
Method Article
رسم خرائط تفاعلات جينوم 3D للثدييات مع Micro-C-XL
In This Article
Summary
يتم تقديم بروتوكول لرسم خرائط تنظيم الجينوم ثلاثي الأبعاد بدقة النيوكليوسوم باستخدام طريقة التقاط التشكل على مستوى الجينوم Micro-C-XL هنا.
Abstract
يعد تنظيم الكروموسوم ثلاثي الأبعاد (3D) عاملا رئيسيا في تنظيم الجينوم ومواصفات نوع الخلية. على سبيل المثال ، يعتقد أن العناصر التنظيمية لرابطة الدول المستقلة ، والمعروفة باسم المعززات ، تنظم نشاط المروجين البعيدين من خلال التفاعل في الفضاء ثلاثي الأبعاد. لقد حولت تقنيات التقاط التشكل الكروموسومي على مستوى الجينوم (3C) ، مثل Hi-C ، فهمنا لكيفية تنظيم الجينوم في الخلايا. الفهم الحالي لتنظيم الجينوم ثلاثي الأبعاد محدود بالدقة التي يمكن من خلالها حل التنظيم الطوبولوجي للكروموسومات في الفضاء ثلاثي الأبعاد. يقيس Micro-C-XL طي الكروموسوم بدقة على مستوى النيوكليوسوم ، الوحدة الأساسية للكروماتين ، من خلال استخدام نوكلياز المكورات الدقيقة (MNase) لتجزئة الجينوم أثناء بروتوكول التقاط تشكل الكروموسوم. وهذا يؤدي إلى تحسين نسبة الإشارة إلى الضوضاء في القياسات ، وبالتالي تسهيل الكشف بشكل أفضل عن مواقع العزل وحلقات الكروموسوم مقارنة بتقنيات 3D الأخرى على مستوى الجينوم. يتم تقديم بروتوكول مفصل ومفصل ومدعوم بصريا لإعداد عينات Micro-C-XL عالية الجودة من خلايا الثدييات في هذه المقالة.
Introduction
Micro-C-XL هي تقنية على مستوى الجينوم لقياس تشكل الجينوم 3D بدقة نيوكليوسوم. يعتمد Micro-C-XL على تقنية Hi-C القائمة على ربط القرب المستخدمة على نطاق واسع ، والتي حولت فهمنا لكيفية تنظيم جينومات 3D1. تم تطوير Micro-C-XL وتكراره الأول ، Micro-C ، في البداية في Saccharomyces cerevisiae 2,3 وتم تكييفه لاحقا مع أنظمة خلايا الثدييات ، والتي أظهر البروتوكول إمكاناته الكاملة في الكشف عن الميزات قصيرة المدى للجينوم ثلاثي الأبعاد ، مثل حلقات الكروموسوم ومواقع العزل. يعتمد هذا الإصدار على منشورات Micro-C-XL الحديثة للثدييات 4,5. نظرا لأن Micro-C-XL يحل محل Micro-C ، يشار إلى Micro-C-XL من الآن فصاعدا باسم Micro-C في المخطوطة.
الاختلافات الرئيسية بين Micro-C و Hi-C6 هي كما يلي: 1) تجزئة الجينوم مع نوكلياز المكورات الدقيقة (MNase) مقارنة بإنزيمات التقييد و 2) روابط متشابكة إضافية مع تباعد ذري أكبر بين المجموعات التفاعلية مقارنة بالفورمالديهايد فقط. تساهم كلتا الخطوتين بشكل كبير في تحسين نسبة الإشارة إلى الضوضاء في Micro-C مقارنة ب Hi-C التقليدي. يحد حجم التجزئة من الدقة التي يمكن من خلالها حل تنظيم الجينوم 3D أثناء بروتوكول ربط القرب. MNase هو نيوكلياز يهضم بشكل تفضيلي الحمض النووي الذي يمكن الوصول إليه ويترك الحمض النووي المحمي بالنيوكوسومات سليما. أظهرت بصمة النيوكليوسوم باستخدام تسلسل MNase أن النيوكليوسومات تغطي بالكامل معظم جينومات حقيقية النواة7. نظرا لأن النيوكليوسومات موزعة في جميع أنحاء الجينوم بمتوسط تباعد يتراوح بين 160 و 220 نقطة أساس ، اعتمادا على الأنواع ونوع الخلية ، فإن MNase هو الإنزيم المثالي لرسم خرائط عالية الدقة لبنية الجينوم.
بالإضافة إلى ذلك ، يؤدي استخدام رابط متشابك إضافي مع الفورمالديهايد (FA) في طريقة Micro-C إلى تحسين نسبة الإشارة إلى الضوضاء 2,8. تسهل الروابط المتشابكة الخاصة بالأمين مع الفواصل الذرية الأطول بين المجموعات التفاعلية الروابط المتقاطعة بين البروتين والبروتين. هذه عادة ما تكون غلوتارات ديسوتشينيميديل (DSG) أو إيثيلين جلايكول بيس-سكسينيميديل سكسينات (EGS) مع فواصل 7.7 Å و 16.1 Å ، على التوالي. يتضح تقليل الضوضاء من خلال EGS أو DSG بشكل خاص في التجارب ذات معدلات التجزئة العالية ، مثل Micro-C ، ويفترض أن يحدث بسبب انخفاض معدل أحداث الربط العشوائي8.
بروتوكول Hi-C 3.0 الذي تم تطويره مؤخرا والذي يستخدم التشابك ESG / DSG ومجموعات متعددة من إنزيمات التقييد يقلل من الضوضاء في تجارب Hi-C ويحسن بشكل كبير من اكتشاف حلقات الكروموسوم ومواقع العزل 8,9. ومع ذلك ، وجدت مقارنة كل موقع على حدة لميزات بيانات التفاعل المختلفة أن Micro-C كان لديه اكتشاف متفوق للميزات قصيرة المدى ، مثل حلقات الكروموسوم ومواقع العزل ، مقارنة بكل من Hi-C 3.0 و Hi-C8 التقليدي. ومع ذلك ، فإن Hi-C 3.0 يحسن اكتشاف الميزات قصيرة المدى ويحافظ على اكتشاف قوي لتجزئة الجينوم مقارنة ب Hi-C التقليدي. باختصار ، يجب تحديد اختيار طريقة التقاط تشكل الكروموسوم من خلال السؤال الموضوعي والبيولوجي.
هنا ، نقدم بروتوكولا خطوة بخطوة لتجارب Micro-C الناجحة التي يمكن أن تكشف عن تنظيم جينوم 3D.
Protocol
1. زراعة الخلايا والتشابك
- خلايا الاستزراع وفقا للاحتياجات التجريبية للحصول على ما لا يقل عن 1 × 107 خلايا. هنا ، نمت الخلايا عند 37 درجة مئوية مع 5٪ CO2 في وسط E14 (DMEM مع البيروفات و L-glutamine ، 15٪ FBS ، 1x LIF ، 1x NEAA ، 1٪ pen-strep ، 0.1 mM β-mercaptoethanol [انظر جدول المواد]) وتمر كل يومين.
ملاحظة: تم تطبيق هذا البروتوكول بنجاح على أنواع مختلفة من الخلايا من أنواع متعددة ، مثل الإنسان العاقلو Mus musculus. في هذا المثال ، تم استخدام الخلايا الجذعية الجنينية للفأر (ES). - حصاد الخلايا عند التقاء 70٪ -80٪ عن طريق استنشاق الوسط. اغسل مرة واحدة ب 5 مل من DPBS ، واحتضن الخلايا ب 3 مل من التربسين المسخن مسبقا بنسبة 0.25٪ لكل طبق 10 سم لمدة 2-3 دقائق عند 37 درجة مئوية.
- قم بإخماد التربسين ب 7 مل من وسط E14 المسخن مسبقا وانقل الخلايا المنفصلة إلى أنبوب سعة 50 مل.
- حبيبات الخلايا عن طريق الطرد المركزي عند 300 × جم لمدة 5 دقائق في درجة حرارة الغرفة (RT). تخلص من المادة الطافية وأعد تعليق حبيبات الخلية في الوسط. عد الخلايا باستخدام عداد خلية.
ملاحظة البروتوكول قوي لبعض الاختلافات في تركيزات الخلايا ، وقد تم استخدام عدادات الخلايا المختلفة لتحديد أعداد الخلايا ؛ ومع ذلك ، كن على دراية بالنطاق الديناميكي لعداد الخلايا المستخدم. هذا هو عادة بين 1 × 105-1 × 107 خلايا / مل. بينما تم اختبار سرعة الطرد المركزي ومدته لهذا النوع من الخلايا ، قد تتطلب الخلايا المختلفة الأصغر سرعة طرد مركزي أعلى أو أوقات دوران أطول ، ويجب تعديل الطرد المركزي وفقا لذلك. - اجمع الخلايا عن طريق الطرد المركزي عند 300 × جم لمدة 5 دقائق في RT وأعد تعليق الخلايا في DPBS بتركيز نهائي قدره 1 × 106 خلايا / مل. على سبيل المثال ، إذا كان العائد 1 × 107 خلايا ، فأعد تعليق الخلايا في 10 مل من DPBS.
- بالنسبة للخطوة الأولى المتشابكة ، أضف 37٪ فورمالديهايد (FA) إلى تركيز نهائي قدره 1٪ إلى تعليق الخلية ، واحتضان تعليق الخلية لمدة 10 دقائق عند RT مع الدوران (15-20 دورة في الدقيقة) ؛ على سبيل المثال ، إذا كان العائد 1 × 107 خلايا ، أضف 270 ميكرولتر من FA إلى تعليق الخلية 10 مل.
