
Method Article
מיפוי אינטראקציות גנום תלת-ממדי של יונקים עם Micro-C-XL
In This Article
Summary
כאן מוצג פרוטוקול למיפוי ארגון הגנום התלת-ממדי ברזולוציית נוקלאוזומים בשיטת לכידת קונפורמציה כרומוזומים רחבת גנום Micro-C-XL.
Abstract
ארגון כרומוזומים תלת-ממדי (תלת-ממדי) הוא גורם מרכזי בוויסות הגנום ובאפיון סוג התא. לדוגמה, אלמנטים CIS-רגולטוריים, המכונים משפרים, נחשבים לווסת את הפעילות של מקדמים דיסטליים באמצעות אינטראקציה במרחב תלת ממדי. טכנולוגיות ללכידת קונפורמציה של כרומוזומים רחבים בגנום (3C), כגון Hi-C, שינו את הבנתנו כיצד גנומים מאורגנים בתאים. ההבנה הנוכחית של ארגון גנום תלת-ממדי מוגבלת על ידי הרזולוציה שבה ניתן לפתור את הארגון הטופולוגי של הכרומוזומים במרחב התלת-ממדי. Micro-C-XL מודד קיפול כרומוזומים ברזולוציה ברמת הנוקלאוזום, היחידה הבסיסית של כרומטין, על ידי שימוש בנוקלאז מיקרוקוקלי (MNase) כדי לפצל גנומים במהלך פרוטוקול לכידת קונפורמציית הכרומוזומים. התוצאה היא יחס אות לרעש משופר במדידות, ובכך מאפשר זיהוי טוב יותר של אתרי בידוד ולולאות כרומוזומים בהשוואה לטכנולוגיות תלת-ממדיות אחרות ברחבי הגנום. במאמר זה מוצג פרוטוקול מפורט ונתמך ויזואלית להכנת דגימות Micro-C-XL באיכות גבוהה מתאי יונקים.
Introduction
Micro-C-XL היא טכניקה כלל-גנומית למדידת קונפורמציה של גנום תלת-ממדי ברזולוציית נוקלאוזומים. Micro-C-XL מתבסס על טכנולוגיית Hi-C המבוססת על קשירת קרבה הנפוצה, אשר שינתה את הבנתנו כיצד גנומים תלת-ממדיים מאורגנים1. Micro-C-XL והאיטרציה הראשונה שלו, Micro-C, פותחו בתחילה ב-Saccharomyces cerevisiae2,3 ומאוחר יותר הותאמו למערכות תאי יונקים, שעבורן הפרוטוקול הוכיח את מלוא הפוטנציאל שלו באיתור תכונות קצרות טווח של הגנום התלת-ממדי, כגון לולאות כרומוזומים ואתרי בידוד. גרסה זו מבוססת על פרסומי Micro-C-XL של יונקים אחרונים 4,5. כפי ש-Micro-C-XL מחליף את Micro-C, Micro-C-XL מכונה מעתה Micro-C בכתב היד.
ההבדלים העיקריים בין Micro-C ו-Hi-C6 הם כדלקמן: 1) פיצול גנום עם נוקלאז מיקרוקוקלי (MNase) בהשוואה לאנזימי הגבלה ו-2) קרוסלינקרים נוספים עם מרווח אטומי גדול יותר בין הקבוצות הריאקטיביות בהשוואה לפורמלדהיד בלבד. שני השלבים תורמים באופן משמעותי לשיפור יחס האות לרעש של Micro-C בהשוואה ל-Hi-C קונבנציונלי. גודל הפיצול מגביל את הרזולוציה שאליה ניתן לפתור את ארגון הגנום התלת-ממדי במהלך פרוטוקול קשירת הקרבה. MNase הוא נוקלאז שמעכל באופן מועדף DNA נגיש ומשאיר DNA מוגן נוקלאוזומים שלם. טביעת רגל נוקלאוזומית באמצעות ריצוף MNase הראתה כי נוקלאוזומים מכסים באופן מלא את רוב הגנומים האיקריוטים7. מכיוון שנוקלאוזומים מפוזרים ברחבי הגנום עם מרווח ממוצע של 160-220 bp, בהתאם למין ולסוג התא, MNase הוא האנזים האידיאלי למיפוי ברזולוציה גבוהה של ארכיטקטורת הגנום.
השימוש בקרוסלינקר נוסף בשילוב עם פורמלדהיד (FA) בשיטת Micro-C משפר בנוסף את יחס האות לרעש 2,8. קרוסלינקרים ספציפיים לאמין עם ספייסרים אטומיים ארוכים יותר בין הקבוצות הריאקטיביות מאפשרים הצלבות חלבון-חלבון. אלה הם בדרך כלל disuccinimidyl גלוטרט (DSG) או אתילן גליקול bis-succinimidyl succinate (EGS) עם 7.7 Å ו 16.1 Å ספייסרים, בהתאמה. הפחתת הרעש באמצעות EGS או DSG בולטת במיוחד בניסויים בעלי קצבי פיצול גבוהים, כגון Micro-C, וככל הנראה מתרחשת עקב ירידה בקצב אירועי קשירה אקראיים8.
פרוטוקול Hi-C 3.0 שפותח לאחרונה המשתמש בקרוסלינקינג ESG/DSG ובשילובים מרובים של אנזימי הגבלה מפחית רעש בניסויי Hi-C ומשפר משמעותית את זיהוי לולאות הכרומוזומים ואתרי בידוד 8,9. עם זאת, השוואה באתר אחר אתר של תכונות נתוני אינטראקציה שונות מצאה כי Micro-C היה זיהוי טוב יותר של תכונות לטווח קצר, כגון לולאות כרומוזומים ואתרי בידוד, בהשוואה הן ל- Hi-C 3.0 והן ל- Hi-C8 קונבנציונלי. עם זאת, Hi-C 3.0 משפר את הזיהוי של תכונות לטווח קצר ושומר על זיהוי חזק של מידור גנום בהשוואה ל- Hi-C קונבנציונלי. לסיכום, הבחירה בשיטת לכידת קונפורמציה כרומוזומלית צריכה להיקבע על ידי השאלה האובייקטיבית והביולוגית.
כאן, אנו מספקים פרוטוקול שלב אחר שלב לניסויי Micro-C מוצלחים שיכולים לפענח ארגון גנום תלת-ממדי.
Protocol
1. תרבית תאים וקרוסלינקינג
- תאי תרבית על פי צרכי הניסוי להשיג מינימום של 1 x 107 תאים. כאן, התאים גודלו ב 37 ° C עם 5% CO2 בתווך E14 (DMEM עם פירובט ו- L-גלוטמין, 15% FBS, 1x LIF, 1x NEAA, 1% pen-strep, 0.1 mM β-mercaptoethanol [ראה טבלה של חומרים]) ועברו כל יום שני.
הערה: פרוטוקול זה יושם בהצלחה על סוגי תאים שונים ממינים רבים, כגון Homo sapiens ו-Mus musculus. בדוגמה זו נעשה שימוש בתאי גזע עובריים של עכברים (ES). - קצרו את התאים במפגש של 70%-80% על ידי שאיפת המדיום. לשטוף פעם אחת עם 5 מ"ל של DPBS, ולדגור את התאים עם 3 מ"ל של 0.25% טריפסין מחומם מראש לכל צלחת 10 ס"מ במשך 2-3 דקות ב 37 ° C.
- להרוות את טריפסין עם 7 מ"ל של מדיום E14 שחומם מראש ולהעביר את התאים מנותקים לצינור 50 מ"ל.
- משחררים את התאים בצנטריפוגה במהירות של 300 x גרם למשך 5 דקות בטמפרטורת החדר (RT). השליכו את הסופרנאטנט והשהו מחדש את גלולת התא בתווך. ספירת התאים באמצעות מונה תאים.
הערה: הפרוטוקול עמיד בפני שינויים מסוימים בריכוזי התא, ומוני תאים שונים שימשו לכימות מספרי התאים; עם זאת, שים לב לטווח הדינמי של מונה התא שבו נעשה שימוש. זה בדרך כלל בין 1 x 105-1 x 107 תאים / מ"ל. בעוד שמהירות הצנטריפוגה ומשך הזמן נבדקו עבור סוג תא זה, תאים שונים וקטנים יותר עשויים לדרוש מהירות צנטריפוגה גבוהה יותר או זמני סיבוב ארוכים יותר, ויש לכוונן את הצנטריפוגה בהתאם. - אסוף את התאים באמצעות צנטריפוגה ב 300 x g במשך 5 דקות ב- RT והשהה מחדש את התאים ב- DPBS בריכוז סופי של 1 x 106 תאים / מ"ל. לדוגמה, אם התשואה היא 1 x 107 תאים, להשעות מחדש את התאים ב 10 מ"ל של DPBS.
