
Method Article
Memeli 3D Genom Etkileşimlerinin Micro-C-XL ile Haritalanması
Bu Makalede
Özet
Genom çapında kromozom konformasyon yakalama yöntemi Micro-C-XL kullanılarak üç boyutlu genom organizasyonunu nükleozom çözünürlüğü ile haritalamak için bir protokol burada sunulmaktadır.
Özet
Üç boyutlu (3D) kromozom organizasyonu, genom regülasyonunda ve hücre tipi spesifikasyonunda önemli bir faktördür. Örneğin, güçlendiriciler olarak bilinen cis-düzenleyici elemanların, 3B uzayda etkileşim yoluyla distal promotörlerin aktivitesini düzenlediği düşünülmektedir. Hi-C gibi genom çapında kromozom konformasyonu yakalama (3C) teknolojileri, genomların hücrelerde nasıl organize edildiğine dair anlayışımızı değiştirdi. 3B genom organizasyonunun mevcut anlayışı, kromozomların 3B uzaydaki topolojik organizasyonunun çözülebileceği çözünürlükle sınırlıdır. Micro-C-XL, kromozom konformasyonu yakalama protokolü sırasında genomları parçalamak için mikrokokal nükleaz (MNaz) kullanarak, kromatinin temel birimi olan nükleozom seviyesinde çözünürlükle kromozom katlanmasını ölçer. Bu, ölçümlerde gelişmiş bir sinyal-gürültü oranı ile sonuçlanır, böylece diğer genom çapında 3D teknolojilere kıyasla yalıtım bölgelerinin ve kromozom döngülerinin daha iyi tespit edilmesini kolaylaştırır. Bu makalede, memeli hücrelerinden yüksek kaliteli Micro-C-XL örnekleri hazırlamak için görsel olarak desteklenen, ayrıntılı, adım adım bir protokol sunulmaktadır.
Giriş
Micro-C-XL, nükleozom çözünürlüğü ile 3D genom konformasyonunu ölçmek için genom çapında bir tekniktir. Micro-C-XL, 3D genomların nasıl organize edildiğine dair anlayışımızı değiştiren, yaygın olarak kullanılan yakınlık ligasyonu tabanlı Hi-C teknolojisine dayanmaktadır1. Micro-C-XL ve ilk yinelemesi olan Micro-C, başlangıçta Saccharomyces cerevisiae 2,3'te geliştirildi ve daha sonra protokolün kromozom döngüleri ve yalıtım bölgeleri gibi 3D genomun kısa menzilli özelliklerini tespit etmede tam potansiyelini gösterdiği memeli hücre sistemlerine uyarlandı. Bu versiyon, son memeli Micro-C-XL yayınlarınadayanmaktadır 4,5. Micro-C-XL, Micro-C'nin yerini aldığından, Micro-C-XL bundan böyle makalede Micro-C olarak anılacaktır.
Micro-C ve Hi-C6 arasındaki başlıca farklar aşağıdaki gibidir: 1) restriksiyon enzimlerine kıyasla mikrokokal nükleaz (MNaz) ile genom fragmantasyonu ve 2) sadece formaldehite kıyasla reaktif gruplar arasında daha büyük atomik aralığa sahip ek çapraz bağlayıcılar. Her iki adım da, geleneksel Hi-C'ye kıyasla Micro-C'nin geliştirilmiş sinyal-gürültü oranına önemli ölçüde katkıda bulunur. Parçalanma boyutu, yakınlık ligasyonu protokolü sırasında 3B genom organizasyonunun çözümlenebileceği çözünürlüğü sınırlar. MNaz, tercihen erişilebilir DNA'yı sindiren ve nükleozomal korumalı DNA'yı sağlam bırakan bir nükleazdır. MNaz dizilimi kullanılarak nükleozom ayak izi, nükleozomların çoğu ökaryotik genomu tamamen kapsadığını göstermiştir7. Nükleozomlar, türlere ve hücre tipine bağlı olarak ortalama 160-220 bp aralıklarla genom boyunca dağıldığından, MNase, genom mimarisinin yüksek çözünürlüklü haritalanması için ideal enzimdir.
Micro-C yönteminde formaldehit (FA) ile kombinasyon halinde ek bir çapraz bağlayıcının kullanılması ayrıca sinyal-gürültü oranını 2,8 iyileştirir. Reaktif gruplar arasında daha uzun atomik ara parçalara sahip amine özgü çapraz bağlayıcılar, protein-protein çapraz bağlarını kolaylaştırır. Bunlar tipik olarak sırasıyla 7.7 şve 16.1 şara parçalı diseksinimidil glutarat (DSG) veya etilen glikol bis-süksinimidil süksinat (EGS)'dir. EGS veya DSG yoluyla gürültünün azaltılması, özellikle Micro-C gibi yüksek parçalanma oranlarına sahip deneylerde belirgindir ve muhtemelen rastgele ligasyon olaylarınınhızındaki bir azalma nedeniyle ortaya çıkar 8.
ESG/DSG çapraz bağlama ve çoklu kısıtlama enzim kombinasyonlarını kullanan yakın zamanda geliştirilen bir Hi-C 3.0 protokolü, Hi-C deneylerinde gürültüyü azaltır ve kromozom döngülerinin ve yalıtım bölgelerinintespitini önemli ölçüde iyileştirir 8,9. Yine de, çeşitli etkileşim verileri özelliklerinin site bazında karşılaştırılması, Micro-C'nin hem Hi-C 3.0 hem de geleneksel Hi-C8'e kıyasla kromozom döngüleri ve yalıtım bölgeleri gibi kısa menzilli özelliklerin daha iyi algılandığını buldu. Bununla birlikte, Hi-C 3.0, kısa menzilli özelliklerin tespitini iyileştirir ve geleneksel Hi-C'ye kıyasla genom bölümlendirmesinin güçlü bir şekilde algılanmasını sağlar. Özetle, bir kromozom konformasyonu yakalama yönteminin seçimi, nesnel ve biyolojik soru tarafından belirlenmelidir.
Burada, 3D genom organizasyonunu çözebilecek başarılı Micro-C deneyleri için adım adım bir protokol sunuyoruz.
Protokol
1. Hücre kültürü ve çapraz bağlama
- Deneye göre kültür hücrelerinin minimum 1 x 107 hücre elde edilmesi gerekir. Burada hücreler 37 °C'de E14 ortamında %5CO2 ile büyütüldü (piruvat ve L-glutamin ile DMEM, %15 FBS, 1x LIF, 1x NEAA, %1 pen-strep, 0.1 mM β-merkaptoetanol [Malzeme Tablosuna bakınız]) ve her iki günde bir geçiştirildi.
NOT: Bu protokol, Homo sapiensve Mus musculus gibi birçok türden çeşitli hücre tiplerine başarıyla uygulanmıştır. Bu örnekte, fare embriyonik kök (ES) hücreleri kullanılmıştır. - Ortamı aspire ederek hücreleri% 70 -% 80 birleşme noktasında hasat edin. 5 mL DPBS ile bir kez yıkayın ve hücreleri 10 cm'lik tabak başına 3 mL önceden ısıtılmış %0.25 tripsin ile 37 °C'de 2-3 dakika inkübe edin.
- Tripsini 7 mL önceden ısıtılmış E14 ortamı ile söndürün ve ayrılan hücreleri 50 mL'lik bir tüpe aktarın.
- Hücreleri, oda sıcaklığında (RT) 5 dakika boyunca 300 x g'da santrifüjleme ile peletleyin. Süpernatanı atın ve ortamdaki hücre peletini yeniden süspanse edin. Hücreleri bir hücre sayacı ile sayın.
NOT: Protokol, hücre konsantrasyonlarındaki bazı değişikliklere karşı sağlamdır ve hücre sayılarını ölçmek için çeşitli hücre sayaçları kullanılmıştır; Ancak, kullanılan hücre sayacının dinamik aralığına dikkat edin. Bu tipik olarak 1 x 105-1 x 107 hücre/mL arasındadır. Santrifüj hızı ve süresi bu hücre tipi için test edilmiş olsa da, farklı, daha küçük hücreler daha yüksek bir santrifüj hızı veya daha uzun sıkma süreleri gerektirebilir ve santrifüjleme buna göre ayarlanmalıdır. - Hücreleri RT'de 5 dakika boyunca 300 x g'de santrifüjleme ile toplayın ve hücreleri DPBS'de 1 x 106 hücre / mL'lik bir nihai konsantrasyonda yeniden süspanse edin. Örneğin, verim 1 x 107 hücre ise, hücreleri 10 mL DPBS'de yeniden süspanse edin.
- İlk çapraz bağlama adımı için, hücre süspansiyonuna% 1'lik bir nihai konsantrasyona% 37 formaldehit (FA) ekleyin ve hücre süspansiyonunu rotasyonla (15-20 rpm) RT'de 10 dakika inkübe edin; Örnek olarak, verim 1 x 107 hücre ise, 10 mL hücre süspansiyonuna 270 μL FA ekleyin.