ملاحظة: عادة ما يكون حل FA مستقرا في RT لمدة تصل إلى 3 أشهر بعد الفتح. - إخماد التفاعل عن طريق إضافة 2.5 M الجلايسين إلى تركيز نهائي من 0.25 M واحتضان لمدة 5 دقائق في RT مع دوران (15-20 دورة في الدقيقة). على سبيل المثال ، إذا كان العائد 1 × 107 خلايا ، أضف 1.027 مل من 2.5 متر جليكاين إلى تعليق الخلية.
- اجمع الخلايا عن طريق الطرد المركزي عند 300 × جم لمدة 5 دقائق في RT. تخلص من المادة الطافية وأعد تعليق الخلايا في 5 مل من DPBS. كرر خطوة الطرد المركزي مرة واحدة وأعد تعليق الخلايا المحببة في DPBS إلى 4 × 106 خلايا / مل. على سبيل المثال ، إذا كان العائد 1 × 107 خلايا ، فأعد تعليق الخلايا في 2.5 مل من DPBS.
- بالنسبة للخطوة الثانية المتشابكة ، قم بإعداد محلول مخزون 0.3 متر من سكسينات الإيثيلين جلايكول بيس-سكسينيميديل (EGS) في DMSO (13.6 مجم من EGS في 100 ميكرولتر من DMSO). أضف EGS بتركيز نهائي قدره 3 mM إلى تعليق الخلية ، واحتضانه في RT لمدة 40 دقيقة مع الدوران (15-20 دورة في الدقيقة). على سبيل المثال ، إذا كان العائد 1 × 107 خلايا ، أضف 25 ميكرولتر من محلول مخزون 0.3 M EGS إلى 2.5 مل من تعليق الخلية.
ملاحظة: قم بموازنة EGS إلى RT لمدة 20 دقيقة على الأقل قبل الوزن لتحضير محلول المخزون. - إخماد التفاعل عن طريق إضافة 2.5 M الجلايسين إلى تركيز نهائي من 0.4 M ، واحتضان لمدة 5 دقائق في RT مع دوران (15-20 دورة في الدقيقة). على سبيل المثال ، إذا كان العائد 1 × 107 خلايا ، أضف 400 ميكرولتر من 2.5 M جليكاين إلى تعليق الخلية.
- اجمع الخلايا عن طريق الطرد المركزي عند 1000 × جم لمدة 5 دقائق في RT ، وأعد تعليق الخلايا المحببة في DPBS إلى 5 × 106 خلايا / مل. توزيع 5 × 10 6 خلايا / أنبوب للمكتبات التحضيرية و 1 × 106 خلايا / أنبوب للمعايرة لهضم MNase.
ملاحظة: يقترح معايرة درجة الهضم المثلى لكل دفعة خلية متشابكة. من الناحية المثالية، اجمع حصصتين إلى ثلاث حصص من 1 × 10 6 خلايا لتجارب معايرة MNase (الخطوة 2) واثنين إلى أربعة حصص من 5 × 106 خلايا للتجارب التحضيرية (الخطوة 3). - اجمع الخلايا عن طريق الطرد المركزي عند 1000 × جم لمدة 5 دقائق في RT وقم بإزالة المادة الطافية. قم بتجميد كريات الخلية في النيتروجين السائل ، وقم بتخزينها في -80 درجة مئوية.
ملاحظة: يمكن إعداد مكتبات Micro-C الناجحة من العينات المخزنة لمدة تصل إلى 3 أشهر.
2. معايرة MNase
ملاحظة: يعد إجراء معايرة MNase ضروريا لتحديد التركيز الأمثل ل MNase قبل معالجة المكتبة التحضيرية للخلايا المتشابكة المزدوجة.
- لإجراء معايرة MNase ، قم بإذابة حبيبات واحدة من 1 × 106 خلايا على الجليد لمدة 10 دقائق وأعد تعليق الخلايا في 500 ميكرولتر من DPBS (أضف 1x BSA إذا التصقت الخلايا بالجدار). احتضان تعليق الخلية على الجليد لمدة 20 دقيقة.
- جمع الخلايا عن طريق الطرد المركزي في 10000 × غرام لمدة 5 دقائق في RT ، وإزالة طاف. أعد تعليق الخلايا المحببة في 500 ميكرولتر من المخزن المؤقت MB # 1 (50 mM NaCl ، 10 mM Tris-HCl ، 5 mM MgCl 2 ، 1 mM CaCl 2 ،0.2٪ NP-40 ، 1x مثبط الأنزيم البروتيني [انظر جدول المواد] ، درجة الحموضة 7.4).
- اجمع الخلايا عن طريق الطرد المركزي عند 10000 × جم لمدة 5 دقائق ، وقم بإزالة المادة الطافية. أعد تعليق الخلايا في 200 ميكرولتر من المخزن المؤقت MB # 1 ، وقسم العينة إلى أربعة أنابيب.
- قم بإذابة قارورة واحدة من MNase (20 وحدة / ميكرولتر) ، وقم بتخفيفها باستخدام 10 mM Tris ، الرقم الهيدروجيني 7.4 ، في سلسلة من 1: 2 ، 1: 4 ، 1: 4 ، و 1: 4 لتحقيق تركيزات 10 U / μL و 2.5 U / μL و 0.625 U / μL و 0.1256 U / μL على التوالي (واحد لكل حالة هضم). مع فترات زمنية مناسبة (10-20 ثانية) ، أضف 1 ميكرولتر من محلول MNase إلى إحدى العينات الأربع ، الدوامة ، واحتضانها على خلاط حراري عند 37 درجة مئوية لمدة 10 دقائق (اهتزاز 800 دورة في الدقيقة). استمر في إضافة 1 ميكرولتر من تخفيفات MNase المتبقية إلى حصص الخلايا المتبقية.
- أوقف عملية الهضم Mnase بإضافة 200 ميكرولتر من محلول STOP المحضر حديثا (150 ميكرولتر من 10 مللي متر Tris ، درجة الحموضة 7.4 ، 25 ميكرولتر من 10٪ SDS ، 25 ميكرولتر من 20 مجم / مل بروتيناز K ، 2 ميكرولتر من 0.5 M EGTA) إلى كل أنبوب بنفس الترتيب وبنفس الفترة الزمنية التي تمت فيها إضافة MNase. احتضان في 65 درجة مئوية لمدة 2 ساعة.
- أضف 500 ميكرولتر من كحول الفينول كلوروفورم إيزو أميل (PCI) إلى كل عينة ، واخلطها جيدا عن طريق الدوامة. جهاز طرد مركزي عند 19800 × جم لمدة 5 دقائق في RT لفصل المراحل ، ونقل الطور المائي إلى أنابيب جديدة (حوالي 200 ميكرولتر / عينة).
تنبيه: يحتوي PCI على العديد من المكونات السامة ويجب التعامل معه فقط في غطاء أمان كيميائي. يرجى استشارة الشركة المصنعة لمزيد من التفاصيل. - تنقية الحمض النووي باستخدام مجموعة تنقية الحمض النووي التجارية (انظر جدول المواد) وفقا لتعليمات الشركة المصنعة وتخفيف العينات في 12 ميكرولتر من المخزن المؤقت للشطف.
ملاحظة: تركيز SDS من خطوة إزالة البروتين (الخطوة 2.5) مثبط لبعض مجموعات تنقية الحمض النووي. تعمل مجموعة تنقية الحمض النووي المستخدمة هنا بشكل مماثل لترسيب الإيثانول. - أضف 2-5 ميكرولتر من صبغة التحميل ، وقم بتشغيل العينات على هلام أغاروز 1.5٪ عند 120 فولت لمدة 30-50 دقيقة (حتى يتم فصلها بشكل صحيح ؛ الشكل 1 أ).
- اختر أفضل درجة هضم للتجربة، واستمر في عملية الهضم التحضيرية ل MNase. تظهر درجة الهضم المثلى شظايا تحت النوكليوسومية قليلة أو معدومة ونسبة 70٪ -90٪ أحادية إلى ثنائية النوكليوسوم.
