
Method Article
Micropatterning Transmission Electron Microscopy Griglie per dirigere il posizionamento delle cellule all'interno dei flussi di lavoro della tomografia crioelettronica a cellule intere
* Questi autori hanno contribuito in egual misura
In questo articolo
Riepilogo
L'obiettivo di questo protocollo è quello di dirigere l'adesione e la crescita cellulare verso aree mirate di griglie per la crio-microscopia elettronica. Ciò si ottiene applicando uno strato antivegetativo che viene ablato in modelli specificati dall'utente seguito dalla deposizione di proteine della matrice extracellulare nelle aree modellate prima della semina cellulare.
Abstract
La tomografia crioelettronica a cellule intere (cryo-ET) è una potente tecnologia che viene utilizzata per produrre strutture di risoluzione a livello nanometrico di macromolecole presenti nel contesto cellulare e conservate in uno stato congelato-idratato quasi nativo. Tuttavia, ci sono sfide associate alla coltivazione e / o all'adesione delle cellule su griglie TEM in un modo che è adatto per la tomografia mantenendo le cellule nel loro stato fisiologico. Qui, viene presentato un protocollo dettagliato passo-passo sull'uso del micropatterning per dirigere e promuovere la crescita delle cellule eucariotiche sulle griglie TEM. Durante il micropatterning, la crescita cellulare è diretta depositando proteine della matrice extracellulare (ECM) all'interno di schemi e posizioni specificati sulla lamina della griglia TEM mentre le altre aree rimangono rivestite con uno strato antivegetativo. La flessibilità nella scelta del rivestimento superficiale e del design del modello rende il micropatterning ampiamente applicabile per una vasta gamma di tipi di celle. Il micropatterning è utile per lo studio delle strutture all'interno delle singole cellule e per sistemi sperimentali più complessi come le interazioni ospite-patogeno o le comunità multicellulari differenziate. La micropatterning può anche essere integrata in molti flussi di lavoro crio-ET a cellule intere a valle, tra cui la microscopia correlata a luce ed elettroni (cryo-CLEM) e la fresatura a fascio di ioni focalizzati (cryo-FIB).
Introduzione
Con lo sviluppo, l'espansione e la versatilità della microscopia crioelettronica (cryo-EM), i ricercatori hanno esaminato una vasta gamma di campioni biologici in uno stato quasi nativo da macromolecolare (~ 1 nm) ad alta risoluzione (~ 2 Å). Le tecniche di crio-EM a singola particella e di diffrazione elettronica sono meglio applicate a macromolecole purificate in soluzione o in uno stato cristallino, rispettivamente1,2. Mentre la tomografia crioelettronica (cryo-ET) è particolarmente adatta per studi strutturali e ultrastrutturali quasi nativi di oggetti grandi ed eterologhi come batteri, virus pleomorfi e cellule eucariotiche3. In crio-ET, le informazioni tridimensionali (3D) si ottengono inclinando fisicamente il campione sullo stadio del microscopio e acquisendo una serie di immagini attraverso il campione a diverse angolazioni. Queste immagini, o serie di inclinazioni, spesso coprono un intervallo di +60/-60 gradi con incrementi da uno a tre gradi. La serie di inclinazioni può quindi essere ricostruita computazionalmente in un volume 3D, noto anche come tomogramma4.
Tutte le tecniche crio-EM richiedono che il campione sia incorporato in un sottile strato di ghiaccio vitreo amorfo, non cristallino. Una delle tecniche di crio-fissazione più comunemente utilizzate è il congelamento a tuffo, in cui il campione viene applicato alla griglia EM, cancellato e rapidamente immerso in etano liquido o in una miscela di etano liquido e propano. Questa tecnica è sufficiente per la vetrificazione di campioni da <100 nm a ~10 μm di spessore, comprese le cellule umane in coltura, come le cellule HeLa5,6. Campioni più spessi, come mini-organoidi o biopsie tissutali, fino a 200 μm di spessore, possono essere vetrificati mediante congelamento ad alta pressione7. Tuttavia, a causa dell'aumento della diffusione elettronica di campioni più spessi, lo spessore del campione e del ghiaccio per il crio-ET è limitato a ~ 0,5 - 1 μm nei microscopi elettronici a trasmissione da 300 kV. Pertanto, la crio-ET a cellule intere di molte cellule eucariotiche è limitata alla periferia cellulare o alle estensioni delle cellule a meno che non vengano utilizzate ulteriori fasi di preparazione del campione, come la criosezione8 o la fresatura a fascio di ioni focalizzati9,10,11.
Una limitazione di molti esperimenti di imaging crio-ET a cellula intera è il throughput di raccolta dei dati12. A differenza della crio-EM a singola particella, in cui migliaia di particelle isolate possono spesso essere riprese da un singolo quadrato della griglia TEM, le cellule sono grandi, diffuse e devono essere coltivate a densità abbastanza bassa da consentire alle cellule di essere conservate in un sottile strato di ghiaccio vitreo. Spesso la regione di interesse è limitata a una particolare caratteristica o sotto-area della cellula. Un'ulteriore limitazione della produttività è la propensione delle cellule a crescere su aree che non sono suscettibili di imaging TEM, ad esempio su o vicino a barre della griglia TEM. A causa di fattori imprevedibili associati alla coltura cellulare sulle griglie TEM, sono necessari sviluppi tecnologici per migliorare l'accessibilità dei campioni e la produttività per l'acquisizione dei dati.
La micropatterning del substrato con proteine aderenti della matrice extracellulare (ECM) è una tecnica consolidata per la microscopia ottica a cellule vive per dirigere la crescita delle cellule su superfici rigide, durevoli e otticamente trasparenti come il vetro e altri substrati di coltura tissutale13,14. Il micropatterning è stato eseguito anche su superfici morbide e/o tridimensionali (3D). Tali tecniche non solo hanno permesso il posizionamento preciso delle cellule; hanno anche supportato la creazione di reti multicellulari, come i circuiti cellulari neurali modellati15. Portare il micropatterning a cryo-ET non solo aumenterà la produttività, ma può anche aprire nuovi studi per esplorare microambienti cellulari complessi e dinamici.
Recentemente, diversi gruppi hanno iniziato a utilizzare tecniche di micropatterning su griglie TEM attraverso approcci multipli16,17. Qui, l'uso di una tecnica di fotopatterning senza maschera per le griglie TEM è descritto utilizzando il sistema di micropatterning Alvéole PRIMO, che presenta pattern ad alta risoluzione e senza contatto. Con questo sistema di micropatterning, uno strato antivegetativo viene applicato sulla parte superiore del substrato, seguito dall'applicazione di un fotocatalizzatore e dall'ablazione dello strato antivegetativo in modelli definiti dall'utente con un laser UV. Le proteine ECM possono quindi essere aggiunte ai modelli per la coltura cellulare appropriata. Questo metodo è stato utilizzato da diversi gruppi per studi crio-ET su epiteliale-1 pigmento retinico (RPE1), rene canino Madin-Darby II (MDCKII), fibroblasti del prepuzio umano (HFF) e linee cellulari endoteliali16,17,18. Questo sistema di micropatterning è compatibile con più substrati di strati antivegetativi e con un reagente fotocatalizzatore liquido o gel. Una varietà di proteine ECM può essere selezionata e adattata per la specificità della linea cellulare, conferendo versatilità all'utente.
Il micropatterning è stato applicato con successo a una serie di progetti all'interno del laboratorio19. Qui viene presentato un protocollo di micropatterning, che include adattamenti specifici per studiare cellule HeLa in coltura, cellule BEAS-2B infette da virus respiratorio sinciziale (RSV) e neuroni primari della Drosophila melanogaster larvale20.
Protocollo
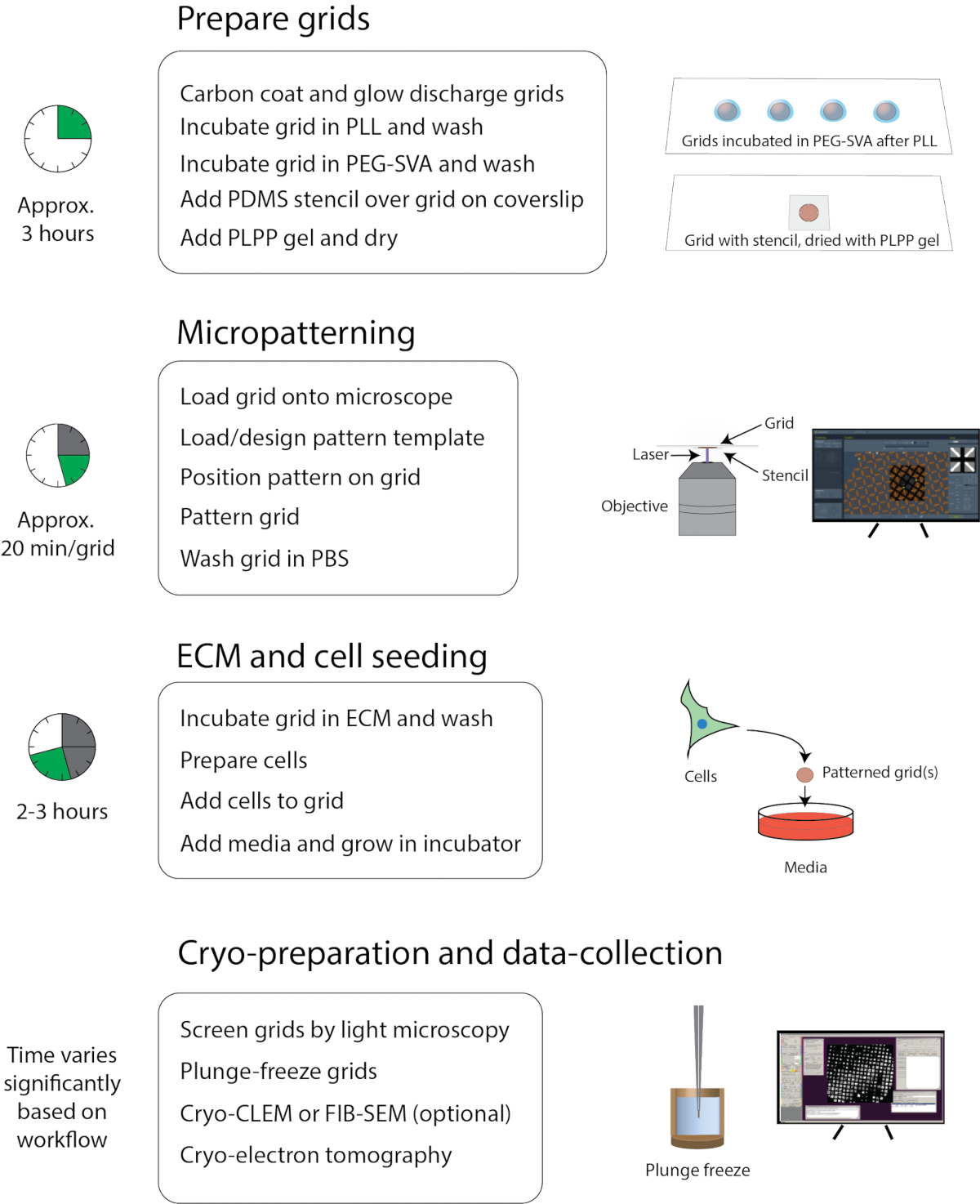
Il protocollo qui descritto è una raccolta dei metodi di coltura cellulare, micropatterning e imaging utilizzati dal laboratorio Wright e dal Cryo-EM Research Center presso l'Università del Wisconsin, Madison. Il flusso di lavoro è illustrato nella Figura 1. Ulteriori materiali di formazione e istruzione sono disponibili presso i seguenti siti: https://cryoem.wisc.edu o https://wrightlab.wisc.edu
1. Preparazione delle griglie per la modellazione
- Trasferire le griglie TEM su una slitta di vetro pulita, lato carbonio verso l'alto (lo spessore standard della lamina di carbonio è di 12 nm). Utilizzando un evaporatore di carbonio, ACE600, evaporare 5-8 nm di carbonio aggiuntivo sulle griglie per aumentare la durata complessiva del film di carbonio.