NOT: FA çözeltisi tipik olarak açıldıktan sonra 3 aya kadar RT'de stabildir. - 0.25 M'lik bir nihai konsantrasyona 2.5 M glisin ekleyerek reaksiyonu söndürün ve RT'de rotasyonla (15-20 rpm) 5 dakika inkübe edin. Örneğin, verim 1 x 107 hücre ise, hücre süspansiyonuna 1.027 mL 2.5 M glisin ekleyin.
- Hücreleri RT'de 5 dakika boyunca 300 x g'da santrifüjleme ile toplayın. Süpernatanı atın ve hücreleri 5 mL DPBS'de yeniden süspanse edin. Santrifüjleme adımını bir kez tekrarlayın ve DPBS'deki peletlenmiş hücreleri 4 x 106 hücre/mL'ye yeniden süspanse edin. Örneğin, verim 1 x 107 hücre ise, hücreleri 2.5 mL DPBS'de yeniden süspanse edin.
- İkinci çapraz bağlama adımı için, DMSO'da (100 μL DMSO'da 13.6 mg EGS) 0.3 M'lik bir etilen glikol bis-süksinimidil süksinat (EGS) stok çözeltisi hazırlayın. Hücre süspansiyonuna 3 mM'lik bir son konsantrasyonda EGS ekleyin ve rotasyonla (15-20 rpm) 40 dakika boyunca RT'de inkübe edin. Örneğin, verim 1 x 107 hücre ise, 2,5 mL hücre süspansiyonuna 25 μL 0,3 M EGS stok çözeltisi ekleyin.
NOT: Stok çözeltiyi hazırlamak için tartımdan önce EGS'yi en az 20 dakika RT olarak dengeleyin. - 0.4 M'lik bir nihai konsantrasyona 2.5 M glisin ekleyerek reaksiyonu söndürün ve RT'de rotasyonla (15-20 rpm) 5 dakika inkübe edin. Örneğin, verim 1 x 107 hücre ise, hücre süspansiyonuna 400 μL 2.5 M glisin ekleyin.
- Hücreleri RT'de 5 dakika boyunca 1.000 x g'da santrifüjleme ile toplayın ve DPBS'de peletlenmiş hücreleri 5 x 106 hücre / mL'ye yeniden süspanse edin. Hazırlayıcı kitaplıklar için 5 x 10 6 hücre/tüp ve MNaz sindirimi için titrasyon için 1 x 106 hücre/tüp dağıtın.
NOT: Her çapraz bağlı hücre partisi için optimum sindirim derecesinin titre edilmesi önerilir. İdeal olarak, MNase titrasyon deneyleri için 1 x 10 6 hücreden iki ila üç alikot toplayın (adım 2) ve hazırlık deneyleri için 5 x 106 hücreden iki ila dört alikot toplayın (adım 3). - Hücreleri RT'de 5 dakika boyunca 1.000 x g'da santrifüjleme ile toplayın ve süpernatanı çıkarın. Hücre peletlerini sıvı nitrojen içinde dondurun ve −80 °C'de saklayın.
NOT: 3 aya kadar saklanan örneklerden başarılı Micro-C kütüphaneleri hazırlanabilir.
2. MNase titrasyonu
NOT: Çift çapraz bağlı hücrelerin hazırlayıcı kitaplığını işlemeden önce optimal MNaz konsantrasyonunu belirlemek için bir MNaz titrasyonu gerçekleştirmek gerekir.
- MNase titrasyonunu gerçekleştirmek için, 1 x 106 hücreden oluşan bir peleti 10 dakika boyunca buz üzerinde çözün ve hücreleri 500 μL DPBS'de yeniden süspanse edin (hücreler duvara yapışırsa 1x BSA ekleyin). Hücre süspansiyonunu buz üzerinde 20 dakika inkübe edin.
- Hücreleri RT'de 5 dakika boyunca 10.000 x g'da santrifüjleme ile toplayın ve süpernatanı çıkarın. Peletlenmiş hücreleri 500 μL MB # 1 tamponunda (50 mM NaCl, 10 mM Tris-HCl, 5 mM MgCl 2, 1 mM CaCl 2,%0.2 NP-40, 1x proteaz inhibitörü [bkz.
- Hücreleri 5 dakika boyunca 10.000 x g'da santrifüjleme ile toplayın ve süpernatanı çıkarın. Hücreleri 200 μL MB # 1 tamponunda yeniden süspanse edin ve numuneyi dört tüpe bölün.
- Bir şişe MNase (20 U / μL) çözün ve 10 U / μL, 2.5 U / μL, 0.625 U / μL konsantrasyonları elde etmek için 1: 2, 1: 4, 1: 4 ve 1: 4 serilerinde 10 mM Tris, pH 7.4 ile seyreltin ve sırasıyla 0.1256 U / μL (her sindirim koşulu için bir tane). Uygun zaman aralıklarında (10-20 s), dört numuneden birine, vortekse 1 μL MNase çözeltisi ekleyin ve bir termomikser üzerinde 37 °C'de 10 dakika (800 rpm çalkalama) inkübe edin. Kalan hücre alikotlarına kalan MNaz dilüsyonlarından 1 μL ekleyerek devam edin.
- 200 μL taze hazırlanmış STOP tamponu (150 μL 10 mM Tris, pH 7.4, 25 μL %10 SDS, 25 μL 20 mg/mL proteinaz K, 2 μL 0.5 M EGTA) ekleyerek Mnase sindirimini durdurun. 65 °C'de 2 saat inkübe edin.
- Her numuneye 500 μL fenol-kloroform-izoamil alkol (PCI) ekleyin ve girdaplama ile iyice karıştırın. Fazları ayırmak için RT'de 5 dakika boyunca 19.800 x g'da santrifüjleyin ve sulu fazı yeni tüplere aktarın (yaklaşık 200 μL/numune).
DİKKAT: PCI çok sayıda toksik bileşen içerir ve yalnızca kimyasal güvenlik başlığı içinde kullanılmalıdır. Daha fazla ayrıntı için lütfen üreticiye danışın. - Üreticinin talimatlarına göre ticari bir DNA saflaştırma kiti (Malzeme Tablosuna bakınız) kullanarak DNA'yı saflaştırın ve numuneleri 12 μL elüsyon tamponunda elüte edin.
NOT: Deproteinizasyon adımından (adım 2.5) gelen SDS konsantrasyonu, bazı DNA saflaştırma kitleri için inhibitördür. Burada kullanılan DNA saflaştırma kiti, etanol çökeltme ile karşılaştırılabilir bir performans gösterir. - 2-5 μL yükleme boyası ekleyin ve numuneleri 120 V'ta 30-50 dakika boyunca %1.5'lik bir agaroz jel üzerinde çalıştırın (uygun şekilde ayrılana kadar; Şekil 1A).
- Deney için en iyi sindirim derecesini seçin ve hazırlayıcı MNase sindirimine devam edin. Optimal bir sindirim derecesi, çok az veya hiç subnükleozomal fragman ve %70-90 mono--dinükleozom oranı gösterir.
NOT: Bu deneyde, Şekil 1A'daki Şerit 4'ün sindirim derecesinin, 2.5 x 10 5 hücre için 0.625-1.25 U Mnaze ile elde edilen optimal sindirim olduğu belirlenmiştir (= 1 x 106 hücre için 2.5-5 U Mnase). Ayrıntılı bir tartışma için temsili sonuçlara bakın.
3. Hazırlayıcı MNaz sindirimi
- Kromatini MNaz sindirimi yoluyla mono-nükleozomlara parçalamak için, daha önce çift çapraz bağlı 5 x 106 hücreli alikotları çözün ve 1 mL DPBS'de yeniden süspanse edin. Buz üzerinde 20 dakika inkübe edin (hücreler tüp duvarına yapışırsa 1x BSA ekleyin).
- Hücreleri RT'de 5 dakika boyunca 10.000 x g'de santrifüjleme ile toplayın ve süpernatanı atın. Peletin 500 μL MB#1 tamponunda yeniden süspanse edilmesi. Santrifüj adımını bir kez tekrarlayın. Peleti 1 mL MB # 1 tamponunda yeniden süspanse edin ve 200 μL alikot yapın (alikot başına 1 x 106 hücre).
- MNase titrasyonuna (2. adımda açıklanmıştır) dayanarak, her bir alikot'a uygun miktarda MNaz (genellikle 1 x 106 hücre başına 2.5-10 U/μL) ekleyerek kromatini sindirin. İyice karıştırın (hızlı bir şekilde girdap yapın ve döndürün) ve bir termomikser üzerinde 37 °C'de 800 rpm çalkalayarak 10 dakika inkübe edin.
