
Method Article
Micropatronaje Transmisión Microscopía Microscopía Rejillas para Dirigir el Posicionamiento Celular dentro de los Flujos de Trabajo de Tomografía Crioelectrónica de Celda Entera
* Estos autores han contribuido por igual
En este artículo
Resumen
El objetivo de este protocolo es dirigir la adhesión y el crecimiento celular a áreas específicas de rejillas para criomicroscopía electrónica. Esto se logra mediante la aplicación de una capa antiincrustante que se ablaciona en patrones especificados por el usuario seguido de la deposición de proteínas de matriz extracelular en las áreas modeladas antes de la siembra celular.
Resumen
La crioelectrónica de células enteras (crio-ET) es una tecnología poderosa que se utiliza para producir estructuras de resolución a nivel nanométrico de macromoléculas presentes en el contexto celular y preservadas en un estado congelado-hidratado casi nativo. Sin embargo, existen desafíos asociados con el cultivo y / o la adhesión de las células a las rejillas TEM de una manera que sea adecuada para la tomografía mientras se retienen las células en su estado fisiológico. Aquí, se presenta un protocolo detallado paso a paso sobre el uso del micropatronaje para dirigir y promover el crecimiento de células eucariotas en las cuadrículas TEM. Durante el micropatronaje, el crecimiento celular se dirige depositando proteínas de matriz extracelular (ECM) dentro de patrones y posiciones especificadas en la lámina de la rejilla TEM, mientras que las otras áreas permanecen recubiertas con una capa antiincrustante. La flexibilidad en la elección del recubrimiento de la superficie y el diseño del patrón hace que el micropatronaje sea ampliamente aplicable para una amplia gama de tipos de células. El micropatronaje es útil para estudios de estructuras dentro de células individuales, así como sistemas experimentales más complejos, como las interacciones huésped-patógeno o las comunidades multicelulares diferenciadas. El micropatronaje también puede integrarse en muchos flujos de trabajo de crio-ET de células enteras aguas abajo, incluida la microscopía electrónica y de luz correlativa (crio-CLEM) y el fresado de haz de iones enfocados (cryo-FIB).
Introducción
Con el desarrollo, la expansión y la versatilidad de la microscopía crioelectrónica (crio-EM), los investigadores han examinado una amplia gama de muestras biológicas en un estado casi nativo desde la resolución macromolecular (~ 1 nm) hasta la alta (~ 2 Å). Las técnicas de crio-EM de una sola partícula y de difracción de electrones se aplican mejor a macromoléculas purificadas en solución o en estado cristalino, respectivamente1,2. Mientras que la crioelectrónica (crio-ET) es especialmente adecuada para estudios estructurales y ultraestructurales casi nativos de objetos grandes y heterólogos como bacterias, virus pleomórficos y células eucariotas3. En crio-ET, la información tridimensional (3D) se obtiene inclinando físicamente la muestra en la etapa del microscopio y adquiriendo una serie de imágenes a través de la muestra en diferentes ángulos. Estas imágenes, o series de inclinación, a menudo cubren un rango de +60/-60 grados en incrementos de uno a tres grados. La serie de inclinación se puede reconstruir computacionalmente en un volumen 3D, también conocido como tomograma4.
Todas las técnicas de crio-EM requieren que la muestra se incruste en una capa delgada de hielo vítreo amorfo, no cristalino. Una de las técnicas de criofijación más utilizadas es la congelación por inmersión, donde la muestra se aplica a la rejilla EM, se borra y se sumerge rápidamente en etano líquido o una mezcla de etano líquido y propano. Esta técnica es suficiente para la vitrificación de muestras de <100 nm a ~10 μm de espesor, incluyendo células humanas cultivadas, como las células HeLa5,6. Las muestras más gruesas, como mini-organoides o biopsias de tejido, de hasta 200 μm de espesor, pueden ser vitrificadas por congelación a alta presión7. Sin embargo, debido al aumento de la dispersión de electrones de muestras más gruesas, el espesor de la muestra y el hielo para crio-ET se limita a ~ 0.5 - 1 μm en microscopios electrónicos de transmisión de 300 kV. Por lo tanto, la crio-ET de células enteras de muchas células eucariotas se limita a la periferia celular o extensiones de células a menos que se utilicen pasos adicionales de preparación de muestras, como la crioseccionamiento8 o el fresado de haz de iones enfocados9,10,11.
Una limitación de muchos experimentos de imágenes crio-ET de células enteras es el rendimiento de la recopilación de datos12. A diferencia de la crio-EM de una sola partícula, donde miles de partículas aisladas a menudo se pueden obtener imágenes desde un solo cuadrado de cuadrícula TEM, las células son grandes, se extienden y deben cultivarse a una densidad lo suficientemente baja como para permitir que las células se conserven en una capa delgada de hielo vítreo. A menudo, la región de interés se limita a una característica o subárea particular de la célula. Lo que limita aún más el rendimiento es la propensión de las células a crecer en áreas que no son susceptibles de imágenes TEM, como en o cerca de las barras de rejilla TEM. Debido a los factores impredecibles asociados con el cultivo celular en las redes TEM, se necesitan desarrollos tecnológicos para mejorar la accesibilidad de las muestras y el rendimiento para la adquisición de datos.
El micropatronaje de sustrato con proteínas adherentes de matriz extracelular (ECM) es una técnica bien establecida para la microscopía de luz de células vivas para dirigir el crecimiento de las células en superficies rígidas, duraderas y ópticamente transparentes, como el vidrio y otros sustratos de cultivo de tejidos13,14. El micropatronaje también se ha realizado en superficies blandas y/o tridimensionales (3D). Tales técnicas no solo han permitido el posicionamiento preciso de las células; también han apoyado la creación de redes multicelulares, como los circuitos celulares neuronales modelados15. Llevar el micropatronaje a la crio-ET no solo aumentará el rendimiento, sino que también puede abrir nuevos estudios para explorar microambientes celulares complejos y dinámicos.
Recientemente, varios grupos han comenzado a utilizar técnicas de micropatronaje en cuadrículas TEM a través de múltiples enfoques16,17. Aquí, se describe el uso de una técnica de fotopatronaje sin máscara para las cuadrículas TEM utilizando el sistema de micropatrones Alvéole PRIMO, que presenta patrones de alta resolución y sin contacto. Con este sistema de micropatronaje, se aplica una capa antiincrustante en la parte superior del sustrato, seguida de la aplicación de un fotocatalizador y la ablación de la capa antiincrustante en patrones definidos por el usuario con un láser UV. Las proteínas ECM se pueden agregar a los patrones para el cultivo celular apropiado. Este método ha sido utilizado por varios grupos para estudios crio-ET de pigmento retiniano epitelial-1 (RPE1), riñón canino Madin-Darby-II (MDCKII), fibroblasto del prepucio humano (HFF) y líneas celulares endoteliales16,17,18. Este sistema de micropatrones es compatible con múltiples sustratos de capa antiincrustante, así como con un reactivo fotocatalizador líquido o en gel. Se puede seleccionar y adaptar una variedad de proteínas ECM para la especificidad de la línea celular, lo que confiere versatilidad al usuario.
El micropatronaje se ha aplicado con éxito a una serie de proyectos dentro del laboratorio19. Aquí, se presenta un protocolo de micropatronaje, que incluye adaptaciones específicas para estudiar células HeLa cultivadas, células BEAS-2B infectadas por el virus sincitial respiratorio (VSR) y neuronas larvales primarias de Drosophila melanogaster20.
Protocolo
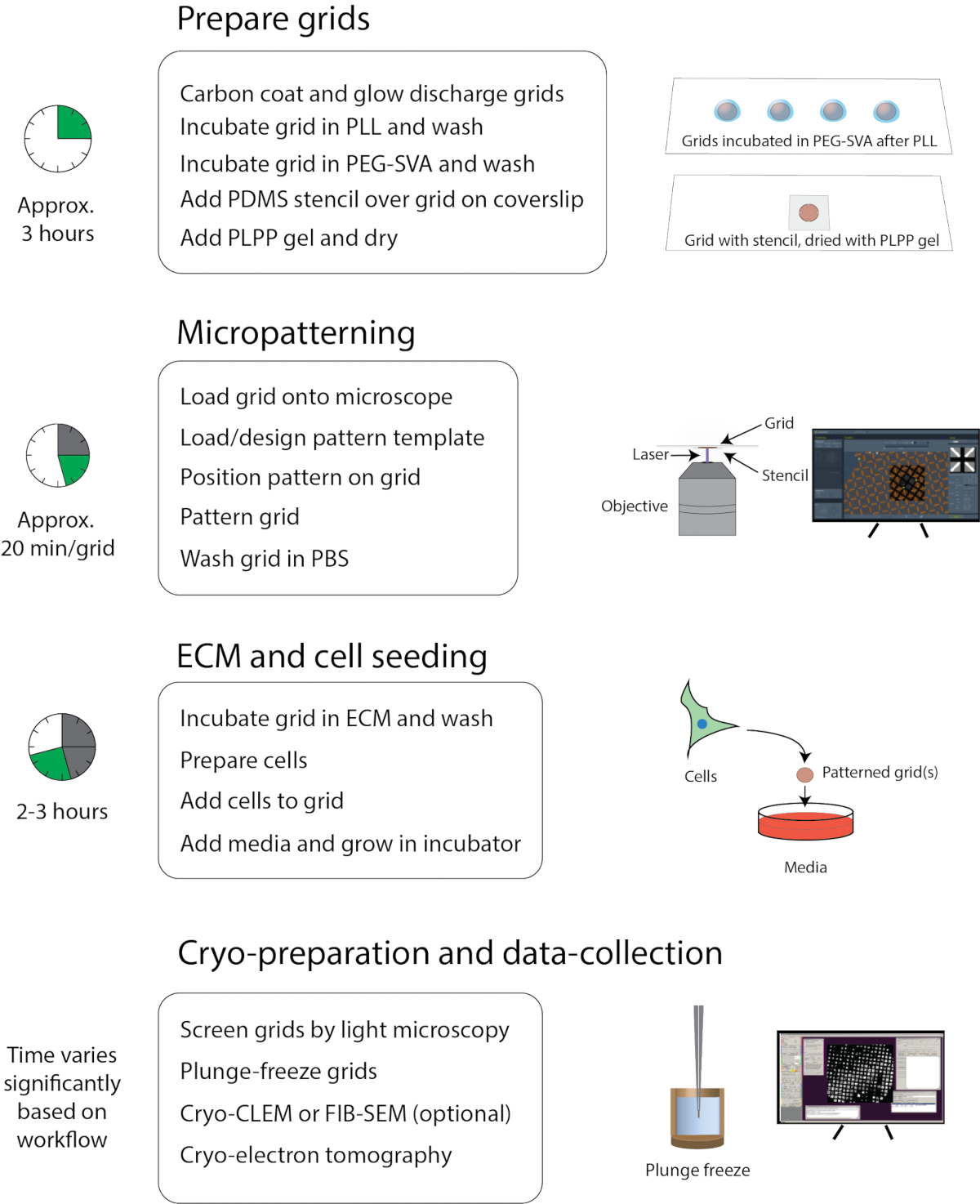
El protocolo descrito aquí es una compilación de los métodos de cultivo celular, micropatronaje e imágenes utilizados por el laboratorio Wright y el Centro de Investigación Cryo-EM de la Universidad de Wisconsin, Madison. El flujo de trabajo se presenta en la Figura 1. Hay capacitación adicional y materiales de instrucción disponibles en los siguientes sitios: https://cryoem.wisc.edu o https://wrightlab.wisc.edu
1. Preparación de rejillas para el modelado
- Transfiera las rejillas TEM a un portaobjetos de vidrio limpio, con el lado de carbono hacia arriba (el grosor estándar de la lámina de carbono es de 12 nm). Usando un evaporador de carbono, ACE600, evapore 5-8 nm de carbono adicional en las rejillas para aumentar la durabilidad general de la película de carbono.
