
Method Article
انقسام الشبكية كتحضير مسطح محسن لدراسة الخلايا العصبية للطبقة النووية الداخلية في شبكية الفقاريات
In This Article
Summary
يقدم هذا العمل إعدادا بديلا لشبكية العين المسطحة حيث تتيح إزالة أجسام الخلايا المستقبلة للضوء انتشارا أسرع للأجسام المضادة وتحسين وصول ماصة التصحيح إلى الخلايا العصبية الداخلية للشبكية للكيمياء المناعية والتهجين في الموقع وتجارب الفيزيولوجيا الكهربية.
Abstract
الخلايا ثنائية القطب والخلايا الأفقية لشبكية الفقاريات هي الخلايا العصبية الأولى التي تعالج المعلومات البصرية بعد اكتشاف الفوتونات بواسطة المستقبلات الضوئية. يؤدون عمليات أساسية مثل التكيف مع الضوء وحساسية التباين والتوافق المكاني واللوني. إن الفهم الكامل للدوائر الدقيقة والآليات الكيميائية الحيوية التي تحكم سلوكهم سيعزز أبحاث علم الأعصاب البصري وطب العيون. ومع ذلك ، فإن الاستعدادات الحالية لفحص الخلايا ثنائية القطب والأفقية (حوامل شبكية العين الكاملة والشرائح الرأسية) محدودة في قدرتها على التقاط تشريح وفسيولوجيا هذه الخلايا. في هذا العمل ، نقدم طريقة لإزالة أجسام الخلايا المستقبلة للضوء من شبكية فأر حية مسطحة ، مما يوفر وصولا معززا إلى الخلايا ثنائية القطب والأفقية من أجل تثبيت التصحيح بكفاءة ووضع العلامات المناعية السريعة. يتم تحضير شبكية العين المنقسمة عن طريق وضع شبكية فأر معزولة بين قطعتين من النيتروسليلوز ، ثم تقشيرها برفق. يقسم الفصل الشبكية فوق الطبقة الضفيرية الخارجية مباشرة لإنتاج قطعتين من النيتروسليلوز ، واحدة تحتوي على أجسام الخلايا المستقبلة للضوء والأخرى تحتوي على الشبكية الداخلية المتبقية. على عكس شرائح الشبكية الرأسية ، فإن تحضير الشبكية المنقسمة لا يقطع العمليات المتغصنة للخلايا العصبية الداخلية للشبكية ، مما يسمح بالتسجيلات من الخلايا ثنائية القطب والأفقية التي تدمج مساهمات الشبكات المقترنة بتقاطع الفجوة وخلايا الأمكرين واسعة المجال. يوضح هذا العمل براعة هذا التحضير لدراسة الخلايا الأفقية وثنائية القطب في الفيزيولوجيا الكهربية والكيمياء الهيستولوجية المناعية وتجارب التهجين في الموقع .
Introduction
شبكية العين هي نسيج عصبي رقيق يقع في العين الخلفية حيث يتم اعتراض الضوء ومعالجته في إشارة كهروكيميائية يمكن تفسيرها من قبل الدماغ. في الجزء الخلفي من شبكية العين ، يتم تحفيز المستقبلات الضوئية للقضيب والمخروط عن طريق الضوء ، مما يقلل من معدل إطلاق منشط الناقل العصبي ، الغلوتامات1. أول الخلايا العصبية التي تختبر وتستجيب لهذا التغيير الناجم عن الضوء في تركيز الغلوتامات هي الخلايا ثنائية القطب (BCs) والخلايا الأفقية (HCs) ، التي توجد سوماتها في المنطقة الخارجية من الطبقة النووية الداخلية (INL). تؤدي هذه الخلايا العصبية من الدرجة الثانية المرحلة الأولى من معالجة الإشارات في شبكية العين وتشكل السمات الحرجة للرؤية مثل التكيف مع الضوء وحساسية التباين والمقابلة المكانية / اللونية2. في حين أن هذه الوظائف قد نسبت إلى BCs و HCs ، فإن الدوائر والآليات الكيميائية الحيوية الكامنة وراء هذه العمليات ليست مفهومة تماما3. لذلك ، فإن تطوير الأدوات والأساليب لاستكشاف فسيولوجيا BC و HC له أهمية قصوى.
أثبتت أقسام شبكية العين الرأسية (المستعرضة) منذ فترة طويلة أنها النموذج الأكثر عملية لدراسة BCs و HCs. ومع ذلك ، لا يمكن الوصول إلى جوانب معينة من فسيولوجيا BC و HC للمجرب بموجب هذا النموذج. لا تعكس التسجيلات المباشرة من HCs أو القياسات غير المباشرة لآثارها على BCs الاتصال الداخلي لشبكية العين حيث يتم قطع العمليات الجانبية لهذه الخلايا أثناء التقطيع. تتحايل مستحضرات شبكية العين الكاملة على هذه المشكلة من خلال الحفاظ على هذه العمليات الجانبية ، لكن طبقات الشبكية المحيطة تشكل تحديا للوصول إلى هذه الخلايا4. في حين أن هناك أمثلة وفيرة على التلوين المناعي5،6،7،8 وتسجيلات المشبكالتصحيح 9 من الخلايا العصبية INL في شبكية العين بأكملها ، هناك فرصة لتسريع وتبسيط جمع هذه البيانات. وبالتالي ، فإن القيود المتأصلة في المقاطع المستعرضة وتحديات نموذج التركيب بأكمله ألهمت تطوير هذا التحضير البديل لشبكية العين المسطحة.
يصف العمل التالي بروتوكولا لإزالة طبقة المستقبلات الضوئية بسهولة من شبكية العين الحية المسطحة لتعزيز الوصول إلى BCs و HCs من أجل لقط رقعة مبسط ووضع العلامات المناعية بشكل أسرع وأكثر كفاءة. يؤدي تقشير قطعتين من غشاء النيتروسليلوز المتصلين بجانبي الشبكية المعزولة إلى تمزيق الأنسجة من خلال محاور مستقبلات الضوء ، تاركا شبكية منقسمة تحتفظ بالطبقة الضفيرية الخارجية (OPL) وجميع طبقات الشبكية الداخلية. بينما وصف آخرون بروتوكولات لفصل طبقات شبكية العين ميكانيكيا ، فإن هذه الطرق إما غير مناسبة لتطبيقات التثبيت والفحص المجهري أو تتطلب معالجة مملة للأنسجة. تتطلب العديد من هذه الطرق أنسجة مجمدة أو مجففة بالتجميد لفصل الطبقة ، مما يجعلها غير متوافقة مع تجارب الفيزيولوجيا الكهربية10،11،12. تم تصميم البعض الآخر للأنسجة الحية ، ولكنه يتطلب 5-15 تقشيرا متتابعا باستخدام ورق الترشيح4،11 أو العلاج بالتربسين13 لإزالة المستقبلات الضوئية. تعمل التقنية الموصوفة هنا على تحسين سابقاتها من خلال تبسيط إجراء إزالة المستقبلات الضوئية وتوسيع ذخيرة التطبيقات النهائية.
Protocol
تم تزويد الفئران بالماء والطعام حسب الطلب وتم الحفاظ عليها في دورة الضوء / الظلام لمدة 12 ساعة. تم القتل الرحيم للفئران عن طريق التعرض للأيزوفلوران متبوعا بخلع عنق الرحم. كانت جميع الإجراءات الحيوانية متوافقة مع إرشادات المعاهد الوطنية للصحة ووافقت عليها لجنة رعاية واستخدام المؤسسية بجامعة أوريغون للصحة والعلوم.
ملاحظة: يجب إجراء استئصال العين وتشريح الشبكية وتقسيم الشبكية في أسرع وقت ممكن للحفاظ على صحة الأنسجة الحية. اهدف إلى إكمال عملية التشريح في < 4 دقائق لكل عين. يجب تنفيذ هذه الخطوات الثلاث بالتتابع. الفئران من النوع البري: تم استخدام الفئران البالغة (>3 أشهر) من الذكور والإناث C57BL / 6J لإجراء التجارب. بالنسبة لمورفولوجيا المشبك العصبي ، تم استخدام الفئران التي تعبر عن بروتين الفلورسنت الأخضر (GFP) تحت محفز Pcp2 (Pcp2-cre / GFP) 14 . الفئران المعدلة وراثيا: لتصور الخلية الأفقي مع GFP أثناء تجارب الكيمياء المناعية أو الفيزيولوجيا الكهربية ، تم استخدام فأر ثلاثي الجينات المحورة: vGATFLPo; vGlut2Cre; Ai80d. سلالات vGATFlpo و vGluT2Cre عبارة عن فئران غير مطروقة تعبر عن Flpo أو Cre recombinase في اتجاه مجرى المروجين المعنيين. الماوس Ai80d هو ماوس مراسل متعدد الجوانب (CatCh / EYFP) وسيعبر فقط عن رودوبسين القناة القابلة للنفاذ Ca2+ (ChR2) في الخلايا التي تعبر عن Cre و Flpo recombinases. وبالتالي ، فإن الماوس الثلاثي المعدل وراثيا يعبر فقط عن ChR2 في الخلايا التي لها تاريخ من تعبير VGAT و vGluT2.
1. إعداد المواد لتشريح الشبكية وتقسيم الشبكية
- تحضير قطع من غشاء النيتروسليلوز
ملاحظة: يؤدي فصل الشبكية المنقسمة عن غشاء النيتروسليلوز إلى تقليل مضان الخلفية في الفحص المجهري ويبسط تسجيل مشبك التصحيح. يمكن إجراء إزالة الغشاء قبل أو بعد تثبيت الأنسجة. بالنسبة لشبكية العين المنقسمة الثابتة ، ليس من الضروري معالجة قطع غشاء النيتروسليلوز. بالنسبة للشبكية الحية المنقسمة ، عالج الغشاء وفقا للخطوات 1.1.3 - 1.1.5 لتسهيل الانفصال اللطيف عن الأنسجة.- قطع 16 قطعة (أو أكثر) من غشاء النيتروسليلوز إلى مربعات 5 مم × 5 مم. يمكن تحضير كمية إضافية بكميات كبيرة وتخزينها للاستخدام في المستقبل.
