
Method Article
طرق محسنة لإعداد المقاطع المستعرضة وحوامل كاملة غير ملفوفة من أوراق الذرة الأولية للتصوير الفلوري والبؤري
In This Article
Summary
أوراق الذرة البدائية مغلفة بعمق وملفوفة ، مما يجعل من الصعب دراستها. هنا ، نقدم طرقا لإعداد المقاطع المستعرضة وحوامل كاملة غير ملفوفة من أوراق الذرة البدائية للتألق والتصوير متحد البؤر.
Abstract
في الذرة (Zea mays) والأعشاب الأخرى (Poaceae) ، تغلف الأوراق البدائية بعمق وتدحرجت داخل دوامة الأوراق ، مما يجعل من الصعب دراسة نمو الأوراق المبكر. هنا ، نصف طرق تحضير المقاطع المستعرضة وحوامل كاملة غير ملفوفة من أوراق الذرة البدائية للتصوير الفلوري والتصوير متحد البؤر. تستخدم الطريقة الأولى متجردا سلكيا لإزالة الأجزاء العلوية من الأوراق القديمة ، مما يعرض طرف الورقة الأولية ويسمح بقياسها لأخذ عينات أكثر دقة من المقطع العرضي. تستخدم الطريقة الثانية شريطا نانويا واضحا على الوجهين لفتح وتركيب بريمورديا كاملة الأوراق للتصوير. نعرض فائدة الطريقتين في تصور وتحليل مراسلي البروتين الفلوري في الذرة. توفر هذه الطرق حلا للتحديات التي يطرحها التشكل المميز لأوراق الذرة البدائية وستكون مفيدة لتصور وقياس السمات التشريحية والتنموية للأوراق في الذرة وأنواع العشب الأخرى.
Introduction
تعد المحاصيل العشبية مصدرا رئيسيا للغذاء والوقود الحيوي لسكان العالم1 ، وتحسين تشريح الأوراق لديه القدرة على زيادة إنتاجيتها 2,3. ومع ذلك ، فإن فهمنا الحالي لكيفية تنظيم تشريح الأوراق في الأعشاب محدود4 ويتطلب تحليل الأوراق البدائية ، حيث يتم تحديد العديد من السمات التشريحية والفسيولوجية للورقة مسبقا في وقت مبكر من التطور5،6،7. لا غنى عن تقنيات التصوير الخلوي ، مثل التألق والتصوير متحد البؤر ، لدراسة تشريح أوراق العشب والسمات الخلوية ، ولكن من الصعب تطبيق هذه التقنيات على بريمورديا أوراق العشب لأنها مغلفة بعمق وملفوفة داخل دوامة الأوراق. لقد عالجنا هذه المشكلة من خلال تطوير طرق لإعداد المقاطع المستعرضة وحوامل الأوراق الكاملة غير الملفوفة للتألق والتحليل البؤري لأوراق الذرة البدائية ، وهو نظام نموذجي لدراسة تشريح أوراق العشب وتطورها2،8.
تتكون ورقة الذرة ، مثل جميع أوراق العشب ، من شفرة تشبه الشريط مع غمد يلتف حول الساق ويتطور في الساق 9،10،11،12،13. تتطور الأوراق من المرستيم القمي للساق (SAM) في نمط distichous ، حيث تبدأ كل ورقة جديدة في الموضع المعاكس للورقة السابقة ، مما ينتج عنه رتبتان من الأوراق على طول المحور الرأسي (الشكل 1 أ) 14 . يتم تحديد المرحلة التنموية لكل ورقة بريمورديوم من خلال موقعها بالنسبة إلى SAM ، مع تعيين أقرب بريمورديوم على أنه plastochron1 (P1) والبريمورديا التالية المعينة على أنها P2 و P3 وما إلى ذلك (الشكل 1B ، C) 2. أثناء التطور (الشكل 1D) ، تظهر ورقة البدائية أولا كدعامة على شكل هلال حول قاعدة SAM (P1) ، ثم تنمو لتصبح بريمورديوم على شكل غطاء يمتد فوق meristem (P2) 9،10،11. ثم تتوسع الحواف القاعدية للغطاء بشكل جانبي وتتداخل مع بعضها البعض مع نمو الطرف لأعلى ، مكونا بدائيا مخروطي الشكل (P3-P5) 10. ثم ينمو طول البدائية بسرعة ، وتصبح حدود شفرة الغمد في القاعدة أكثر بروزا مع تكوين الرباط ، وهو الإسقاط الشبيه بالهامش على الجانب المحوري للورقة (P6 / P7). أخيرا ، تنفتح الورقة عند خروجها من الزهرة أثناء نمو الحالة المستقرة ، حيث يتم تقييد الخلايا المنقسمة داخل المنطقة القاعدية الصغيرة للشفرة ، وتشكيل تدرج مع خلايا متوسعة ومميزة على طول المحور القريب البعيد (P7 / P8) 15. تحتوي قمة تبادل لشتلة الذرة على العديد من البدائيات في مراحل مختلفة من التطور ، مما يجعلها نموذجا ممتازا لدراسة نمو الأوراق8.
يتطلب التحليل الدقيق لتطور الأوراق المبكر التدريج أو استخدام معايير موحدة لتحديد مراحل متميزة من تطور البدائية فيما يتعلق بالنمو أو المعلمات المورفولوجية الأخرى. نظرا لأن ورقة البدائية مخفية داخل ساق العشب ، يستخدم الباحثون عادة معلمات مثل عمر النبات أو حجم الأوراق الناشئة كتنبؤات لمراحل وأحجام ورقة البدائية 9,16. في الذرة ، يتم تحديد العمر الزمني للنبات إما بعدد الأيام بعد الزراعة أو الإنبات (DAP / DAG)17,18. يتم تحديد المرحلة الخضرية (المرحلة V) من خلال الورقة العلوية ذات الياقة المرئية ، وخط شاحب على الجانب المحوري بين النصل والغمد يتوافق مع موضع الرباط والأذنين ، وزوج من المناطق على شكل إسفين عند قاعدة النصل (الشكل 1 أ ، ب)17,19 . بين 20 و 25 DAG ، ينتقل SAM إلى مرستيم الإزهار ويتوقف عن إنتاج أوراق جديدة20. يمكن أن تختلف معدلات نمو نبات الذرة البدائية اعتمادا على البيئة والنمط الوراثي للنبات. لهذا السبب ، لا يمكن لعمر النبات وحجم الأوراق الناشئة التنبؤ بدقة بأحجام الأوراق البدائية ؛ ومع ذلك ، يمكن أن يساعد استخدام هذه المعلمات في التنبؤ بنطاق مراحل وأحجام Primordia للأغراض التجريبية.
يعد تحليل المقطع المستعرض طريقة شائعة لفحص تشريح الأوراق وتطورها في الذرة والأعشاب الأخرى لأنه يسمح بأخذ عينات من البلاستوكرونات المتعددة في قسم واحد عبر الساق21،22،23. هذه الطريقة ملائمة أيضا للتصوير الخلوي للعينات الطازجة ، حيث تعمل الأوراق المحيطة كسقالة تحافظ على بدائية الأوراق في مكانها أثناء التقسيم والتركيب24. ومع ذلك ، فإن عيب هذه الطريقة هو أنه قد يكون من الصعب تحديد موقع البلاستوكرون المستهدف والمنطقة بدقة داخل البدائية عند التقسيم من لقطة سليمة. علاوة على ذلك ، نظرا لأن نمو الأوراق يختلف عبر البلاستوكرون وعلى طول المحور القريبالبعيد 2,5 ، فقد يؤدي أخذ العينات غير الدقيق إلى تفسير غير صحيح لمرحلة نمو ومنطقة البدائية في قسم معين. لذلك ، فإن تطوير طريقة لأخذ عينات دقيقة من المقطع المستعرض أمر بالغ الأهمية لضمان دقة وتكرار التحليلات التشريحية والتنموية لأوراق العشب البدائية.
يتيح تحليل جبل الورقة الكاملة التحقيق الشامل والتكاملي للأنسجة والعمليات الخلوية التي تحدث على نطاق العضو بالكامل ، مثل النمو التكاثري25 ونمط الوريد26،27،28. توفر الطريقة نظرة عامة على الورقة ، مما يسمح باكتشاف عمليات وأنماط مميزة يصعب اكتشافها باستخدام تحليل المقطع العرضي24،27. على عكس أرابيدوبسيس ، حيث توجد بالفعل طرق راسخة لتصوير حوامل الأوراق الكاملة29,30 ، لا توجد حاليا طريقة قياسية لتصوير حوامل الأوراق الكاملة غير المطوية في الأعشاب. تضمن بروتوكول سابق لفك أوراق الذرة المعزولة مواد غير شائعة ولم يكن مناسبا للتصوير الخلوي31. يمكن لتقنيات التصوير المتقدمة ، مثل التصوير المقطعي المحوسب (CT) والتصوير بالرنين المغناطيسي (MRI) ، الحصول على معلومات تشريحية ثلاثية الأبعاد دون عزل وفتح البدائية11،32،33 ، لكنها باهظة الثمن وتتطلب معدات متخصصة. إن تطوير تقنية للتغلب على القيود التي يفرضها التشكل المدلفن والمخروطي للأوراق البدائية في الذرة والأعشاب الأخرى من شأنه أن يعزز التحقيقات في سماتها التشريحية والتنموية.
هنا ، نقدم طرقا لإعداد المقاطع المستعرضة وحوامل كاملة غير ملفوفة من أوراق الذرة البدائية للتألق والتصوير متحد البؤر. استخدمنا هذه الطرق لتحديد عدد الوريد ورسم خريطة لتوزيع الهرمونات المكانية والزمانية في أوراق الذرة البدائية مع البروتينات الفلورية (FPs)24. تتضمن الطريقة الأولى إزالة الجزء العلوي من الأوراق القديمة من شتلات الذرة باستخدام متجرد سلكي (الشكل 1E). من خلال تعريض طرف البدائية (P5-P7) ، يصبح من الممكن تحديد طوله دون الحاجة إلى إزالة الأوراق المحيطة القديمة تماما ، مما يتيح التقسيم السهل والدقيق. تتضمن الطريقة الثانية فك وتركيب بريمورديا كاملة الأوراق (P3-P7) بشريط نانو شفاف على الوجهين (الشكل 1F). هذه الطرق مناسبة لتصور مختلف FPs24 ولكنها تحتاج إلى تحسين لاستخدام الأصباغ الفلورية وكواشف الإزالة. بالإضافة إلى ذلك ، نوضح بعض الإجراءات لتسطيح مكدسات z ، وخياطة الصور ، ودمج القنوات في ImageJ / FIJI34 ، والتي تنطبق على الصور التي تنتجها الطريقتان. هذه الطرق مفيدة للتألق الروتيني أو التصوير البؤري لأوراق الذرة ، ولكن يمكن أيضا تكييفها مع أنواع العشب النموذجية الأخرى ، مثل الأرز و Setaria و Brachypodium.