NOTA: Este paso no es necesario para las cuadrículas de SiO2 . Este paso también se puede hacer con anticipación; almacene las rejillas recubiertas en un ambiente de baja humedad, como un desecador al vacío. - Transfiera las rejillas a un soporte de preparación de la red y descargue el vidrio de las rejillas hacia arriba. Utilizando un sistema de descarga de resplandor, descargue las rejillas de resplandor durante 60 s a 10 mA con una distancia de trabajo de 80 mm y una presión de vacío de 1.0 x 10-3 mbar. Haga esto dentro de los 15-30 minutos del siguiente paso.
NOTA: Los soportes de preparación de rejilla se pueden comprar comercialmente o hacer en casa con un trozo de papel de filtro en una pequeña placa de Petri.
2. Aplicación de la capa antiincrustante
NOTA: Se debe utilizar una técnica estéril adecuada al manipular las rejillas, y todas las soluciones deben ser estériles y / o esterilizadas por filtro.
- Transfiera las rejillas (lado de carbono hacia arriba) a un portaobjetos o cubiertas de vidrio limpio con al menos 1 cm de separación entre las rejillas. Pipetear 10 μL de poli-L-lisina (PLL) al 0,05% en cada rejilla. Incube las rejillas en una cámara húmeda, como una caja de plástico cerrada con toallas de papel húmedas, durante al menos 30 minutos.
NOTA: Este paso se puede extender a la noche. Asegúrese de que el nivel de humedad en la cámara sea suficiente para evitar que las rejillas se sequen. - Lavar cada rejilla tres veces con 15 μL de 0,1 M HEPES pH 8,5. Para cada lavado, retire la mayor parte del líquido de la rejilla con una pipeta sin dejar que la rejilla se seque. Añadir 15 μL de tampón fresco, incubar durante al menos 30 s y repetir. Dejar cada rejilla en 15 μL de 0,1 M DE HEPES después del lavado final.
NOTA: En este paso y en los pasos futuros, es importante mantener la rejilla húmeda y evitar el contacto entre la pipeta y la rejilla. - Preparar 10 μL de 100 mg/ml de polietilenglicol-succinimidilato de valerato (PEG-SVA) en 0,1 M HEPES pH 8,5 para cada rejilla. El PEG-SVA se disolverá rápidamente con una mezcla suave que dará como resultado una solución clara.
NOTA: No prepare la solución PEG-SVA con antelación. PEG-SVA tiene una vida media de 10 min a pH 8.5. Evite exponer el material de PEG-SVA a una humedad excesiva almacenándolo en un desecador o ambiente seco a -20 °C y calentándolo a temperatura ambiente antes de abrirlo. - Inmediatamente después de preparar la solución de PEG-SVA, retire la gota de 15 μL de HEPES pH 8.5 de cada rejilla (teniendo cuidado de no secar la rejilla) y agregue una gota de 10 μL de la solución de PEG-SVA. Incubar las rejillas en una cámara húmeda durante al menos 1 h.
NOTA: Este paso se puede extender a la noche. Asegúrese de que la humedad en la cámara sea suficiente para evitar que las rejillas se sequen. - Lave cada rejilla tres veces con 15 μL de agua estéril. Para cada lavado, retire la mayor parte del líquido de la rejilla con una pipeta sin dejar que la rejilla se seque, agregue 15 μL de agua dulce, incube durante al menos 30 s y repita. Deje cada rejilla en 15 μL de agua después del lavado final.
3. Aplicación de gel PLPP
- Prepare una cubierta limpia para el microscopio para cada rejilla. Complete los siguientes pasos para cada rejilla, una rejilla a la vez, para minimizar la posibilidad de que la rejilla se seque.
- Coloque una gota de agua de 1.0 μL en el centro de la cubierta para ayudar a colocar la rejilla en la cubierta y mantener la rejilla húmeda. Transfiera cuidadosamente la rejilla de la gota de agua de 15 μL a la gota de agua de 1.0 μL en el coverlip. Asegúrese de colocar el lado de carbono de la red hacia arriba.
- Coloque cuidadosamente una plantilla de polidimetilsiloxano (PDMS) sobre la rejilla, teniendo cuidado de mantener la rejilla centrada y minimizar el contacto de la plantilla con la lámina de carbono de la rejilla.
- Añadir 1,0 μL de gel de cloruro de 4-benzoilbencil-trimetilamonio (PLPP) a la rejilla. Pipete suavemente para mezclar (no toque la rejilla con la punta de la pipeta).
- Mueva el recubierto con la rejilla a un lugar oscuro para que se seque. El gel se secará en aproximadamente 15-30 min.
4. Calibración y diseño del micropatrones
- Colorea un lado de una funda de vidrio con un resaltador. Agregue líneas negras de un marcador permanente de punta fina para facilitar el enfoque. Coloque el lape de cubierta en el microscopio de tal manera que el lado coloreado se enfrente a la lente del objetivo. Usando el modo de campo brillante, concéntrate en el resaltador.
- Asegúrese de que el microscopio y el sistema de micropatronaje estén encendidos y que se establezca la ruta de luz correcta. Abra Micromanager y el software Leonardo (Plugins > Leonardo) en la computadora del microscopio.
- Seleccione calibrar y siga las instrucciones en pantalla. Ajuste el enfoque del microscopio para que la imagen proyectada en la diapositiva esté enfocada. Es posible que sea necesario reducir el tiempo de exposición. Después de la calibración, seleccione Patrón ahora.
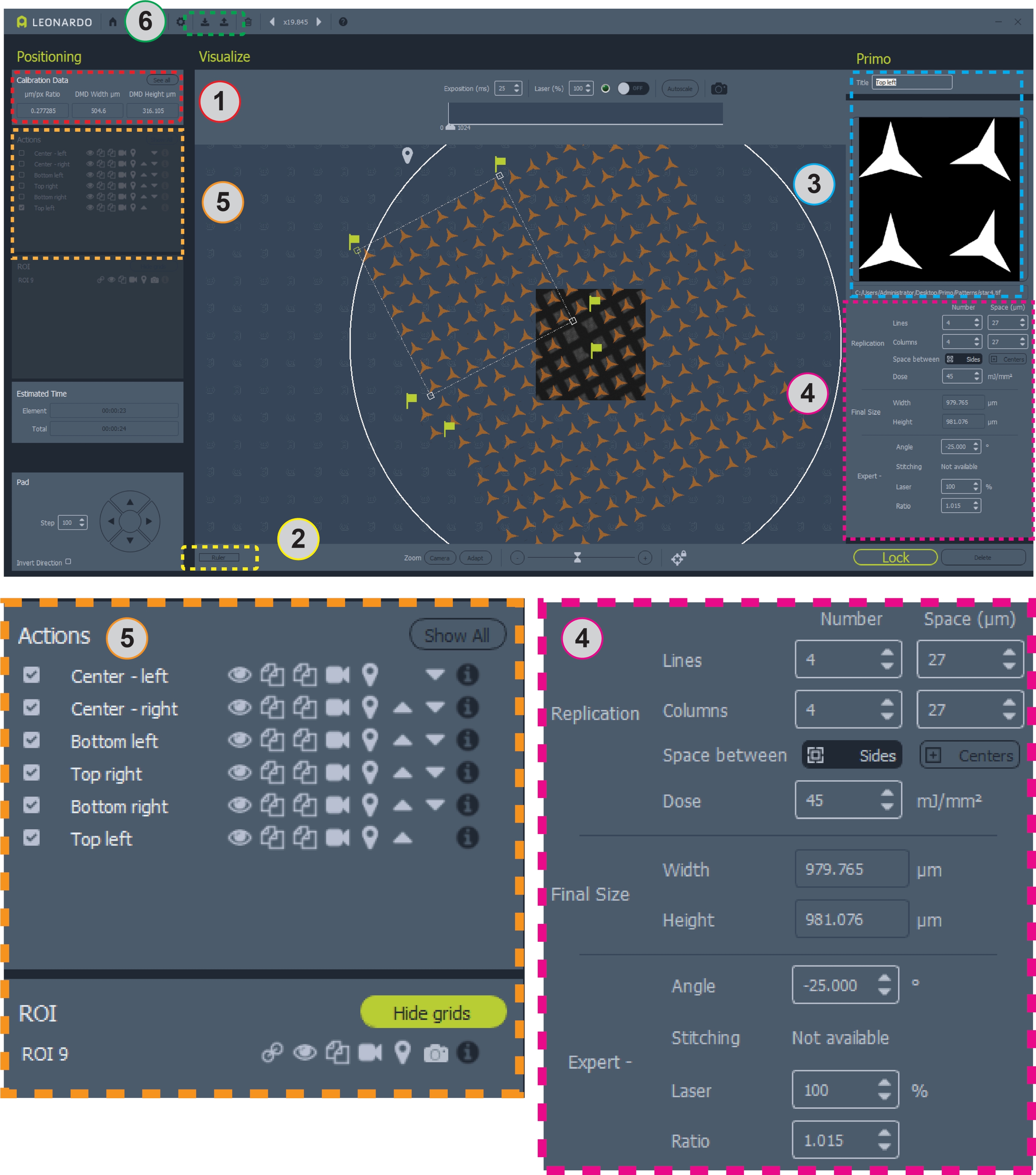
- Registre la relación micrómetro/píxel (μm/px) reportada bajo los datos de calibración en la ventana superior izquierda del programa (Figura 2, área 1). Utilice esta relación para determinar el número de píxeles que se utilizarán por micrómetro al diseñar un patrón.
- Después de la calibración, asegúrese de que el software esté abierto ahora con una vista de campo brillante en vivo desde el microscopio. Cargue una rejilla preparada en un cobertor (sección 3) en el escenario con la rejilla frente a la lente del objetivo. Coloque el escenario y ajuste el enfoque para que la cuadrícula sea visible en la ventana del software.
- Mida el tamaño de los cuadrados de cuadrícula y las barras de cuadrícula en micrómetros. El software incluye una regla activada por el botón cerca de la esquina inferior izquierda para medir la cuadrícula (Figura 2, área 2). Por ejemplo, los patrones utilizados aquí para una cuadrícula de malla de 200 corresponden a ~ 87 cuadrados de cuadrícula de × 87 μm y barras de cuadrícula de ~ 36 μm.
NOTA: El software ofrece flexibilidad para cambiar el tamaño de los patrones sobre la marcha, por lo que se pueden tolerar inexactitudes menores en la medición. - Basado en las medidas y proporciones anteriores, cree patrones con cualquier software de creación de imágenes. El tamaño mínimo de la característica con un objetivo de 20× es de 1,2 μm. Los patrones deben guardarse como archivos .tiff de 8 bits sin comprimir.
- Asegúrese de que el software no reescale las imágenes a un tamaño de píxel diferente al guardarlas. El patrón debe caber dentro de una caja de 800 × 800 píxeles, suficiente para cubrir cuatro cuadrados de cuadrícula.
NOTA: Los píxeles con un valor de 255 (blanco) se modelarán a la intensidad más alta (dosis total del láser) y los píxeles con un valor de cero (negro) no se modelarán. Cualquier píxel con un valor intermedio se modelará con una dosis de aproximadamente (X/255)*dosis total. En la Figura 3A, se utilizaron valores de píxeles de 255 y 129 para los patrones de escala de grises. Una vez que se diseña el patrón, se puede guardar y reutilizar sin modificaciones.