- לשלב הקרוסלינקינג הראשון, הוסיפו 37% פורמלדהיד (FA) לריכוז סופי של 1% לתרחיף התא, ודגרו על מתלה התא למשך 10 דקות ב-RT עם סיבוב (15-20 סל"ד); לדוגמה, אם התפוקה היא 1 x 107 תאים, הוסף 270 μL של FA להשעיית התא של 10 מ"ל.
הערה: פתרון FA יציב בדרך כלל ב-RT עד 3 חודשים לאחר הפתיחה. - להרוות את התגובה על ידי הוספת 2.5 M גליצין לריכוז סופי של 0.25 M ולדגור במשך 5 דקות ב RT עם סיבוב (15-20 סל"ד). לדוגמה, אם התפוקה היא 1 x 107 תאים, הוסף 1.027 מ"ל של 2.5 M גליצין לתרחיף התא.
- לאסוף את התאים על ידי צנטריפוגה ב 300 x גרם במשך 5 דקות ב RT. להשליך את supernatant ו resuspend את התאים ב 5 מ"ל של DPBS. חזור על שלב הצנטריפוגה פעם אחת והשהה מחדש את התאים הכדוריים ב- DPBS ל- 4 x 106 תאים / מ"ל. לדוגמה, אם התשואה היא 1 x 107 תאים, להשעות מחדש את התאים ב 2.5 מ"ל של DPBS.
- לשלב הקרוסלינקינג השני, הכינו תמיסת מלאי באורך 0.3 מ' של אתילן גליקול ביס-סוקסינימידיל סוקצינאט (EGS) ב-DMSO (13.6 מ"ג EGS ב-100 מיקרוליטר DMSO). הוסף EGS בריכוז סופי של 3 mM לתרחיף התא, ודגר ב- RT במשך 40 דקות עם סיבוב (15-20 סל"ד). לדוגמה, אם התשואה היא 1 x 107 תאים, הוסף 25 μL של 0.3 M EGS תמיסת מלאי ל 2.5 מ"ל של השעיית התא.
הערה: אזן את ה-EGS ל-RT למשך 20 דקות לפחות לפני השקילה להכנת פתרון המלאי. - להרוות את התגובה על ידי הוספת 2.5 M גליצין לריכוז סופי של 0.4 M, ולדגור במשך 5 דקות ב RT עם סיבוב (15-20 סל"ד). לדוגמה, אם התפוקה היא 1 x 107 תאים, הוסף 400 μL של 2.5 M גליצין לתרחיף התא.
- אסוף את התאים באמצעות צנטריפוגה במהירות של 1,000 x גרם למשך 5 דקות ב- RT, והשהה מחדש את התאים הכדוריים ב- DPBS ל- 5 x 106 תאים / מ"ל. להפיץ 5 x 10 6 תאים / צינור עבור ספריות הכנה ו 1 x 106 תאים / צינור עבור טיטרציה עבור עיכול MNase.
הערה: מומלץ לבצע טיטרציה של דרגת העיכול האופטימלית עבור כל אצווה של תאים צולבים. באופן אידיאלי, לאסוף שניים עד שלושה aliquots של 1 x 10 6 תאים עבור ניסויי טיטרציה MNase (שלב 2) ושניים עד ארבעה aliquots של 5 x 106 תאים עבור ניסויים הכנה (שלב 3). - אספו את התאים באמצעות צנטריפוגה במהירות של 1,000 x גרם למשך 5 דקות ב-RT והוציאו את הסופרנטנט. הקפיאו בזק את כדורי התא בחנקן נוזלי, ואחסנו אותם בטמפרטורה של -80°C.
הערה: ניתן להכין ספריות Micro-C מוצלחות מדגימות המאוחסנות עד 3 חודשים.
2. טיטרציה MNase
הערה: ביצוע טיטרציה של MNase נחוץ כדי לקבוע את הריכוז האופטימלי של MNase לפני עיבוד ספריית ההכנה של התאים המוצלבים הכפולים.
- כדי לבצע את הטיטרציה MNase, הפשיר גלולה אחת של 1 x 106 תאים על קרח במשך 10 דקות והשהה מחדש את התאים ב 500 μL של DPBS (הוסף 1x BSA אם התאים נדבקים לקיר). דוגרים על תרחיף התא על קרח למשך 20 דקות.
- לאסוף את התאים על ידי צנטריפוגה ב 10,000 x גרם במשך 5 דקות ב RT, ולהסיר את supernatant. השהה מחדש את התאים הכדוריים ב-500 μL של מאגר MB#1 (50 mM NaCl, 10 mM Tris-HCl, 5 mM MgCl 2, 1 mM CaCl 2,0.2% NP-40, 1x מעכב פרוטאז [ראה טבלת חומרים], pH 7.4).
- לאסוף את התאים על ידי צנטריפוגה ב 10,000 x גרם במשך 5 דקות, ולהסיר את supernatant. השהה מחדש את התאים ב 200 μL של מאגר MB#1, ופצל את הדגימה לארבע צינורות.
- הפשירו בקבוקון אחד של MNase (20 U/μL), ודללו עם 10 mM Tris, pH 7.4, בסדרה של 1:2, 1:4, 1:4 ו-1:4 כדי להשיג ריכוזים של 10 U/μL, 2.5 U/μL, 0.625 U/μL ו-0.1256 U/μL בהתאמה (אחד לכל תנאי עיכול). במרווחי זמן מתאימים (10-20 שניות), יש להוסיף 1 μL של תמיסת MNase לאחת מארבע הדגימות, מערבולת, ולדגור על תרמומיקסר בטמפרטורה של 37°C למשך 10 דקות (טלטול של 800 סל"ד). המשך עם הוספת 1 μL מדילולי MNase הנותרים לאליציטוטים הנותרים של התא.
- עצור את עיכול Mnase על ידי הוספת 200 μL של חיץ STOP טרי מוכן (150 μL של 10 mM Tris, pH 7.4, 25 μL של 10% SDS, 25 μL של 20 מ"ג / מ"ל פרוטאינאז K, 2 μL של 0.5 M EGTA) לכל צינור באותו סדר ועם מרווח זמן זהה כי MNase נוסף. לדגור ב 65 ° C במשך 2 שעות.
- הוסף 500 μL של פנול-כלורופורם-איזואמיל אלכוהול (PCI) לכל דגימה, וערבב היטב על ידי מערבולת. צנטריפוגה ב 19,800 x גרם במשך 5 דקות ב RT כדי להפריד את השלבים, ולהעביר את הפאזה המימית לצינורות חדשים (כ 200 μL / דגימה).
התראה: PCI מכיל רכיבים רעילים רבים ויש לטפל בו רק במכסה מנוע בטיחות כימי. אנא פנה ליצרן לקבלת פרטים נוספים. - לטהר את הדנ"א באמצעות ערכת טיהור DNA מסחרית (ראה טבלת חומרים) בהתאם להוראות היצרן ולחקות את הדגימות ב-12 μL של חיץ אלוציה.
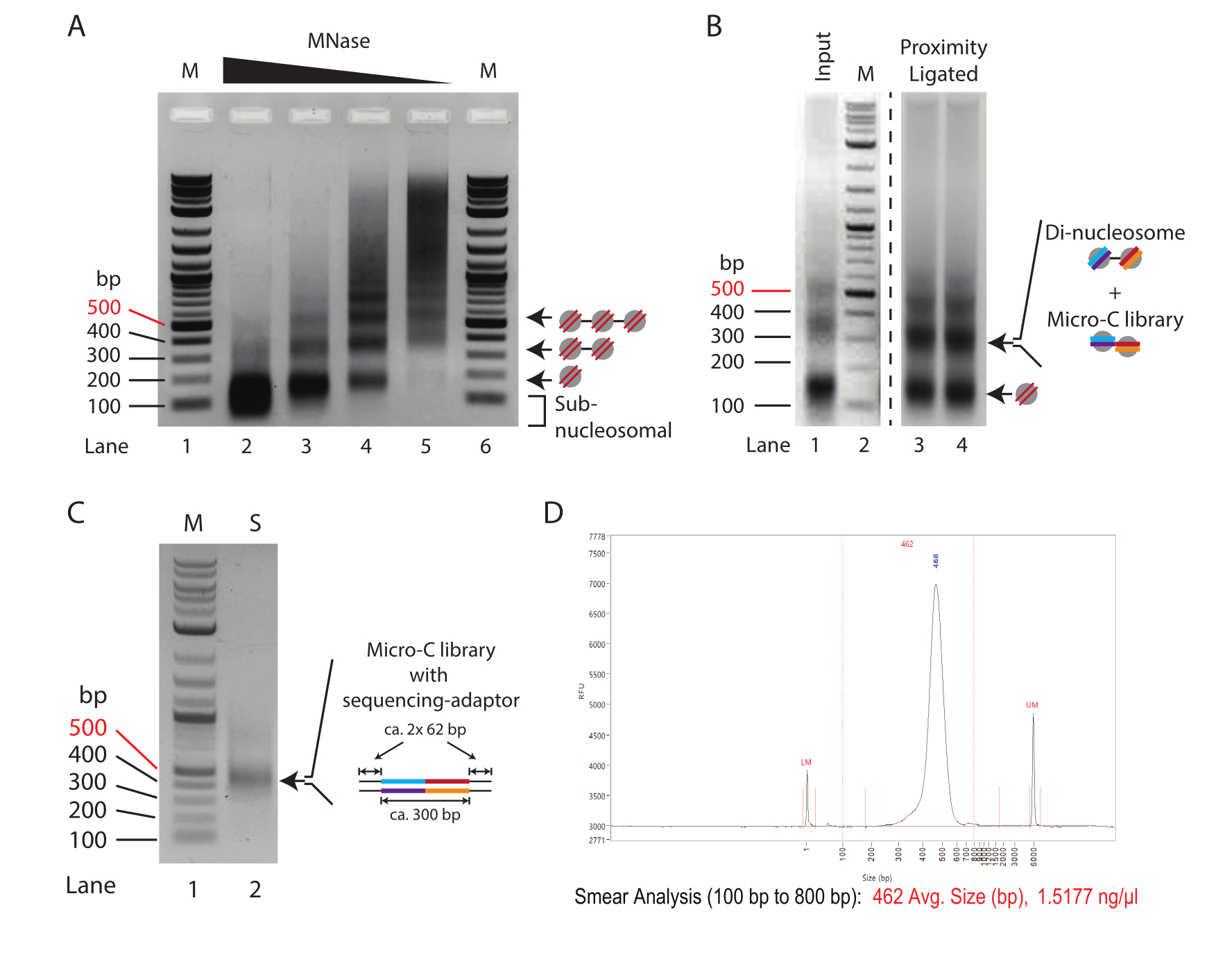
הערה: ריכוז SDS משלב הדה-פרוטאינציה (שלב 2.5) מעכב עבור ערכות טיהור DNA מסוימות. ערכת טיהור הדנ"א המשמשת כאן מתפקדת באופן דומה למשקעים אתנוליים. - מוסיפים 2-5 μL של צבע העמסה, ומריצים את הדגימות על ג'ל אגרוז 1.5% במתח של 120 וולט למשך 30-50 דקות (עד להפרדה נכונה; איור 1A).