- ضع جانبا نصف قطع الغشاء لاستخدامها لاحقا. لن يتم التعامل مع هذه القطع بمحلول حظر.
- احتضن القطع المتبقية في محلول مانع IHC خال من المنظفات (مثل 3٪ مصل حصان + 0.025٪ NaN3 مخفف في PBS) لمدة 10 دقائق في درجة حرارة الغرفة ، مع رجها برفق.
تنبيه: استخدم معدات الوقاية الشخصية المناسبة عند التعامل مع NaN3 ، لأنه سم قوي. - اغسل قطع الغشاء جيدا عن طريق الحضانة في وسائط أميس المخزنة بالبيكربونات لمدة 10 دقائق في درجة حرارة الغرفة ، مع رجها برفق.
- جفف تماما قطع الغشاء المسدودة بالهواء (~ 20 دقيقة). قم بتسمية وتخزين قطع الغشاء في درجة حرارة الغرفة ، مع إبقائها منفصلة عن قطع الغشاء غير المعالجة.
- إعداد وسائل الإعلام أميس
- تحضير وسائط Ames المخزنة بالبيكربونات والحفاظ على المحلول في درجة حرارة الغرفة تحت كربوجين ثابت (95٪ O 2 و 5٪ CO2).
2. استئصال عين الفأر
- القتل الرحيم للفأر بأي طريقة متاحة وفقا لإرشادات IACUC المؤسسية.
- اقلب الماوس على جانب واحد واستخدم إصبعين للضغط برفق حول مقبس العين. سيؤدي ذلك إلى انتفاخ العين من الجمجمة.
- باستخدام مقص تشريح منحني ، قم بقص أسفل العين المنتفخة لقطع العصب البصري وفصل العين عن الجمجمة.
- اغرف العين بالمقص وضعها في طبق بتري مليء بوسائط أميس الباردة المثلجة.
ملاحظة: بالنسبة للتطبيقات النهائية التي سيتم فيها تثبيت الأنسجة بعد الانقسام ، يمكن استخدام برنامج تلفزيوني بارد مثلج بدلا من وسائط أميس. - كرر الخطوات من 2.1 إلى 2.4 للعين المتبقية.
3. تشريح الشبكية
- استخدم ماصة نقل الزجاج المخصصة لنقل عين واحدة إلى طبق بتري جديد يحتوي على وسائط أميس طازجة مثلجة.
ملاحظة: تمنع الفتحة الواسعة لماصة النقل المخصصة السحق العرضي للأنسجة ، ويقلل استخدام الزجاج من التصاق الأنسجة بجدران الماصة. ومع ذلك ، فإن ماصة نقل البلاستيك واسعة الفم مقبولة أيضا إذا كان المجرب بارعا بالفعل في استخدام هذه الأداة. - استخدم الملقط لتثبيت العين عن طريق تثبيت نسيجها الضام الإضافي في قاع طبق بتري. بعد ذلك ، قم بثقب العين على طول خط ora serrata باستخدام إبرة 25G لإنشاء نقطة دخول لمقص Vannas.
- استخدم مقص Vannas لقطع خط ora serrata حتى تتحرر القرنية من بقية العين (الشكل التكميلي 1A). قم بإزالة العدسة من فنجان العين باستخدام الملقط (الشكل التكميلي 1 ب).
- استخدم الماصة الزجاجية المخصصة لنقل فنجان العين إلى حجم كبير (≥100 مل) من Ames الكربوجين وكرر الخطوات 3.1 - 3.3 مع العين المتبقية.
ملاحظة: يتم وضع أكواب العين في أميس الكربوجينية للحفاظ على صحة الأنسجة بينما يتم إجراء التشريح على العين الأخرى. - انقل كوبا واحدا إلى طبق بتري مملوء بالكربوجين الطازج.
- باستخدام مقص Vannas ، قم بعمل قصاصة صغيرة إلى الداخل من حافة الصلبة ، ثم استخدم زوجين من الملقط لتقشير الصلبة بعيدا عن شبكية العين (الشكل التكميلي 1C). تجنب الاستيلاء على شبكية العين بالملقط. بدلا من ذلك ، اسحب اللوحات الصلبة الناتجة عن قصاصة المقص.
- استخدم مقص Vannas لقص العصب البصري الذي يربط الصلبة والشبكية (الشكل التكميلي 1D) ، ثم أخرج الشبكية برفق من الصلبة باستخدام المقص أو الملقط لعزل الشبكية. (الشكل 1 أ).
ملاحظة: على الرغم من أن RPE سيبقى عادة متصلا بفنجان العين ، فلا يلزم اتخاذ خطوات إضافية لإزالة RPE في حالة توصيله بالشبكية. في هذه المرحلة ، يمكن تقليم حواف الشبكية اختياريا بمشرط لمنع التجعيد أثناء خطوة التسطيح (الشكل 1 ب). - استخدم مشرطا لقطع شبكية العين إلى نصفين أو أرباع (الشكل 1C) ، ثم استخدم ماصة النقل المخصصة لإعادة القطع إلى حجم كبير (≥ 100 مل) من وسائط Ames المكركرة باستمرار.
ملاحظة: اختيار النصفين أو الأرباع غير موضوعي. اختر الخيار الأفضل للتطبيق المطلوب. - كرر الخطوات من 3.5 إلى 3.8 للعين المتبقية قبل الشروع في شق الشبكية.
4. تقسيم شبكية العين
- تخلص من وسائط Ames من أطباق Petri واستبدلها ب Ames الطازج.
ملاحظة: للحفاظ على الكربوجين طوال الفترة المتبقية من إجراء تقسيم الشبكية ، استبدل الوسائط الموجودة في طبق بتري بأميس الطازج كل 5 دقائق تقريبا. - باستخدام ماصة النقل المخصصة ، ضع قطعة من شبكية العين على شريحة زجاجية (7.5 سم × 5 سم) ، وجانب خلية العقدة لأعلى ، ثم قم بتسويتها عن طريق إزالة السائل المحيط بمسح مهمة دقيق (الشكل 1 د). إذا لزم الأمر ، اسحب حواف الشبكية برفق باستخدام فرشاة رسم ذات طرف دقيق تحت مجهر التشريح.
- استخدم الملقط لخفض قطعة جافة 5 مم × 5 مم من غشاء النيتروسليلوز على شبكية العين ، مما يؤدي إلى التصاق جانب الخلية العقدية (الشكل 1E).
ملاحظة: إذا كانت إزالة الغشاء من الأنسجة الحية مطلوبة (أي الفيزيولوجيا الكهربية) ، فاستخدم قطعة جافة من الغشاء المعالج بالمصل لهذه الخطوة (انظر الخطوات 1.1.3 - 1.1.5 للحصول على التفاصيل). هذا يقلل من قوة الالتصاق بطبقة الخلايا العقدية ، مما يسهل إزالة شبكية العين من النيتروسليلوز بعد الانقسام. - اقلب الشبكية بحيث يستقر النيتروسليلوز على الشريحة الزجاجية وضع قطعة جافة من غشاء 5 مم × 5 مم على جانب مستقبلات الضوء من شبكية العين (الشكل 1F).
- المس الطرف المبلل لفرشاة الرسم بالفراغ بين الغشاءين واسمح للعمل الشعري بامتصاص أميس في الساندويتش (الشكل 1 ج). هذا يقلل من التصاق الأغشية بالشبكية وهو ضروري فقط إذا تم تجفيف شبكية العين بشكل مفرط بمسح المهمة الحساسة.
ملاحظة: إذا فقدت شبكية العين مظهرها اللامع ، فقد تم تجفيفها بشكل مفرط ، والخطوة 4.5 ضرورية. - لضمان التصاق موحد ، قم بتطبيق ضغط خفيف لأسفل على الغشاء العلوي باستخدام فرشاة طلاء مبللة (الشكل 1H).
- أثناء تثبيت الغشاء السفلي على الزجاج بزوج واحد من الملقط ، استخدم حركة بطيئة وثابتة لتقشير الغشاء العلوي برفق باستخدام زوج ثان من الملقط. سيؤدي ذلك إلى انقسام شبكية العين فوق OPL مباشرة (الشكل 1I).
- تخلص من الغشاء العلوي الذي يحتوي على المستقبلات الضوئية (الشكل 1J ، يسار). يحتوي الغشاء السفلي على الشبكية الداخلية ، ويشار إليها من الآن فصاعدا باسم الشبكية المنقسمة (الشكل 1J ، يمين).
- أعد الشبكية المنقسمة على الفور إلى وسائط Ames الكربوجينية.
ملاحظة: بالنسبة للتجارب على الأنسجة الحية ، قد تستفيد شبكية العين من فترة نقاهة تتراوح بين 15 و 30 دقيقة في أميس الكربوجينية بعد الانقسام.

