 수 있게 합니다. 마이크로패터닝은 개별 세포 내의 구조뿐만 아니라 숙주 병원체 상호 작용 또는 분화 된 다세포 커뮤니티와 같은 보다 복잡한 실험 시스템에 유용합니다. 마이크로패터닝은 또한 코르타상대광 및 전자 현미경검사법(cryo-CLEM) 및 집중이온 빔 밀링(cryo-FIB)을 포함한 많은 하류 전세포 냉동고-ET 워크플로우에 통합될 수 있다.
서문
저온 전자 현미경 검사법(cryo-EM)의 개발, 확장 및 다기능성으로 연구자들은 거시분자(~1nm)에서 고(~2Å) 해상도에 가까운 네이티브 상태의 광범위한 생물학적 샘플을 조사했습니다. 단일 입자 저온-EM 및 전자 회절 기술은 용액 또는 결정 상태에서 각각 1,2에서 정제 된 거대 분자에 가장 잘 적용됩니다. 냉동 전자 단층 촬영 (극저온-ET)은 박테리아, 흉피 바이러스 및 진핵 세포와 같은 대형 이종 물체의 거의 네이티브 구조 및 초구조 적 연구에 독특하게 적합합니다3. 저온-ET에서, 3차원(3D) 정보는 현미경 단계에서 샘플을 물리적으로 기울이고 샘플시리즈를 상이한 각도로 수집하여 얻어진다. 이러한 이미지 또는 틸트 계열은 종종 1~3도 단위로 +60/-60도의 범위를 다룹니다. 그런 다음 틸트 계열은 토모그램4라고도 하는 3D 볼륨으로 계산하여 재구성할 수 있습니다.
모든 저저온-EM 기술은 비정질, 비결정, 유리체 얼음의 얇은 층에 샘플을 포함시켜야 합니다. 가장 일반적으로 사용되는 극저온 고정 기술 중 하나는 샘플이 EM 그리드에 적용되고, 블로드되고, 액체 에탄 또는 액체 에탄과 프로판의 혼합물로 빠르게 급락하는 급락 동결입니다. 이 기술은 HeLa 세포5,6와 같은 배양 인간 세포를 포함하여 두께가 <100 nm에서 ~10 μm까지의 시료의 진동에 충분하다. 두께가 최대 200 μm인 미니 오가노이드 또는 조직 생검과 같은 두꺼운 샘플은 고압 동결에 의해 진동될 수 있다7. 그러나, 두꺼운 시료의 전자 산란 증가로 인해, 극저온-ET에 대한 샘플 및 얼음 두께는 300kV 전송 전자 현미경에서 ~0.5-1 μm로 제한됩니다. 따라서, 많은 진핵세포의 전세포 극저온-ET는 극저온-절개8 또는 집중이온 빔 밀링9,10,11과 같은 추가 시료 준비 단계가 사용되지 않는 한 세포 주변 또는 세포의 확장에 한정된다.
많은 전세포 저온-ET 이미징 실험의 한계는 데이터 수집 처리량12입니다. 수천 개의 격리된 입자가 단일 TEM 그리드 정사각형에서 종종 이미지화될 수 있는 단일 입자 저온-EM과 달리 세포는 크고 확산되며, 세포가 얇은 유리체 층에서 보존될 수 있도록 충분히 낮은 밀도로 재배되어야 합니다. 종종 관심 영역은 셀의 특정 피쳐 또는 하위 영역으로 제한됩니다. 추가 처리량을 제한하는 것은 TEM 그리드 바 의 온 또는 그 근처에서와 같은 TEM 이미징에 대해 불가능한 영역에서 성장하는 세포의 성향입니다. TEM 그리드의 세포 배양과 관련된 예측할 수 없는 요인으로 인해 데이터 수집을 위한 샘플 접근성 및 처리량을 개선하기 위해 기술 개발이 필요합니다.
부착 된 세포 매트릭스 (ECM) 단백질을 사용한 기판 미세 패턴은 유리 및 기타 조직 배양 기판과 같은 경직되고 내구성이 뛰어나며 광학적으로 투명한 표면에서 세포의 성장을 지시하기 위한 살아있는 세포 광 현미경 검사법을 위한 잘 확립된 기술입니다13,14. 마이크로패터닝은 또한 연약및/또는 3차원(3D) 표면에서 수행되었습니다. 이러한 기술은 세포의 정확한 위치 지정을 허용했을 뿐만 아니라; 그들은 또한 패턴 신경 세포 회로15와 같은 다세포 네트워크의 생성을 지원했습니다. 저온 ET에 마이크로패턴을 도입하면 처리량이 증가할 뿐만 아니라 복잡하고 역동적인 세포 마이크로환경을 탐색하기 위한 새로운 연구도 열 수 있습니다.
최근에는 여러 접근 방식을 통해 TEM 그리드에서 마이크로패터닝 기술을 사용하기 시작한 여러 그룹이 16,17입니다. 여기서, TEM 그리드용 마스크리스 포토패팅 기법을 사용하는 것은 고해상도 및 비접촉식 패터닝을 특징으로 하는 Alvéole PRIMO 마이크로패터닝 시스템을 사용하여 설명된다. 이 마이크로패턴 시스템을 사용하면 기판 상단에 안티 폴링 층이 적용되고, 이어서 자외선 레이저를 사용하는 사용자 정의 패턴에서 광촉매 및 안티 폴링 레이어의 절제를 적용한다. ECM 단백질은 적절한 세포 배양을 위한 패턴에 첨가될 수 있다. 이 방법은 망막 안료 상피-1 (RPE1), Madin-Darby 개 신장 II (MDCKII), 인간 포피 피부 섬유아세포 (HFF), 및 내피 세포 주 16,17,18의 저온 ET 연구를 위해 여러 그룹에 의해 사용되었습니다. 이 마이크로패턴 시스템은 다중 항오염층 기판뿐만 아니라 액체 또는 젤 광촉매 시약과 호환됩니다. 다양한 ECM 단백질을 선택 및 세포선의 특이성에 맞게 조정될 수 있으며, 사용자에게 다재다능함을 부여한다.
마이크로패터닝은 실험실내에서 다수의 프로젝트에 성공적으로 적용되었습니다19. 여기서, 마이크로패턴 프로토콜은 배양된 HeLa 세포를 연구하기 위한 특정 적응, 호흡 싱크성 바이러스(RSV)-감염된 BEAS-2B 세포, 및 1차 애벌레 드로소필라 멜라노가스터 뉴런20을 포함한다.
프로토콜
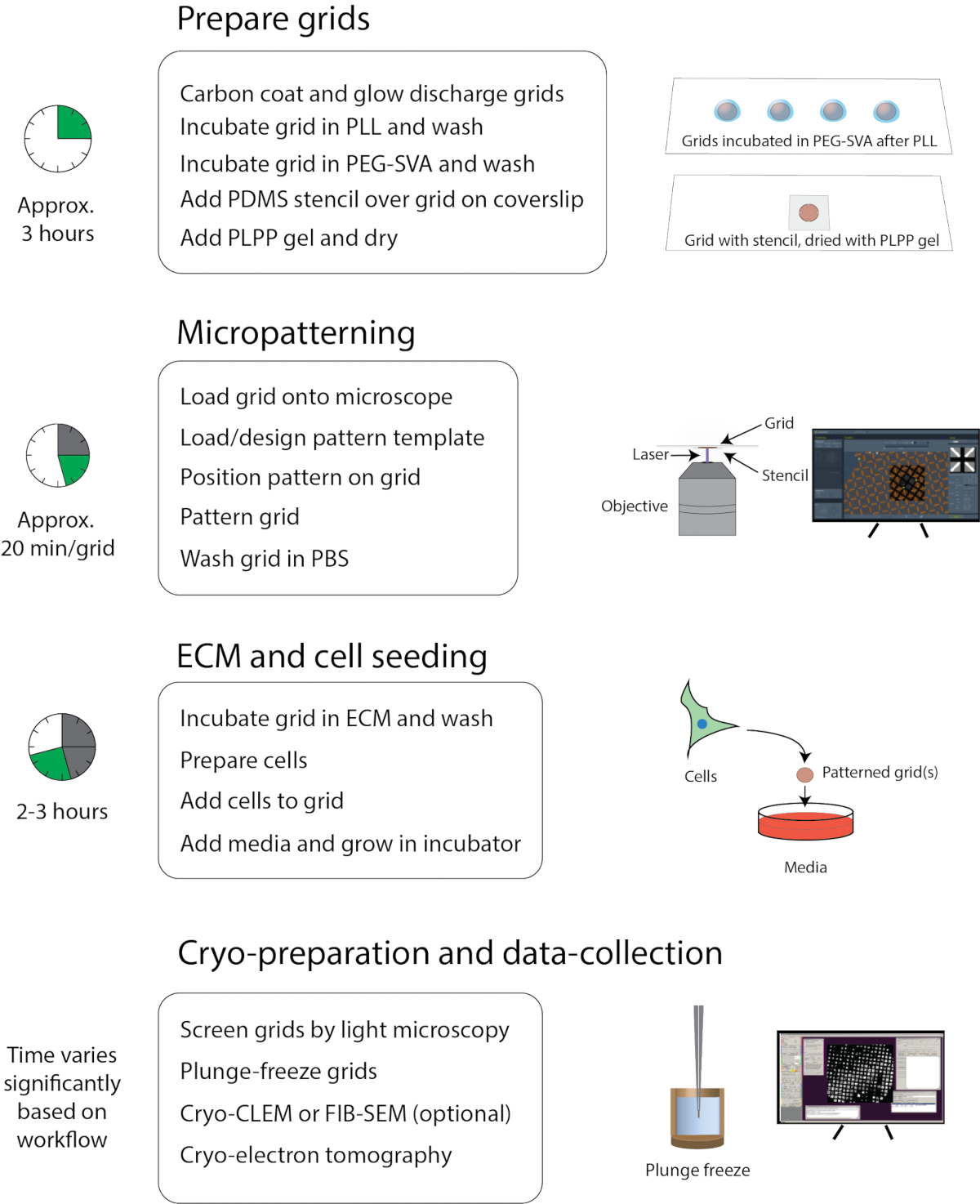
여기에 설명된 프로토콜은 매디슨 위스콘신 대학의 라이트 연구소와 Cryo-EM 연구 센터에서 사용하는 세포 배양, 마이크로 패턴 및 이미징 방법의 편집입니다. 워크플로는 그림 1에 표시됩니다. 추가 교육 및 교육 자료는 다음 사이트에서 사용할 수 있습니다: https://cryoem.wisc.edu 또는 https://wrightlab.wisc.edu
1. 패터닝을 위한 그리드 준비
- TEM 그리드를 깨끗한 유리 슬라이드로 옮기고, 탄소 측위로 (탄소 호일의 표준 두께는 12 nm입니다). 탄소 증발기인 ACE600을 사용하여 5-8nm의 추가 탄소를 그리드에 증발하여 전체 탄소 필름 내구성을 향상시춥니다.
참고: SiO2 그리드에는 이 단계가 필요하지 않습니다. 이 단계는 사전에 수행 될 수있다; 진공 건조기와 같은 습도가 낮은 환경에 코팅된 그리드를 저장합니다. - 그리드 준비 홀더로 그리드를 전송하고 광채 배출 그리드 탄소 측위로. 글로우 방전 시스템을 사용하여 10mA에서 60s의 그리드를 배출하며 80mm 작동 거리와 1.0 x 10-3 mbar의 진공 압력으로 그리드를 배출합니다. 다음 단계의 15-30 분 이내에이 작업을 수행하십시오.
참고: 그리드 준비 홀더는 작은 페트리 접시에 필터 종이 조각으로 상업적으로 구입하거나 수제 할 수 있습니다.
2. 안티 파울 링 층의 응용 프로그램
참고: 그리드를 처리할 때 적절한 멸균 기술을 사용해야 하며 모든 솔루션은 멸균 및/또는 필터 를 살균해야 합니다.
- 그리드(카본 사이드 업)를 깨끗한 유리 슬라이드 또는 커버슬립으로 옮기고 그리드 간에 최소 1cm의 분리를 합니다. 파이펫 10 μL000% 폴리-L-리신(PLL)을 각 그리드에 올보고 있습니다. 촉촉한 종이 타월이 있는 밀폐된 플라스틱 상자와 같이 습한 챔버에서 그리드를 최소 30분 동안 배양합니다.