- Asegúrese de que el software no reescale las imágenes a un tamaño de píxel diferente al guardarlas. El patrón debe caber dentro de una caja de 800 × 800 píxeles, suficiente para cubrir cuatro cuadrados de cuadrícula.
5. Micropatronaje
- Después de la calibración, asegúrese de que el software esté abierto ahora con una vista de campo brillante en vivo desde el microscopio. Cargue una rejilla preparada en un cobertor (sección 3) en el escenario con la rejilla frente a la lente del objetivo. Coloque el escenario y ajuste el enfoque para ver la cuadrícula en el software.
- Para una ejecución inicial, diseñe una nueva plantilla. En el software, seleccione Agregar ROI (no se muestra, en la ubicación del área 3 de la Figura 2 ) y elija un círculo de 3.000 μm. Coloque el ROI del círculo sobre la cuadrícula utilizando la imagen de campo brillante en la pantalla como guía. Presione bloquear para asegurar el ROI.
- Después de bloquear el ROI en su lugar, seleccione Agregar patrón (no se muestra, en la ubicación del área 3 de la Figura 2 ). Elija el patrón diseñado en la sección 4. Divida la cuadrícula en seis regiones para permitir que el enfoque y el posicionamiento independientes en cada región tengan en cuenta las cuadrículas desiguales. Una región cuadrada de cuadrícula de 8 × 8 para cada esquina de la cuadrícula y una región cuadrada de cuadrícula de 2 × 8 a cada lado del centro, dejando el centro cuatro cuadrados de cuadrícula sin patrón (Figura 2, imagen central).
- Utilice las opciones de replicación (Figura 2, área 4) para generar copias del patrón inicial para alcanzar el número deseado de copias totales del patrón. Ajuste el espaciado entre copias para que coincida con el espaciado entre cuadrados de cuadrícula si es necesario.
- Establezca la dosis total para el patrón. 30 mJ/mm2 es un buen punto de partida. Consulte la sección de discusión para obtener más detalles.
- En Opciones de experto (Figura 2, área 4) ajuste el ángulo de la región para que coincida con el de los cuadrados de la cuadrícula. Las regiones se pueden reposicionar con el ratón. La relación (tamaño) de los patrones también se puede ajustar. Itera ajustando el ángulo, la posición, el espacio entre y la proporción del patrón hasta que los patrones se alineen con los cuadrados de la cuadrícula. Mueva la etapa del microscopio para cambiar la región de la cuadrícula en la pantalla de campo brillante en vivo.
- Pulse Bloquear para guardar los cambios realizados en la región.
- Para copiar una región, haga clic en el botón Duplicar (Figura 2, área 5, icono de dos hojas de papel) junto a su nombre en el panel Acciones de la izquierda. Para cambiar la posición, cambiar el nombre o editar la copia, haga clic en su nombre en el panel Acción.
- Repita los pasos 5.4 a 5.9 según sea necesario para rellenar todas las regiones deseadas.
- Una vez que la plantilla completa esté diseñada y posicionada, guarde el archivo de plantilla dentro del software (Figura 2, área 6, barra con icono de flecha hacia arriba en la barra de herramientas superior).
- Al cargar una plantilla guardada previamente (barra con icono de flecha hacia abajo) centre el ROI sobre la cuadrícula y presione Bloquear. Haga clic en cada región del Panel de acciones para cambiar el ángulo, la posición, la dosis y/o el archivo de patrón.
- Una vez que la plantilla y los patrones estén posicionados, desmarque todas las regiones menos una en el Panel de acción del software.
- Use la etapa del microscopio para navegar a esa región y enfocarse en la lámina de carbono. Al hacer clic en el icono Globo ocular en el panel Acción (Figura 2, área 5) se activará o desactivará la visualización de la superposición de patrón.
- Una vez que la cuadrícula esté enfocada, cierre el obturador de campo brillante y presione el ícono Reproducir en la esquina inferior derecha del software para comenzar el proceso de modelado, que se puede monitorear en vivo.
- En el panel de acciones, seleccione la casilla correspondiente a la siguiente región. Abra el obturador de campo brillante para que la cuadrícula sea visible y centre esa región usando la etapa del microscopio. Repita los pasos 5.13 a 5.14 para cada región del Panel de acciones.
- Retire la cubierta con la rejilla del microscopio e inmediatamente pipete 10 μL de solución salina estéril tamponada con fosfato (PBS) en la rejilla.
- Después de 10 minutos, retire la plantilla con pinzas, luego lave la rejilla 3 veces con 15 μL de PBS. Después del lavado final, coloque cada rejilla en 15 μL de PBS y mueva las rejillas a un lugar oscuro.
6. Deposición de proteínas ECM
- Para las células cultivadas, siga los pasos 6.2-6.5; para las neuronas primarias de Drosophila , siga los pasos 6.6-6.10.
- Prepare al menos 15 μL de ECM para cada rejilla. Para las células BEAS-2B, preparar una concentración final de 0,01 mg/ml de fibronectina bovina y 0,01 mg/ml de fibrinógeno conjugado con fluoróforo en PBS estéril. Para las células HeLa, preparar 0,01 mg/ml de colágeno I bovino y 0,1 mg/ml de fibrinógeno conjugado con fluoróforo en PBS estéril.
- Retire la mayor parte del PBS de cada rejilla y aplique 15 μL de la ECM. Incubar la rejilla en una cámara húmeda a temperatura ambiente durante al menos 1 h.
NOTA: Este paso puede extenderse a toda la noche a 4 °C. - Después de la incubación en ECM, lave cada rejilla 5 veces con PBS estéril. Para cada lavado, retire la mayor parte del líquido con una pipeta sin dejar que la rejilla se seque, agregue 15 μL de PBS fresco, incube durante al menos 30 s y repita. Deje cada rejilla en PBS después del lavado final.
NOTA: Las rejillas se pueden almacenar hasta por una semana en PBS a 4 °C sin deterioro observado en la calidad. - Use un microscopio de fluorescencia para detectar el fluoróforo en el ECM para confirmar el patrón y que la lámina de carbono permaneció intacta. Unos pocos cuadrados rotos son generalmente tolerables.
- Para las neuronas primarias de Drosophila , mueva las rejillas estampadas a un plato con fondo de vidrio de 30 mm que contenga PBS estéril.
- Aspire el PBS del plato y aplique 2 ml de concanavalina A conjugada con fluoróforo de 0,5 mg/ml. Incubar durante la noche a 25 °C en un ambiente estéril.
- Retire la solución de concanavalina A del plato (sin secar las rejillas) y lave las rejillas 3x con PBS. Para cada lavado, agregue y retire 2 ml de PBS del plato.
- Use un microscopio de fluorescencia para detectar el fluoróforo en el ECM para confirmar el patrón y que la lámina de carbono permaneció intacta. Unos pocos cuadrados rotos son generalmente tolerables.
- Después del lavado final, retire el PBS de la placa con fondo de vidrio y agregue 2 ml de Drosophila media de Schneider suplementada con filtro estéril recién preparada21, que contenga un 20% de suero fetal bovino (FBS) inactivado por calor, 5 μg/ ml de insulina, 100 μg / ml de penicilina, 100 μg / ml de estreptomicina y 10 μg / ml de tetraciclina. Incubar a 25 °C en un ambiente estéril hasta que las neuronas estén listas para ser chapadas.
7. Preparación de células primarias de Drosophila antes de la siembra
- Esterilizar una placa de disección de 55 mm con 70% de EtOH, y luego agregar a la placa 2-3 mL de solución salina de disección 1× filtrada estéril (9.9 mM HEPES pH 7.5, 137 mM NaCl, 5.4 mM KCl, 0.17 mM NaH2PO4, 0.22 mM KH2PO4, 3.3 mM de glucosa, 43.8 mM de sacarosa)21.
- Elija 30-40 larvas de 3rd instar suavemente de la comida usando un par de pinzas.
- Coloque las larvas en el tubo con 1× PBS, luego transfiéralas al segundo tubo con 1× PBS para lavar las larvas.
- Transfiera las larvas al tubo con un 70% de EtOH para lavar el PBS, luego transfiéralas al segundo tubo con un 70% de EtOH. Deje las larvas en el segundo tubo durante 2-3 minutos para esterilizar las larvas.
- Transfiera las larvas a un tubo con solución salina de disección al 1×, luego transfiéralas inmediatamente al segundo tubo con solución salina de disección al 1×.
- Transfiera las larvas individuales al plato de disección que contiene solución salina de disección 1×. Con un par de fórceps y un microscopio de disección, desgarra rápidamente cada larva para extraer el cerebro y transferirlo al tercer tubo con solución salina de disección 1×. Repita hasta que se extraigan todos los cerebros.
- Centrifugar el tubo que contiene los cerebros a 300 x g durante 1 min.
- Deseche el sobrenadante, y lave con 1 ml de solución salina de disección 1× y centrifugue el tubo a 300 x g durante 1 min. Repita este paso una vez más.
- Deseche el sobrenadante hasta que queden 200-250 μL en el tubo y agregue 20 μL de 2.5 mg / ml liberasa en 1x solución salina de disección.
- Gire el tubo en un rotador durante 1 h a temperatura ambiente; durante esta hora, pipetee la solución 25-30 veces cada 10 minutos. Al final, la solución debe ser ligeramente opaca.
- Centrifugar las células a 300 × g durante 5 min.
- Deseche el sobrenadante, luego agregue 1 ml de medios suplementados de Schneider. Pipetear la solución 30 veces para mezclar.
- Centrifugar las células a 300 × g durante 5 min.
- Deseche el sobrenadante y lave el pellet celular agregando 1 ml de medios de Schneider suplementados. Pipetear la solución 30 veces para mezclar.
- Centrifugar las células a 300 × g durante 5 min.
- Deseche el sobrenadante, luego vuelva a suspender el pellet celular con 300 μL de medios de Schneider suplementados. Pipetear la solución 30-40 veces para mezclar.
8. Cultivo e infección por VRS de células BEAS-2B y HeLa
- Mantener las células HeLa y las células BEAS-2B en matraces T75 a 37 °C y al 5 % de CO2. Las células de paso cada 3-4 días una vez que alcanzan aproximadamente el 80% de confluencia. Mantener las células HeLa en DMEM + 10% FBS + 1× Antibiótico-Antimicótico. Mantener BEAS-2B en RPMI + 10% FBS + 1× Antibiótico-Antimicótico6,22,23.
- Para la siembra de células no infectadas, vaya a la sección 9. Las células BEAS-2B y HeLa son susceptibles a la infección por VRS; Las células BEAS-2B se utilizaron para todos los experimentos con VRS que se muestran aquí.
NOTA: Realice todos los pasos de BSL-2 de acuerdo con los protocolos institucionales utilizando un gabinete de bioseguridad (BSC) apropiado y equipo de protección personal (PPE). - Antes de la infección de células por VRS, pase 5 × 104 células por pozo en una placa de 6 pocillos (área de superficie ~ 9.6 cm2) con 2 ml de medios de crecimiento e incube durante la noche.
- Tripsinizar y contar un pozo de células. Para tripsinizar, aspire los medios de un pozo y lave con 2 ml de PBS estéril sin Mg2+ y Ca2+ para eliminar los medios residuales. Añadir 500 μL de solución de tripsina al 0,25%. Incubar a 37 °C durante 5-10 min. Revise periódicamente las células para ver si se liberan de la superficie. Una vez que se liberan las células, agregue 1.5 ml de medios de cultivo.
- Mezclar 100 μL de células tripsinizadas con 100 μL de azul de tripano. Pipetear 10 μL de mezcla celular diluida en un hemocitómetro. Cuente las células y calcule el número de células por pozo. Utilice este número para calcular el MOI a continuación.
