
Method Article
脊椎動物の網膜の内核層ニューロンを研究するための改良されたフラットマウント調製としてのスプリット網膜
要約
この研究は、光受容体細胞体を除去することで、抗体の拡散が速くなり、免疫組織化学、 in situ ハイブリダイゼーション、および電気生理学実験のための網膜内ニューロンへのパッチピペットアクセスが改善される、代替のフラットマウント網膜製剤を提示します。
要約
脊椎動物の網膜の双極細胞と水平細胞は、光子が光受容体によって検出された後、視覚情報を処理する最初のニューロンです。これらは、光順応、コントラスト感度、空間および色のオポネンシーなどの基本的な操作を実行します。その行動を支配する精密な回路と生化学的メカニズムを完全に理解することは、視覚神経科学の研究と眼医学を前進させるでしょう。しかし、双極細胞や水平細胞(網膜全体マウントや垂直スライス)を検査するための現在の調製物は、これらの細胞の解剖学的構造や生理学的特徴を捉える能力が限られています。この研究では、生きたフラットマウントマウス網膜から光受容体細胞体を除去する方法を提示し、効率的なパッチクランプと迅速な免疫標識のために、バイポーラ細胞および水平細胞へのアクセスを強化します。裂けた網膜は、単離されたマウス網膜を2枚のニトロセルロースで挟み込み、そっと剥がすことによって作製されます。分離により、外側の網状層のすぐ上で網膜が分割され、2つのニトロセルロースが得られ、1つは視細胞体を含み、もう1つは残りの内側の網膜を含みます。垂直網膜スライスとは異なり、スプリット網膜調製物は網膜内ニューロンの樹状突起を切断しないため、ギャップ結合結合ネットワークと広視野アマクリン細胞の寄与を統合する双極細胞および水平細胞からの記録が可能になります。この研究は、電気生理学、免疫組織化学、および in situ ハイブリダイゼーション実験における水平細胞および双極細胞の研究のためのこの調製物の汎用性を実証しています。
概要
網膜は、後眼にある薄い神経組織で、光が遮断され、脳が解釈できる電気化学信号に処理されます。網膜の後ろでは、桿体と錐体の光受容体が光によって刺激され、神経伝達物質であるグルタミン酸1の強直性放出速度が低下します。この光によるグルタミン酸濃度の変化を経験して反応する最初のニューロンは、双極細胞(BC)と水平細胞(HC)であり、その体細胞は内核層(INL)の最外層に存在します。これらの2次ニューロンは、網膜で信号処理の第1段階を実行し、光順応、コントラスト感度、空間/色のopponencyなどの視覚の重要な特徴を形成します2。これらの機能はBCとHCに起因していると考えられてきましたが、これらのプロセスの根底にある回路や生化学的メカニズムは完全には解明されていません3。したがって、BCおよびHC生理学を探索するためのツールと方法の進歩が最も重要です。
垂直(横)網膜切片は、BCおよびHCを研究するための最も実用的なモデルであると長い間証明されてきました。ただし、BCおよびHC生理学の特定の側面は、このモデルでは実験者がアクセスできません。HCからの直接記録またはBCへの影響の間接的な測定は、これらの細胞の側方突起がスライス中に切断されるため、網膜の内因性接続を反映していません。網膜全体の製剤は、これらの側方突起を保存することでこの問題を回避しますが、周囲の網膜層はこれらの細胞にアクセスするための課題を提起します4。網膜全体のINLニューロンからの免疫染色5,6,7,8およびパッチクランプ記録9の例は豊富にありますが、これらのデータの収集を迅速化および簡素化する機会があります。したがって、横断面の固有の制限とマウントモデル全体の課題は、この代替のフラットマウント網膜準備の開発に影響を与えました。