참고: 이 단계는 하룻밤 사이에 연장할 수 있습니다. 챔버의 습도 수준이 그리드가 건조되는 것을 방지하기에 충분해야 합니다. - 0.1 M HEPES pH 8.5의 15 μL로 각 그리드를 세 번 세척합니다. 각 세척에 대해 그리드를 건조시키지 않고 파이펫으로 그리드에서 대부분의 액체를 제거합니다. 신선한 버퍼 15 μL을 추가하고 적어도 30 s에 대한 배양하고 반복합니다. 최종 세척 후 각 그리드를 0.1 M HEPES의 15 μL에 둡니다.
참고: 이 단계와 향후 단계에서는 그리드를 적시게 하고 파이펫과 그리드 간의 접촉을 방지하는 것이 중요합니다. - 각 그리드당 100 mg/mL 폴리에틸렌 글리콜-수치니미딜 발레염(PEG-SVA)의 10μL을 0.1M HEPES pH 8.5로 준비한다. PEG-SVA는 부드러운 혼합으로 빠르게 녹아 선명한 솔루션을 제공합니다.
참고: PEG-SVA 솔루션을 미리 준비하지 마십시오. PEG-SVA는 pH 8.5에서 10분의 반감기를 가지고 있습니다. -20°C의 건조기 또는 건조한 환경에 저장하여 PEG-SVA 스톡을 과도한 수분에 노출시키고 개봉 전에 실온으로 따뜻하게 하는 것을 피하십시오. - PEG-SVA 용액을 준비한 직후 각 그리드에서 HEPES pH 8.5의 15 μL 낙하를 제거하고 PEG-SVA 용액의 10 μL 낙하를 추가합니다. 개습 챔버에서 그리드를 적어도 1 시간 동안 배양합니다.
참고: 이 단계는 하룻밤 사이에 연장할 수 있습니다. 챔버의 습도가 그리드가 건조되는 것을 방지하기에 충분해야 합니다. - 멸균 수의 15 μL로 각 그리드를 세 번 세척합니다. 각 세척에 대해, 그리드건조하지 않고 파이펫으로 그리드에서 액체의 대부분을 제거하고, 담수 의 15 μL을 추가하고, 적어도 30 s에 대한 배양하고 반복합니다. 마지막 세척 후 각 그리드를 15 μL의 물에 둡니다.
3. PLPP 젤 을 적용
- 각 그리드에 대해 깨끗한 현미경 커버슬립을 준비합니다. 각 그리드에 대해 다음 단계를 완료하여 한 번에 하나의 그리드를 사용하여 그리드가 건조될 가능성을 최소화합니다.
- 커버슬립 중앙에 1.0 μL 방울의 물을 놓아 그리드를 커버슬립에 배치하고 그리드를 적시게 합니다. 15 μL 물 방울에서 덮개 슬립의 1.0 μL 물 방울로 그리드를 조심스럽게 이송합니다. 그리드 탄소 면을 위로 올려 놓으십시오.
- 그리드 위에 폴리디메틸실록산(PDMS) 스텐실을 조심스럽게 배치하여 그리드를 중심으로 유지하고 그리드의 탄소 호일과스텐실 접촉을 최소화합니다.
- 그리드에 4-벤조일벤질 트리메틸람모늄(PLPP) 젤의 1.0 μL을 넣습니다. 파이펫을 부드럽게 섞어줍니다(파이펫 팁으로 그리드를 만지지 않음).
- 그리드가 있는 커버슬립을 어두운 위치로 이동하여 건조시다. 젤은 약 15-30 분 안에 건조합니다.
4. 마이크로 패턴의 교정 및 설계
- 하이라이터가 있는 유리 커버슬립의 한쪽에 색을 새는 다. 미세 기울어진 영구 마커에서 검은색 선을 추가하여 초점을 쉽게 맞출 수 있습니다. 컬러 측이 객관적인 렌즈를 향할 수 있도록 현미경에 커버슬립을 놓습니다. 브라이트필드 모드를 사용하여 형광펜에 초점을 맞춥니다.
- 현미경과 마이크로 패턴 시스템이 켜져 있는지 확인하고 올바른 조명 경로가 설정됩니다. 마이크로스코프 컴퓨터에서 마이크로매니저와 레오나르도 소프트웨어(플러그인 > 레오나르도)를 엽니다.
- 보정을 선택하고 화면 의 지침을 따릅니다. 슬라이드에 투사된 이미지가 초점에 초점을 맞추도록 현미경 포커스를 조정합니다. 노출 시간을 줄여야 할 수도 있습니다. 교정 후 지금 패턴을 선택합니다.
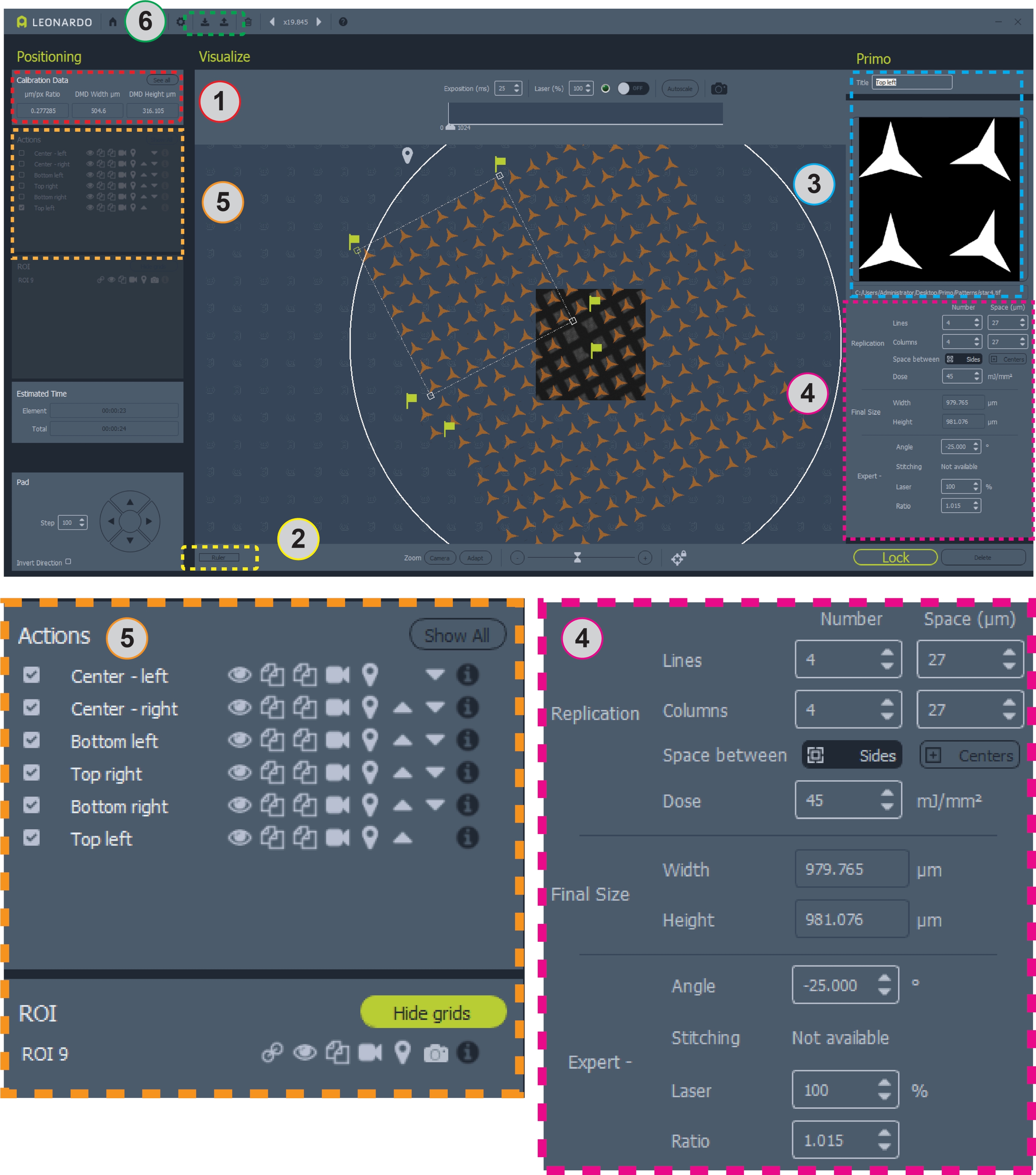
- 프로그램의 왼쪽 상단 창에서 교정 데이터 아래에 보고된 마이크로미터/픽셀(μm/px) 비율을 기록합니다(그림 2, 영역 1). 이 비율을 사용하여 패턴을 디자인할 때 마이크로미터당 사용할 픽셀 수를 결정합니다.
- 교정 후, 소프트웨어가 현미경에서 라이브 밝은 필드 보기와 함께 열려 있는지 확인합니다. 준비된 그리드를 커버슬립(섹션 3)에 적재하여 그리드가 목표 렌즈를 향하도록 무대에 로드합니다. 스테이지를 배치하고 포커스를 조정하여 그리드가 소프트웨어 창에 표시되도록 합니다.
- 마이크로미터에서 그리드 사각형 및 그리드 막대의 크기를 측정합니다. 이 소프트웨어에는 그리드를 측정하기 위해 왼쪽 아래 모서리 근처의 버튼에 의해 활성화된 눈금표가 포함되어 있습니다(그림 2, 영역 2). 예를 들어, 200 메쉬 그리드에 사용되는 패턴은 ~87× 87 μm 그리드 사각형 및 ~36 μm 그리드 막대에 해당한다.
참고: 이 소프트웨어는 즉석에서 패턴을 크기 조정하는 유연성을 제공하므로 측정에 사소한 부정확성이 허용될 수 있습니다. - 위의 측정 값과 비율을 기반으로 이미지 생성 소프트웨어로 패턴을 만듭니다. 20× 목표가 있는 최소 피쳐 크기는 1.2 μm입니다. 패턴은 압축되지 않은 8비트 .tiff 파일로 저장되어야 합니다.
- 소프트웨어를 저장할 때 이미지를 다른 픽셀 크기로 다시 조정하지 않도록 합니다. 패턴은 800 × 800 픽셀 상자 내에 맞아야 하며, 4개의 그리드 사각형을 덮기에 충분합니다.
참고: 255(흰색)의 값을 가진 픽셀은 가장 높은 강도(레이저의 총 용량)로 패턴화되고 값이 0(검정)인 픽셀은 패턴화되지 않습니다. 중간 값을 가진 모든 픽셀은 약 (X/255)*총 용량의 용량으로 패턴화됩니다. 그림 3A에서 255 및 129의 픽셀 값이 그레이스케일 패턴에 사용되었습니다. 패턴이 디자인되면 수정없이 저장하고 다시 사용할 수 있습니다.
- 소프트웨어를 저장할 때 이미지를 다른 픽셀 크기로 다시 조정하지 않도록 합니다. 패턴은 800 × 800 픽셀 상자 내에 맞아야 하며, 4개의 그리드 사각형을 덮기에 충분합니다.
5. 마이크로 패턴화
- 교정 후, 소프트웨어가 현미경에서 라이브 밝은 필드 보기와 함께 열려 있는지 확인합니다. 준비된 그리드를 커버슬립(섹션 3)에 적재하여 그리드가 목표 렌즈를 향하도록 무대에 로드합니다. 스테이지를 배치하고 포커스를 조정하여 소프트웨어의 그리드를 볼 수 있습니다.
- 초기 실행의 경우 새 템플릿을 디자인합니다. 소프트웨어에서 ROI 추가 ( 그림 2 영역 3의 위치에 표시되지 않음)를 선택하고 3,000 μm 원을 선택합니다. 화면의 밝은 필드 이미지를 사용하여 그리드 위에 원 ROI를 배치합니다. ROI를 고정하려면 잠금 장치를 누릅니다.
- ROI를 제자리에 고정한 후 패턴 추가 를 선택합니다( 그림 2 영역 3의 위치에 표시되지 않음). 섹션 4에서 디자인 된 패턴을 선택합니다. 그리드를 6개 영역으로 나누어 각 지역의 독립적인 초점 과 포지셔닝을 통해 고르지 않은 그리드를 고려합니다. 그리드의 각 모서리에 대해 8개의 × 8그리드 정사각형 영역과 중앙 양쪽에 2× 8그리드 정사각형 영역이 있으며, 중앙 4개의 그리드 사각형패턴(그림 2, 가운데 이미지)을 그대로 둡니다.