- בחר את מידת העיכול הטובה ביותר עבור הניסוי, ולהמשיך עם עיכול MNase הכנה. דרגת עיכול אופטימלית מציגה מעט מאוד מקטעים תת-נוקלאוזומליים ויחס מונו-די-נוקלאוזומי של 70%-90%.
הערה: בניסוי זה, דרגת העיכול של נתיב 4 באיור 1A נקבעה כעיכול האופטימלי שהתקבל עם 0.625-1.25U של מנאז עבור 2.5 x 105 תאים (= 2.5-5 U של מנאז עבור 1 x 106 תאים). לדיון מפורט, ראו את התוצאות המייצגות.
3. עיכול MNase מכין
- כדי לפרק את הכרומטין למונו-נוקלאוזומים באמצעות עיכול MNase, הפשיר בעבר כפול 5 x 106 aliquots תאים, והשהה מחדש ב 1 מ"ל של DPBS. דוגרים על קרח במשך 20 דקות (מוסיפים 1x BSA אם התאים נדבקים לדופן הצינור).
- לאסוף את התאים על ידי צנטריפוגה ב 10,000 x גרם במשך 5 דקות ב RT, ולהשליך את supernatant. השהה מחדש את הגלולה ב 500 μL של מאגר MB#1. חזור על שלב הצנטריפוגה פעם אחת. השהה מחדש את הגלולה ב 1 מ"ל של מאגר MB#1, ולעשות 200 μL aliquots (1 x 106 תאים לכל aliquot).
- בהתבסס על טיטרציה MNase (המתואר בשלב 2), לעכל את הכרומטין על ידי הוספת הכמות המתאימה של MNase (בדרך כלל 2.5-10 U/μL לכל 1 x 106 תאים) לכל aliquot. מערבבים היטב (מערבלים ומסתובבים במהירות), ודגרים על תרמיקסר בטמפרטורה של 37°C למשך 10 דקות עם רעידות של 800 סל"ד.
- עצור את העיכול MNase על ידי הוספת 1.6 μL של 0.5 M EGTA (4 mM סופי) לכל aliquot, ודגור על thermomixer ב 65 ° C במשך 10 דקות עם 800 סל"ד רעד.
- אספו את הדגימה באמצעות צנטריפוגה במהירות של 10,000 x גרם למשך 5 דקות ב-RT, והשליכו את הסופרנטנט. להשעות מחדש את גלולת התא ב 500 μL של 1x NEBuffer 2.1.
- דגימות מאגר שוות ערך לקלט של 5 x 106 תאים או פחות לעיבוד נוסף.
הערה: אם יש לעבד יותר מ- 5 x 106 תאים, עבד דגימות אלה במקביל, מכיוון שהתנאים האנזימטיים ממוטבים עבור 5 x 106 תאים. - לפני שתמשיך לשלבי קשירת הקרבה, העבר 10% מהדגימה כבקרת קלט לבקרה על רמת העיכול של MNase. הוסף 150 μL של 10 mM Tris, pH 7.4, 25 μL של 10% SDS, ו 25 μL של 20 מ"ג / מ"ל פרוטאינאז K לדגימה זו, ודגור לילה ב 65 ° C.
4. עיבוד קצה DNA וקשירת קרבה
- אספו את הדגימה הנותרת באמצעות צנטריפוגה בטמפרטורה של 10,000 x גרם למשך 5 דקות בטמפרטורה של 4°C, והשליכו את הסופרנטנט. יש להשהות מחדש את הגלולה ב-90 מיקרוליטר של תערובת מאסטר 1 טרייה של מיקרו-C (טבלה 1), ולדגור על תרמומיקסר למשך 15 דקות ב-37°C עם רעידות של 800 סל"ד.
- מוסיפים 10 μL של 5 U/μL Klenow Fragment, ודגרים על תרמומיקסר למשך 15 דקות ב-37°C עם טלטול של 800 סל"ד.
- הוסיפו 100 μL של תערובת מאסטר Micro-C טרייה 2 (טבלה 2), ודגרו על תרמומיקסר למשך 45 דקות ב-25°C עם ניעור של 800 סל"ד. לאחר הדגירה, להרוות את התגובה האנזימטית על ידי הוספת EDTA לריכוז סופי של 30 mM. יש לדגור על תרמומיקסר למשך 20 דקות ב-65°C עם רעידות של 800 סל"ד.
- אספו את הדגימה באמצעות צנטריפוגה במהירות של 10,000 x גרם למשך 5 דקות ב-4°C, והשליכו את הסופר-נטנט. השהה מחדש את הדגימה ב- 500 μL של תערובת מאסטר Micro-C טרייה 3 (טבלה 3), ודגר במשך 2.5 שעות ב- RT עם סיבוב (15-20 סל"ד).
- אספו את הדגימה באמצעות צנטריפוגה במהירות של 10,000 x גרם למשך 5 דקות ב-4°C, והשליכו את הסופר-נטנט. יש להשהות מחדש את הדגימה ב-200 מיקרוליטר של תערובת מאסטר 4 טרייה של מיקרו-C (טבלה 4), ולדגור על תרמומיקסר למשך 15 דקות ב-37°C עם טלטול של 800 סל"ד.
- להצלבה הפוכה ודה-פרוטאינציה, יש להוסיף 25 μL של 20 מ"ג/מ"ל פרוטאינאז K ו-25 μL של 10% SDS לדגימה, ולדגור ב-65°C למשך הלילה עם ערבוב לסירוגין.
5. טיהור DNA די-נוקלאוזומלי ובחירת גודל
- הוסף 500 μL של PCI לדגימות ולבקרת הקלט, וערבב על ידי מערבולת. מפרידים את השלבים על ידי צנטריפוגה ב 19,800 x גרם למשך 5 דקות, ומעבירים את הפאזה המימית העליונה לצינור טרי.
- רכז את הדנ"א באמצעות ערכת טיהור DNA או על ידי משקעי אתנול. האזינו את הדגימות ב-30 μL (שלב 5.3) ואת פקדי הקלט (שלב 3.11) ב-15 μL, והפעילו ג'ל אגרוז ב-1.5% כדי להפריד את המונו-נוקלאוזומים והדי-נוקלאוזומים (איור 1B).
הערה: ריכוז SDS משלב הדה-פרוטאיזציה מעכב עבור ערכות טיהור DNA מסוימות. ערכת טיהור הדנ"א המשמשת כאן (ראו טבלת חומרים) מתפקדת באופן דומה למשקעי אתנול. בהתאם למספר התאים המשמשים, הקלט יכול לנוע בין 100 ng ל 10 מיקרוגרם. בדרך כלל, 1-5 מיקרוגרם של DNA מופק מ 5 x 106 תאים. - הבלו את מקטעי ה- DNA שיש להם גודל di-nucleosomal (כ 300 bp). השתמש בערכת ג'ל DNA זמינה מסחרית (ראה טבלת חומרים) כדי לחלץ את הדנ"א מג'ל האגרוז, וללוט ב 150 μL.