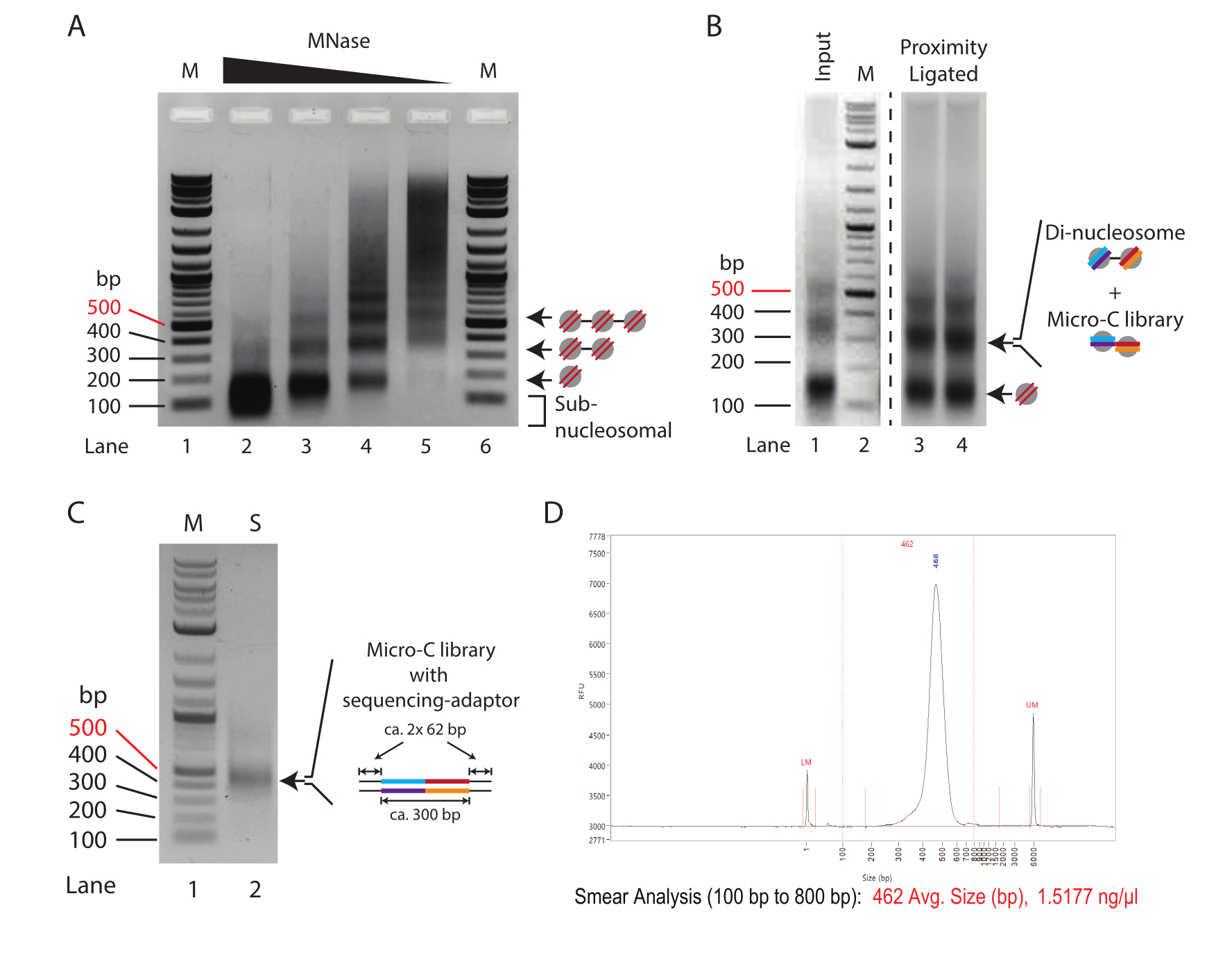
ملاحظة: في هذه التجربة ، تم تحديد درجة الهضم في Lane 4 في الشكل 1A لتكون عملية الهضم المثلى التي تم الحصول عليها باستخدام 0.625-1.25 U من Mnase ل 2.5 × 10 5 خلايا (= 2.5-5 U من Mnase ل 1 × 106 خلايا). للاطلاع على مناقشة مفصلة، انظر النتائج التمثيلية.
3. هضم MNase التحضيري
- لتجزئة الكروماتين إلى النوكليوسومات الأحادية عن طريق هضم MNase ، قم بإذابة قسمة الخلايا المزدوجة المتشابكة سابقا 5 × 106 ، وأعد التعليق في 1 مل من DPBS. احتضان على الجليد لمدة 20 دقيقة (أضف 1x BSA إذا التصقت الخلايا بجدار الأنبوب).
- اجمع الخلايا عن طريق الطرد المركزي عند 10000 × جم لمدة 5 دقائق في RT ، وتخلص من المادة الطافية. أعد تعليق الحبيبات في 500 ميكرولتر من المخزن المؤقت MB # 1. كرر خطوة الطرد المركزي مرة واحدة. أعد تعليق الحبيبات في 1 مل من المخزن المؤقت MB # 1 ، وقم بعمل 200 ميكرولتر من القسمة (1 × 106 خلايا لكل قسامة).
- استنادا إلى معايرة MNase (الموصوفة في الخطوة 2) ، قم بهضم الكروماتين عن طريق إضافة الكمية المناسبة من MNase (عادة 2.5-10 وحدة / ميكرولتر لكل 1 × 106 خلايا) إلى كل قسمة . تخلط جيدا (دوامة وتدور بسرعة) ، واحتضانها على خلاط حراري عند 37 درجة مئوية لمدة 10 دقائق مع اهتزاز 800 دورة في الدقيقة.
- أوقف هضم MNase بإضافة 1.6 ميكرولتر من 0.5 M EGTA (4 mM النهائية) إلى كل حصة ، واحتضانها على خلاط حراري عند 65 درجة مئوية لمدة 10 دقائق مع اهتزاز 800 دورة في الدقيقة.
- اجمع العينة عن طريق الطرد المركزي عند 10000 × جم لمدة 5 دقائق في RT ، وتخلص من المادة الطافية. أعد تعليق حبيبات الخلية في 500 ميكرولتر من 1x NEBuffer 2.1.
- تجميع عينات تعادل إدخال 5 × 106 خلايا أو أقل لمزيد من المعالجة.
ملاحظة: إذا كان سيتم معالجة أكثر من 5 × 10 6 خلايا ، فقم بمعالجة هذه العينات بالتوازي ، حيث يتم تحسين الظروف الأنزيمية لخلايا 5 × 106. - قبل الشروع في خطوات ربط القرب ، انقل 10٪ من العينة كعنصر تحكم في الإدخال للتحكم في مستوى هضم MNase. أضف 150 ميكرولتر من 10 mM Tris ، ودرجة الحموضة 7.4 ، و 25 ميكرولتر من 10٪ SDS ، و 25 ميكرولتر من 20 مجم / مل من البروتيناز K إلى هذه العينة ، واحتضانها طوال الليل عند 65 درجة مئوية.
4. معالجة نهاية الحمض النووي وربط القرب
- اجمع العينة المتبقية عن طريق الطرد المركزي عند 10000 × جم لمدة 5 دقائق عند 4 درجات مئوية ، وتخلص من المادة الطافية. أعد تعليق الحبيبات في 90 ميكرولتر من المزيج الرئيسي Micro-C المحضر حديثا 1 (الجدول 1) ، واحتضانها على خلاط حراري لمدة 15 دقيقة عند 37 درجة مئوية مع اهتزاز 800 دورة في الدقيقة.
- أضف 10 ميكرولتر من 5 وحدة / ميكرولتر من جزء Klenow ، واحتضنها على خلاط حراري لمدة 15 دقيقة عند 37 درجة مئوية مع اهتزاز 800 دورة في الدقيقة.
- أضف 100 ميكرولتر من Micro-C master mix 2 المحضر حديثا (الجدول 2) ، واحتضنه على خلاط حراري لمدة 45 دقيقة عند 25 درجة مئوية مع اهتزاز 800 دورة في الدقيقة. بعد الحضانة ، قم بإخماد التفاعل الأنزيمي بإضافة EDTA إلى تركيز نهائي يبلغ 30 mM. احتضان على خلاط حراري لمدة 20 دقيقة عند 65 درجة مئوية مع اهتزاز 800 دورة في الدقيقة.
- اجمع العينة عن طريق الطرد المركزي عند 10000 × جم لمدة 5 دقائق عند 4 درجات مئوية ، وتخلص من المادة الطافية. أعد تعليق العينة في 500 ميكرولتر من المزيج الرئيسي Micro-C 3 المحضر حديثا (الجدول 3) ، واحتضانها لمدة 2.5 ساعة عند RT مع الدوران (15-20 دورة في الدقيقة).
- اجمع العينة عن طريق الطرد المركزي عند 10000 × جم لمدة 5 دقائق عند 4 درجات مئوية ، وتخلص من المادة الطافية. أعد تعليق العينة في 200 ميكرولتر من المزيج الرئيسي Micro-C 4 المحضر حديثا (الجدول 4) ، واحتضانها على خلاط حراري لمدة 15 دقيقة عند 37 درجة مئوية مع اهتزاز 800 دورة في الدقيقة.
- للتشابك العكسي وإزالة البروتين، أضف 25 ميكرولتر من 20 ملغم/مل بروتيناز K و25 ميكرولتر من 10٪ SDS إلى العينة، واحتضانها عند 65 درجة مئوية طوال الليل مع الخلط المتقطع.
5. تنقية الحمض النووي ثنائي النواة واختيار الحجم
- أضف 500 ميكرولتر من PCI إلى العينات والتحكم في الإدخال ، واخلطها عن طريق الدوامة. افصل المراحل عن طريق الطرد المركزي عند 19800 × جم لمدة 5 دقائق ، وانقل المرحلة المائية العليا إلى أنبوب جديد.
- ركز الحمض النووي إما باستخدام مجموعة تنقية الحمض النووي أو عن طريق ترسيب الإيثانول. قم بسحب العينات في 30 ميكرولتر (الخطوة 5.3) وعناصر التحكم في الإدخال (الخطوة 3.11) في 15 ميكرولتر ، وقم بتشغيل هلام أغاروز بنسبة 1.5٪ لفصل النوكليوسومات الأحادية والنيوكليوسومات الثنائية (الشكل 1 ب).
ملاحظة: تركيز SDS من خطوة إزالة البروتين مثبط لبعض مجموعات تنقية الحمض النووي. تعمل مجموعة تنقية الحمض النووي المستخدمة هنا (انظر جدول المواد) بشكل مماثل لترسيب الإيثانول. اعتمادا على عدد الخلايا المستخدمة ، يمكن أن يتراوح الإدخال من 100 نانوغرام إلى 10 ميكروغرام. عادة ، يتم استخراج 1-5 ميكروغرام من الحمض النووي من 5 × 106 خلايا. - استئصال شظايا الحمض النووي التي لها حجم ثنائي النوكليوسومال (حوالي 300 نقطة أساس). استخدم مجموعة شطف هلام الحمض النووي المتاحة تجاريا (انظر جدول المواد) لاستخراج الحمض النووي من هلام الأغاروز ، و elute في 150 ميكرولتر.