- 복제 옵션(그림 2, 영역 4)을 사용하여 초기 패턴의 복사본을 생성하여 패턴의 원하는 총 복사본 수에 도달합니다. 필요한 경우 복사본 간의 간격을 조정하여 그리드 사각형 사이의 간격과 일치합니다.
- 패턴에 대한 총 용량을 설정합니다. 30 mJ / mm2 는 좋은 출발점입니다. 자세한 내용은 토론 섹션을 참조하십시오.
- 전문가 옵션(그림 2, 영역 4)에서 영역의 각도를 조정하여 그리드 사각형의 각도와 일치합니다. 영역은 마우스를 사용하여 위치를 지정할 수 있습니다. 패턴의 비율(크기)도 조정할 수 있습니다. 패턴이 그리드 사각형과 정렬될 때까지 패턴의 각도, 위치, 공간 및 비율을 조정하여 반복합니다. 현미경 스테이지를 이동하여 라이브 브라이트필드 디스플레이에서 그리드 영역을 변경합니다.
- 잠금을 눌러 지역에 대한 변경 사항을 저장합니다.
- 지역을 복사하려면 왼쪽의 작업 패널에서 이름 옆에 중복 단추(그림 2, 영역 5, 용지 아이콘 2장)를 클릭합니다. 복사본의 위치를 변경, 이름 바꾸기 또는 편집하려면 작업 패널에서 이름을 클릭합니다.
- 원하는 모든 영역을 채우기 위해 필요에 따라 5.4-5.9 단계를 반복합니다.
- 전체 템플릿이 설계되고 배치되면 소프트웨어 내에서 템플릿 파일을 저장 합니다(그림 2, 영역 6, 상단 도구 모음에 최대 화살표 아이콘이 있는 막대).
- 이전에 저장된 템플릿(아래쪽 화살표 아이콘이 있는 막대)을 로드할 때 는 그리드 위에 ROI를 중앙으로 표시하고 잠금을 누릅니다. 동작 패널 의 각 영역을 클릭하여 각도, 위치, 용량 및/또는 패턴 파일을 변경합니다.
- 템플릿과 패턴이 배치되면 소프트웨어의 작업 패널 의 영역 중 하나를 제외한 모든 영역을 선택 취소합니다.
- 현미경 단계를 사용하여 해당 영역으로 이동하여 탄소 호일에 집중하십시오. 액션 패널에서 아이볼 아이콘을 클릭하면 패턴 오버레이의 표시가 켜지거나 꺼집니다.
- 그리드가 포커스가 되면 밝은 필드 셔터를 닫고 소프트웨어 오른쪽 하단에 있는 Play 아이콘을 눌러 실시간으로 모니터링할 수 있는 패터닝 프로세스를 시작합니다.
- 작업 패널에서 다음 영역에 대한 상자를 선택합니다. 그리드가 표시되도록 밝은 필드 셔터를 열고 현미경 스테이지를 사용하여 해당 영역을 중심으로 합니다. 작업 패널의 각 영역에 대해 5.13-5.14 단계를 반복합니다.
- 현미경에서 그리드로 커버슬립을 제거하고 멸균 인산염 완충식식염(PBS)의 10 μL을 그리드에 즉시 피펫합니다.
- 10분 후 핀셋으로 스텐실을 제거한 다음 15μL의 PBS로 그리드 3x를 세척합니다. 마지막 세척 후 각 그리드를 PBS의 15 μL에 배치하고 그리드를 어두운 위치로 이동합니다.
6. ECM 단백질의 증착
- 배양 된 세포의 경우 6.2-6.5 단계를 따르십시오. 1 차적인 Drosophila 신경을 위해, 단계를 따르십시오 6.6-6.10.
- 각 그리드에 대해 최소 15μL의 ECM을 준비합니다. BEAS-2B 세포의 경우 멸균 PBS에서 0.01 mg/mL 소 섬유네틴 및 0.01 mg/mL 플루오로포레-공주 피브리노겐의 최종 농도를 준비한다. HeLa 세포의 경우 멸균 PBS에서 0.01 mg/mL 소 콜라겐 I 및 0.1 mg/mL 플루오로포어-컨쥬게이트 피브리노겐을 준비하십시오.
- 각 그리드에서 대부분의 PBS를 제거하고 ECM의 15 μL을 적용합니다. 적어도 1 시간 동안 실온에서 습한 챔버에서 그리드를 배양한다.
참고: 이 단계는 4°C에서 하룻밤사이에 연장될 수 있다. - ECM에서 인큐베이션 후 멸균 PBS로 각 그리드 5배세척합니다. 각 세척에 대해 그리드를 건조시키지 않고 파이펫으로 액체의 대부분을 제거하고 신선한 PBS 15 μL을 추가하고 적어도 30 s의 배양하고 반복하십시오. 마지막 세척 후 각 그리드를 PBS에 둡니다.
참고: 그리드는 4°C에서 PBS에서 최대 1주일 동안 저장할 수 있으며 품질 저하가 관찰되지 않습니다. - 형광 현미경을 사용하여 ECM의 형광소를 감지하여 패터닝을 확인하고 탄소 호일이 그대로 유지된다는 것을 확인하십시오. 깨진 사각형 몇 개는 일반적으로 견딜 수 있습니다.
- 1차 Drosophila 뉴런의 경우 패턴 그리드를 멸균 PBS를 포함하는 30mm 유리 바닥 접시로 이동합니다.
- 접시에서 PBS를 흡인하고 멸균 환경에서 25 °C에서 하룻밤 동안 0.5 mg / mL 플루오로포레 -컨쥬게이트 공수 의 2 mL을 적용합니다.
- 접시에서 concanavalin A 용액을 제거하고 (그리드를 건조하지 않고) PBS로 그리드 3배 세척합니다. 각 세척에 대해 접시에서 2mL PBS를 넣고 제거합니다.
- 형광 현미경을 사용하여 ECM의 형광소를 감지하여 패터닝을 확인하고 탄소 호일이 그대로 유지된다는 것을 확인하십시오. 깨진 사각형 몇 개는 일반적으로 견딜 수 있습니다.
- 마지막 세척 후, 유리 바닥 접시에서 PBS를 제거하고 2mL의 신선하게 준비된 멸균 여과 된 슈나이더의 Drosophila media21을 추가하여 20% 열 불활성 태아 소 혈청(FBS), 5 μg/mL 인슐린, 100 μg/mL 페니실린, 100 μg/mL stl, 100 μg/mL stl 뉴런이 도금될 때까지 멸균 환경에서 25°C에서 배양한다.
7. 시드 전에 1 차 적인 Drosophila 세포의 준비
- 55mm 해부 접시를 70% EtOH로 살균하고, 이어서 멸균 여과 1× 해부 염벽(9.9m HEPES pH 7.5, 137m NaCl, 5.4mM KCl, 0.17 mM NaH2PO4, 0.22m M KH2PO4, 3.3mM 포도당, 43.8mm)의 접시2-3mL을 첨가한다.
- 핀셋을 사용하여 음식에서 30-40 3번째 인스타 애벌레를 부드럽게 선택하십시오.
- 1× PBS로 튜브에 애벌레를 넣고 1× PBS로 두 번째 튜브로 옮겨 애벌레를 씻으십시오.
- 유충을 70%의 EtOH로 튜브로 옮겨 PBS를 씻어낸 다음 70%의 EtOH로 두 번째 튜브로 옮겨 넣습니다. 애벌레를 살균하기 위해 2-3 분 동안 두 번째 튜브에 애벌레를 둡니다.
- 애벌레를 1× 해부 식염수로 튜브로 옮은 다음 즉시 1× 해부 식염수로 두 번째 튜브로 옮기습니다.
- 개별 애벌레를 1× 해부 식염수로 함유된 해부 접시에 옮기습니다. 한 쌍의 집게와 해부 현미경으로 각 애벌레를 빠르게 찢어 뇌를 추출하고 1× 해부 식염수로 제 3 튜브로 옮습니다. 모든 뇌가 추출될 때까지 반복합니다.
- 뇌를 함유하는 튜브를 300 x g 에서 1분 동안 원심분리합니다.
- 상류체를 버리고 1mL의 1mL× 해부 식염수로 씻고 튜브를 300 x g에서 1 분 동안 원심 분리합니다. 이 단계를 한 번 더 반복합니다.
- 200-250 μL이 튜브에 남아있을 때까지 상체를 버리고 1 x 해부 식염수에 2.5 mg / mL 리베라제20 μL을 추가하십시오.
- 실온에서 1 시간 동안 회전기에서 튜브를 회전하는 경우; 이 시간 동안, 파이펫 용액은 10 분마다 25-30 배. 끝으로, 솔루션은 약간 불투명해야합니다.
- 300 × g 에서 세포를 원심 분리하여 5 분 동안.
- 상체를 폐기한 다음 1mL의 보충제 슈나이더 미디어를 추가합니다. 파이펫을 30번 섞습니다.
- 300 × g 에서 세포를 원심 분리하여 5 분 동안.
- 상체를 버리고 슈나이더의 미디어를 보충 한 1 mL을 추가하여 셀 펠릿을 씻으십시오. 파이펫을 30번 섞습니다.
- 300 × g 에서 세포를 원심 분리하여 5 분 동안.
- 상체를 폐기한 다음 슈나이더의 미디어를 보충한 300μL로 셀 펠릿을 다시 놓습니다. 파이펫용액을 30-40번 섞는다.
8. BEAS-2B 및 HeLa 세포의 배양 및 RSV 감염
- 37°C 및 5% CO2에서 T75 플라스크에서 HeLa 세포 및 BEAS-2B 세포를 유지합니다. 통로 세포마다 3-4 일 한 번 약 80 %의 합류에 도달합니다. DMEM + 10% FBS + 1× 항생제 항진균제에서 HeLa 세포를 유지합니다. RPMI + 10% FBS + 1× 항생제-항진균6,22,23에서 BEAS-2B를 유지하십시오.
- 감염되지 않은 세포의 파종은 9절로 건너뜁니다. BEAS-2B 및 HeLa 세포는 RSV 감염에 취약합니다; BEAS-2B 세포는 여기에 도시된 RSV를 포함하는 모든 실험에 사용되었다.
참고: 적절한 생물안전 캐비닛(BSC) 및 개인 보호 장비(PPE)를 사용하여 기관 프로토콜을 준수하여 모든 BSL-2 단계를 수행합니다. - 세포의 RSV 감염 이전에는, 5 × 104 세포를 6웰 플레이트(표면적 ~9.6 cm2)로 통과하고 하룻밤 사이에 배양한다.
- 트립시네이니즈하고 세포의 하나의 우물을 계산합니다. 트립시니화하려면, 하나의 우물에서 미디어를 흡습하고 Mg2 + 및 Ca2 + 없이 멸균 PBS의 2 mL로 세척하여 잔류 매체를 제거합니다. 0.25% 트립신 용액의 500 μL을 추가합니다. 5-10 분 동안 37 °C에서 배양하십시오. 주기적으로 세포를 확인하여 표면에서 방출되는지 확인합니다. 세포가 방출되면 배양 배지의 1.5mL을 추가하십시오.
- 트립판 블루의 100 μL와 트립시네화된 세포의 100 μL을 섞는다. 희석된 세포 믹스의 파이펫 10 μL이 혈류계로 혼합됩니다. 셀을 계산하고 잘 셀 수를 계산합니다. 이 번호를 사용하여 아래 MOI를 계산합니다.
- 성장 매체에서 RSV-A2mK+24의 희석을 준비하여 750 μL의 미디어에서 웰당 10개의 MOI를 달성합니다. RSV-A2mK+의 MOI는 주식의 형광 초점 장치(FFU) 티터(예: 우물당 1.0 × 105셀 및 RSV 주식 1.0 × 108 FFU/mL로 계산할 수 있으며, 바이러스 주식 1:75 ~ 1× 106 FFU/750 μL 또는 1.30 μL)×.×.×.
- 6웰 접시의 세포로부터 미디어를 흡인하고 위에서 각 우물까지 바이러스 용액의 750 μL을 추가합니다.
- 1 시간 동안 실온에서 접시를 흔들어.