الشكل 1: إجراء انقسام الشبكية . (أ) بعد الاستئصال وإعداد فنجان العين في وسائط PBS أو Ames الباردة ، اعزل شبكية الفأر عن فنجان العين واستبدل PBS بدرجة حرارة الغرفة ، وسائط Ames المكربة. (ب) باستخدام مشرط ، قم بقص حواف الشبكية بعيدا حتى لا تكون هناك مناطق بها تجعيد داخلي (اختياري). ج: قطع الشبكية إلى أرباع أو أنصاف باستخدام مشرط. (د) ضع قطعة واحدة من شبكية العين على شريحة زجاجية (جانب خلية العقدة لأعلى) باستخدام ماصة النقل المخصصة وقم بإزالة جميع أميس الزائدة باستخدام مسح المهام الدقيق. تأكد من أن شبكية العين شبه الجافة ملقاة على الزجاج قبل الانتقال إلى الخطوة التالية. استخدم طرف فرشاة الرسم المبلل من Ames لفتح مناطق الشبكية غير المسطحة برفق. (ه) باستخدام الملقط ، ضع قطعة مقطوعة مسبقا من غشاء النيتروسليلوز الجاف (5 مم × 5 مم) على الشبكية المسطحة. (و) اقلب قطعة النيتروسليلوز بحيث يكون جانب المستقبلات الضوئية من الشبكية متجها لأعلى. ثم ضع قطعة جافة أخرى من الغشاء على شبكية العين. (ز) المس الطرف المبلل للفرشاة بالفراغ بين الغشاءين واسمح للشعيرات الدموية بامتصاص أميس في الساندويتش. هذا يقلل من التصاق الأغشية بالشبكية وهو ضروري فقط إذا تم تجفيف الشبكية بشكل مفرط بمسح المهمة الحساسة. (H) استخدم طرف فرشاة الرسم المبلل للضغط برفق لأسفل على مركز الشبكية المحصورة. (I) استخدم زوجا واحدا من الملقط لتثبيت القطعة السفلية من الغشاء على الشريحة الزجاجية ، مع استخدام زوج آخر من الملقط لتقشير القطعة العلوية من الغشاء برفق بعيدا عن الجزء السفلي. (J) تظل الشبكية الداخلية (يسار) على الغشاء السفلي بينما يتم سحب المستقبلات الضوئية (اليمنى) بعيدا مع الغشاء العلوي. تم الحصول على الألواح (أ) و (ب) و (ج) و (د) و (ياء) باستخدام مجهر التشريح. يمثل شريط المقياس حوالي 1 مم ؛ تم الحصول على اللوحات (E-I) بكاميرا هاتف ذكي بدون تكبير. يرجى النقر هنا لعرض نسخة أكبر من هذا الرقم.
{kind=link}
5. تحضير الشبكية المنقسمة لتجارب التألق المناعي
ملاحظة: ستظل الشبكية المنقسمة متصلة بغشاء النيتروسليلوز حتى الخطوة 5.5. أكمل إما الخطوات 5.1 أو 5.2 أو 5.3 أو 5.4 ، وليس الأربعة كلها لأنها لتجارب مختلفة.
تنبيه: استخدم معدات الوقاية الشخصية المناسبة وتابع بحذر عند التعامل مع بارافورمالدهيد (مثبت).
- التحضير للتألق المناعي المسطح
- احتضان الشبكية المنقسمة في 4٪ بارافورمالدهيد على الثلج لمدة 30 دقيقة باستخدام محلول كاف لتغطية الشبكية بالكامل.
- اغسل شبكية العين المنقسمة 3x في 5-10 مل من درجة حرارة الغرفة PBS. وقفة اختيارية: يمكن ترك شبكية العين المنقسمة في PBS عند 4 درجات مئوية لمدة تصل إلى 24 ساعة.
- التحضير للتألق المناعي مع المقاطع الرأسية للشبكية المنقسمة
- احتضان الشبكية المنقسمة في 4٪ بارافورمالدهيد على الثلج لمدة 30 دقيقة باستخدام محلول كاف لتغطية الشبكية بالكامل.
- اغسل شبكية العين المنقسمة 3x في 5-10 مل من درجة حرارة الغرفة PBS. وقفة اختيارية: يمكن ترك شبكية العين المنقسمة في PBS عند 4 درجات مئوية لمدة تصل إلى 24 ساعة.
- مع استمرار الغشاء متصلا ، اغمر الشبكية المنقسمة بالتتابع في 10٪ و 20٪ و 30٪ سكروز عند 4 درجات مئوية لمدة 1 ساعة لكل منها لحماية الأنسجة بالتبريد.
- قم بتضمين شبكية العين المنقسمة المحمية بالتبريد في مركب درجة حرارة القطع المثلى (O.C.T.) وقم بتخزينها في -80 درجة مئوية (حتى 6 أشهر) حتى التقسيم بالتبريد.
- قم بإزالة شبكية العين المنقسمة المدمجة من -80 درجة مئوية واستخدم cryostat لقطع الأجزاء بسمك 20 ميكرومتر. قم بتركيب الأقسام على شرائح مجهر زجاجية مشحونة بالكهرباء الساكنة ، واتركها تجف في الهواء ، ثم قم بتخزينها في -20 درجة مئوية لمدة تصل إلى 6 أشهر.
- التحضير للتهجين الفلوري المزدوج في الموقع والكيمياء الهيستولوجية المناعية
- احتضان الشبكية المنقسمة في 4٪ بارافورمالدهيد على الجليد لمدة 2 ساعة باستخدام محلول كاف لتغطية شبكية العين بالكامل.
- اغسل شبكية العين المنقسمة 3x في 5-10 مل من درجة حرارة الغرفة PBS. وقفة اختيارية: يمكن ترك شبكية العين المنقسمة في PBS عند 4 درجات مئوية لمدة تصل إلى 24 ساعة.
- التحضير للفيزيولوجيا الكهربية
- تحضير ماصات التصحيح عن طريق سحب ماصات زجاج البورسليكات سميكة الجدران مع خيوط باستخدام مجتذب micropipite. استخدم فقط الماصات ذات المقاومة المقاسة بين 6-10 MΩ.
- املأ الماصات المسحوبة مرة أخرى بمحلول داخلي يحتوي (بالملليمتر): 125 K-gluconate ، 8 KCl ، 5 HEPES ، 1 MgCl 2 ، 1 CaCl 2 ،0.2 EGTA ، 3 ATP-Mg ، و 0.5 GTP-Na.
- إزالة الشبكية المنقسمة من غشاء النيتروسليلوز
- باستخدام قلم حاجز مسعور ، قم بإعداد الآبار الدائرية على شريحة مجهرية (قطرها ~ 1 سم) واتركها تجف في الهواء لمدة 5-10 دقائق.
- ضع شبكية العين المنقسمة داخل آبار القلم الحاجز الكارهة للماء المحضرة وأضف ما يكفي من برنامج تلفزيوني لتغطيتها بالكامل.
- تحت مجهر التشريح ، ادفع شعيرات فرشاة الرسم الدقيقة أسفل حواف الأنسجة وارفعها برفق لأعلى. بهذه الطريقة ، اعمل حول الشبكية في دائرة لرفعها بعيدا عن الغشاء.
- استخدم ملقط لإزالة الغشاء من تحت القطعة العائمة من شبكية العين.
- قم بشفط PBS المتبقي بعناية بحيث تستقر قطعة الشبكية على شريحة المجهر ، وجانب خلية العقدة لأسفل.
ملاحظة: لا يتم تنفيذ الخطوات التالية بالتتابع. اختر البروتوكول المناسب للتطبيق المطلوب (أي التلوين المناعي أو التألق المزدوج في الموقع [FISH] والكيمياء الهيستولوجية المناعية [IHC] أو الفيزيولوجيا الكهربية).
6. التلوين المناعي
- إذا لم تكن مستعدة بعد ، استخدم قلم حاجز كاره للماء لإنشاء آبار دائرية على شريحة مجهرية (قطرها ~ 1 سم) واتركها تجف في الهواء لمدة 5-10 دقائق. سيتم تنفيذ جميع خطوات الحضانة وخطوات الغسيل داخل آبار الحظائر هذه.
- احتضان الشبكية المنقسمة أو أقسام الشبكية المنقسمة الرأسية في محلول حضانة الأجسام المضادة (AIS: 3٪ مصل حصان ، 0.5٪ Triton X-100 ، 0.025٪ NaN3 في PBS) لمدة 30 دقيقة في درجة حرارة الغرفة.
- احتضان الشبكية المنقسمة أو أقسام الشبكية المنقسمة العمودية بالأجسام المضادة الأولية المخففة في AIS لمدة 1 ساعة في درجة حرارة الغرفة.
ملاحظة: سيتطلب وقت حضانة الأجسام المضادة الأولية تحسينا لأهداف البروتين والأجسام المضادة المختلفة. - اغسل المنديل 3x في درجة حرارة الغرفة PBS.
- احتضان الأنسجة مع الأجسام المضادة الثانوية المخففة في AIS لمدة 1 ساعة في درجة حرارة الغرفة. اغسل المنديل 3x في درجة حرارة الغرفة PBS.
- إذا كان التلوين النووي مطلوبا ، فقم باحتضان الأنسجة باستخدام DAPI المخفف في برنامج تلفزيوني لمدة 30 ثانية في درجة حرارة الغرفة. اغسل المنديل 1x في درجة حرارة الغرفة PBS.
- ضع قطرة من وسائط تثبيت الشريحة على كل قطعة من الأنسجة وقم بتركيب غطاء زجاجي.
- ضع طلاء الأظافر حول حواف غطاء الغطاء لإغلاق العينة. قم بتخزين الشريحة على حرارة 4 درجات مئوية.
7. FISH المزدوج و IHC
- تخبز شبكية العين المنقسمة على حرارة 40 درجة مئوية لمدة 30 دقيقة في فرن تهجين لزيادة الالتصاق بالشريحة.
- أكمل بروتوكول RNAscope FISH وفقا لبروتوكول الشركة المصنعة مع الاستثناءات والتعديلات التالية:
- لا يلزم اتخاذ خطوة استرجاع المستضد. استخدم البروتياز الثالث مع وقت حضانة 18 دقيقة في درجة حرارة الغرفة.
- قم بإجراء جميع خطوات الغسيل على الشريحة داخل الآبار المصنوعة بواسطة قلم حاجز كاره للماء.
- احتضان العينات في الأجسام المضادة الأولية المخففة (انظر جدول المواد) في برنامج تلفزيوني لمدة 30 دقيقة عند 40 درجة مئوية في فرن التهجين. اغسل العينات 3x في درجة حرارة الغرفة PBS.
- احتضان العينات في الأجسام المضادة الثانوية المخففة (انظر جدول المواد) في برنامج تلفزيوني لمدة 30 دقيقة عند 40 درجة مئوية في فرن التهجين. اغسل العينات 3x في درجة حرارة الغرفة PBS.
- احتضان العينات في 1x DAPI لمدة 30 ثانية في درجة حرارة الغرفة. اغسل العينات 1x في درجة حرارة الغرفة PBS.
- ضع قطرة من وسائط التثبيت المضادة للتلاشي على كل قطعة من الأنسجة وقم بتركيب غطاء زجاجي.