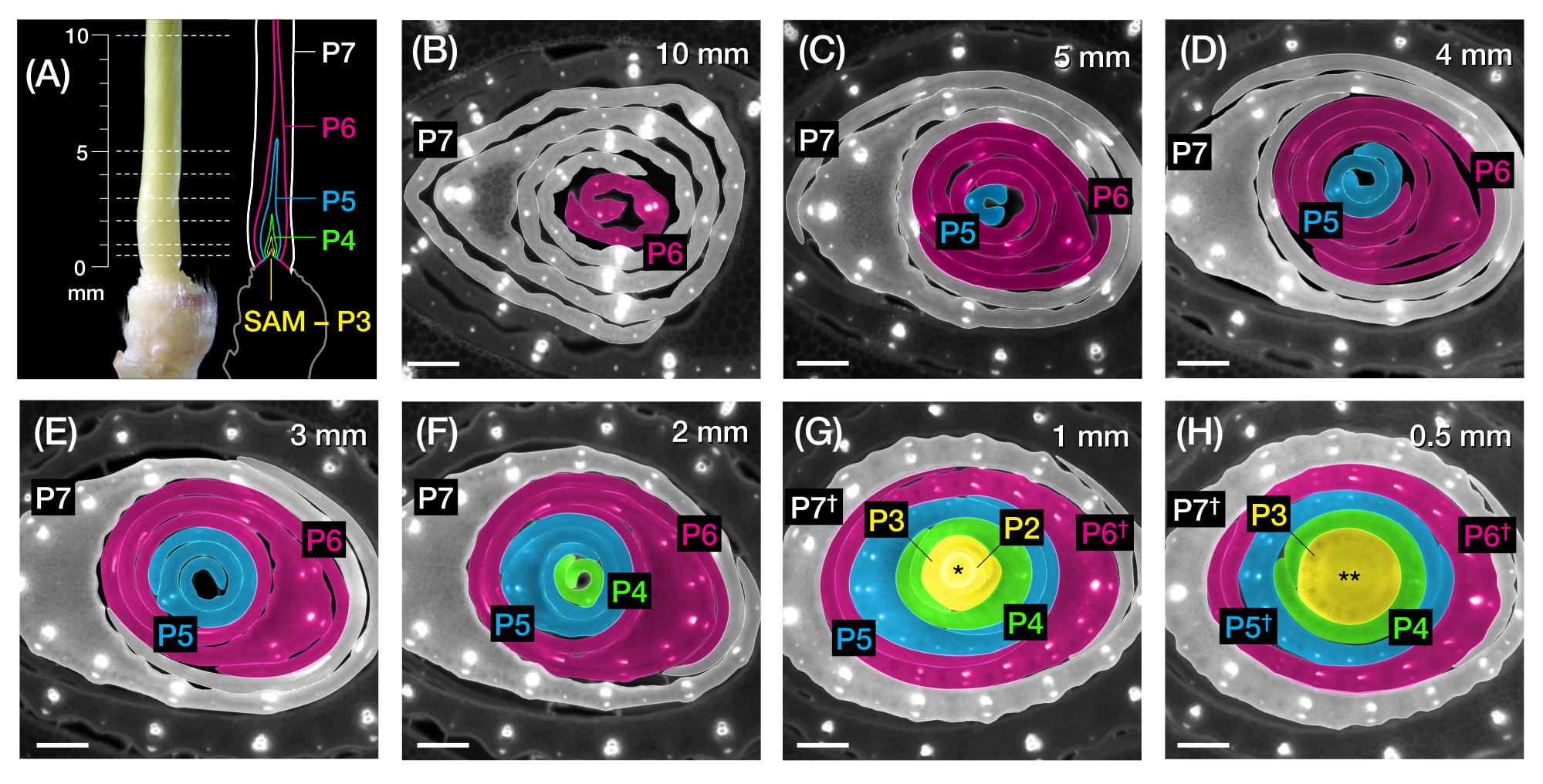
الشكل 1: تنظيم ومورفولوجيا أوراق الذرة البدائية ونظرة عامة على الطرق . (أ) تمثيل تخطيطي لشتلة الذرة. الذرة لها سلالة ديستيكوس ، حيث تبدأ الورقة الجديدة في الموضع المعاكس للورقة السابقة. يشير رقم الورقة إلى الترتيب الزمني الذي ظهرت به الأوراق من الإنبات (أي الورقة الأولى ، L1 ؛ الورقة الثانية ، L2 ؛ الورقة الثالثة ، L3 ؛ إلخ). تحتوي كل ورقة على شفرة بعيدة وغمد قاعدي محدد بواسطة طوق يتوافق مع الرباط والأذن. تشير الورقة العلوية ذات الياقة المرئية إلى المرحلة الخضرية. الشتلات في هذا المثال في مرحلة V2 ، مع طوق L2 (رأس السهم) مرئية. يشير رمز المقص إلى الموقع في mesocotyl (أنا) حيث يجب قطع الشتلات من أجل جمعها. (ب) تمثيل تخطيطي للساق المقطعة يظهر L1 إلى L4 معزولا، مع عرض الورقة الأولية L5 إلى L9 كصورة مكبرة في (C). يشير رقم البلاستوكرون إلى موضع البدائية بالنسبة إلى SAM ، مع أصغر ورقة بريمورديوم (P1) الأقرب إلى SAM والأوراق الأقدم بريمورديا (P2 ، P3 ، P4 ، وهكذا) على التواليأبعد 2. (د) تمثيل تخطيطي لمورفولوجيا أوراق الذرة الأولية من P1 إلى P5. (ه) نظرة عامة تخطيطية لطريقة تحليل المقطع المستعرض لبدائية أوراق الذرة. (1) تقليم الأوراق القديمة باستخدام متجرد سلكي. (2) قياس البدائية وقسم تبادل لاطلاق النار. (3) قم بتركيب القسم على شريحة للتصوير والمعالجة (4 ، 5). (و) نظرة عامة تخطيطية لطريقة التحليل الكامل لأوراق الذرة البدائية. (1) قم بإزالة الأوراق المحيطة لاستخراج البدائية. (2) قص وفك البدائية المسطحة على شريط النانو. (3) تركيب العينة للتصوير والمعالجة (4 ، 5). الاختصارات: L = ورقة ؛ bl = شفرة ؛ ش = غمد ؛ ثاني أكسيد الكربون = طوق ؛ أنا = ميزوكوتيل. V = نباتي ؛ P = بلاستوكرون ؛ SAM = تبادل لاطلاق النار meristem القمي. يرجى النقر هنا لعرض نسخة أكبر من هذا الرقم.
{kind=link}
Protocol
1. تنظيم تطوير أوراق الذرة وتصميم التجربة
- زراعة النباتات وتنظيم تطوير الأوراق
- تحديد العمر ومرحلة النمو للنباتات التي سيتم استخدامها للتجربة وإجراء مرحلة مفصلة لتطور الأوراق.
ملاحظة: يوصى بإجراء التدريج على النباتات التي لها نفس الخلفية الوراثية مثل الخطوط الطافرة أو المعدلة وراثيا التي سيتم استخدامها في التجارب. يتوقف SAM عن إنتاج أوراق جديدة بين 20 و 25 DAG ، اعتمادا على الخلفية الوراثية وظروف النمو. لهذا السبب ، فإن الطرق الموضحة هنا مثالية لشتلات الذرة التي تتراوح أعمارها بين 7-14 DAG. - قم بزراعة نباتات الذرة في دفيئة أو غرفة نمو باتباع بروتوكول مناسب للعناية بالنباتات35.
- اجمع النباتات في العمر المطلوب أو المرحلة V بسكين صغير أو مقص عن طريق قطع الميزوكوتيل ، وهو الجذع الموجود أسفل سطح التربة (الشكل 1 أ). أدخل أداة القطع بعناية في التربة بجانب الشتلات وحركها لأسفل إلى القاعدة بزاوية 45 درجة لقطع الميزوكوتيل.
- ارفع الشتلات من التربة وقم بإزالة أي أوساخ أو جزيئات تربة متبقية قد تتشبث بالنبات. قم بإزالة الغلاف الورقي ، وهو الغلاف الواقي الذي يغطي اللقطة الناشئة في الشتلات الصغيرة ، يدويا.
- لكل نبات ، سجل المرحلة V وعدد الأوراق وطول الورقة الأخيرة الخارجة من الزهرة. التقط صورا للنباتات للرجوع إليها في المستقبل.
- قم بإزالة الأوراق القديمة واحدة تلو الأخرى. للقيام بذلك ، أمسك النبات ببقايا الميزوكوتيل أو الساق ، واستأصل كل ورقة من قاعدة الغمد وافتح الغمد برفق بطريقة دائرية باستخدام مسبار أسنان (الشكل 1 ب).
ملاحظة: راجع الخطوة 2.1 للحصول على طريقة سريعة لإزالة الأوراق المحيطة. - تحت مجهر ستيريو عند تكبير 2x تقريبا ، قم بتشريح بقية قمة اللقطة بعناية على مناديل مبللة لمنع العينة من الجفاف. قم باستئصال كل ورقة بدائية برفق وفتحها باستخدام مسبار أسنان بإبرة منحنية أو زوج من الملقط الدقيق حتى تصبح SAM و P1 و P2 مرئية (الشكل 1 د). سجل مظهر وقياس كل بلاستوكرون.
ملاحظة: انظر مثالا على تطور الأوراق في شتلات الذرة في الملف التكميلي 1.
- تحديد العمر ومرحلة النمو للنباتات التي سيتم استخدامها للتجربة وإجراء مرحلة مفصلة لتطور الأوراق.
- التخطيط للتجربة
ملاحظة: يمكن أن يؤدي التخطيط للتجربة بعناية إلى تحسين الكفاءة. على سبيل المثال ، في تحليل المقطع المستعرض ، يمكن أن يؤدي تحديد الموقع التقريبي للقطع عند تصوير بلاستوكرونات أو مناطق معينة إلى تقليل وقت التشريح. يوضح الشكل 2 مثالا على أخذ العينات القياسي. علاوة على ذلك ، يمكن أن يؤدي تحسين معلمات التصوير بناء على خصائص FPs أو مجسات الفلورسنت إلى تحسين كفاءة التجارب.- استخدم معلومات التدريج كدليل لتركيز التجربة على بلاستوكرونات محددة ومناطق بدائية وأنسجة. استخدم FPs أو مجسات الفلورسنت الأخرى ذات الأهمية لمتابعة الأقسام 2 و / أو 3 أدناه في بعض العينات.
ملاحظة: انظر مخزون بذور علامة FP المتاح من الذرة 36-42 عند https://www.maizegdb.org/data_center/stock. - حدد المعلمات المثلى للحصول على الصور لكل مراسل أو مسبار FP ، بما في ذلك الكاشف أو المرشح المناسب ، والأطوال الموجية للإثارة والانبعاث ، وحجم الثقب ، والإعدادات الأخرى. احفظ الإعدادات لإعادة إنتاج ظروف العمل لكل تجربة.
ملاحظة: انظر Linh and Scarpella30 للحصول على بعض الإرشادات الفنية للمسح المجهري بالليزر متحد البؤر للأوراق النامية. - خطط لوقت التصوير بناء على عمر النصف والخصائص الأخرى لمراسل FP43.
ملاحظة: يجب أيضا مراعاة الوقت بين التشريح والتصوير عند التخطيط للتجربة ، حيث تتعرض عينات النباتات الطازجة للجفاف والتغيرات الخلوية خلال هذه الفترة.
- استخدم معلومات التدريج كدليل لتركيز التجربة على بلاستوكرونات محددة ومناطق بدائية وأنسجة. استخدم FPs أو مجسات الفلورسنت الأخرى ذات الأهمية لمتابعة الأقسام 2 و / أو 3 أدناه في بعض العينات.