6. إعداد حبات الستربتافيدين
- نقل 10 ميكرولتر من حبات الستربتافيدين (انظر جدول المواد) لكل عينة إلى أنبوب تفاعل.
- ضعه في مغناطيس مناسب لأنابيب سعة 1.5 مل (انظر جدول المواد). بعد مسح المحلول (1-2 دقيقة) ، قم بإزالة المادة الطافية ، وأعد تعليق الخرزات في 300 ميكرولتر من 1x TBW (5 mM Tris-HCl ، درجة الحموضة 7.5 ، 0.5 mM EDTA ، 1 M NaCl ، 0.05٪ Tween 20) لكل عينة. كرر هذه الخطوة مرة واحدة.
- أعد تعليق الخرزات في 150 ميكرولتر من المخزن المؤقت 2x B&W (10 mM Tris-HCl ، الرقم الهيدروجيني 7.5 ، 1 mM EDTA ، 2 M NaCl) لكل عينة تمت معالجتها في الخطوة 7.
7. الستربتافيدين المنسدلة وإعداد مكتبة الخرز
- أضف 150 ميكرولتر من الخرز المعد مسبقا (الخطوة 6.3) إلى 150 ميكرولتر من العينة (الخطوة 5.3). احتضان لمدة 20 دقيقة في RT مع الدوران (15-20 دورة في الدقيقة).
- ضع الأنابيب في مغناطيس مناسب ، وانتظر حتى يزول المحلول (1-2 دقيقة). قم بإزالة المادة الطافية وأعد تعليق الخرزات في 300 ميكرولتر من 1x TBW. كرر هذه الخطوة.
- ضع الأنابيب في مغناطيس مناسب وانتظر حتى يزول المحلول (1-2 دقيقة). قم بإزالة المادة الطافية وأعد تعليق الحبيبات في 100 ميكرولتر من 0.1x TE (1 mM Tris ، 0.1 mM EDTA ، درجة الحموضة 8.0).
- ضع الأنابيب في مغناطيس مناسب وانتظر حتى يزول المحلول (1-2 دقيقة). قم بإزالة المادة الطافية ، وأعد تعليق الخرز في 50 ميكرولتر من 0.1x TE ، وانقلها إلى أنابيب PCR.
ملاحظة: يتوافق حجم 0.1x TE المستخدم (50 ميكرولتر) مع حجم الإدخال لمجموعة إعداد مكتبة تسلسل الحمض النووي (انظر جدول المواد) المستخدمة في هذا البروتوكول. إذا تم استخدام مجموعة أو استراتيجية مختلفة ، فاضبط مستوى الصوت وفقا لذلك. - قم بإجراء خطوات معالجة الحمض النووي لمجموعة إعداد مكتبة التسلسل وفقا لبروتوكول الشركة المصنعة. تتضمن هذه الخطوات عادة تقشير الحمض النووي ، و A-tailing ، وربط المحول ، واستئصال U. الخطوة الأخيرة (U excision) خاصة بالمجموعة المستخدمة في هذه الدراسة. في حالة استخدام هذه المجموعة ، اتبع تعليمات الشركة المصنعة من الخطوة 1 إلى الخطوة 2.6 في بروتوكول المجموعة باستخدام محولات غير مخففة ، ولا تفكر في التخزين عند -20 درجة مئوية بعد الخطوة 2.6.
ملاحظة: يجب استبدال تغييرات المخزن المؤقت التي تتطلبها مجموعات التسلسل عن طريق ربط الخرزات بالمغناطيس والغسل (الخطوات من 7.3 إلى 7.4) ، حيث لا يزال الحمض النووي مرتبطا بحبات الستربتافدين. بالإضافة إلى ذلك ، نظرا لأن الحمض النووي مرتبط بالخرز المغناطيسي ، فتجاهل أي خطوات تنقية واختيار الحجم بعد ربط المحول من بروتوكول المجموعة. تابع إلى الخطوة 7.6 (هذا البروتوكول). - بعد اكتمال ربط المحول، اغسل العينة كما هو موضح في الخطوة 7.2. تخلص من المادة الطافية وأعد تعليق الخرزات في 20 ميكرولتر من 0.1x TE.
8. تقدير دورات PCR المطلوبة
ملاحظة: ينصح بتقدير دورات تفاعل البوليميراز المتسلسل المطلوبة لتضخيم المكتبة. عادة ، تتطلب مكتبة Micro-C 8-15 دورة من تفاعل البوليميراز المتسلسل. على الرغم من أن الخطوة ليست ضرورية ، إلا أنها تساعد على تجنب الإفراط في التضخيم وتقلل من خطر تكرار تفاعل البوليميراز المتسلسل.
- لتحديد الحد الأدنى لعدد دورات تفاعل البوليميراز المتسلسل المطلوبة ، قم بإجراء تفاعل البوليميراز المتسلسل مع 1 ميكرولتر من عينة ستربتافيدين - بيوتين - الحمض النووي (الخطوة 7.6). للقيام بذلك ، أضف مزيج PCR الرئيسي (3.2 ميكرولتر من H2O ، 0.4 ميكرولتر من i5 التمهيدي ، 0.4 ميكرولتر من i7 التمهيدي ، و 5 ميكرولتر من بوليميراز الحمض النووي عالي الدقة Q5) إلى العينة. قم بإجراء تفاعل البوليميراز المتسلسل مع 16 دورة وفقا لتعليمات الشركة المصنعة لمجموعة تحضير مكتبة الحمض النووي المستخدمة.
ملاحظة: يتم شراء البادئات المستخدمة هنا في مجموعة منفصلة (انظر جدول المواد) يجب أن تكون متوافقة مع مجموعة إعداد المكتبة المستخدمة. - بعد تفاعل البوليميراز المتسلسل ، اجمع الخرزات بمغناطيس مناسب ، وقم بقياس تركيز الحمض النووي البالغ 1 ميكرولتر من المادة الطافية باستخدام أداة تقدير كمية الحمض النووي عالية الحساسية (انظر جدول المواد). للحصول على إجمالي تركيز الحمض النووي ، اضرب هذه القيمة في 10 (الحجم الكلي لتفاعل PCR). تقدير العدد المطلوب من دورات تفاعل البوليميراز المتسلسل كما هو موضح في النتائج التمثيلية.
- أضف 2 ميكرولتر من مخزن التحميل المؤقت 6x إلى 9 ميكرولتر المتبقية من خليط تفاعل البوليميراز المتسلسل. قم بحل مكتبة PCR المضخمة على هلام أغاروز 1٪ لتحديد ربط المحول الناجح ، مما ينتج عنه حجم 450 نقطة أساس تقريبا (الشكل 1C).
ملاحظة: يجب أن يظهر الجل شريطا مميزا يبلغ حوالي 420 نقطة أساس (مكتبة Micro-C بالإضافة إلى محولات). تمثل نطاقات 120 bp دايمرات المحول وتدل على مكتبات منخفضة التعقيد. قد تظهر نطاقات ذات أوزان جزيئية أقل (أقل من 100 نقطة أساس) ، وهذه مواد أولية غير مستخدمة من تفاعل تفاعل تفاعل البوليميراز المتسلسل.
9. تسلسل تضخيم المكتبة
- أضف مزيج PCR الرئيسي (65 ميكرولتر من H2O ، 100 ميكرولتر من بوليميراز الحمض النووي عالي الدقة Q5 ، 8 ميكرولتر من i5 التمهيدي ، 8 ميكرولتر من i7 التمهيدي) إلى 19 ميكرولتر المتبقية من عينة الحمض النووي للستربتافيدين والبيوتين. قسم خليط التفاعل إلى 50-100 ميكرولتر من القسامة.
ملاحظة: يعتمد الحجم الأمثل ل PCR على جهاز PCR ؛ عادة ، يوفر 50 ميكرولتر التضخيم الأكثر قابلية للتكرار في أجهزة PCR الشائعة. - قم بإجراء تفاعل البوليميراز المتسلسل وفقا لتعليمات الشركة المصنعة لمجموعة أدوات إعداد مكتبة الحمض النووي مع عدد الدورات المحددة في الخطوة 8. إذا تم حذف الخطوة 8 ، يوصى باستخدام 14 دورة ل PCR.
- تنقية الحمض النووي مع حبات البارامغناطيسية (انظر جدول المواد) بنسبة 1: 0.9 وفقا لبروتوكول الشركة المصنعة. Elute في 20 ميكرولتر من 0.1 ٪ TE.
ملاحظة: إذا تم اكتشاف دايمرات المحول في الخطوة 8.5 ، فقم بإجراء التنقية مرتين باستخدام نفس النسبة. - تحديد تركيز الحمض النووي وتشغيل العينات على نظام مراقبة الجودة (انظر جدول المواد). تأكد من الجودة الجيدة لمكتبة Micro-C من خلال وجود نطاق دقيق واحد (الشكل 1D).
ملاحظة: دايمرات المحول غير شائعة في تحضيرات المكتبة على الخرز بسبب غسل العينة قبل تفاعل البوليميراز المتسلسل. وبالتالي ، فإن ظهور دايمرات المحول يشير إلى انخفاض تعقيد المكتبة. إذا لوحظت ثنائيات المحول ، يوصى بشدة بمراقبة جودة العينة مع تسلسل مدخلات منخفض.
10. تسلسل الحمض النووي ومعالجة البيانات
- تسلسل مكتبة Micro-C مع تسلسل الطرف المزدوج وفقا لمتطلبات موفر التسلسل.