6. הכנת חרוזי סטרפטווידין
- מעבירים 10 μL של חרוזי סטרפטאבידין (ראו טבלת חומרים) לכל דגימה לתוך צינור תגובה.
- הכניסו אותו למגנט מתאים לשפופרות של 1.5 מ"ל (ראו טבלת חומרים). לאחר ניקוי התמיסה (1-2 דקות), הסר את הסופרנטנט, והשהה מחדש את החרוזים ב 300 μL של 1x TBW (5 mM Tris-HCl, pH 7.5, 0.5 mM EDTA, 1 M NaCl, 0.05% Tween 20) לכל דגימה. חזור על שלב זה פעם אחת.
- השהה מחדש את החרוזים ב-150 מיקרוליטר של 2x B&W buffer (10 mM Tris-HCl, pH 7.5, 1 mM EDTA, 2 M NaCl) לכל דגימה המעובדת בשלב 7.
7. סטרפטווידין נפתח והכנת ספריית חרוזים
- הוסף 150 μL של חרוזים מוכנים מראש (שלב 6.3) ל 150 μL של המדגם (שלב 5.3). דגירה במשך 20 דקות ב- RT עם סיבוב (15-20 סל"ד).
- הכניסו את הצינורות למגנט מתאים, והמתינו עד שהתמיסה תתנקה (1-2 דקות). הסר את supernatant ו resuspend את החרוזים ב 300 μL של 1x TBW. חזור על שלב זה.
- הכניסו את הצינורות למגנט מתאים והמתינו עד שהתמיסה תתנקה (1-2 דקות). הסר את supernatant ו resuspend את החרוזים ב 100 μL של 0.1x TE (1 mM Tris, 0.1 mM EDTA, pH 8.0).
- הכניסו את הצינורות למגנט מתאים והמתינו עד שהתמיסה תתנקה (1-2 דקות). הסר את supernatant, להשעות מחדש את החרוזים ב 50 μL של 0.1x TE, ולהעביר אותם צינורות PCR.
הערה: הנפח של 0.1x TE המשמש (50 μL) תואם לנפח הקלט של ערכת הכנת ספריית ריצוף ה- DNA (ראה טבלת חומרים) המשמשת בפרוטוקול זה. אם נעשה שימוש בערכה או אסטרטגיה אחרת, כוונן את עוצמת הקול בהתאם. - בצע את שלבי מניפולציית הדנ"א של ערכת הכנת ספריית הריצוף בהתאם לפרוטוקול היצרן. שלבים אלה כוללים בדרך כלל קהות DNA, זנב A, קשירת מתאם וכריתת U. השלב האחרון (כריתת U) הוא ספציפי לערכה ששימשה במחקר זה. אם אתה משתמש בערכה זו, בצע את הוראות היצרן משלב 1 עד שלב 2.6 בפרוטוקול הערכה באמצעות מתאמים לא מדוללים, ואל תשקול אחסון של -20°C לאחר שלב 2.6.
הערה: שינויי החיץ הנדרשים על ידי ערכות הריצוף חייבים להיות מוחלפים על ידי קשירת החרוזים למגנט ושטיפות (שלבים 7.3 עד 7.4), מכיוון שהדנ"א עדיין קשור לחרוזי הסטרפטווידין. בנוסף, מכיוון שהדנ"א קשור לחרוזים המגנטיים, התעלם מכל שלבי הטיהור ובחירת הגודל לאחר קשירת המתאם מפרוטוקול הערכה. המשך לשלב 7.6 (פרוטוקול זה). - לאחר השלמת קשירת המתאם, שטוף את הדגימה כמתואר בשלב 7.2. השליכו את הסופרנאטנט והשהו מחדש את החרוזים ב-20 מיקרוליטר של 0.1x TE.
8. הערכת מחזורי ה-PCR הנדרשים
הערה: מומלץ להעריך את מחזורי ה-PCR הנדרשים להגברה בספרייה. בדרך כלל, ספריית Micro-C דורשת 8-15 מחזורים של PCR. למרות שהצעד אינו חיוני, הוא מסייע למנוע הגברה יתר ומקטין את הסיכון לכפילויות PCR.
- כדי להגדיר את המספר המינימלי של מחזורי PCR נדרשים, בצע PCR עם 1 μL של דגימת סטרפטווידין-ביוטין-DNA (שלב 7.6). לשם כך, הוסף תערובת מאסטר PCR (3.2 μL של H2O, 0.4 μL של פריימר i5, 0.4 μL של פריימר i7, ו 5 μL של Q5 פולימראז DNA באיכות גבוהה) לדגימה. ביצוע PCR עם 16 מחזורים על פי הוראות היצרן עבור ערכת הכנת ספריית DNA בשימוש.
הערה: הפריימרים המשמשים כאן נרכשים בערכה נפרדת (ראה טבלת חומרים) שצריכה להיות תואמת לערכת הכנת הספרייה שבה נעשה שימוש. - לאחר ה-PCR, אספו את החרוזים בעזרת מגנט מתאים, ומדדו את ריכוז הדנ"א של 1 מיקרוליטר של הסופרנאטנט באמצעות מכשיר לכימות DNA בעל רגישות גבוהה (ראו טבלת חומרים). כדי להשיג את ריכוז ה- DNA הכולל, הכפל ערך זה ב -10 (נפח כולל של תגובת PCR). להעריך את המספר הנדרש של מחזורי PCR כפי שנדון בתוצאות המייצגות.
- הוסף 2 μL של מאגר העמסה 6x ל- 9 μL הנותרים של תערובת PCR. פתרו את ספריית ה-PCR המוגברת על ג'ל אגרוז 1% כדי לקבוע קשירת מתאם מוצלחת, והתוצאה היא גודל של כ-450 bp (איור 1C).
הערה: הג'ל צריך להראות רצועה ברורה של כ-420 bp (ספריית Micro-C בתוספת מתאמים). רצועות של 120 bp מייצגות דימרים של מתאמים ומעידות על ספריות בעלות מורכבות נמוכה. רצועות של משקלים מולקולריים נמוכים יותר עשויים להופיע (פחות מ -100 bp), ואלה הם פריימרים שאינם בשימוש מתגובת PCR.
9. הגברה של ספריית רצף
- הוסף תערובת מאסטר PCR (65 μL של H2O, 100 μL של Q5 DNA פולימראז באיכות גבוהה, 8 μL של פריימר i5, 8 μL של פריימר i7) ל -19 μL הנותרים של דגימת סטרפטווידין-ביוטין-DNA. לפצל את תערובת התגובה לתוך 50-100 μL aliquots.
הערה: עוצמת הקול האופטימלית עבור PCR תלויה במכשיר ה-PCR; בדרך כלל, 50 μL מציע את ההגברה הניתנת ביותר לשחזור במכונות PCR נפוצות. - בצע PCR בהתאם להוראות היצרן של ערכת הכנת ספריית ה- DNA עם מספר המחזורים שנקבע בשלב 8. אם שלב 8 הושמט, מומלץ לבצע 14 מחזורים עבור PCR.
- לטהר את הדנ"א בחרוזים פאראמגנטיים (ראה טבלת חומרים) ביחס של 1:0.9 בהתאם לפרוטוקול היצרן. Elute ב 20 μL של 0.1% TE.
הערה: אם זיהו דימרים של מתאם בשלב 8.5, בצע טיהור פעמיים באותו יחס. - לקבוע את ריכוז הדנ"א ולהריץ את הדגימות על מערכת בקרת איכות (ראה טבלת חומרים). ודא איכות טובה של ספריית Micro-C על-ידי נוכחות של פס מדויק יחיד (איור 1D).
הערה: דימרים מתאמים אינם נפוצים בהכנות לספריית חרוזים עקב שטיפות הדגימה לפני ה-PCR. לפיכך, המראה של דימרים מתאמים מצביע על מורכבות ספרייה נמוכה. אם נצפו דימרים מתאמים, מומלץ מאוד לשלוט באיכות הדגימה עם רצף קלט נמוך.
10. ריצוף DNA ועיבוד נתונים
- רצף את ספריית Micro-C עם ריצוף משויך בהתאם לדרישות ספק הרצף.
הערה: באופן אידיאלי, הדגימות מסודרות בפלטפורמה במצב קצה משויך עם 50 bp לכל קריאה. פלטפורמות ישנות יותר המציעות אורכי קריאה קצרים יותר, כגון 2 x 35 bp, שימשו גם הם בהצלחה. חשוב לציין, אם אזורים גנומיים חוזרים נחקרים, ייתכן שיהיה מומלץ לרצף עם אורך קריאה ארוך יותר. - כדי להעריך את איכות ספריית Micro-C, בצע ריצוף קלט נמוך עם 5 x 106 עד 1 x 107 קריאות לכל דגימה.
- עבד את קובצי הרצף (קובצי fastq) באמצעות Distiller10. מפה את הקריאות כנגד גנום הייחוס המתאים, כאן mm10.