الشكل 2: مخطط أخذ العينات لتحليل المقطع العرضي لأوراق الذرة البدائية. (أ، يسار) نبتة قريبة من شتلة ذرة 7 DAG ، تظهر ورقة رابعة مكشوفة (L4) في P7. تشير الخطوط المتقطعة إلى نقاط أخذ العينات السبع على طول البدائية ، من 0.5 مم إلى 10 مم. (أ ، يمين) تمثيل تخطيطي للورقة البدائية ، مع الحجم والموضع المتوقعين لكل بلاستوكرون: P7 (أبيض) ؛ P6 (أرجواني)؛ P5 (أزرق) ؛ P4 (أخضر) ؛ و P3 حتى SAM (أصفر). (ب-ح) صور مضان للمقاطع المستعرضة التي تمثل نقاط أخذ العينات الموضحة في A من 10 مم (B) إلى 0.5 مم (H). البدائية هي ألوان زائفة وفقا لنظام ألوان البلاستوكرون في (أ). تم تصوير المقاطع باستخدام مجهر التألق باستخدام مرشح الأشعة فوق البنفسجية ذات الانبعاث الطويل للتألق الذاتي. شريط المقياس = 200 ميكرومتر (B-H). تم تعديل هذا الرقم واستنساخه بإذن من Robil و McSteen24. الاختصارات: DAG = أيام بعد الإنبات ؛ P = بلاستوكرون ؛ SAM = تبادل لاطلاق النار meristem القمي. † = غمد الأوراق أو ما قبل الرباط المناسب ؛ * = تبادل لاطلاق النار meristem القمي. ** = الجذعية. يرجى النقر هنا لعرض نسخة أكبر من هذا الرقم.
{kind=link}
2. تصوير المقاطع المستعرضة لأوراق الذرة البدائية
- تجريد الأوراق المحيطة القديمة وقياس ورقة البدائية
- اجمع الشتلات عن طريق قطعها بعناية في الميزوكوتيل تحت سطح التربة بسكين صغير أو مقص ، كما هو موضح في الخطوات 1.1.3-1.1.4 (الشكل 1 أ).
- حدد حجم فتحة متجرد الأسلاك الصحيح الذي سيتم استخدامه ومكان إجراء القطع على طول اللقطة. تأكد من أن الفتحة كبيرة بما يكفي لتناسب البدائية المستهدفة.
- ابدأ بالقطع في الجزء البعيد من الساق واعمل تدريجيا باتجاه القاعدة حتى ينكشف طرف البدائية.
- لإجراء القطع ، أمسك متجرد الأسلاك مع مواجهة الفكين للتصوير. ضع اللقطة في الفتحة المحددة واضغط على مقابض المتجرد معا لقطع دوامة الورقة.
ملاحظة: لتجنب قطع البدائية المستهدفة عن طريق الخطأ ، قم بمحاذاة مركز اللقطة بعناية مع الفتحة الموجودة في متجرد الأسلاك. بالنسبة للنباتات الكبيرة ، يوصى بإزالة بعض الأوراق المحيطة يدويا يدويا قبل استخدام المتجرد.
تنبيه: تحتوي أدوات تقشير الأسلاك على فكوك حادة يمكن أن تسبب جروحا أو إصابات أخرى إذا لم يتم استخدامها بعناية. - مع الاستمرار في الضغط على المقابض معا ، حرك المتجرد بعيدا عن اللقطة لتقليم الجزء العلوي من الأوراق. تأكد من أن المنطقة المستهدفة في البدائية تظل مغلفة بالأوراق المحيطة (الشكل التكميلي S1C ، D).
ملاحظة: بعد تقليم الأوراق العلوية ، يتم ترك اللقطة بطرف بريمورديوم مكشوف (P5 أو P6 أو P7 ، اعتمادا على موضع القطع وحجم فتحة متجرد الأسلاك المستخدمة ؛ الشكل 1E). سيكون هذا البدائية بمثابة مرجع لتقدير حجم ومرحلة البدائية الأصغر سنا. P6 هو المرجع الأكثر عملية للاستخدام في شتلات الذرة نظرا لنطاق حجمه ومورفولوجيته الشبيهة بالدبوس. - استخدم مسطرة لقياس الطول من قمة البدائية إلى قاعدة الساق. سجل القياس.
ملاحظة: سيتم المبالغة في تقدير طول البدائية قليلا حيث يوجد بضعة ملليمترات من الساق عند قاعدة البدائية (الشكل 2 أ).
- التقسيم اليدوي وتصوير ورقة البدائية
- تحت مجهر ستيريو عند تكبير 0.8x تقريبا ، أمسك اللقطة ثابتة على سطح أملس ، مثل لوح تقطيع الزجاج أو أي مادة أخرى مقاومة للخدش. استخدم شفرة حلاقة أو مشرط لقطع مقاطع رقيقة (0.25-0.8 مم) من اللقطة عند النقاط المرغوبة على طول البدائية.
- قم بعمل قطع أولي أعلى قليلا من المنطقة المستهدفة للتخلص من الجزء البعيد من اللقطة ، ثم احصل على أقسام رفيعة حول المنطقة المستهدفة. لعمل جروح نظيفة ، أمسك الشفرة بشكل عمودي على اللقطة وحركها للداخل بحركة سلسة ومتساوية.
ملاحظة: قد يكون هناك بضعة ملليمترات من فسحة التقسيم للبدائية الأكبر (P6 أو P7). ومع ذلك ، بالنسبة للبريمورديا الأصغر (P5 وما قبلها) ، يمكن أن يحدث 1 مم فرقا كبيرا في أخذ العينات ، حيث يتم تكديس هذه البدائية خلال المليمترات القليلة الأولى من قاعدة اللقطة (انظر الشكل 2A ، F-H). يوصى باستخدام شفرة حلاقة ذات حدين للتقسيم الأدق.
تنبيه: توخ الحذر عند استخدام شفرات الحلاقة والمشارط لتجنب أي جروح أو إصابات عرضية. - استبدل شفرة الحلاقة بانتظام ، لأن أغلفة الأوراق يمكن أن تضعف الشفرة بسهولة. قم بتغطية اللقطة بمناديل مبللة بعد كل جولة من التقسيم لمنع جفاف العينة.
- قم بتركيب قسم الورقة على شريحة زجاجية نظيفة (75 مم × 25 مم) ، واستخدم ماصة لوضع قطرة ماء (أو 50٪ جلسرين) مباشرة على القسم ، وضع غطاء (22 مم × 22 مم) في الأعلى.
- في حالة استخدام صبغة الفلورسنت ، ضع محلول الصبغة على القسم ، وضع غطاء في الأعلى ، واحتضانه حسب الحاجة.
ملاحظة: اعتمادا على نوع الصبغة ، قد تكون من الضروري اتخاذ خطوات إضافية لمعالجة أقسام الأوراق قبل المتابعة إلى الخطوة التالية. توفر هذه المراجع44،45،46،47 تفاصيل عامة واستخدامات الأصباغ الفلورية في تصوير الخلايا النباتية. تم استخدام صبغة فلورية ملطخة بالنواة ، 5-ethynyl-2′-deoxyuridine (EdU) ، بشكل فعال في المقاطع المستعرضة لأوراق الذرة البدائية48. على النقيض من ذلك ، فإن أصباغ تلطيخ غشاء البلازما ، مثل Fei Mao 4-64 (FM 4-64) ويوديد البروبيديوم ، لا تنتج نتائج مرضية (الشكل 3A-D). - ضع الشريحة على مرحلة مجهر المسح بالليزر الفلوري أو متحد البؤر واضبط التركيز والإعدادات حسب الحاجة لتصور الفلوروفور30. انظر الجدول 1 للاطلاع على إعدادات الإضاءة والحصول على الصور المستخدمة في خطوط مراسل الذرة FP المختارة.
ملاحظة: قد تنتج المقاطع السميكة مستويات عالية من التألق الذاتي في الخلفية. ضع في اعتبارك بعض استراتيجيات التصوير وإعداد العينات التي يمكن أن تساعد في التخفيف من هذه المشكلة30,49 (انظر الجدول 2).
تنبيه: توخي الحذر وارتداء النظارات الواقية عند العمل مع مصادر الضوء القوية والليزر لتقليل خطر تلف العين. - صورة من خلال قسم الورقة بأشكال تكبير وقنوات كشف مختلفة (بما في ذلك إضاءة المجال الساطع)30. استخدم تكبير 4x لتصوير المقطع العرضي بأكمله وتكبير 20x أو 40x لتصوير أنسجة معينة. تقليل وقت التعرض عند التكبير العالي للتصوير فوق الفلوري لحماية العينات من التبييض الضوئي السريع (انظر الجدول 1).
- التقط صورا للقسم بأكمله أو مناطق اهتمام محددة (ROIs). إذا لزم الأمر ، احصل على مكدس z ، وهو عبارة عن سلسلة من الصور التي تم التقاطها على أعماق بؤرية مختلفة30.
- للحصول على مكدس z ، حدد أولا المواضع العلوية والسفلية في العينة ، ثم عدد المقاطع البصرية التي يجب التقاطها بين هذه المواضع. ينتج عن عدد أقل من المقاطع البصرية حجم خطوة z أكبر ، وهو المسافة بين كل قسم. اعتمادا على خصائص FP والتكبير المستخدم ، التقط ثلاثة إلى 25 مقطعا بصريا بأحجام خطوة z بين 1 و 12 ميكرومتر لتصوير المقاطع المستعرضة لأوراق الذرة البدائية.
ملاحظة: لتسطيح مكدس z، استخدم تقنية عرض الحجمالمناسبة 50 التي يمكن تحقيقها من خلال برنامج المجهر أو باستخدام ImageJ/FIJI34 (انظر الخطوتين 4.1 و4.2). - احفظ جميع ملفات الصور والبيانات الوصفية المرتبطة بها ، إن وجدت.
الجدول 1: إعدادات الإضاءة والحصول على الصور المستخدمة في التصوير الفلوري ومتحد البؤر لمراسلي FP مختارين من الذرة. الاختصارات: FP = بروتين الفلورسنت. TRITC = رباعي ميثيل رودامين. FITC = فلوريسئين إيزوثيوسيانات ؛ WLL = ليزر الضوء الأبيض ؛ Ar-ion = ليزر أيون الأرجون ؛ HyD = كاشف هجين ؛ AU = وحدة جيدة التهوية ؛ هرتز = هرتز ، خط المسح في الثانية. الرجاء الضغط هنا لتنزيل هذا الجدول.
3. تصوير حوامل كاملة من أوراق الذرة البدائية
- تشريح البدائية وإعداد الشريحة الزجاجية بشريط نانو
- اجمع الشتلات عن طريق قطعها بعناية في الميزوكوتيل تحت سطح التربة بسكين صغير أو مقص ، كما هو موضح في الخطوات 1.1.3-1.1.4 (الشكل 1 أ).
- قم بإعداد الشريحة الزجاجية (75 مم × 25 مم) بشريط نانو شفاف على الوجهين (الشكل التكميلي S1H) قبل تشريح النبات. اقطع قطعة مستطيلة من شريط النانو وألصقها بوسط شريحة زجاجية نظيفة. لا تقم بإزالة الفيلم البلاستيكي الواقي الموجود على الجانب العلوي من الشريط.