- Her bir alikot için 1,6 μL 0,5 M EGTA (son 4 mM) ekleyerek MNase sindirimini durdurun ve bir termomikser üzerinde 65 °C'de 10 dakika boyunca 800 rpm çalkalama ile inkübe edin.
- Numuneyi RT'de 5 dakika boyunca 10.000 x g'da santrifüjleme ile toplayın ve süpernatanı atın. Hücre peletini 500 μL 1x NEBuffer 2.1'de yeniden süspanse edin.
- Daha fazla işlem için 5 x 106 hücreli veya daha az bir girişe eşdeğer havuz örnekleri.
NOT: 5 x 10 6'dan fazla hücre işlenecekse, enzimatik koşullar 5 x 106 hücre için optimize edildiğinden bu numuneleri paralel olarak işleyin. - Yakınlık ligasyonu adımlarına geçmeden önce, MNase sindirim düzeyini kontrol etmek için numunenin %10'unu giriş kontrolü olarak aktarın. Bu örneğe 150 μL 10 mM Tris, pH 7.4, 25 μL %10 SDS ve 25 μL 20 mg/mL proteinaz K ekleyin ve gece boyunca 65 °C'de inkübe edin.
4. DNA son işleme ve yakınlık ligasyonu
- Kalan numuneyi 4 °C'de 5 dakika boyunca 10.000 x g'da santrifüjleme ile toplayın ve süpernatanı atın. Peletin 90 μL'lik taze hazırlanmış Micro-C ana karışım 1'de (Tablo 1) yeniden süspanse edilmesi ve bir termomikser üzerinde 37 °C'de 800 rpm çalkalama ile 15 dakika inkübe edilmesi.
- 10 μL 5 U/μL Klenow Parçası ekleyin ve bir termomikser üzerinde 37 °C'de 800 rpm çalkalayarak 15 dakika inkübe edin.
- 100 μL taze hazırlanmış Micro-C ana karışım 2 (Tablo 2) ekleyin ve bir termomikser üzerinde 25 °C'de 800 rpm çalkalayarak 45 dakika inkübe edin. İnkübasyondan sonra, 30 mM'lik bir nihai konsantrasyona EDTA ekleyerek enzimatik reaksiyonu söndürün. Bir termomikser üzerinde 65 °C'de 800 rpm çalkalayarak 20 dakika inkübe edin.
- Numuneyi 4 °C'de 5 dakika boyunca 10.000 x g'da santrifüjleme yoluyla toplayın ve süpernatanı atın. Numuneyi 500 μL taze hazırlanmış Micro-C ana karışımı 3 (Tablo 3) içinde yeniden süspanse edin ve RT'de rotasyonla (15-20 rpm) 2,5 saat inkübe edin.
- Numuneyi 4 °C'de 5 dakika boyunca 10.000 x g'da santrifüjleme yoluyla toplayın ve süpernatanı atın. Numuneyi 200 μL taze hazırlanmış Micro-C ana karışım 4 (Tablo 4) içinde yeniden süspanse edin ve bir termomikser üzerinde 37 °C'de 800 rpm çalkalama ile 15 dakika inkübe edin.
- Ters çapraz bağlama ve deproteinasyon için, numuneye 25 μL 20 mg/mL proteinaz K ve 25 μL %10 SDS ekleyin ve aralıklı karıştırma ile gece boyunca 65 °C'de inkübe edin.
5. Di-nükleozomal DNA saflaştırma ve boyut seçimi
- Örneklere ve giriş kontrolüne 500 μL PCI ekleyin ve vorteksleme ile karıştırın. Fazları 5 dakika boyunca 19.800 x g'da santrifüjleme ile ayırın ve üst sulu fazı taze bir tüpe aktarın.
- DNA'yı bir DNA saflaştırma kiti kullanarak veya etanol çökeltme ile konsantre edin. Numuneleri 30 μL'de (adım 5.3) ve giriş kontrollerini (adım 3.11) 15 μL'de kaldırın ve mono-nükleozomları ve di-nükleozomları ayırmak için %1.5'lik bir agaroz jeli çalıştırın (Şekil 1B).
NOT: Deproteinizasyon adımından gelen SDS konsantrasyonu, bazı DNA saflaştırma kitleri için inhibitördür. Burada kullanılan DNA saflaştırma kiti (Malzeme Tablosuna bakınız) etanol çökeltmesine benzer şekilde çalışır. Kullanılan hücre sayısına bağlı olarak, giriş 100 ng ila 10 μg arasında değişebilir. Tipik olarak, 5 x 106 hücreden 1-5 μg DNA ekstrakte edilir. - Di-nükleozomal boyuta (yaklaşık 300 bp) sahip DNA parçalarını kesin. DNA'yı agaroz jelden çıkarmak için ticari olarak temin edilebilen bir DNA jel elüsyon kiti (Malzeme Tablosuna bakınız) kullanın ve 150 μL'de elüte edin.
6. Streptavidin boncuklarının hazırlanması
- Numune başına 10 μL streptavidin boncukları ( Malzeme Tablosuna bakınız) bir reaksiyon tüpüne aktarın.
- 1,5 mL'lik tüpler için uygun bir mıknatısa yerleştirin (Malzeme Tablosuna bakın). Çözelti temizlendikten sonra (1-2 dakika), süpernatanı çıkarın ve boncukları 300 μL 1x TBW (5 mM Tris-HCl, pH 7.5, 0.5 mM EDTA, 1 M NaCl, %0.05 Tween 20) içinde yeniden süspanse edin. Bu adımı bir kez tekrarlayın.
- Boncukları, 7. adımda işlenen numune başına 150 μL 2x S&B tampon (10 mM Tris-HCl, pH 7.5, 1 mM EDTA, 2 M NaCl) içinde yeniden süspanse edin.
7. Streptavidin pull-down ve boncuk kütüphanesi hazırlığı
- Numunenin 150 μL'sine (adım 5.3) 150 μL önceden hazırlanmış boncuk (adım 6.3) ekleyin. RT'de rotasyonla (15-20 rpm) 20 dakika inkübe edin.
- Tüpleri uygun bir mıknatısa yerleştirin ve çözelti temizlenene kadar bekleyin (1-2 dakika). Süpernatanı çıkarın ve boncukları 300 μL 1x TBW'de yeniden süspanse edin. Bu adımı tekrarlayın.
- Tüpleri uygun bir mıknatısa yerleştirin ve çözelti temizlenene kadar bekleyin (1-2 dakika). Süpernatanı çıkarın ve boncukları 100 μL 0.1x TE (1 mM Tris, 0.1 mM EDTA, pH 8.0) içinde yeniden süspanse edin.
- Tüpleri uygun bir mıknatısa yerleştirin ve çözelti temizlenene kadar bekleyin (1-2 dakika). Süpernatanı çıkarın, boncukları 50 μL 0.1x TE'de yeniden süspanse edin ve PCR tüplerine aktarın.
NOT: Kullanılan 0.1x TE hacmi (50 μL), bu protokolde kullanılan DNA dizileme kütüphanesi hazırlama kitinin (bkz. Malzeme Tablosu) giriş hacmine karşılık gelir. Farklı bir kit veya strateji kullanılıyorsa, ses seviyesini buna göre ayarlayın. - Dizileme kütüphanesi hazırlama kitinin DNA manipülasyon adımlarını üreticinin protokolüne göre gerçekleştirin. Bu adımlar tipik olarak DNA köreltme, A-kuyruklama, adaptör ligasyonu ve U eksizyonu içerir. Son adım (U eksizyonu) bu çalışmada kullanılan kite özgüdür. Bu kiti kullanıyorsanız, seyreltilmemiş adaptörler kullanarak kit protokolünde 1. adımdan 2.6. adıma kadar üreticinin talimatlarını izleyin ve 2.6. adımdan sonra −20 °C'de saklamayı düşünmeyin.
NOT: DNA hala streptavidin boncuklarına bağlı olduğundan, dizileme kitlerinin gerektirdiği tampon değişiklikleri, boncukların bir mıknatısa bağlanması ve yıkanmasıyla değiştirilmelidir (7.3 ila 7.4. adımlar). Ek olarak, DNA manyetik boncuklara bağlı olduğundan, kit protokolünden adaptör ligasyonundan sonra herhangi bir saflaştırma ve boyut seçim adımını dikkate almayın. Adım 7.6'ya (bu protokol) geçin. - Adaptör ligasyonu tamamlandıktan sonra, numuneyi adım 7.2'de açıklandığı gibi yıkayın. Süpernatanı atın ve boncukları 20 μL 0.1x TE'de yeniden süspanse edin.
8. Gerekli PCR döngülerinin tahmini
NOT: Kütüphane amplifikasyonu için gerekli PCR döngülerinin tahmin edilmesi tavsiye edilir. Tipik olarak, bir Micro-C kütüphanesi 8-15 döngü PCR gerektirir. Adım gerekli olmasa da, aşırı amplifikasyonun önlenmesine yardımcı olur ve PCR kopyaları riskini azaltır.