- 1시간 후, 37°C로 미리 데워진 성장 매체로 최대 2mL당 의 총 부피를 가져와 서 플레이트를 37°C로 설정하여 6h당 5% CO2 로 설정한다.
- 세포를 풀어 놓기 위해 트립시닝하고 아래에 설명된 대로 파종으로 진행합니다. 시드 후, 급락 동결 전에 추가 18 h에 대한 그리드를 배양 (총 24 시간 감염 후).
9. 마이크로 패턴 그리드에 셀 시드
- 배양 된 세포의 경우 단계를 따르십시오 9.2-9.8; 1 차적인 Drosophila 신경을 위해, 따라 9.9-9.11.
- 세포를 풀어 놓기 위해 트립시니즈(위의 섹션 8에서 4단계 참조). 세포 집계를 줄이려면 60% 이하의 수렴성으로 세포를 트립시닉합니다.
- 트립판 블루의 100 μL와 트립시네화된 세포의 100 μL을 섞는다. 희석된 세포의 파이펫 10 μL은 혈류계로 혼합되어 세포를 계산합니다.
- 2 × 104 세포 /mL로 매체의 세포를 희석.
- 30mm 유리 바닥 접시의 중앙에 1 μL의 매체를 추가하여 그리드를 배치하고 건조를 방지합니다. 커버슬립의 PBS에서 유리 바닥 접시의 중심으로 그리드를 전송합니다. 그리드에 셀 용액 10μL을 추가합니다.
- 밝은 필드 현미경을 사용하여 5 분 후에 그리드에 세포 접착력을 관찰하십시오. 패턴의 대부분이 비어 있는 상태로 유지되면 셀 용액의 10 μL 드롭을 추가합니다. 인큐베이션 중에 그리드 및 셀 용액을 37°C로 유지합니다.
- 대부분의 패턴이 점유되거나 많은 점유 패턴이 여러 셀을 갖기 시작할 때까지 9.6 단계를 반복합니다. 인큐베이터(37°C, 5% CO2)에서 2h의 그리드를 배양한다.
- 미리 데워진 2mL의 전동 된 매체로 접시를 범람하고 하룻밤 동안 배양 (37 °C, 5 % CO2).
- 1차 Drosophila 뉴런의 경우 그리드 함유 접시에서 미디어를 제거하고 세포를 접시에 접시에 접시로 플레이트합니다.
- 세포가 부착될 때까지 30-60분 기다린 다음 2mL의 보충제 슈나이더 미디어를 추가합니다.
- 급락 동결 전에 최소 2-3 일 동안 25 °C 인큐베이터에서 뉴런을 배양.
10. 패턴 그리드의 이미징 및 진동
- 형광 현미경에 패턴 그리드 및 배양 된 세포를 포함하는 유리 바닥 접시를 놓습니다.
- 밝은 필드와 적절한 형광 채널을 사용하여 그리드의 이미지를 획득하여 세포내의 패턴 및 기타 라벨링을 감지합니다. 세포 밀도와 위치 지정이 다운스트림 이미징 및 분석에 적합한지 확인합니다.
참고: 브라이트필드와 형광 이미지는 FIJI 소프트웨어 패키지25에서 처리되었습니다. - 냉동고 플런지 냉동고를 준비하십시오. 동결 장치의 유형은 샘플에 가장 적합한 가용성, 비용 및 기능에 따라 달라집니다.
참고: 1차 드로소필라 뉴런은 자동 플런지-냉동고에 제조되었고, BEAS-2B 세포는 반자동 플런지-냉동고를 사용하여 제조하였다. - 틸트 계열의 적절한 정렬을 위해 샘플에 금 신탁을 적용합니다. Blot 샘플은 과잉 매체를 제거하기 위해 샘플을 액체 질소에 의해 냉각된 액체 에탄과 같은 극저온으로 얼게크-고정합니다. 1 차적인 Drosophila 뉴런을 위해, 뒤에서 4 s에 대한 얼룩. HeLa 및 BEAS-2B 세포의 경우, 4-6 s에 대한 양쪽에서 얼룩. 냉동 그리드는 추가 사용이 있을 때까지 액체 질소에 보관할 수 있습니다.
- 직구 검출기 카메라로 300kV로 작동되는 저온 전자 현미경으로 이미지 진동 세포. cryo-EM/cryo-ET 데이터 수집을 위한 SerialEM26 과 같은 소프트웨어로 관심 있는 각 영역에 대한 틸트 시리즈 컬렉션을 설정합니다.
참고: 1차 드로소필라 뉴런의 틸트 시리즈는 -60°에서 60°까지 양방향으로 -8 μm 디포커스에서 2°의 속도로 양방향으로 수집되었으며, 픽셀 크기는 4.628Å로 70-75 e-/Å2의 총 용량으로 수집하였다. RSV-감염된 BEAS-2B의 틸트 시리즈는 -5 μm 디포커스에서 에너지 필터(20 eV 슬릿)를 가진 직접 전자 검출기에 수집되었으며, 총 용량은 ~80 e-/Å2이다. - 틸트 계열을 처리하여 토모그램을 재구성합니다.
참고: 여기에 제시 된 토모그램은 IMOD 패키지를 사용하여 재구성되었다27; 로우패스 필터링은 EMAN2 소프트웨어 패키지를 사용하여 수행되었다28.
결과
이 절차는 전체 셀 극저온-ET 실험에 대한 EM 그리드를 패턴화하는 데 사용되었습니다. 초기 세포 배양 제제, 마이크로패턴(그림 1) 및 이미징을 포함하여 이 연구에서 발표된 전체 워크플로우는 3-7일을 포괄합니다. 2단계 절차는 그리드에 PLL을 적용하고 반응성 PEG-SVA를 추가하여 PEG를 연결하여 안티 폴링 층을 생성하는 데 사용되었다. 한 배큐베이션에 PLL-g-PEG를 추가하여 단 단단에 적용할 수도 있습니다. PLPP 젤은 UV 마이크로패터닝을 위한 촉매제이며, 이는 또한 덜 농축된 액체로도 이용된다. 젤은 액체에 비해 현저하게 감소 된 용량으로 패터닝을 허용하여 훨씬 빠른 패터닝을 초래합니다. 이 시스템을 사용하면 전체 TEM 그리드의 실제 패터닝 시간은 ~2분이었습니다. 마이크로패턴 워크플로우만으로도 일반적으로 5-6시간 동안 이며 개인이 TEM 그리드에서 표준 셀 배양을 위해 8개의 그리드를 패턴화할 수 있습니다.
마이크로패턴화 공정 동안 여러 단계의 단계는 긴 배양 시간이 필요합니다(단계 2.1, 2.3, 6.4 참조). 편리하게, PLL 패시베이(2.1) 또는 PEG-SVA 패시베이(2.3)와 같은 이러한 단계 중 일부는 하룻밤 잠복기로 확장될 수 있다. 또한, 그리드는 나중에 사용하기 위해 ECM 단백질 또는 PBS의 용액에 미리 패턴화되고 저장될 수 있다. 우리의 연구 결과에서는, 이 선택권은 세포 준비 및 파종의 타이밍이 BEAS-2B 세포의 1 차적인 Drosophila 신경및 RSV 감염과 같은 중요한 경우에 중요했습니다.
그리드는 클린 공구, 멸균 용액을 사용하여 일반 생물 안전 수준 2(BSL-2) 실험실 설정에서 제조되었으며 성장 미디어에 항생제/항진제를 포함시켰습니다6,22,29,30. 미생물 오염에 특히 민감한 시료의 경우, 항 오염 층 및 ECM은 조직 배양 후드 또는 기타 멸균 환경에서 적용될 수 있다. 또한 패터닝과 ECM 응용 프로그램 간에 그리드를 에탄올로 세척할 수 있습니다. 전염성 제제와 협력하는 경우 적절한 생물 안전 프로토콜을 준수하기 위해 절차를 조정하는 것이 중요합니다.
이러한 워크플로우 및 제시된 절차(도 1)는 HeLa 세포(도 4), RSV-감염된 BEAS-2B 세포(도 3, 도 5) 및 1차 드로소필라 애벌레 뉴런(도 6, 도 7)을 최적의 저저온 ET 데이터 수집을 위해 패턴 EM 그리드에 시드되도록 허용하였다.
마이크로패턴 TEM 그리드에 시드된 HeLa 세포는 칼세인-AM 및 에티듐 호모이머-1 계 세포 생존성 분석(도 4A, B)을 이용하여 형광 염색에 의해 결정된 바와 같이 실행 가능한 상태를 유지한다. 콜라겐과 피브리노겐의 혼합 ECM을 사용하여 HeLa 세포는 그리드 전체의 패턴을 쉽게 부착합니다(도 4A, C). 패턴을 따라 확장 세포의 전반적인 형태는 패턴되지 않은 그리드에서 자란 세포의 형태와 유사합니다(그림 4C,D). HeLa 세포의 경우, 총 세포 두께는 셀 주변 부근의 두께가 < ~< 1 μm인 10 μm을 가진 ~< 10 μm로 남아 있다(도 4E, F).
RSV 연구의 경우, 우리는 그라데이션을 사용하여 전체 그리드 사각형을 패턴화, 가장자리에 낮은 용량 노출과 센터를 향해 더 높은 용량 패턴 (그림 3A). 그라데이션 패턴은 세포 주변 근처에 존재하는 방출된 바이러스를 검색할 때 더 나은 결과를 산출했습니다. 이러한 패턴으로, 세포는 높은 ECM 농도를 우선적으로 준수하는 것으로 나타났지만, 또한 낮은 ECM 농도에서 부착하고 성장할 수 있다. 지역 간의 상대적인 복용량 여러 복용량을 필요로 하는 패턴을 사용 하는 경우 최적화 해야 합니다. 복용량과 따라서 ECM 농도 는 서로 너무 유사 하거나 너무 이질 적인 경우, 여러 복용량을 사용 하 여 효과 손실 됩니다.
도 3에서, TEM 그리드는 패턴화되었고, 그 후 RSV감염 BEAS-2B 세포로 시드되고 극저온-EM 데이터 수집에 사용되었다. 도 4A는 그라데이션 패턴을 사용하여 TEM 그리드에 패턴화된 ECM의 형광 이미지이다. 패턴의 중앙 영역을 따라 세포 접착 및 성장은 도 3B에서 세포의 밝은 필드 이미지로서 볼 수 있다 18 시간 후 시드. 도 3C에서 RSV-A2mK+의 복제로부터의 형광 신호(빨간색)는 ECM의 신호와 겹쳐져 있다. 감염된 세포의 대다수는 그라데이션 패턴의 고밀도 중앙 영역을 따라 배치된다. 그리드 포스트 저온 고정의 저막트 TEM 맵은 그리드 사각형의 중심 근처의 탄소 호일에 위치한 RSV 감염 세포를 포함한 다수의 세포를 보여줍니다. 이전에 표준 TEM grids22에서 자란 세포에 대해 나타난 바와 같이, 틸트 시리즈는 마이크로패턴 그리드에서 자란 감염된 BEAS-2B 세포의 주변부근에 근접하여 RSV 비리온을 위치시키고 수집하였다(그림 5A,B). RSV 구조 단백질의 대부분은 뉴클레오캡시드(N) 및 바이러스 융합 단백질(F)을 포함하는 토모그램 내에서 확인할 수 있다(도 5C, 블루 및 붉은 화살표).
1차 Drosophila 뉴런 연구의 경우, 소프트웨어가 제공하는 해상도 한계(패턴의 두께가 2 μm인 경우) 그리드 사각형 내에서 단리되도록 허용된 좁은 패턴이 발견되었다(도 6). 신경 소마 패턴 내에서 며칠의 기간 동안 그것의 neurites를 확장할 수 있었다. 이를 통해 무패턴 그리드에서 배양된 뉴런에 비해 중성염의 쉬운 식별 및 틸트 시리즈 획득이 허용되었습니다(그림 7). 또한 형광으로 표지된 콩카나베일 A, 시험관 내 드로소필라 뉴런 배양에 대한 ECM로 사용되어 온 렉틴이 패터닝에 적합하다는 것을 발견했습니다.