ملاحظة: حدد حجم الشريط المراد استخدامه وفقا لحجم العينات المراد تصويرها (انظر أمثلة العينات كبيرة الحجم في الشكل التكميلي S1I وJ). يتوفر شريط النانو إما في مادة هلام الأكريليك أو البولي يوريثين. كلا النوعين فعالان للتصوير الفلوري والبؤري24. لمنع تغير لون الشريط ، قم بحمايته من التعرض الطويل للضوء عن طريق تخزينه في حاوية مظلمة. - ابدأ في تشريح اللقطة عن طريق إزالة الجزء العلوي من الأوراق القديمة باستخدام متجرد سلكي ، كما هو موضح في الخطوة 2.1.
- امسك اللقطة بواسطة mesocotyl وقم بإزالة الأوراق المحيطة بعناية باستخدام مسبار أسنان لاستخراج البدائية. للقيام بذلك ، قم بإزالة الأوراق من قاعدة اللقطة وفتحها واحدة تلو الأخرى حتى يتم الكشف عن البدائية ذات الاهتمام.
- بدلا من ذلك ، استخدم متجرد الأسلاك لاستخراج ورقة البدائية مباشرة عن طريق إجراء قطع في المنطقة القاعدية من اللقطة (الشكل التكميلي S1E-G).
ملاحظة: تنطوي هذه الطريقة على مخاطر أكبر لالتقاط البدائية.
- تركيب وتصوير البدائية.
- قم بإزالة الفيلم الواقي من الشريط. ضع البدائية المكشوفة على الشريط. استخدم شفرة حلاقة لقطع البدائية في القاعدة ، والتخلص من الجذع القاعدي و hypocotyl (الشكل 1F).
- قم بفك البدائية باستخدام مسبار أسنان بإبرة منحنية (قطر الطرف 0.25-0.6 مم). استخدم مسبارا دقيقا (قطر الطرف 0.15-0.2 مم) للبدائية الأصغر من 3 مم (P3 إلى P4 المبكر).
- ضع طرف الإبرة بالتوازي مع السطح وافتح الهامش القاعدي الخارجي ، واضغط عليه برفق على الشريط.
- لفك أكثر سلاسة ، قم بتليين السطح الداخلي (adaxial) للورقة عن طريق غمس طرف الإبرة في 100٪ جلسرين.
- مع التصاق الهامش الخارجي بالشريط ، قم بمحاذاة طرف الإبرة بالتوازي مع المحور الطويل للورقة. حرك الإبرة برفق جانبا لفك الورقة وتسطيحها على الشريط (الشكل 1F).
ملاحظة: يعد كسر الورقة أو إتلافها أثناء اللف أمرا شائعا ، خاصة في البريمورديا الأكبر حجما ذات الضلع الأوسط القوي (انظر الشكل 3E-G). يمكن أن يؤدي فتح الورقة بقوة إلى كدمات سطحها وإنتاج قطع أثرية أثناء التصوير (الشكل 3H). انظر الجدول 2 والمناقشة للاطلاع على الحلول الموصى بها لهذه المشاكل. - ضع قطرة ماء على البدائية غير المتداولة. ضع على الفور غطاء فوق قطرة الماء والبدائية. اضغط برفق على حواف غطاء الغطاء حتى تلتصق بالشريط.
ملاحظة: يوصى باستخدام أغطية مستطيلة كبيرة (50 مم × 24 مم أو 60 مم × 24 مم) مع مساحات إضافية للالتصاق بالشريط. قلل من تكوين فقاعات الهواء ، والتي يمكن أن تشوه الورقة وتسبب انكسارا غير متساو للضوء (انظر الشكل 3I-K). تأكد من أن غطاء الغطاء يلتصق تماما بالشريط ، حيث قد تتدحرج هوامش البدائية المركبة مرة أخرى تحت غطاء ملتصق بشكل فضفاض (انظر الشكل 3L). - ضع الشريحة على مرحلة التألق (انظر الشكل التكميلي S1K) أو مجهر المسح بالليزر متحد البؤر واضبط التركيز والإعدادات حسب الحاجة لتصور الفلوروفور30. انظر الجدول 1 للاطلاع على إعدادات الإضاءة والحصول على الصور المستخدمة لخطوط مراسل الذرة FP المختارة.
- تخيل الورقة بأكملها أو عائد استثمار محدد من خلال قنوات الكشف المختلفة30. لتصوير الورقة بأكملها ، إما أن تلتقط صور الفسيفساء للورقة يدويا أو استخدم عملية التجانب للمجهر عند التكبير المنخفض (4x أو 10x).
ملاحظة: حتى عند أدنى تكبير ، قد تكون بريمورديا أوراق الذرة كبيرة جدا بحيث لا يمكن تصويرها باستخدام مجهر المسح بالليزر أو مجهر المسح بالليزر متحد البؤر ، مما يؤدي إلى أوقات تصوير أطول قد تؤدي إلى التبييض الضوئي (انظر الشكل 3M) أو زيادة التألق الذاتي في الخلفية. لذلك ، يوصى باستخدام مجهر ستيريو مضان لتصوير عينات الأوراق الكاملة التي يزيد حجمها عن 5 مم. - بالنسبة للتصوير متحد البؤر ، سيكون من الضروري الحصول على مداخن z التي تشمل سمك الورقة (انظر الخطوة 2.2.9).
- احفظ جميع ملفات الصور والبيانات الوصفية المرتبطة بها ، إن وجدت.
الجدول 2: استكشاف المشكلات الشائعة وإصلاحها في تصوير المقاطع المستعرضة والجبال الكاملة لأوراق الذرة البدائية. الرجاء الضغط هنا لتنزيل هذا الجدول.

الشكل 3: مقطع عرضي دون المستوى الأمثل وتحضير كامل التركيب لأوراق الذرة البدائية. (A-D) صور تمثيلية متحدة البؤر لمقاطع عرضية من ورقة البدائية مع علامة غشاء البلازما ، PIP2-1-CFP (A) ، وصبغة فلورية ملزمة بغشاء البلازما ، FM 4-64 (B) ، مع صور المجال الساطع المقابلة (C ، D). عند مقارنته ب PIP2-1-CFP ، يعرض FM 4-64 تصورا دون المستوى الأمثل لمخططات الخلايا. (E-M) صور مضان تمثيلية لحوامل كاملة من الأوراق البدائية تظهر وجود تمزق (E-G) ، وأسطح كدمات † (H) ، وفقاعات هواء * (I-K) ، وهوامش دحرجة (L) ، ومناطق مبيضة ضوئيا ‡ (M). تعبر الورقة الأولية عن DII-Venus (E-G) و GAR2-YFP (H-J) و mDII-Venus (K) و PIN1a-YFP (L) و DR5-RFP (M). شريط المقياس = 200 ميكرومتر (A-D) ؛ 500 ميكرومتر (E-M). تم تعديل الشكل 3A واستنساخه بإذن من Robil و McSteen24 ، في حين أن الشكل 5B-M عبارة عن بيانات غير منشورة من المؤلفين. الاختصارات: P = بلاستوكرون. YFP = بروتين الفلورسنت الأصفر ؛ RFP = بروتين الفلورسنت الأحمر ؛ CFP = بروتين الفلورسنت السماوي ؛ PIP2-1-CFP =pZm PIP2-1::ZmPIP2-1:CFP ؛ DII-فينوس = pZmUbi:DII:YFP-NLS; GAR2-YFP = p Zm GAR2::ZmGAR2:YFP; mDII-فينوس = pzmUbi:mDII:YFP-NLS; PIN1a-YFP = p Zm PIN1a::ZmPIN1a:YFP; DR5-RFP = DR5rev::mRFPer; BF = حقل مشرق. يرجى النقر هنا لعرض نسخة أكبر من هذا الرقم.
{kind=link}
4. معالجة الصور باستخدام ImageJ / FIJI
- تسطيح مكدسات z باستخدام الإسقاط الأقصى للكثافة (MIP)
- قم بتشغيل ImageJ/FIJI وافتح كومة الصور، أو أنشئ مكدسا جديدا من عدة صور منفصلة عن طريق تحديد صورة | مداخن | الصور المراد تكديسها من القائمة.
- حدد مكدس الصور. من القائمة، حدد صورة | مداخن | Z-المشروع.... في شاشة Z-Project ، اختر أقصى كثافة من القائمة المنسدلة نوع الإسقاط . انقر فوق موافق لإنشاء أقصى كثافة z-projection.
ملاحظة: الصورة المسطحة هي إسقاط 2D من وحدات البكسل مع أعلى كثافة عبر مكدسات z50. هذه الطريقة مناسبة لتصور الفلوروفور ولكن ليس السمات التشريحية في صور المجال الساطع للمقاطع المستعرضة السميكة وحوامل الأوراق الكاملة. - احفظ الصورة باسم TIFF (تنسيق ملف الصورة الموسوم) عن طريق تحديد ملف | حفظ باسم | المشاجرة. أدخل اسما للملف وانقر فوق حفظ.
ملاحظة: يوصى باستخدام TIFF لتخزين الصور الفلورية ومتحدة البؤر ، حيث يحتفظ بالجودة والتفاصيل عند حفظ الصورة.
- تسطيح مكدسات z باستخدام المكون الإضافي لعمق المجال الممتد (EDF)51
- قم بتثبيت المكون الإضافي EDF52. قم بتنزيل ملف Extended_Depth_Field.jar (الإصدار 17.05.2021) من http://bigwww.epfl.ch/demo/edf/ وضعه في مجلد "المكونات الإضافية" في ImageJ / FIJI.
- قم بتشغيل ImageJ/FIJI وافتح كومة الصور، أو أنشئ مكدسا جديدا من عدة صور منفصلة عن طريق تحديد صورة | مداخن | الصور المراد تكديسها من القائمة.
- حدد مكدس الصور. من القائمة، حدد المكونات الإضافية | عمق المجال الممتد | EDF الوضع السهل. في مربع الحوار عمق المجال الممتد ، استخدم أشرطة التمرير الموجودة ضمن السرعة/ الجودة لضبط الجودة المطلوبة وسرعة المعالجة، وسجل خريطة الارتفاع لتعيين مستوى النعومة للتضاريس. انقر فوق تشغيل لإنشاء الصورة المعاد بناؤها.
ملاحظة: يجمع EDF المكدس في صورة 2D حادة ومركبة من خلال عرض أفضل تركيزبؤري 51. هذه التقنية مفيدة للتغلب على عمق التركيز المحدود وهي مناسبة لإعادة بناء صور المجال الساطع للمقاطع المستعرضة السميكة أو حوامل الأوراق الكاملة. توخ الحذر عند تطبيق هذا المرشح لتصور الفلوروفور لأنه ، على عكس MIP ، يقوم EDF بضبط تباين الصورة ، مما يؤثر على كثافة البكسل. - احفظ الصورة كملف TIFF عن طريق تحديد ملف | حفظ باسم | المشاجرة. أدخل اسما للملف وانقر فوق حفظ.
- تجميع الصور معا باستخدام المكون الإضافي Grid / Collection Stitch53
- احفظ مجموعة الصور أو مكدسات الصور ليتم تجميعها معا في دليل واحد.
- قم بتشغيل ImageJ / FIJI وحدد المكونات الإضافية | خياطة | خياطة الشبكة / المجموعة من القائمة. في مربع الحوار ربط الشبكة/المجموعة ، اختر موضع غير معروف من القائمة المنسدلة النوع، ثم كل الملفات في الدليل من الترتيب. انقر فوق موافق.
- في مربع الحوار خياطة الشبكة: موضع غير معروف، كل الملفات في الدليل ، حدد موقع الصور عن طريق كتابة مسار الدليل أو لصقه في حقل نص الدليل أو بالنقر فوق استعراض ... لتحديد موقع الدليل. انقر فوق "موافق " لبدء عملية الخياطة.