- ضع طلاء الأظافر حول حواف غطاء الغطاء لإغلاق العينة. قم بتخزين الشريحة على حرارة 4 درجات مئوية.
8. الفيزيولوجيا الكهربية
- بعد إزالة غشاء النيتروسليلوز ، انقل شبكية العين المنقسمة إلى غرفة تسجيل المشبك الرقعة وقم بتثبيتها برفق في مكانها باستخدام قيثارة بلاتينية.
- طوال التجربة ، قم باستمرار بتغذية شبكية العين المنقسمة بمحلول Ames المحتفي ب 95٪ O 2 و 5٪ CO2. الحفاظ على الحل بين 32-34 درجة مئوية.
ملاحظة: أثناء التجربة ، يمكن تصور الأنسجة باستخدام مجهر تباين التدرج Dodt. - تحت إضاءة الغرفة ، قم بإجراء تثبيت جهد الخلية بالكامل للتسجيل من الخلايا العصبية INL.
- أثناء التسجيل ، قم بمحاكاة استجابات الضوء باستخدام وحدة حقن ميكروخلوية لتطبيق المركبات الصيدلانية ، أو مصباح LED 470 نانومتر لتحفيز قناة رودوبسين (ChR2).
ملاحظة: يمكن قياس شدة الضوء باستخدام مقياس طاقة ضوئي رقمي.
- أثناء التسجيل ، قم بمحاكاة استجابات الضوء باستخدام وحدة حقن ميكروخلوية لتطبيق المركبات الصيدلانية ، أو مصباح LED 470 نانومتر لتحفيز قناة رودوبسين (ChR2).
9. الفحص المجهري متحد البؤر
- بالنسبة للتألق المناعي متحد البؤر ، التقط صورا باستخدام مجهر متحد البؤر باستخدام هدف غمر الزيت 40x / 1.3 أو 63x / 1.40. استخدم FIJI لضبط السطوع والتباين ولإنشاء إسقاطات Z من مكدسات الصور.
النتائج
انقسام الشبكية يحافظ على المحطات المستقبلة للضوء
للتأكد من أن انقسام الشبكية لا يضر بالتشعبات العصبية من الدرجة الثانية في OPL ، تم تلطيخ المقاطع الرأسية من شبكية العين المنقسمة بالأجسام المضادة ضد بروتين الحويصلة المشبكية synaptophysin (الأخضر) ، وبروتين كيناز C alpha (PKCα ؛ أحمر). يشير النطاق المكثف لوضع العلامات على المشابك عبر الجزء العلوي من الشبكية المنقسمة إلى الاحتفاظ بالنهايات المشبكية المستقبلة للضوء (الشكل 2). علاوة على ذلك ، يكشف تلطيخ PKCα عن مورفولوجيا طبيعية للخلايا ثنائية القطب للقضيب (RBCs). لا توجد نوى مستقبلات ضوئية مرئية ، مما يشير إلى أن شبكية العين مقسمة بين OPL والصف الأعمق من أجسام الخلايا المستقبلة للضوء (الشكل 2).

الشكل 2: تحتفظ الشبكية المنقسمة بالنهايات المستقبلة للضوء. صور مجهرية متحدة البؤر فلورية تظهر مقطعا عرضيا رأسيا لشبكية البصاق تم قطعها بالتبريد (سمك 20 ميكرومتر) بعد إجراء الانقسام. كل صورة هي أقصى إسقاط لمكدس z متحد البؤر. تم تصنيف القسم مناعيا بأجسام مضادة ضد PKCα (أعلى الوسط) و synaptophysin (أعلى اليمين) لتصور كرات الدم الحمراء والحويصلات المشبكية ، على التوالي. تظهر الصورة المدمجة (أسفل) حويصلات متشابكة (خضراء) ، موجودة في أطراف المستقبلات الضوئية ، فوق العمليات القمية لكرات الدم الحمراء (الحمراء) في OPL. يتم تمييز نوى الخلية ب DAPI (أزرق). لا توجد نوى مستقبلات ضوئية مرئية داخل ONL. الاختصارات: ONL = الطبقة النووية الخارجية ؛ OPL = طبقة شبكية خارجية ؛ INL = الطبقة النووية الداخلية ؛ IPL = طبقة الضفيرة الداخلية ؛ GC = خلايا العقدة. قضبان المقياس = 10 ميكرومتر. يرجى النقر هنا لعرض نسخة أكبر من هذا الشكل.
{kind=link}
يتم الحفاظ على مورفولوجيا المشبك في OPL بعد تقسيم الشبكية
باستخدام الماوس الذي يعبر عن GFP في كرات الدم الحمراء تحت مروج pcp2، تم تصنيف 14 بروتينا قبل وبعد المشبكي في OPL لتقييم سلامة هذه الطبقة المشبكية بعد الانقسام14. على الرغم من قوى القص التي تحدث من خلال محاور المستقبلات الضوئية ، فإن الانقسام لا يزعج مورفولوجيا المشابك المستقبلة للضوء BC في OPL ، حيث لوحظ تحديد المواقع الطبيعية لتغصنات كرات الدم الحمراء ، المسمى ل RGS11 ، وشرائط متشابك للمستقبلات الضوئية ، المسمى ل CtBP215 (الشكل 3). لكل اتصال متشابك بين القضبان وكرات الدم الحمراء ، يمكن رؤية RGS11 على أنه نقطة حمراء تقع داخل شكل حدوة الحصان للشرائط المشبكية (خضراء). في تجربة لاحقة ، تم استخدام جسم مضاد ل GPR179 16 لتسمية الأطراف المتغصنة ON-BC بعد المشبكي16 ، وتم استخدام جسم مضاد ل PSD-95 لتسمية أطراف المستقبلات الضوئية للقضيب قبل المشبكي (الشكل التكميلي 2). تؤكد هذه النتائج مرة أخرى استقرار OPL في تحضير الشبكية المنقسمة ، حيث يظهر أن التشعبات الحمراء الحمراء ترتبط ارتباطا وثيقا بشريكها قبل المشبكي ، أطراف القضيب.

الشكل 3: يتم الحفاظ على مورفولوجيا المشبك العصبي في OPL بعد انقسام الشبكية. صور التألق المناعي متحد البؤر لشبكية منقسمة من فأر معدل وراثيا يعبر عن GFP في كرات الدم الحمراء تحت مروج Pcp2. تختلف مستويات تعبير GFP (الأزرق) عبر كرات الدم الحمراء في شبكية العين. بعد الانقسام ، تم إصلاح شبكية العين ، ثم حضنها بأجسام مضادة ضد CtBP2 (أخضر) و RGS11 (أحمر) لتسمية شرائط متشابك للمستقبلات الضوئية ونصائح تغصنية ON-BC ، على التوالي. يمثل كل زوج من الأخضر الأحمر اتصالا متشابكا بين قضيب و ON-BC. قضبان المقياس = 10 ميكرومتر. يرجى النقر هنا لعرض نسخة أكبر من هذا الشكل.
{kind=link}
انقسام شبكية العين يحافظ على جدوى كرات الدم الحمراء
لتقييم صلاحية الخلايا العصبية داخل الشبكية بعد الانقسام ، تم استخدام صبغة نووية غشائية غير منفذة وقريبة من الأشعة تحت الحمراء (MI-NIR) ، والتي تمكن من تحديد الخلايا الميتة. بعد الحضانة مع MI-NIR ، تم إصلاح شبكية العين المنقسمة ، ثم تم تصنيفها بمضادات PKCα لتحديد كرات الدم الحمراء. تكشف الصور المجهرية متحدة البؤر للشبكية المنقسمة عن تباين إقليمي في صلاحية الخلية عبر الأنسجة ، حيث تشهد بعض المناطق معدلات موت الخلايا أعلى من غيرها. قد ينتج هذا التباين عن الضرر الذي يلحق بمناطق معينة من شبكية العين أثناء إجراءات التشريح أو الانقسام أو المناولة (الشكل 4). بالنظر إلى أن أجسام خلايا كرات الدم الحمراء موجودة في المنطقة الخارجية من INL ، بالقرب من موقع الانقسام ، كان هناك ما يبرر إجراء تقييم دقيق لصلاحيتها. أكد التمركز المشترك النادر ل PKCα و MI-NIR أن معظم كرات الدم الحمراء تظل قابلة للحياة بعد انقسام الشبكية (الشكل 4).

الشكل 4: الخلايا ثنائية القطب قابلة للحياة بعد انقسام الشبكية. صور مجهرية متحدة البؤر فلورية تظهر منطقة من شبكية العين المنقسمة في منظور مسطح. بعد الانقسام ، تم تحضين الشبكية الحية بصبغة MI-NIR (حمراء) لمدة 30 دقيقة عند 37 درجة مئوية. ثم تم تثبيت شبكية العين وتصنيفها المناعي بأجسام مضادة ضد PKCα لتصور كرات الدم الحمراء. في هذه المنطقة من شبكية العين ، يكون التمركز المشترك ل PKCα و MI-NIR نادرا. MI-NIR يتزامن مع النوى (الزرقاء) التي لا تنتمي إلى كرات الدم الحمراء. الاختصارات: MI-NIR = غشاء غير منفذ NIR صبغة حية / ميتة. قضبان المقياس = 10 ميكرومتر. يرجى النقر هنا لعرض نسخة أكبر من هذا الشكل.