- Preparar una dilución de RSV-A2mK+24 en medios de crecimiento para lograr un MOI de 10 por pocillo en 750 μL de medio. El MOI de RSV-A2mK+ se puede calcular a partir de títulos de unidades de enfoque fluorescente (FFU) del stock (por ejemplo: para 1.0 × 105 células por pozo y un stock de RSV de 1.0 × 108 FFU/mL, diluir el stock viral 1:75 a 1 × 106 FFU/750 μL o 1.33 × 106 FFU/mL).
- Aspire los medios de las células en el plato de 6 pocillos y agregue 750 μL de la solución viral desde arriba a cada pozo.
- Balancee la placa a temperatura ambiente durante 1 h.
- Después de 1 h, lleve el volumen total por pozo hasta 2 ml con los medios de crecimiento precalentados a 37 ° C y coloque la placa en una incubadora a 37 ° C con 5% de CO2 durante 6 h.
- Tripsinizar las células para liberarlas y proceder a la siembra como se describe a continuación. Después de la siembra, incube las rejillas durante 18 horas adicionales antes de la congelación por inmersión (para un total de 24 h después de la infección).
9. Siembra de células en cuadrículas con micropatrones
- Para las células cultivadas, siga los pasos 9.2-9.8; para las neuronas primarias de Drosophila , siga 9.9-9.11.
- Tripsinizar las células para liberarlas (ver paso 4 en la sección 8 anterior). Para reducir la agregación celular, tripsinizar las células al 60 % o menos de confluencia.
- Mezclar 100 μL de células tripsinizadas con 100 μL de azul de tripano. Pipetear 10 μL de la mezcla de células diluidas en un hemocitómetro y contar las células.
- Diluir las células en medios a 2 × 104 células/ml.
- Agregue 1 μL de medio al centro de un plato con fondo de vidrio de 30 mm para ayudar a colocar la rejilla y evitar que se seque. Transfiera la rejilla desde el PBS en la cubierta hasta el centro del plato con fondo de vidrio. Añadir 10 μL de solución celular a la rejilla.
- Usando un microscopio de campo brillante, observe la adhesión celular a la rejilla después de 5 minutos. Si la mayoría de los patrones permanecen desocupados, agregue una gota adicional de 10 μL de la solución celular. Mantenga las rejillas y la solución celular a 37 °C durante las incubaciones.
- Repita el paso 9.6 hasta que la mayoría de los patrones estén ocupados o muchos patrones ocupados comiencen a tener varias celdas. Incubar la rejilla durante 2 h en la incubadora (37 °C, 5% CO2).
- Inundar el plato con 2 ml de medios precalentados e incubar durante la noche (37 °C, 5% de CO2).
- Para las neuronas primarias de Drosophila , retire los medios del plato que contiene la rejilla y coloque las células en el plato.
- Espere 30-60 minutos para que las celdas se conecten, luego agregue 2 ml de medios de Schneider suplementados.
- Cultive las neuronas en una incubadora de 25 °C durante un mínimo de 2-3 días antes de la congelación por inmersión.
10. Obtención de imágenes y vitrificación de cuadrículas estampadas
- Coloque el plato con fondo de vidrio que contiene la rejilla estampada y las células cultivadas en el microscopio de fluorescencia.
- Adquiera imágenes de la cuadrícula utilizando el campo brillante y los canales fluorescentes apropiados para detectar el patrón y cualquier otro etiquetado en las celdas. Asegúrese de que la densidad y el posicionamiento de la célula sean adecuados para la obtención de imágenes y el análisis posteriores.
NOTA: Las imágenes brightfield y fluorescentes se procesaron en el paquete de software FIJI25. - Prepare un congelador crio-inmersión; el tipo de dispositivo de congelación dependerá de la disponibilidad, el costo y las características que sean más adecuadas para la muestra.
NOTA: Las neuronas primarias de Drosophila se prepararon en un congelador automatizado, y las células BEAS-2B se prepararon utilizando un congelador semiautomatizado. - Aplique fiduciales de oro a las muestras para una alineación adecuada de la serie de inclinación. Seque muestras para eliminar el exceso de medios, luego congele las muestras en un criógeno, como el etano líquido enfriado por nitrógeno líquido. Para las neuronas primarias de Drosophila , seque durante 4 s de la parte posterior. Para las células HeLa y BEAS-2B, seque desde ambos lados durante 4-6 s. Las rejillas congeladas se pueden almacenar en nitrógeno líquido hasta su uso posterior.
- Imagen de células vitrificadas en un microscopio crioelectrónico, operado a 300 kV con una cámara detectora de electrones directa. Configure la recopilación de series basculantes para cada región de interés con software como SerialEM26 para la recopilación de datos cryo-EM/cryo-ET.
NOTA: La serie de inclinación de las neuronas primarias de Drosophila se recogió en un detector de electrones directo de -60 ° a 60 ° bidireccionalmente a incrementos de 2 ° a -8 μm de desenfoque con un tamaño de píxel de 4.628 Å para una dosis total de 70-75 e-/ Å2. La serie Tilt de BEAS-2B infectado por RSV se recolectó en un detector de electrones directo con un filtro de energía (hendidura de 20 eV) a -5 μm de desenfoque con un tamaño de píxel de 4.603 Å y una dosis total de ~ 80 e-/ Å2. - Procese la serie de inclinación para reconstruir tomogramas.
NOTA: Los tomogramas presentados aquí fueron reconstruidos utilizando el paquete IMOD27; El filtrado lowpass se realizó utilizando el paquete de software EMAN228.
Resultados
Este procedimiento se utilizó para modelar rejillas EM para experimentos crio-ET de células enteras. Todo el flujo de trabajo presentado en este estudio, incluidas las preparaciones iniciales de cultivo celular, el micropatronaje (Figura 1) y las imágenes, abarca de 3 a 7 días. Se utilizó un procedimiento de dos pasos para generar la capa antiincrustante aplicando PLL a la rejilla y posteriormente vinculando PEG mediante la adición del PEG-SVA reactivo. La capa antiincrustante también se puede aplicar en un solo paso agregando PLL-g-PEG en una incubación. El gel PLPP es un catalizador para el micropatronaje UV, que también está disponible como un líquido menos concentrado. El gel permite el modelado a una dosis significativamente reducida en comparación con el líquido, lo que resulta en un patrón mucho más rápido. Con este sistema, el tiempo real de modelado de una cuadrícula TEM completa fue de ~ 2 minutos. El flujo de trabajo de micropatronaje por sí solo generalmente abarca de 5 a 6 horas y permite a un individuo modelar ocho cuadrículas para el cultivo celular estándar en cuadrículas TEM.
Varios de los pasos durante el proceso de micropatronaje requieren largos tiempos de incubación (ver pasos 2.1, 2.3, 6.4). Convenientemente, algunos de estos pasos, como la pasivación PLL (2.1) o la pasivación PEG-SVA (2.3), pueden extenderse a una incubación nocturna. Además, las rejillas pueden ser modeladas por adelantado y almacenadas en una solución de la proteína ECM o PBS para su uso posterior. En nuestro estudio, estas opciones fueron valiosas en los casos en que el momento de la preparación y siembra celular es crítico, como las neuronas primarias de Drosophila y la infección por VRS de las células BEAS-2B.
Las rejillas se prepararon en un entorno de laboratorio de nivel 2 de bioseguridad general (BSL-2) utilizando herramientas limpias, soluciones estériles e incluyeron antibióticos / antimicóticos en el medio de crecimiento6,22,29,30. Para muestras particularmente sensibles a la contaminación microbiana, la capa antiincrustante y la ECM se pueden aplicar en una campana de cultivo de tejidos u otro entorno estéril. Además, la rejilla podría lavarse en etanol entre el patrón y la aplicación de ECM. Si se trabaja con agentes infecciosos, es importante adaptar el procedimiento para cumplir con los protocolos de bioseguridad adecuados.
Este flujo de trabajo y los procedimientos presentados (Figura 1) permitieron que las células HeLa (Figura 4), las células BEAS-2B infectadas por RSV (Figura 3, Figura 5) y las neuronas larvales primarias de Drosophila (Figura 6, Figura 7) se sembraran en rejillas EM con patrones para una recopilación óptima de datos crio-ET.
Las células HeLa sembradas en rejillas TEM con micropatrones siguen siendo viables según lo determinado por la tinción fluorescente utilizando un ensayo de viabilidad celular basado en calceína-AM y homodímero de etidio-1 (Figura 4A, B). Usando una ECM mixta de colágeno y fibrinógeno, las células HeLa se adhieren fácilmente a patrones a través de la cuadrícula (Figura 4A, C). La morfología general de las células que se expanden a lo largo del patrón es similar a la de las células cultivadas en cuadrículas sin patrones (Figura 4C, D). En el caso de las células HeLa, el grosor total de la célula permanece ~ < 10 μm con áreas significativamente más delgadas ~ < 1 μm de espesor cerca de la periferia celular (Figura 4E, F).
Para los estudios de VSR, modelamos cuadrados de cuadrícula completos utilizando un gradiente, con una exposición a dosis bajas en los bordes y un patrón de dosis más alto hacia el centro (Figura 3A). Los patrones de gradiente produjeron mejores resultados al buscar virus liberados presentes cerca de la periferia de las células. Con estos patrones, se encontró que las células se adhieren preferentemente a la concentración más alta de ECM, pero también son capaces de adherirse y crecer en las concentraciones más bajas de ECM. La dosis relativa entre áreas deberá optimizarse cuando se utilicen patrones que requieran dosis múltiples. Si las dosis y, por lo tanto, las concentraciones de ECM son demasiado similares o demasiado dispares entre sí, se perderá el efecto de usar dosis múltiples.