ملاحظة: يقوم خيار الموضع غير المعروف في المكون الإضافي Grid/Collection Stitch بإعادة بناء صورة مركبة من مجموعة من الصور المتجاورة عن طريق تحليل مناطقها المتداخلة. لذلك ، يكون هذا الخيار مفيدا لخياطة فسيفساء صور المقاطع المستعرضة أو حوامل الأوراق الكاملة التي لم يتم الحصول عليها باستخدام عملية مسح البلاط. - عند الانتهاء من الخياطة ، ستظهر نافذة جديدة مع الصورة المخيطة. احفظ الصورة كملف TIFF عن طريق تحديد ملف | حفظ باسم | المشاجرة. أدخل اسما للملف وانقر فوق حفظ.
ملاحظة: يمكن أيضا إجراء خياطة مونتاج كبير للصور 2D و 3D باستخدام أوامر إعادة بناء الصور في برنامج المجهر أو من خلال خيارات استيراد التنسيقات الحيوية في ImageJ / FIJI.
- دمج قنوات متعددة باستخدام أمر دمج القنوات
- قم بتشغيل ImageJ / FIJI وافتح الصور أو مكدسات الصور المراد دمجها.
ملاحظة: يمكن تسوية مكدسات الصور أولا باستخدام الخطوتين 4.1 أو 4.2. يوصى باستخدام MIP بشكل عام لتسطيح قنوات الفلوروفور ، في حين أن EDF مناسب بشكل أفضل لتسطيح قنوات المجال الساطع أو القنوات الأخرى ذات عمق التركيز المتفاوت. - اضبط سطوع الصور وتباينها باستخدام Image | ضبط | السطوع / التباين. استخدم أشرطة التمرير أو حقول الإدخال لضبط المعلمات، ثم انقر على تطبيق. بدلا من ذلك ، استخدم طريقة تمدد الرسم البياني الخطي عن طريق تحديد عملية | تعزيز التباين.... قم بتعيين النسبة المئوية للبيكسلات المشبعة، حدد تطبيع، ثم انقر على موافق.
ملاحظة: عند تحديد كثافة إشارة الفلوروفور أو مقارنتها بين الصور ، من الأفضل عموما عدم ضبط السطوع أو التباين لأن هذا يمكن أن يغير كثافة البكسل النسبية في الصورة ، مما قد يؤثر على القياسات والمقارنات. - للتمييز بين القنوات، قم بتطبيق الألوان الزائفة على صور معينة باستخدام جدول بحث (LUT). للقيام بذلك ، حدد الصورة ، ثم من قائمة ImageJ / FIJI ، حدد صورة ، وانقر فوق جداول البحث ، واختر جدول البحث المناسب من القائمة المنسدلة.
- من القائمة، حدد صورة | اللون | دمج القنوات. لكل قناة (C1 ، C2 ، C3 ...) ، استخدم القائمة المنسدلة لتحديد الصورة المراد تعيينها لتلك القناة. انقر فوق موافق لدمج القنوات.
- احفظ الصورة الجديدة بتنسيق TIFF عن طريق تحديد ملف | حفظ باسم | المشاجرة. أدخل اسما للملف وانقر فوق حفظ.
- قم بتشغيل ImageJ / FIJI وافتح الصور أو مكدسات الصور المراد دمجها.
النتائج
تحليل المقطع المستعرض لأوراق الذرة البدائية
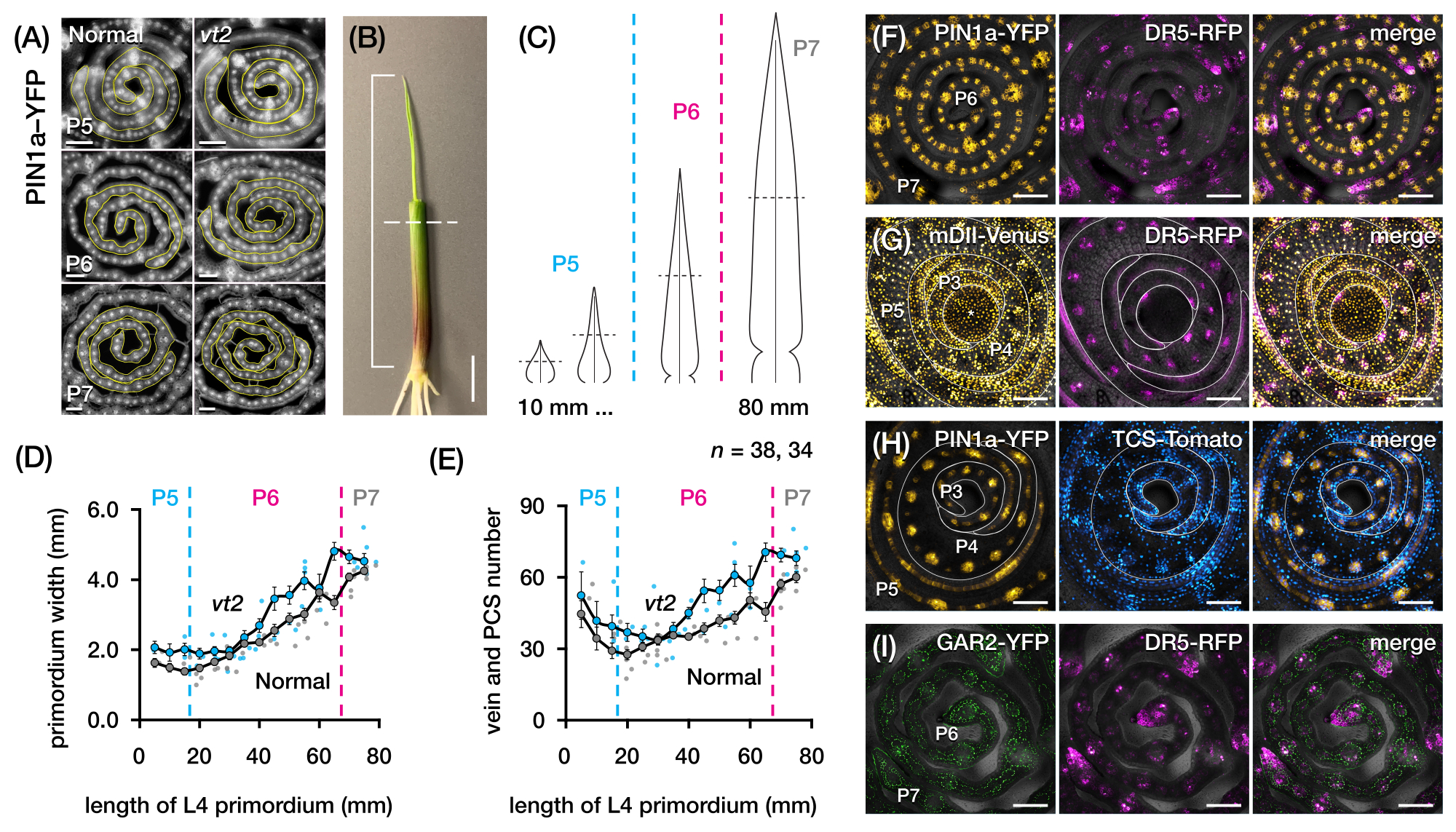
استخدمنا قسم البروتوكول 2 لتحديد عدد الوريد وتوصيف أنماط الاستجابة الهرمونية في المقاطع المستعرضة لأوراق الذرة الأولية مع FPs (الشكل 4) 24. لتقييم دور هرمون الأوكسين النباتي في نمو الأوراق وتكوين الوريد، حددنا عدد الأوردة في الورقة البدائية لطافرة الذرة التي تعاني من نقص الأوكسين، والتي تتلاشى شرابة254. في البلاستوكرونات المبكرة ، تظهر الأوردة النامية خلوية مميزة في طبقة الخلايا المتوسطة لأوراق الذرة البدائية21,22. ومع ذلك ، فإن تحديد الأوردة وعدها باستخدام التقنيات النسيجية التقليدية22 يمكن أن يكون كثيف العمالة ويستغرق وقتا طويلا. وهكذا، لتحديد كمية الأوردة، استخدمنا علامة بروتين تدفق أوكسين الذرة p Zm PIN1a::ZmPIN1a:YFP41 (PIN1a-YFP فيما بعد)، والتي تشير إلى الأوردة النامية والخيوط الشفافة (PCSs; الشكل 4 أ). باستخدام قسم البروتوكول 2 ، تمكنا من توحيد أخذ عينات المقطع المستعرض عن طريق قياس البدائية قبل التقسيم (الشكل 4B ، C). اكتشفنا اتجاها يحتوي فيه vt2 على بدائية أوسع وأوردة أكثر من المعتاد (الشكل 4D ، E) 24 ، وهو ما يتوافق مع البيانات من الأوراق الموسعة بالكامل55 ، مما يشير إلى أن العيب في vt2 بدأ في وقت مبكر من تطور الأوراق. باستخدام قسم البروتوكول 2 ، تمكنا أيضا من فحص أنماط التعبير عن مراسلي FP للاستجابة الهرمونية بشكل منهجي في ورقة البدائية (انظر أمثلة الصور في الشكل 4F-I). من خلال مخطط موحد لأخذ عينات المقطع المستعرض ، قمنا بتعيين توزيع استجابات الأوكسين والسيتوكينين (CK) وحمض الجبريليك (GA) عبر البلاستوكرونات والمناطق المختلفة من الأوراق البدائية ، واكتشفنا أنماط استجابة جديدة نفترض أن لها آثارا على نمو الأوراق وتكوين الوريد24. لذلك ، توضح هذه النتائج التمثيلية فائدة قسم البروتوكول 2 لتحليل المقطع العرضي لأوراق الذرة البدائية.

الشكل 4: النتائج التمثيلية لتحليل المقطع العرضي لأوراق الذرة البدائية. (A-E) القياس الكمي لعدد الوريد وعرض البدائيات في ورقة البدائية للشرابة العادية والمتلاشية2 باستخدام علامة بروتين تدفق الأوكسين PIN1a-YFP. (أ) صور مضان تمثيلية للمقاطع المستعرضة من P5 إلى P7 التي تعبر عن PIN1a-YFP في الأوردة النامية والخيوط المفصلية. تم تصوير المقاطع المستعرضة باستخدام مجهر التألق باستخدام مرشح FITC (إثارة 495-519 نانومتر). تم تحديد عدد الأوردة وعرض البدائية باستخدام أدوات الخط متعدد النقاط والخط الحر في FIJI / ImageJ ، على التوالي. (ب) شتلة ذرة مع إزالة زهور الورقة العلوية بمتجرد سلكي لكشف قمة الورقة الرابعة (L4). يمتد القوس على طول البدائية المتوقع ، مع الخط المتقطع الذي يشير إلى منتصف الطول. (ج) رسم تخطيطي لأشكال البدائية المختلفة من P5 إلى P7 ، يوضح كيف يمكن أن يختلف عدد الوريد وعرض البدائية عند الطول المتوسط (الخط الأفقي المتقطع) اعتمادا على مرحلة تطور البدائية. (د، ه) قياس عرض البدائية (D) ورقم الوريد (E) في قسم منتصف الطول من L4 العادي و vt2. يمثل خط الاتجاه متوسطات التدحرج 10 مم للقياسات ± الخطأ القياسي للمتوسط (SEM). (واو-ط) صور تمثيلية متحدة البؤر لمقاطع عرضية من أوراق البدائية التي تعبر عن مجموعات من PIN1a-YFP ، مراسل استجابة الأوكسين ، DR5-RFP ، مراسل استجابة السيتوكينين ، TCS-Tomato ، علامة استجابة حمض الجبريليك ، GAR2-YFP ، و mDII-Venus ، نسخة متحولة من مراسل إدخال إشارات الأوكسين DII-Venus. يتم تثبيت قنوات FP على قناة المجال الساطع في كل صورة. شريط المقياس = 200 ميكرومتر (أ) ؛ 10 مم (ب) ؛ 100 ميكرومتر (F-I). تم تعديل هذا الرقم واستنساخه بإذن من Robil و McSteen24. الاختصارات: DAG = أيام بعد الإنبات ؛ P = بلاستوكرون ؛ vt2 = شرابة التلاشي 2 ؛ PCS = حبلا بروكاميال ؛ YFP = بروتين الفلورسنت الأصفر ؛ FITC = فلوريسئين إيزوثيوسيانات ؛ RFP = بروتين الفلورسنت الأحمر ؛ PIN1a-YFP = p Zm PIN1a::ZmPIN1a:YFP; DR5-RFP = DR5rev::mRFPer; TCS-الطماطم = TCSv2::NLS-tdالطماطم; GAR2-YFP = p Zm GAR2::ZmGAR2:YFP; mDII-فينوس = pzmUbi:mDII:YFP-NLS. يرجى النقر هنا لعرض نسخة أكبر من هذا الرقم.