以下の研究では、生きているフラットマウント網膜から光受容体層を簡単に除去して、BCおよびHCへのアクセスを強化し、パッチクランプを簡素化し、より迅速で効率的な免疫標識を行うためのプロトコルについて説明します。孤立した網膜の両側に取り付けられた2枚のニトロセルロース膜を剥がすと、視細胞の軸索を介して組織が引き裂かれ、外側の網状層(OPL)とすべての内側の網膜層を保持する裂けた網膜が残ります。網膜の層を機械的に分離するためのプロトコルを説明している人もいますが、これらの方法はパッチクランプや顕微鏡検査の用途に適していないか、組織の面倒な操作が必要です。これらの方法のいくつかは、層分離のために凍結または凍結乾燥された組織を必要とするため、電気生理学実験とは互換性がありません10、11、12。その他は生体組織用に設計されているが、光受容体を除去するために濾紙4,11で5〜15回連続して剥離するか、トリプシン13で処理する必要がある。ここで説明する技術は、光受容体の除去手順を簡素化し、下流のアプリケーションのレパートリーを拡大することにより、以前の技術を改善します。
プロトコル
マウスに水と餌を自由に与え、12時間の明暗サイクルで維持しました。マウスは、イソフルランへの曝露とそれに続く子宮頸部脱臼によって安楽死させられました。すべての動物処置は、米国国立衛生研究所のガイドラインに従っており、オレゴン健康科学大学の動物実験委員会によって承認されました。
注:眼球摘出術、網膜解剖、網膜分割は、生体組織の健康を維持するためにできるだけ早く行う必要があります。片目4分<で解剖を完了することを目指します。これら 3 つのステップは順番に実行されます。野生型マウス:成体(>3ヶ月)の雌雄のC57BL/6Jマウスを実験に用いた。シナプス形態については、Pcp2プロモーター(Pcp2-cre/GFP)14 で緑色蛍光タンパク質(GFP)を発現するマウスを用いた。トランスジェニックマウス:免疫組織化学または電気生理学実験中のGFPによる水平細胞の可視化には、トリプルトランスジェニックマウスvGATFLPoを使用しました。vGlut2Cre;Ai80dです。vGATFlpo株およびvGluT2Cre株は、それぞれのプロモーターの下流にFlpoまたはCreリコンビナーゼを発現するノックインマウスです。Ai80dマウスは交差レポーターマウス(CatCh/EYFP)であり、CreおよびFlpoリコンビナーゼを発現する細胞においてのみCa2+ 透過性チャネルロドプシン(ChR2)を発現します。したがって、トリプルトランスジェニックマウスは、VGATとvGluT2の両方の発現歴を持つ細胞でのみChR2を発現します。
1.網膜解離および網膜裂片のための材料調製
- ニトロセルロースメンブレンのピースを準備します
注:分割網膜をニトロセルロースメンブレンから剥離すると、顕微鏡でのバックグラウンド蛍光が減少し、パッチクランプによる記録が簡素化されます。膜除去は、組織固定の前または後に行うことができます。固定された分割網膜の場合、ニトロセルロース膜の断片を処理する必要はありません。生きている裂けた網膜の場合、組織からの穏やかな剥離を容易にするために、ステップ1.1.3〜1.1.5に従って膜を処理します。- 16個(またはそれ以上)のニトロセルロースメンブレンを5mm x 5mmの正方形にカットします。余分なものはまとめて準備し、将来の使用のために保管することができます。
- 後で使用するために、メンブレンの半分を取っておきます。これらの破片はブロッキング溶液で処理されません。
- 残りの断片を界面活性剤を含まないIHCブロッキング溶液(PBSで希釈した3%ウマ血清+0.025%NaN3 など)で室温で10分間、穏やかに振とうしながらインキュベートします。
注意: NaN3 は強力な毒素であるため、取り扱いには適切な PPE を使用してください。 - メンブレン片を、重炭酸緩衝エーム培地中で室温で10分間、穏やかに振とうしてインキュベートすることにより、完全に洗浄します。
- 閉塞したメンブレン片を完全に風乾します(~20分)。メンブレン片にラベルを貼り、室温で保管し、未処理のメンブレン片から分離してください。
- エイムズ培地の準備
- 重炭酸緩衝エーム培地を調製し、一定のカルボゲン化(95%O2および5%CO2)下で溶液を室温に維持します。
2.マウス眼球摘出術
- 機関のIACUCガイドラインに従って、利用可能な方法でマウスを安楽死させます。
- マウスを片側にひっくり返し、2本の指で眼窩の周りをそっと押し下げます。これにより、目が頭蓋骨から膨らみます。
- 湾曲した解剖ハサミを使用して、膨らんだ目の下を切り取り、視神経を切断し、目を頭蓋骨から分離します。
- ハサミで目をすくい、氷冷したエイムズ培地で満たされたペトリ皿に入れます。