הערה: ניתן לעבד את קבצי הרצף באמצעות צינורות שונים במחשבים מקומיים או באשכולות מחשוב. עבור דגימות עם עומק רצף נמוך, פחים גדולים יותר, כגון 10,000 bp, 50,000 bp, 100,000 bp ו- 500,000 bp, יכולים להפחית את דרישות המחשוב ואת גודל הקובץ. Distiller (המשמש במחקר זה) מייצר את כל סוגי הקבצים הדרושים כדי להעריך את האיכות של ספריית Micro-C. קובץ *.stats שנוצר מכיל את המידע על קצב המפה, יחסי cis-trans וכיוון קריאה מרובד על ידי המרחק בין זוגות הקריאה. פרמטרים אלה מוצגים באופן חזותי באיור 2, והערכת איכות ספריית Micro-C נדונה בתוצאות המייצגות. תוכנת העיבוד מייצרת גם קבצי mcool שניתן לטעון ישירות לתוך HiGlass (https://docs.higlass.io/) כדי ליצור מטריצות אינטראקציה9.
תוצאות
ניתן להעריך את ההכנה המוצלחת של ספריות Micro-C במספר שלבים של הפרוטוקול. השלב החשוב ביותר הוא הבחירה של תואר עיכול MNase תקין. לכן, ריכוז MNase חייב להיות titrated כדי להניב באופן עקבי 70%-90% mono-nucleosomes על di-nucleosomes עבור כל דגימה. חשוב לציין כי עיכול הכרומטין שונה עבור כרומטין, כאשר MNase מעכל הטרוכרומטין בצורה פחות יעילה. לפיכך, מידת העיכול האופטימלית תלויה באזור הכרומטין המעניין ובסוג התא הנחקר, שכן החלק היחסי של הכרומטין האירופי וההטרו-כרומטין הוא ספציפי לסוג התא. לכן, מומלץ לבצע טיטרציה זהירה של ריכוז ה-MNase הנדרש ולהעריך תחילה את הצלחת ניסוי ה-Micro-C על ידי ריצוף בעל קלט נמוך.
דפוס טיטרציה טיפוסי של MNase של כרומטין שטופל בכמויות הולכות ופוחתות של MNase מוצג באיור 1A. כאן, כרומטין מ -250,000 תאים לכל תגובה מתעכל עם דילול פי ארבעה של MNase. הריכוז הגבוה ביותר (10 U של MNase, Lane 2) מראה כרומטין מעוכל יתר על המידה המורכב כמעט אך ורק מדנ"א מונו-נוקלאוזומלי (~150 bp). יש לציין כי מרכז הרצועה המונו-נוקלאוזומלית נמוך יותר בג'ל האגרוז בהשוואה לרצועות המקבילות בדגימות עם ריכוזי MNase מופחתים, דבר המצביע על עיכול יתר של DNA נוקלאוזומי. נוקלאוזומים מעוכלים יתר על המידה קשורים באופן לא יעיל בתגובת קשירת הקרבה; לכן, הדגימה בנתיב 2 אינה אופטימלית לניסויי מיקרו-C. נתיב 3 (2.5 U של MNase) מציג דרגת עיכול כמעט מתאימה לניסויי Micro-C. כאן, הרצועה המונו-נוקלאוזומלית היא המין הדומיננטי, והכתם התת-נוקלאוזומי, המעיד על נוקלאוזומים מעוכלים יתר על המידה, מצטמצם; עם זאת, הוא עדיין נוכח. דרגת העיכול בנתיב 4 (0.635 U של MNase) היא תנאי אידיאלי לניסוי Micro-C בדוגמת טיטרציה זו. קיימת רצועה מונו-נוקלאוזומלית ברורה ללא DNA תת-נוקלאוזומי. עוצמת הפס של הדנ"א המונו-נוקלאוזומי והדי-נוקלאוזומי כמעט שווה, מה שמצביע על תפוקה חד-נוקלאוזומית של 66% ומעלה. ראוי לציין כי הדנ"א הדי-נוקלאוזומלי הוא בערך כפול מגודלו של הדנ"א החד-נוקלאוזומלי (~320 bp לעומת ~150 bp), כך שעוצמת הפס שלו לכל שומה של DNA גבוהה פי שניים בהשוואה למקבילו המונו-נוקלאוזולי. דרגת העיכול בנתיב 5 (0.156 U של MNase) מראה כרומטין לא מעוכל כמעט ללא DNA נוקלאוזומי, ולכן זה מייצג דגימה תת-אופטימלית.
לסיכום, בדוגמה זו, העיכול של 2.5 x 10 5 תאי ES עכבר עם 0.625 U של MNase (המקביל ל-2.5 U של MNase עבור 1 x 106 תאים ב- 200 μL) מציע את נקודת ההתחלה המבטיחה ביותר לעיכול מוכן בניסויי Micro-C. עם זאת, יש לשקול גם ריכוז MNase ביניים בין התנאים המשמשים לדגימות בנתיב 3 ונתיב 4 (המקביל ל- 5 U של MNase עבור 1 x 106 תאים ב- 200 μL). חשוב לציין, עיכול כרומטין עם MNase לא ניתן בקנה מידה ליניארי, ולא מומלץ לשדרג את העיכול הכנה יותר מ 4x. כדי להכין ספריות Micro-C מיותר מ 1 x 10 6 תאים, מומלץ לעכל את הכרומטין ב aliquots של 1 x 106 תאים ולצרף אותם לאחר השבתת MNase.
כדי להעריך את ההצלחה של פרוטוקול קשירת הקרבה, יש להשוות את בקרת הקלט, שהיא MNase-digested ולא קשירת קרבה (שלב 3.8), לדגימת קשירת הקרבה (שלב 5.3) באלקטרופורזה של ג'ל אגרוז ב-1.5% (איור 1B). לרצועה מונו-נוקלאוזומים קרבת קרבה יש גודל משוער של 300 bp, דומה לזה של di-nucleosomes. לכן, יחס אות הפס מונו-נוקלאוזומים לדו-נוקלאוזומים צריך להשתנות ממונו-נוקלאוזומים בעיקרם (נתיב 1) לכיוון די-נוקלאוזומים (נתיב 3 ונתיב 4). מכיוון שג'ל האגרוז בשלב זה הוא ה- DNA הדי-נוקלאוזומלי שנכרת ומטוהר, מומלץ לפצל את הדגימות למספר נתיבים כדי למנוע עומס יתר.
מומלץ להעריך את האיכות והכמות של ספריית הריצוף המוכנה על ידי PCR מינימלי. כאן, DNA מ 1 μL של חרוזים (1/20 מכלל הדגימה) מוגבר במשך 16 מחזורים ב 10 μL של תגובת PCR. הריכוז הכולל של ספריית ה-PCR המינימלית נע בדרך כלל בין 50-500 ננוגרם לאחר 16 מחזורי PCR. בתיאוריה, זה מתאים לספרייה של 1-10 מיקרוגרם מהדגימה הנותרת של 19 μL אם היא גם הייתה מוגברת במשך 16 מחזורים. מומלץ להשתמש במספר המינימלי של מחזורי PCR הדרושים ליצירת ספרייה של כ-100 ננוגרם מסך הדנ"א. בהנחה של הגברה לוגריתמית ב-PCR, ניתן לחלק את הריכוז התיאורטי של הדנ"א המתקבל מקלט 19 μL ב-16 מחזורים ברצף בשניים כדי לחשב את מספר מחזורי ה-PCR הדרושים ליצירת ספריית 100 ננוגרם. לדוגמה, תפוקה של 100 ננוגרם מ-1 מיקרוליטר לאחר 16 מחזורים מקבילה לתפוקה של 1,900 ננוגרם מוגברת מ-19 מיקרוליטר. בתרחיש זה, 12 מחזורים אמורים ליצור באופן אידיאלי ספריית ריצוף של 118 ננוגרם מכלל הדנ"א (1,900 ננוגרם/[2 × 2 × 2 × 2] = 118 ננוגרם). לאחר מכן ניתן להשתמש בדגימה הנותרת של 9 μL מה-PCR המינימלי כדי להעריך את איכות הספרייה על-ידי אלקטרופורזה של ג'ל אגרוז (איור 1C). תצוגה חזותית צריכה להציג פס אחד נפרד ב- 420 bp וללא פסים עבור דימרים מתאמים (120 bp). שברים קטנים יותר עשויים להופיע גם, ואלה תואמים פריימרים PCR שאינם בשימוש.