{kind=link}
تحليل كامل لأوراق الذرة البدائية
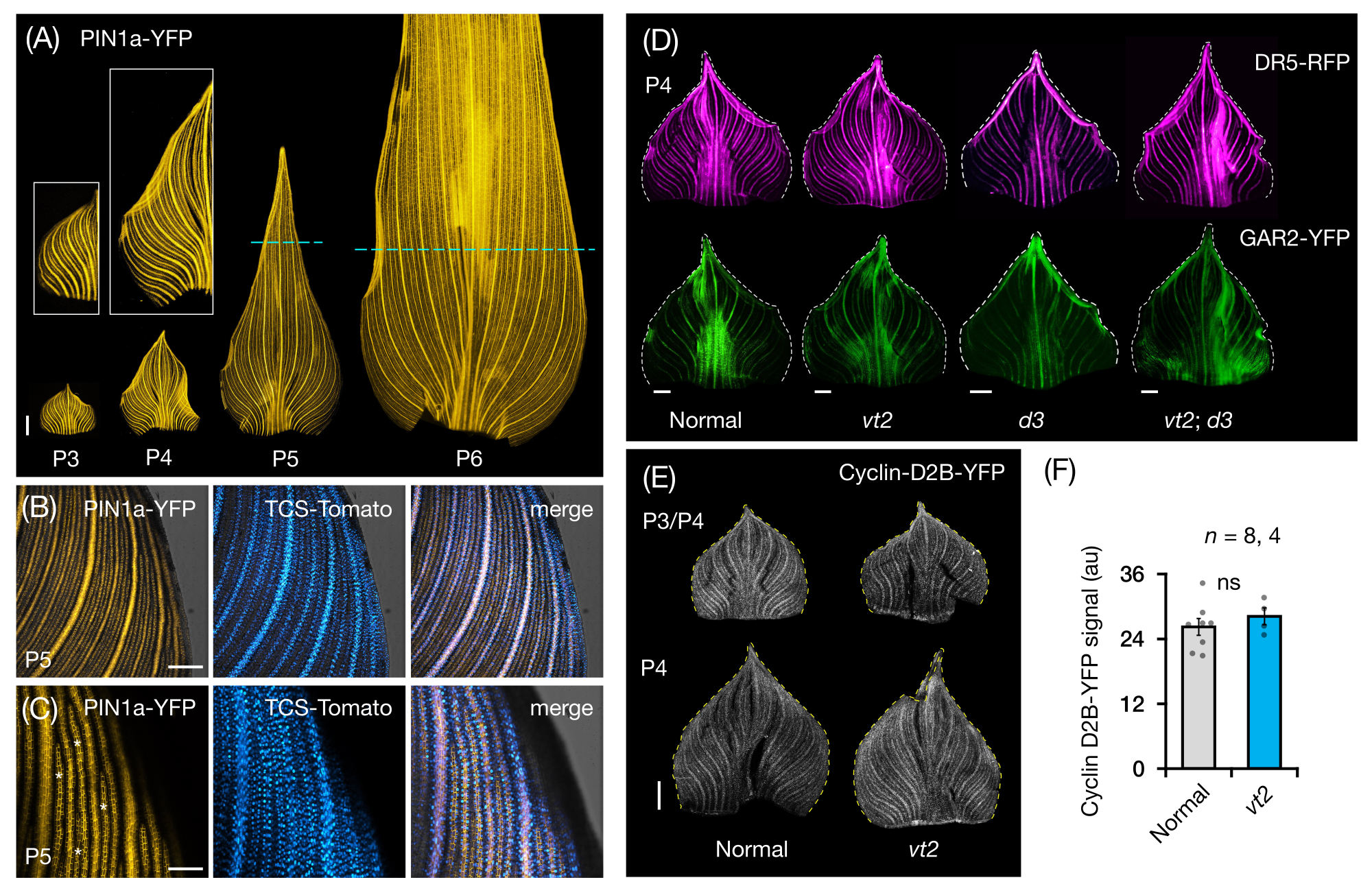
اتبعنا قسم البروتوكول 3 لتصور وتحليل تعبير FP في حوامل الأوراق الكاملة لأوراق الذرة البدائية (الشكل 5). من خلال تصور أنماط الوريد باستخدام PIN1a-YFP ، وجدنا أن تكوين الأوردة يحدث في البدائية بأكملها خلال البلاستوكرونات المبكرة ، لكن هذه العملية تصبح مقيدة في المناطق القريبة لاحقا في التطور (الشكل 5 أ)24. استكمالا لتحليلات المقطع العرضي ، كشفت تحليلات جبل الورقة الكاملة عن أنماط خاصة بالأنسجة والمرحلة لاستجابة الهرمونات أثناء تكوين الوريد24. أحد الأمثلة على ذلك هو نمط التعبير لمراسل استجابة CK TCSv2::NLS-tdTomato37 (TCS-Tomato) ، بالنسبة إلى تعبير PIN1a-YFP (الشكل 5B)24. باتباع قسم البروتوكول 3 ، تمكنا من إجراء تحليلات نوعية وكمية لتعبير FP في ورقة البدائية (الشكل 5D-F ؛ بيانات غير منشورة). قمنا بفحص أنماط التعبير لمراسل استجابة الأوكسين ، DR5rev :: mRFPer41 (DR5-RFP) ، وعلامة استجابة GA ، p Zm GAR2::ZmGAR2: YFP39 (GAR2-YFP) ، في حوامل كاملة الأوراق من طفرات مفردة ومزدوجة من vt2 والنبات القزم 3 (d3) ، طافر ناقصGA 56 (الشكل 5D). قارنا أيضا المستويات النسبية لتكاثر الخلايا بين البدائية العادية وأوراق vt2 باستخدام p Zm Cyclin-D2B::ZmCyclin-D2B:YFP42 (Cyclin-D2B-YFP) ، وهي علامة على انتقال G1 / S في دورة الخلية 57 (الشكل 5E ، F). في حين لم يكن هناك فرق كبير بين الطبيعي و vt2 ، كان تعبير Cyclin-D2B-YFP متسقا مع ملف تكاثر الخلايا المعروف للبلاستوكروناتالمبكرة 31. نستنتج أن قسم البروتوكول 3 هو طريقة فعالة لتحليل حوامل كاملة من أوراق الذرة البدائية ، والتي يصعب تصويرها بسبب مورفولوجيتها الملفوفة.

الشكل 5: النتائج التمثيلية للتحليل الكامل لأوراق الذرة البدائية. (أ) صور مضان تمثيلية للأوراق البدائية ل 7 شتلة ذرة من DAG تظهر الأوردة النامية والخيوط البروكمبيالية ، كما هو موضح بعلامة بروتين تدفق الأوكسين PIN1a-YFP. تم استئصال P3-P6 primordia من القاعدة ، وفكها ، وتسويتها مع الجانب المحوري لأعلى. تظهر الأجزاء الداخلية لقطات مقربة ل P3 و P4. في P5 و P6 ، تحدد الخطوط المتقطعة النهاية البعيدة للمناطق التكاثرية ، حيث لا تزال غالبية الخيوط الاستباقية تتطور وتمتد. (ب، ج) صور تمثيلية متحدة البؤر تظهر تعبير PIN1a-YFP ومراسل استجابة السيتوكينين ، TCS-Tomato ، في المنطقة الهامشية القريبة من بدائية P5. (د) صور مضان تمثيلية ل P4 primordia تظهر تعبير مراسل استجابة الأوكسين ، DR5-RFP ، وعلامة استجابة حمض الجبريليك ، GAR2-YFP في الطفرات العادية والمفردة والمزدوجة للشرابة المتلاشية 2 والنبات القزم 3. (ه) صور تمثيلية متحدة البؤر ل P3 و / أو P4 تظهر تعبير Cyclin-D2B-YFP ، مراسل انتقال G1 / S في دورة الخلية. (F) الكميات النسبية لتكاثر الخلايا في ورقة P3 / P4 الأولية العادية و vt2 ، محددة كميا عن طريق قياس الكثافة المتكاملة لإشارة Cyclin-D2B-YFP فوق منطقة البدائية باستخدام ImageJ / FIJI. تمثل الأشرطة متوسط القياسات ± الخطأ المعياري للمتوسط. شريط المقياس = 500 ميكرومتر (A ، D ، E) ؛ 200 ميكرومتر (ب) ؛ 100 ميكرومتر (C). تم تعديل الشكل 4A-C واستنساخه بإذن من Robil و McSteen24 ، في حين أن الشكل 4D-F هو بيانات غير منشورة من المؤلفين. الاختصارات: DAG = أيام بعد الإنبات ؛ P = بلاستوكرون ؛ vt2 = شرابة التلاشي 2 ؛ d3 = نبات قزم 3 ؛ YFP = بروتين الفلورسنت الأصفر ؛ RFP = بروتين الفلورسنت الأحمر ؛ PIN1a-YFP = p Zm PIN1a::ZmPIN1a:YFP; TCS-الطماطم = TCSv2::NLS-tdالطماطم; DR5-RFP = DR5rev::mRFPer; GAR2-YFP = p Zm GAR2::ZmGAR2:YFP; سيكلين-D2B-YFP = p Zm سيكلين-D2B::Zmسيكلين-D2B:YFP; ns = لا يوجد فرق كبير ؛ au = وحدة تعسفية. يرجى النقر هنا لعرض نسخة أكبر من هذا الرقم.
{kind=link}
الملف التكميلي 1: أمثلة على مراحل تطور الأوراق في شتلات الذرة. الرجاء الضغط هنا لتنزيل هذا الملف.