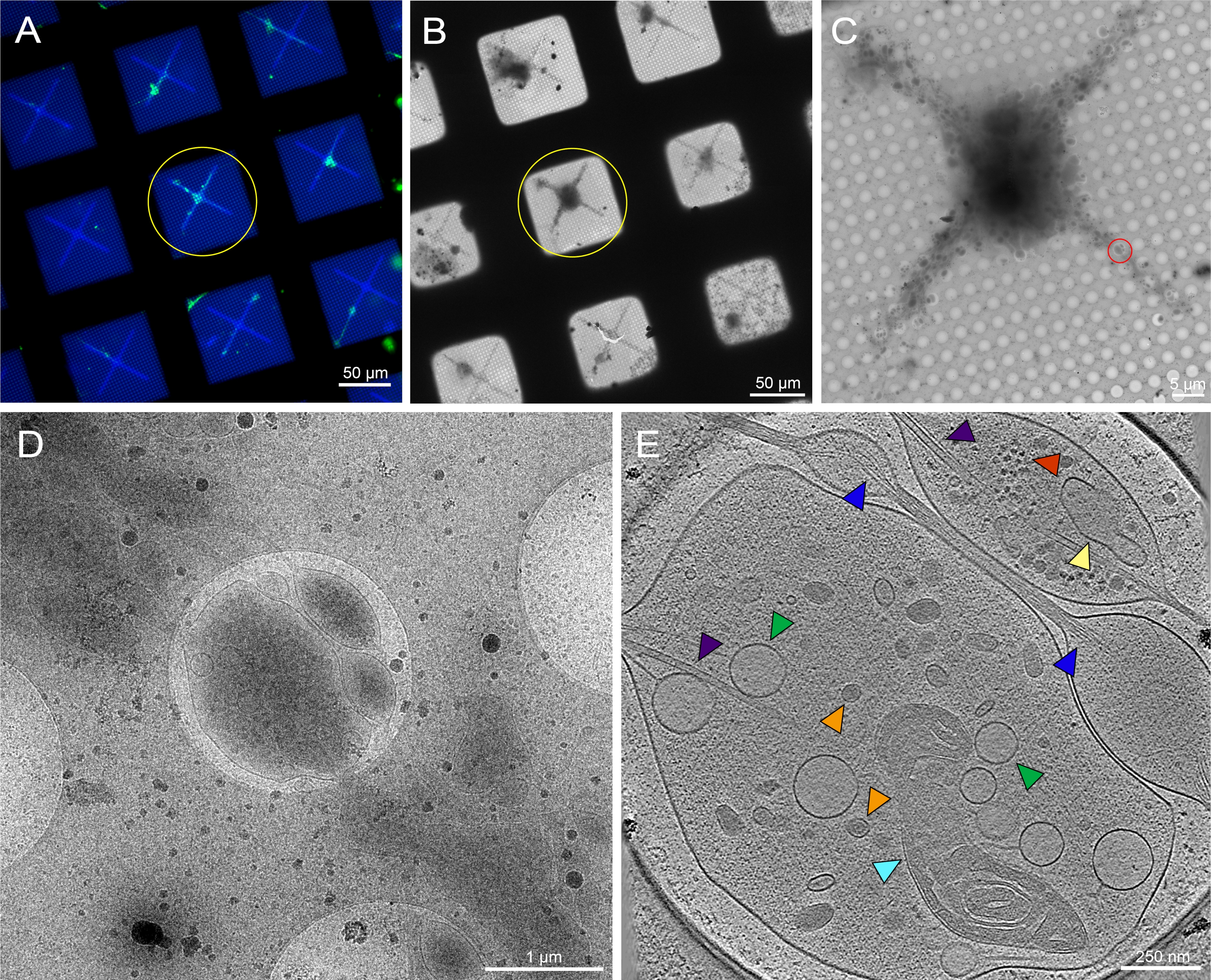
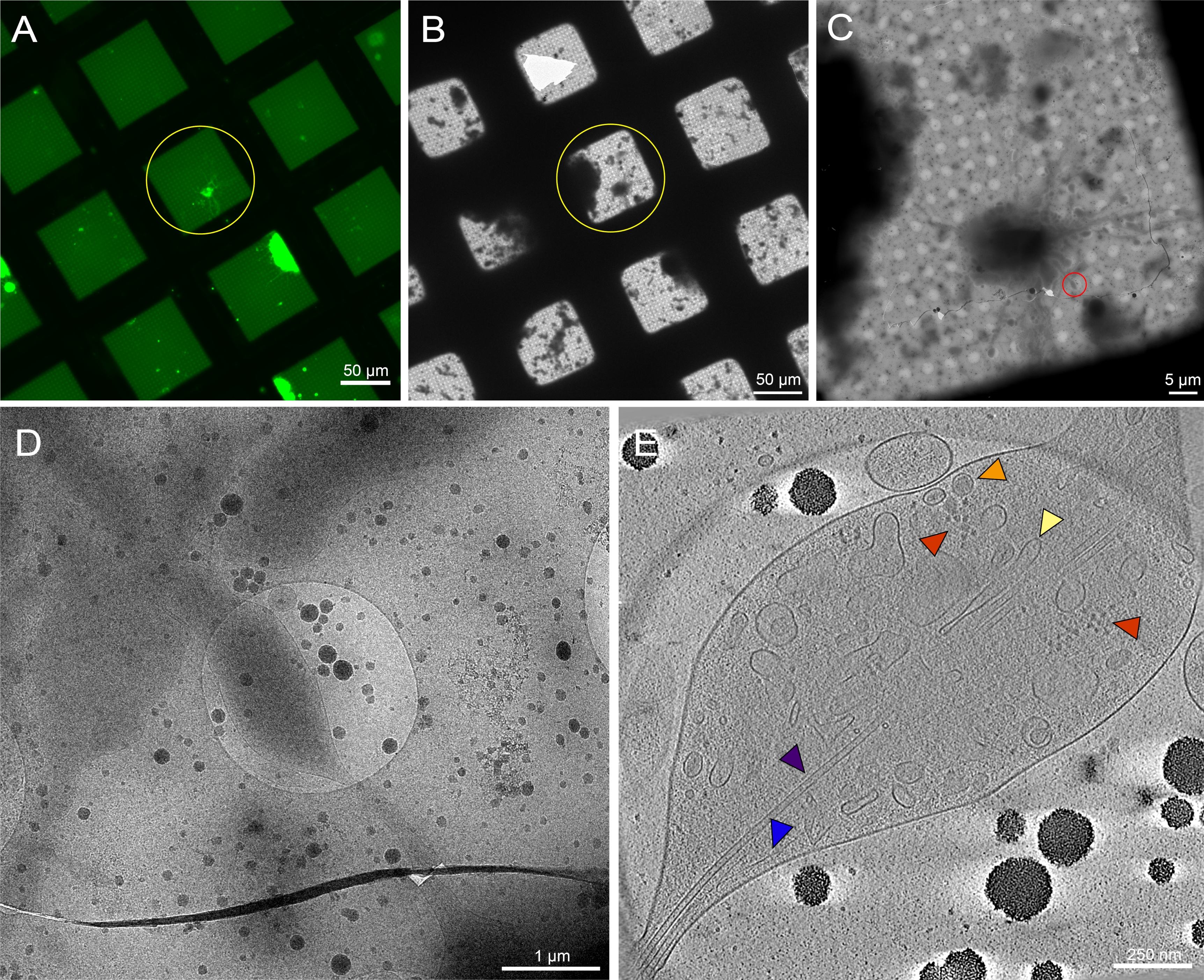
세 번째 인스타 애벌레에서 드로소필라 뉴런은 이전에 출판 된 프로토콜에 따라 격리되었다20,21,31. 신경 제제는 마이크로패턴 극저온-EM 그리드에 적용되었으며, 여기서 concanavalin A는 세포 배치, 확산 및 조직을 조절하기 위해 패턴에 증착되었다. 패턴 또는 무패턴 그리드의 뉴런은 최소 48-72시간 동안 배양할 수 있었고 그리드는 동결되었습니다. 패턴 영역을 통해 분포된 여러 드로소필라 뉴런을 가진 마이크로패턴 EM 그리드의 대표적인 이미지가 도 6A에 도시된다. 막에 범신경 GFP 발현이 있는 형질 선질 플라이 스트레인에서 유래한 이러한 뉴런은 형광 라벨링뿐만 아니라 마이크로패턴 내의 위치 때문에 가벼운 현미경 검사법에 의해 쉽게 추적할 수 있습니다. 무패턴 그리드에서 배양된 뉴런은 광 현미경 검사법(도 7A, 노란색 원)에 의한 GFP 신호를 통해 추적할 수 있지만, 극저온 EM에서 이를 찾는 것은 세포 이물질및 매체의 오염으로 인해 실질적으로 더 어려워졌다(그림 7B, 황원). 이러한 존재는 패턴 그리드에 뉴런에 대한 감소되었다, 비 패턴 영역의 안티 파울 층에서 PEG로 인해 준수에서 세포 파편을 격퇴. 뉴런 세포 체체의 치수및 확장된 중성염(도 6A, B, 황색 원)으로 인해, 극저온-ET 틸트 계열은 세포의 더 얇은 영역을 따라 수집되었다(도 6C, D, 적색 원). 뉴런 세포막, 미토콘드리온(cyan), 마이크로투부(purple), 액틴 필라멘트(파란색), 혈관 구조(주황색 및 녹색), 리보솜(red)과 같은 거대 분자는 3D 토모그램(도 6E)을 통해 배율화 영상 몽타주와 슬라이스로 잘 해결되었다. 유사한 서브 셀룰러 특징은 패턴이 없는 뉴런(그림 7E)의 3D 토모그램에서 볼 수 있지만, 데이터 수집에 대한 실행 가능한 세포 표적을 찾는 데 어려움이 크게 감소했습니다.
그림 8에서는 이러한 문제 중 일부를 가진 그리드의 대표 이미지가 수집되어 식별 및 문제 해결을 지원합니다. 최적의 조건이 결정되면 마이크로패터닝은 극저온-TEM을 위해 그리드에 셀을 배치하는 신뢰할 수 있고 재현 가능한 방법입니다.

그림 1: 저온-EM용 마이크로패터닝의 일반적인 워크플로우입니다. 워크플로는 그리드 준비, 마이크로 패턴화, ECM 및 셀 시드링, 저온 준비 및 데이터 수집의 네 부분으로 대략 분할될 수 있습니다. 각 섹션의 주요 단계는 제목 아래에 나열되며 각 섹션을 완료하는 대략적인 시간은 왼쪽에 표시됩니다. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
{kind=link}

그림 2: 그리드에 패턴이 배치된 소프트웨어의 스크린 샷입니다. 영역 1에는 패턴 디자인용 μm/pix 비율이 포함되어 있습니다. 영역 2는 그리드를 측정하기 위한 눈금자입니다. 영역 3은 패턴 및 ROI를 추가하거나 변경하는 곳입니다. 영역 4에는 패턴 포지셔닝 및 용량에 대한 모든 정보가 포함되어 있습니다. 영역 5에는 오버레이 토깅, 패턴 복사 또는 삭제, 마이크로패턴 에 대한 패턴 선택 등 패턴에 대한 옵션이 포함되어 있습니다. 영역 6은 템플릿을 저장하고 로드할 수 있는 곳입니다. 4구역과 5구역의 더 큰 전망은 명확성을 위해 아래와 같습니다. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
{kind=link}

도 3: RSV-감염된 BEAS-2B 세포패턴 극저온-TEM 그리드에 . (A) 형광표 ECM을 첨가한 후 패턴 그리드의 형광 이미지. 입력 패턴이 왼쪽 아래 모서리에 표시됩니다. (B) A.(C)에서 그리드에서 자란 BEAS-2B 세포의 브라이트필드 이미지는 급락 동결 직전에 RSV 감염 세포(red)의 형광 이미지와 A(cyan) 및 B(grey)에서 이미지를 병합하는 단계; 감염된 세포는 mKate-2를 표현합니다. 스케일 바는 급락 동결 후 B의 그리드의 500 μm.(D) 저배율 극저온-TEM 맵입니다. 형광 이미지는 가성비색입니다. 스케일 바는 500 μm입니다. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
{kind=link}

그림 4: 패턴 및 패턴되지 않은 셀의 라이브/데드 스테인링. (A) 패턴 그리드에서 자라서 칼신-AM(라이브 셀 얼룩, 녹색) 및 에티듐 호모디머-1(죽은 세포 얼룩, 빨강)으로 염색된 HeLa 세포의 형광 이미지. (B) HeLa 세포는 무패턴 그리드에서 재배되고 0.01 mg/mL 콜라겐 및 피브리노겐 647 ECM(빨간색)을 가진 패턴이 있는 콴티포일 R2/2 그리드에서 HeLa 세포의 공초점 z 스택의 프로젝션에서와 같이 염색된다. 세포는 칼신-AM(녹색) 및 Hoechst-33342(파란색)로 염색하였다. (D) 0.01 mg/mL 콜라겐 및 피브리노겐 647 ECM으로 배양된 무패턴 그리드에 있는 HeLa 세포는 칼신-AM 및 Hoecsht-33342로 배양및 염색된다. 형광 이미지는 전송된 빛(grayscale)과 병합되었습니다. (E) X, Z 프로젝션의 C. (F) X, Z 프로젝션의 D. 이미지는 가시변입니다. (A) 및 (B)의 스케일 바는 500 μm; (C) - (F)의 스케일 바는 10 μm입니다.