注:分裂後に組織を固定するダウンストリームアプリケーションでは、Ames培地の代わりに氷冷PBSを使用できます。 - 残りの目について、手順2.1〜2.4を繰り返します。
3.網膜解剖
- カスタムガラストランスファーピペットを使用して、片方の目を新鮮で氷冷したエイム培地が入った新しいペトリ皿に移します。
注:カスタムトランスファーピペットの開口部が広いため、組織の偶発的な押しつぶしを防ぎ、ガラスを使用することでピペットの壁への組織の付着を最小限に抑えます。ただし、広口プラスチックトランスファーピペットは、実験者がすでにこのツールの使用に習熟している場合にも許容されます。 - 鉗子を使用して、余分な結合組織をペトリ皿の底に固定することにより、目を安定させます。次に、25Gの針を使用して オラセラータ ラインに沿って目を穿刺し、バンナハサミのエントリーポイントを作成します。
- バンナハサミを使用して、角膜が目の残りの部分から解放されるまで、オラセラータラインに沿ってカットします(補足図1A)。鉗子を使用してアイカップからレンズを取り外します(補足図1B)。
- カスタムガラスピペットを使用して、アイカップを大容量(≥100 mL)のカーボゲン化エイムに移し、残りの目で手順3.1〜3.3を繰り返します。
注:アイカップは、解剖がもう一方の眼で行われる間、組織の健康を維持するためにカルボゲン化されたエイムに入れられます。 - 1つのアイカップを、新たにカルボゲン化したエイムで満たされたペトリ皿に移します。
- バンナハサミを使用して、強膜の端から内側に小さな切り込みを入れ、次に2対の鉗子を使用して強膜を網膜から剥がします(補足図1C)。鉗子で網膜をつかまないでください。代わりに、はさみ切り取りによって作成された強膜のフラップを引き離します。
- バンナハサミを使用して強膜と網膜をつなぐ視神経を切り取り(補足図1D)、ハサミまたは鉗子を使用して網膜を強膜からそっとこじ開けて網膜を分離します。(図1A)。
注意: RPEは通常、アイカップに取り付けられたままになりますが、網膜に取り付けられている場合、RPEを取り外すために追加の手順は必要ありません。この時点で、網膜の縁は、平坦化工程中のカールを防ぐためにメスで任意にトリミングされてもよい(図1B)。 - メスを使用して網膜を半分または4分の1に切断し(図1C)、カスタムトランスファーピペットを使用して、断片を大容量(≥100 mL)の連続カーボゲン化Ames培地に戻します。
注:半分または四分の一の選択は主観的です。目的のアプリケーションに最適なオプションを選択してください。 - 網膜の分割に進む前に、残りの目に対して手順3.5〜3.8を繰り返します。
4.網膜の分裂
- ペトリ皿からエイムズ培地を廃棄し、新たにカルボゲン化したエイムと交換します。
注:網膜分裂手順の残りの期間にわたってカルボゲン化を維持するには、ペトリ皿内の培地を約5分ごとに新たにカルボゲン化したエイムと交換します。 - カスタムトランスファーピペットを使用して、神経節細胞側を上にしてスライドガラス(7.5 cm x 5 cm)に網膜片を置き、デリケートなタスクワイプで周囲の液体を取り除き、平らにします(図1D)。必要に応じて、解剖顕微鏡の下で細い先端の絵筆で網膜の端をそっと引っ張ります。
- 鉗子を使用して、乾燥した5 mm x 5 mmのニトロセルロース膜を網膜に下げ、神経節細胞側に接着させます(図1E)。
注:生きた組織からの膜除去が必要な場合(つまり、電気生理学の場合)、このステップには血清処理された膜の乾燥片を使用します(詳細については、手順1.1.3〜1.1.5を参照してください)。これにより、神経節細胞層への接着強度が低下し、分裂後のニトロセルロースから網膜を除去しやすくなります。 - ニトロセルロースがスライドガラスの上に載るように網膜を裏返し、5 mm x 5 mmの乾いた膜片を網膜の感光体側に置きます(図1F)。
- 濡れた絵筆の先端を2つの膜の間のスペースに触れ、毛細管現象を起こしてエイムをサンドイッチに吸い込みます(図1G)。これにより、網膜への膜の付着が減少し、網膜がデリケートなタスクワイプで過度に乾燥した場合にのみ必要です。