ملاحظة: من الناحية المثالية ، يتم تسلسل العينات على منصة في وضع النهاية المزدوجة مع 50 نقطة أساس لكل قراءة. كما تم استخدام الأنظمة الأساسية القديمة التي تقدم أطوال قراءة أقصر ، مثل 2 × 35 نقطة أساس ، بنجاح. الأهم من ذلك ، إذا تمت دراسة المناطق الجينومية المتكررة ، فقد يكون من المستحسن التسلسل بطول قراءة أطول. - لتقييم جودة مكتبة Micro-C ، قم بإجراء تسلسل إدخال منخفض مع 5 × 106 إلى 1 × 107 قراءات لكل عينة.
- معالجة ملفات التسلسل (ملفات fastq) باستخدام Distiller10. قم بتعيين القراءات مقابل الجينوم المرجعي المناسب ، هنا mm10.
ملاحظة: يمكن معالجة ملفات التسلسل باستخدام خطوط أنابيب مختلفة على أجهزة الكمبيوتر المحلية أو مجموعات الحوسبة. بالنسبة للعينات ذات أعماق التسلسل المنخفضة ، يمكن أن تقلل أحجام الحاويات الأكبر ، مثل 10000 نقطة أساس و 50000 نقطة أساس و 100000 نقطة أساس و 500000 نقطة أساس ، من متطلبات الحوسبة وأحجام الملفات. يقوم Distiller (المستخدم في هذه الدراسة) بإنشاء جميع أنواع الملفات المطلوبة لتقييم جودة مكتبة Micro-C. يحتوي ملف * .stats الذي تم إنشاؤه على معلومات حول معدل الخريطة ونسب cis-trans واتجاه القراءة الطبقي حسب المسافة بين أزواج القراءة. يتم تصور هذه المعلمات في الشكل 2 ، وتتم مناقشة تقييم جودة مكتبة Micro-C في النتائج التمثيلية. يقوم برنامج المعالجة أيضا بإنشاء ملفات mcool التي يمكن تحميلها مباشرة في HiGlass (https://docs.higlass.io/) لإنشاء مصفوفات التفاعل9.
النتائج
يمكن تقييم الإعداد الناجح لمكتبات Micro-C في عدة خطوات من البروتوكول. الخطوة الأكثر أهمية هي اختيار درجة هضم MNase المناسبة. لذلك ، يجب معايرة تركيز MNase لإنتاج 70٪ -90٪ من النوكليوسومات الأحادية باستمرار على ثنائي النيوكليوسومات لكل عينة. من المهم ملاحظة أن هضم الكروماتين يختلف بالنسبة للكروماتين eu وغير المتجانس ، حيث يهضم MNase heterochromatin بشكل أقل كفاءة. وبالتالي ، فإن درجة الهضم المثلى تعتمد على منطقة الكروماتين ذات الاهتمام ونوع الخلية التي تمت دراستها لأن النسبة النسبية للكروماتين eu وغير المتجانس هي نوع الخلية المحددة. لذلك ، ينصح بمعايرة تركيز MNase المطلوب بعناية وتقييم نجاح تجربة Micro-C أولا عن طريق تسلسل منخفض المدخلات.
يوضح الشكل 1 أ نمط معايرة MNase نموذجي للكروماتين المعالج بكميات متناقصة من MNase. هنا ، يتم هضم الكروماتين من 250000 خلية لكل تفاعل بتخفيف أربعة أضعاف من MNase. أعلى تركيز (10 U من MNase ، Lane 2) يظهر الكروماتين المهضوم بشكل حصري تقريبا يتكون من الحمض النووي أحادي النواة (~ 150 bp). والجدير بالذكر أن مركز النطاق أحادي النواة ينخفض في هلام الأغاروز مقارنة بالعصابات المقابلة في العينات ذات تركيزات MNase المنخفضة ، مما يشير إلى الإفراط في هضم الحمض النووي النووي. يتم ربط النيوكليوسومات المفرطة الهضم بشكل غير فعال في تفاعل ربط القرب. لذلك ، فإن العينة في الحارة 2 دون المستوى الأمثل لتجارب Micro-C. يعرض Lane 3 (2.5 U من MNase) درجة هضم مناسبة تقريبا لتجارب Micro-C. هنا ، يكون النطاق أحادي النواة هو النوع السائد ، ويتم تقليل اللطاخة تحت النوكليوسومية ، التي تدل على النيوكليوسومات المفرطة الهضم ؛ ومع ذلك ، فإنه لا يزال موجودا. تعد درجة الهضم في الحارة 4 (0.635 U من MNase) حالة مثالية لتجربة Micro-C في مثال المعايرة بالتحليل الحجمي هذا. يوجد شريط أحادي النواة واضح بدون الحمض النووي دون النووي. كثافة النطاق للحمض النووي أحادي وثنائي النوكليوسوم متساوية تقريبا ، مما يشير إلى عائد أحادي النوكليوسوم بنسبة 66٪ أو أعلى. تجدر الإشارة إلى أن الحمض النووي ثنائي النواة هو ما يقرب من ضعف حجم الحمض النووي أحادي النواة (~ 320 bp مقابل ~ 150 bp) ، لذا فإن شدة نطاقه لكل مول من الحمض النووي أعلى مرتين مقارنة بنظيره أحادي النواة. تظهر درجة الهضم في الحارة 5 (0.156 U من MNase) كروماتين غير مهضوم مع عدم وجود حمض نووي نووي تقريبا ، وبالتالي يمثل هذا عينة دون المستوى الأمثل.
في الختام ، في هذا المثال ، فإن هضم 2.5 × 10 5 خلايا ES للفأر مع 0.625 U من MNase (المقابلة ل2.5 U من MNase ل 1 × 106 خلايا في 200 ميكرولتر) يوفر نقطة البداية الواعدة لعمليات الهضم التحضيرية في تجارب Micro-C. ومع ذلك ، ينبغي أيضا مراعاة تركيز MNase الوسيط بين الظروف المستخدمة للعينات في Lane 3 و Lane 4 (المقابلة ل 5 U من MNase لخلايا 1 × 106 في 200 ميكرولتر). الأهم من ذلك ، لا يمكن تحجيم هضم الكروماتين مع MNase خطيا ، ولا ينصح برفع مستوى الهضم التحضيري أكثر من 4x. لإعداد مكتبات Micro-C من أكثر من 1 × 10 6 خلايا ، ينصح بهضم الكروماتين في قسمة من 1 × 106 خلايا وتجميعها بعد تعطيل MNase.
لتقييم نجاح بروتوكول ربط القرب ، يجب مقارنة عنصر التحكم في الإدخال ، وهو مهضوم MNase وليس مرتبطا بالقرب (الخطوة 3.8) ، بالعينة المرتبطة بالقرب (الخطوة 5.3) بنسبة 1.5٪ من هلام الأغاروز الكهربائي (الشكل 1 ب). يبلغ حجم نطاق النوكليوسوم الأحادي المرتبط بالقرب 300 bp تقريبا ، على غرار حجم ثنائي النيوكليوسومات. لذلك ، يجب أن تتحول نسبة إشارة النطاق أحادي إلى ثنائي النواة من النوكليوسومات الأحادية في الغالب (الحارة 1) إلى النوكليوسومات الثنائية (الحارة 3 والحارة 4). نظرا لأن هلام الأغاروز في هذه الخطوة هو الحمض النووي ثنائي النواة الذي يتم استئصاله وتنقيته ، فمن المستحسن تقسيم العينات إلى ممرات متعددة لتجنب التحميل الزائد.
يوصى بتقييم جودة وكمية مكتبة التسلسل المعدة بأقل قدر من تفاعل البوليميراز المتسلسل. هنا ، يتم تضخيم الحمض النووي من 1 ميكرولتر من الخرز (1/20 من العينة الكلية) لمدة 16 دورة في 10 ميكرولتر من تفاعل تفاعل البوليميراز المتسلسل. يتراوح التركيز الكلي لمكتبة الحد الأدنى من تفاعل البوليميراز المتسلسل عادة من 50-500 نانوغرام بعد 16 دورة تفاعل البوليميراز المتسلسل. من الناحية النظرية ، يتوافق هذا مع مكتبة 1-10 ميكروغرام من عينة 19 ميكرولتر المتبقية إذا تم تضخيمها أيضا لمدة 16 دورة. يوصى باستخدام الحد الأدنى لعدد دورات تفاعل البوليميراز المتسلسل اللازمة لإنشاء مكتبة تبلغ حوالي 100 نانوغرام من إجمالي الحمض النووي. بافتراض التضخيم اللوغاريتمي في تفاعل البوليميراز المتسلسل ، يمكن تقسيم التركيز النظري للحمض النووي الذي تم الحصول عليه من مدخلات 19 ميكرولتر في 16 دورة على التوالي على اثنين لحساب عدد دورات تفاعل البوليميراز المتسلسل المطلوبة لإنشاء مكتبة 100 نانوغرام. على سبيل المثال ، العائد 100 نانوغرام من 1 ميكرولتر بعد 16 دورة يتوافق مع عائد 1,900 نانوغرام تضخيم من 19 ميكرولتر. في هذا السيناريو ، يجب أن تولد 12 دورة بشكل مثالي مكتبة تسلسل 118 نانوغرام من إجمالي الحمض النووي (1,900 نانوغرام / [2 × 2 × 2 × 2] = 118 نانوغرام). يمكن بعد ذلك استخدام العينة المتبقية 9 ميكرولتر من الحد الأدنى من تفاعل البوليميراز المتسلسل لتقييم جودة المكتبة عن طريق الرحلان الكهربائي لهلام الأغاروز (الشكل 1C). يجب أن يظهر التصور نطاقا مميزا واحدا عند 420 نقطة أساس ولا توجد نطاقات لثنائيات المحول (120 نقطة أساس). قد تظهر أيضا شظايا أصغر ، وهذه تتوافق مع بادئات PCR غير المستخدمة.