NOTA: questo passaggio non è necessario per le griglie SiO2 . Questo passaggio può anche essere fatto in anticipo; conservare le griglie rivestite in un ambiente a bassa umidità come un essiccatore sottovuoto. - Trasferire le griglie a un supporto per la preparazione della griglia e scaricare il bagliore delle griglie di carbonio verso l'alto. Utilizzando un sistema di scarica a bagliore, scarica le griglie per 60 s a 10 mA con una distanza di lavoro di 80 mm e una pressione del vuoto di 1,0 x 10-3 mbar. Fallo entro 15-30 minuti dal passaggio successivo.
NOTA: i portaprepista a griglia possono essere acquistati commercialmente o fatti in casa con un pezzo di carta da filtro su una piccola capsula di Petri.
2. Applicazione dello strato antivegetativo
NOTA: quando si maneggiano le griglie deve essere utilizzata una tecnica sterile adeguata e tutte le soluzioni devono essere sterili e/o sterilizzate con filtro.
- Trasferire le griglie (lato carbonio verso l'alto) su una slitta di vetro pulita o una slitta di copertura con almeno 1 cm di separazione tra le griglie. Pipetta 10 μL di 0,05% di poli-L-lisina (PLL) su ogni griglia. Incubare le griglie in una camera umida, come una scatola di plastica chiusa con asciugamani di carta umidi, per almeno 30 minuti.
NOTA: questo passaggio può essere esteso a durante la notte. Assicurarsi che il livello di umidità nella camera sia sufficiente per evitare che le griglie si secchino. - Lavare ogni griglia tre volte con 15 μL di 0,1 M HEPES pH 8,5. Per ogni lavaggio, rimuovere la maggior parte del liquido dalla griglia con una pipetta senza lasciare asciugare la griglia. Aggiungere 15 μL di tampone fresco, incubare per almeno 30 s e ripetere. Lasciare ogni griglia in 15 μL di 0,1 M HEPES dopo il lavaggio finale.
NOTA: in questa fase e nei passaggi futuri, è importante mantenere la griglia bagnata ed evitare il contatto tra la pipetta e la griglia. - Preparare 10 μL di 100 mg/mL di polietilenglicole-succinimidil valerato (PEG-SVA) in 0,1 M HEPES pH 8,5 per ogni griglia. Il PEG-SVA si dissolve rapidamente con una miscelazione delicata risultando in una soluzione limpida.
NOTA: non preparare la soluzione PEG-SVA in anticipo. PEG-SVA ha un'emivita di 10 minuti a pH 8,5. Evitare di esporre lo stock PEG-SVA a un'umidità eccessiva conservandolo in un essiccatore o in un ambiente asciutto a -20 °C e riscaldandolo a temperatura ambiente prima dell'apertura. - Subito dopo aver preparato la soluzione PEG-SVA, rimuovere la goccia di 15 μL di HEPES pH 8,5 da ciascuna griglia (facendo attenzione a non asciugare la griglia) e aggiungere una goccia di 10 μL della soluzione PEG-SVA. Incubare le griglie in una camera umida per almeno 1 ora.
NOTA: questo passaggio può essere esteso a durante la notte. Assicurarsi che l'umidità nella camera sia sufficiente per evitare che le griglie si secchino. - Lavare ogni griglia tre volte con 15 μL di acqua sterile. Per ogni lavaggio, rimuovere la maggior parte del liquido dalla griglia con una pipetta senza lasciare asciugare la griglia, aggiungere 15 μL di acqua dolce, incubare per almeno 30 s e ripetere. Lasciare ogni griglia in 15 μL di acqua dopo il lavaggio finale.
3. Applicazione del gel PLPP
- Preparare un coprimicroscopio pulito per ogni griglia. Completare i passaggi seguenti per ogni griglia, una griglia alla volta, per ridurre al minimo la possibilità che la griglia si secchi.
- Posizionare una goccia d'acqua da 1,0 μL al centro del coverslip per aiutare a posizionare la griglia sul coverslip e mantenere la griglia bagnata. Trasferire con attenzione la griglia dalla goccia d'acqua da 15 μL alla goccia d'acqua da 1,0 μL sulla coverslip. Assicurati di posizionare il lato del carbonio della griglia verso l'alto.
- Posizionare con attenzione uno stencil di polidimetilsilossano (PDMS) sulla griglia, avendo cura di mantenere la griglia centrata e di ridurre al minimo il contatto dello stencil con la lamina di carbonio della griglia.
- Aggiungere 1,0 μL di gel di cloruro di 4-benzoilbenzil-trimetilammonio (PLPP) sulla griglia. Pipettare delicatamente per mescolare (non toccare la griglia con la punta della pipetta).
- Spostare il coverslip con la griglia in una posizione buia per asciugare. Il gel si asciugherà in circa 15-30 minuti.
4. Taratura e progettazione del micropattern
- Colora un lato di una copertina di vetro con un evidenziatore. Aggiungi linee nere da un marcatore permanente a punta fine per facilitare la messa a fuoco. Posizionare il coverslip sul microscopio in modo tale che il lato colorato sia rivolto verso la lente dell'obiettivo. Utilizzando la modalità brightfield, concentrati sull'evidenziatore.
- Assicurarsi che il microscopio e il sistema di micropatterning siano accesi e che sia impostato il percorso luminoso corretto. Aprire Micromanager e il software Leonardo (Plugin > Leonardo) sul computer del microscopio.
- Selezionare Calibra e seguire le istruzioni visualizzate. Regolare la messa a fuoco del microscopio in modo che l'immagine proiettata sul vetrino sia a fuoco. Potrebbe essere necessario ridurre il tempo di esposizione. Dopo la calibrazione, selezionare Crea serie ora.
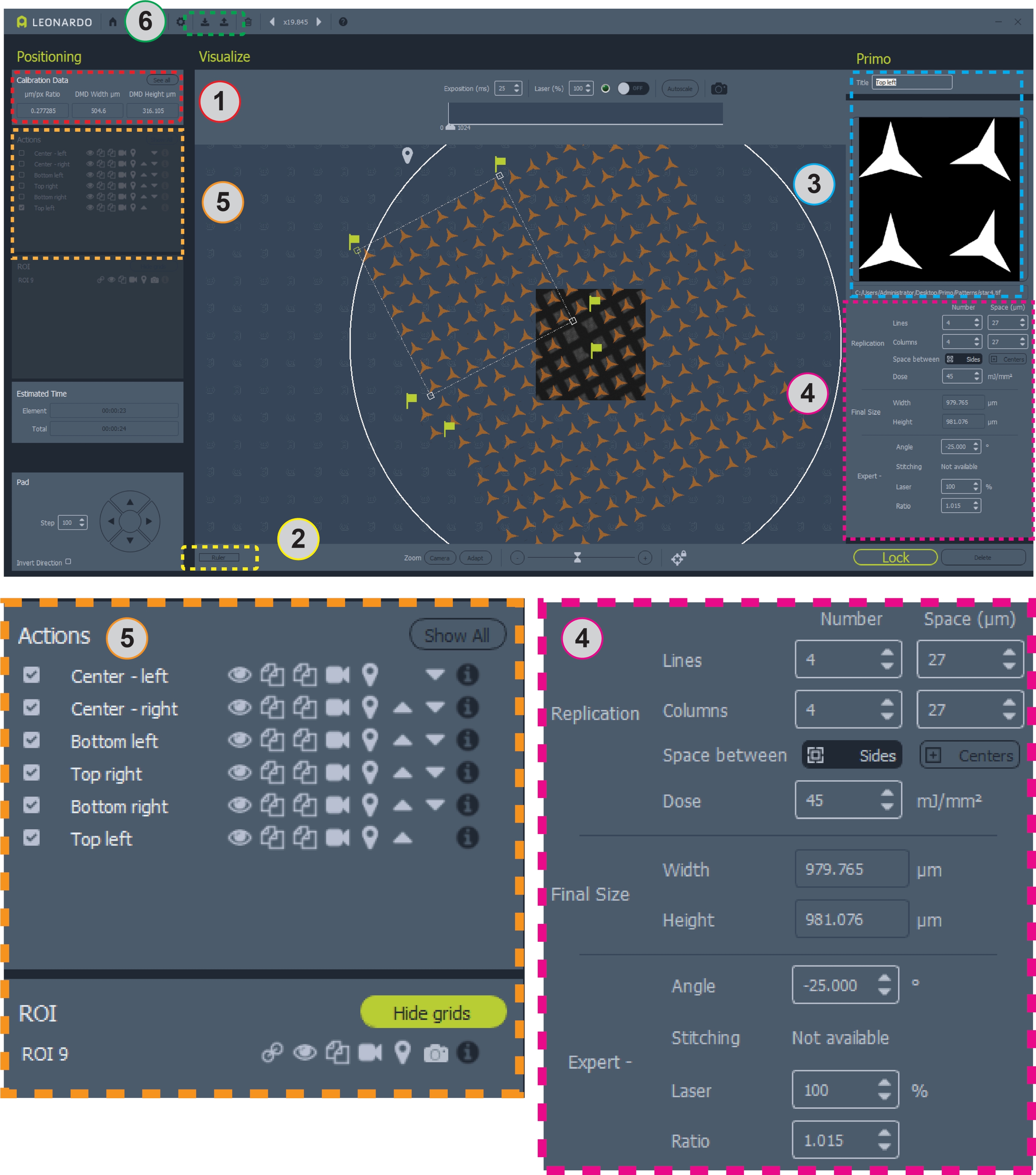
- Registrare il rapporto micrometro/pixel (μm/px) riportato sotto i dati di calibrazione nella finestra in alto a sinistra del programma (Figura 2, area 1). Utilizzare questo rapporto per determinare il numero di pixel da utilizzare per micrometro durante la progettazione di un pattern.
- Dopo la calibrazione, assicurarsi che il software sia ora aperto con una vista dal vivo del campo luminoso dal microscopio. Carica una griglia preparata su una copertina (sezione 3) sul palco con la griglia rivolta verso l'obiettivo. Posizionare lo stage e regolare lo stato attivo in modo che la griglia sia visibile nella finestra del software.
- Misurare la dimensione dei quadrati della griglia e delle barre della griglia in micrometri. Il software include un righello attivato dal pulsante vicino all'angolo in basso a sinistra per misurare la griglia (Figura 2, area 2). Ad esempio, i modelli utilizzati qui per una griglia a 200 mesh corrispondono a ~ 87 × quadrati di griglia da 87 μm e barre della griglia da ~ 36 μm.
NOTA: il software offre flessibilità nel ridimensionamento dei modelli al volo, quindi possono essere tollerate piccole imprecisioni nella misurazione. - In base alle misurazioni e ai rapporti di cui sopra, crea modelli con qualsiasi software di creazione di immagini. La dimensione minima della feature con un obiettivo di 20× è 1,2 μm. I modelli devono essere salvati come file .tiff a 8 bit non compressi.
- Assicurarsi che il software non ridimensiona le immagini a una dimensione di pixel diversa durante il salvataggio. Il modello dovrebbe adattarsi a una scatola da 800 × 800 pixel, sufficiente a coprire quattro quadrati della griglia.
NOTA: i pixel con un valore di 255 (bianco) verranno modellati alla massima intensità (dose totale del laser) e i pixel con un valore pari a zero (nero) non verranno modellati. Tutti i pixel con un valore intermedio saranno modellati con una dose di circa (X/255)*dose totale. Nella Figura 3A, i valori dei pixel di 255 e 129 sono stati utilizzati per i modelli in scala di grigi. Una volta che il modello è stato progettato, può essere salvato e riutilizzato senza modifiche.
- Assicurarsi che il software non ridimensiona le immagini a una dimensione di pixel diversa durante il salvataggio. Il modello dovrebbe adattarsi a una scatola da 800 × 800 pixel, sufficiente a coprire quattro quadrati della griglia.