الشكل التكميلي S1: العينات النباتية والمواد المستخدمة في البروتوكولات. (أ، ب) شتلة الذرة 7-8 DAG مع إزالة دوامة الورقة العلوية باستخدام متجرد سلكي. (ب) يظهر الجزء الداخلي لقطة مقربة للساق باستخدام بدائية P6 المكشوفة. (ج-ز) براعم شتلات الذرة مع إزالة الزهرات العلوية لتحليل المقطع العرضي (C ، D) وإزالة الأوراق المحيطة بالكامل لتحليل التركيب الكامل (E-G). (H) لفافة من هلام البولي يوريثين شريط نانوي شفاف على الوجهين. (ط، ي) P6 ورقة primordium (I) والمنطقة القريبة من ورقة P8 (J) غير ملفوفة ومثبتة على شرائح زجاجية بشريط نانو. (ك) شريحة زجاجية ذات ورقة بريمورديوم غير ملفوفة مثبتة على مرحلة مجهر فوق التألق. اختصار: DAG = أيام بعد الإنبات. الرجاء الضغط هنا لتنزيل هذا الملف.
Discussion
نقدم طريقتين لإعداد بريمورديا أوراق الذرة للتصوير الخلوي. تسمح الطريقة الأولى (قسم البروتوكول 2) بقياس البدائية لتحليل المقطع العرضي ، بينما تتيح الطريقة الثانية (قسم البروتوكول 3) فتح وتسطيح البدائية لتحليل التركيب الكامل. تسهل هذه الطرق التصوير الخلوي ل FPs في أوراق الذرة الأولية24 (كما هو موضح في الشكل 4 والشكل 5) وتوفر حلولا بسيطة لتحديات تصوير أوراق الذرة النامية. يقلل قسم البروتوكول 2 من وقت التشريح ويحسن دقة أخذ العينات عن طريق قياس البدائية قبل التقسيم بدلا من الاعتماد فقط على معلمات التدريج 9,16. مع شريط نانو متاح تجاريا ، يحل قسم البروتوكول 3 المشكلة طويلة الأمد المتمثلة في تصوير البدائية ذات الأوراق الكاملة في الذرة. يحسن هذا البروتوكول الطريقة السابقة ، التي تستخدم أنابيب غسيل الكلى 31 ، وهو بديل أرخص بكثير للتصوير المقطعي المحوسب والتصوير بالرنين المغناطيسي11،32،33. ومع ذلك ، عندما يتعلق الأمر بتصور السمات التشريحية للأوراق وتحقيق أفضل النتائج ، فإن كلا البروتوكولين لهما بعض القيود ، الموضحة في الجدول 2 وتتم مناقشتها بمزيد من التفصيل أدناه.
في قسم البروتوكول 2 ، واجهنا صعوبة في تصور الخطوط العريضة للخلايا في أقسام عرضية سميكة من البدائية للأوراق ، ولم يقدم التلطيخ المضاد بأصباغ الفلورسنت المرتبطة بجدار الخلية أو غشاء البلازما نتائج مرضية. على سبيل المثال ، أنتجت FM 4-64 نتائج دون المستوى الأمثل مقارنة بعلامة FP لغشاء البلازما ، p Zm PIP2-1::ZmPIP2-1:CFP39 (PIP2-1-CFP; الشكل 3 أ - د). للتغلب على هذا القيد ، نوصي باستخدام اهتزاز لإنتاج أقسام أنسجة أرق (~ 0.1 مم)58 والتي ستسمح بتصوير مجال مشرق حيوي لمخططات الخلايا أو تحسين بروتوكول التلوينالمضاد 47,59.
في قسم البروتوكول 3 ، يتمثل القيد الرئيسي في صعوبة تركيب الورقة دون تمزق أو تلف أو فقاعات هواء ، كما هو مفصل في خطوات البروتوكول 3.2.5-3.2.6 (الشكل 3E-K). نظرا لأن ورقة الذرة متناظرة ثنائيا ، فقد يكون حامل نصف ورقة بدلا من حامل ورقة كاملة كافيا للتصور9. للقيام بذلك ، يمكن قطع البدائية بشفرة حلاقة على طول المحور الطولي بعد فكها حتى الوسط ، مما يسمح بتركيب نصف الورقة فقط. هناك قيد آخر لقسم البروتوكول 3 وهو أن سمك الورقة يمكن أن يحد من الدقة البصرية لإشارة الفلوروفور أثناء التصوير العميق. لمعالجة هذه المشكلة ، من الممكن استخدام تقنية إزالة الأنسجة60. ومع ذلك ، وجدنا أن ClearSee61 ، وهو كاشف إزالة شائع الاستخدام لتصوير الأنسجة النباتية ، غير متوافق مع البروتوكول لأنه يتسبب في انفصال العينة والغطاء عن الشريط النانوي. يمكن أن يكون الحل المحتمل لهذه المشكلة هو تطبيق غشاء شبه منفذ31 على عينة الورقة ، مما يسمح بمعالجتها بمحلول المقاصة أثناء تثبيتها في مكانها بواسطة الشريط النانوي. يمكن أيضا استخدام مثل هذه الطريقة التي تسمح بتطبيق المحاليل السائلة على الورقة غير الملفوفة في تقنيات التهجين الكامل للحمض النووي الريبي في الموقع وتقنيات التوطين المناعي ، والتي تم تحسينها سابقا لتطوير نورات الذرة ولكن ليس للأوراق الكاملةالبدائية 62,63.
وصفنا بروتوكولات الذرة ، التي تحتوي على أوراق بريمورديا كبيرة حتى في مرحلة الشتلات. قد تتطلب أنواع الحشائش الأخرى ذات الأوراق الأولية الأصغر بكثير ، مثل الأرز والشعير والقمح و Setaria و Brachypodium16،23،64،65،66 ، استخدام أدوات دقيقة إضافية لتطبيق هذه البروتوكولات بشكل فعال. علاوة على ذلك ، لم تكن هذه البروتوكولات مخصصة لتصوير الخلايا الحية ، والتي تلتقط العمليات الديناميكية في الوقت الفعلي لتكوين الأنسجة والاستجابات الخلوية. ومع ذلك ، مع استمرار تقدم مجسات الفلورسنت وتقنيات التصوير وقدرات الحوسبة في تصوير الخلايا الحية للنباتات67 ، يمكن أن تبني الأبحاث المستقبلية على هذه البروتوكولات لتطوير استراتيجيات تصوير الخلايا الحية المصممة خصيصا للميزات الفريدة لأوراق العشب البدائية.
Disclosures
ليس لدى المؤلفين أي تضارب في المصالح للكشف عنه.
Acknowledgements
يود المؤلفون أن يشكروا التعاون في علم الوراثة الذرة ، ومشروع جينوم خلايا الذرة ، وديف جاكسون (مختبر كولد سبرينج هاربور ، نيويورك) ، وآن دبليو سيلفستر (المختبر البيولوجي البحري ، جامعة شيكاغو ، إلينوي) ، وأندريا جالافوتي (جامعة روتجرز ، نيوجيرسي) ، وكارولين جي راسموسن (جامعة كاليفورنيا ، ريفرسايد) لتوفير المخزونات الطافرة والمحورة وراثيا ، وكذلك روبرت ف. بيكر وألكسندر جوركيفيتش من مركز المجهر الضوئي المتقدم في جامعة ميسوري كولومبيا لمساعدتهم في الفحص المجهري متحد البؤر. تم دعم JMR من قبل زمالة J. William Fulbright ، وصندوق Diane P. و Robert E. Sharp ، وبرنامج أبحاث الجينوم النباتي التابع لمؤسسة العلوم الوطنية (IOS-1546873) إلى PM. يتم دعم CDTC و CMRV و EDCDP و RJRR من قبل برنامج المنح الدراسية في كلية أتينيو. يتم دعم CDTC و EDCDP و RJRR من خلال منحة DOST-SEI S &T الجامعية. يتم دعم DODL من قبل منحة الأب توماس شتاينبوغلر SJ الأكاديمية. يتم دعم RJRR من قبل Aiducation International - Pathways to Higher Education Scholarship. تم دعم هذا العمل من قبل كلية العلوم والهندسة ومكتبة ريزال ، جامعة أتينيو دي مانيلا.
Materials
| Name | Company | Catalog Number | Comments |
| Acrylic Gel Clear Double Sided Nano Tape 16.5 ft x 1.2 in, 2 mm thick | EZlifego Store (Amazon) | B07YB1ZXG6 | 1 roll |
| Bellucci Pick Curved micro probe 16.8 cm, 6.6 in | Bausch & Lomb | N1692 9 | 1 pc |
| Clayman guide microprobe Sinskey hook angled shaft, 11.6 cm, 4.6 in | Storz Opthalmic Instruments | E0542 | 1 pc |
| Dental Probe, Bent Needle, 14 cm (5.5 in) | Ted Pella | 13553 | 1 pc |
| DOWELL 10-22 AWG Wire Stripper | Dowell Store (Amazon) | 10-22 AWG | 1 pc |
| Feather Double Edge Carbon Steel Blades | Ted Pella | 121-9 | pkg/10; for fine sectioning |
| Frosted End Glass Microscope Slides, 75 mm x 25 mm x 1-1.2 mm | Ted Pella | 260442 | pkg/144 |
| GEM Single Edge, Stainless Steel Uncoated Blades | Ted Pella | 121-1 | box/200; for general cutting/sectioning |
| Glycerol | Thermo Scientific | PI17904 | 1 liter |
| ImageJ/FIJI with EDF plugin (version 17.05.2021) and Grid/Collection Stitching plugin | National Institutes of Health (NIH) USA | version 2.9.0/1.54s | The EDF plugin was developed by Alex Prudencio, Jesse Berent, and Daniel Sage for the Biomedical Imaging Group, École Polytechnique Fédérale de Lausanne (EPFL; http://bigwww.epfl.ch/demo/edf/). The grid/collection stitching software was developed by Stephan Preibisch for the Max Planck Institute of Molecular Cell Biology and Genetics (MPI-CBG). |
| Kimwipes Ex-L Small 111.76 mm x 213.36 mm | Kimtech Science | 34155 | box/280 ply |
| Micro Cover Glasses, 22 mm x 22 mm x 0.13 - 0.16 mm thick | Ted Pella | 260140 | 1 ounce |
| PU Gel Clear Double Sided Nano Tape 29.5 ft x 1.18 in, 1 mm thick | Yecaye Store (Amazon) | L354 W1.18 | 2 rolls |
| Superslip Cover Glasses, 24 mm x 50 mm x 0.13 - 0.16 mm thick | Ted Pella | 260166 | 1 ounce |
| Superslip Cover Glasses, 24 mm x 60 mm x 0.13 - 0.16 mm thick | Ted Pella | 260168 | 1 ounce |
| Tempered Glass Cutting Board | Hacaroa (Amazon) | B09XMXBT5S | 4 pc |
References
- McSteen, P., Kellogg, E. A. Molecular, cellular, and developmental foundations of grass diversity. Science. 377 (6606), 599-602 (2022).
- Wang, P., Vlad, D., Langdale, J. A. Finding the genes to build C4 rice. Current Opinion in Plant Biology. 31, 44-50 (2016).
- Wang, P., et al. Candidate regulators of early leaf development in maize perturb hormone signalling and secondary cell wall formation when constitutively expressed in rice. Scientific Reports. 7 (1), 4535 (2017).
- Perico, C., Tan, S., Langdale, J. A. Developmental regulation of leaf venation patterns: Monocot versus eudicots and the role of auxin. New Phytologist. 234 (3), 783-803 (2022).
- Wang, P., Kelly, S., Fouracre, J. P., Langdale, J. A. Genome-wide transcript analysis of early maize leaf development reveals gene cohorts associated with the differentiation of C4 Kranz anatomy. The Plant Journal. 75 (4), 656-670 (2013).