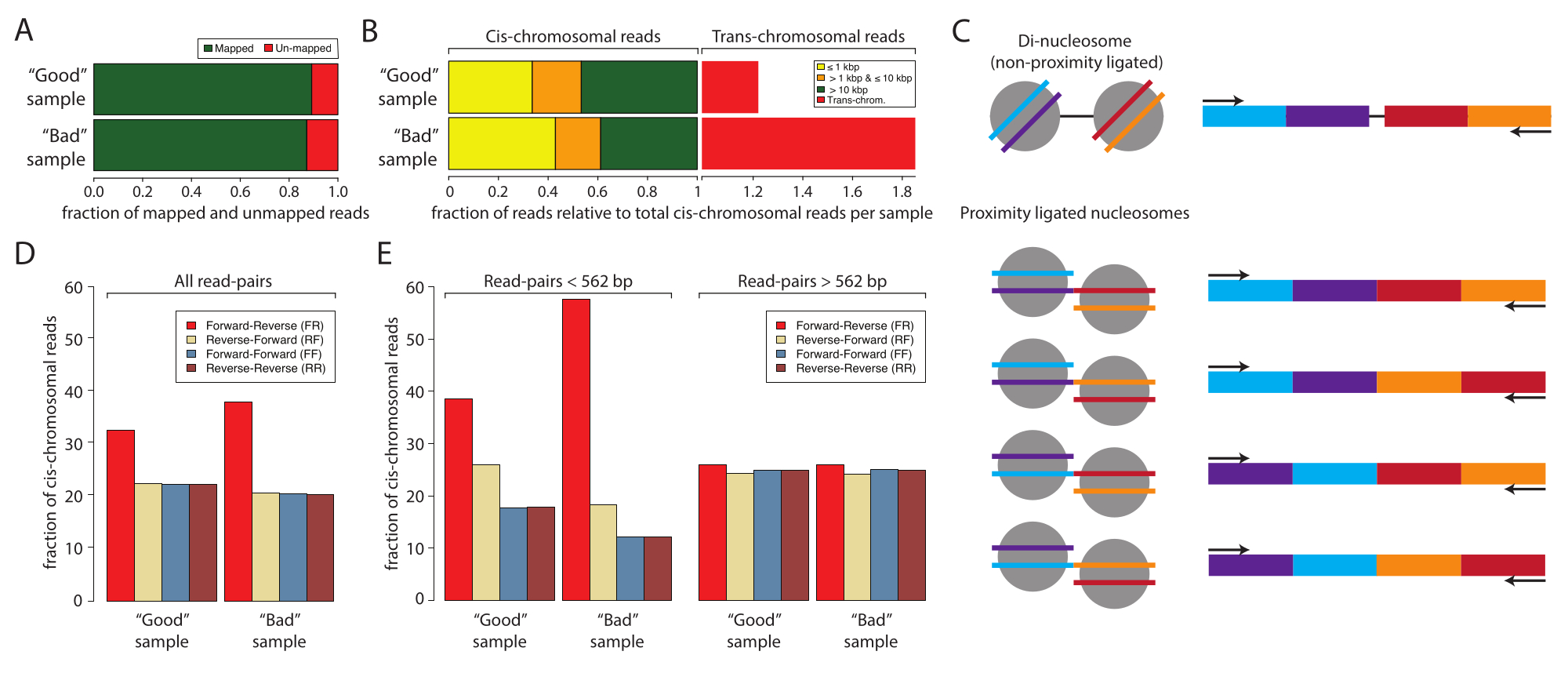
بعد ذلك ، يوصى بتحليل وتأكيد التحضير الناجح لعينة Micro-C عن طريق تسلسل منخفض المدخلات قبل الالتزام بالتسلسل العميق كثيف الموارد. عادة ، يتم تسلسل المكتبات إلى عمق قراءة من 5 × 106 إلى 1 × 107 ويتم تقييمها بناء على المعايير التالية: معدل تكرار قراءة التسلسل ، ومعدل تفاعل رابطة الدول المستقلة مقابل الكروموسومات العابرة ، وتردد اتجاه قراءة التسلسل. تتم معالجة مكتبات Micro-C باستخدام Distiller ، وهو خط أنابيب كامل الخدمات يعالج البيانات من تسلسل ملفات القراءة (تنسيق Fastq) إلى ملفات أزواج القراءة (تنسيق Bedpe) ومصفوفات التفاعل القابلة للتطوير (تنسيقات Cool و Mcool) باستخدام cooler و pairtools و cooltools10،11،12. يقوم خط الأنابيب أيضا بإنشاء ملف ملخص مثالي لتقييم جودة مكتبات Micro-C10 (https://github.com/open2c/distiller-nf). يوفر معدل تكرار PCR معلومات حول تعقيد مكتبة التسلسل ويمكن استخراجه من ملف * .stats الذي تم إنشاؤه. تحتوي مكتبات Micro-C عالية الجودة على معدلات تكرار PCR أقل من 5٪ -10٪ عند إنشائها من 5 ملايين خلية أو أكثر. والجدير بالذكر أن بعض منصات التسلسل تولد تكرارات PCR أثناء تكوين الكتلة بشكل مستقل عن تعقيد مكتبة التسلسل. يوضح الشكل 2 أ معدلات الازدواجية النسبية لتجربتين: واحدة نعتبرها عينة جيدة وعينة سيئة. في هذا المثال، عرضت كلتا العينتين معدلات خريطة مقبولة. المعايير التالية لتقييم جودة مكتبات Micro-C هي نسبة رابطة الدول المستقلة مقابل الترانس وترددات اتجاه القراءة. داخل النواة ، تعيش الكروموسومات في مناطق كروموسوم فردية ، وبالتالي نادرا ما تتفاعل مع الكروموسومات الأخرى. يشير المعدل المرتفع للتفاعلات عبر الكروموسومات المكتشفة إلى ارتفاع معدل الربط العشوائي. وتجدر الإشارة إلى أنه في هذا المستوى من التحليل ، أظهرت العينة السيئة معدلا مرتفعا من التفاعلات عبر الكروموسومات مقارنة بالعينة الجيدة (الشكل 2 ب). بالنسبة إلى Micro-C ، من المستحسن الحصول على معدل تفاعل كروموسومي بنسبة 70٪ أو أعلى.
تحتوي مكتبة Micro-C على حجم جزء مشابه لشريط الحمض النووي ثنائي النواة ، والذي يمكن أن ينقي بشكل مشترك مع العينة المرتبطة القريبة ويلوث التجربة. هذه الملوثات هي دائما تفاعلات كروموسومية رابطة الدول المستقلة. لذلك ، من المهم أيضا تقييم معدلات اتجاه القراءة. يمكن تقدير معدل التلوث ثنائي النيوكليوسومال عن طريق تسلسل المدخلات المنخفضة. ينبع الحمض النووي ثنائي النواة من نيوكليوسومات متجاورة لم يتم شقها بواسطة MNase. وبالتالي ، فإن قراءات التسلسل الناتجة ستعرض دائما اتجاه قراءة أمامي عكسي (F و R) ، وستكون المسافة بين أزواج القراءة حوالي 320 نقطة أساس. بالمقارنة ، يمكن ربط الأجزاء المرتبطة بالقرب في أربعة اتجاهات ، مما ينتج عنه أزواج قراءة مع F-R و R-R و R-F و F-F ، من الناحية المثالية بوفرة متساوية (الشكل 2C). بالإضافة إلى ذلك ، فإنها تعرض مسافات مختلفة بين أزواج القراءة. لتقدير كمية الملوثات ثنائية النيوكليوسومات ، يمكن حساب تواتر اتجاهات القراءة من ملفات * stats التي تم إنشاؤها بواسطة التقطير (الشكل 2D). والجدير بالذكر أنه في هذا العمل ، كان جزء قراءات F-R (الأحمر) أعلى في العينة السيئة مقارنة بالعينة الجيدة ، وأصبح هذا أكثر وضوحا عندما تم تقسيم اتجاهات القراءة إلى طبقات حسب المسافة (الشكل 2E). تهيمن شظايا ثنائية النيوكليوسومال على جزء F-R مقارنة بمكتبات Micro-C عندما يتم تقسيم أزواج القراءة إلى قراءات بمسافات <562 bp أو ≥562 bp. هنا ، تهيمن قراءات F-R على جزء القراءات بمسافة <562 نقطة أساس ، في حين أن الكسر ذو المسافات ≥562 نقطة أساس يعرض توزيعا متساويا بين الاتجاهات الأربعة المحتملة ، مما يشير إلى أن التمثيل المفرط العالمي لقراءات F-R ينبع من الملوثات ثنائية النيوكليوسومال. يتم تحديد اختيار 562 نقطة أساس كحد أدنى للإعداد الفرعي بواسطة binning في ملف * stats الذي تم إنشاؤه. على الرغم من أنه ليس ضروريا لمراقبة الجودة هذه ، إلا أنه يمكن تحقيق إعداد فرعي أكثر تحديدا عن طريق استخراج المسافات من ملف * pairs ، والذي يتم إنشاؤه أيضا بواسطة جهاز التقطير. من المهم ملاحظة أن قراءات ثنائي النيوكليوسومال لا تقلل من جودة عينة Micro-C حيث يمكن تحديدها وتجاهلها أثناء معالجة البيانات. ومع ذلك ، فهي لا تحتوي على معلومات قيمة حول تفاعلات 3D ، وتخفف من القراءات الإعلامية.