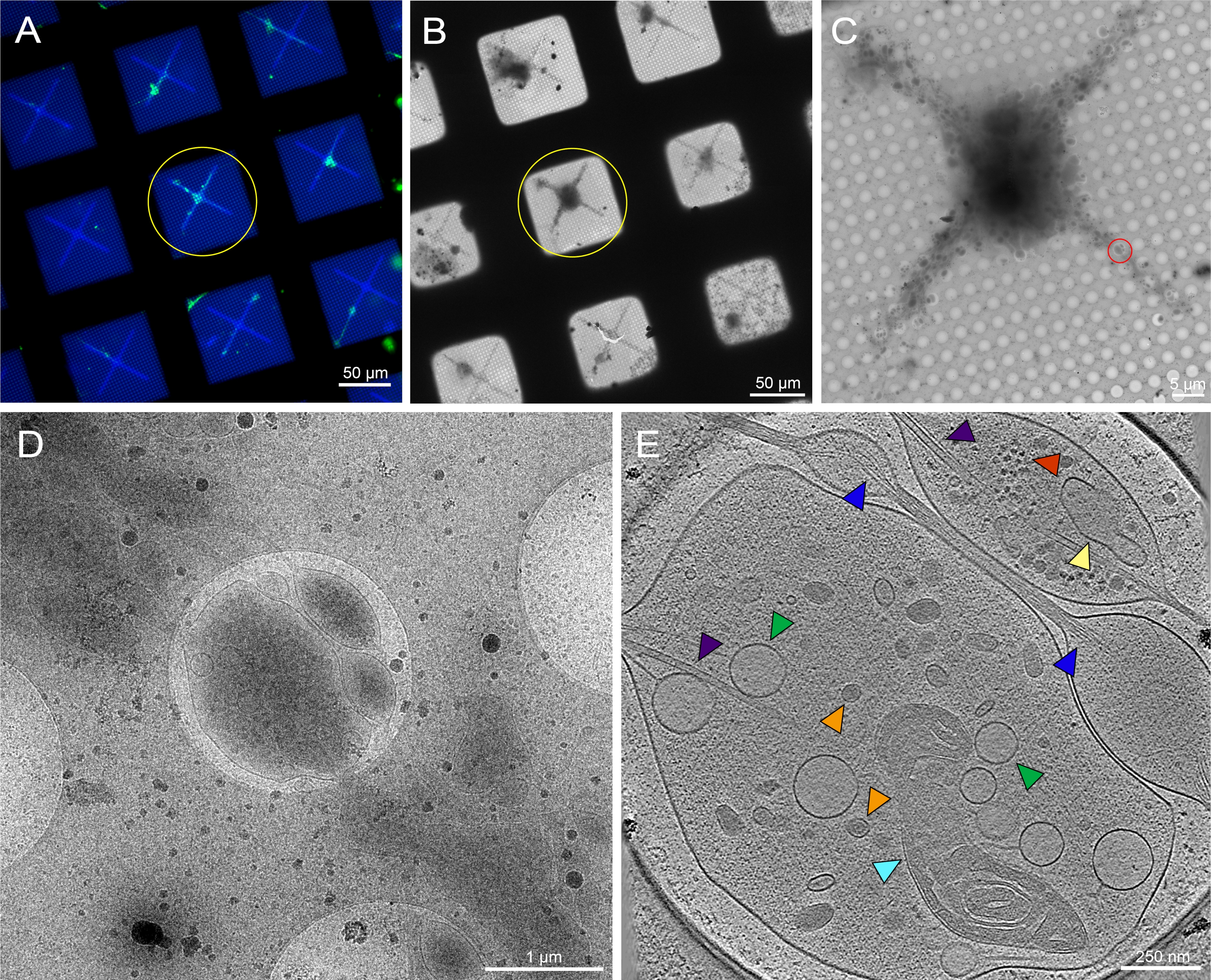
En la Figura 3, se modeló una cuadrícula TEM y posteriormente se sembró con células BEAS-2B infectadas por RSV y se utilizó para la recopilación de datos crio-EM. La Figura 4A es una imagen fluorescente de ECM estampada en una cuadrícula TEM utilizando un patrón de gradiente. La adhesión y el crecimiento celular a lo largo de la región central del patrón se pueden ver en la Figura 3B como una imagen de campo brillante de las células 18 horas después de la siembra. En la Figura 3C, la señal fluorescente (roja) de la replicación del RSV-A2mK+ se superpone con la señal del ECM. La mayoría de las células infectadas se colocan a lo largo de la región central de mayor densidad del patrón de gradiente. Un mapa TEM de bajo mag de la criofijación posterior a la cuadrícula revela una serie de células, incluidas las células infectadas por RSV, colocadas en la lámina de carbono cerca del centro de los cuadrados de la cuadrícula. Como se mostró anteriormente para las células cultivadas en cuadrículas TEM estándar22, las series inclinables se localizaron y recolectaron de viriones RSV muy cerca de la periferia de las células BEAS-2B infectadas cultivadas en rejillas micromodeladas (Figura 5A, B). Muchas de las proteínas estructurales del VSR se pueden identificar dentro de los tomogramas, incluyendo la nucleocápside (N) y la proteína de fusión viral (F) (Figura 5C, flechas azules y rojas respectivamente).
Para los estudios primarios de neuronas de Drosophila, se encontró que el patrón estrecho, cerca del límite de resolución ofrecido por el software (donde el grosor del patrón era de 2 μm), permitía aislar de una a unas pocas células dentro de un cuadrado de cuadrícula (Figura 6). El soma neuronal fue capaz de extender sus neuritas durante un período de varios días dentro del patrón. Esto permitió una fácil identificación y adquisición de series inclinadas de las neuritas en comparación con las neuronas cultivadas en cuadrículas sin patrones (Figura 7). También se encontró que la concanavalina A marcada fluorescentemente, una lectina que se ha utilizado como ECM para cultivos neuronales in vitro de Drosophila20,21, es susceptible de patronaje.
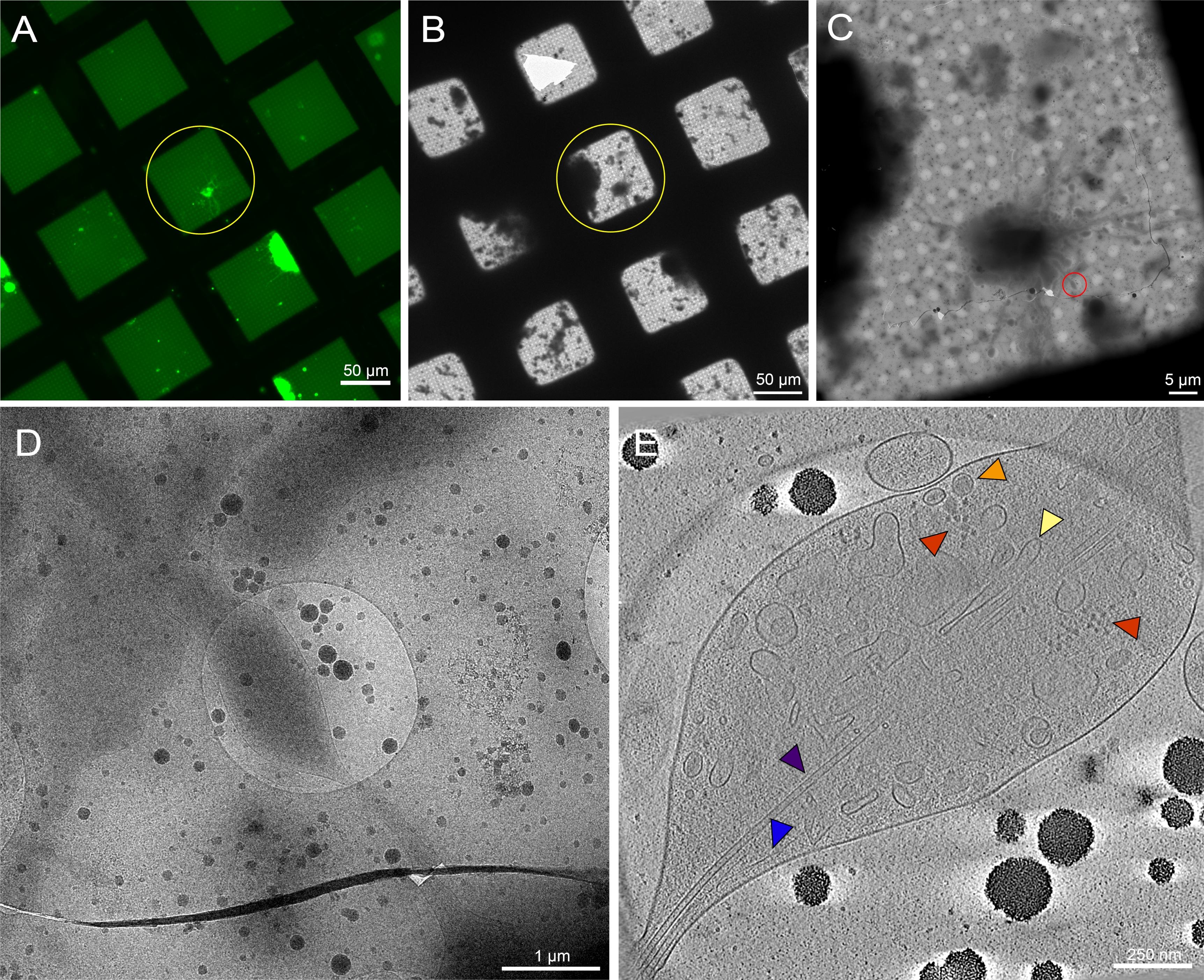
Las neuronas de Drosophila de tercera larvas de instar fueron aisladas según protocolos previamente publicados20,21,31. Las preparaciones neuronales se aplicaron a rejillas crio-EM con micropatrones donde la concanavalina A se depositó en el patrón para regular la colocación, propagación y organización celular. Se permitió que las neuronas en rejillas modeladas o sin patrones se incubaran durante un mínimo de 48-72 horas, y las rejillas se congelaron. En la Figura 6A se muestra una imagen representativa de una cuadrícula EM micromodelada con varias neuronas de Drosophila distribuidas en las regiones modeladas. Estas neuronas, derivadas de una cepa de mosca transgénica que tiene expresión pan-neuronal de GFP en la membrana, pueden ser fácilmente rastreadas por microscopía de luz no solo debido a su marcado fluorescente, sino también debido a su ubicación dentro de los micropatrones. Si bien las neuronas cultivadas en rejillas sin patrón también se pueden rastrear a través de su señalización GFP mediante microscopía de luz (Figura 7A, círculo amarillo), ubicarlas en crio-EM se volvió sustancialmente más difícil debido a la presencia de desechos celulares y la contaminación de los medios (Figura 7B, círculo amarillo). Tal presencia se redujo para las neuronas en rejillas modeladas, probablemente debido al PEG en la capa antiincrustante de las regiones no modeladas que repelen los desechos celulares de la adhesión. Debido a las dimensiones del cuerpo celular de la neurona y las neuritas extendidas (Figura 6A, B, círculo amarillo), las series de inclinación crio-ET se recolectaron a lo largo de regiones más delgadas de las células (Figura 6C, D, círculo rojo). La membrana celular neuronal, una mitocondria (cian), microtúbulos (púrpura), filamentos de actina (azul), estructuras vesiculares (naranja y verde) y macromoléculas como ribosomas (rojo) se resolvieron bien en montajes de imágenes de mayor aumento y rodajas a través del tomograma 3D (Figura 6E). Si bien se pueden ver características subcelulares similares en los tomogramas 3D de neuronas sin patrones (Figura 7E), la dificultad para localizar objetivos celulares viables para la recopilación de datos disminuyó sustancialmente el rendimiento.
En la Figura 8, se han ensamblado imágenes representativas de cuadrículas con algunos de estos problemas para ayudar en su identificación y solución de problemas. Una vez que se determinan las condiciones óptimas, el micropatronaje es un método confiable y reproducible para el posicionamiento de células en rejillas para crio-TEM.