- Gerekli minimum PCR döngüsü sayısını tanımlamak için 1 μL streptavidin-biotin-DNA örneği ile PCR gerçekleştirin (adım 7.6). Bunu yapmak için, numuneye PCR ana karışımı (3.2 μLH2O, 0.4 μL i5 primer, 0.4 μL i7 primer ve 5 μL Q5 yüksek kaliteli DNA polimeraz) ekleyin. Kullanılan DNA kütüphanesi hazırlama kiti için üreticinin talimatlarına göre 16 döngü ile PCR gerçekleştirin.
NOT: Burada kullanılan primerler, kullanılan kütüphane hazırlama kiti ile uyumlu olması gereken ayrı bir kit (Malzeme Tablosuna bakınız) içinde satın alınır. - PCR'den sonra, boncukları uygun bir mıknatısla toplayın ve yüksek hassasiyetli bir DNA niceleme cihazı kullanarak süpernatantın 1 μL'lik DNA konsantrasyonunu ölçün (bkz. Toplam DNA konsantrasyonunu elde etmek için bu değeri 10 ile çarpın (PCR reaksiyonunun toplam hacmi). Temsili sonuçlarda tartışıldığı gibi gerekli PCR döngüsü sayısını tahmin edin.
- Kalan 9 μL PCR karışımına 2 μL 6x yükleme tamponu ekleyin. Başarılı adaptör ligasyonunu belirlemek için PCR amplifiye edilmiş kitaplığı %1'lik bir agaroz jel üzerinde çözün, bu da yaklaşık 450 bp boyutuyla sonuçlanır (Şekil 1C).
NOT: Jel, yaklaşık 420 bp'lik (Micro-C kütüphanesi artı adaptörler) farklı bir bant göstermelidir. 120 bp'lik bantlar, bağdaştırıcı dimerlerini temsil eder ve düşük karmaşıklıktaki kitaplıkların göstergesidir. Daha düşük moleküler ağırlıklı bantlar görünebilir (100 bp'den az) ve bunlar PCR reaksiyonundan kullanılmayan primerlerdir.
9. Sıralama kitaplığı amplifikasyonu
- Streptavidin-biotin-DNA örneğinin kalan 19 μL'sine PCR ana karışımı (65 μLH2O, 100 μL Q5 yüksek kaliteli DNA polimeraz, 8 μL i5 primer, 8 μL i7 primer) ekleyin. Reaksiyon karışımını 50-100 μL'lik alikotlara bölün.
NOT: PCR için en uygun hacim, PCR makinesine bağlıdır; tipik olarak, 50 μL, yaygın PCR makinelerinde en tekrarlanabilir amplifikasyonu sunar. - PCR'yi 8. adımda belirlenen döngü sayısı ile DNA kütüphanesi hazırlama kitinin üreticisinin talimatlarına göre gerçekleştirin. 8. adım atlanırsa, PCR için 14 döngü önerilir.
- DNA'yı, üreticinin protokolüne göre 1:0.9 oranında paramanyetik boncuklarla ( Malzeme Tablosuna bakınız) saflaştırın. 20 μL% 0.1 TE'de elüte.
NOT: Adım 8.5'te adaptör dimerleri algılanırsa, aynı oranı kullanarak iki kez saflaştırma gerçekleştirin. - DNA konsantrasyonunu belirleyin ve numuneleri bir kalite kontrol sisteminde çalıştırın (bkz. Tek bir hassas bant varlığıyla Micro-C kitaplığının iyi kalitede olmasını sağlayın (Şekil 1D).
NOT: Adaptör dimerleri, PCR'den önce numune yıkamaları nedeniyle boncuk üzerinde kitaplık preparatlarında nadirdir. Bu nedenle, adaptör dimerlerinin görünümü, düşük kütüphane karmaşıklığını gösterir. Adaptör dimerleri gözlenirse, numunenin düşük giriş sıralaması ile kalite kontrolünün yapılması şiddetle tavsiye edilir.
10. DNA dizilimi ve veri işleme
- Micro-C kitaplığını, sıralama sağlayıcısının gereksinimlerine göre eşleştirilmiş uç dizileme ile sıralayın.
NOT: İdeal olarak, numuneler, okuma başına 50 bp ile eşleştirilmiş uç modunda bir platformda sıralanır. 2 x 35 bp gibi daha kısa okuma uzunlukları sunan eski platformlar da başarıyla kullanılmıştır. Daha da önemlisi, tekrarlayan genomik bölgeler incelenirse, daha uzun bir okuma uzunluğu ile dizileme yapılması tavsiye edilebilir. - Micro-C kitaplığının kalitesini değerlendirmek için, örnek başına 5 x 106 ila 1 x 107 okuma ile düşük giriş sıralaması gerçekleştirin.
- Sıralama dosyalarını (fastq dosyaları) Distiller10 ile işleyin. Okumaları uygun referans genomuna göre eşleyin, burada mm10.
NOT: Sıralama dosyaları, yerel bilgisayarlarda veya bilgi işlem kümelerinde çeşitli ardışık düzenler kullanılarak işlenebilir. Düşük dizileme derinliklerine sahip numuneler için 10.000 bp, 50.000 bp, 100.000 bp ve 500.000 bp gibi daha büyük kutu boyutları, bilgi işlem taleplerini ve dosya boyutlarını azaltabilir. Distiller (bu çalışmada kullanılmıştır), Micro-C kitaplığının kalitesini değerlendirmek için gerekli tüm dosya türlerini oluşturur. Oluşturulan *.stats dosyası, okuma çiftleri arasındaki mesafeye göre katmanlandırılmış harita hızı, cis-trans oranları ve okuma yönü hakkındaki bilgileri içerir. Bu parametreler Şekil 2'de görselleştirilmiştir ve Micro-C kütüphane kalitesinin değerlendirilmesi temsili sonuçlarda tartışılmıştır. İşleme yazılımı ayrıca, etkileşim matrisleri9 oluşturmak için doğrudan HiGlass'a (https://docs.higlass.io/) yüklenebilen mcool dosyaları oluşturur.
Sonuçlar
Mikro-C kütüphanelerinin başarılı bir şekilde hazırlanması, protokolün birkaç adımında değerlendirilebilir. En önemli adım, uygun bir MNase sindirim derecesinin seçilmesidir. Bu nedenle, MNaz konsantrasyonu, her numune için di-nükleozomlar üzerinde tutarlı bir şekilde %70-90 mono-nükleozom verecek şekilde titre edilmelidir. Kromatin sindiriminin eu- ve hetero-kromatin için farklı olduğuna ve MNaz'ın heterokromatini daha az verimli bir şekilde sindirdiğine dikkat etmek önemlidir. Bu nedenle, optimal sindirim derecesi, ilgilenilen kromatin bölgesine ve incelenen hücre tipine bağlıdır, çünkü eu- ve hetero-kromatinin nispi oranı hücre tipine özgüdür. Bu nedenle, gerekli MNaz konsantrasyonunun dikkatli bir şekilde titre edilmesi ve önce Micro-C deneyinin başarısının düşük girdi dizilimi ile değerlendirilmesi tavsiye edilir.