5. Micropatterning
- Dopo la calibrazione, assicurarsi che il software sia ora aperto con una vista dal vivo del campo luminoso dal microscopio. Carica una griglia preparata su una copertina (sezione 3) sul palco con la griglia rivolta verso l'obiettivo. Posizionare lo stage e regolare la messa a fuoco per vedere la griglia nel software.
- Per un'esecuzione iniziale, progettare un nuovo modello. Nel software, selezionare Aggiungi ROI (non mostrato, nella posizione della Figura 2 area 3) e scegliere un cerchio da 3.000 μm. Posiziona il ROI del cerchio sulla griglia utilizzando l'immagine a campo luminoso sullo schermo come guida. Premere Bloc per garantire il ROI.
- Dopo aver bloccato il ROI, selezionare Aggiungi modello (non visualizzato, nella posizione dell'area 3 della Figura 2 ). Scegliete il modello disegnato nella sezione 4. Dividere la griglia in sei regioni per consentire la messa a fuoco e il posizionamento indipendenti in ogni regione per tenere conto delle griglie irregolari. Un'area quadrata a griglia 8 × 8 per ogni angolo della griglia e una regione quadrata a griglia 2 × 8 su ciascun lato del centro, lasciando i quattro quadrati della griglia centrale non stampati (Figura 2, immagine centrale).
- Utilizzare le opzioni di replica (Figura 2, area 4) per generare copie del modello iniziale per raggiungere il numero desiderato di copie totali del modello. Se necessario, regolate la spaziatura tra le copie in modo che corrisponda alla spaziatura tra i quadrati della griglia.
- Impostare la dose totale per il modello. 30 mJ/mm2 è un buon punto di partenza. Vedere la sezione di discussione per maggiori dettagli.
- In Opzioni esperti (Figura 2, area 4) regolare l'angolo della regione in modo che corrisponda a quello dei quadrati della griglia. Le regioni possono essere riposizionate utilizzando il mouse. Anche il rapporto (dimensione) dei modelli può essere regolato. Scorrere regolando l'angolo, la posizione, lo spazio tra e il rapporto del modello fino a quando i modelli non si allineano con i quadrati della griglia. Spostare lo stadio del microscopio per modificare la regione della griglia nella visualizzazione in campo luminoso dal vivo.
- Premere Blocca per salvare le modifiche apportate all'area.
- Per copiare un'area, fate clic sul pulsante Duplica (Figura 2, area 5, icona a due fogli di carta) accanto al suo nome nel pannello Azioni a sinistra. Per riposizionare, rinominare o modificare la copia, fate clic sul suo nome nel pannello Azione.
- Ripetere i passaggi da 5,4 a 5,9 se necessario per riempire tutte le regioni desiderate.
- Una volta progettato e posizionato il modello completo, salvare il file modello all'interno del software (Figura 2, area 6, barra con icona freccia su nella barra degli strumenti superiore).
- Quando si carica un modello salvato in precedenza (barra con icona a freccia in giù) centrare il ROI sulla griglia e premere Blocca. Fate clic su ciascuna regione nel pannello Azione per modificare l'angolo, la posizione, la dose e/o il file di pattern.
- Una volta posizionati il modello e i modelli, deselezionare tutte le aree tranne una nel Pannello azioni del software.
- Usa lo stadio del microscopio per navigare in quella regione e concentrarti sul foglio di carbonio. Facendo clic sull'icona Bulbo oculare nel pannello Azione (Figura 2, area 5) si attiva o disattiva la visualizzazione della sovrapposizione del pattern.
- Una volta che la griglia è a fuoco, chiudi l'otturatore a campo luminoso e premi l'icona Riproduci nell'angolo in basso a destra del software per iniziare il processo di creazione di modelli, che può essere monitorato dal vivo.
- Nel pannello delle azioni, selezionate la casella per l'area successiva. Aprire l'otturatore a campo luminoso in modo che la griglia sia visibile e centrare quella regione utilizzando lo stadio del microscopio. Ripetere i passaggi da 5.13 a 5.14 per ogni regione del Pannello azioni.
- Rimuovere il coperchio con la griglia dal microscopio e pipettare immediatamente 10 μL di soluzione salina sterile tamponata con fosfato (PBS) sulla griglia.
- Dopo 10 minuti, rimuovere lo stencil con una pinzetta, quindi lavare la griglia 3x con 15 μL di PBS. Dopo il lavaggio finale, posizionare ogni griglia in 15 μL di PBS e spostare le griglie in una posizione buia.
6. Deposizione di proteine ECM
- Per le cellule in coltura, seguire i passaggi 6.2-6.5; per i neuroni primari della Drosophila , seguire i passaggi 6.6-6.10.
- Preparare almeno 15 μL di ECM per ogni griglia. Per le cellule BEAS-2B, preparare una concentrazione finale di 0,01 mg/mL di fibronectina bovina e 0,01 mg/mL di fibrinogeno coniugato con fluoroforo in PBS sterile. Per le cellule HeLa, preparare 0,01 mg/mL di collagene bovino I e 0,1 mg/mL di fibrinogeno coniugato con fluoroforo in PBS sterile.
- Rimuovere la maggior parte del PBS da ogni griglia e applicare 15 μL di ECM. Incubare la griglia in una camera umida a temperatura ambiente per almeno 1 ora.
NOTA: questo passaggio può essere esteso alla notte a 4 °C. - Dopo l'incubazione in ECM, lavare ogni griglia 5 volte con PBS sterile. Per ogni lavaggio, rimuovere la maggior parte del liquido con una pipetta senza lasciare asciugare la griglia, aggiungere 15 μL di PBS fresco, incubare per almeno 30 s e ripetere. Lasciare ogni griglia in PBS dopo il lavaggio finale.
NOTA: le griglie possono essere conservate fino a una settimana in PBS a 4 °C senza alcun deterioramento della qualità osservato. - Utilizzare un microscopio a fluorescenza per rilevare il fluoroforo nell'ECM per confermare il pattern e che il foglio di carbonio è rimasto intatto. Alcuni quadrati rotti sono generalmente tollerabili.
- Per i neuroni primari della Drosophila , spostare le griglie modellate su un piatto di fondo di vetro da 30 mm contenente PBS sterile.
- Aspirare il PBS dal piatto e applicare 2 mL di concanavalina coniugata con fluoroforo da 0,5 mg/mL A. Incubare durante la notte a 25 °C in ambiente sterile.
- Rimuovere la soluzione di concanavalina A dal piatto (senza asciugare le griglie) e lavare le griglie 3 volte con PBS. Per ogni lavaggio, aggiungere e rimuovere 2 ml di PBS dal piatto.
- Utilizzare un microscopio a fluorescenza per rilevare il fluoroforo nell'ECM per confermare il pattern e che il foglio di carbonio è rimasto intatto. Alcuni quadrati rotti sono generalmente tollerabili.
- Dopo il lavaggio finale, rimuovere il PBS dal piatto con fondo di vetro e aggiungere 2 ml di Drosophila media di Schneider appena preparato e filtrato sterile21, contenente il 20% di siero bovino fetale inattivato dal calore (FBS), 5 μg/mL di insulina, 100 μg/mL di penicillina, 100 μg/mL di streptomicina e 10 μg/mL di tetraciclina. Incubare a 25 °C in un ambiente sterile fino a quando i neuroni sono pronti per essere placcati.
7. Preparazione delle cellule primarie di Drosophila prima della semina
- Sterilizzare una piastra di dissezione da 55 mm con il 70% di EtOH, quindi aggiungere al piatto 2-3 mL di soluzione salina di dissezione 1× filtrata sterile (9,9 mM HEPES pH 7,5, 137 mM NaCl, 5,4 mM KCl, 0,17 mM NaH2PO4, 0,22 mM KH2PO4, 3,3 mM glucosio, 43,8 mM di saccarosio)21.
- Raccogli delicatamente 30-40 larve di 3 ° instar dal cibo usando un paio di pinzette.
- Posizionare le larve nel tubo con 1× PBS, quindi trasferirle nel secondo tubo con 1× PBS per lavare le larve.
- Trasferire le larve nel tubo con il 70% di EtOH per lavare via il PBS, quindi trasferirle nel secondo tubo con il 70% di EtOH. Lasciare le larve nel secondo tubo per 2-3 minuti per sterilizzare le larve.
- Trasferire le larve in un tubo con 1× soluzione salina di dissezione, quindi trasferirle immediatamente nel secondo tubo con 1× soluzione salina di dissezione.
- Trasferire le singole larve sul piatto di dissezione contenente 1× soluzione salina di dissezione. Con un paio di pinze e un microscopio di dissezione, strappare rapidamente ogni larva per estrarre il cervello e trasferirlo al terzo tubo con 1× soluzione salina di dissezione. Ripeti fino a quando tutti i cervelli vengono estratti.
- Centrifugare il tubo contenente il cervello a 300 x g per 1 min.
- Scartare il surnatante e lavare con 1 mL di 1× soluzione salina di dissezione e centrifugare il tubo a 300 x g per 1 min. Ripetere questo passaggio ancora una volta.
- Scartare il surnatante fino a quando 200-250 μL non viene lasciato nel tubo e aggiungere 20 μL di 2,5 mg/mL liberasi in 1x soluzione salina di dissezione.
- Ruotare il tubo su un rotatore per 1 ora a temperatura ambiente; durante quest'ora, pipettare la soluzione 25-30 volte ogni 10 minuti. Alla fine, la soluzione dovrebbe essere leggermente opaca.
- Centrifugare le celle a 300 × g per 5 min.
- Scartare il surnatante, quindi aggiungere 1 mL di supporti Schneider integrati. Pipettare la soluzione 30 volte per mescolare.
- Centrifugare le celle a 300 × g per 5 min.
- Scartare il surnatante e lavare il pellet cellulare aggiungendo 1 mL di supporto Schneider integrato. Pipettare la soluzione 30 volte per mescolare.
- Centrifugare le celle a 300 × g per 5 min.
- Scartare il surnatante, quindi risospesciare il pellet cellulare con 300 μL di media di Schneider integrati. Pipettare la soluzione 30-40 volte per mescolare.
8. Coltura e infezione da RSV di cellule BEAS-2B e HeLa
- Mantenere le celle HeLa e le celle BEAS-2B nei palloni T75 a 37 °C e al 5 % di CO2. Cellule di passaggio ogni 3-4 giorni una volta raggiunto circa l'80% di confluenza. Mantenere le cellule HeLa in DMEM + 10% FBS + 1× Antibiotico-Antimicotico. Mantenere BEAS-2B in RPMI + 10% FBS + 1× Antibiotico-Antimicotico6,22,23.
- Per la semina di cellule non infette, saltare al paragrafo 9. Le cellule BEAS-2B e HeLa sono suscettibili all'infezione da RSV; Le cellule BEAS-2B sono state utilizzate per tutti gli esperimenti che coinvolgono RSV mostrati qui.
NOTA: Eseguire tutte le fasi BSL-2 in conformità con i protocolli istituzionali utilizzando un apposito armadietto di biosicurezza (BSC) e dispositivi di protezione individuale (DPI). - Prima dell'infezione da RSV delle cellule, il passaggio 5 × 104 cellule per pozzo in una piastra a 6 pozzetti (superficie ~ 9,6 cm2) con 2 ml di mezzi di crescita e incubare durante la notte.
- Tripsinizzare e contare un pozzo di cellule. Per tripsinizzare, aspirare i mezzi da un pozzetto e lavare con 2 ml di PBS sterile senza Mg2+ e Ca2+ per rimuovere i residui. Aggiungere 500 μL di soluzione di tripsina allo 0,25%. Incubare a 37 °C per 5-10 min. Controllare periodicamente le celle per vedere se vengono rilasciate dalla superficie. Una volta rilasciate le cellule, aggiungere 1,5 ml di terreno di coltura.
- Mescolare 100 μL di cellule tripsinizzate con 100 μL di tripano blu. Pipettare 10 μL di miscela cellulare diluita in un emocitometro. Contare le celle e calcolare il numero di celle per pozzetto. Usa questo numero per calcolare il MOI di seguito.