Figura 1: Flujo de trabajo general de micropatronaje para crio-EM. El flujo de trabajo se puede dividir aproximadamente en cuatro partes: preparación de la red, micromodelado, ECM y siembra celular, y criopreparación y recopilación de datos. Los pasos principales de cada sección se enumeran debajo de los encabezados y el tiempo aproximado para completar cada sección se muestra a la izquierda. Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}

Figura 2: Captura de pantalla del software con patrón colocado en cuadrícula. El área 1 contiene la relación μm/pix para el diseño de patrones. El área 2 es la regla para medir una cuadrícula. El Área 3 es donde agregar o cambiar patrones y ROI. El Área 4 contiene toda la información para el posicionamiento del patrón y la dosis. El área 5 contiene opciones para patrones, incluida la alternancia de superposiciones, la copia o eliminación de patrones y la selección de patrones para el micropatronaje. El Área 6 es donde se pueden guardar y cargar las plantillas. A continuación se muestran vistas más amplias de las áreas 4 y 5 para mayor claridad. Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}

Figura 3: Células BEAS-2B infectadas por RSV en la cuadrícula crio-TEM con patrón. (A) Imagen fluorescente de la cuadrícula con patrón después de la adición de ECM marcada fluorescentemente. El patrón de entrada se muestra en la esquina inferior izquierda. (B) Imagen de campo brillante de células BEAS-2B cultivadas en la cuadrícula en A. (C) Fusión de la imagen en A (cian) y B (gris) con imagen fluorescente de células infectadas por RSV (rojo) inmediatamente antes de la congelación por inmersión; las células infectadas expresan mKate-2. Las barras de escala son de 500 μm. (D) Mapa crio-TEM de bajo aumento de la cuadrícula en B después de la congelación por inmersión. Las imágenes fluorescentes son pseudocolores. Las barras de escala son de 500 μm. Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}

Figura 4: Tinción viva/muerta de células estampadas y sin patrones. (A) Imagen fluorescente de células HeLa cultivadas en una rejilla estampada y teñidas con calceína-AM (tinción de células vivas, verde) y homodímero de etidio-1 (tinción de células muertas, rojo). (B) Células HeLa cultivadas en una rejilla sin patrón y teñidas como en A. (C) Proyección de pilas z confocales de una célula HeLa en una rejilla Quantifoil R2/2 estampada con 0,01 mg/ml de colágeno y fibrinógeno 647 ECM (rojo). La célula fue teñida con calceína-AM (verde) y Hoechst-33342 (azul). (D) Células HeLa en rejilla sin patrón incubadas con 0,01 mg/ml de colágeno y fibrinógeno 647 ECM, incubadas y teñidas con calceína-AM y Hoecsht-33342. Las imágenes fluorescentes se fusionaron con la luz transmitida (escala de grises). (E) Proyección X,Z de C. (F) Proyección X,Z de D. Las imágenes son pseudocoloreadas. Las barras de escala en (A) y (B) son de 500 μm; las barras de escala en (C) - (F) son de 10 μm. Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}

Figura 5: Cryo-ET de la célula BEAS-2B infectada por el VSR en la cuadrícula crio-TEM con patrón. (A) Mapa cuadrado de la cuadrícula Cryo-EM de la célula BEAS-2B infectada por el VSR. El límite aproximado de la celda se indica mediante la línea verde discontinua. (B) Imagen de mayor resolución del área encajonada en rojo en (A). El límite aproximado de la celda se indica mediante una línea verde discontinua. Los viriones del VSR se pueden ver cerca de la periferia celular (flecha blanca y caja amarilla). (C) Rebanada z simple del tomograma recogido en el área de la caja amarilla en (B). Las flechas rojas apuntan a la proteína de fusión RSV F, las flechas azules apuntan al complejo ribonucleoproteína (RNP). Las barras de escala en (A)-(C) están incrustadas en la imagen. Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}

Figura 6: Neuronas primarias derivadas de los cerebros de larvas de 3rd instar Drosophila melanogaster en la cuadrícula crio-TEM con patrón. (A) Montaje de rejilla de microscopía de fluorescencia de células vivas superpuestas de neuronas de Drosophila que expresan GFP dirigida a la membrana en cuadrados de cuadrícula modelados con concanavalina fluorescente A de 0,5 mg / ml. Verde: Neuronas de Drosophila . Azul: Fotopatrones. (B) Montaje de imagen crio-EM de la cuadrícula en (A) después de la criopreservación. El círculo amarillo anota el mismo cuadrado de cuadrícula que en (A). (C) Montaje de imagen crio-EM del cuadrado resaltado por el círculo amarillo en (A) y (B). (D) Imagen de mayor aumento del área delimitada por el círculo rojo en (C), donde se recogió una serie de inclinación en las neuritas de la célula. E. Rebanada de 25 nm de espesor de un tomograma reconstruido a partir de la serie de inclinación que se adquirió del círculo rojo en (C). Varios orgánulos se pueden ver en este tomograma, como las mitocondrias (cian), los microtúbulos (púrpura), las vesículas centrales densas (naranja), las vesículas claras (verde), el retículo endoplásmico (amarillo) y la actina (azul). Las macromoléculas, como los ribosomas (rojos), también se pueden ver en la esquina superior derecha. Las imágenes fluorescentes son pseudocolores. Las barras de escala en (A)-(E) están incrustadas en la imagen. Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}

Figura 7: Neuronas primarias derivadas de los cerebros de larvas de 3rd instar Drosophila melanogaster en rejillas sin patrón. (A) Montaje en rejilla de microscopía de fluorescencia de células vivas de neuronas de Drosophila que expresan GFP dirigida a membrana en cuadrados de cuadrícula con 0,5 mg/ ml de concanavalina A. Verde: Neuronas de Drosophila . (B) Montaje de rejilla Cryo-EM de la misma rejilla en (A) después de la congelación por inmersión. El círculo amarillo muestra el mismo cuadrado de cuadrícula que en (A). Tenga en cuenta la presencia de desechos celulares y contaminación de los medios, lo que dificultó la identificación del objetivo en comparación con las rejillas con patrones. (C) Montaje de imagen crio-EM del cuadrado resaltado por los círculos amarillos en los mapas (A) y (B). (D) Imagen de mayor aumento del área delimitada por el círculo rojo en (C), donde se recogió una serie de inclinación en las neuritas de la célula. (E) Rebanada de 25 nm de espesor del tomograma reconstruido a partir de la serie de inclinación de (C) y (D). Varios orgánulos son visibles en este tomograma, como microtúbulos (púrpura), actina (azul), el retículo endoplásmico (amarillo) y vesículas de núcleo denso (naranja). También se pueden ver macromoléculas, como los ribosomas (rojos). Las imágenes fluorescentes son pseudocolores. Las barras de escala en (A)-(E) están incrustadas en la imagen. Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}

Figura 8: Ejemplos de posibles problemas con el modelado. Imágenes fluorescentes de ECM etiquetado depositadas en rejillas con micropatrones. (A) Patrón desigual a través de la red debido a la distribución desigual del gel PLPP. (B) La ECM no puede adherirse a las áreas cubiertas por la plantilla PDMS durante el modelado. (C) Patrón de gradiente saturado (lado derecho) o patrón invertido (izquierda) en una cuadrícula con patrón con una dosis total demasiado alta. (D) ECM se adhiere a áreas en las barras de la rejilla, así como al área modelada debido a los reflejos del láser UV durante el modelado. Las imágenes son pseudocolores; el patrón de entrada se muestra en la parte inferior izquierda; las barras de escala son de 100 μm. Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}
| Emitir | Causa(s) potencial(es) | Solución de problemas |
| Micropatronaje | ||
| No se puede ver la iluminación del láser PRIMO | • La trayectoria de la luz no está configurada correctamente | • Compruebe que la trayectoria de la luz del microscopio esté configurada correctamente |
| • El láser PRIMO no está encendido o el láser está entrelazado | ||
| Muchos cuadrados de cuadrícula rotos | • Tocar la lámina de rejilla con pinzas o pipetas durante la manipulación | • Manejar las rejillas con cuidado |
| • Rejilla seca durante las incubaciones o el lavado | • No permita que la rejilla se seque durante los lavados e incubaciones | |
| Grandes áreas sin patrones | • Cobertura insuficiente del gel | • Asegúrese de que el gel se extienda uniformemente sobre la rejilla mientras agrega |
| • Lámina de rejilla fuera de foco durante el modelado | • Añadir un microlitro adicional de gel | |
| •Área cubierta por plantilla | • Compruebe el enfoque antes de modelar cada región | |
| • Centrar cuidadosamente la cuadrícula en la plantilla | ||
| Patrón saturado o invertido | • Dosis incorrecta | • Pruebe un rango de dosis totales para el patrón |
| • Cobertura insuficiente del gel | • Asegúrese de que la rejilla esté cubierta uniformemente con gel | |
| • Pruebe diferentes valores para patrones de escala de grises | ||
| Patrón borroso | • Enfoque deficiente durante el modelado | • Repita la calibración primo a la misma altura que la muestra |
| • Calibración incorrecta | • Concéntrese en la lámina de rejilla antes de modelar | |
| • Dividir el patrón en regiones adicionales para el modelado | ||
| ECM que se adhiere fuera del patrón | • Reflejos de gel o polvo | • Asegúrese de que el gel esté seco antes de modelar |
| • Asegúrese de que la cubierta y la lente del objetivo estén limpias | ||
| ECM no visible después del modelado | • Blanqueamiento fotográfico | • Minimizar la exposición a la luz a la ECM antes de la toma de imágenes |
| • Dosis incorrecta durante el modelado | • Pruebe un rango de valores de dosis total para el patrón | |
| • Tiempo de incubación de ECM insuficiente | • Aumentar el tiempo de incubación para ECM | |
| Siembra celular | ||
| Aglomeración de células | • Sobre digestión | • Use un porcentaje más bajo de tripsina o tiempo para la liberación de células adherentes |
| • Alta densidad celular | • Pasar y/o digerir las células a menor confluencia | |
| • No agitar las células durante la liberación | ||
| • Pipetee suavemente la solución celular o use coladores celulares | ||
| Células que no se adhieren a áreas modeladas | • ECM no es adecuado para el tipo de célula | • Pruebe diferentes concentraciones y composición de ECM |
| • La viabilidad de las células disminuye antes de la siembra | • Asegurar que el cultivo celular y las condiciones de liberación celular no dañen las células | |
| Células que no se expanden después de la adhesión | • ECM o patrón no adecuado para el tipo de célula | • Pruebe diferentes patrones y ECM |
| • En algunos casos, una lámina más continua (R1.2/20 vs R2/1) puede promover la expansión celular |
Tabla 1: Posibles problemas durante el micropatronaje. En esta tabla se describen algunos problemas que puede experimentar un usuario durante el micropatronaje o la siembra de celdas. Se proporcionan las posibles causas y la solución de problemas para cada problema. Las imágenes representativas de algunos problemas se pueden ver en la Figura 8.
Discusión
Los microscopios electrónicos modernos y avanzados y los paquetes de software ahora admiten la recopilación automatizada optimizada de datos crio-EM y crio-ET donde se pueden apuntar y obtener imágenes de cientos a miles de posiciones en unos pocos días32,33,34,35. Un factor limitante significativo para los flujos de trabajo crio-ET de celdas enteras ha sido la obtención de un número suficiente de objetivos coleccionables por cuadrícula. Recientemente, varios grupos han desarrollado protocolos para redes de micropatrones para crio-EM, con una ventaja en la mejora de la eficiencia de la recopilación de datos16,17,18. Aquí se presenta un protocolo para el uso de un sistema de micropatronaje disponible comercialmente para micromodelar cuadrículas TEM para estudios crio-ET de neuronas primarias de Drosophila y líneas celulares humanas cultivadas (no infectadas o infectadas por RSV). Este sistema de micropatrones es versátil y muchos pasos se pueden optimizar y adaptar para adaptarse a objetivos experimentales específicos. Un usuario con experiencia en TEM y microscopía de fluorescencia puede convertirse rápidamente en experto en preparación de rejillas y micropatrones. Con una práctica cuidadosa, los buenos resultados deben ser alcanzables después de unas pocas iteraciones. A continuación, se discuten algunas de las opciones disponibles, las consideraciones del usuario, los beneficios potenciales y las aplicaciones futuras del micropatronaje para crio-EM.