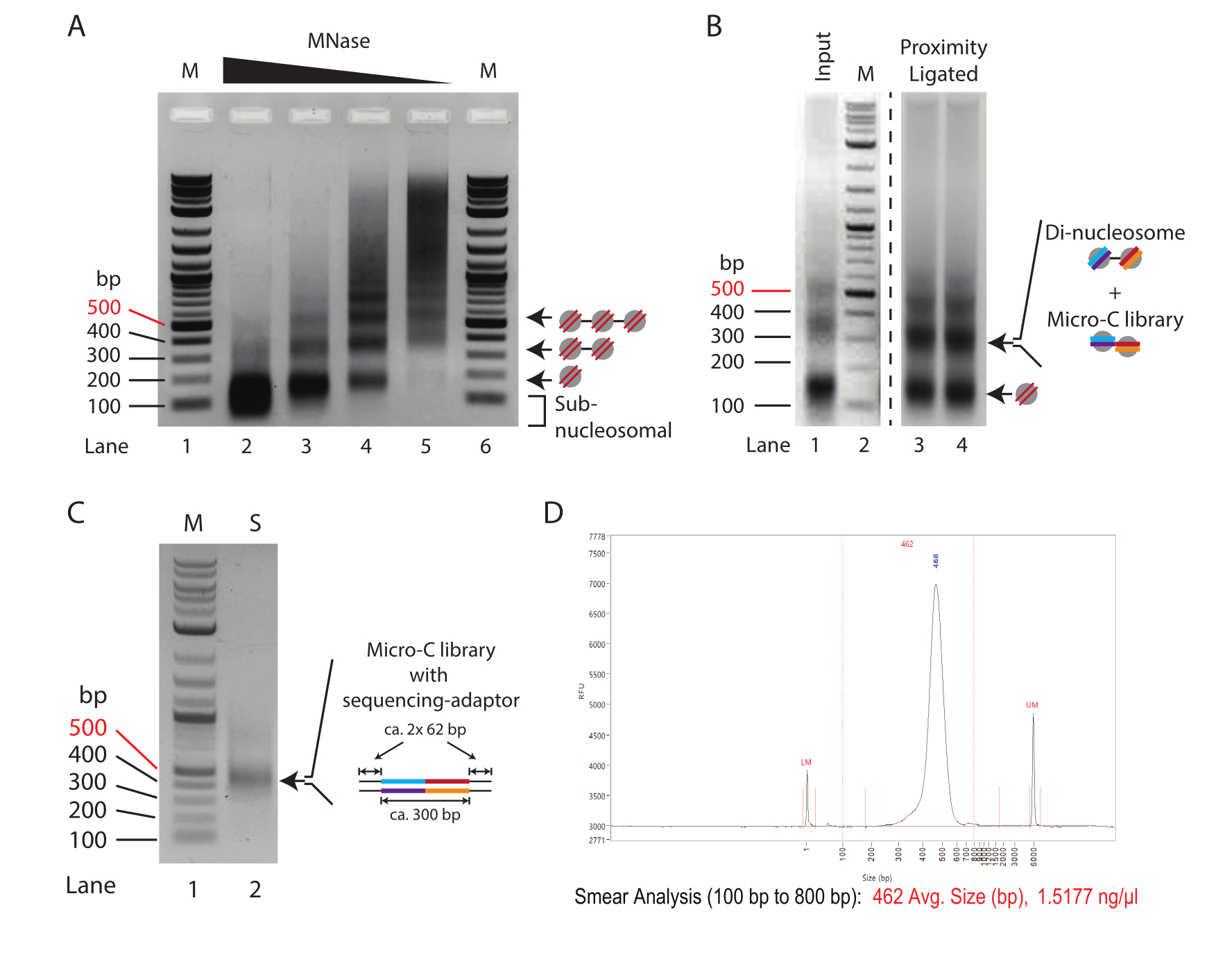
Azalan miktarlarda MNaz ile muamele edilen kromatinin tipik bir MNaz titrasyon modeli Şekil 1A'da gösterilmektedir. Burada, reaksiyon başına 250.000 hücreden elde edilen kromatin, MNaz'ın dört kat seyreltilmesiyle sindirilir. En yüksek konsantrasyon (10 U MNase, Lane 2), neredeyse sadece mono-nükleozomal DNA'dan (~ 150 bp) oluşan aşırı sindirilmiş kromatini gösterir. Özellikle, mono-nükleozomal bandın merkezi, nükleozomal DNA'nın aşırı sindirimini gösteren, azaltılmış MNaz konsantrasyonlarına sahip numunelerdeki karşılık gelen bantlara kıyasla agaroz jelde daha düşük çalışır. Aşırı sindirilmiş nükleozomlar, yakınlık ligasyon reaksiyonunda verimsiz bir şekilde bağlanır; bu nedenle, Şerit 2'deki örnek, Micro-C deneyleri için yetersizdir. Şerit 3 (2,5 U MNase), Micro-C deneyleri için neredeyse uygun bir sindirim derecesi gösterir. Burada, mono-nükleozomal bant baskın türdür ve aşırı sindirilmiş nükleozomların göstergesi olan subnükleozomal yayma azalır; Ancak, hala mevcuttur. Şerit 4'teki sindirim derecesi (0,635 U MNaz), bu titrasyon örneğinde bir Micro-C deneyi için ideal bir koşuldur. Sub-nükleozomal DNA'sı olmayan berrak bir mono-nükleozomal bant mevcuttur. Mono- ve di-nükleozom DNA'sı için bant yoğunluğu hemen hemen eşittir, bu da %66 veya daha yüksek bir mono-nükleozom verimini gösterir. Di-nükleozomal DNA'nın, mono-nükleozomal DNA'nın yaklaşık iki katı büyüklüğünde olduğunu (~320 bp'ye karşı ~150 bp), bu nedenle DNA'nın molü başına bant yoğunluğunun, mono-nükleozomal muadili ile karşılaştırıldığında iki kat daha yüksek olduğunu belirtmekte fayda var. Şerit 5'teki sindirim derecesi (0.156 U MNaz), neredeyse hiç nükleozomal DNA içermeyen yetersiz sindirilmiş kromatin gösterir ve bu nedenle bu, optimal olmayan bir numuneyi temsil eder.
Sonuç olarak, bu örnekte, 2.5 x 10 5 fare ES hücrelerinin 0.625 U MNaz ile sindirimi (200 μL'de 1 x 106 hücre için2.5 U MNaz'a karşılık gelir), Mikro-C deneylerinde hazırlayıcı sindirimler için en umut verici başlangıç noktasını sunar. Bununla birlikte, Şerit 3 ve Şerit 4'teki numuneler için kullanılan koşullar arasında bir ara MNaz konsantrasyonu (200 μL'de 1 x 106 hücre için 5 U MNaz'a karşılık gelir) de dikkate alınmalıdır. Daha da önemlisi, MNaz ile kromatin sindirimi doğrusal olarak ölçeklendirilemez ve hazırlayıcı sindirimin 4 kattan fazla yükseltilmesi önerilmez. 1 x 10 6 hücreden daha fazla Mikro-C kütüphaneleri hazırlamak için, kromatinin 1 x 106 hücreli alikotlarda sindirilmesi ve MNaz inaktivasyonundan sonra havuzlanması tavsiye edilir.
Yakınlık ligasyon protokolünün başarısını değerlendirmek için, MNaz ile sindirilmiş ve yakınlık bağlanmamış (adım 3.8) giriş kontrolü, yakınlık bağlanmış numune (adım 5.3) ile %1.5 agaroz jel elektroforezi ile karşılaştırılmalıdır (Şekil 1B). Yakınlık bağlı mono-nükleozom bandı, di-nükleozomlarınkine benzer şekilde yaklaşık 300 bp'lik bir boyuta sahiptir. Bu nedenle, mono- ila dinükleozomal bant sinyal oranı, ağırlıklı olarak mono-nükleozomlardan (Şerit 1) di-nükleozomlara (Şerit 3 ve Şerit 4) doğru kaymalıdır. Bu adımdaki agaroz jel, eksize edilen ve saflaştırılan di-nükleozomal DNA olduğundan, aşırı yüklemeyi önlemek için numunelerin birden fazla şeride bölünmesi tavsiye edilir.
Hazırlanan dizileme kütüphanesinin nitelik ve niceliğinin minimal PCR ile değerlendirilmesi önerilir. Burada, 1 μL boncuktan (toplam numunenin 1/20'si) alınan DNA, 10 μL PCR reaksiyonunda 16 döngü için amplifiye edilir. Minimal PCR kütüphanesinin toplam konsantrasyonu, 16 PCR döngüsünden sonra tipik olarak 50-500 ng arasında değişir. Teorik olarak, bu, 16 döngü için de amplifiye edilmişse, kalan 19 μL numuneden 1-10 μg'lık bir kütüphaneye karşılık gelir. Toplam DNA'dan yaklaşık 100 ng'lik bir kütüphane oluşturmak için gereken minimum PCR döngüsü sayısının kullanılması önerilir. PCR'de logaritmik amplifikasyon varsayıldığında, 16 döngüde 19 μL girişinden elde edilen DNA'nın teorik konsantrasyonu, 100 ng'lik bir kütüphane oluşturmak için gereken PCR döngülerinin sayısını hesaplamak için art arda ikiye bölünebilir. Örneğin, 16 döngüden sonra 1 μL'den 100 ng'lik bir verim, 19 μL'den amplifiye edilmiş 1.900 ng'lik bir verime karşılık gelir. Bu senaryoda, 12 döngü ideal olarak toplam DNA'dan 118 ng'lik bir dizileme kütüphanesi oluşturmalıdır (1.900 ng/[2 × 2 × 2 × 2] = 118 ng). Minimal PCR'den kalan 9 μL numune daha sonra agaroz jel elektroforezi ile kütüphanenin kalitesini değerlendirmek için kullanılabilir (Şekil 1C). Görselleştirme, 420 bp'de ayrı bir bant göstermeli ve adaptör dimerleri (120 bp) için bant göstermemelidir. Daha küçük fragmanlar da görünebilir ve bunlar kullanılmayan PCR primerlerine karşılık gelir.