- Preparare una diluizione di RSV-A2mK+24 nei mezzi di crescita per ottenere un MOI di 10 per pozzetto in 750 μL di media. Il MOI di RSV-A2mK+ può essere calcolato dai titoli delle unità di focalizzazione fluorescenti (FFU) dello stock (ad esempio: per 1,0 × 105 celle per pozzetto e uno stock RSV di 1,0 × 108 FFU/mL, diluire lo stock virale da 1:75 a 1 × 106 FFU/750 μL o 1,33 × 106 FFU/mL).
- Aspirare il mezzo dalle cellule nel piatto a 6 pozzetti e aggiungere 750 μL della soluzione virale dall'alto a ciascun pozzetto.
- Scuotere la piastra a temperatura ambiente per 1 ora.
- Dopo 1 ora, portare il volume totale per pozzetto fino a 2 ml con terreno di crescita preriscaldato a 37 °C e posizionare la piastra in un incubatore impostato a 37 °C con il 5% di CO2 per 6 ore.
- Tripsinizzare le cellule per rilasciarle e procedere alla semina come descritto di seguito. Dopo la semina, incubare le griglie per ulteriori 18 ore prima del congelamento a tuffo (per un totale di 24 ore dopo l'infezione).
9. Semina cellulare su griglie micropatternate
- Per le cellule coltivate, seguire i passaggi 9.2-9.8; per i neuroni primari della Drosophila , seguire 9.9-9.11.
- Tripsinizzare le cellule per rilasciarle (vedere il passaggio 4 nella sezione 8 sopra). Per ridurre l'aggregazione cellulare, tripsinalizzare le cellule al 60 % o meno di confluenza.
- Mescolare 100 μL di cellule tripsinizzate con 100 μL di tripano blu. Pipettare 10 μL della miscela cellulare diluita in un emocitometro e contare le cellule.
- Diluire le cellule in media a 2 × 104 cellule/ml.
- Aggiungere 1 μL di materiale al centro di una parabola con fondo di vetro da 30 mm per facilitare il posizionamento della griglia e impedire che si asciughi. Trasferire la griglia dal PBS sul coperchio al centro del piatto con fondo di vetro. Aggiungere 10 μL di soluzione cellulare alla griglia.
- Utilizzando un microscopio a campo luminoso, osservare l'adesione cellulare alla griglia dopo 5 minuti. Se la maggior parte dei modelli rimane non occupata, aggiungere un'ulteriore goccia di 10 μL della soluzione cellulare. Mantenere le griglie e la soluzione cellulare a 37 °C durante le incubazioni.
- Ripetere il passaggio 9.6 fino a quando la maggior parte dei modelli non è occupata o molti modelli occupati iniziano ad avere più celle. Incubare la griglia per 2 ore nell'incubatore (37 °C, 5% CO2).
- Inondare il piatto con 2 ml di mezzi preriscaldati e incubare durante la notte (37 °C, 5% CO2).
- Per i neuroni primari della Drosophila , rimuovere il supporto dal piatto contenente griglia e placcare le cellule sul piatto.
- Attendere 30-60 minuti affinché le celle si attacchino, quindi aggiungere 2 ml di supporti Schneider integrati.
- Coltivare i neuroni in un incubatore a 25 °C per un minimo di 2-3 giorni prima del congelamento a immersione.
10. Imaging e vitrificazione di griglie modellate
- Posizionare il piatto di fondo di vetro contenente la griglia modellata e le cellule coltivate sul microscopio a fluorescenza.
- Acquisire immagini della griglia utilizzando il campo luminoso e i canali fluorescenti appropriati per rilevare il modello e qualsiasi altra etichettatura nelle celle. Assicurarsi che la densità e il posizionamento della cella siano adatti per l'imaging e l'analisi a valle.
NOTA: le immagini Brightfield e fluorescenti sono state elaborate nel pacchetto software FIJI25. - Preparare un congelatore crio-tuffo; il tipo di dispositivo di congelamento dipenderà dalla disponibilità, dal costo e dalle caratteristiche più adatte al campione.
NOTA: I neuroni primari della Drosophila sono stati preparati su un congelatore automatico e le cellule BEAS-2B sono state preparate utilizzando un congelatore semi-automatico. - Applicare fiduciali dorati ai campioni per un corretto allineamento della serie di inclinazioni. Tamponare i campioni per rimuovere i mezzi in eccesso, quindi immergere e congelare i campioni in un criogeno, come l'etano liquido raffreddato da azoto liquido. Per i neuroni primari della Drosophila , macchia per 4 s dal retro. Per le celle HeLa e BEAS-2B, macchia da entrambi i lati per 4-6 s. Le griglie congelate possono quindi essere conservate in azoto liquido fino a un ulteriore utilizzo.
- Immagini di cellule vetrificate in un microscopio crioelettronico, azionate a 300 kV con una telecamera con rivelatore di elettroni diretto. Imposta la raccolta tilt-series per ogni regione di interesse con software come SerialEM26 per la raccolta di dati cryo-EM/cryo-ET.
NOTA: La serie di inclinabili dei neuroni primari di Drosophila è stata raccolta su un rivelatore di elettroni diretto da -60° a 60° bidirezionalmente con incrementi di 2° a -8 μm di sfocatura con una dimensione dei pixel di 4,628 Å per una dose totale di 70-75 e-/Å2. La serie tilt-series di BEAS-2B infettati da RSV sono stati raccolti su un rivelatore di elettroni diretto con un filtro di energia (fessura 20 eV) a -5 μm di sfocatura con una dimensione dei pixel di 4.603 Å e una dose totale di ~ 80 e-/Å2. - Elaborare la serie di inclinazione per ricostruire i tomogrammi.
NOTA: i tomogrammi qui presentati sono stati ricostruiti utilizzando il pacchetto IMOD27; Il filtraggio passa-basso è stato eseguito utilizzando il pacchetto software EMAN228.
Risultati
Questa procedura è stata utilizzata per modellare le griglie EM per esperimenti crio-ET a cellule intere. L'intero flusso di lavoro presentato in questo studio, compresi i preparativi iniziali della coltura cellulare, il micropatterning (Figura 1) e l'imaging, comprende 3-7 giorni. È stata utilizzata una procedura in due fasi per generare lo strato antivegetativo applicando PLL alla griglia e successivamente collegando PEG mediante aggiunta del PEG-SVA reattivo. Lo strato antivegetativo può anche essere applicato in un unico passaggio aggiungendo PLL-g-PEG in un'incubazione. Il gel PLPP è un catalizzatore per il micropatterning UV, che è disponibile anche come liquido meno concentrato. Il gel consente il patterning a una dose significativamente ridotta rispetto al liquido, il che si traduce in un pattern molto più veloce. Con questo sistema, il tempo di pattern effettivo di una griglia TEM completa era di ~ 2 minuti. Il flusso di lavoro di micropatterning da solo dura generalmente 5-6 ore e consente a un individuo di modellare otto griglie per la coltura cellulare standard su griglie TEM.
Alcune fasi durante il processo di micropatterning richiedono lunghi tempi di incubazione (vedere i passaggi 2.1, 2.3, 6.4). Convenientemente, alcuni di questi passaggi, come la passivazione PLL (2.1) o la passivazione PEG-SVA (2.3), possono essere estesi a un'incubazione notturna. Inoltre, le griglie possono essere modellate in anticipo e conservate in una soluzione della proteina ECM o PBS per un uso successivo. Nel nostro studio, queste opzioni sono state preziose nei casi in cui i tempi di preparazione e semina cellulare sono critici, come i neuroni primari della Drosophila e l'infezione da RSV delle cellule BEAS-2B.
Le griglie sono state preparate in un ambiente di laboratorio di livello di biosicurezza generale 2 (BSL-2) utilizzando strumenti puliti, soluzioni sterili e includendo antibiotici / antimicotici nei mezzi di crescita6,22,29,30. Per i campioni particolarmente sensibili alla contaminazione microbica, lo strato antivegetativo e l'ECM possono essere applicati in una cappa di coltura tissutale o in un altro ambiente sterile. Inoltre, la griglia potrebbe essere lavata in etanolo tra la modellazione e l'applicazione ECM. Se si lavora con agenti infettivi, è importante adattare la procedura per conformarsi ai protocolli di biosicurezza appropriati.
Questo flusso di lavoro e le procedure presentate (Figura 1) hanno permesso alle cellule HeLa (Figura 4), alle cellule BEAS-2B infette da RSV (Figura 3, Figura 5) e ai neuroni larvali primari di Drosophila (Figura 6, Figura 7) di essere seminati su griglie EM modellate per una raccolta ottimale dei dati crio-ET.
Le cellule HeLa seminate su griglie TEM micropatterate rimangono vitali come determinato dalla colorazione fluorescente utilizzando un test di vitalità cellulare basato su calceina-AM e omodimero-1 di etidio (Figura 4A, B). Utilizzando un ECM misto di collagene e fibrinogeno, le cellule HeLa aderiscono prontamente ai modelli attraverso la griglia (Figura 4A, C). La morfologia complessiva delle cellule che si espandono lungo il modello è simile a quella delle cellule cresciute su griglie non modellate (Figura 4C, D). Nel caso delle celle HeLa, lo spessore totale della cella rimane ~< 10 μm con aree significativamente più sottili ~< 1 μm di spessore vicino alla periferia cellulare (Figura 4E,F).
Per gli studi RSV, abbiamo modellato interi quadrati della griglia usando un gradiente, con un'esposizione a basse dosi sui bordi e un modello di dose più elevato verso il centro (Figura 3A). I modelli di gradiente hanno prodotto risultati migliori durante la ricerca di virus rilasciati presenti vicino alla periferia delle cellule. Con questi modelli, le cellule sono state trovate per aderire preferenzialmente alla più alta concentrazione di ECM, ma sono anche in grado di aderire e crescere sulle concentrazioni ECM più basse. La dose relativa tra le aree dovrà essere ottimizzata quando si utilizzano modelli che richiedono dosi multiple. Se le dosi e quindi le concentrazioni di ECM sono troppo simili o troppo disparate l'una con l'altra, l'effetto dell'uso di dosi multiple andrà perso.
Nella Figura 3, una griglia TEM è stata modellata e successivamente seminata con cellule BEAS-2B infette da RSV e utilizzata per la raccolta di dati crio-EM. La Figura 4A è un'immagine fluorescente di ECM modellata su una griglia TEM utilizzando un modello di gradiente. L'adesione e la crescita cellulare lungo la regione centrale del modello possono essere viste nella Figura 3B come un'immagine a campo luminoso delle cellule 18 ore dopo la semina. Nella Figura 3C, il segnale fluorescente (rosso) dalla replica di RSV-A2mK+ è sovrapposto al segnale dell'ECM. La maggior parte delle cellule infette sono posizionate lungo la regione centrale a densità più elevata del modello di gradiente. Una mappa TEM a basso mag della griglia post crio-fissazione rivela un certo numero di cellule, comprese le cellule infette da RSV, posizionate sulla lamina di carbonio vicino al centro dei quadrati della griglia. Come precedentemente mostrato per le cellule cresciute su griglie TEM standard22, le serie tilt sono state localizzate e raccolte di virioni RSV in prossimità della periferia di cellule BEAS-2B infette cresciute su griglie micropatternate (Figura 5A, B). Molte delle proteine strutturali RSV possono essere identificate all'interno dei tomogrammi, tra cui nucleocapside (N) e la proteina di fusione virale (F) (Figura 5C, frecce blu e rosse rispettivamente).
Per gli studi primari sui neuroni Drosophila, è stato riscontrato che il modello stretto, vicino al limite di risoluzione offerto dal software (dove lo spessore del modello era di 2 μm), consentiva di isolare da una a poche cellule all'interno di un quadrato della griglia (Figura 6). Il soma neuronale è stato in grado di estendere i suoi neuriti per un periodo di diversi giorni all'interno del modello. Ciò ha permesso una facile identificazione e acquisizione in serie di inclinazione dei neuriti rispetto ai neuroni coltivati su griglie non modellate (Figura 7). È stato anche scoperto che la concanavalina A marcata fluorescentemente, una lectina che è stata utilizzata come ECM per colture neuronali di Drosophila in vitro20,21, è suscettibile di patterning.