注意: 網膜が光沢を失った場合は、過度に乾燥しているため、手順4.5が必要です。 - 均一な接着性を確保するには、濡れた絵筆で上膜に軽い下向きの圧力をかけます(図1H)。
- 1組の鉗子で下膜をガラスに固定しながら、ゆっくりと安定した動きで、2組目の鉗子で上膜をそっと剥がします。これにより、網膜がOPLのすぐ上で裂けます(図1I)。
- 光受容体を含む上膜を廃棄します(図1J、左)。下膜には内側の網膜があり、以下、裂網膜と呼ぶ(図1J、右)。
- 直ちに裂けた網膜をカーボゲン化エーム培地に戻す。
注:生体組織での実験では、網膜は分割後のカルボゲン化エイムスで15〜30分の回復期間の恩恵を受ける可能性があります。

図1:網膜の分割手術 。 (A)冷たいPBSまたはAmes培地での眼球摘出および眼杯調製後、マウス網膜を眼杯から分離し、PBSを室温の炭酸添加Ames培地と交換します。(B)メスを使用して、内側にカールしている領域がなくなるまで網膜の端を切り取ります(オプション)。(C)メスを使って網膜を四分の一または半分に切ります。(D)カスタムトランスファーピペットを使用してスライドガラス(神経節細胞側を上にして)に網膜を1枚置き、デリケートなタスクワイプを使用して余分なエイムをすべて取り除きます。次のステップに進む前に、セミドライ網膜がガラスの上に平らに横たわっていることを確認してください。エイムズで濡らした絵筆の先端を使用して、網膜の平らでない領域をそっと広げます。(E)鉗子を使用して、事前にカットした乾燥ニトロセルロース膜(5 mm x 5 mm)を平らな網膜に置きます。(F)網膜の視細胞側が上を向くようにニトロセルロース片を裏返します。次に、別の乾いた膜を網膜に置きます。(G)ブラシの濡れた先端を2つの膜の間のスペースに触れ、毛細管現象を起こしてエイムズをサンドイッチに吸い込みます。これにより、網膜への膜の付着が減少し、網膜がデリケートなタスクワイプで過度に乾燥した場合にのみ必要です。(H)濡れた絵筆の先端を使用して、挟んだ網膜の中心を下向きにそっと押します。(I)1組の鉗子を使用して下部の膜をスライドガラスに固定し、もう1組の鉗子を使用して上部の膜を下部の膜からそっと剥がします。(J)内側の網膜(左)は下の膜に残り、視細胞(右)は上の膜で引き離されます。パネル(A)、(B)、(C)、(D)、および(J)は、解剖顕微鏡を使用して取得しました。スケールバーは約 1 mm を表します。パネル(E-I)は、拡大せずにスマートフォンのカメラで取得しました。 この図の拡大版をご覧になるには、ここをクリックしてください。
{kind=link}
5. 免疫蛍光実験のための割れ目網膜の作製
注:分割網膜は、ステップ5.5までニトロセルロースメンブレンに付着したままです。ステップ 5.1、5.2、5.3、5.4 のいずれかを実行しますが、これらは異なる実験用であるため、4 つすべてではありません。
注意: パラホルムアルデヒド(固定剤)を取り扱うときは、適切なPPEを使用し、慎重に進めてください。
- フラットマウント免疫蛍光法の調製
- 網膜を完全に覆うのに十分な溶液を使用して、分割網膜を氷上で4%パラホルムアルデヒドで30分間インキュベートします。
- 裂けた網膜を5〜10 mLの室温PBSで3回洗浄します。オプションの一時停止:裂けた網膜は、4°CのPBSに最大24時間放置することができます。
- 裂けた網膜の垂直切片による免疫蛍光法の調製
- 網膜を完全に覆うのに十分な溶液を使用して、分割網膜を氷上で4%パラホルムアルデヒドで30分間インキュベートします。
- 裂けた網膜を5〜10 mLの室温PBSで3回洗浄します。オプションの一時停止:裂けた網膜は、4°CのPBSに最大24時間放置することができます。
- メンブレンを付着したまま、分割網膜を10%、20%、30%のスクロースに4°Cでそれぞれ1時間順次浸漬し、組織を凍結保護します。
- 凍結保護された分割網膜を最適な切断温度(O.C.T.)コンパウンドに包埋し、凍結切片まで-80°C(最大6か月)で保存します。
- 埋め込まれた裂網膜を-80°Cから除去し、クライオスタットを使用して厚さ20μmの切片を切断します。切片を静電気を帯びたガラス顕微鏡スライドに取り付け、風乾させてから、-20°Cで最大6ヶ月間保管します。