{kind=link}
شبكية العين المنقسمة قابلة للازدواج FISH و IHC
من خلال تمديد وقت التثبيت ل IHC القياسي ، يمكن معالجة شبكية العين المنقسمة بالتتابع بواسطة FISH و IHC لتسمية mRNAs والبروتينات في وقت واحد17,18. أكدت التجارب أن تثبيت 2 ساعة في 4٪ بارافورمالدهيد ينتج عنه وضع علامات قوية على mRNA مع الحفاظ على حوامل البروتين لربط الأجسام المضادة. تم إجراء FISH على شبكية العين المنقسمة متبوعة ب IHC لتصور تعبير الوحدة الفرعية لمستقبلات GABAA δ (GABRD ؛ مجسات mRNA المضادة للحس) فيما يتعلق بموضع كرات الدم الحمراء (الجسم المضاد ل PKCα) في INL الخارجي (الشكل 5A). يبدو تعبير GABRD mRNA نادرا في كرات الدم الحمراء (الشكل 5 أ) ؛ ومع ذلك ، يتم التعبير عن النص بكثرة بواسطة خلايا الأماكرين والخلايا العقدية كما يتضح من نمط وضع العلامات على المقاطع المستعرضة من شبكية سليمة (الشكل 5 ب). في INL الخارجي (الشكل 5A) ، يتم توزيع GABRD mRNA بشكل متساو مقارنة ب INL الداخلي (الشكل 5C) حيث يتركز في خلايا متميزة. تنتج مجسات Antisense التي تستهدف الوحدات الفرعية لمستقبلات GABA الأخرى أنماطا مميزة لوضع العلامات ، مما يدل على خصوصية المجسات (البيانات غير معروضة).

الشكل 5: FISH المزدوج وIHC في شبكية منقسمة وشبكية سليمة. (أ) الصور المجهرية متحدة البؤر لشبكية منقسمة مسطحة و(ب) مقطع رأسي من شبكية سليمة. الصور في (A) و (C) هي أقصى إسقاطات للمقاطع البصرية في المناطق العلوية والسفلية من INL على التوالي. تمثل المستطيلات المنقطة في (B) الحدود التقريبية المستخدمة لإنشاء الإسقاطات الموضحة في (A) و (C). تم تثبيت الشبكية المنقسمة (A ، C) لمدة 2 ساعة ، ثم تم تصنيفها بمجسات mRNA المضادة للحساسية ضد GABRD (أحمر). بعد ذلك ، تم تلطيخ شبكية العين المنقسمة بالأجسام المضادة ضد PKCα لتسمية كرات الدم الحمراء (الخضراء). تم حذف قناة PKCα من إسقاطات INL السفلية من أجل الوضوح. تم تثبيت الشبكية السليمة في (B) لمدة 24 ساعة قبل التقسيم. بعد ذلك ، تم تمييز شبكية العين الثابتة بتحقيقات mRNA المضادة للحساسية ضد GABRD (أحمر). تم تلطيخ جميع العينات ب DAPI (أزرق) لمدة 20 ثانية قبل تركيب الغطاء. الاختصارات: INL = الطبقة النووية الداخلية. قضبان المقياس = 10 ميكرومتر. الرجاء الضغط هنا لعرض نسخة أكبر من هذا الشكل.
{kind=link}
شبكية العين المنقسمة مناسبة تماما لتسجيل الفيزيولوجيا الكهربية ذات المشبك التصحيحي من BCs و HCs
لتصحيح سوما BC أو HC في شبكية العين التقليدية الكاملة ، يجب أن تقترب الماصة إما من جانب خلية العقدة أو جانب المستقبل الضوئي. يتطلب كلا النهجين اجتياز عدة طبقات من شبكية العين للوصول إلى INL ، حيث غالبا ما يتم إعاقة طرف الماصة بواسطة الحطام. في إعداد شريحة الاهتزاز ، يمكن الوصول بسهولة إلى BC و HC somas ، ولكن قد يتم قطع عملياتهما المتغصنة ، مما يعطل اتصالاتهما الجانبية. ومع ذلك ، في شبكية العين المنقسمة ، تجلس أجسام الخلايا من كرات الدم الحمراء و HCs على سطح الأنسجة ، مما يوفر وصولا محسنا بشكل كبير إلى ماصات التصحيح مع الحفاظ على الدوائر الجانبية ل OPL.
يوضح الشكل 6 استجابات ضوئية محاكاة كيميائيا مسجلة من BCs في شبكية منقسمة. تم استكمال وسط أميس Perfused ب L-AP4 (4 μM) ، وهو ناهض mGluR من المجموعة الثالثة ، لمحاكاة إطلاق الغلوتامات من المستقبلات الضوئية في الظلام. تم نفخ مضاد mGluR6 ، CPPG (600 ميكرومتر ، في أميس) ، على التشعبات في الخلية المرقعة (مثبتة عند -60 مللي فولت) لمحاكاة وميض ضوئي عن طريق تثبيط mGluR6. استجابت الخلايا لنفث CPPG بنوعين من التيارات الداخلية. يظهر أحد الأنواع تيارا عابرا متبوعا بهضبة (الشكل 6 أ) ، على غرار التيارات المتعارف عليها التي يستحثها الضوء المسجلة من كرات الدم الحمراء في شرائح الشبكية19. يبقى النوع الآخر مستمرا طوال مدة النفخة (الشكل 6B) ، يشبه التيارات المسجلة من الخلايا ثنائية القطب المخروطية ON (ON-CBC) 19.
تم إجراء تجربة منفصلة لاستهداف HCs ، وهو نوع من الخلايا ذات مجال شجيري واسع يصعب الحفاظ عليه غالبا في مستحضرات الشرائح. تم استخدام خط الماوس الذي يعبر عن قناة رودوبسين (ChR2) و GFP في HCs لتسهيل التعرف بسهولة تحت المجهر الفلوري. أولا ، تم تسجيل التيارات من HCs استجابة لسلسلة من خطوات إزالة الاستقطاب (-100 mV إلى 50 mV ، حجم الخطوة = 15 mV) والتي استجابت لها بتيارات داخلية متبوعة بتيارات خارجية (الشكل 6C). ثم تم تحفيز هذه الخلايا بنبضة ضوء أزرق قصيرة (200 مللي ثانية ، 470 نانومتر) تنتج تيارات داخلية كبيرة مدفوعة ب ChR2 في خليتين (الشكل 6D).

الشكل 6: تسجيلات المشبك الرقعي من الخلايا العصبية INL في شبكية العين المنقسمة. (أ) تم تثبيت كرات الدم الحمراء المفترضة و (ب) CBC بالجهد عند -60 mV في وسائط Ames المثقوبة التي تحتوي على L-AP4 (4 ميكرومتر). أدى نفخ CPPG (600 ميكرومتر) على الزوائد الشجيرية للخلايا المثبتة إلى استدعاء تيار داخلي كان عابرا في كرات الدم الحمراء ولكنه استمر في CBC. تسجيل كرات الدم الحمراء في (A) هو تتبع واحد بينما يمثل تسجيل CBC في (B) متوسط 3 تتبعات. (ج) تسجيل مشبك رقعة من HC في vGATFLPo ؛ vGlut2Cre; Ai80d الماوس. يظهر الخط الأحمر مدة نبضة ضوئية تبلغ 200 مللي ثانية و 470 نانومتر تستخدم لاستدعاء التيار الداخلي الكبير عبر ChR2. (د) استجابات التيار المحقون من HC الذي تم تثبيته بالجهد عند -60 mV ، ثم تدرج بين -70 mV و +35 mV في فترات 15 mV وعاد إلى -60 mV. يظهر الجزء الداخلي نفس الآثار في نافذة 6 مللي ثانية تحيط ببداية خطوة الجهد. (ه) صورة مجهرية منعية فلورية لشبكية منقسمة مسطحة تظهر خلايا أفقية تعبر عن GFP في vGATFLPo؛ vGlut2Cre; Ai80d الماوس. شريط المقياس = 20 ميكرومتر. تم جمع بيانات الفيزيولوجيا الكهربية بمعدل أخذ عينات 20 كيلو هرتز وترشيحها باستخدام مرشح بيسيل منخفض الترددات عند 5 كيلو هرتز. ثم تم تصدير البيانات ، وتم إجراء التصور والتحليل في وضع عدم الاتصال باستخدام Python 3. يرجى النقر هنا لعرض نسخة أكبر من هذا الرقم.
{kind=link}
يتيح تقسيم شبكية العين الاستجواب السريع لتشريح INL و OPL
يتكون الغشاء الخارجي المحدد للشبكية (ELM) و ONL من حاجز ~ 90 ميكرومتر ، مما يعيق انتشار الأجسام المضادة في شبكية العين الداخلية ويخلق ظروف تلطيخ مناعي دون المستوى الأمثل20،21،22. لذلك ، تتطلب أهداف وضع العلامات المناعية في OPL أو INL باستخدام شبكية العين المسطحة التقليدية بروتوكولات تلطيخ تستغرق وقتا طويلا والتي تتطلب غالبا حضانات الأجسام المضادة 48-96 ساعة5،6،7،8،20،22.
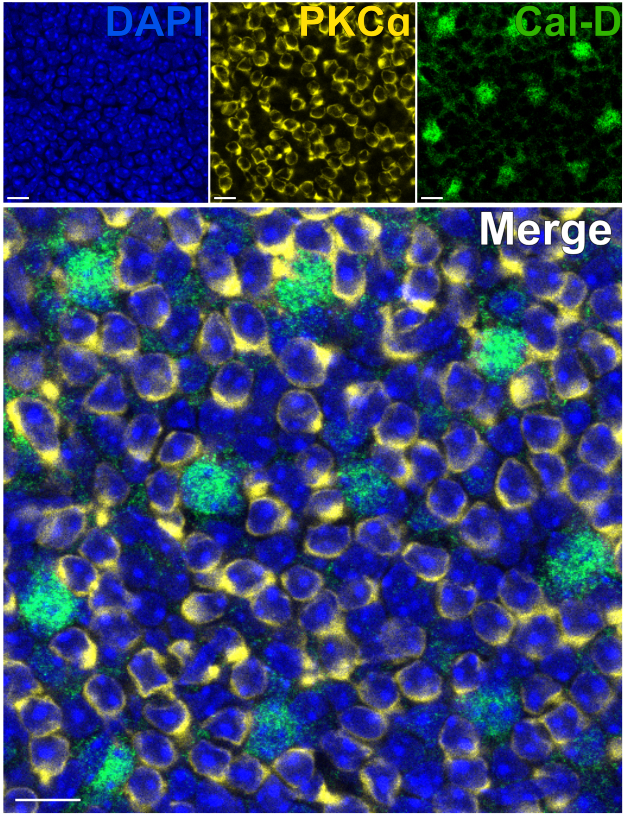
تسمح إزالة المستقبلات الضوئية باختراق سريع للأجسام المضادة للخلايا العصبية الداخلية للشبكية. نتيجة لذلك ، يمكن وضع العلامات على أهداف البروتين داخل الشبكية في أقل من 1 ساعة باستخدام الأجسام المضادة الأولية المترافقة بالصبغة. تم استخدام الأجسام المضادة ضد PKCα و Calbindin-D لتسمية كرات الدم الحمراء و HCs من INL على التوالي (الشكل 7). على عكس أقسام الشبكية الرأسية التقليدية التي تقتطع العمليات الجانبية للخلايا العصبية واسعة المجال ، يتيح تحضير الشبكية المنقسمة تصور الشجرة المتغصنة الكاملة للخلايا واسعة المجال مثل HCs (الشكل 6E ، الشكل 7).