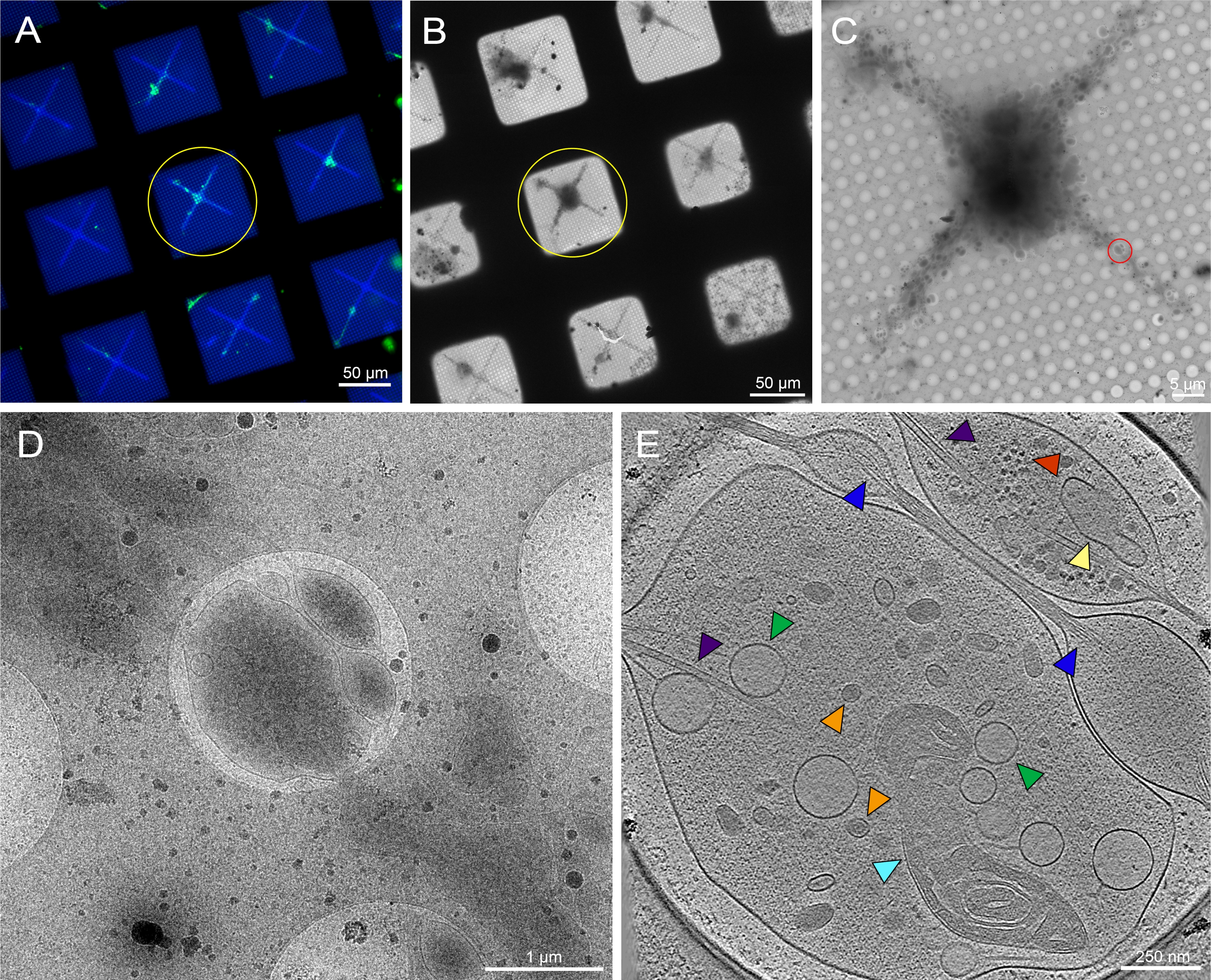
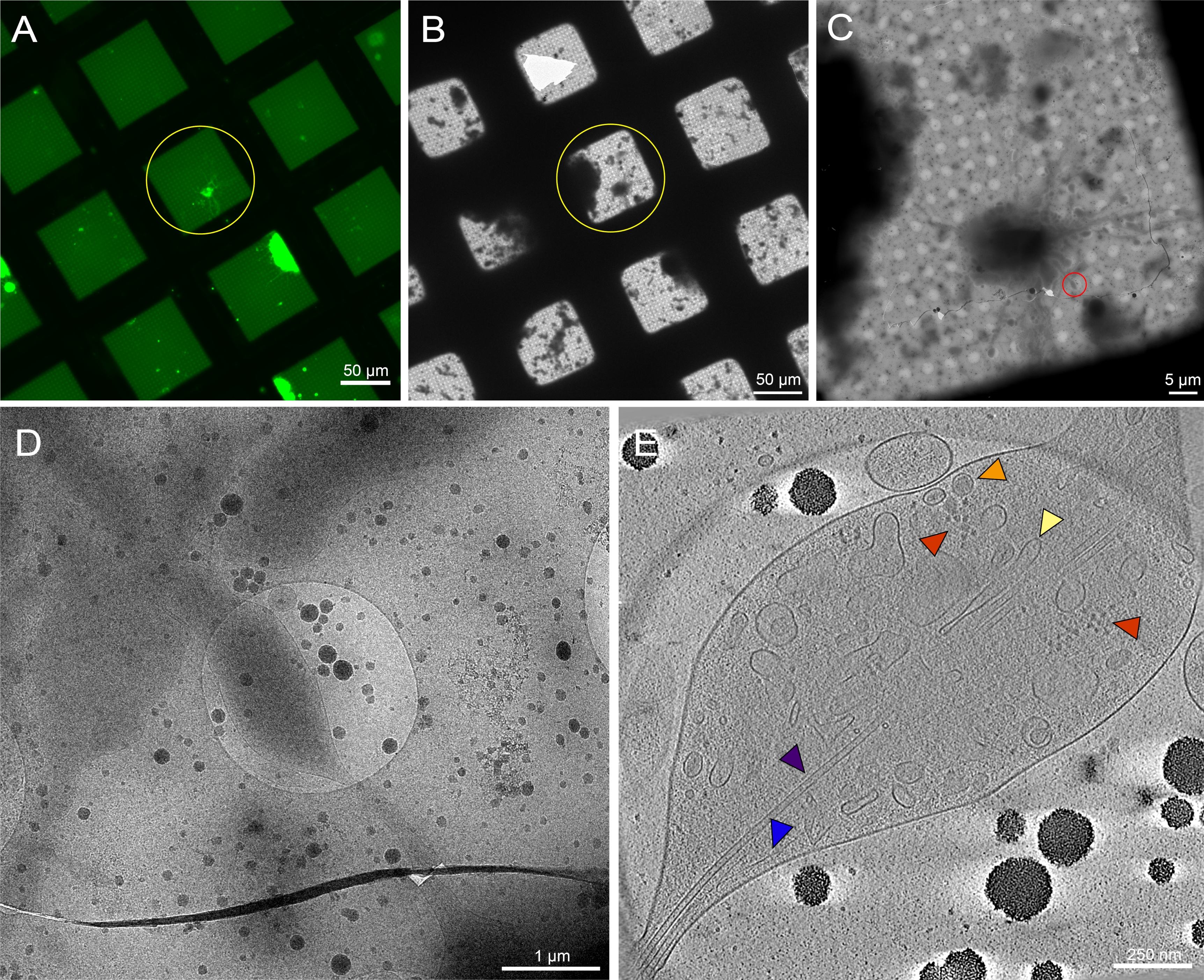
I neuroni della Drosophila delle larve della terza stella sono stati isolati secondo i protocolli precedentemente pubblicati20,21,31. I preparati neuronali sono stati applicati a griglie crio-EM micropatternate in cui la concanavalina A è stata depositata sul modello per regolare il posizionamento, la diffusione e l'organizzazione delle cellule. I neuroni su griglie modellate o non modellate sono stati autorizzati a incubare per un minimo di 48-72 ore e le griglie sono state quindi congelate. Un'immagine rappresentativa di una griglia EM micropatternata con diversi neuroni Drosophila distribuiti nelle regioni modellate è mostrata nella Figura 6A. Questi neuroni, derivati da un ceppo di mosca transgenica che ha espressione GFP panneurnalale nella membrana, possono essere facilmente monitorati dalla microscopia ottica non solo grazie alla sua etichettatura fluorescente, ma anche a causa della sua posizione all'interno dei micropattern. Mentre i neuroni coltivati su griglie non modellate possono anche essere tracciati attraverso la sua segnalazione GFP al microscopio ottico (Figura 7A, cerchio giallo), localizzarli in crio-EM è diventato sostanzialmente più difficile a causa della presenza di detriti cellulari e della contaminazione dai mezzi (Figura 7B, cerchio giallo). Tale presenza è stata ridotta per i neuroni su griglie modellate, probabilmente a causa del PEG nello strato antivegetativo delle regioni non modellate che respingono i detriti cellulari dall'adesione. A causa delle dimensioni del corpo cellulare neuronale e dei neuriti estesi (Figura 6A, B, cerchio giallo), sono state raccolte serie di inclinazione crio-ET lungo regioni più sottili delle cellule (Figura 6C, D, cerchio rosso). La membrana cellulare neuronale, un mitocondrio (ciano), microtubuli (viola), filamenti di actina (blu), strutture vescicolari (arancione e verde) e macromolecole come i ribosomi (rosso) sono stati ben risolti in montaggi di immagini ad alto ingrandimento e fette attraverso il tomogramma 3D (Figura 6E). Mentre caratteristiche subcellulari simili possono essere viste da tomogrammi 3D di neuroni non modellati (Figura 7E), la difficoltà nel localizzare bersagli cellulari vitali per la raccolta dei dati ha ridotto sostanzialmente il throughput.
Nella Figura 8, sono state assemblate immagini rappresentative di griglie con alcuni di questi problemi per facilitarne l'identificazione e la risoluzione dei problemi. Una volta determinate le condizioni ottimali, il micropatterning è un metodo affidabile e riproducibile per il posizionamento delle cellule su griglie per crio-TEM.

Figura 1: Flusso di lavoro generale di micropatterning per cryo-EM. Il flusso di lavoro può essere approssimativamente diviso in quattro parti: preparazione della griglia, micropatterning, ECM e semina cellulare, crio-preparazione e raccolta dei dati. I passaggi principali di ogni sezione sono elencati sotto le intestazioni e il tempo approssimativo per completare ogni sezione è mostrato a sinistra. Fare clic qui per visualizzare una versione più grande di questa figura.
{kind=link}

Figura 2: Cattura di schermata del software con pattern posizionato sulla griglia. L'area 1 contiene il rapporto μm/pix per la progettazione del modello. L'area 2 è il righello per misurare una griglia. L'area 3 è dove aggiungere o modificare modelli e ROI. L'area 4 contiene tutte le informazioni per il posizionamento e la dose del modello. L'area 5 contiene opzioni per i modelli, tra cui l'attivazione o disattivazione delle sovrapposizioni, la copia o l'eliminazione dei modelli e la selezione dei modelli per il micropatterning. L'area 6 è dove i modelli possono essere salvati e caricati. Viste più ampie delle aree 4 e 5 sono mostrate di seguito per chiarezza. Fare clic qui per visualizzare una versione più grande di questa figura.