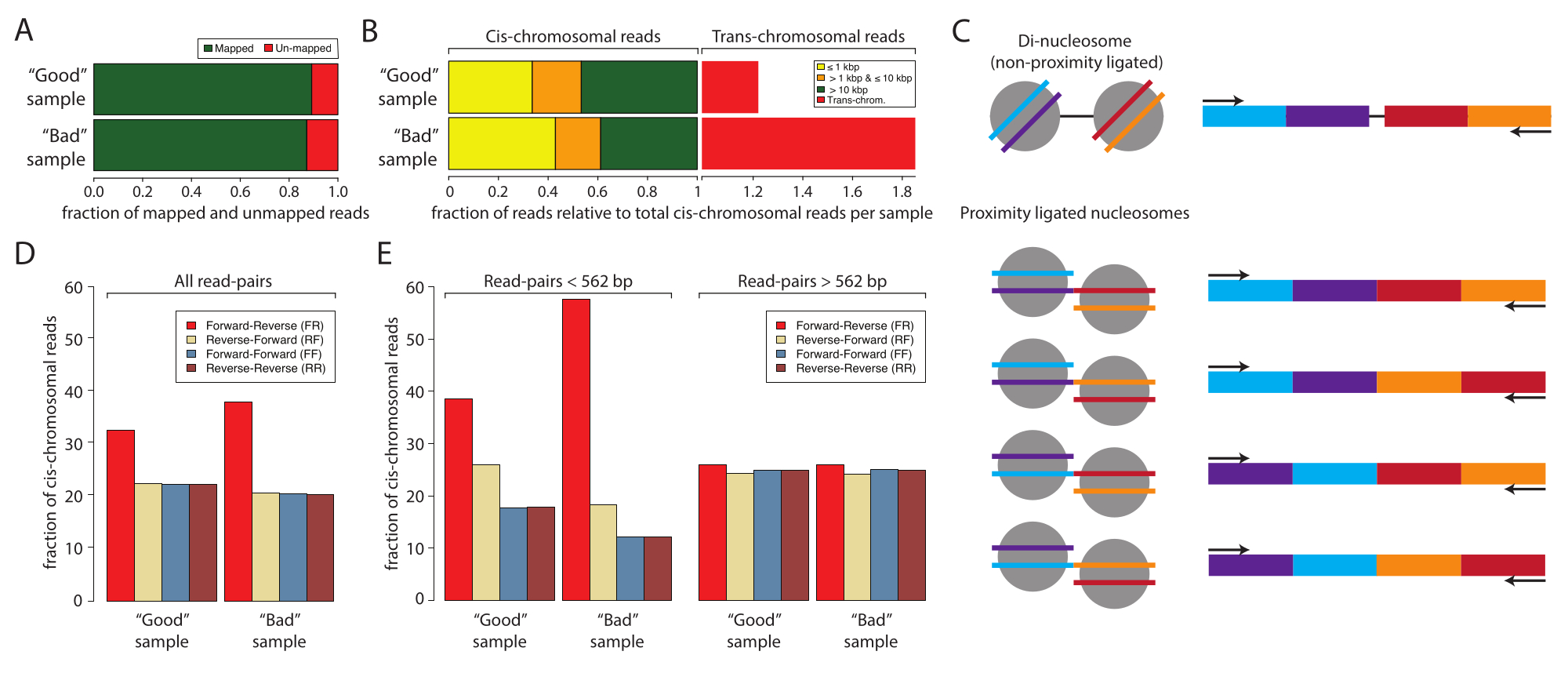
לאחר מכן, מומלץ לנתח ולאשר הכנת דגימת Micro-C מוצלחת על ידי ריצוף בעל קלט נמוך לפני שמתחייבים לריצוף עמוק עתיר משאבים. בדרך כלל, ספריות מרוצפות לעומק קריאה של 5 x 106 עד 1 x 107 ומוערכות על סמך הקריטריונים הבאים: קצב שכפול קריאת הרצף, קצב האינטראקציה הטרנס-כרומוזומלית לעומת CIS, ותדירות כיוון הקריאה הרצף. ספריות Micro-C מעובדות באמצעות Distiller, צינור בשירות מלא המעבד את הנתונים החל מרצף קבצי קריאה (פורמט Fastq) לקובצי זוגות קריאה (פורמט Bedpe) ומטריצות אינטראקציה ניתנות להרחבה (פורמטים Cool ו-Mcool) באמצעות cooler, pairtools ו-cooltools10,11,12. הצינור גם מייצר קובץ סיכום אידיאלי להערכת האיכות של ספריות Micro-C10 (https://github.com/open2c/distiller-nf). קצב שכפול ה-PCR מספק מידע על מורכבות ספריית הרצף וניתן לחלץ אותו מקובץ *.stats שנוצר. לספריות Micro-C באיכות גבוהה יש שיעורי שכפול PCR של פחות מ-5%-10% כאשר הן מיוצרות מ-5 מיליון תאים או יותר. יש לציין כי חלק מפלטפורמות הרצף מייצרות כפילויות PCR במהלך היווצרות אשכולות ללא תלות במורכבות ספריית הרצף. איור 2A מראה את שיעורי הכפילות היחסיים של שני ניסויים: אחד שאנו מחשיבים כמדגם טוב ואחד מדגם רע. בדוגמה זו, שתי הדגימות הציגו שיעורי מפה קבילים. הקריטריון הבא להערכת האיכות של ספריות מיקרו-C הוא יחס cis לעומת טרנס ותדרי כיוון קריאה. בתוך הגרעין, כרומוזומים מאכלסים טריטוריות כרומוזומים בודדות, ולכן לעתים רחוקות אינטראקציה עם כרומוזומים אחרים. שיעור גבוה של אינטראקציות טרנס-כרומוזומליות שזוהו מצביע על שיעור גבוה של קשירה אקראית. יש לציין שברמת ניתוח זו, הדגימה הרעה הראתה שיעור גבוה של אינטראקציות טרנס-כרומוזומליות בהשוואה למדגם הטוב (איור 2B). עבור Micro-C, שיעור אינטראקציה cis-כרומוזומלי של 70% ומעלה רצוי.
לספריית מיקרו-C יש גודל מקטע הדומה לפס הדנ"א הדי-נוקלאוזומי, שיכול לטהר יחד עם דגימת קשירת הקרבה ולזהם את הניסוי. זיהומים אלה הם תמיד אינטראקציות cis-כרומוזומליות. לכן, חשוב להעריך גם את שיעורי כיוון הקריאה. ניתן להעריך את שיעור הזיהום הדי-נוקלאוזומלי על ידי ריצוף בעל קלט נמוך. DNA די-נוקלאוזומלי נובע משני נוקלאוזומים שכנים שלא נבקעו על ידי MNase. לפיכך, קריאות הרצף המתקבלות יציגו תמיד כיוון קריאה קדימה-הפוך (F ו- R), והמרחק בין זוגות הקריאה יהיה סביב 320 bp. לשם השוואה, ניתן לקשור מקטעים בעלי קרבה בארבעה כיוונים, וליצור זוגות קריאה עם F-R, R-R, R-F ו-F-F, באופן אידיאלי עם שפע שווה (איור 2C). בנוסף, הם מציגים מרחקים שונים בין שני זוגות הקריאה. כדי להעריך את כמות הזיהומים הדו-נוקלאוזומליים ניתן לחשב את תדירות כיווני הקריאה מקובצי *סטטיסטיקה שנוצרו על-ידי המזקקה (איור 2D). יש לציין שבעבודה זו, החלק של קריאות F-R (אדום) היה גבוה יותר במדגם הרע בהשוואה למדגם הטוב, וזה נעשה ברור יותר כאשר כיווני הקריאה היו מרובדים לפי מרחק (איור 2E). שבר F-R נשלט על ידי מקטעים די-נוקלאוזומליים בהשוואה לספריות מיקרו-C כאשר זוגות הקריאה מרובדים לקריאות עם מרחקים <562 bp או ≥562 bp. כאן, שבר הקריאות עם מרחק <562 bp נשלט על ידי קריאות F-R, ואילו השבר עם מרחקים ≥562 bp מציג התפלגות שווה בין ארבעת הכיוונים האפשריים, מה שמצביע על כך שייצוג היתר העולמי של קריאות F-R נובע ממזהמים די-נוקלאוזומליים. הבחירה של 562 bp כסף עבור תת-הגדרה מוגדרת על-ידי binning בקובץ *stats שנוצר. למרות שאין צורך בבקרת איכות זו, ניתן להשיג תת-הגדרה מוגדרת יותר על ידי חילוץ המרחקים מקובץ הזוגות *, שנוצר גם על ידי מזקקה. חשוב לציין כי קריאות דו-נוקלאוזומליות אינן מפחיתות את איכות דגימת המיקרו-C מכיוון שניתן לזהות אותן ולהתעלם מהן במהלך עיבוד הנתונים. עם זאת, הם אינם מכילים מידע רב ערך על אינטראקציות תלת ממדיות, והם מדללים את הקריאות האינפורמטיביות.
לפיכך, טיטרציה זהירה של MNase ובקרת איכות יסודית עם רצף קלט נמוך הם הכלים הטובים ביותר לייעל את איכות ניסויי Micro-C.