- 二重蛍光 in situ ハイブリダイゼーションと免疫組織化学の調製
- 網膜を完全に覆うのに十分な溶液を使用して、分割網膜を4%パラホルムアルデヒドで氷上で2時間インキュベートします。
- 裂けた網膜を5〜10 mLの室温PBSで3回洗浄します。オプションの一時停止:裂けた網膜は、4°CのPBSに最大24時間放置することができます。
- 電気生理学の準備
- マイクロピペットプーラーを使用して、フィラメント付きの厚肉ホウケイ酸ガラスピペットを引っ張ってパッチピペットを調製します。測定抵抗が6〜10MΩのピペットのみを使用してください。
- プルしたピペットを、125 K-グルコン酸、8 KCl、5 HEPES、1 MgCl 2、1 CaCl 2、0.2 EGTA、3 ATP-Mg、および0.5 GTP-Naを含む内部溶液(単位:mM)でバックフィルします。
- ニトロセルロース膜からの裂けた網膜の除去
- 疎水性バリアペンを使用して、顕微鏡スライド(直径~1cm)に円形のウェルを準備し、5〜10分間風乾させます。
- 準備した疎水性バリアペンウェル内に分割網膜を配置し、それらを完全に覆うのに十分なPBSを追加します。
- 解剖顕微鏡で、細い絵筆の毛を組織の端の下に押し込み、ゆっくりと上に持ち上げます。このようにして、網膜の周りを円を描くように動かして、網膜から持ち上げます。
- 鉗子を使用して、浮遊する網膜の下から膜を取り除きます。
- 網膜片が神経節細胞側を下にして顕微鏡スライド上に静止するように、残りのPBSを慎重に吸引します。
メモ: 次の手順は、順番に実行しないでください。目的のアプリケーション(免疫染色または二重蛍光in situハイブリダイゼーション[FISH]および免疫組織化学[IHC]または電気生理学)に適したプロトコルを選択します。
6. 免疫染色
- まだ準備していない場合は、疎水性バリアペンを使用して、顕微鏡スライド(直径~1cm)に円形のウェルを作成し、5〜10分間自然乾燥させます。すべてのインキュベーションステップと洗浄ステップは、これらのペンウェル内で実行されます。
- 網膜の分割または網膜の垂直分割切片を抗体インキュベーション溶液(AIS:3%ウマ血清、0.5%Triton X-100、0.025%NaN3 in PBS)で室温で30分間インキュベートします。
- 網膜の分割または垂直網膜の分割切片を、AISで希釈した一次抗体と室温で1時間インキュベートします。
注:一次抗体のインキュベーション時間には、さまざまなタンパク質ターゲットと抗体の最適化が必要です。 - ティッシュを室温PBSで3回洗浄します。
- AISで希釈した二次抗体と組織を室温で1時間インキュベートします。ティッシュを室温PBSで3回洗浄します。
- 核染色が必要な場合は、PBSで希釈したDAPIで組織を室温で30秒間インキュベートします。ティッシュを室温PBSで1回洗浄します。
- 各ティッシュにスライド封入剤を一滴垂らし、ガラスカバーガラスを貼付します。
- カバーガラスの端にマニキュアを塗って、サンプルを密封します。スライドは4°Cで保管してください。
7. デュアルFISHとIHC
- スプリット網膜を40°Cのハイブリダイゼーションオーブンで30分間焼き、スライドへの密着性を高めます。
- RNAscope FISHプロトコルは、メーカーのプロトコルに従って完了しますが、以下の例外と変更があります。
- 抗原賦活化ステップは必要ありません。プロテアーゼIIIは、室温で18分のインキュベーション時間で使用します。
- 疎水性バリアペンで作られたウェル内のスライドですべての洗浄手順を実行します。
- サンプルをPBSで希釈した一次抗体( 材料表参照)中で、ハイブリダイゼーションオーブンで40°Cで30分間インキュベートします。サンプルを室温PBSで3回洗浄します。
- サンプルをPBSで希釈した二次抗体( 材料表を参照)中で、ハイブリダイゼーションオーブンで40°Cで30分間インキュベートします。サンプルを室温PBSで3回洗浄します。
- サンプルを1x DAPIで室温で30秒間インキュベートします。サンプルを室温PBSで1回洗浄します。
- ティッシュの各部分に褪色防止封入剤を一滴垂らし、ガラスカバーガラスを覆布します。