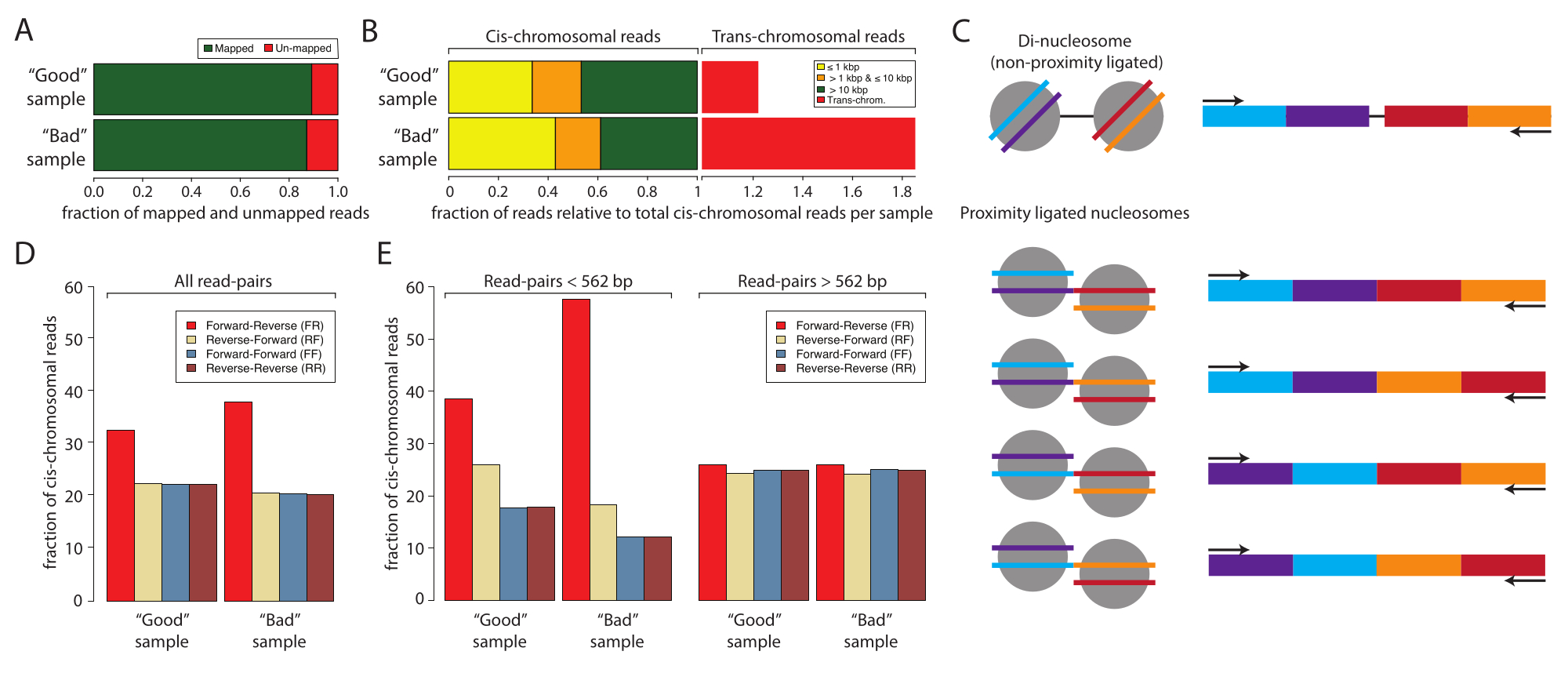
Daha sonra, kaynak yoğun derin dizilemeye geçmeden önce düşük girdili dizileme ile başarılı Micro-C numune hazırlamanın analiz edilmesi ve onaylanması önerilir. Tipik olarak, kitaplıklar 5 x 106 ila 1 x 107 okuma derinliğine göre sıralanır ve aşağıdaki kriterlere göre değerlendirilir: sıralama okuma çoğaltma oranı, cis- ve trans-kromozomal etkileşim hızı ve sıralama okuma oryantasyon frekansı. Micro-C kitaplıkları, okuma dosyalarını (Fastq formatı) sıralamadan okuma çiftleri dosyalarına (Bedpe formatı) ve ölçeklenebilir etkileşim matrislerine (Cool ve Mcool formatları) kadar verileri işleyen tam hizmetli bir boru hattı olan Distiller ile işlenir ve soğutucu, pairtools ve cooltools10,11,12. İşlem hattı ayrıca Micro-C kitaplıklarının10 (https://github.com/open2c/distiller-nf) kalitesini değerlendirmek için ideal olan bir özet dosyası oluşturur. PCR çoğaltma oranı, sıralama kitaplığı karmaşıklığı hakkında bilgi sağlar ve oluşturulan *.stats dosyasından çıkarılabilir. Yüksek kaliteli Micro-C kitaplıkları, 5 milyon veya daha fazla hücreden oluşturulduğunda %5-10'dan daha az PCR çoğaltma oranlarına sahiptir. Özellikle, bazı dizileme platformları, dizileme kitaplığının karmaşıklığından bağımsız olarak küme oluşumu sırasında PCR kopyaları üretir. Şekil 2A, iki deneyin nispi çoğaltma oranlarını göstermektedir: biri iyi bir örnek ve diğeri kötü bir örnek olarak kabul ediyoruz. Bu örnekte, her iki örnek de kabul edilebilir harita hızlarını görüntüledi. Micro-C kütüphanelerinin kalitesini değerlendirmek için bir sonraki kriter, cis-trans oranı ve okuma oryantasyon frekanslarıdır. Çekirdek içinde, kromozomlar bireysel kromozom bölgelerinde yaşar ve bu nedenle nadiren diğer kromozomlarla etkileşime girer. Tespit edilen trans-kromozomal etkileşimlerin yüksek oranı, yüksek oranda rastgele ligasyonları gösterir. Bu analiz seviyesinde, kötü numunenin, iyi numuneye kıyasla yüksek oranda trans-kromozomal etkileşim gösterdiğine dikkat edilmelidir (Şekil 2B). Micro-C için,% 70 veya daha yüksek bir cis-kromozomal etkileşim oranı arzu edilir.
Bir Mikro-C kütüphanesi, yakınlık bağlı numune ile birlikte saflaştırılabilen ve deneyi kontamine edebilen di-nükleozomal DNA bandına benzer bir parça boyutuna sahiptir. Bu kirleticiler her zaman cis-kromozomal etkileşimlerdir. Bu nedenle, okuma oryantasyon oranlarını da değerlendirmek önemlidir. Di-nükleozomal kontaminasyon oranı, düşük girdi dizileme ile tahmin edilebilir. Di-nükleozomal DNA, MNaz tarafından parçalanmamış iki komşu nükleozomdan kaynaklanır. Böylece, ortaya çıkan sıralama okumaları her zaman bir ileri-geri okuma yönü (F ve R) gösterecek ve okuma çiftleri arasındaki mesafe yaklaşık 320 bp olacaktır. Yakınlık bağlanmış fragmanlar, karşılaştırıldığında, ideal olarak eşit bollukta F-R, R-R, R-F ve F-F ile okuma çiftleri verecek şekilde dört yönde bağlanabilir (Şekil 2C). Ek olarak, iki okuma çifti arasında çeşitli mesafeler gösterirler. Di-nükleozomal kirleticilerin miktarını tahmin etmek için, okuma yönelimlerinin sıklığı, damıtıcı tarafından oluşturulan *stats dosyalarından hesaplanabilir (Şekil 2D). Özellikle, bu çalışmada, F-R okumalarının oranı (kırmızı) kötü örnekte iyi örneğe kıyasla daha yüksekti ve bu, okuma yönelimleri mesafeye göre tabakalandırıldığında daha belirgin hale geldi (Şekil 2E). Okuma çiftleri <562 bp veya ≥562 bp mesafeli okumalar halinde tabakalandırıldığında, FR fraksiyonuna Micro-C kütüphanelerine kıyasla di-nükleozomal fragmanlar hakimdir. Burada, <562 bp mesafeli okumaların fraksiyonuna F-R okumaları hakimdir, oysa ≥562 bp mesafeli fraksiyon, dört olası yönelim arasında eşit bir dağılım gösterir, bu da F-R okumalarının küresel aşırı temsilinin di-nükleozomal kirleticilerden kaynaklandığını gösterir. Alt kümeleme eşiği olarak 562 bp seçimi, oluşturulan *stats dosyasındaki gruplama tarafından tanımlanır. Bu kalite kontrol için gerekli olmasa da, damıtıcı tarafından da oluşturulan *pairs dosyasından mesafeler çıkarılarak daha tanımlanmış alt kümeleme elde edilebilir. Di-nükleozomal okumaların, veri işleme sırasında tanımlanabilecekleri ve göz ardı edilebilecekleri için Micro-C numunesinin kalitesini düşürmediğine dikkat etmek önemlidir. Ancak, 3D etkileşimler hakkında değerli bilgiler içermezler ve bilgilendirici okumaları sulandırırlar.
Bu nedenle, dikkatli MNase titrasyonu ve düşük girdili dizileme ile kapsamlı kalite kontrolü, Micro-C deneylerinin kalitesini optimize etmek için en iyi araçlardır.