Una de las consideraciones importantes para el crio-ET de células enteras es la selección de la rejilla EM. Las rejillas EM se componen de dos partes: un marco de malla (o soporte estructural) y la lámina (o película), que es la superficie de película continua o agujereada en la que crecerán las células. Las rejillas de malla de cobre se utilizan comúnmente para crio-EM de proteínas y complejos aislados. Sin embargo, no son adecuados para la crio-ET de células enteras debido a la citotoxicidad del cobre. En cambio, una malla de oro se usa comúnmente para la tomografía celular. Otras opciones incluyen níquel o titanio, que pueden proporcionar beneficios sobre el oro, como una mayor rigidez16. Las rejillas EM están disponibles con diferentes dimensiones de malla para admitir una amplia gama de aplicaciones. Los tamaños de malla más grandes proporcionan más espacio para que las células crezcan entre las barras de la rejilla y más áreas que son susceptibles de recolección de series inclinables, aunque a costa de una mayor fragilidad general de la muestra. La lámina más utilizada es el carbono amorfo perforado o agujereado, como Quantifoils o rejillas C-planas. Los objetivos biológicos se pueden obtener imágenes a través de los agujeros en el carbono o a través del carbono translúcido de electrones. Las rejillas como R 2/1 o R 2/2, donde los orificios tienen 2 μm de ancho y están espaciados a 1 y 2 μm respectivamente, proporcionan una gran cantidad de agujeros y, por lo tanto, un gran número de áreas potenciales para la recopilación de datos. Sin embargo, algunas células pueden crecer y expandirse mejor en superficies más uniformes, como rejillas R 1.2/20 o carbono continuo. Para el procesamiento de muestras aguas abajo mediante fresado de haz de iones enfocados (crio-FIB), la lámina se elimina a través del fresado, lo que reduce las preocupaciones sobre la presencia continua de la película subyacente. Al igual que con la malla, también están disponibles láminas de otros materiales, y el protocolo de modelado presentado aquí es igualmente adecuado para las rejillas de SiO2 . Las rejillas de uso común incluyen Quantifoil de oro, carbono continuo o rejillas de 200 mallas de película de SiO2 (~ 90 μm de espaciado entre las barras de las rejillas) para crio-ET de celda entera.
Hay una serie de consideraciones al diseñar un patrón. La mayoría de estas decisiones están guiadas por el tipo de célula y el propósito del experimento. Un buen punto de partida es elegir un patrón que se aproxime a la forma y dimensiones de las células en cultivo. Muchos estudios han demostrado efectos significativos de la forma del patrón sobre el crecimiento celular y la disposición citoesquelética13,36,37. Se debe tener especial cuidado durante el diseño del patrón si esto podría alterar el objetivo de interés. Se probaron varios patrones para cada tipo de célula para determinar qué patrones promovían la adhesión y el crecimiento celular. La flexibilidad del sistema de micropatrones permite probar múltiples patrones en una sola cuadrícula y cambiar los patrones para diferentes cuadrículas dentro de un solo experimento. Los patrones más grandes (~ 50-90 μm), como los utilizados aquí, aumentan la probabilidad de que múltiples células se adhieran a una sola región del patrón y permitan que las células se expandan y se extiendan después de la adhesión. Patrones más restringidos (20-30 μm) pueden ser apropiados en experimentos donde el aislamiento celular es más crítico que la expansión celular, como para los experimentos de fresado de haz de iones enfocados (crio-FIB). Para aplicaciones de tomografía, es posible que deba considerar el impacto del eje de inclinación. Si se coloca un patrón de tal manera que todas las células crezcan paralelas entre sí en una sola dirección, es posible que todas las células sean perpendiculares al eje de inclinación cuando se carguen en la etapa del microscopio, lo que resulta en una menor calidad de los datos.
En las cuadrículas sin patrones, las celdas a menudo se adhieren preferentemente a las barras de la cuadrícula, donde TEM no puede obtener imágenes. Incluso en las rejillas con patrones, a menudo se observa que las celdas se colocan en las esquinas de los cuadrados de la cuadrícula parcialmente tanto en la lámina de carbono modelada como en la barra de la rejilla. Recientemente, se utilizó el micropatronaje para colocar intencionalmente parte de la celda sobre la barra de cuadrícula18. Esto podría considerarse para experimentos en los que no es crítico tener toda la periferia celular en la lámina. Esto puede ser especialmente importante para las células que pueden crecer más grandes que un solo cuadrado de cuadrícula, como las neuronas primarias que crecen durante varios días.
Hay muchas herramientas que se pueden utilizar para diseñar un patrón. Aquí, el patrón se limitó a menos de 800 píxeles en cualquier dimensión, de modo que el patrón se puede girar a cualquier ángulo y aún así encajar dentro del área máxima que puede ser modelada en una sola proyección por este sistema de micropatronaje. Esto permite al usuario girar el patrón para orientarlo correctamente con la rejilla independientemente de la orientación de la rejilla en el microscopio. Aquí, la cuadrícula se dividió en seis áreas de patrones. Principalmente, esto permite el ajuste del enfoque entre diferentes regiones de la cuadrícula. Las rejillas doradas, en particular, son muy maleables y no pueden colocarse completamente planas sobre el vidrio. El enfoque adecuado es esencial para obtener resultados de patrones limpios y refinados. Mediante el uso de patrones segmentados, solo se deben realizar ajustes menores en la posición del patrón si la cuadrícula cambia ligeramente durante el proceso de modelado, aunque esto generalmente no es un problema cuando se usa el gel PLPP con las plantillas PDMS. Finalmente, los cuatro cuadrados centrales de la cuadrícula permanecieron sin patrones. Esto permite que un usuario pueda identificar claramente el centro de la cuadrícula, lo que es muy útil para experimentos de imágenes correlativas.
El software de modelado para este sistema de micropatrones, Leonardo, también tiene características más avanzadas como costuras y la capacidad de importar patrones como PDF, que están más allá del alcance de este protocolo. Este software también incluye detección de microestructuras y posicionamiento automatizado de patrones que se pueden utilizar en cuadrículas TEM. Esta característica es más útil cuando la cuadrícula es muy plana y se puede modelar sin la necesidad de ajustar el enfoque entre diferentes áreas.
La selección de una proteína ECM puede tener un impacto significativo en la adhesión y expansión celular. Se sabe que algunas células experimentan cambios fisiológicos cuando se cultivan en sustratos específicos38. Se probaron múltiples proteínas y concentraciones de ECM para cualquier tipo de célula nueva basada en trabajos previos reportados en la literatura. La laminina, el fibrinógeno, la fibronectina y el colágeno se usan ampliamente para las células cultivadas y se pueden usar como punto de partida si no se dispone de otros datos. Sin embargo, otras proteínas ECM también deben considerarse si las proteínas ECM comúnmente utilizadas no confieren propiedades de adherencia adecuadas para las células. Esto fue particularmente cierto para las neuronas primarias de Drosophila , ya que una alta concentración de la planta lectina concanavalina A era necesaria para la adherencia celular adecuada. La compatibilidad de la adhesión celular y el crecimiento con el ECM se puede probar mediante patrones en platos de vidrio o diapositivas antes de la transición a las rejillas TEM. Este enfoque de preselección es efectivo en tiempo y costo si se necesita examinar un gran número de combinaciones. La inclusión de una proteína ECM conjugada fluorescentemente es valiosa para evaluar el éxito y la calidad del modelado.
La siembra celular es uno de los pasos más importantes para la crio-ET de células enteras, ya sea con o sin micropatrones6,16,39. Para la Drosophila primaria u otras neuronas, que son frágiles, inestables en suspensión y pueden ser limitadas en cantidad, se prefieren los enfoques de siembra única sobre la siembra celular secuencial monitoreada. Un solo paso de siembra en una densidad celular optimizada, como se describe en el protocolo para las neuronas de Drosophila, es una opción viable para la mayoría de los tipos de células. Sin embargo, también es posible sembrar células en el sustrato a una concentración inicial más baja y agregar más células de manera monitoreada como se describe aquí y en otra literatura18. Esta siembra secuencial puede proporcionar resultados más consistentes en algunos casos. Al igual que en el cultivo celular estándar, siempre se debe tener cuidado para mantener la viabilidad celular y minimizar la aglomeración celular durante el aislamiento.
Al comenzar por primera vez con el micropatronaje, hay algunas trampas potenciales que son perjudiciales para el resultado final. El manejo cuidadoso de la rejilla y la técnica estéril, una distribución uniforme del gel PLPP, la dosis y el enfoque adecuados durante el patrón, y el mantenimiento de la viabilidad celular antes de la siembra se encuentran entre las consideraciones más importantes para el éxito. En la Tabla 1 se reunió una lista de algunos de los posibles problemas, así como de las soluciones.
Las rejillas con micropatrones se pueden utilizar para ayudar a posicionar las celdas a establecer una densidad celular consistente en toda la cuadrícula y para posicionar regiones de interés en áreas adecuadas para la recolección de series inclinadas16,18. La colocación y el posicionamiento de las células se pueden utilizar como marcadores fiduciarios para la correlación en experimentos crio-CLEM, reduciendo la necesidad de rejillas de búsqueda frágiles y marcadores fiduciarios fluorescentes. Sin embargo, cabe señalar que tales marcadores fiduciarios aún pueden ser útiles para la correlación de precisión submicrométrica29,40. Además, una distribución uniforme de células aisladas también es altamente beneficiosa para los experimentos de fresado de haz de iones enfocados (crio-FIB) para maximizar el número de células de las que se pueden cortar las láminas16.
La adición de micropatrones a los flujos de trabajo de crio-EM dará como resultado mejoras medibles en el rendimiento de datos y potencialmente permitirá nuevos experimentos. A medida que la técnica se adopte y desarrolle aún más, las aplicaciones más avanzadas de micropatronaje, incluidos los gradientes de ECM, las múltiples deposiciones de ECM y el ensamblaje de microestructuras, ampliarán aún más las capacidades de crio-ET para estudiar objetivos y procesos biológicos en un contexto celular completo.
Divulgaciones
Los autores no tienen nada que revelar.
Agradecimientos
Agradecemos a la Dra. Jill Wildonger, al Dr. Sihui Z. Yang y a la Sra. Josephine W. Mitchell en el Departamento de Bioquímica de la Universidad de Wisconsin, Madison por compartir generosamente la cepa de mosca elav-Gal4, UAS-CD8::GFP (Bloomington stock center, #5146). También nos gustaría agradecer al Dr. Aurélien Duboin, al Sr. Laurent Siquier y a la Sra. Marie-Charlotte Manus de Alvéole y al Sr. Serge Kaddoura de Nanoscale Labs por su generoso apoyo durante este proyecto. Este trabajo fue apoyado en parte por la Universidad de Wisconsin, Madison, el Departamento de Bioquímica de la Universidad de Wisconsin, Madison, y las subvenciones del servicio de salud pública R01 GM114561, R01 GM104540, R01 GM104540-03W1 y U24 GM139168 a E.R.W. y R01 AI150475 a P.W.S. de los NIH. Una parte de esta investigación fue apoyada por la subvención U24 GM129547 de los NIH y realizada en el PNCC en OHSU y se accedió a través de EMSL (grid.436923.9), una Instalación de Usuario de la Oficina de Ciencia del DOE patrocinada por la Oficina de Investigación Biológica y Ambiental. También estamos agradecidos por el uso de las instalaciones y la instrumentación en el Centro de Investigación Cryo-EM en el Departamento de Bioquímica de la Universidad de Wisconsin, Madison.