- カバーガラスの端にマニキュアを塗って、サンプルを密封します。スライドは4°Cで保管してください。
8.電気生理学
- ニトロセルロースメンブレンを取り外した後、分割された網膜をパッチクランプ記録チャンバーに移し、プラチナハープで静かに固定します。
- 実験全体を通して、95%O2 および5%CO2で炭化したエームス溶液で分割網膜を連続的に灌流します。溶液を32〜34°Cに維持します。
注:実験中、組織はDodtグラジエントコントラスト顕微鏡を使用して視覚化できます。 - 室内照明の下で、セル全体の電圧クランプを実行して、INLニューロンから記録します。
- 記録中に、マイクロセルラー注入ユニットを使用して医薬品化合物を塗布するか、チャネルロドプシン(ChR2)を刺激するために470 nm LEDを使用して光応答をシミュレートします。
注意: 光強度は、デジタル光パワーメーターを使用して測定できます。
- 記録中に、マイクロセルラー注入ユニットを使用して医薬品化合物を塗布するか、チャネルロドプシン(ChR2)を刺激するために470 nm LEDを使用して光応答をシミュレートします。
9. 共焦点顕微鏡
- 共焦点免疫蛍光法では、40倍/1.3倍または63倍/1.40の油浸対物レンズを使用して共焦点顕微鏡で画像を撮影します。FIJI を使用して、明るさとコントラストを調整し、イメージ スタックから Z 投影を生成します。
結果
網膜分裂は光受容体末端を温存する
網膜分裂がOPLの2次ニューロンの樹状突起を損傷しないことを確認するために、分裂した網膜の垂直切片をシナプス小胞タンパク質シナプトフィジン(緑)およびプロテインキナーゼCα(PKCα;赤)に対する抗体で染色しました。裂けた網膜の上部を横切るシナプトフィジン標識の強いバンドは、視細胞のシナプス末端が保持されていることを示しています(図2)。さらに、PKCα染色により、桿体双極細胞(RBC)の正常な形態が明らかになります。視細胞核は見えず、網膜がOPLと視細胞体の最も内側の列に分かれていることを示しています(図2)。

図2:分割網膜は視細胞末端を保持します。 分裂手順後に凍結切片した唾液網膜の垂直断面を示す蛍光共焦点顕微鏡写真(厚さ20μm)。各画像は、共焦点 z スタックの最大投影です。この切片は、PKCα(中央上)とシナプトフィジン(右上)に対する抗体で免疫標識され、それぞれ赤血球とシナプス小胞を可視化しました。マージされた画像(下)は、視細胞末端に存在するシナプス小胞(緑)を示しており、OPLの赤血球(赤)の頂端突起のすぐ上にあります。細胞核はDAPI(青色)で標識されています。ONL内には視細胞核は見えません。略語:ONL =外側核層;OPL =外側の網状層;INL = 内側核層;IPL = 内側の網状層;GC = 神経節細胞。スケールバー = 10 μm。 この図の拡大版をご覧になるには、ここをクリックしてください。
{kind=link}
OPLのシナプス形態は網膜分裂後も維持される
pcp2プロモーター14のシナプス前部および後部シナプスタンパク質をpcp2プロモーター下で赤血球にGFPを発現するマウスを用いて、分裂後のこのシナプス層の完全性を評価するために免疫標識を行った14。視細胞の軸索を介して発生するせん断力にもかかわらず、RGS11で標識された赤血球樹状突起とCtBP2 15で標識された視細胞シナプスリボンの正常な位置が観察されるため、分裂はOPLにおける視細胞-BCシナプスの形態を乱しません(図3)。桿体と赤血球の間の各シナプス接触について、RGS11はシナプスリボンの馬蹄形(緑)内にある赤い点状として見ることができます。その後の実験では、抗GPR179抗体16を用いてシナプス後ON-BC樹状突起チップ16を標識し、抗PSD-95抗体を用いてシナプス前桿体視細胞末端を標識した(補足図2)。これらの結果は、赤血球樹状突起がシナプス前パートナーである桿体末端と密接に関連していることが示されているため、分割網膜準備におけるOPLの安定性を再び確認しています。