איור 1: שלבי ביניים של פרוטוקול Micro-C . (A) אלקטרופורזה של ג'ל אגרוז של כרומטין מ 2.5 x 105 תאי ES עכבר מעוכלים עם ריכוזי MNase משתנים. הפסים מונו, די ותלת-נוקלאוזומלי מסומנים בחצים. M: סולם DNA (נתיב 1/6); 10U של MNase לכל 250,000 תאים (נתיב 2); 2.5U של MNase לכל 250,000 תאים (נתיב 3); 0.625 U של MNase לכל 250,000 תאים (נתיב 4); 0.156 U של MNase לכל 250,000 תאים (נתיב 2). (B) אלקטרופורזה של ג'ל אגרוז 1.0% של הדגימות המוכנות של Micro-C (נתיב 3 ונתיב 4) ובקרת הקלט המעוכלת MNase (נתיב 1). נתיב 1 ונתיב 2 (M: סולם DNA) משופרים כדי להדגיש את השינוי היחסי בעוצמת המקטעים מונו-נוקלאוזומליים עד דו-נוקלאוזומליים. הפסים מונו-נוקלאוזומליים ודי-נוקלאוזומליים מסומנים על ידי חצים. הרצועה הדי-נוקלאוזומלית בדגימת קשירת הקרבה משלבת DNA של ספריית di-nucleosomal ו-Micro-C. (C) אלקטרופורזה של ג'ל אגרוז 1.0% של ספריות ריצוף Micro-C מוגברת מדגימה של 1 μL כדי להעריך את האיכות. נתיב 1 (M): סולם DNA; נתיב 2 (S): ספריית Mirco-C. (D) עקבות מנתח הפרגמנטים של ספריית Micro-C הסופית. אנא לחץ כאן כדי להציג גרסה גדולה יותר של איור זה.
{kind=link}

איור 2: סטטיסטיקה מדגמית עבור ריצוף קלט נמוך של מדגם טוב ומדגם רע . (A) תרשים עמודות של האחוזים הממופים (ירוק) והלא ממופים (אדום) קוראים. (B) מקטע מנורמל של קריאות מיפוי סיס ואינטראקציות טרנס-כרומוזומליות. מערכי הנתונים נורמלו לקריאת מיפוי CIS. קריאות המיפוי של CIS רובדו לפי המרחק בין הקריאה הראשונה לקריאה השנייה של הדגימות המרוצות הזוגיות: ≤1 kbp (צהוב), >1 kbp ו- ≤10 kbp (כתום), ו- >10 kbp (אדום). (C) סכמה של המינים המולקולריים הפוטנציאליים עם הגדלים הדו-נוקלאוזומלים. (D) אחוזים של כיווני קריאה זוגית של כל הקריאות של המדגם הטוב והמדגם הרע. (E) זהה ללוח (D) אך מרובד לפי מרחקים (משמאל, <562 bp ומימין, ≥562 bp). אנא לחץ כאן כדי להציג גרסה גדולה יותר של איור זה.
{kind=link}
| רכיבים | 1x | פי 4.4 |
| 10x NEBuffer 2.1, | 10 מיקרוליטר | 44 מיקרוליטר |
| 2 μL 100 mM ATP | 2 μL | 8.8 מיקרוליטר |
| 100 mM DTT | 5 מיקרוליטר | 22 מיקרוליטר |
| ח2O | 68 מיקרוליטר | 299.2 מיקרוליטר |
| 10 U/μL T4 PNK | 5 מיקרוליטר | 22 מיקרוליטר |
| סך | 90 מיקרוליטר | 396 μL |
טבלה 1: תערובת מאסטר מיקרו-C 1. הרכב תערובת האב לתגובת הלעיסה הסופית.
| רכיבים | 1x | פי 4.4 |
| 1 מ"מ ביוטין-dATP | 10 מיקרוליטר | 44 מיקרוליטר |
| 1 מ"מ ביוטין-dCTP | 10 מיקרוליטר | 44 מיקרוליטר |
| תערובת של 10 mM של dTTP ו-dGTP | 1 μL | 4.4 מיקרוליטר |
| 10x T4 DNA Ligase Buffer | 5 מיקרוליטר | 22 מיקרוליטר |
| 200x BSA | 0.25 מיקרוליטר | 1.1 מיקרוליטר |
| ח2O | 23.75 מיקרוליטר | 104.5 מיקרוליטר |
טבלה 2: מיקרו-C מאסטר מיקס 2. הרכב תערובת האב לתגובת התיוג הסופית.
| רכיבים | 1x | פי 4.4 |
| 10x NEB T4 מאגר תגובה Ligase | 50 מיקרוליטר | 220 מיקרוליטר |
| ח2O | 422,5 μL | 1859 מיקרוליטר |
| ליגאז DNA T4 | 25 מיקרוליטר | 110μL |
טבלה 3: מיקרו-C מאסטר מיקס 3. הרכב תערובת האב לתגובת קשירת הקרבה.
| רכיבים | 1x | פי 4.4 |
| 10x NEBuffer 1.1 | 20 מיקרוליטר | 88 מיקרוליטר |
| ח2O | 180 מיקרוליטר | 792 מיקרוליטר |
| נוקלאז ExoIII | 10 מיקרוליטר | 44 מיקרוליטר |
טבלה 4: מיקרו-C מאסטר מיקס 4. הרכב תערובת האב לתגובת הסרת ביוטין.
Discussion
הצלחתו של ניסוי Micro-C תלויה בכמה שלבים קריטיים בפרוטוקול שיש לבצע בזהירות. ראשית, crosslinking עם crosslinker נוסף DSG או EGS יכול להוביל צבירה של תאים, בהתאם לסוג התא. הוספת BSA של 0.1%-0.5% לתגובת הקרוסלינקינג מפחיתה באופן משמעותי את הצבירה מבלי להשפיע על יעילות הקרוסלינקינג. הצלבה לא יעילה עלולה לגרום לשיעורים מוגברים של אינטראקציות טרנס-כרומוזומליות המעידות על קשירה אקראית. השלב השני, אך החשוב ביותר, בפרוטוקול זה הוא עיכול הכרומטין עם MNase. עיכול כרומטין תת-אופטימלי מוביל לקשירת קרבה לא יעילה (עיכול יתר) או לשיעורים מוגברים של די-נוקלאוזומים שאינם קשורים לקרבה (תת-עיכול). ניתן להעריך את יעילות תגובת הקשירה על-ידי אלקטרופורזה של ג'ל אגרוז (איור 1B), ובנוסף ניתן להעריך אותה בצורה הטובה ביותר על-ידי ריצוף בעל קלט נמוך. אם ריצוף הקלט הנמוך מגלה שיעור שכפול גבוה (קשירה לא יעילה) או קצב די-נוקלאוזומים מוגבר, יש להעריך מחדש את דרגת העיכול של MNase. יש לציין כי אובדן הדגימה בעת הפעלת הפרוטוקול יכול להוביל להפחתת מורכבות הספרייה. ריכוז הדגימה מוערך בצורה הטובה ביותר לאחר טיהור DNA (שלב 5.3) או על ידי PCR מינימלי (שלב 8). התשואה הכוללת של DNA מ 5 x 106 תאי יונקים לאחר טיהור DNA הוא בדרך כלל >2 מיקרוגרם. ריכוז ה- DNA צריך להיות מבוקר לאחר עיכול MNase, עיכול ExoIII וטיהור DNA. נוקלאזות אנדוגניות, שהשפע שלהן הוא ספציפי לסוג התא וספציפי למין, יכול להיות מקור לפירוק DNA. בנוסף, טיהור DNA מבוסס עמודות יכול להוביל לאובדן דגימה עקב חוסר תאימות עם SDS מתגובות דה-פרוטאינציה. משקעי אתנול יכולים להיחשב אם ריכוז ה- DNA נמוך בשלב זה.
מכיוון שמיקרו-C דורש טיטרציה ספציפית של MNase, מאתגר ליישם מיקרו-C על אוכלוסיות תאים קטנות, כגון עם איברים קטנים מאורגניזמים שונים במודל, עוברים ותאים בודדים, אורגנואידים או ביופסיות של מטופלים. כאן, Hi-C 3.0 מציע חלופה מבוססת היטב באמצעות תגובת נקודת קצה על ידי אנדונוקלאז הגבלה ספציפי לרצף 8,9.
Micro-C היא טכנולוגיית קונפורמציה של כרומוזומים ברזולוציה גבוהה עם טווח דינמי גבוה ויחס אות לרעש נמוך, מה שהופך אותה למתאימה במיוחד לחקר תכונות כרומוזום לטווח קצר 4,5,8, כגון לולאות כרומוזומים. הרזולוציה של Micro-C מאפשרת לכידת לולאות מקדם-מקדם, שהן מעבר לגבול הזיהוי של Hi-C, ומאפשרת ניתוח מפורט יותר של הקשר בין ארגון הגנום לבין רגולציה13,14,15. יתר על כן, אסטרטגיות לכידת DNA שולבו לאחרונה עם Micro-C כדי להגדיל את הרזולוציה הספציפית למוקד של האתרים הגנומיים הממוקדים לרמות חסרות תקדים, וחשפו תובנות חדשות על מבנה העל של הגנום התלת-ממדי16,17,18. לסיכום, אנו צופים כי Micro-C ונגזרותיו יהוו טכנולוגיית מפתח לניתוח תפקידו של הגנום התלת-ממדי בוויסות שעתוק, וכתוצאה מכך, התמיינות ותחזוקת סוג התא.