Şekil 1: Micro-C protokolünün ara aşamaları . (A) Değişen MNaz konsantrasyonları ile sindirilen 2.5 x 105 fare ES hücrelerinden kromatinin agaroz jel elektroforezi. Mono-, di- ve tri-nükleozomal bantlar oklarla gösterilir. M: DNA merdiveni (Şerit 1/6); 250.000 hücre başına 10 U MNaz (Şerit 2); 250.000 hücre başına 2.5 U MNaz (Şerit 3); 250.000 hücre başına 0.625 U MNaz (Şerit 4); 250.000 hücre başına 0.156 U MNaz (Şerit 2). (B) Micro-C ile hazırlanan numunelerin (Şerit 3 ve Şerit 4) %1.0 agaroz jel elektroforezi ve MNase sindirilmiş giriş kontrolü (Şerit 1). Şerit 1 ve Şerit 2 (M: DNA merdiveni), mono- ila di-nükleozomal fragman yoğunluğundaki nispi değişimi vurgulamak için geliştirilmiştir. Mono- ve di-nükleozomal bantlar oklarla gösterilir. Yakınlık bağlanmış numunedeki di-nükleozomal bant, di-nükleozomal ve Micro-C kütüphane DNA'sını birleştirir. (C) Kaliteyi değerlendirmek için 1 μL numuneden amplifiye edilen Micro-C dizileme kitaplıklarının% 1.0 agaroz jel elektroforezi. Şerit 1 (M): DNA merdiveni; Şerit 2 (S): Mirco-C kütüphanesi. (D) Son Micro-C kütüphanesinin Fragment Analyzer izi. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.
{kind=link}

Şekil 2: İyi bir numunenin ve kötü bir numunenin düşük girdili sıralaması için numune istatistikleri. (A) Eşlenen (yeşil) ve eşlenmemiş (kırmızı) okuma yüzdesinin çubuk grafiği. (B) Cis ve kromozomal ötesi etkileşimleri haritalayan okumaların normalleştirilmiş fraksiyonu. Veri kümeleri cis-mapping okumasına normalleştirildi. Cis-eşleme okumaları, eşleştirilmiş uçlu dizilenmiş örneklerin birinci ve ikinci okumaları arasındaki mesafeye göre katmanlandırıldı: ≤1 kbp (sarı), >1 kbp ve ≤10 kbp (turuncu) ve >10 kbp (kırmızı). (C) Di-nükleozomal boyutlara sahip potansiyel moleküler türlerin şeması. (D) İyi numunenin ve kötü numunenin tüm okumalarının okuma çifti yönelimlerinin yüzdeleri. (E) Panel (D) ile aynıdır, ancak mesafelere göre katmanlandırılmıştır (sol, <562 bp ve sağ, ≥562 bp). Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.
{kind=link}
| Bileşen | 1 adet | 4,4 kat |
| 10 adet NEBuffer 2.1, | 10 μL | 44 μL |
| 2 μL 100 mM ATP | 2 μL | 8,8 μL |
| 100 mM DTT | 5 μL | 22 μL |
| H2O | 68 μL | 299,2 μL |
| 10 U/μL T4 PNK | 5 μL | 22 μL |
| Toplam | 90 μL | 396 μL |
Tablo 1: Micro-C ana karışımı 1. Son çiğneme reaksiyonu için ana karışımın bileşimi.
| Bileşen | 1 adet | 4,4 kat |
| 1 mM Biotin-dATP | 10 μL | 44 μL |
| 1 mM Biotin-dCTP | 10 μL | 44 μL |
| 10 mM dTTP ve dGTP karışımı | 1 μL | 4,4 μL |
| 10x T4 DNA Ligaz Tamponu | 5 μL | 22 μL |
| 200 adet BSA | 0,25 μL | 1,1 μL |
| H2O | 23,75 μL | 104,5 μL |
Tablo 2: Micro-C ana karışımı 2. Son etiketleme reaksiyonu için ana karışımın bileşimi.
| Bileşen | 1 adet | 4,4 kat |
| 10x NEB T4 Ligaz reaksiyon tamponu | 50 μL | 220 μL |
| H2O | 422,5 μL | 1859 μL |
| T4 DNA Ligaz | 25 μL | 110μL |
Tablo 3: Micro-C ana karışımı 3. Yakınlık ligasyon reaksiyonu için ana karışımın bileşimi.
| Bileşen | 1 adet | 4,4 kat |
| 10 adet NEBuffer 1.1 | 20 μL | 88 μL |
| H2O | 180 μL | 792 μL |
| ExoIII nükleaz | 10 μL | 44 μL |
Tablo 4: Micro-C ana karışımı 4. Biyotin giderme reaksiyonu için ana karışımın bileşimi.
Tartışmalar
Bir Micro-C deneyinin başarısı, protokolde dikkatli bir şekilde yürütülmesi gereken birkaç kritik adıma bağlıdır. İlk olarak, ek çapraz bağlayıcı DSG veya EGS ile çapraz bağlama, hücre tipine bağlı olarak hücrelerin toplanmasına yol açabilir. Çapraz bağlama reaksiyonuna %0.1-%0.5 BSA eklenmesi, çapraz bağlama verimliliğini etkilemeden agregasyonu önemli ölçüde azaltır. Verimsiz çapraz bağlama, rastgele ligasyonların göstergesi olan trans-kromozomal etkileşim oranlarının artmasına neden olabilir. Bu protokoldeki ikinci, ancak en önemli adım, kromatinin MNaz ile sindirilmesidir. Optimal olmayan kromatin sindirimi, verimsiz yakınlık ligasyonuna (aşırı sindirim) veya yakınlık bağlanmamış di-nükleozom oranlarının artmasına (yetersiz sindirim) yol açar. Ligasyon reaksiyonunun etkinliği, agaroz jel elektroforezi ile değerlendirilebilir (Şekil 1B) ve ayrıca en iyi düşük girdili dizileme ile tahmin edilir. Düşük girdi dizilimi, yüksek bir duplikasyon oranı (verimsiz ligasyon) veya artmış di-nükleozom oranları ortaya çıkarırsa, MNaz sindirim derecesi yeniden değerlendirilmelidir. Özellikle, protokol yürütülürken örnek kaybı, kütüphane karmaşıklığının azalmasına neden olabilir. Bir numunenin konsantrasyonu en iyi DNA saflaştırmasından sonra (adım 5.3) veya minimum PCR (adım 8) ile değerlendirilir. DNA saflaştırmasından sonra 5 x 106 memeli hücresinden elde edilen toplam DNA verimi tipik olarak >2 μg'dır. DNA konsantrasyonu, MNase sindirimi, ExoIII sindirimi ve DNA saflaştırmasından sonra kontrol edilmelidir. Bolluğu hücre tipine özgü ve türe özgü olan endojen nükleazlar, DNA bozulmasının bir kaynağı olabilir. Ek olarak, kolon bazlı DNA saflaştırması, deproteinasyon reaksiyonlarından kaynaklanan SDS ile uyumsuzluk nedeniyle numune kaybına yol açabilir. Bu adımda DNA konsantrasyonu düşükse etanol çökelmesi düşünülebilir.
Micro-C, numuneye özgü MNaz titrasyonu gerektirdiğinden, Micro-C'yi çeşitli model organizmalardan alınan küçük organlar, embriyolar ve tek hücreler, organoidler veya hasta biyopsileri gibi küçük hücre popülasyonlarına uygulamak zordur. Burada, Hi-C 3.0, diziye özgü restriksiyon endonükleazları 8,9 ile bir son nokta reaksiyonu kullanan köklü bir alternatif sunar.
Micro-C, yüksek dinamik aralığa ve düşük sinyal-gürültü oranına sahip, yaygın olarak uygulanabilir yüksek çözünürlüklü bir kromozom konformasyon teknolojisidir, bu da onu özellikle kromozom döngüleri gibi kısa menzilli kromozom özelliklerini 4,5,8 araştırmak için uygun hale getirir. Micro-C'nin çözünürlüğü, Hi-C'nin tespit sınırının ötesinde olan promotör-arttırıcı döngülerin yakalanmasına izin vererek, genom organizasyonu ile düzenleme13,14,15 arasındaki ilişkinin daha ayrıntılı bir analizini verimli bir şekilde sağlar. Ayrıca, DNA yakalama stratejileri, hedeflenen genomik lokusların lokusa özgü çözünürlüğünü benzeri görülmemiş seviyelere çıkarmak için yakın zamanda Micro-C ile birleştirildi ve 3D genomun üst yapısına ilişkin yeni bilgiler ortaya çıktı16,17,18. Özetle, Micro-C ve türevlerinin, 3D genomun transkripsiyonel düzenlemedeki rolünü ve sonuç olarak hücre tipi farklılaşması ve bakımını incelemek için anahtar bir teknoloji olacağını öngörüyoruz.
Açıklamalar
Yazarların açıklayacak hiçbir şeyi yok.