- Liu, W. Y., et al. Regulators of early maize leaf development inferred from transcriptomes of laser capture microdissection (LCM)-isolated embryonic leaf cells. Proceedings of the National Academy of Sciences. 119 (35), e2208795119 (2022).
- Liu, W. Y., et al. Anatomical and transcriptional dynamics of maize embryonic leaves during seed germination. Proceedings of the National Academy of Sciences. 110 (10), 3979-3984 (2013).
- Strable, J., Nelissen, H. The dynamics of maize leaf development: Patterned to grow while growing a pattern. Current Opinion in Plant Biology. 63, 102038 (2021).
- Reynolds, J. O., Eisses, J. F., Sylvester, A. W. Balancing division and expansion during maize leaf morphogenesis: Analysis of the mutant, warty-1. Development. 125 (2), 259-268 (1998).
- Richardson, A. E., et al. Evolution of the grass leaf by primordium extension and petiole-lamina remodeling. Science. 374 (6573), 1377-1381 (2021).
- Johnston, R., Leiboff, S., Scanlon, M. J. Ontogeny of the sheathing leaf base in maize (Zea mays). New Phytologist. 205 (1), 306-315 (2015).
- Sharman, B. C. Developmental anatomy of the shoot of Zea mays L. Annals of Botany. 6 (22), 245-282 (1942).
- Johnston, R., et al. Transcriptomic analyses indicate that maize ligule development recapitulates gene expression patterns that occur during lateral organ initiation. The Plant Cell. 26 (12), 4718-4732 (2014).
- Sharman, B. C. Leaf and bud initiation in the Gramineae. Botanical Gazette. 106 (3), 269-289 (1945).
- Nelissen, H., et al. A local maximum in gibberellin levels regulates maize leaf growth by spatial control of cell division. Current Biology. 22 (13), 1183-1187 (2012).
- Itoh, J., et al. Rice plant development: From zygote to spikelet. Plant and Cell Physiology. 46 (1), 23-47 (2005).
- Ritchie, S. W., Hanway, J. J., Benson, G. O. How a corn plant develops. Iowa State University of Science and Technology. , (1986).
- Freeling, M., Walbot, V. . The Maize Handbook. , (2013).
- Freeling, M., Hake, S. Developmental genetics of mutants that specify knotted leaves in maize. Genetics. 111 (3), 617-634 (1985).
- Durbak, A. R., et al. Transport of boron by the tassel-less1 aquaporin is critical for vegetative and reproductive development in maize. The Plant Cell. 26 (7), 2978-2995 (2014).
- Langdale, J. A., Lane, B., Freeling, M., Nelson, T. Cell lineage analysis of maize bundle sheath and mesophyll cells. Developmental Biology. 133 (1), 128-139 (1989).
- Bosabalidis, A. M., Evert, R. F., Russin, W. A. Ontogeny of the vascular bundles and contiguous tissues in the maize leaf blade. American Journal of Botany. 81 (6), 745-752 (1994).
- Junqueira, N. E. G., et al. Anatomy and ultrastructure of embryonic leaves of the C4 species Setaria viridis. Annals of Botany. 121 (6), 1163-1172 (2018).
- Robil, J. M., McSteen, P. Hormonal control of medial-lateral growth and vein formation in the maize leaf. New Phytologist. 238 (1), 125-141 (2023).
- Donnelly, P. M., Bonetta, D., Tsukaya, H., Dengler, R. E., Dengler, N. G. Cell cycling and cell enlargement in developing leaves of Arabidopsis. Developmental Biology. 215 (2), 407-419 (1999).
- Govindaraju, P., Verna, C., Zhu, T., Scarpella, E. Vein patterning by tissue-specific auxin transport. Development. 147 (13), (2020).
- Linh, N. M., Scarpella, E. Leaf vein patterning is regulated by the aperture of plasmodesmata intercellular channels. PLoS Biology. 20 (9), e3001781 (2022).
- Verna, C., Ravichandran, S. J., Sawchuk, M. G., Linh, N. M., Scarpella, E. Coordination of tissue cell polarity by auxin transport and signaling. eLife. 8, e51061 (2019).
- Sawchuk, M. G., Head, P., Donner, T. J., Scarpella, E. Time-lapse imaging of Arabidopsis leaf development shows dynamic patterns of procambium formation. New Phytologist. 176 (3), 560-571 (2007).
- Linh, N. M., Scarpella, E. Confocal imaging of developing leaves. Current Protocols. 2 (1), e349 (2022).
- Poethig, R. S., Szymkowiak, E. J. Clonal analysis of leaf development in maize. Maydica. 40, 67-76 (1995).
- Sprangers, K., Thys, S., van Dusschoten, D., Beemster, G. T. S. Gibberellin enhances the anisotropy of cell expansion in the growth zone of the maize leaf. Frontiers in Plant Science. 11, 1163 (2020).
- Tsuda, K., et al. KNOTTED1 cofactors, BLH12 and BLH14, regulate internode patterning and vein anastomosis in maize. The Plant Cell. 29 (5), 1105-1118 (2017).
- Schindelin, J., et al. FIJI: An open-source platform for biological-image analysis. Nature Methods. 9 (7), 676-682 (2012).
- Plant Care Protocols - Maize. The Donald Danforth Plant Science Center's Plant Growth Facility Available from: https://www.danforthcenter.org/our-work/core-facilities/plant-growth/ (2019)
- Mir, R., et al. A DII domain-based auxin reporter uncovers low auxin signaling during telophase and early G1. Plant Physiology. 173 (1), 863-871 (2017).
- DeBlasio, S. L., Sylvester, A. W., Jackson, D. Illuminating plant biology: Using fluorescent proteins for high-throughput analysis of protein localization and function in plants. Briefings in Functional Genomics. 9 (2), 129-138 (2010).
- Wu, Q., Luo, A., Zadrozny, T., Sylvester, A., Jackson, D. Fluorescent protein marker lines in maize: Generation and applications. The International Journal of Developmental Biology. 57 (6-8), 535-543 (2013).
- Mohanty, A., et al. Advancing cell biology and functional genomics in maize using fluorescent protein-tagged lines. Plant Physiology. 149 (2), 601-605 (2009).
- Baker, R. F., et al. Sucrose transporter ZmSut1 expression and localization uncover new insights into sucrose phloem loading. Plant Physiology. 172 (3), 1876-1898 (2016).
- Gallavotti, A., Yang, Y., Schmidt, R. J., Jackson, D. The relationship between auxin transport and maize branching. Plant Physiology. 147 (4), 1913-1923 (2008).
- Krishnakumar, V., et al. A maize database resource that captures tissue-specific and subcellular-localized gene expression, via fluorescent tags and confocal imaging (Maize Cell Genomics Database). Plant and Cell Physiology. 56 (1), 12 (2015).
- Snapp, E. L. Fluorescent proteins: A cell biologist's user guide. Trends in Cell Biology. 19 (11), 649-655 (2009).
- Geng, Y., Zhou, Y. Confocal live imaging of shoot apical meristems from different plant species. Journal of Visualized Experiments. (145), e59369 (2019).
- Stanislas, T., Hamant, O., Traas, J. In-vivo analysis of morphogenesis in plants. Methods in Cell Biology. 139, 203-223 (2017).
- Fodor, E., Ayaydin, F. Fluorescent probes and live imaging of plant cells. Advances in Plant Ecophysiology Techniques. , 241-251 (2018).
- Grandjean, O., et al. In vivo analysis of cell division, cell growth, and differentiation at the shoot apical meristem in Arabidopsis. The Plant Cell. 16 (1), 74-87 (2004).
- Conklin, P. A., Johnston, R., Conlon, B. R., Shimizu, R., Scanlon, M. J. Plant homeodomain proteins provide a mechanism for how leaves grow wide. Development. 147 (20), (2020).
- Shaw, S. L. Imaging the live plant cell. The Plant Journal. 45 (4), 573-598 (2006).
- Klaus, A. V., Schawaroch, V., Frischmann, K. J. Confocal imaging and three-dimensional visualization of thick autofluorescent specimens. Methods in Molecular Biology. 1075, 213-225 (2014).
- Forster, B., Van De Ville, D., Berent, J., Sage, D., Unser, M. Extended depth-of-focus for multi-channel microscopy images: a complex wavelet approach. 2004 2nd IEEE International Symposium on Biomedical Imaging: Nano to Macro (IEEE Cat No. 04EX821). IEEE. , 660-663 (2004).
- Preibisch, S., Saalfeld, S., Tomancak, P. Globally optimal stitching of tiled 3D microscopic image acquisitions. Bioinformatics. 25 (11), 1463-1465 (2009).
- Phillips, K. A., et al. vanishing tassel2 encodes a grass-specific tryptophan aminotransferase required for vegetative and reproductive development in maize. The Plant Cell. 23 (2), 550-566 (2011).
- Robil, J. M., et al. GrasVIQ: An image analysis framework for automatically quantifying vein number and morphology in grass leaves. The Plant Journal. 107 (2), 629-648 (2021).
- Helliwell, C. A., Chandler, P. M., Poole, A., Dennis, E. S., Peacock, W. J. The CYP88A cytochrome P450, ent-kaurenoic acid oxidase, catalyzes three steps of the gibberellin biosynthesis pathway. Proceedings of the National Academy of Sciences. 98 (4), 2065-2070 (2001).
- Gutierrez, R., Quiroz-Figueroa, F., Vazquez-Ramos, J. M. Maize cyclin D2 expression, associated kinase activity and effect of phytohormones during germination. Plant and Cell Physiology. 46 (1), 166-173 (2005).
- Atkinson, J. A., Wells, D. M. An updated protocol for high throughput plant tissue sectioning. Frontiers in Plant Science. 8, 1721 (2017).
- Lux, A., Morita, S., Abe, J., Ito, K. An improved method for clearing and staining free-hand sections and whole-mount samples. Annals of Botany. 96 (6), 989-996 (2005).
- Heriche, M., Arnould, C., Wipf, D., Courty, P. E. Imaging plant tissues: Advances and promising clearing practices. Trends in Plant Science. 27 (6), 601-615 (2022).
- Kurihara, D., Mizuta, Y., Sato, Y., Higashiyama, T. ClearSee: A rapid optical clearing reagent for whole-plant fluorescence imaging. Development. 142 (23), 4168-4179 (2015).
- Chuck, G., Muszynski, M., Kellogg, E., Hake, S., Schmidt, R. J. The control of spikelet meristem identity by the branched silkless1 gene in maize. Science. 298 (5596), 1238-1241 (2002).
- Tran, T. M., et al. An optimized whole-mount immunofluorescence method for shoot apices. Current Protocols. 1 (4), e101 (2021).
- O'Connor, D. L. PINs Lost and PINs Gained: Auxin-Transport Mediated Patterning in the Grasses. University of California, Berkeley. , (2012).
- Sharman, B. C., Hitch, P. A. Initiation of procambial strands in leaf primordia of bread wheat, Triticum aestivum L. Annals of Botany. 31 (2), 229-243 (1967).
- Serra, L., Tan, S., Robinson, S., Langdale, J. A. Flip-flap: A simple dual-view imaging method for 3D reconstruction of thick plant samples. Plants. 11 (4), 506 (2022).
- Colin, L., et al. Imaging the living plant cell: From probes to quantification. The Plant Cell. 34 (1), 247-272 (2022).
Reprints and Permissions
Request permission to reuse the text or figures of this JoVE article
Request PermissionExplore More Articles
This article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. All rights reserved