Disclosures
למחברים אין מה לחשוף.
Acknowledgements
אנו מודים לכריסטל גאוביץ' ולקתלין סטיוארט-מורגן על קריאתם הביקורתית של כתב היד. אנו מודים לאניה גרות' ולמעבדת גרות' על תמיכתן בהקמת המעבדה שלנו. אנו מודים לצוות פלטפורמת החייאה/reNEW Genomics על התמיכה: ה. וולמן, מ. מיכאוט וא. קלוויזה. המרכז לרפואת תאי גזע של קרן נובו נורדיסק (reNEW) נתמך על ידי מענק מספר NNF21CC0073729 של קרן נובו נורדיסק. מרכז נובו נורדיסק לחקר חלבונים (CPR) נתמך על ידי מענק מספר NNF14CC0001 של קרן נובו נורדיסק. אנו מודים למעבדת בריקמן במרכז נובו נורדיסק לרפואת תאי גזע, reNEW קופנהגן, עבור תאי ES של עכבר.
Materials
| Name | Company | Catalog Number | Comments |
| 1 mM Biotin dATP | Jenna Bioscience | NU-835-Bio14-S | |
| 1 mM Biotin dCTP | Jenna Bioscience | NU-809-BioX-S | |
| 10 mM dGTP | NEB | N0442S | |
| 10 mM dTTP | NEB | N0443S | |
| 10 U/ml T4 PNK | NEB | M0201L | |
| 100 U/L Exonuclease III | NEB | M0206L | |
| 10x NEBuffer 1.1 | NEB | B7001S | |
| 10x NEBuffer 2.1 | NEB | B7202S | |
| 10x T4 DNA Ligase buffer | NEB | B0202A | |
| 1x DPBS w/o Mg2+ and Ca2+ | ThermoFisher | 14190144 | |
| 1x LIF | |||
| 2_Mercaptoethanol 50 mM | Gibco | 31350010 | 0.1 mM b-mercaptoethanol |
| 37% Formaldehyde | Sigma Aldrich | 252549-500ML | Caution. See manufactures MSDS |
| 400 U/ml T4 DNA Ligase | NEB | M0202L | |
| 5 U/ml Klenow Fragment | NEB | M0210L | |
| Agarose | BIO-RAD | 1613102 | Caution. See manufactures MSDS |
| BSA 20mg/ml | NEB | B9000S | |
| CaCl2 | |||
| cell counter | |||
| Dimethyl Sulfoxide (DMSO) | Sigma Aldrich | D8418-100ML | Caution. See manufactures MSDS |
| Dynabeads MyOne Streptavidin C1 | Invitrogen | 65001 | |
| DynaMag-2 Magnet | Invitrogen | 12321D | refered to as: magnet magnet for 1.5 ml tubes |
| DynaMag-PCR Magnet | Invitrogen | 492025 | refered to as: magnet magnet for PCR tubes |
| EDTA Ultrapure 0.5M pH 8.0 | Invitrogen | 15575-038 | |
| EGTA Ultrapure 0.5M pH 8.0 | BioWorld | 40121266-1 | |
| Ethanol 96% | VWR Chemicals | 20824365 | quality control system |
| Ethidium Bromide | Invitrogen | 15585-011 | |
| Ethylene glycol bis(succinimidyl succinate) (EGS) | ThermoFisher | 21565 | |
| Fetl Bovin Serum | Sigma Aldrich | F7524 | 15% FBS |
| Gel Loading dye purple (6X) | NEB | B7024S | |
| Glycine | PanReac AppliChem | A1067.0500 | |
| Halt Proteinase inhibitor (100x) | ThermoFisher | 78430 | Caution. See manufactures MSDS |
| IGEPAL CA-630 (NP-40) | Sigma Aldrich | 18896-50ML | |
| MgCl 1 M | Invitrogen | AM9530G | |
| Micrococcal Nuclease (MNase) | Worthington | LS004798 | |
| mouse embryonic stem cells | |||
| NaCl | Sigma Aldrich | S9888-1KG | |
| NEBNext Multiplex Oligos for Illumina (Dual Index primers) | NEB | E7600S | amplification primers for sequencing libraries |
| NEBNext Ultra II DNA library prep kit for Illumina | NEB | E7645L | sequencing library preparation kit |
| NEBNext Ultra II Q5 Master mix | NEB | M0544S | Caution. See manufactures MSDS |
| Non-Essential Amino Acids Solution | Gibco | 11140050 | 1x NEAA |
| Penicillin-Streptomycin (10,000 U/mL) | Gibco | 15140148 | 1% Pen-Strep |
| Proteinase K (40 mg/ml) | GoldBio | P-480-1 | Caution. See manufactures MSDS |
| QIAquick Gel extraction kit | QIAgen | 28706 | refered to as: DNA gel elution kit |
| QIAquick PCR purification kit | QIAgen | 28106 | refered to as: commercial DNA purification kit |
| Qubit dsDNA HS Assay kit | Invitrogen | Q32854 | high sensitivity DNA quantification instrument |
| Quick load purple 1kb plus DNA Ladder | NEB | N0550S | |
| SPRIselect size selection beads | Beckman Coulter | B23319 | paramagnetic beads |
| ThermoMixer C | Eppendorf | 5382000015 | refered to as: thermomixer |
| Tris | Merck | 10708976001 | |
| Trypsin | |||
| Tween20 | Sigma Aldrich | P7949-100ML | |
| Ultrapure 10% SDS | Invitrogen | 15553-035 | |
| Ultrapure Phenol Chloroform Isoamyl Alcohol (PCI) | Invitrogen | 15593-031 | |
| Fragment Analyzer |
References
- Lieberman-Aiden, E., et al. Comprehensive mapping of long-range interactions reveals folding principles of the human genome. Science. 326 (5950), 289-293 (2009).
- Hsieh, T. -. H. S., Fudenberg, G., Goloborodko, A., Rando, O. J. Micro-C XL: Assaying chromosome conformation from the nucleosome to the entire genome. Nature Methods. 13 (12), 1009-1011 (2016).
- Hsieh, T. -. H. S., et al. Mapping nucleosome resolution chromosome folding in yeast by Micro-C. Cell. 162 (1), 108-119 (2015).
- Krietenstein, N., et al. Ultrastructural details of mammalian chromosome architecture. Molecular Cell. 78 (3), 554-565 (2020).
- Hsieh, T. -. H. S., et al. Resolving the 3D landscape of transcription-linked mammalian chromatin folding. Molecular Cell. 78 (3), 539-553 (2020).
- Belaghzal, H., Dekker, J., Gibcus, J. H. Hi-C 2.0: An optimized Hi-C procedure for high-resolution genome-wide mapping of chromosome conformation. Methods. 123, 56-65 (2017).
- van Holde, K. E. . Chromatin. , (1989).
- Oksuz, B. A. Systematic evaluation of chromosome conformation capture assays. Nature Methods. 18, 1046-1055 (2021).
- Lafontaine, D. L., Yang, L., Dekker, J., Gibcus, J. H. Hi-C 3.0: Improved protocol for genome-wide chromosome conformation capture. Current Protocols. 1 (7), 198 (2021).
- Goloborodko, A., Venev, S., Abdennur, N., Di Tommaso, P. Mirnylab/distiller-nf; v033. Zenodo. , (2019).
- Venev, S., et al. Open2c/cooltools: v0.4.1. Zenodo. , (2021).
- Abdennur, N., Mirny, L. A. Cooler: Scalable storage for Hi-C data and other genomically labeled arrays. Bioinformatics. 36 (1), 311-316 (2019).
- Zhang, S., Übelmesser, N., Barbieri, M., Papantonis, A. Enhancer-promoter contact formation requires RNAPII and antagonizes loop extrusion. BioRxiv. , (2022).
- Barshad, G., et al. RNA polymerase II and PARP1 shape enhancer-promoter contacts. BioRxiv. , (2022).
- Hansen, A. S., et al. Distinct classes of chromatin loops revealed by deletion of an RNA-binding region in CTCF. Molecular Cell. 76 (3), 395-411 (2019).
- Hua, P., et al. Defining genome architecture at base-pair resolution. Nature. 595 (7865), 125-129 (2021).
- Downes, D. J. High-resolution targeted 3C interrogation of cis-regulatory element organization at genome-wide scale. Nature Communications. 12, 531 (2021).
- Goel, V. Y., Huseyin, M. K., Hansen, A. S. Region capture Micro-C reveals coalescence of enhancers and promoters into nested microcompartments. BioRxiv. , (2022).
Reprints and Permissions
Request permission to reuse the text or figures of this JoVE article
Request PermissionThis article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. All rights reserved