Teşekkürler
Christl Gaubitz ve Kathleen Stewart-Morgan'a el yazmasını eleştirel okumaları için teşekkür ederiz. Laboratuvarımızı kurmamızdaki destekleri için Anja Groth'a ve Groth laboratuvarına teşekkür ederiz. CPR/reNEW Genomik Platformu personeline destekleri için teşekkür ederiz: H. Wollmann, M. Michaut ve A. Kalvisa. Novo Nordisk Vakfı Kök Hücre Tıbbı Merkezi (reNEW), Novo Nordisk Vakfı'nın NNF21CC0073729 numaralı hibesi tarafından desteklenmektedir. Novo Nordisk Vakfı Protein Araştırmaları Merkezi (CPR), Novo Nordisk Vakfı'nın NNF14CC0001 numaralı hibesi tarafından desteklenmektedir. Novo Nordisk Kök Hücre Tıbbı Merkezi'ndeki (reNEW Copenhag) Brickman laboratuvarına fare ES hücreleri için teşekkür ederiz.
Malzemeler
| Name | Company | Catalog Number | Comments |
| 1 mM Biotin dATP | Jenna Bioscience | NU-835-Bio14-S | |
| 1 mM Biotin dCTP | Jenna Bioscience | NU-809-BioX-S | |
| 10 mM dGTP | NEB | N0442S | |
| 10 mM dTTP | NEB | N0443S | |
| 10 U/ml T4 PNK | NEB | M0201L | |
| 100 U/L Exonuclease III | NEB | M0206L | |
| 10x NEBuffer 1.1 | NEB | B7001S | |
| 10x NEBuffer 2.1 | NEB | B7202S | |
| 10x T4 DNA Ligase buffer | NEB | B0202A | |
| 1x DPBS w/o Mg2+ and Ca2+ | ThermoFisher | 14190144 | |
| 1x LIF | |||
| 2_Mercaptoethanol 50 mM | Gibco | 31350010 | 0.1 mM b-mercaptoethanol |
| 37% Formaldehyde | Sigma Aldrich | 252549-500ML | Caution. See manufactures MSDS |
| 400 U/ml T4 DNA Ligase | NEB | M0202L | |
| 5 U/ml Klenow Fragment | NEB | M0210L | |
| Agarose | BIO-RAD | 1613102 | Caution. See manufactures MSDS |
| BSA 20mg/ml | NEB | B9000S | |
| CaCl2 | |||
| cell counter | |||
| Dimethyl Sulfoxide (DMSO) | Sigma Aldrich | D8418-100ML | Caution. See manufactures MSDS |
| Dynabeads MyOne Streptavidin C1 | Invitrogen | 65001 | |
| DynaMag-2 Magnet | Invitrogen | 12321D | refered to as: magnet magnet for 1.5 ml tubes |
| DynaMag-PCR Magnet | Invitrogen | 492025 | refered to as: magnet magnet for PCR tubes |
| EDTA Ultrapure 0.5M pH 8.0 | Invitrogen | 15575-038 | |
| EGTA Ultrapure 0.5M pH 8.0 | BioWorld | 40121266-1 | |
| Ethanol 96% | VWR Chemicals | 20824365 | quality control system |
| Ethidium Bromide | Invitrogen | 15585-011 | |
| Ethylene glycol bis(succinimidyl succinate) (EGS) | ThermoFisher | 21565 | |
| Fetl Bovin Serum | Sigma Aldrich | F7524 | 15% FBS |
| Gel Loading dye purple (6X) | NEB | B7024S | |
| Glycine | PanReac AppliChem | A1067.0500 | |
| Halt Proteinase inhibitor (100x) | ThermoFisher | 78430 | Caution. See manufactures MSDS |
| IGEPAL CA-630 (NP-40) | Sigma Aldrich | 18896-50ML | |
| MgCl 1 M | Invitrogen | AM9530G | |
| Micrococcal Nuclease (MNase) | Worthington | LS004798 | |
| mouse embryonic stem cells | |||
| NaCl | Sigma Aldrich | S9888-1KG | |
| NEBNext Multiplex Oligos for Illumina (Dual Index primers) | NEB | E7600S | amplification primers for sequencing libraries |
| NEBNext Ultra II DNA library prep kit for Illumina | NEB | E7645L | sequencing library preparation kit |
| NEBNext Ultra II Q5 Master mix | NEB | M0544S | Caution. See manufactures MSDS |
| Non-Essential Amino Acids Solution | Gibco | 11140050 | 1x NEAA |
| Penicillin-Streptomycin (10,000 U/mL) | Gibco | 15140148 | 1% Pen-Strep |
| Proteinase K (40 mg/ml) | GoldBio | P-480-1 | Caution. See manufactures MSDS |
| QIAquick Gel extraction kit | QIAgen | 28706 | refered to as: DNA gel elution kit |
| QIAquick PCR purification kit | QIAgen | 28106 | refered to as: commercial DNA purification kit |
| Qubit dsDNA HS Assay kit | Invitrogen | Q32854 | high sensitivity DNA quantification instrument |
| Quick load purple 1kb plus DNA Ladder | NEB | N0550S | |
| SPRIselect size selection beads | Beckman Coulter | B23319 | paramagnetic beads |
| ThermoMixer C | Eppendorf | 5382000015 | refered to as: thermomixer |
| Tris | Merck | 10708976001 | |
| Trypsin | |||
| Tween20 | Sigma Aldrich | P7949-100ML | |
| Ultrapure 10% SDS | Invitrogen | 15553-035 | |
| Ultrapure Phenol Chloroform Isoamyl Alcohol (PCI) | Invitrogen | 15593-031 | |
| Fragment Analyzer |
Referanslar
- Lieberman-Aiden, E., et al. Comprehensive mapping of long-range interactions reveals folding principles of the human genome. Science. 326 (5950), 289-293 (2009).
- Hsieh, T. -. H. S., Fudenberg, G., Goloborodko, A., Rando, O. J. Micro-C XL: Assaying chromosome conformation from the nucleosome to the entire genome. Nature Methods. 13 (12), 1009-1011 (2016).
- Hsieh, T. -. H. S., et al. Mapping nucleosome resolution chromosome folding in yeast by Micro-C. Cell. 162 (1), 108-119 (2015).
- Krietenstein, N., et al. Ultrastructural details of mammalian chromosome architecture. Molecular Cell. 78 (3), 554-565 (2020).
- Hsieh, T. -. H. S., et al. Resolving the 3D landscape of transcription-linked mammalian chromatin folding. Molecular Cell. 78 (3), 539-553 (2020).
- Belaghzal, H., Dekker, J., Gibcus, J. H. Hi-C 2.0: An optimized Hi-C procedure for high-resolution genome-wide mapping of chromosome conformation. Methods. 123, 56-65 (2017).
- van Holde, K. E. . Chromatin. , (1989).
- Oksuz, B. A. Systematic evaluation of chromosome conformation capture assays. Nature Methods. 18, 1046-1055 (2021).
- Lafontaine, D. L., Yang, L., Dekker, J., Gibcus, J. H. Hi-C 3.0: Improved protocol for genome-wide chromosome conformation capture. Current Protocols. 1 (7), 198 (2021).
- Goloborodko, A., Venev, S., Abdennur, N., Di Tommaso, P. Mirnylab/distiller-nf; v033. Zenodo. , (2019).
- Venev, S., et al. Open2c/cooltools: v0.4.1. Zenodo. , (2021).
- Abdennur, N., Mirny, L. A. Cooler: Scalable storage for Hi-C data and other genomically labeled arrays. Bioinformatics. 36 (1), 311-316 (2019).
- Zhang, S., Übelmesser, N., Barbieri, M., Papantonis, A. Enhancer-promoter contact formation requires RNAPII and antagonizes loop extrusion. BioRxiv. , (2022).
- Barshad, G., et al. RNA polymerase II and PARP1 shape enhancer-promoter contacts. BioRxiv. , (2022).
- Hansen, A. S., et al. Distinct classes of chromatin loops revealed by deletion of an RNA-binding region in CTCF. Molecular Cell. 76 (3), 395-411 (2019).
- Hua, P., et al. Defining genome architecture at base-pair resolution. Nature. 595 (7865), 125-129 (2021).
- Downes, D. J. High-resolution targeted 3C interrogation of cis-regulatory element organization at genome-wide scale. Nature Communications. 12, 531 (2021).
- Goel, V. Y., Huseyin, M. K., Hansen, A. S. Region capture Micro-C reveals coalescence of enhancers and promoters into nested microcompartments. BioRxiv. , (2022).
Yeniden Basımlar ve İzinler
Bu JoVE makalesinin metnini veya resimlerini yeniden kullanma izni talebi
Izin talebiThis article has been published
Video Coming Soon
JoVE Hakkında
Telif Hakkı © 2020 MyJove Corporation. Tüm hakları saklıdır