الشكل 7: التسمية المناعية السريعة لبروتينات الشبكية الداخلية في الشبكية المنقسمة. صور التألق المناعي متحد البؤر لشبكية منقسمة من منظور مسطح. تم تحضين الشبكية المنقسمة بأجسام مضادة ضد PKCα (أصفر) و Calbindin-D (أخضر) لمدة 1 ساعة في درجة حرارة الغرفة لتسمية ON-BCs و HCs على التوالي. (أ) كل صورة قناة واحدة هي إسقاط Z متوسط يتكون من أربعة أقسام بصرية: DAPI ، متوسط z10-13 ؛ كالبندين- D ، متوسط z11-14 ؛ PKCα ، متوسط z11-14. (ب) في الصورة المدمجة، يتم تراكب نفس الإسقاطات. قضبان المقياس = 10 ميكرومتر. يرجى النقر هنا لعرض نسخة أكبر من هذا الشكل.
{kind=link}
الشكل التكميلي 1: المراحل الرئيسية لتشريح الشبكية. تم التقاط جميع الصور بكاميرا هاتف ذكي مثبتة على عدسات العين لمجهر التشريح. أ: صورة من أعلى لأسفل لعين فأر بعد إزالة القرنية. ( ب) صورة من أعلى لأسفل لفنجان عين الفأر بعد إزالة العدسة. (ج) يجرى شق صغير في الصلبة على فنجان عين الفأر. تشير الأسهم إلى اثنين من اللوحات من الصلبة التي يتم سحبها في اتجاهين متعاكسين بواسطة ملقط لبدء فصل شبكية العين عن الصلبة. (د) بعد سحب الصلبة جزئيا بعيدا عن الشبكية، يدخل مقص فانا بين الصلبة والشبكية، ويقطع العصب البصري، مما يؤدي إلى تحرير الشبكية. تظهر الدائرة الحمراء المنقطة رأس العصب البصري ، ويوضح المقص مسار القطع الصحيح (أدخل مقصا بين الصلبة والشبكية). يتم التخلص من الشبكية المعزولة بعد الصلبة. الرجاء الضغط هنا لتنزيل هذا الملف.
الشكل التكميلي 2: توصيف المكونات قبل وبعد المشبكي ل OPL في الشبكية المنقسمة. صور التألق المناعي متحد البؤر من OPL في شبكية منقسمة. تم تحضين شبكية العين المنقسمة بأجسام مضادة ضد GPR179 و PSD95 لمدة 1 ساعة في درجة حرارة الغرفة لوضع علامة على الأطراف المتغصنة ل ON-BCs وأطراف المستقبلات الضوئية للقضيب ، على التوالي. الصور اليسرى والوسطى هي أقصى إسقاطات لعدة أقسام بصرية. يتم تثبيت نفس الإسقاطات في الصورة الموجودة في أقصى اليمين. ينظر إلى نقطة GPR179 في الأطراف المتغصنة ON-BC على أنها مرتبطة ارتباطا وثيقا بأطراف المستقبلات الضوئية للقضيب ، مما يدل على اتصالات متشابكة سليمة داخل OPL. قضبان المقياس = 10 ميكرومتر. الرجاء النقر هنا لتنزيل هذا الملف.
الشكل التكميلي 3: استكشاف الأخطاء وإصلاحها: تقييم جودة الشبكية المنقسمة. صور مجهرية فلورية لشبكية منقسمة ملطخة ب DAPI لتصور نوى الخلية. يمكن تحديد الخلايا بناء على قطر النواة وعمق نسيجها. (أ) نوى المستقبلات الضوئية أصغر حجما وأكثر سطوعا وسطحية، في حين أن نوى (ب) قبل الميلاد أكبر وأكثر خفوتا وأعمق. ج: صورة تكبير منخفضة لمنطقة أزيلت فيها مستقبلات ضوئية بشكل غير كامل. النوى التي تظهر في التركيز هي من BCs ، والتي هي أعمق من نوى مستقبلات الضوء على حواف الصورة التي تظهر خارج نطاق التركيز. أشرطة المقياس ل (A) و (B) = 20 ميكرومتر. شريط المقياس ل (C) = 50 ميكرومتر. الرجاء الضغط هنا لتنزيل هذا الملف.
Discussion
بعد أن تقوم المستقبلات الضوئية بتحويل امتصاص الفوتون إلى إطلاق ناقل عصبي ، تعد BCs و HCs أول خلايا عصبية في شبكية العين تعالج الإشارة البصرية23. في حين أن أهمية هذه الخلايا العصبية تحظى بتقدير كبير ، فإن العديد من وظائفها غير مفهومة بشكل كامل أو غير مستكشفة تماما. من المحتمل أن تستفيد العديد من دراسات علم وظائف الأعضاء BC و HC من إعداد شبكية العين المسطح الذي يحسن الوصول إلى الخلايا العصبية INL مع الحفاظ على الاتصال الجانبي. يمثل تطوير طريقة تقسيم الشبكية جهدا لتوفير بروتوكول سهل للحصول على تسجيلات فيزيولوجية كهربية عالية الجودة وبيانات مجهرية من BCs و HCs في اتجاه مسطح. يمكن إجراء تحضير الشبكية المنقسمة الموصوف هنا في حوالي 20 دقيقة لكل فأر (10 دقائق لكل شبكية) بعد عزل الشبكية ، دون استخدام معدات متخصصة. تستمد الطريقة الإلهام من إجراءات إزالة المستقبلات الضوئية الحالية ولكنها تقدم تحسينات كبيرة في البساطة والسرعة والتنوع4،10،11،12،13. على عكس الطرق السابقة لفصل طبقات الشبكية ، لا يتطلب تقسيم الشبكية التجميد أو التجفيد أو التطبيق المتكرر للمواد اللاصقة على شبكية العين. مع الممارسة ، يمكن إزالة جميع المستقبلات الضوئية تقريبا في تمزق واحد باستخدام غشاء النيتروسليلوز. تسمح سرعة وسهولة هذا النهج للمرء بتقليل الوقت الذي تقضيه شبكية العين خارج Ames الكربوجينية ، مما يتيح بقاء الخلايا العالية لفترات طويلة. يمكن الحفاظ على شبكية العين المنقسمة في وسائط أميس الكربوجينية لعدة ساعات بعد الانقسام. كدليل على صحة الخلايا العصبية INL في هذا المستحضر ، تؤكد بقعة الخلايا الحية / الميتة (الشكل 4) والفيزيولوجيا الكهربية لمشبك التصحيح (الشكل 6) صلاحية كرات الدم الحمراء و HCs بعد الانقسام.
إن إزالة طبقة المستقبلات الضوئية في شبكية العين المنقسمة تنقل ميزة كبيرة أثناء وضع العلامات المناعية عن طريق تقليل وقت انتشار الأجسام المضادة في INL بشكل كبير. يمكن إكمال وضع العلامات على الأجسام المضادة الأولية والثانوية في غضون 2 ساعة ، وهو تحسن كبير في تلطيخ التثبيت المسطح التقليدي الذي يمكن أن يستغرق 72 ساعة أو أكثر اعتمادا على الهدف5،6،7،8،20،22. نتيجة لذلك ، يمكن الحصول على بيانات الفحص المجهري في نفس يوم تحضير الأنسجة ، مما يسرع بشكل كبير من وتيرة تجارب التألق المناعي. لتسهيل تلدين مسبار mRNA ، توصي تجارب FISH عادة بأوقات تثبيت أطول بكثير (~ 24 ساعة) من وضع العلامات المناعية18. ومع ذلك ، توضح التجارب المقدمة هنا أن التثبيت لمدة 2 ساعة لا يزال ينتج ملصقات FISH استثنائية (الشكل 5). على الرغم من تمديد وقت التثبيت من 30 دقيقة إلى 2 ساعة ، لم يكن من الضروري إجراء خطوات استرجاع المستضد للحصول على علامات مناعية ممتازة ، ولكن هذا قد يختلف مع الجسم المضاد أو المستضد. قد يتداخل علاج الأنزيم البروتيني في بروتوكول FISH مع وضع العلامات على الأجسام المضادة ، ويرجع ذلك على الأرجح إلى تدمير الحواتم المستهدفة. تم التحايل على هذه المشكلة باستخدام الأجسام المضادة متعددة النسيلة التي تستهدف حواتم متعددة ، مما يقلل من احتمال أن يؤدي تدمير الحواتم إلى إعاقة وضع العلامات المناعية. بالإضافة إلى ذلك ، تم استخدام علاج البروتياز المعتدل (ACD protease III) لمنع التغيير المفرط في الحاتم مع توفير اختراق كاف للأنسجة.