وبالتالي ، فإن معايرة MNase الدقيقة ومراقبة الجودة الشاملة مع تسلسل المدخلات المنخفضة هي أفضل الأدوات لتحسين جودة تجارب Micro-C.

الشكل 1: المراحل المتوسطة لبروتوكول Micro-C . (أ) الرحلان الكهربائي لهلام الأغاروز للكروماتين من 2.5 × 105 خلايا ES فأر مهضومة بتركيزات مختلفة من MNase. يشار إلى النطاقات أحادية وثنائية وثلاثية النواة بالأسهم. M: سلم الحمض النووي (حارة 1/6) ؛ 10 U من MNase لكل 250000 خلية (Lane 2) ؛ 2.5 U من MNase لكل 250000 خلية (الحارة 3) ؛ 0.625 U من MNase لكل 250000 خلية (Lane 4) ؛ 0.156 U من MNase لكل 250000 خلية (Lane 2). (ب) الرحلان الكهربائي لهلام الأغاروز بنسبة 1.0٪ للعينات المحضرة من Micro-C (Lane 3 و Lane 4) والتحكم في الإدخال المهضوم MNase (Lane 1). تم تحسين الحارة 1 والحارة 2 (M: سلم الحمض النووي) للتأكيد على التغير النسبي في شدة الشظايا أحادية إلى ثنائية النواة. يشار إلى النطاقات أحادية وثنائية النواة بالأسهم. يجمع النطاق ثنائي النوكليوسومال في العينة المرتبطة بالقرب بين الحمض النووي لمكتبة ثنائي النوكليوسومال ومكتبة Micro-C. (C) تم تضخيم الرحلان الكهربائي لهلام الأغاروز بنسبة 1.0٪ لمكتبات تسلسل Micro-C من عينة 1 ميكرولتر لتقييم الجودة. الحارة 1 (M): سلم الحمض النووي. الممر 2 (S): مكتبة Mirco-C. (د) تتبع محلل الأجزاء لمكتبة Micro-C النهائية. يرجى النقر هنا لعرض نسخة أكبر من هذا الرقم.
{kind=link}

الشكل 2: إحصائيات العينة للتسلسل منخفض المدخلات لعينة جيدة وعينة سيئة. (أ) يقرأ التمثيل البياني بالأعمدة للنسبة المئوية المعينة (الخضراء) وغير المعينة (الحمراء). (ب) جزء طبيعي من القراءات يرسم خرائط رابطة الدول المستقلة والتفاعلات عبر الكروموسومات. تم تطبيع مجموعات البيانات إلى قراءة رسم خرائط رابطة الدول المستقلة. تم تقسيم قراءات رسم خرائط رابطة الدول المستقلة إلى طبقات حسب المسافة بين القراءات الأولى والثانية للعينات المتسلسلة ذات النهاية المزدوجة: ≤1 كيلو بايت (أصفر) و >1 كيلو بايت و ≤10 كيلو بايت (برتقالي) و >10 كيلو بايت (أحمر). ج: رسم تخطيطي للأنواع الجزيئية المحتملة ذات الأحجام الثنائية النيوكليوسومية. (د) النسب المئوية لاتجاهات أزواج القراءة لجميع قراءات العينة الجيدة والعينة السيئة. (ه) نفس اللوحة (D) ولكن مقسمة حسب المسافات (يسار ، <562 نقطة أساس ويمين ، ≥562 نقطة أساس). يرجى النقر هنا لعرض نسخة أكبر من هذا الرقم.
{kind=link}
| مكونات | 1x | 4.4 أضعاف |
| 10x NEBuffer 2.1 ، | 10 ميكرولتر | 44 ميكرولتر |
| 2 ميكرولتر 100 مللي متر ATP | 2 ميكرولتر | 8.8 ميكرولتر |
| 100 مللي متر DTT | 5 ميكرولتر | 22 ميكرولتر |
| H2O | 68 ميكرولتر | 299.2 ميكرولتر |
| 10 وحدة / ميكرولتر T4 PNK | 5 ميكرولتر | 22 ميكرولتر |
| مجموع | 90 ميكرولتر | 396 ميكرولتر |
الجدول 1: مزيج رئيسي Micro-C 1. تكوين المزيج الرئيسي لرد فعل المضغ النهائي.
| مكونات | 1x | 4.4 أضعاف |
| 1 مللي متر البيوتين-dATP | 10 ميكرولتر | 44 ميكرولتر |
| 1 مللي متر البيوتين-dCTP | 10 ميكرولتر | 44 ميكرولتر |
| مزيج 10 مللي متر من dTTP و dGTP | 1 ميكرولتر | 4.4 ميكرولتر |
| 10x T4 الحمض النووي ليجاز العازلة | 5 ميكرولتر | 22 ميكرولتر |
| 200x BSA | 0.25 ميكرولتر | 1.1 ميكرولتر |
| H2O | 23.75 ميكرولتر | 104.5 ميكرولتر |
الجدول 2: مايكرو سي ماستر ميكس 2. تكوين المزيج الرئيسي لتفاعل وضع العلامات النهائي.
| مكونات | 1x | 4.4 أضعاف |
| 10x NEB T4 Ligase تفاعل العازلة | 50 ميكرولتر | 220 ميكرولتر |
| H2O | 422,5 ميكرولتر | 1859 ميكرولتر |
| T4 الحمض النووي ليجاز | 25 ميكرولتر | 110 ميكرولتر |
الجدول 3: مزيج رئيسي Micro-C 3. تكوين المزيج الرئيسي لتفاعل ربط القرب.
| مكونات | 1x | 4.4 أضعاف |
| 10x NEBuffer 1.1 | 20 ميكرولتر | 88 ميكرولتر |
| H2O | 180 ميكرولتر | 792 ميكرولتر |
| نوكلياز ExoIII | 10 ميكرولتر | 44 ميكرولتر |
الجدول 4: مزيج رئيسي Micro-C 4. تكوين المزيج الرئيسي لتفاعل إزالة البيوتين.
Discussion
يعتمد نجاح تجربة Micro-C على بعض الخطوات الحاسمة في البروتوكول التي يجب تنفيذها بعناية. أولا ، يمكن أن يؤدي التشابك مع DSG أو EGS الإضافي إلى تجميع الخلايا ، اعتمادا على نوع الخلية. إن إضافة 0.1٪ -0.5٪ BSA إلى تفاعل التشابك يقلل بشكل كبير من التجميع دون التأثير على كفاءة التشابك. يمكن أن يؤدي التشابك غير الفعال إلى زيادة معدلات التفاعلات عبر الكروموسومات التي تدل على الروابط العشوائية. الخطوة الثانية ، ولكن الأكثر أهمية ، في هذا البروتوكول هي هضم الكروماتين مع MNase. يؤدي هضم الكروماتين دون المستوى الأمثل إلى عدم كفاءة ربط القرب (الإفراط في الهضم) أو زيادة معدلات ثنائي النيوكليوسومات غير القريب (نقص الهضم). يمكن تقييم كفاءة تفاعل الربط عن طريق الرحلان الكهربائي لهلام الأغاروز (الشكل 1 ب) ويتم تقديره بشكل أفضل من خلال التسلسل منخفض المدخلات. إذا كشف تسلسل المدخلات المنخفضة إما عن معدل تكرار مرتفع (ربط غير فعال) أو زيادة معدلات ثنائي النوكليوسوم ، فيجب إعادة تقييم درجة هضم MNase. والجدير بالذكر أن فقدان العينة عند تنفيذ البروتوكول يمكن أن يؤدي إلى تقليل تعقيد المكتبة. من الأفضل تقييم تركيز العينة بعد تنقية الحمض النووي (الخطوة 5.3) أو عن طريق الحد الأدنى من تفاعل البوليميراز المتسلسل (الخطوة 8). العائد الإجمالي للحمض النووي من 5 × 106 خلايا الثدييات بعد تنقية الحمض النووي هو عادة >2 ميكروغرام. يجب التحكم في تركيز الحمض النووي بعد هضم MNase وهضم ExoIII وتنقية الحمض النووي. يمكن أن تكون النيوكليازات الداخلية ، التي تكون وفرتها خاصة بنوع الخلية والأنواع الخاصة بها ، مصدرا لتدهور الحمض النووي. بالإضافة إلى ذلك ، يمكن أن تؤدي تنقية الحمض النووي المستندة إلى العمود إلى فقدان العينة بسبب عدم التوافق مع SDS من تفاعلات إزالة البروتين. يمكن اعتبار ترسيب الإيثانول إذا كان تركيز الحمض النووي منخفضا في هذه الخطوة.
نظرا لأن Micro-C يتطلب معايرة MNase الخاصة بالعينة ، فمن الصعب تطبيق Micro-C على مجموعات الخلايا الصغيرة ، مثل الأعضاء الصغيرة من الكائنات الحية النموذجية المختلفة أو الأجنة والخلايا المفردة أو المواد العضوية أو خزعات المريض. هنا ، يقدم Hi-C 3.0 بديلا راسخا باستخدام تفاعل نقطة النهاية عن طريق نوكلياز التقييد الخاص بالتسلسل 8,9.