図3:OPLのシナプス形態は、網膜分裂後も維持される。 Pcp2プロモーター下で赤血球にGFPを発現するトランスジェニックマウスの網膜の分裂した共焦点免疫蛍光画像。GFP発現レベル(青)は、網膜の赤血球によって異なります。分裂後、網膜を固定し、CtBP2(緑)およびRGS11(赤)に対する抗体とインキュベートして、それぞれ視細胞シナプスリボンおよびON-BC樹状突起チップを標識した。各赤-緑のペアは、桿体とON-BCの間のシナプス接触を表す。スケールバー = 10 μm。 この図の拡大版をご覧になるには、ここをクリックしてください。
{kind=link}
網膜の分裂により、赤血球の生存率が維持されます
分裂後の網膜内ニューロンの生存率を評価するために、膜不透過性の近赤外核色素(MI-NIR)が使用され、死細胞の同定が可能になりました。MI-NIRでインキュベートした後、分割網膜を固定し、抗PKCαで標識して赤血球を同定した。このばらつきは、解剖、分割、または取り扱い手順中に網膜の特定の領域に加えられた損傷に起因する可能性があります(図4)。赤血球の細胞体が分裂部位に近いINLの最も外側の領域に存在することを考えると、その生存率の慎重な評価が必要でした。PKCαとMI-NIRの共局在が乏しいことから、ほとんどの赤血球は網膜分裂後も生存可能であることが確認されました(図4)。

図4:桿体双極細胞は網膜分裂後も生存可能。 フラットマウントの視点で網膜の裂けた領域を示す蛍光共焦点顕微鏡写真。分割後、生きた網膜をMI-NIR色素(赤色)と37°Cで30分間インキュベートしました。 次に、網膜を固定し、PKCαに対する抗体で免疫標識して、赤血球を可視化しました。網膜のこの領域では、PKCαとMI-NIRの共局在はまれです。MI-NIRは、赤血球に属さない核(青)と共局在します。 略語:MI-NIR = 膜不透過性近赤外生/死染色。スケールバー = 10 μm。 この図の拡大版をご覧になるには、ここをクリックしてください。
{kind=link}
スプリット網膜は、デュアルFISHとIHCに適しています
標準的なIHCの固定時間を延長することにより、分割網膜をFISHとIHCで連続的に処理して、mRNAとタンパク質を同時に標識することができます17,18。実験により、4%パラホルムアルデヒドに2時間固定すると、抗体結合のためのタンパク質エピトープを維持しながら、堅牢なmRNA標識が得られることが確認されました。FISHを裂網膜で実施し、続いてIHCで実施し、GABAA受容体サブユニットδ(GABRD;抗センスmRNAプローブ)の発現を、外側INLのRBC(抗PKCα抗体)の位置との関係で可視化しました(図5A)。GABRD mRNAの発現は赤血球ではまれである(図5A)。しかし、転写産物はアマクリン細胞と神経節細胞によって豊富に発現しており、無傷の網膜からの横断面の標識パターンによって証明されています(図5B)。外側のINL(図5A)では、GABRD mRNAは、異なる細胞に集中している内側のINL(図5C)と比較して、より均一に分布しています。他のGABA受容体サブユニットを標的とするアンチセンスプローブは、異なる標識パターンを産生し、プローブの特異性を実証する(データは示さず)。

図5:裂けた網膜と無傷の網膜における二重FISHとIHC 。 (A、C)フラットマウント分割網膜の共焦点顕微鏡写真、(B)無傷の網膜からの垂直切片。(A)と(C)の画像は、それぞれINLの上部領域と下部領域における光学セクションの最大投影です。(B) の点線の四角形は、(A) と (C) に示す投影の作成に使用されるおおよその境界を表します。裂けた網膜(A、C)を2時間固定し、GABRD(赤色)に対するアンチセンスmRNAプローブで標識した。その後、裂けた網膜をPKCαに対する抗体で染色し、赤血球(緑色)を標識した。PKCαチャネルは、明瞭にするために、より低いINLの予測から省略しました。(B)の無傷の網膜は、切片形成前に24時間固定した。その後、固定網膜をGABRD(赤)に対するアンチセンスmRNAプローブで標識した。すべてのサンプルをDAPI(青色)で20秒間染色してから、カバーガラスを封入しました。略語:INL =内側核層。スケールバー = 10 μm。 この図の拡大版をご覧になるには、ここをクリックしてください。
{kind=link}
スプリット網膜は、BCおよびHCからのパッチクランプ電気生理学的記録に適しています