في بعض الأحيان ، تنقسم شبكية العين بدلا من ذلك عبر الطبقة النووية الخارجية (ONL) ، تاركة وراءها طبقات من الأورام المستقبلة للضوء مع عدم وجود خلايا INL مرئية. لمنع ذلك ، يجب على المرء التأكد من أن شبكية العين مسطحة تماما على الزجاج ، وأن أي سائل متبقي من حول الشبكية قد تمت إزالته. قد يساعد الضغط بقوة أكبر على النيتروسليلوز باستخدام فرشاة الرسم أيضا في منع الانقسام من خلال ONL. إذا أصبح الغشاء رطبا جدا أو تم طي الشبكية على نفسها ، فإن فرص الانقسام الناجح ستتضاءل بشكل كبير. يعد استخدام DAPI لتلطيخ نوى الخلايا مفيدا لتقييم جودة الانقسام وتحديد تغطية المستقبلات الضوئية المتبقية. تكون نوى المستقبلات الضوئية أصغر حجما وأكثر إشراقا وسطحية (الشكل التكميلي 3 أ) ، في حين أن نوى BC أكبر وأكثر خفوتا وأعمق (الشكل التكميلي 3 ب). في بعض الحالات ، يختلف مستوى التمزق قليلا عبر قطعة الشبكية ، مما يؤدي إلى ظهور بقع حيث لم تتم إزالة أجسام خلايا المستقبلات الضوئية بالكامل (الشكل التكميلي 3C). بالنسبة للتطبيقات في الفحص المجهري والفيزيولوجيا الكهربية ، فإن هذا لا يعيق القدرة على جمع بيانات عالية الجودة من المناطق التي تمت فيها إزالة المستقبلات الضوئية بشكل صحيح ؛ يمكن بسهولة العثور على مجالات كبيرة من شبكية العين الداخلية المكشوفة عند التصوير أو التسجيل باستخدام ماصة رقعة. إذا كانت هناك رغبة في إزالة مستقبلات ضوئية أكثر اكتمالا ، فيمكن إجراء تمزق ثان بقطعة إضافية من غشاء النيتروسليلوز ، على الرغم من أن إزالة المستقبلات الضوئية بنسبة 100٪ غير مضمونة. لذلك ينصح بالحذر عند استخدام شبكية العين المنقسمة في دراسات التعبير الجيني أو البروتينات حيث يمكن أن تؤثر المواد المستقبلة للضوء المتبقية على النتائج. بالنسبة لتطبيقات الخلية الواحدة ، لا مبرر لهذا القلق ، حيث يمكن استبعاد البيانات من المستقبلات الضوئية من التحليل.
ربما تكون مزايا تحضير الشبكية المنقسمة أكثر بروزا في التسجيلات الفيزيولوجية الكهربية للخلايا العصبية البينية واسعة المجال. في حين أن الشرائح الرأسية التقليدية تقطع العمليات الواسعة للخلايا واسعة المجال ، فإن تحضير شبكية العين المنقسمة يترك OPL و IPL سليمين ، مما يسمح للمرء بالتقاط المدخلات من الخلايا واسعة المجال مثل HCs 24 و A17s25 و TH ACs 26 و NOS-1 ACs27 التي يمكن تجاهلها في شرائح عمودية. لذلك ، يتطلب تفسير النتائج والمقارنة مع البيانات السابقة التي تم جمعها من شرائح الشبكية تفكيرا دقيقا. ومع ذلك ، في التجارب التي تستخدم المحاكاة الدوائية لتحفيز الضوء ، تشبه هذه النتائج البيانات المسجلة من شرائح الشبكية19. من خلال التعبير عن ChR2 تحت المروجين الخاصين بالخلية ، يمكن للمرء تحفيز عدد الخلايا المرغوب فيه أثناء التسجيل من BCs في INL للتحقيق في تأثير الخلية المطلوبة على مسار المعلومات العمودي. التسجيل مباشرة من الخلايا العصبية INL العميقة ، مثل خلايا amacrine ، ممكن أيضا في شبكية العين المنقسمة. بينما في هذه الحالة يجب أن ينتقل قطب التصحيح أولا عبر الخلايا العصبية INL الأكثر سطحية ، إلا أن هناك أنسجة أقل بكثير تعيق مسارها عند مقارنتها بإعداد التثبيت الكامل التقليدي.
بالإضافة إلى قياس تأثير الخلايا واسعة المجال على الخلايا العصبية الأخرى ، تتيح هذه الطريقة التثبيت المباشر لرقعة الخلية الواحدة من HCs ، التي تشكل الزوائد الشجيرية شبكة مقترنة واسعة النطاق في OPL28. ترسل الخلايا الأفقية ردود فعل مهمة إلى المستقبلات الضوئية التي تشكل نقل المعلومات الرأسية عبر شبكية العين. ومع ذلك ، نظرا لأن الحقول المتغصنة ل HCs يتم اقتطاعها في شرائح رأسية ، فإن بيانات تسجيل الخلية المفردة غير متوفرة. يقدم هذا العمل HCs سليمة تشريحيا وفسيولوجيا يتم تسجيل التيارات المستثارة ChR2 منها في خط فأر معدل وراثيا ثلاثي (الشكل 6 C-E). خارج تحفيز ChR2 ، يمكن استخدام الشبكية المنقسمة لدراسة تيارات HC الداخلية واقتران تقاطع الفجوة28. بينما توفر شبكية العين المنقسمة نموذجا مناسبا لدراسة الاتصال المشبكي والنشاط العصبي الناجم عن التطبيق الكيميائي أو تحفيز ChR2 ، فإن نقص المستقبلات الضوئية يحول دون أي استكشاف مباشر لاستجابات الضوء الطبيعي أو آليات التكيف مع الضوء.
حقق التصوير في الموقع في شبكية العين تقدما مثيرا للإعجاب في السنوات الأخيرة. ومع ذلك ، فإن غالبية دراسات التصوير تقتصر على طبقة الخلايا العقدية في مستحضرات شبكية العين الكاملة29. يتصور المؤلفون أن غياب المستقبلات الضوئية في شبكية العين المنقسمة سيجعلها نموذجا مثاليا لتصوير الكالسيوم الحي في OPL و INL. بالإضافة إلى تصوير الكالسيوم ، يتمتع هذا النموذج بإمكانية كبيرة للاستخدام مع أجهزة الاستشعار الحيوية المشفرة وراثيا مثل iGluSnFR 30,31 و iGABASnFR32 و pHluorin 33. إلى جانب تحضير الشبكية المنقسمة ، قد توفر هذه الأدوات القوية نهجا فعالا لاستكشاف التفاعلات المشبكية والخصائص الفيزيائية الحيوية ل BCs و HCs التي تساهم في معالجة الضوء في شبكية العين.
Disclosures
يعلن أصحاب البلاغ عدم وجود مصالح مالية متنافسة.
Acknowledgements
تم دعم هذا العمل من خلال منح المعاهد الوطنية للصحة التالية: منحة المعاهد الوطنية للصحة R01EY031596 (إلى C.M.) ؛ منحة المعاهد الوطنية للصحة R01EY029985 (إلى C.M.) ؛ منحة المعاهد الوطنية للصحة P30EY010572 (إلى C.M.) ؛ منحة المعاهد الوطنية للصحة R01EY032564 (إلى BS). نشكر تامي هالي على دعمها الفني في إعداد أقسام شبكية العين ، والدكتور تشارلز ألين لمساهمته السخية في مجسات mRNA FISH المستخدمة في هذا العمل.
Materials
| Name | Company | Catalog Number | Comments |
| #1.5 glass coverslips | Fisherbrand | 12544E | |
| 2 pairs of Dumont #5 forceps | Ted Pella | 38125 | |
| 25 gauge needle | Becton Dickenson | 305122 | |
| 470 nm LED | THORLABS | M470L2 | |
| 5-306 curved scissors | Miltex | 5-306 | |
| 9" disposable pasteur pipetes | Fisherbrand | 13-678-20D | for constructing custom transfer pipette |
| Ai80d mouse | Jackson Laboratories | 25109 | RRID: IMSR_JAX:025109 |
| Ames Medium w/L-Glutamate | US Biological | A1372-25 | |
| amplifier control software | Molecular Devices | Clampex 10.3 software | |
| anti-calbindin D28K antibody | Invitrogen | PA-5 85669 | RRID: AB_2792808, host species = rabbit; 1:100 dilution |
| anti-CtBP2 antibody | BD Biosciences | 612044 | RRID: AB_399431, host species = mouse; 1:5000 dilution |
| anti-GPR179 antibody | NA | NA | gift from Kirill Martemyanov; Scripps Research Institute, Jupiter, FL; host species = sheep; 1:1000 dilution |
| anti-PKC alpha antibody | Sigma-Aldrich | P4334 | RRID: AB_477345, host species = rabbit; 1:5000 dilution |
| anti-PKC alpha antibody | Santa Cruz Biotechnology | sc8393 AF594 | RRID: AB_628142, host species = mouse; 1:1000 dilution |
| anti-PSD95 antibody | BD Transduction Laboratories | 610495 | RRID: AB_397862, host species = mouse; 1:1000 dilution |
| anti-RGS11 antibody | NA | NA | gift from Ted Wensel; Baylor College of Medicine, Houston, TX; host species = rabbit; between 1:1000 and 1:5000 dilution |
| anti-Synaptophysin P38 antibody | Sigma | S-S5768 | RRID: AB_477523, host species = mouse; 1:1000 dilution |
| Aquamount mounting media | Epredia | 13800 | slide mounting media |
| C57BL/6J mouse | Jackson Laboratories | 000664 | RRID: IMSR_JAX:000664 |
| carbogen tank | Matheson | NA | 95% O2 and 5% CO2 |
| custom transfer pipette | custom build | NA | Instructions: use scissors to cut off the tip of a plasitc transfer pipette at the point it begins to taper. Use pliers to safely break off the last 2-3 inches of a glass pasteur pipette. Fit the narrow end of the glass pasteur pipette into the wide tip of the plastic transfer pipete. Wrap parafilm around the joint of the two pieces to enhance the seal. |
| Digitical optical power meter | THORLABS | PM100D | |
| dissection microscope | Zeiss | Stemi 2000 | |
| electrophysiology amplifier | Molecular Devices | Axopatch 200B | |
| electrophysiology microscope | Olympus | OLYMPUS, BX50WI | Dodt gradient contrast microscopy |
| Fluoromount-G | SouthernBiotech | 0100-01 | |
| HC PL APO CS2 40x/1.3 | Leica | 506358 | |
| HC PL APO CS2 63x/1.40 | Leica | 15506350 | |
| Hybridization oven | Robbins Scientific | Model 1000 | for RNAscope protocol only |
| Immedge hydrophobic barrier pen | Vector Laboratories | H-4000 | |
| isoflurane | Piramal Critical Care | 66794-017-25 | |
| Kimwipe (delicate task wipe) | Kimtech Science | 34155 | |
| Leica HC PL APO CS2 40x/1.3 oil immersion objective | Leica | 506358 | |
| Leica HC PL APO CS2 63x/1.40 oil immersion objective | Leica | 15506350 | |
| Leica TCS SP8 X confocal microscope | Leica | discontinued | |
| medium 15 mm petri dish | Corning | 25060-60 | eyes are kept here during retina dissection |
| Merit 97-275 steel scissors | Merit | 97-275 | |
| Micropipette Puller | Sutter Instrument | p-97 | |
| Mm-Gabrd-C2 mRNA probe | ACD | 459481-C2 | |
| mouse euthanasia chamber | NA | NA | custom build; glass petri dish covering a small glass jar. |
| nitrocellulose membrane filters | GE Healthcare Life Sciences; Whatman | 7184-005 | 0.45 µm pore size |
| Picospritzer | General Valve Corporation | Picospritzer II | referred to in the text as microcellular injection unit |
| plastic transfer pipets | Fisherbrand | 13-711-7M | for constructing custom transfer pipette |
| Plastic tubing | Tygon | R-603 | for connection to carbogen tank |
| platinum harp | custom build | NA | for anchoring split retinas within the electrophysiology recording chamber. |
| size 0 paint brush | generic | NA | for flattening retina during splitting. |
| SlowFade Gold antifade reagent | Molecular Probes | S36937 | referred to in the text as anti-fade mounting media |
| small 10 mm petri dish | Falcon | 353001 | eyes are placed here following enucleation |
| small glass pane (7.5 cm x 5 cm) | generic | NA | isolatd retina pieces are placed onto this for the splitting procedure |
| Superfrost plus microscope slides | Fisherbrand | 12-550-15 | electrostatically-charged glass microscope slides |
| Thick-walled borosilicate glass pipettes with filament | Sutter Instrument | BF150-86-10HP | |
| Vannas Scissors; straight | Titan Medical | TMS121 | not brand specific; any comparable scissors will work |
| vGATFLPo mouse | Jackson Laboratories | 29591 | RRID: IMSR_JAX:029591 |
| vGlut2Cre mouse | Jackson Laboratories | 28863, 016963 | RRID: IMSR_JAX:028863, RRID: IMSR_JAX:016963 |
| Zombie NIR Fixable Viability Kit | BioLegend | 423105 | referred to in the text as MI-NIR |
References
- Morgans, C. W. Neurotransmitter release at ribbon synapses in the retina. Immunology & Cell Biology. 78 (4), 442-446 (2000).