{kind=link}

Figura 3: Cellule BEAS-2B infette da RSV sulla griglia crio-TEM modellata. (A) Immagine fluorescente della griglia modellata dopo l'aggiunta di ECM etichettato fluorescentemente. Il modello di input viene visualizzato nell'angolo in basso a sinistra. (B) Immagine a campo luminoso di cellule BEAS-2B cresciute sulla griglia in A. (C) Unione dell'immagine in A (ciano) e B (grigio) con immagine fluorescente di cellule infette da RSV (rosso) immediatamente prima del congelamento a tuffo; le cellule infette esprimono mKate-2. Le barre di scala sono 500 μm. (D) Mappa crio-TEM a basso ingrandimento della griglia in B dopo il congelamento a immersione. Le immagini fluorescenti sono pseudocolori. Le barre della scala sono 500 μm. Fare clic qui per visualizzare una versione più grande di questa figura.
{kind=link}

Figura 4: Colorazione viva/morta di cellule modellate e non modellate. (A) Immagine fluorescente di cellule HeLa cresciute su una griglia modellata e colorate con calceina-AM (macchia di cellule vive, verde) e omodimero-1 di etidio (macchia di cellule morte, rosso). (B) Cellule HeLa cresciute su una griglia non modellata e colorate come in A. (C) Proiezione di z-stack confocali di una cellula HeLa su una griglia Quantfoil R2/2 modellata con 0,01 mg/mL di collagene e fibrinogeno 647 ECM (rosso). La cellula è stata macchiata con calcein-AM (verde) e Hoechst-33342 (blu). (D) Cellule HeLa su griglia non modellata incubate con 0,01 mg/mL di collagene e fibrinogeno 647 ECM, incubate e colorate con calceina-AM e Hoecsht-33342. Le immagini fluorescenti sono state unite alla luce trasmessa (scala di grigi). (E) Proiezione X,Z di C. (F) X,Z proiezione di D. Le immagini sono pseudocolori. Le barre di scala in (A) e (B) sono 500 μm; le barre della scala in (C) - (F) sono 10 μm. Fare clic qui per visualizzare una versione più grande di questa figura.
{kind=link}

Figura 5: Crio-ET della cella BEAS-2B infetta da RSV sulla griglia crio-TEM modellata. (A) Mappa quadrata della griglia Cryo-EM della cella BEAS-2B infetta da RSV. Il limite approssimativo della cella è indicato dalla linea verde tratteggiata. (B) Immagine ad alta risoluzione dell'area inscatolata in rosso in (A). Il limite approssimativo della cella è indicato dalla linea verde tratteggiata. I virioni RSV possono essere visti vicino alla periferia cellulare (freccia bianca e scatola gialla). (C) Singola fetta z dal tomogramma raccolta nell'area della casella gialla di cui alla lettera B). Le frecce rosse puntano alla proteina di fusione RSV F, le frecce blu puntano al complesso delle ribonucleoproteine (RNP). Le barre di scala in (A)-(C) sono incorporate nell'immagine. Fare clic qui per visualizzare una versione più grande di questa figura.
{kind=link}

Figura 6: Neuroni primari derivati dal cervello delle larve di Drosophila melanogaster della 3a stella sulla griglia crio-TEM modellata. (A) Montaggio della griglia della griglia di microscopia a fluorescenza a cellule vive sovrapposte di neuroni Drosophila che esprimono GFP mirata alla membrana su quadrati di griglia modellati con concanavalina fluorescente da 0,5 mg / mL A. Verde: neuroni Drosophila. Blu: Photopattern. (B) Montaggio dell'immagine Cryo-EM della griglia in (A) dopo la crioconservazione. Il cerchio giallo indica lo stesso quadrato della griglia di cui alla (A). (C) Montaggio dell'immagine Cryo-EM del quadrato evidenziato dal cerchio giallo in (A) e (B). (D) Immagine di ingrandimento più elevato dell'area delimitata dal cerchio rosso in (C), dove è stata raccolta una serie di inclinazioni sui neuriti della cellula. E. fetta spessa 25 nm di un tomogramma ricostruito dalla serie di inclinazione che è stata acquisita dal cerchio rosso in (C). Vari organelli possono essere visti in questo tomogramma, come i mitocondri (ciano), i microtubuli (viola), le vescicole dense (arancione), le vescicole chiare (verde), il reticolo endoplasmatico (giallo) e l'actina (blu). Le macromolecole, come i ribosomi (rosso), possono anche essere viste nell'angolo in alto a destra. Le immagini fluorescenti sono pseudocolori. Le barre di scala in (A)-(E) sono incorporate nell'immagine. Fare clic qui per visualizzare una versione più grande di questa figura.
{kind=link}

Figura 7: Neuroni primari derivati dal cervello delle larve di Drosophila melanogaster della 3a stella su griglie non modellate. (A) Montaggio della griglia della griglia di microscopia a fluorescenza a cellule vive dei neuroni Drosophila che esprimono GFP mirata alla membrana su quadrati di griglia con concanavalina 0,5 mg / mL A. Verde: neuroni Drosophila. (B) Montaggio della griglia Cryo-EM della stessa griglia in (A) dopo il congelamento a immersione. Il cerchio giallo mostra lo stesso quadrato della griglia di cui al punto (A). Si noti la presenza di detriti cellulari e contaminazione dei media, che ha reso difficile l'identificazione del bersaglio rispetto alle griglie modellate. (C) Montaggio di immagini Cryo-EM del quadrato evidenziato dai cerchi gialli nelle mappe (A) e (B). (D) Immagine di ingrandimento più elevato dell'area delimitata dal cerchio rosso in (C), dove è stata raccolta una serie di inclinazioni sui neuriti della cellula. (E) fetta di 25 nm di spessore del tomogramma ricostruito della serie di inclinazione da (C) e (D). Un certo numero di organelli sono visibili in questo tomogramma, come i microtubuli (viola), l'actina (blu), il reticolo endoplasmatico (giallo) e le vescicole dense (arancione). Si possono vedere anche macromolecole, come i ribosomi (rossi). Le immagini fluorescenti sono pseudocolori. Le barre di scala in (A)-(E) sono incorporate nell'immagine. Fare clic qui per visualizzare una versione più grande di questa figura.
{kind=link}

Figura 8: Esempi di possibili problemi con la creazione di modelli. Immagini fluorescenti di ECM etichettato depositate su griglie micropatterned. (A) Pattern non uniforme attraverso la griglia a causa della distribuzione non uniforme del gel PLPP. (B) L'ECM non può aderire alle aree coperte dallo stencil PDMS durante la creazione di modelli. (C) Modello di gradiente saturo (lato destro) o motivo invertito (a sinistra) su una griglia modellata con una dose totale troppo elevata. (D) ECM aderisce alle aree sulle barre della griglia e all'area modellata a causa dei riflessi del laser UV durante la modellazione. Le immagini sono pseudocolori; il modello di input è mostrato in basso a sinistra; le barre della scala sono 100 μm. Fare clic qui per visualizzare una versione più grande di questa figura.
{kind=link}
| Questione | Cause potenziali | Risoluzione dei problemi |
| Micropatterning | ||
| Impossibile vedere l'illuminazione dal laser PRIMO | • Il percorso della luce non è impostato correttamente | • Controllare che il percorso luminoso del microscopio sia impostato correttamente |
| • Il laser PRIMO non è acceso o il laser è interbloccato | ||
| Molti quadrati a griglia spezzati | • Toccare la lamina della griglia con pinzette o pipetta durante la manipolazione | • Maneggiare le griglie con cura |
| • Griglia asciugata durante le incubazioni o il lavaggio | • Non lasciare asciugare la griglia durante lavaggi e incubazioni | |
| Grandi aree non tagliate | • Insufficiente copertura del gel | • Assicurarsi che il gel si diffonda uniformemente sulla griglia durante l'aggiunta |
| • Lamina della griglia sfocata durante la creazione di modelli | • Aggiungere un ulteriore microlitro di gel | |
| •Area coperta da stencil | • Controlla lo stato attivo prima di modellare ogni regione | |
| • Griglia accuratamente centrale nello stencil | ||
| Modello saturo o invertito | • Dose errata | • Prova una gamma di dosi totali per il modello |
| • Insufficiente copertura del gel | • Assicurarsi che la griglia sia uniformemente coperta di gel | |
| • Prova valori diversi per i modelli in scala di grigi | ||
| Motivo sfocato | • Scarsa messa a fuoco durante il patterning | • Ripetere la calibrazione PRIMO alla stessa altezza del campione |
| • Calibrazione errata | • Concentrati sulla lamina della griglia prima di modellare | |
| • Dividere il modello in regioni aggiuntive per la creazione di modelli | ||
| ECM che aderisce al di fuori del modello | • Riflessi da gel o polvere | • Assicurarsi che il gel sia asciutto prima della modellatura |
| • Assicurati che il coverslip e l'obiettivo siano puliti | ||
| ECM non visibile dopo la creazione di serie | • Sbiancamento foto | • Ridurre al minimo l'esposizione alla luce all'ECM prima dell'imaging |
| • Dose errata durante il patterning | • Prova un intervallo di valori di dose totale per il modello | |
| • Tempo di incubazione ECM insufficiente | • Aumentare il tempo di incubazione per ECM | |
| Semina cellulare | ||
| Aggregazione delle cellule | • Eccessiva digestione | • Utilizzare una percentuale inferiore di tripsina o tempo per il rilascio di cellule aderenti |
| • Alta densità di celle | • Cellule di passaggio e/o digestione a confluenza inferiore | |
| • Non agitare le cellule durante il rilascio | ||
| • Pipettare delicatamente la soluzione cellulare o utilizzare filtri cellulari | ||
| Celle che non aderiscono alle aree modellate | • ECM non è adatto per il tipo di cella | • Prova diverse concentrazioni e composizione ECM |
| • La vitalità delle cellule è diminuita prima della semina | • Garantire che la coltura cellulare e le condizioni di rilascio cellulare non danneggino le cellule | |
| Cellule che non si espandono dopo l'adesione | • ECM o modello non adatto al tipo di cella | • Prova diversi modelli ed ECM |
| • In alcuni casi una lamina più continua (R1.2/20 vs R2/1) può favorire l'espansione cellulare |
Tabella 1: Potenziali problemi durante il micropatterning. In questa tabella vengono descritti alcuni problemi che un utente può riscontrare durante il micropatterning o il cell-seeding. Per ogni problema vengono fornite le cause potenziali e la risoluzione dei problemi. Immagini rappresentative di alcuni problemi possono essere viste nella Figura 8.
Discussione
Microscopi elettronici moderni e avanzati e pacchetti software ora supportano la raccolta automatizzata di dati crio-EM e crio-ET in cui centinaia o migliaia di posizioni possono essere prese di mira e visualizzate in pochi giorni32,33,34,35. Un fattore limitante significativo per i flussi di lavoro crio-ET a cellula intera è stato l'ottenimento di un numero sufficiente di bersagli collezionabili per griglia. Recentemente, un certo numero di gruppi ha sviluppato protocolli per griglie di micropatterning per crio-EM, con un vantaggio di migliorare l'efficienza di raccolta dei dati16,17,18. Qui viene presentato un protocollo per l'utilizzo di un sistema di micropatterning disponibile in commercio per micropattern TEM griglie per studi crio-ET di neuroni Drosophila primari e linee cellulari umane in coltura (non infette o infettate da RSV). Questo sistema di micropatterning è versatile e molti passaggi possono essere ottimizzati e adattati per adattarsi a specifici obiettivi sperimentali. Un utente con esperienza teM e microscopia a fluorescenza può diventare rapidamente esperto nella preparazione della griglia e nella micropatterning. Con un'attenta pratica, buoni risultati dovrebbero essere raggiungibili dopo alcune iterazioni. Di seguito, vengono discusse alcune delle opzioni disponibili, le considerazioni per gli utenti, i potenziali benefici e le future applicazioni del micropatterning per la crio-EM.