Materiales
| Name | Company | Catalog Number | Comments |
| 0.1% (w/v) Poly-L-Lysine | Sigma | P8920-100ML | |
| 0.22 µm syringe filters PVDF membrane | Genesee | 25-240 | |
| 22x60-1 Glass cover slip | Fisher | 12545F | |
| 5/15 Tweezers | EMS (Dumont) | 0203-5/15-PO | |
| Antibiotic-Antimycotic (100X) | ThermoFisher (Gibco) | 15240096 | |
| BEAS-2B cells | ATCC | CRL-9609 | |
| Collagen I, bovine | ThermoFisher (Gibco) | A1064401 | |
| Concanavalin A, Alexa Fluor 350 Conjugate | ThermoFisher (Invitrogen) | C11254 | |
| DMEM | Fisher (Lonza) | BW12-604F | |
| EtOH | Fisher (Decon Labs) | 22-032-600 | |
| Fetal Bovine Serum | ATCC | 30-2020 | |
| Fibrinogen From Human Plasma, Alexa Fluor 647 Conjugate | ThermoFisher (Invitrogen) | F35200 | |
| Fibronectin Bovine Protein, Plasma | ThermoFisher (Gibco) | 33010018 | |
| Glass bottom dish | MatTek | P35G-1.5-20-C | |
| Glucose | VWR | 0643-1KG | |
| Grid prep holder | EMS | 71175-01 | |
| HeLa cells | ATCC | CCL-2 | |
| Hemacytometer | Fisher (SKC, Inc.) | 22600100 | |
| HEPES | Fisher (ACROS Organics) | AC172572500 | |
| Hoechst 33342 | ThermoFisher (Invitrogen) | H3570 | |
| Insulin | Fisher (Sigma Aldrich) | NC0520015 | |
| KCl | MP Bio | 194844 | |
| KH2PO4 | Fisher (ACROS Organics) | AC212595000 | |
| Leica-DMi8 | Leica Microsystems | Can be customized with camera, stage, and objective attachments | |
| Leonardo | Alvéole | https://www.alveolelab.com/our-products/leonardo-photopatterning-software/ | |
| Liberase Research Grade | Fisher (Supply Solutions) | 50-100-3280 | |
| LIVE/DEAD Viability/Cytotoxicity Kit | ThermoFisher (Invitrogen) | L3224 | |
| Microscope camera | Hammamatsu | C13440-20CU | |
| Motorized stage | Märzhäuser Wetzlar | 00-24-599-0000 | |
| NaCl | Fisher (Fisher BioReagents) | BP358-1 | |
| NaH2PO4 | Fisher (ACROS Organics) | AC207802500 | |
| NaOH | Fisher (Alfa Aesar) | AAA1603736 | |
| PBS | Corning | 21-040-CV | |
| PDMS stencils | nanoscaleLABS | PDMS_STENCILS_EM | https://www.alveolelab.com/our-products/pdms-stencil-multiwell-plate/ |
| PEG-SVA | nanoscaleLABS | PEG-SVA-1GR | mPEG-Succinimidyl Valerate, MW 5,000 |
| Penicillin | Fisher (Research Products International Corp) | 50-213-641 | |
| pH strips | Fisher (Millipore Sigma) | M1095350001 | pH probe can also be used |
| PLPP gel | nanoscaleLABS | PLPP-GEL-300UL | https://www.alveolelab.com/our-products/plpp-photoactivatable-reagent/ |
| PRIMO | Alvéole | https://www.alveolelab.com/our-products/primo-micropatterning/ | |
| pSynkRSV-I19F (BAC containing RSV A2-mK+ antigenomic cDNA ) | BEI Resources | NR-36460 | https://www.beiresources.org/Catalog/BEIPlasmidVectors/NR-36460.aspx |
| Quantifoil grids | EMS (Quantifoil) | Q2100AR1 | 2 µm holes spaced 1 µm apart, other dimensions are available |
| RPMI | Fisher (Lonza) | BW12-702F | |
| RSV A2-mK+ | see entry for pSynkRSV-19F | - | Described in Hotard et al. [22]. Can be generated from pSynkRSV-ll9F |
| Schneider's Media | ThermoFisher (Gibco) | 21720-024 | |
| SerialEM | SerialEM (https://bio3d.colorado.edu/SerialEM/ ) | https://bio3d.colorado.edu/SerialEM/ | |
| Straight tweezers | EMS (Dumont) | 72812-D | |
| Streptomycin | Fisher (Fisher BioReagents) | BP910-50 | |
| Sucrose | Avantor | 4097-04 | |
| Tetracycline | Sigma | T8032-10MG | |
| Titan Krios electron microscope | ThermoFisher | 300kV, with direct electron detector camera and energy filter | |
| Trypsin | ThermoFisher (Gibco) | 15090046 | |
| Tube Revolver/Rotator | Fisher (Thermo Scientific) | 11676341 | |
| UAS:mcD8:GFP Drosophila fly strain | Bloomington Drosophila Stock Center | 5146 | http://flybase.org/reports/FBtp0002652.html |
Referencias
- Nogales, E., Scheres, S. H. Cryo-EM: A unique tool for the visualization of macromolecular complexity. Molecular Cell. 58 (4), 677-689 (2015).
- Martynowycz, M. W., Gonen, T. From electron crystallography of 2D crystals to MicroED of 3D crystals. Current Opinion in Colloid and Interface Science. 34, 9-16 (2018).
- Wagner, J., Schaffer, M., Fernandez-Busnadiego, R. Cryo-electron tomography-the cell biology that came in from the cold. FEBS Letters. 591 (17), 2520-2533 (2017).
- Wan, W., Briggs, J. A. Cryo-electron tomography and subtomogram averaging. Methods in Enzymology. 579, 329-367 (2016).
- Bäuerlein, F. J., Pastor-Pareja, J. C., Fernández-Busnadiego, R. Cryo-electron tomography of native Drosophila tissues vitrified by plunge freezing. bioRxiv. , 437159(2021).
- Hampton, C. M., et al. Correlated fluorescence microscopy and cryo-electron tomography of virus-infected or transfected mammalian cells. Nature Protocols. 12 (1), 150-167 (2017).
- Hsieh, C. E., Leith, A., Mannella, C. A., Frank, J., Marko, M. Towards high-resolution three-dimensional imaging of native mammalian tissue: Electron tomography of frozen-hydrated rat liver sections. Journal of Structural Biology. 153 (1), 1-13 (2006).
- Al-Amoudi, A., Norlen, L. P., Dubochet, J. Cryo-electron microscopy of vitreous sections of native biological cells and tissues. Journal of Structural Biolology. 148 (1), 131-135 (2004).
- Rigort, A., et al. Focused ion beam micromachining of eukaryotic cells for cryoelectron tomography. Proceedings of the National Academy of Sciences. 109 (12), 4449-4454 (2012).
- Gorelick, S., et al. PIE-scope, integrated cryo-correlative light and FIB/SEM microscopy. Elife. 8, 45919(2019).
- Wu, G. H., et al. Multi-scale 3D cryo-correlative microscopy for vitrified cells. Structure. 28 (11), 1231-1237 (2020).
- Turk, M., Baumeister, W. The promise and the challenges of cryo-electron tomography. FEBS Letters. 594 (20), 3243-3261 (2020).
- Théry, M. Micropatterning as a tool to decipher cell morphogenesis and functions. Journal of Cell Science. 123 (24), 4201-4213 (2010).
- Tseng, Q., et al. Spatial organization of the extracellular matrix regulates cell-cell junction positioning. Proceedings of the National Academy of Sciences. 109 (5), 1506-1511 (2012).
- Hardelauf, H., et al. Micropatterning neuronal networks. Analyst. 139 (13), 3256-3264 (2014).
- Toro-Nahuelpan, M., et al. Tailoring cryo-electron microscopy grids by photo-micropatterning for in-cell structural studies. Nature Methods. 17 (1), 50-54 (2020).
- Engel, L., et al. Extracellular matrix micropatterning technology for whole cell cryogenic electron microscopy studies. Journal of Micromechanics and Microengineering. 29 (11), (2019).
- Engel, L., et al. Lattice micropatterning for cryo-electron tomography studies of cell-cell contacts. bioRxiv. , 272237(2021).
- Sibert, B. S., Kim, J. Y., Yang, J. E., Wright, E. R. Whole-cell cryo-electron tomography of cultured and primary eukaryotic cells on micropatterned TEM grids. bioRxiv. , 447251(2021).
- Egger, B., van Giesen, L., Moraru, M., Sprecher, S. G. In vitro imaging of primary neural cell culture from Drosophila. Nature Protocols. 8 (5), 958-965 (2013).
- Lu, W., Lakonishok, M., Gelfand, V. I. Kinesin-1-powered microtubule sliding initiates axonal regeneration in Drosophila cultured neurons. Molecular Biology of the Cell. 26 (7), 1296-1307 (2015).
- Ke, Z., et al. The morphology and assembly of respiratory syncytial virus revealed by cryo-electron tomography. Viruses. 10 (8), (2018).
- Stobart, C. C., et al. A live RSV vaccine with engineered thermostability is immunogenic in cotton rats despite high attenuation. Nature Communications. 7, 13916(2016).
- Hotard, A. L., et al. A stabilized respiratory syncytial virus reverse genetics system amenable to recombination-mediated mutagenesis. Virology. 434 (1), 129-136 (2012).
- Schindelin, J., et al. Fiji: an open-source platform for biological-image analysis. Nature Methods. 9 (7), 676-682 (2012).
- Mastronarde, D. N. Automated electron microscope tomography using robust prediction of specimen movements. Journal of Structural Biology. 152 (1), 36-51 (2005).
- Kremer, J. R., Mastronarde, D. N., McIntosh, J. R. Computer visualization of three-dimensional image data using IMOD. Journal of Structural Biology. 116 (1), 71-76 (1996).
- Tang, G., et al. EMAN2: an extensible image processing suite for electron microscopy. Journal of Structural Biology. 157 (1), 38-46 (2007).
- Yang, J. E., Larson, M. R., Sibert, B. S., Shrum, S., Wright, E. R. CorRelator: Interactive software for real-time high precision cryo-correlative light and electron microscopy. Journal of Structural Biology. , 107709(2021).
- Ke, Z., et al. Promotion of virus assembly and organization by the measles virus matrix protein. Nature Communications. 9 (1), 1736(2018).
- Kim, J., Yang, S., Wildonger, J., Wright, E. A new in situ neuronal model for cryo-ET. Microscopy and Microanalysis. 26 (2), 130-132 (2020).
- Bouvette, J., et al. Beam image-shift accelerated data acquisition for near-atomic resolution single-particle cryo-electron tomography. Nature Communications. 12 (1), 1957(2021).
- Schorb, M., Haberbosch, I., Hagen, W. J. H., Schwab, Y., Mastronarde, D. N. Software tools for automated transmission electron microscopy. Nature Methods. 16 (6), 471-477 (2019).
- Weis, F., Hagen, W. J. H., Schorb, M., Mattei, S. Strategies for optimization of cryogenic electron tomography data acquisition. Journal of Visual Experiments. (169), e62383(2021).
- Chreifi, G., Chen, S., Jensen, G. J. Rapid tilt-series method for cryo-electron tomography: Characterizing stage behavior during FISE acquisition. Journal of Structural Biology. 213 (2), 107716(2021).
- Anderson, D. E., Hinds, M. T. Endothelial cell micropatterning: methods, effects, and applications. Annals of Biomedical Engineering. 39 (9), 2329-2345 (2011).
- McWhorter, F. Y., Wang, T., Nguyen, P., Chung, T., Liu, W. F. Modulation of macrophage phenotype by cell shape. Proceedings of the National Academy of Sciences. 110 (43), 17253-17258 (2013).
- Kleinman, H. K., Luckenbill-Edds, L., Cannon, F. W., Sephel, G. C. Use of extracellular matrix components for cell culture. Analytical Biochemistry. 166 (1), 1-13 (1987).
- Fassler, F., Zens, B., Hauschild, R., Schur, F. K. M. 3D printed cell culture grid holders for improved cellular specimen preparation in cryo-electron microscopy. Journal of Structural Biology. 212 (3), 107633(2020).
- Schellenberger, P., et al. High-precision correlative fluorescence and electron cryo microscopy using two independent alignment markers. Ultramicroscopy. 143, 41-51 (2014).
Reimpresiones y Permisos
Solicitar permiso para reutilizar el texto o las figuras de este JoVE artículos
Solicitar permisoThis article has been published
Video Coming Soon
ACERCA DE JoVE
Copyright © 2025 MyJoVE Corporation. Todos los derechos reservados