Micro-C هي تقنية تشكيل كروموسوم عالية الدقة قابلة للتطبيق على نطاق واسع مع نطاق ديناميكي عالي ونسبة إشارة إلى ضوضاء منخفضة ، مما يجعلها مناسبة بشكل خاص للتحقيق في ميزات الكروموسوم قصيرة المدى4،5،8 ، مثل حلقات الكروموسوم. تسمح دقة Micro-C بالتقاط حلقات محسن المروج ، والتي تتجاوز حد الكشف عن Hi-C ، مما يتيح بكفاءة تحليلا أكثر تفصيلا للعلاقة بين تنظيم الجينوم واللائحة13،14،15. علاوة على ذلك ، تم مؤخرا دمج استراتيجيات التقاط الحمض النووي مع Micro-C لزيادة الدقة الخاصة بالموقع للمواقع الجينومية المستهدفة إلى مستويات غير مسبوقة ، مما يكشف عن رؤى جديدة حول البنية التحتية للجينوم ثلاثي الأبعاد16،17،18. باختصار ، نتصور أن Micro-C ومشتقاته ستكون تقنية رئيسية لتشريح دور جينوم 3D في تنظيم النسخ ، وبالتالي تمايز نوع الخلية وصيانتها.
Disclosures
ليس لدى المؤلفين ما يكشفون عنه.
Acknowledgements
نشكر كريستل غوبيتز وكاثلين ستيوارت مورغان على قراءتهما النقدية للمخطوطة. نشكر أنجا غروث ومختبر غروث على دعمهم في إنشاء مختبرنا. نشكر موظفي منصة CPR / reNEW Genomics على الدعم: H. Wollmann و M. Michaut و A. Kalvisa. يتم دعم مركز مؤسسة نوفو نورديسك لطب الخلايا الجذعية (reNEW) من قبل مؤسسة نوفو نورديسك رقم المنحة NNF21CC0073729. يتم دعم مركز مؤسسة نوفو نورديسك لأبحاث البروتين (CPR) من قبل مؤسسة نوفو نورديسك رقم المنحة NNF14CC0001. نشكر مختبر بريكمان في مركز نوفو نورديسك لطب الخلايا الجذعية ، reNEW كوبنهاغن ، على الخلايا الجذعية الجنينية للفأر.
Materials
| Name | Company | Catalog Number | Comments |
| 1 mM Biotin dATP | Jenna Bioscience | NU-835-Bio14-S | |
| 1 mM Biotin dCTP | Jenna Bioscience | NU-809-BioX-S | |
| 10 mM dGTP | NEB | N0442S | |
| 10 mM dTTP | NEB | N0443S | |
| 10 U/ml T4 PNK | NEB | M0201L | |
| 100 U/L Exonuclease III | NEB | M0206L | |
| 10x NEBuffer 1.1 | NEB | B7001S | |
| 10x NEBuffer 2.1 | NEB | B7202S | |
| 10x T4 DNA Ligase buffer | NEB | B0202A | |
| 1x DPBS w/o Mg2+ and Ca2+ | ThermoFisher | 14190144 | |
| 1x LIF | |||
| 2_Mercaptoethanol 50 mM | Gibco | 31350010 | 0.1 mM b-mercaptoethanol |
| 37% Formaldehyde | Sigma Aldrich | 252549-500ML | Caution. See manufactures MSDS |
| 400 U/ml T4 DNA Ligase | NEB | M0202L | |
| 5 U/ml Klenow Fragment | NEB | M0210L | |
| Agarose | BIO-RAD | 1613102 | Caution. See manufactures MSDS |
| BSA 20mg/ml | NEB | B9000S | |
| CaCl2 | |||
| cell counter | |||
| Dimethyl Sulfoxide (DMSO) | Sigma Aldrich | D8418-100ML | Caution. See manufactures MSDS |
| Dynabeads MyOne Streptavidin C1 | Invitrogen | 65001 | |
| DynaMag-2 Magnet | Invitrogen | 12321D | refered to as: magnet magnet for 1.5 ml tubes |
| DynaMag-PCR Magnet | Invitrogen | 492025 | refered to as: magnet magnet for PCR tubes |
| EDTA Ultrapure 0.5M pH 8.0 | Invitrogen | 15575-038 | |
| EGTA Ultrapure 0.5M pH 8.0 | BioWorld | 40121266-1 | |
| Ethanol 96% | VWR Chemicals | 20824365 | quality control system |
| Ethidium Bromide | Invitrogen | 15585-011 | |
| Ethylene glycol bis(succinimidyl succinate) (EGS) | ThermoFisher | 21565 | |
| Fetl Bovin Serum | Sigma Aldrich | F7524 | 15% FBS |
| Gel Loading dye purple (6X) | NEB | B7024S | |
| Glycine | PanReac AppliChem | A1067.0500 | |
| Halt Proteinase inhibitor (100x) | ThermoFisher | 78430 | Caution. See manufactures MSDS |
| IGEPAL CA-630 (NP-40) | Sigma Aldrich | 18896-50ML | |
| MgCl 1 M | Invitrogen | AM9530G | |
| Micrococcal Nuclease (MNase) | Worthington | LS004798 | |
| mouse embryonic stem cells | |||
| NaCl | Sigma Aldrich | S9888-1KG | |
| NEBNext Multiplex Oligos for Illumina (Dual Index primers) | NEB | E7600S | amplification primers for sequencing libraries |
| NEBNext Ultra II DNA library prep kit for Illumina | NEB | E7645L | sequencing library preparation kit |
| NEBNext Ultra II Q5 Master mix | NEB | M0544S | Caution. See manufactures MSDS |
| Non-Essential Amino Acids Solution | Gibco | 11140050 | 1x NEAA |
| Penicillin-Streptomycin (10,000 U/mL) | Gibco | 15140148 | 1% Pen-Strep |
| Proteinase K (40 mg/ml) | GoldBio | P-480-1 | Caution. See manufactures MSDS |
| QIAquick Gel extraction kit | QIAgen | 28706 | refered to as: DNA gel elution kit |
| QIAquick PCR purification kit | QIAgen | 28106 | refered to as: commercial DNA purification kit |
| Qubit dsDNA HS Assay kit | Invitrogen | Q32854 | high sensitivity DNA quantification instrument |
| Quick load purple 1kb plus DNA Ladder | NEB | N0550S | |
| SPRIselect size selection beads | Beckman Coulter | B23319 | paramagnetic beads |
| ThermoMixer C | Eppendorf | 5382000015 | refered to as: thermomixer |
| Tris | Merck | 10708976001 | |
| Trypsin | |||
| Tween20 | Sigma Aldrich | P7949-100ML | |
| Ultrapure 10% SDS | Invitrogen | 15553-035 | |
| Ultrapure Phenol Chloroform Isoamyl Alcohol (PCI) | Invitrogen | 15593-031 | |
| Fragment Analyzer |
References
- Lieberman-Aiden, E., et al. Comprehensive mapping of long-range interactions reveals folding principles of the human genome. Science. 326 (5950), 289-293 (2009).
- Hsieh, T. -. H. S., Fudenberg, G., Goloborodko, A., Rando, O. J. Micro-C XL: Assaying chromosome conformation from the nucleosome to the entire genome. Nature Methods. 13 (12), 1009-1011 (2016).
- Hsieh, T. -. H. S., et al. Mapping nucleosome resolution chromosome folding in yeast by Micro-C. Cell. 162 (1), 108-119 (2015).
- Krietenstein, N., et al. Ultrastructural details of mammalian chromosome architecture. Molecular Cell. 78 (3), 554-565 (2020).
- Hsieh, T. -. H. S., et al. Resolving the 3D landscape of transcription-linked mammalian chromatin folding. Molecular Cell. 78 (3), 539-553 (2020).
- Belaghzal, H., Dekker, J., Gibcus, J. H. Hi-C 2.0: An optimized Hi-C procedure for high-resolution genome-wide mapping of chromosome conformation. Methods. 123, 56-65 (2017).
- van Holde, K. E. . Chromatin. , (1989).
- Oksuz, B. A. Systematic evaluation of chromosome conformation capture assays. Nature Methods. 18, 1046-1055 (2021).
- Lafontaine, D. L., Yang, L., Dekker, J., Gibcus, J. H. Hi-C 3.0: Improved protocol for genome-wide chromosome conformation capture. Current Protocols. 1 (7), 198 (2021).
- Goloborodko, A., Venev, S., Abdennur, N., Di Tommaso, P. Mirnylab/distiller-nf; v033. Zenodo. , (2019).
- Venev, S., et al. Open2c/cooltools: v0.4.1. Zenodo. , (2021).
- Abdennur, N., Mirny, L. A. Cooler: Scalable storage for Hi-C data and other genomically labeled arrays. Bioinformatics. 36 (1), 311-316 (2019).
- Zhang, S., Übelmesser, N., Barbieri, M., Papantonis, A. Enhancer-promoter contact formation requires RNAPII and antagonizes loop extrusion. BioRxiv. , (2022).
- Barshad, G., et al. RNA polymerase II and PARP1 shape enhancer-promoter contacts. BioRxiv. , (2022).
- Hansen, A. S., et al. Distinct classes of chromatin loops revealed by deletion of an RNA-binding region in CTCF. Molecular Cell. 76 (3), 395-411 (2019).
- Hua, P., et al. Defining genome architecture at base-pair resolution. Nature. 595 (7865), 125-129 (2021).
- Downes, D. J. High-resolution targeted 3C interrogation of cis-regulatory element organization at genome-wide scale. Nature Communications. 12, 531 (2021).
- Goel, V. Y., Huseyin, M. K., Hansen, A. S. Region capture Micro-C reveals coalescence of enhancers and promoters into nested microcompartments. BioRxiv. , (2022).
Reprints and Permissions
Request permission to reuse the text or figures of this JoVE article
Request PermissionExplore More Articles
This article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. All rights reserved