Una delle considerazioni importanti per la crio-ET a cellula intera è la selezione della griglia EM. Le griglie EM sono composte da due parti: un telaio a rete (o supporto strutturale) e la lamina (o film), che è la superficie continua o bucata su cui cresceranno le cellule. Le griglie a rete di rame sono comunemente usate per la crio-EM di proteine e complessi isolati. Tuttavia, non sono adatti per la crio-ET a cellule intere a causa della citotossicità del rame. Invece, una rete d'oro è comunemente usata per la tomografia cellulare. Altre opzioni includono nichel o titanio, che possono fornire vantaggi rispetto all'oro come una maggiore rigidità16. Le griglie EM sono disponibili con diverse dimensioni di mesh per supportare una vasta gamma di applicazioni. Le dimensioni delle maglie più grandi offrono più spazio per le celle per crescere tra le barre della griglia e più aree che sono suscettibili di raccolta di serie tilt, anche se al costo di una maggiore fragilità complessiva del campione. La lamina più comunemente usata è il carbonio amorfo perforato o bucato, come i Quantifoils o le griglie C-flat. I bersagli biologici possono essere ripresi attraverso i fori nel carbonio o attraverso il carbonio elettrone-traslucido. Griglie come R 2/1 o R 2/2, dove i fori sono larghi 2 μm che sono distanziati rispettivamente di 1 e 2 μm, forniscono un gran numero di fori e quindi un gran numero di aree potenziali per la raccolta dei dati. Tuttavia, alcune cellule possono crescere ed espandersi meglio su superfici più uniformi come griglie R 1.2/20 o carbonio continuo. Per l'elaborazione dei campioni a valle mediante fresatura a fascio di ioni focalizzati (cryo-FIB), la lamina viene rimossa attraverso la fresatura, riducendo le preoccupazioni sulla presenza continua del film sottostante. Come per la rete, sono disponibili anche pellicole di altri materiali, con il protocollo di pattern qui presentato ugualmente adatto per le griglie SiO2 . Le griglie comunemente usate includono griglie a 200 maglie a 200 squadre in carbonio continuo o film siO2 (spaziatura di ~ 90 μm tra le barre delle griglie) per crio-ET a cellule intere.
Ci sono una serie di considerazioni quando si progetta un modello. La maggior parte di queste decisioni sono guidate dal tipo di cellula e dallo scopo dell'esperimento. Un buon punto di partenza è scegliere un modello che approssima la forma e le dimensioni delle cellule in coltura. Molti studi hanno dimostrato effetti significativi della forma del modello sulla crescita cellulare e sulla disposizione citoscheletrica13,36,37. Prestare particolare attenzione durante la progettazione del modello se ciò potrebbe alterare il target di interesse. Diversi modelli per ogni tipo di cellula sono stati testati per determinare quali modelli hanno promosso l'adesione e la crescita cellulare. La flessibilità del sistema di micropatterning consente di testare più modelli su una singola griglia e di cambiare modelli per diverse griglie all'interno di un singolo esperimento. Modelli più grandi (~ 50-90 μm), come quelli usati qui, aumentano la probabilità che più cellule aderiscano a una singola regione del modello e consentono alle cellule di espandersi ed estendersi dopo l'adesione. Modelli più vincolati (20-30 μm) possono essere appropriati in esperimenti in cui l'isolamento cellulare è più critico dell'espansione cellulare, come per esperimenti di fresatura a fascio di ioni focalizzati (crio-FIB). Per le applicazioni di tomografia, potrebbe essere necessario considerare l'impatto dell'asse di inclinazione. Se un modello è posizionato in modo tale che tutte le cellule crescano parallele l'una all'altra in un'unica direzione, è possibile che tutte le cellule siano perpendicolari all'asse di inclinazione quando caricate sullo stadio del microscopio, con conseguente minore qualità dei dati.
Sulle griglie non formattate, le celle spesso aderiscono preferenzialmente alle barre della griglia, dove non possono essere visualizzate da TEM. Anche su griglie modellate, le celle sono spesso osservate per essere posizionate negli angoli dei quadrati della griglia parzialmente sia sulla lamina di carbonio modellata che sulla barra della griglia. Recentemente, il micropatterning è stato utilizzato per posizionare intenzionalmente parte della cella sulla barra della griglia18. Questo potrebbe essere preso in considerazione per esperimenti in cui non è fondamentale avere l'intera periferia cellulare sulla lamina. Questo può essere particolarmente importante per le cellule che possono crescere più grandi di un singolo quadrato a griglia, come i neuroni primari che crescono in più giorni.
Esistono molti strumenti che possono essere utilizzati per progettare un modello. Qui, il modello era limitato a meno di 800 pixel in qualsiasi dimensione in modo tale che il modello potesse essere ruotato a qualsiasi angolo e comunque adattarsi all'area massima che può essere modellata in una singola proiezione da questo sistema di micropatterning. Ciò consente all'utente di ruotare il modello per essere correttamente orientato con la griglia indipendentemente dall'orientamento della griglia sul microscopio. Qui, la griglia è stata divisa in sei aree di pattern. In primo luogo, ciò consente la regolazione della messa a fuoco tra le diverse regioni della griglia. Le griglie dorate, in particolare, sono molto malleabili e potrebbero non essere completamente piatte sul vetro. Una corretta messa a fuoco è essenziale per risultati di pattern puliti e raffinati. Utilizzando modelli segmentati, è necessario apportare solo piccole modifiche alla posizione del modello se la griglia si sposta leggermente durante il processo di creazione della creazione di modelli, anche se questo di solito non è un problema quando si utilizza il gel PLPP con gli stencil PDMS. Infine, i quattro quadrati centrali della griglia sono rimasti invariati. Ciò supporta un utente in grado di identificare chiaramente il centro della griglia, il che è molto utile per gli esperimenti di imaging correlativo.
Il software di patterning per questo sistema di micropatterning, Leonardo, ha anche caratteristiche più avanzate come la cucitura e la possibilità di importare modelli come PDF, che esulano dallo scopo di questo protocollo. Questo software include anche il rilevamento di microstrutture e il posizionamento automatizzato del modello che può essere utilizzato sulle griglie TEM. Questa funzione è particolarmente utile quando la griglia è molto piatta e può essere modellata senza la necessità di regolare la messa a fuoco tra diverse aree.
La selezione di una proteina ECM può avere un impatto significativo sull'adesione e l'espansione cellulare. È noto che alcune cellule subiscono cambiamenti fisiologici se coltivate su substrati specifici38. Più proteine e concentrazioni di ECM sono state testate per qualsiasi nuovo tipo di cellula sulla base di precedenti lavori riportati in letteratura. Laminina, il fibrinogeno, la fibronectina e il collagene sono ampiamente utilizzati per le cellule coltivate e possono essere utilizzati come punto di partenza se non sono disponibili altri dati. Tuttavia, anche altre proteine ECM devono essere considerate se le proteine ECM comunemente usate non riescono a conferire proprietà di aderenza adeguate per le cellule. Ciò era particolarmente vero per i neuroni primari della Drosophila , poiché un'alta concentrazione della pianta lectina concanavalina A era necessaria per una corretta aderenza cellulare. La compatibilità dell'adesione e della crescita cellulare con l'ECM può essere testata modellando su piatti o vetrini prima di passare alle griglie TEM. Questo approccio di pre-screening è efficace in termini di tempo e di costi se è necessario esaminare un gran numero di combinazioni. L'inclusione di una proteina ECM coniugata fluorescentemente è preziosa per valutare il successo e la qualità del patterning.
La semina cellulare è uno dei passaggi più importanti per la crio-ET a cellule intere, con o senza micropatterning6,16,39. Per la Drosophila primaria o altri neuroni, che sono fragili, instabili in sospensione e possono essere limitati in quantità, gli approcci di semina singola sono preferiti rispetto alla semina cellulare sequenziale monitorata. Una singola fase di semina a una densità cellulare ottimizzata, come descritto nel protocollo per i neuroni Drosophila, è un'opzione praticabile per la maggior parte dei tipi di cellule. Tuttavia, è anche possibile seminare cellule sul substrato a una concentrazione iniziale inferiore e aggiungere più cellule in modo monitorato, come descritto qui e in altra letteratura18. Questa semina sequenziale può fornire risultati più coerenti in alcuni casi. Simile alla coltura cellulare standard, si dovrebbe sempre prestare attenzione a mantenere la vitalità cellulare e ridurre al minimo l'aggregazione cellulare durante l'isolamento.
Quando si inizia con il micropatterning, ci sono alcune potenziali insidie che sono dannose per il risultato finale. Un'attenta gestione della griglia e una tecnica sterile, una distribuzione uniforme del gel PLPP, una dose e una messa a fuoco adeguate durante il pattern e il mantenimento della vitalità cellulare prima della semina sono tra le considerazioni più importanti per il successo. Un elenco di alcuni dei potenziali problemi e delle soluzioni è stato raccolto nella Tabella 1.
Le griglie micropatternate possono essere utilizzate per aiutare a posizionare le celle per stabilire una densità di cella coerente attraverso la griglia e per posizionare le regioni di interesse in aree adatte alla raccolta di serie di inclinazioni16,18. Il posizionamento e il posizionamento delle cellule possono essere utilizzati come marcatori fiduciali per la correlazione negli esperimenti crio-CLEM, riducendo la necessità di fragili griglie di ricerca e marcatori fiduciali fluorescenti. Tuttavia, va notato che tali marcatori fiduciali possono ancora essere utili per la correlazione dell'accuratezza sub-micrometrica29,40. Inoltre, una distribuzione uniforme delle cellule isolate è anche molto vantaggiosa per gli esperimenti di fresatura a fascio di ioni focalizzati (crio-FIB) per massimizzare il numero di cellule da cui è possibile tagliare la lamella16.
L'aggiunta del micropatterning ai flussi di lavoro crio-EM comporterà miglioramenti misurabili nel throughput dei dati e potenzialmente consentirà nuovi esperimenti. Man mano che la tecnica viene ulteriormente adottata e sviluppata, applicazioni più avanzate di micropatterning, tra cui gradienti ECM, deposizioni ECM multiple e assemblaggio di microstrutture, espanderanno ulteriormente le capacità di crio-ET per studiare bersagli e processi biologici in pieno contesto cellulare.
Divulgazioni
Gli autori non hanno nulla da rivelare.
Riconoscimenti
Ringraziamo la dott.ssa Jill Wildonger, la dott.ssa Sihui Z. Yang e la signora Josephine W. Mitchell del Dipartimento di Biochimica dell'Università del Wisconsin, Madison per aver generosamente condiviso il ceppo di mosca elav-Gal4, UAS-CD8::GFP (Bloomington stock center, #5146). Vorremmo anche ringraziare il Dr. Aurélien Duboin, il Signor Laurent Siquier e la Signora Marie-Charlotte Manus di Alvéole e il Signor Serge Kaddoura di Nanoscale Labs per il loro generoso supporto durante questo progetto. Questo lavoro è stato supportato in parte dall'Università del Wisconsin, Madison, dal Dipartimento di Biochimica dell'Università del Wisconsin, Madison, e il servizio sanitario pubblico concede R01 GM114561, R01 GM104540, R01 GM104540-03W1 e U24 GM139168 a E.R.W. e R01 AI150475 a P.W.S. dal NIH. Una parte di questa ricerca è stata supportata dalla sovvenzione NIH U24 GM129547 ed eseguita presso il PNCC dell'OHSU e accessibile tramite EMSL (grid.436923.9), un DOE Office of Science User Facility sponsorizzato dall'Office of Biological and Environmental Research. Siamo anche grati per l'uso di strutture e strumentazione presso il Cryo-EM Research Center nel Dipartimento di Biochimica dell'Università del Wisconsin, Madison.