{kind=link}

도 5: RSV에 감염된 BEAS-2B 세포의 극저온-ET 패턴 극저온-TEM 그리드. (A) KRV 감염 BEAS-2B 셀의 Cryo-EM 그리드 사각 지도. 대략적인 셀 경계는 파선된 녹색 선으로 표시됩니다. (B) 빨간색으로 박스된 영역의 고해상도 이미지(A). 대략적인 셀 경계는 파선 된 녹색 선으로 표시됩니다. RSV 비리온은 세포 주변(흰색 화살표 및 노란색 상자) 근처에서 볼 수 있습니다. (C) 노란색 상자의 영역에서 수집된 토모그램으로부터의 단일 z 슬라이스(B). 적색 화살표는 RSV F 융합 단백질을 가리키며, 푸른 화살표는 리보뉴클레오단백질(RNP) 복합체를 가리킨다. (A)-(C)의 축척 막대가 이미지에 내장되어 있습니다. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
{kind=link}

그림 6: 패턴 극저온-TEM 그리드에서 제3회 인스타 드로소필라 멜라노가스터 애벌레의 뇌에서 파생된 1차 뉴런. (A) Drosophila 뉴런의 오버레이 라이브 세포 형광 현미경 그리드 몽타주 0.5 mg/mL 형광 콘카발레인 A. 그린: 드로소필라 뉴런 패턴 그리드 사각형에 막 표적 GFP를 발현. 파란색: 포토패턴. (B) 극저온 보존 후 그리드(A)의 Cryo-EM 이미지 몽타주. 노란색 원은 (A)와 동일한 그리드 사각형을 기록합니다. (C) 황색 원(A)과 (B)로 강조 표시된 사각형의 Cryo-EM 이미지 몽타주. (D) 세포의 중성염에 기울기 계열이 수집된(C)의 적색 원에 의해 경계된 영역의 배율 이미지가 높았다. E. (C)에서 레드 원으로부터 획득한 틸트 시리즈에서 재구성된 토모그램의 25nm 두께의 슬라이스. 미토콘드리아(시안), 마이크로투부(보라색), 조밀한 코어 소포(주황색), 가벼운 소포(녹색), 내피 성 망상(노랑), 액틴(blue)과 같은 다양한 세포기관에서 다양한 세포기관을 볼 수 있다. 리보좀(빨간색)과 같은 거대 분자도 오른쪽 상단 모서리에서 볼 수 있습니다. 형광 이미지는 가성비색입니다. (A)-(E)의 축척 막대가 이미지에 내장되어 있습니다. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
{kind=link}

그림 7: 패턴되지 않은 그리드에 3 번째 인스타 Drosophila 멜라노가스터 애벌레의 뇌에서 파생 된 기본 뉴런. (A) Drosophila 뉴런의 라이브 세포 형광 현미경 그리드 몽타주 0.5 mg/mL concanavalin A. Green: Drosophila 뉴런에 막 표적 GFP를 발현. (B) 급락 동결 후 (A)에서 동일한 그리드의 Cryo-EM 그리드 몽타주. 노란색 원은 (A)와 동일한 그리드 사각형을 표시합니다. 셀룰러 이물질과 미디어 오염의 존재로 패턴 그리드에 비해 대상 식별이 어려워진 점에 유의하십시오. (C) (A) 및 (B) 맵의 노란색 원이 강조 표시된 사각형의 Cryo-EM 이미지 몽타주. (D) 세포의 중성염에 기울기 계열이 수집된(C)의 적색 원에 의해 경계된 영역의 배율 이미지가 높았다. (E) (C) 및 (D)에서 틸트 시리즈에서 재구성 된 토모그램의 25 nm 두께의 슬라이스. 마이크로투블러(보라색), 액틴(파란색), 내피 망상(노란색), 조밀한 코어 소포(주황색)와 같은 여러 세포기관이 이 토모그램에서 볼 수 있습니다. 리보솜(빨간색)과 같은 거대 분자도 볼 수 있습니다. 형광 이미지는 가성비색입니다. (A)-(E)의 축척 막대가 이미지에 내장되어 있습니다. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
{kind=link}

그림 8: 패터닝에 문제가 발생할 수 있는 예입니다. 마이크로패턴 그리드에 증착된 라벨이 부착된 ECM의 형광 이미지. (A) PLPP 젤의 고르지 않은 분포로 인해 그리드 전체의 고르지 않은 패터닝. (B) ECM은 패터닝 중에 PDMS 스텐실이 적용되는 영역을 부착할 수 없습니다. (C) 포화 그라데이션 패턴(오른쪽) 또는 반전된 패턴(왼쪽)이 너무 높은 총 용량으로 패턴화된 그리드에 있습니다. (D) ECM은 패터닝 중 UV 레이저의 반사로 인해 그리드 바의 영역뿐만 아니라 패턴 영역을 준수한다. 이미지는 의사 변색; 입력 패턴은 왼쪽 아래쪽에 표시됩니다. 스케일 바는 100 μm 입니다.
{kind=link}
| 출판하다 | 잠재적 인 원인 (들) | 문제 해결 |
| 마이크로패턴화 | ||
| PRIMO 레이저에서 조명을 볼 수 없습니다. | • 라이트 경로가 올바르게 설정되지 않음 | • 현미경 광 경로가 제대로 설정되어 있는지 확인 |
| • PRIMO 레이저가 켜지지 않거나 레이저가 연동되어 있지 않습니다. | ||
| 많은 깨진 그리드 사각형 | • 핸들링 하는 동안 핀셋 또는 파이펫으로 그리드 호일을 만지고 | • 주의그리드 처리 |
| • 인큐베이션 이나 세척 중에 그리드 건조 | • 세차 및 배양 중에 그리드가 건조하지 않도록 하십시오. | |
| 패턴이 없는 대형 영역 | • 불충분한 젤 커버리지 | • 젤이 그리드 위에 고르게 퍼지도록 하는 동시에 |
| • 패터닝 중 초점에서 그리드 호일 | • 젤의 추가 마이크로 리터를 추가 | |
| • 스텐실로 덮인 영역 | • 각 영역을 패킹하기 전에 포커스를 확인 | |
| • 스텐실의 중앙 그리드 | ||
| 포화 또는 반전 패턴 | • 잘못된 용량 | • 패턴에 대한 총 복용량의 범위를 시도 |
| • 불충분한 젤 커버리지 | • 그리드가 젤로 균일하게 덮여 있는지 확인 | |
| • 그레이스케일 패턴에 대해 다른 값을 시도하십시오. | ||
| 흐릿한 패턴 | • 패터닝 중 집중력 저하 | • 샘플과 동일한 높이에서 반복 PRIMO 교정 |
| • 잘못된 교정 | • 패터닝 하기 전에 그리드 호일에 초점 | |
| • 패터닝을 위해 패턴을 추가 영역으로 분할 | ||
| 패턴 외부의 ECM 부착 | • 젤 또는 먼지의 반사 | • 패턴화 전에 젤이 건조해지 확인 |
| • 커버슬립과 객관적인 렌즈가 깨끗한지 확인 | ||
| 패턴화 후 보이지 않는 ECM | • 사진 표백 | • 이미징 전에 ECM에 대한 광 노출 최소화 |
| • 패터닝 중 잘못된 용량 | • 패턴에 대한 총 용량 값의 범위를 시도 | |
| • ECM 잠복기 부족 | • ECM의 인큐베이션 시간 증가 | |
| 셀 시드 | ||
| 세포 응집 | • 소화 과잉 | • 부착 셀의 방출을 위해 트립신 또는 시간의 낮은 비율을 사용 |
| • 높은 세포 밀도 | • 더 낮은 컨플루엔티지에서 세포를 통로 및/또는 소화 | |
| • 방출 중에 세포를 동요하지 마십시오. | ||
| • 부드럽게 피펫 셀 솔루션 또는 셀 스트레이너를 사용 | ||
| 패턴 영역을 준수하지 않는 셀 | • ECM은 세포 유형에 적합하지 않습니다. | • 다른 ECM 농도 및 구성을 시도 |
| • 세포 생존율은 종자 전에 감소 | • 세포 배양 및 세포 방출 조건이 세포를 손상시키지 않도록 | |
| 접착 후 팽창하지 않는 셀 | • ECM 또는 패턴은 세포 유형에 적합하지 않습니다. | • 다른 패턴과 ECM을 시도 |
| • 경우에 따라 더 연속 호일 (R1.2/20 대 R2/1) 세포 확장을 촉진 할 수 있습니다 |
표 1: 마이크로패터닝 중 잠재적인 문제. 이 표는 사용자가 마이크로패터닝 또는 셀 시드 중에 발생할 수 있는 몇 가지 문제를 설명합니다. 각 문제에 대해 잠재적인 원인과 문제 해결이 제공됩니다. 도 8에서는 일부 문제의 대표적인 이미지를 볼 수 있습니다.
토론
현대식 고급 전자 현미경 및 소프트웨어 패키지는 이제 수백~수천 개의 위치를 표적으로 하고 이미지화할 수 있는 간소화된 자동 냉동 저온-EM 및 cryo-ET 데이터 수집을 지원합니다32,33,34,35. 전셀 cryo-ET 워크플로우에 대한 한 가지 중요한 제한 요소는 그리드당 수집 가능한 대상을 충분히 확보하는 것입니다. 최근에는 저온-EM용 마이크로패터닝 그리드용 프로토콜을 개발했으며, 데이터 수집 효율이 향상되어 데이터 수집 효율이 향상되었습니다16,17,18. 여기서 프로토콜은 1차 드로소필라 뉴런및 배양된 인간 세포주(감염되지 않은 또는 RSV-감염)의 저온-ET 연구를 위해 마이크로패턴 TEM 그리드에 상용 마이크로패턴 시스템을 사용하기 위한 것이다. 이 마이크로패턴 시스템은 다재다능하며, 특정 실험 목표에 맞게 최적화하고 조정할 수 있는 다양한 단계를 제공합니다. TEM 및 형광 현미경 검사를 가진 사용자는 그리드 준비 및 마이크로 패터닝에 신속하게 숙련 될 수 있습니다. 주의 깊은 연습을 통해 몇 번의 반복 후에 좋은 결과를 달성할 수 있어야 합니다. 아래에서 사용 가능한 옵션, 사용자 고려 사항, 잠재적 이점 및 cryo-EM용 마이크로패터닝의 향후 응용 프로그램에 대해 설명합니다.