- Euler, T., Haverkamp, S., Schubert, T., Baden, T. Retinal bipolar cells: elementary building blocks of vision. Nature Reviews Neuroscience. 15 (8), 507-519 (2014).
- Barnes, S., Grove, J. C. R., McHugh, C. F., Hirano, A. A., Brecha, N. C. Horizontal Cell Feedback to Cone Photoreceptors in Mammalian Retina: Novel Insights From the GABA-pH Hybrid Model. Frontiers in Cellular Neuroscience. 14, (2020).
- Walston, S. T., Chang, Y. C., Weiland, J. D., Chow, R. H. Method to remove photoreceptors from whole mount retina in vitro. Journal of Neurophysiology. 118 (5), 2763-2769 (2017).
- Stefanov, A., Novelli, E., Strettoi, E. Inner retinal preservation in the photoinducible I307N rhodopsin mutant mouse, a model of autosomal dominant retinitis pigmentosa. Journal of Comparative Neurology. 528 (9), 1502-1522 (2020).
- Matsuoka, R. L., Nguyen-Ba-Charvet, K. T., Parray, A., Badea, T. C., Chédotal, A., Kolodkin, A. L. Transmembrane semaphorin signaling controls laminar stratification in the mammalian retina. Nature. 470 (7333), 259-263 (2011).
- Matsuoka, R. L., et al. Guidance-Cue Control of Horizontal Cell Morphology, Lamination, and Synapse Formation in the Mammalian Outer Retina. Journal of Neuroscience. 32 (20), 6859-6868 (2012).
- Wässle, H., Puller, C., Müller, F., Haverkamp, S. Cone Contacts, Mosaics, and Territories of Bipolar Cells in the Mouse Retina. Journal of Neuroscience. 29 (1), 106-117 (2009).
- Thoreson, W. B., Dacey, D. M. Diverse Cell Types, Circuits, and Mechanisms for Color Vision in the Vertebrate Retina. Physiological Reviews. 99 (3), 1527-1573 (2019).
- Guido, M. E., et al. A simple method to obtain retinal cell preparations highly enriched in specific cell types. Suitability for lipid metabolism studies. Brain Research Protocols. 4 (2), 147-155 (1999).
- Rose, K., Walston, S. T., Chen, J. Separation of photoreceptor cell compartments in mouse retina for protein analysis. Molecular Neurodegeneration. 12 (1), 28(2017).
- Todorova, V., et al. Retinal Layer Separation (ReLayS) method enables the molecular analysis of photoreceptor segments and cell bodies, as well as the inner retina. Scientific Reports. 12 (1), 20195(2022).
- Shiosaka, S., Kiyama, H., Tohyama, M. A simple method for the separation of retinal sublayers from the entire retina with special reference to application for cell culture. Journal of Neuroscience Methods. 10 (3), 229-235 (1984).
- Ivanova, E., Hwang, G. S., Pan, Z. H. Characterization of transgenic mouse lines expressing Cre-recombinase in the retina. Neuroscience. 165 (1), 233-243 (2010).
- Sarria, I., Orlandi, C., McCall, M. A., Gregg, R. G., Martemyanov, K. A. Intermolecular Interaction between Anchoring Subunits Specify Subcellular Targeting and Function of RGS Proteins in Retina ON-Bipolar Neurons. The Journal of Neuroscience. 36 (10), 2915-2925 (2016).
- Orlandi, C., Cao, Y., Martemyanov, K. A. Orphan Receptor GPR179 Forms Macromolecular Complexes With Components of Metabotropic Signaling Cascade in Retina ON-Bipolar Neurons. Investigative Ophthalmology & Visual Science. 54 (10), 7153-7161 (2013).
- Dikshit, A., Zong, H., Anderson, C., Zhang, B., Ma, X. -J. Simultaneous Visualization of RNA and Protein Expression in Tissue Using a Combined RNAscopeTM In Situ Hybridization and Immunofluorescence Protocol. Methods in Molecular Biology. 2148, Clifton, N.J. 301-312 (2020).
- Wang, F., et al. RNAscope. The Journal of Molecular Diagnostics. 14 (1), 22-29 (2012).
- Morgans, C. W., et al. TRPM1 is required for the depolarizing light response in retinal ON-bipolar cells. Proceedings of the National Academy of Sciences of the United States of America. 106 (45), 19174-19178 (2009).
- Alessio, E., Zhang, D. Q. Immunostaining of whole-mount retinas with the CLARITY tissue clearing method. Investigative Ophthalmology & Visual Science. 61 (7), 5054(2020).
- Ferguson, L. R., Dominguez, J. M., Balaiya, S., Grover, S., Chalam, K. V. Retinal Thickness Normative Data in Wild-Type Mice Using Customized Miniature SD-OCT. PLoS ONE. 8 (6), e67265(2013).
- Ivanova, E., Toychiev, A. H., Yee, C. W., Sagdullaev, B. T. Optimized Protocol for Retinal Wholemount Preparation for Imaging and Immunohistochemistry. Journal of Visualized Experiments JoVE. (82), e51018(2013).
- Kolb, H. Neurotransmitters in the Retina. Webvision: The Organization of the Retina and Visual System. , University of Utah Health Sciences Center. (1995).
- Chaya, T., et al. Versatile functional roles of horizontal cells in the retinal circuit. Scientific Reports. 7 (1), 5540(2017).
- Egger, V., Diamond, J. S. A17 Amacrine Cells and Olfactory Granule Cells: Parallel Processors of Early Sensory Information. Frontiers in Cellular Neuroscience. 14, 600537(2020).
- Dacey, D. M. The dopaminergic amacrine cell. The Journal of Comparative Neurology. 301 (3), 461-489 (1990).
- Park, S. J., et al. Connectomic analysis reveals an interneuron with an integral role in the retinal circuit for night vision. eLife. 9, 56077(2020).
- Janssen-Bienhold, U., et al. Connexin57 is expressed in dendro-dendritic and axo-axonal gap junctions of mouse horizontal cells and its distribution is modulated by light. The Journal of Comparative Neurology. 513 (4), 363-374 (2009).
- Jain, V., et al. The functional organization of excitation and inhibition in the dendrites of mouse direction-selective ganglion cells. eLife. 9, 52949(2020).
- Marvin, J. S., et al. Stability, affinity, and chromatic variants of the glutamate sensor iGluSnFR. Nature Methods. 15 (11), 936-939 (2018).
- Strauss, S., et al. Center-surround interactions underlie bipolar cell motion sensitivity in the mouse retina. Nature Communications. 13 (1), 5574(2022).
- Marvin, J. S., et al. A genetically encoded fluorescent sensor for in vivo imaging of GABA. Nature Methods. 16 (8), 763-770 (2019).
- Beckwith-Cohen, B., Holzhausen, L. C., Wang, T. M., Rajappa, R., Kramer, R. H. Localizing Proton-Mediated Inhibitory Feedback at the Retinal Horizontal Cell-Cone Synapse with Genetically-Encoded pH Probes. The Journal of Neuroscience: The Official Journal of the Society for Neuroscience. 39 (4), 651-662 (2019).
Reprints and Permissions
Request permission to reuse the text or figures of this JoVE article
Request PermissionExplore More Articles
This article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. All rights reserved