Materiali
| Name | Company | Catalog Number | Comments |
| 0.1% (w/v) Poly-L-Lysine | Sigma | P8920-100ML | |
| 0.22 µm syringe filters PVDF membrane | Genesee | 25-240 | |
| 22x60-1 Glass cover slip | Fisher | 12545F | |
| 5/15 Tweezers | EMS (Dumont) | 0203-5/15-PO | |
| Antibiotic-Antimycotic (100X) | ThermoFisher (Gibco) | 15240096 | |
| BEAS-2B cells | ATCC | CRL-9609 | |
| Collagen I, bovine | ThermoFisher (Gibco) | A1064401 | |
| Concanavalin A, Alexa Fluor 350 Conjugate | ThermoFisher (Invitrogen) | C11254 | |
| DMEM | Fisher (Lonza) | BW12-604F | |
| EtOH | Fisher (Decon Labs) | 22-032-600 | |
| Fetal Bovine Serum | ATCC | 30-2020 | |
| Fibrinogen From Human Plasma, Alexa Fluor 647 Conjugate | ThermoFisher (Invitrogen) | F35200 | |
| Fibronectin Bovine Protein, Plasma | ThermoFisher (Gibco) | 33010018 | |
| Glass bottom dish | MatTek | P35G-1.5-20-C | |
| Glucose | VWR | 0643-1KG | |
| Grid prep holder | EMS | 71175-01 | |
| HeLa cells | ATCC | CCL-2 | |
| Hemacytometer | Fisher (SKC, Inc.) | 22600100 | |
| HEPES | Fisher (ACROS Organics) | AC172572500 | |
| Hoechst 33342 | ThermoFisher (Invitrogen) | H3570 | |
| Insulin | Fisher (Sigma Aldrich) | NC0520015 | |
| KCl | MP Bio | 194844 | |
| KH2PO4 | Fisher (ACROS Organics) | AC212595000 | |
| Leica-DMi8 | Leica Microsystems | Can be customized with camera, stage, and objective attachments | |
| Leonardo | Alvéole | https://www.alveolelab.com/our-products/leonardo-photopatterning-software/ | |
| Liberase Research Grade | Fisher (Supply Solutions) | 50-100-3280 | |
| LIVE/DEAD Viability/Cytotoxicity Kit | ThermoFisher (Invitrogen) | L3224 | |
| Microscope camera | Hammamatsu | C13440-20CU | |
| Motorized stage | Märzhäuser Wetzlar | 00-24-599-0000 | |
| NaCl | Fisher (Fisher BioReagents) | BP358-1 | |
| NaH2PO4 | Fisher (ACROS Organics) | AC207802500 | |
| NaOH | Fisher (Alfa Aesar) | AAA1603736 | |
| PBS | Corning | 21-040-CV | |
| PDMS stencils | nanoscaleLABS | PDMS_STENCILS_EM | https://www.alveolelab.com/our-products/pdms-stencil-multiwell-plate/ |
| PEG-SVA | nanoscaleLABS | PEG-SVA-1GR | mPEG-Succinimidyl Valerate, MW 5,000 |
| Penicillin | Fisher (Research Products International Corp) | 50-213-641 | |
| pH strips | Fisher (Millipore Sigma) | M1095350001 | pH probe can also be used |
| PLPP gel | nanoscaleLABS | PLPP-GEL-300UL | https://www.alveolelab.com/our-products/plpp-photoactivatable-reagent/ |
| PRIMO | Alvéole | https://www.alveolelab.com/our-products/primo-micropatterning/ | |
| pSynkRSV-I19F (BAC containing RSV A2-mK+ antigenomic cDNA ) | BEI Resources | NR-36460 | https://www.beiresources.org/Catalog/BEIPlasmidVectors/NR-36460.aspx |
| Quantifoil grids | EMS (Quantifoil) | Q2100AR1 | 2 µm holes spaced 1 µm apart, other dimensions are available |
| RPMI | Fisher (Lonza) | BW12-702F | |
| RSV A2-mK+ | see entry for pSynkRSV-19F | - | Described in Hotard et al. [22]. Can be generated from pSynkRSV-ll9F |
| Schneider's Media | ThermoFisher (Gibco) | 21720-024 | |
| SerialEM | SerialEM (https://bio3d.colorado.edu/SerialEM/ ) | https://bio3d.colorado.edu/SerialEM/ | |
| Straight tweezers | EMS (Dumont) | 72812-D | |
| Streptomycin | Fisher (Fisher BioReagents) | BP910-50 | |
| Sucrose | Avantor | 4097-04 | |
| Tetracycline | Sigma | T8032-10MG | |
| Titan Krios electron microscope | ThermoFisher | 300kV, with direct electron detector camera and energy filter | |
| Trypsin | ThermoFisher (Gibco) | 15090046 | |
| Tube Revolver/Rotator | Fisher (Thermo Scientific) | 11676341 | |
| UAS:mcD8:GFP Drosophila fly strain | Bloomington Drosophila Stock Center | 5146 | http://flybase.org/reports/FBtp0002652.html |
Riferimenti
- Nogales, E., Scheres, S. H. Cryo-EM: A unique tool for the visualization of macromolecular complexity. Molecular Cell. 58 (4), 677-689 (2015).
- Martynowycz, M. W., Gonen, T. From electron crystallography of 2D crystals to MicroED of 3D crystals. Current Opinion in Colloid and Interface Science. 34, 9-16 (2018).
- Wagner, J., Schaffer, M., Fernandez-Busnadiego, R. Cryo-electron tomography-the cell biology that came in from the cold. FEBS Letters. 591 (17), 2520-2533 (2017).
- Wan, W., Briggs, J. A. Cryo-electron tomography and subtomogram averaging. Methods in Enzymology. 579, 329-367 (2016).
- Bäuerlein, F. J., Pastor-Pareja, J. C., Fernández-Busnadiego, R. Cryo-electron tomography of native Drosophila tissues vitrified by plunge freezing. bioRxiv. , 437159(2021).
- Hampton, C. M., et al. Correlated fluorescence microscopy and cryo-electron tomography of virus-infected or transfected mammalian cells. Nature Protocols. 12 (1), 150-167 (2017).
- Hsieh, C. E., Leith, A., Mannella, C. A., Frank, J., Marko, M. Towards high-resolution three-dimensional imaging of native mammalian tissue: Electron tomography of frozen-hydrated rat liver sections. Journal of Structural Biology. 153 (1), 1-13 (2006).
- Al-Amoudi, A., Norlen, L. P., Dubochet, J. Cryo-electron microscopy of vitreous sections of native biological cells and tissues. Journal of Structural Biolology. 148 (1), 131-135 (2004).
- Rigort, A., et al. Focused ion beam micromachining of eukaryotic cells for cryoelectron tomography. Proceedings of the National Academy of Sciences. 109 (12), 4449-4454 (2012).
- Gorelick, S., et al. PIE-scope, integrated cryo-correlative light and FIB/SEM microscopy. Elife. 8, 45919(2019).
- Wu, G. H., et al. Multi-scale 3D cryo-correlative microscopy for vitrified cells. Structure. 28 (11), 1231-1237 (2020).
- Turk, M., Baumeister, W. The promise and the challenges of cryo-electron tomography. FEBS Letters. 594 (20), 3243-3261 (2020).
- Théry, M. Micropatterning as a tool to decipher cell morphogenesis and functions. Journal of Cell Science. 123 (24), 4201-4213 (2010).
- Tseng, Q., et al. Spatial organization of the extracellular matrix regulates cell-cell junction positioning. Proceedings of the National Academy of Sciences. 109 (5), 1506-1511 (2012).
- Hardelauf, H., et al. Micropatterning neuronal networks. Analyst. 139 (13), 3256-3264 (2014).
- Toro-Nahuelpan, M., et al. Tailoring cryo-electron microscopy grids by photo-micropatterning for in-cell structural studies. Nature Methods. 17 (1), 50-54 (2020).
- Engel, L., et al. Extracellular matrix micropatterning technology for whole cell cryogenic electron microscopy studies. Journal of Micromechanics and Microengineering. 29 (11), (2019).
- Engel, L., et al. Lattice micropatterning for cryo-electron tomography studies of cell-cell contacts. bioRxiv. , 272237(2021).
- Sibert, B. S., Kim, J. Y., Yang, J. E., Wright, E. R. Whole-cell cryo-electron tomography of cultured and primary eukaryotic cells on micropatterned TEM grids. bioRxiv. , 447251(2021).
- Egger, B., van Giesen, L., Moraru, M., Sprecher, S. G. In vitro imaging of primary neural cell culture from Drosophila. Nature Protocols. 8 (5), 958-965 (2013).
- Lu, W., Lakonishok, M., Gelfand, V. I. Kinesin-1-powered microtubule sliding initiates axonal regeneration in Drosophila cultured neurons. Molecular Biology of the Cell. 26 (7), 1296-1307 (2015).
- Ke, Z., et al. The morphology and assembly of respiratory syncytial virus revealed by cryo-electron tomography. Viruses. 10 (8), (2018).
- Stobart, C. C., et al. A live RSV vaccine with engineered thermostability is immunogenic in cotton rats despite high attenuation. Nature Communications. 7, 13916(2016).
- Hotard, A. L., et al. A stabilized respiratory syncytial virus reverse genetics system amenable to recombination-mediated mutagenesis. Virology. 434 (1), 129-136 (2012).
- Schindelin, J., et al. Fiji: an open-source platform for biological-image analysis. Nature Methods. 9 (7), 676-682 (2012).
- Mastronarde, D. N. Automated electron microscope tomography using robust prediction of specimen movements. Journal of Structural Biology. 152 (1), 36-51 (2005).
- Kremer, J. R., Mastronarde, D. N., McIntosh, J. R. Computer visualization of three-dimensional image data using IMOD. Journal of Structural Biology. 116 (1), 71-76 (1996).
- Tang, G., et al. EMAN2: an extensible image processing suite for electron microscopy. Journal of Structural Biology. 157 (1), 38-46 (2007).
- Yang, J. E., Larson, M. R., Sibert, B. S., Shrum, S., Wright, E. R. CorRelator: Interactive software for real-time high precision cryo-correlative light and electron microscopy. Journal of Structural Biology. , 107709(2021).
- Ke, Z., et al. Promotion of virus assembly and organization by the measles virus matrix protein. Nature Communications. 9 (1), 1736(2018).
- Kim, J., Yang, S., Wildonger, J., Wright, E. A new in situ neuronal model for cryo-ET. Microscopy and Microanalysis. 26 (2), 130-132 (2020).
- Bouvette, J., et al. Beam image-shift accelerated data acquisition for near-atomic resolution single-particle cryo-electron tomography. Nature Communications. 12 (1), 1957(2021).
- Schorb, M., Haberbosch, I., Hagen, W. J. H., Schwab, Y., Mastronarde, D. N. Software tools for automated transmission electron microscopy. Nature Methods. 16 (6), 471-477 (2019).
- Weis, F., Hagen, W. J. H., Schorb, M., Mattei, S. Strategies for optimization of cryogenic electron tomography data acquisition. Journal of Visual Experiments. (169), e62383(2021).
- Chreifi, G., Chen, S., Jensen, G. J. Rapid tilt-series method for cryo-electron tomography: Characterizing stage behavior during FISE acquisition. Journal of Structural Biology. 213 (2), 107716(2021).
- Anderson, D. E., Hinds, M. T. Endothelial cell micropatterning: methods, effects, and applications. Annals of Biomedical Engineering. 39 (9), 2329-2345 (2011).
- McWhorter, F. Y., Wang, T., Nguyen, P., Chung, T., Liu, W. F. Modulation of macrophage phenotype by cell shape. Proceedings of the National Academy of Sciences. 110 (43), 17253-17258 (2013).
- Kleinman, H. K., Luckenbill-Edds, L., Cannon, F. W., Sephel, G. C. Use of extracellular matrix components for cell culture. Analytical Biochemistry. 166 (1), 1-13 (1987).
- Fassler, F., Zens, B., Hauschild, R., Schur, F. K. M. 3D printed cell culture grid holders for improved cellular specimen preparation in cryo-electron microscopy. Journal of Structural Biology. 212 (3), 107633(2020).
- Schellenberger, P., et al. High-precision correlative fluorescence and electron cryo microscopy using two independent alignment markers. Ultramicroscopy. 143, 41-51 (2014).
Ristampe e Autorizzazioni
Richiedi autorizzazione per utilizzare il testo o le figure di questo articolo JoVE
Richiedi AutorizzazioneThis article has been published
Video Coming Soon
Personale delle biblioteche
Copyright © 2025 MyJoVE Corporation. Tutti i diritti riservati