전체 셀 저온-ET의 중요한 고려 사항 중 하나는 EM 그리드 선택입니다. EM 그리드는 메시 프레임(또는 구조지지대)과 호일(또는 필름)의 두 부분으로 구성되며, 이는 세포가 성장할 연속 또는 구멍이 뚫리는 필름 표면이다. 구리 메쉬 그리드는 일반적으로 단백질과 고립 된 복합체의 저온-EM에 사용됩니다. 그러나, 그들은 구리의 세포 독성으로 인해 전세포 극저온-ET에 적합하지 않습니다. 대신, 금 메쉬는 일반적으로 세포 단층 촬영에 사용됩니다. 다른 옵션으로는 니켈 또는 티타늄이 포함되며, 이는 강성 증가와 같은 금에 대한 이점을 제공할 수 있습니다16. EM 그리드는 다양한 응용 프로그램을 지원하기 위해 서로 다른 메시 치수로 사용할 수 있습니다. 메시 크기가 클수록 전체 표본 취약성이 증가하는 비용이 들지만 그리드 바와 틸트 시리즈 수집을 위해 사용할 수 있는 영역 사이에 셀이 더 많이 성장할 수 있는 공간을 제공합니다. 가장 일반적으로 사용되는 호일은 천공 또는 구멍이 없는 무정형 탄소(예: 퀀티포일 또는 C-플랫 그리드)입니다. 생물학적 표적은 탄소의 구멍을 통해 또는 전자 반투명 탄소를 통해 서 이미지될 수 있다. 구멍이 각각 1과 2 μm 간격이 있는 2 μm 너비인 R 2/1 또는 R 2/2와 같은 그리드는 많은 수의 구멍을 제공하므로 데이터 수집을 위한 많은 수의 잠재적 영역을 제공합니다. 그러나 일부 세포는 R 1.2/20 그리드 또는 연속 탄소와 같은 더 균일한 표면에서 더 잘 성장하고 확장될 수 있습니다. 집중 이온 빔 밀링(cryo-FIB)에 의한 다운스트림 샘플 처리의 경우, 호일은 밀링을 통해 제거되어 기본 필름의 지속적인 존재에 대한 우려를 줄입니다. 메쉬와 마찬가지로 다른 재료의 호일도 사용할 수 있으며, 여기에 제공되는 패터닝 프로토콜은 SiO2 그리드에 동등하게 적합합니다. 일반적으로 사용되는 그리드에는 전신 냉동고-ET용 골드 콴티포일, 연속 탄소 또는 SiO2 필름 200 메쉬 그리드(그리드 바 간~90μm 간격)가 포함됩니다.
패턴을 디자인할 때 고려해야 할 사항이 여러 가지가 있습니다. 이러한 결정의 대부분은 실험의 세포 유형 및 목적에 의해 인도된다. 좋은 출발점은 배양에서 세포의 모양과 치수를 근사화하는 패턴을 선택하는 것입니다. 많은 연구는 세포 성장과 사이토셀레탈 배열에 패턴 모양의 중요 한 효과 입증 13,36,37. 이 관심의 대상을 변경할 수 있다면 패턴 디자인 중에 특별한주의를 기울여야합니다. 각 세포 유형에 대한 여러 패턴은 세포 접착 및 성장을 촉진하는 패턴을 결정하기 위해 테스트되었습니다. 마이크로패턴 시스템의 유연성은 단일 그리드에서 여러 패턴을 테스트하고 단일 실험 내에서 서로 다른 그리드에 대한 패턴을 변경할 수 있습니다. 여기에 사용되는 패턴과 같은 더 큰 패턴 (~50-90 μm)은 여러 세포가 패턴의 단일 영역에 부착할 가능성을 높이고 세포가 접착 후 확장 및 확장 할 수 있게합니다. 더 제한된 패턴(20-30 μm)은 집중이온 빔 밀링(cryo-FIB) 실험과 같이 세포 분리가 세포 팽창보다 더 중요한 실험에 적합할 수 있다. 단층 촬영 응용 프로그램의 경우 기울기 축의 영향을 고려해야 할 수 있습니다. 패턴이 배치되어 모든 세포가 한 방향으로 서로 평행하게 자라도록 하는 경우, 모든 세포가 현미경 단계에 로드될 때 틸트축에 수직으로 되어 데이터의 품질이 낮아질 수 있다.
패턴이 없는 그리드에서 셀은 종종 TEM에 의해 이미지를 식별할 수 없는 그리드 막대를 우선적으로 부착합니다. 패턴 그리드에서도 세포는 패턴이 있는 탄소 호일과 그리드 바 모두에 부분적으로 그리드 사각형의 모서리에 위치하는 것으로 관찰됩니다. 최근에는 마이크로패터닝이 그리드 bar18 위에 셀의 일부를 의도적으로 배치하는 데 사용되었습니다. 이것은 호일에 전체 세포 주변을 갖는 것이 중요하지 않은 실험에 대해 고려될 수 있습니다. 이것은 다일 동안 증가하는 1 차적인 뉴런과 같은 단 하나 그리드 광장 보다는 더 큰 성장할 수 있는 세포에 특히 중요할 수 있습니다.
패턴을 디자인하는 데 사용할 수 있는 많은 도구가 있습니다. 여기서, 패턴은 임의의 차원에서 800픽셀 미만으로 제한되어 패턴을 임의의 각도로 회전할 수 있고 이 마이크로패터닝 시스템에 의해 단일 프로젝션으로 패턴화할 수 있는 최대 영역 내에 여전히 맞출 수 있었다. 이를 통해 사용자는 현미경의 그리드 방향에 관계없이 그리드와 함께 패턴을 올바르게 지향할 수 있습니다. 여기서 그리드는 6개의 패터닝 영역으로 나뉘었다. 주로 그리드의 다른 영역 간에 포커스 조정이 가능합니다. 특히 금 격자는 매우 가단적이며 유리에 완전히 평평하게 놓이지 않을 수 있습니다. 깨끗하고 세련된 패터닝 결과에 적합한 초점이 필수적입니다. 분할된 패턴을 사용함으로써 패턴 처리 과정에서 그리드가 약간 이동하는 경우 패턴 위치에 대한 사소한 조정만 이루어져야 하지만 일반적으로 PDMS 스텐실을 사용하여 PLPP 젤을 사용할 때는 문제가 되지 않습니다. 마지막으로 그리드의 중앙 4개 그리드 사각형은 패턴이 없는 상태로 유지되었습니다. 이는 사용자가 그리드의 중심을 명확하게 식별할 수 있도록 지원하며, 이는 상관 관계 이미징 실험에 매우 유용합니다.
이 마이크로 패턴 화 시스템의 패터닝 소프트웨어인 Leonardo는 스티치및 이 프로토콜의 범위를 벗어난 PDF로 패턴을 가져올 수 있는 기능과 같은 고급 기능을 갖추고 있습니다. 이 소프트웨어에는 TEM 그리드에서 사용할 수 있는 미세 구조 감지 및 자동화된 패턴 위치 지정도 포함되어 있습니다. 이 기능은 그리드가 매우 평평하고 서로 다른 영역 간의 초점을 조정할 필요 없이 패턴을 조정할 수 있을 때 가장 유용합니다.
ECM 단백질의 선택은 세포 접착 및 확장에 중요한 영향을 미칠 수 있습니다. 일부 세포는 특정 기판에 성장 할 때 생리적 변화를 겪는 것으로 알려져있습니다38. 다중 ECM 단백질과 농도는 문헌에 보고된 사전 작업을 기반으로 모든 새로운 세포 유형에 대해 테스트되었다. 라미닌, 피브리노겐, 섬유넥틴 및 콜라겐은 배양 된 세포에 널리 사용되며 다른 데이터를 사용할 수없는 경우 출발점으로 사용할 수 있습니다. 그러나, 다른 ECM 단백질은 또한 일반적으로 사용되는 ECM 단백질이 세포에 대한 적절한 준수 특성을 부여하지 못하는 경우에 고려되어야 합니다. 이것은 1 차적인 Drosophila 뉴런을 위해 특히 사실이었습니다, 식물 렉틴 concanavalin A의 고농도는 적당한 세포 준수를 위해 필요했기 때문에. ECM과의 세포 접착 및 성장의 호환성은 TEM 그리드로 전환하기 전에 유리 접시 또는 슬라이드의 패턴화로 테스트할 수 있습니다. 이 사전 선별 접근 방식은 많은 수의 조합을 검사해야 하는 경우 시간과 비용 효율적입니다. 형광으로 공주된 ECM 단백질의 포함은 패터닝의 성공과 품질을 평가하는 데 유용합니다.
세포 시싱은 마이크로패싱6,16,39의 유무에 관계없이 전체 세포 저온-ET를 위한 가장 중요한 단계 중 하나입니다. 1 차 적인 Drosophila 또는 그밖 뉴런을 위해, 연약하고, 현탁액에 있는 불안정하고, 양에서 제한될 수 있고, 단 하나 종자 접근은 감시되고, 순차적인 세포 파종이상 선호됩니다. Drosophila 뉴런에 대한 프로토콜에 설명된 바와 같이 최적화된 세포 밀도에서 단일 파종 단계는 대부분의 세포 유형에 대한 실행 가능한 옵션입니다. 그러나, 또한 낮은 초기 농도에서 기판에 세포를 종자하고 여기에 다른 문헌18에 설명된 바와 같이 모니터링된 방식으로 더 많은 세포를 추가할 수 있다. 이 순차적 시드는 경우에 따라 보다 일관된 결과를 제공할 수 있습니다. 표준 세포 배양과 마찬가지로 세포 생존가능성을 유지하고 격리 시 세포 응집력을 최소화하기 위해 항상 주의를 기울여야 합니다.
마이크로패터닝으로 처음 시작할 때 최종 결과에 해로운 몇 가지 잠재적 인 함정이 있습니다. 신중한 그리드 처리 및 멸균 기술, PLPP 젤의 균일한 분포, 패터닝 중 적절한 용량 및 초점, 시드 전에 세포 생존가능성의 유지가 성공을 위한 가장 중요한 고려 사항 중 하나입니다. 일부 잠재적 인 문제 및 해결 방법 목록이 표 1에 모였다.
마이크로패턴 그리드는 세포를 위치시켜 그리드 전반에 걸쳐 일관된 셀 밀도를 설정하고 틸트 시리즈 컬렉션16,18에 적합한 영역에 관심 영역을 배치하는 데 사용할 수 있습니다. 세포의 배치 및 위치는 극저온-CLEM 실험에서 상관 관계를 위한 신탁 마커로 사용될 수 있으며, 깨지기 쉬운 파인더 그리드 및 형광 선량마커의 필요성을 감소시킵니다. 그러나, 그러한 서두체 마커는 여전히 서브 마이크로미터 정확도 상관관계에 유용할 수 있다는 점에 유의해야 한다29,40. 더욱이, 격리된 세포의 균등한 분포는 또한 라멜라절단할 수 있는 세포의 수를 최대화하기 위하여 집중이온 빔 밀링(cryo-FIB) 실험에 매우 유익하다16.
저온-EM 워크플로우에 마이크로패턴이 추가되면 데이터 처리량이 측정 가능한 개선이 가능하고 잠재적으로 새로운 실험을 가능하게 할 수 있습니다. 기술이 더욱 채택되고 개발됨에 따라 ECM 그라데이션, 다중 ECM 기탁 및 미세 구조 어셈블리를 포함한 마이크로패턴의 고급 응용 분야는 극저온-ET의 기능을 더욱 확장하여 전체 세포 맥락에서 생물학적 표적과 공정을 연구할 것입니다.
공개
저자는 공개 할 것이 없습니다.
감사의 말
우리는 박사 질 Wildonger, 박사 시희 Z. 양, 그리고 유세핀 W. 미첼 박사생화학의 학과, 위스콘신 대학, 매디슨은 아낌없이 elav-Gal4, UAS-CD8::GFP 플라이 스트레인 (블루밍턴 주식 센터, #5146)을 공유해 주셔서 감사합니다. 또한 오렐리엔 두보인 박사, 로랑 시퀴어 박사, 알베올의 마리 샬럿 마누스 박사, 나노스케일 연구소의 세르주 카두라 씨에게 이 프로젝트를 통해 후한 지원을 부탁드립니다. 이 작품은 위스콘신 대학에 의해 부분적으로 지원되었다, 매디슨, 위스콘신 대학의 생화학학과, 매디슨, 공중 보건 서비스 보조금 R01 GM114561, R01 GM104540, R01 GM104540-03W1, U24 GM139168 E.R.W. 및 R01 AI150475 이 연구의 일부는 NIH 보조금 U24 GM129547에 의해 지원되고 OHSU에서 PNCC에서 수행하고 EMSL (grid.436923.9), 생물 및 환경 연구 사무실후원 과학 사용자 시설의 DOE 사무실을 통해 액세스. 우리는 또한 위스콘신 대학, 매디슨의 생화학학과의 Cryo-EM 연구 센터에서 시설과 계측의 사용에 감사드립니다.
자료
| Name | Company | Catalog Number | Comments |
| 0.1% (w/v) Poly-L-Lysine | Sigma | P8920-100ML | |
| 0.22 µm syringe filters PVDF membrane | Genesee | 25-240 | |
| 22x60-1 Glass cover slip | Fisher | 12545F | |
| 5/15 Tweezers | EMS (Dumont) | 0203-5/15-PO | |
| Antibiotic-Antimycotic (100X) | ThermoFisher (Gibco) | 15240096 | |
| BEAS-2B cells | ATCC | CRL-9609 | |
| Collagen I, bovine | ThermoFisher (Gibco) | A1064401 | |
| Concanavalin A, Alexa Fluor 350 Conjugate | ThermoFisher (Invitrogen) | C11254 | |
| DMEM | Fisher (Lonza) | BW12-604F | |
| EtOH | Fisher (Decon Labs) | 22-032-600 | |
| Fetal Bovine Serum | ATCC | 30-2020 | |
| Fibrinogen From Human Plasma, Alexa Fluor 647 Conjugate | ThermoFisher (Invitrogen) | F35200 | |
| Fibronectin Bovine Protein, Plasma | ThermoFisher (Gibco) | 33010018 | |
| Glass bottom dish | MatTek | P35G-1.5-20-C | |
| Glucose | VWR | 0643-1KG | |
| Grid prep holder | EMS | 71175-01 | |
| HeLa cells | ATCC | CCL-2 | |
| Hemacytometer | Fisher (SKC, Inc.) | 22600100 | |
| HEPES | Fisher (ACROS Organics) | AC172572500 | |
| Hoechst 33342 | ThermoFisher (Invitrogen) | H3570 | |
| Insulin | Fisher (Sigma Aldrich) | NC0520015 | |
| KCl | MP Bio | 194844 | |
| KH2PO4 | Fisher (ACROS Organics) | AC212595000 | |
| Leica-DMi8 | Leica Microsystems | Can be customized with camera, stage, and objective attachments | |
| Leonardo | Alvéole | https://www.alveolelab.com/our-products/leonardo-photopatterning-software/ | |
| Liberase Research Grade | Fisher (Supply Solutions) | 50-100-3280 | |
| LIVE/DEAD Viability/Cytotoxicity Kit | ThermoFisher (Invitrogen) | L3224 | |
| Microscope camera | Hammamatsu | C13440-20CU | |
| Motorized stage | Märzhäuser Wetzlar | 00-24-599-0000 | |
| NaCl | Fisher (Fisher BioReagents) | BP358-1 | |
| NaH2PO4 | Fisher (ACROS Organics) | AC207802500 | |
| NaOH | Fisher (Alfa Aesar) | AAA1603736 | |
| PBS | Corning | 21-040-CV | |
| PDMS stencils | nanoscaleLABS | PDMS_STENCILS_EM | https://www.alveolelab.com/our-products/pdms-stencil-multiwell-plate/ |
| PEG-SVA | nanoscaleLABS | PEG-SVA-1GR | mPEG-Succinimidyl Valerate, MW 5,000 |
| Penicillin | Fisher (Research Products International Corp) | 50-213-641 | |
| pH strips | Fisher (Millipore Sigma) | M1095350001 | pH probe can also be used |
| PLPP gel | nanoscaleLABS | PLPP-GEL-300UL | https://www.alveolelab.com/our-products/plpp-photoactivatable-reagent/ |
| PRIMO | Alvéole | https://www.alveolelab.com/our-products/primo-micropatterning/ | |
| pSynkRSV-I19F (BAC containing RSV A2-mK+ antigenomic cDNA ) | BEI Resources | NR-36460 | https://www.beiresources.org/Catalog/BEIPlasmidVectors/NR-36460.aspx |
| Quantifoil grids | EMS (Quantifoil) | Q2100AR1 | 2 µm holes spaced 1 µm apart, other dimensions are available |
| RPMI | Fisher (Lonza) | BW12-702F | |
| RSV A2-mK+ | see entry for pSynkRSV-19F | - | Described in Hotard et al. [22]. Can be generated from pSynkRSV-ll9F |
| Schneider's Media | ThermoFisher (Gibco) | 21720-024 | |
| SerialEM | SerialEM (https://bio3d.colorado.edu/SerialEM/ ) | https://bio3d.colorado.edu/SerialEM/ | |
| Straight tweezers | EMS (Dumont) | 72812-D | |
| Streptomycin | Fisher (Fisher BioReagents) | BP910-50 | |
| Sucrose | Avantor | 4097-04 | |
| Tetracycline | Sigma | T8032-10MG | |
| Titan Krios electron microscope | ThermoFisher | 300kV, with direct electron detector camera and energy filter | |
| Trypsin | ThermoFisher (Gibco) | 15090046 | |
| Tube Revolver/Rotator | Fisher (Thermo Scientific) | 11676341 | |
| UAS:mcD8:GFP Drosophila fly strain | Bloomington Drosophila Stock Center | 5146 | http://flybase.org/reports/FBtp0002652.html |
참고문헌
- Nogales, E., Scheres, S. H. Cryo-EM: A unique tool for the visualization of macromolecular complexity. Molecular Cell. 58 (4), 677-689 (2015).
- Martynowycz, M. W., Gonen, T. From electron crystallography of 2D crystals to MicroED of 3D crystals. Current Opinion in Colloid and Interface Science. 34, 9-16 (2018).
- Wagner, J., Schaffer, M., Fernandez-Busnadiego, R. Cryo-electron tomography-the cell biology that came in from the cold. FEBS Letters. 591 (17), 2520-2533 (2017).
- Wan, W., Briggs, J. A. Cryo-electron tomography and subtomogram averaging. Methods in Enzymology. 579, 329-367 (2016).
- Bäuerlein, F. J., Pastor-Pareja, J. C., Fernández-Busnadiego, R. Cryo-electron tomography of native Drosophila tissues vitrified by plunge freezing. bioRxiv. , 437159(2021).
- Hampton, C. M., et al. Correlated fluorescence microscopy and cryo-electron tomography of virus-infected or transfected mammalian cells. Nature Protocols. 12 (1), 150-167 (2017).
- Hsieh, C. E., Leith, A., Mannella, C. A., Frank, J., Marko, M. Towards high-resolution three-dimensional imaging of native mammalian tissue: Electron tomography of frozen-hydrated rat liver sections. Journal of Structural Biology. 153 (1), 1-13 (2006).
- Al-Amoudi, A., Norlen, L. P., Dubochet, J. Cryo-electron microscopy of vitreous sections of native biological cells and tissues. Journal of Structural Biolology. 148 (1), 131-135 (2004).
- Rigort, A., et al. Focused ion beam micromachining of eukaryotic cells for cryoelectron tomography. Proceedings of the National Academy of Sciences. 109 (12), 4449-4454 (2012).
- Gorelick, S., et al. PIE-scope, integrated cryo-correlative light and FIB/SEM microscopy. Elife. 8, 45919(2019).
- Wu, G. H., et al. Multi-scale 3D cryo-correlative microscopy for vitrified cells. Structure. 28 (11), 1231-1237 (2020).
- Turk, M., Baumeister, W. The promise and the challenges of cryo-electron tomography. FEBS Letters. 594 (20), 3243-3261 (2020).
- Théry, M. Micropatterning as a tool to decipher cell morphogenesis and functions. Journal of Cell Science. 123 (24), 4201-4213 (2010).
- Tseng, Q., et al. Spatial organization of the extracellular matrix regulates cell-cell junction positioning. Proceedings of the National Academy of Sciences. 109 (5), 1506-1511 (2012).
- Hardelauf, H., et al. Micropatterning neuronal networks. Analyst. 139 (13), 3256-3264 (2014).
- Toro-Nahuelpan, M., et al. Tailoring cryo-electron microscopy grids by photo-micropatterning for in-cell structural studies. Nature Methods. 17 (1), 50-54 (2020).
- Engel, L., et al. Extracellular matrix micropatterning technology for whole cell cryogenic electron microscopy studies. Journal of Micromechanics and Microengineering. 29 (11), (2019).
- Engel, L., et al. Lattice micropatterning for cryo-electron tomography studies of cell-cell contacts. bioRxiv. , 272237(2021).
- Sibert, B. S., Kim, J. Y., Yang, J. E., Wright, E. R. Whole-cell cryo-electron tomography of cultured and primary eukaryotic cells on micropatterned TEM grids. bioRxiv. , 447251(2021).
- Egger, B., van Giesen, L., Moraru, M., Sprecher, S. G. In vitro imaging of primary neural cell culture from Drosophila. Nature Protocols. 8 (5), 958-965 (2013).
- Lu, W., Lakonishok, M., Gelfand, V. I. Kinesin-1-powered microtubule sliding initiates axonal regeneration in Drosophila cultured neurons. Molecular Biology of the Cell. 26 (7), 1296-1307 (2015).
- Ke, Z., et al. The morphology and assembly of respiratory syncytial virus revealed by cryo-electron tomography. Viruses. 10 (8), (2018).
- Stobart, C. C., et al. A live RSV vaccine with engineered thermostability is immunogenic in cotton rats despite high attenuation. Nature Communications. 7, 13916(2016).
- Hotard, A. L., et al. A stabilized respiratory syncytial virus reverse genetics system amenable to recombination-mediated mutagenesis. Virology. 434 (1), 129-136 (2012).
- Schindelin, J., et al. Fiji: an open-source platform for biological-image analysis. Nature Methods. 9 (7), 676-682 (2012).
- Mastronarde, D. N. Automated electron microscope tomography using robust prediction of specimen movements. Journal of Structural Biology. 152 (1), 36-51 (2005).
- Kremer, J. R., Mastronarde, D. N., McIntosh, J. R. Computer visualization of three-dimensional image data using IMOD. Journal of Structural Biology. 116 (1), 71-76 (1996).
- Tang, G., et al. EMAN2: an extensible image processing suite for electron microscopy. Journal of Structural Biology. 157 (1), 38-46 (2007).
- Yang, J. E., Larson, M. R., Sibert, B. S., Shrum, S., Wright, E. R. CorRelator: Interactive software for real-time high precision cryo-correlative light and electron microscopy. Journal of Structural Biology. , 107709(2021).
- Ke, Z., et al. Promotion of virus assembly and organization by the measles virus matrix protein. Nature Communications. 9 (1), 1736(2018).
- Kim, J., Yang, S., Wildonger, J., Wright, E. A new in situ neuronal model for cryo-ET. Microscopy and Microanalysis. 26 (2), 130-132 (2020).
- Bouvette, J., et al. Beam image-shift accelerated data acquisition for near-atomic resolution single-particle cryo-electron tomography. Nature Communications. 12 (1), 1957(2021).
- Schorb, M., Haberbosch, I., Hagen, W. J. H., Schwab, Y., Mastronarde, D. N. Software tools for automated transmission electron microscopy. Nature Methods. 16 (6), 471-477 (2019).
- Weis, F., Hagen, W. J. H., Schorb, M., Mattei, S. Strategies for optimization of cryogenic electron tomography data acquisition. Journal of Visual Experiments. (169), e62383(2021).
- Chreifi, G., Chen, S., Jensen, G. J. Rapid tilt-series method for cryo-electron tomography: Characterizing stage behavior during FISE acquisition. Journal of Structural Biology. 213 (2), 107716(2021).
- Anderson, D. E., Hinds, M. T. Endothelial cell micropatterning: methods, effects, and applications. Annals of Biomedical Engineering. 39 (9), 2329-2345 (2011).
- McWhorter, F. Y., Wang, T., Nguyen, P., Chung, T., Liu, W. F. Modulation of macrophage phenotype by cell shape. Proceedings of the National Academy of Sciences. 110 (43), 17253-17258 (2013).
- Kleinman, H. K., Luckenbill-Edds, L., Cannon, F. W., Sephel, G. C. Use of extracellular matrix components for cell culture. Analytical Biochemistry. 166 (1), 1-13 (1987).
- Fassler, F., Zens, B., Hauschild, R., Schur, F. K. M. 3D printed cell culture grid holders for improved cellular specimen preparation in cryo-electron microscopy. Journal of Structural Biology. 212 (3), 107633(2020).
- Schellenberger, P., et al. High-precision correlative fluorescence and electron cryo microscopy using two independent alignment markers. Ultramicroscopy. 143, 41-51 (2014).
재인쇄 및 허가
JoVE'article의 텍스트 или 그림을 다시 사용하시려면 허가 살펴보기
허가 살펴보기This article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. 판권 소유