従来の全マウント網膜のBCまたはHC体細胞にパッチを貼るには、ピペットを神経節細胞側または視細胞側からアプローチする必要があります。どちらのアプローチでも、INLに到達するためにいくつかの網膜層を横断する必要があり、その間、ピペットチップが破片で塞がれることがよくあります。ビブラトームスライスの調製では、BC体細胞とHC体細胞に容易にアクセスできますが、樹状突起が切断され、横方向の接続が破壊される可能性があります。しかし、網膜が裂けた状態では、赤血球とHCの細胞体が組織の表面にあるため、OPLの側方回路を維持しながら、パッチピペットへのアクセスが大幅に改善されます。
図6は、分裂した網膜のBCから記録された化学的にシミュレートされた光応答を示しています。灌流エーム培地にグループIIIのmGluRアゴニストであるL-AP4(4 μM)を添加し、暗闇での視細胞からのグルタミン酸放出をシミュレートしました。mGluR6 アンタゴニストである CPPG(600 μM、エイムズ中)をパッチを貼った細胞の樹状突起(-60 mV に保持)に吹き付けて、mGluR6 の阻害による閃光をシミュレートしました。細胞はCPPGパフに2種類の内向き電流で応答した。1つのタイプは、網膜切片の赤血球から記録された標準的な光誘発電流と同様に、過渡電流とそれに続くプラトー(図6A)を示しています19。もう1つのタイプは、パフ持続時間全体にわたって持続し(図6B)、ONコーンバイポーラセル(ON-CBC)から記録された電流に類似しています19。
スライス調製物では保存が困難なことが多い樹状突起野が広い細胞タイプであるHCを標的とする別の実験が行われました。HC中のチャネルロドプシン(ChR2)およびGFPを発現するマウス系統を用いて、蛍光顕微鏡下での同定を容易にした。まず、HCからの電流は、一連の脱分極ステップ(-100mV〜50mV、ステップサイズ= 15mV)に応答して記録され、内向きの電流とそれに続く外向きの電流で応答しました(図6C)。次に、これらの細胞を短時間の青色光パルス(200 ms、470 nm)で刺激し、2つの細胞にChR2駆動の大きな内向き電流を生成しました(図6D)。

図6:裂けた網膜のINLニューロンからのパッチクランプ記録 。 (A)推定 RBC および(B)CBC を、L-AP4 (4 μM) を含む灌流エーム培地中で -60 mV で電圧クランプしました。クランプした細胞の樹状突起にCPPG(600μM)をパフすると、赤血球では一過性であるがCBCでは持続する内向きの電流が呼び出された。(A)のRBC記録は1トレースですが、(B)のCBC記録は3トレースの平均を表します。(C)vGATFLPo内のHCからのパッチクランプ記録。vGlut2Cre;Ai80dマウス。赤い線は、ChR2を流れる大きな内向きの電流を呼び出すために使用される200ms、470nmの光パルスの持続時間を示しています。 (D)-60mVで電圧クランプされたHCから注入された電流応答は、15mV間隔で-70mVと+35mVの間をステップアップし、-60mVに戻しました。挿入図は、電圧ステップの開始を囲む6msのウィンドウに同じトレースを示しています。(E)vGATFLPoでGFPを発現する水平細胞を示すフラットマウント分割網膜の免疫蛍光顕微鏡写真;vGlut2Cre;Ai80dマウス。スケールバー = 20 μm。電気生理学データは 20 kHz のサンプリング レートで収集し、5 kHz のローパス ベッセル フィルターでフィルタリングしました。その後、データをエクスポートし、Python 3 を使用してオフラインの視覚化と分析を実行しました。 この図の拡大版をご覧になるには、ここをクリックしてください。
{kind=link}
網膜分割により、INLおよびOPLの解剖学的構造の迅速な調査が可能
網膜の外部制限膜(ELM)とONLは厚さ~90μmのバリアで構成されており、網膜内への抗体の拡散を妨げ、最適ではない免疫染色条件を作り出します20,21,22。したがって、従来のフラットマウント網膜を使用したOPLまたはINLの免疫標識ターゲットには、時間のかかる染色プロトコルが必要であり、多くの場合、48〜96時間の抗体インキュベーションが必要です5、6、7、8、20、22。
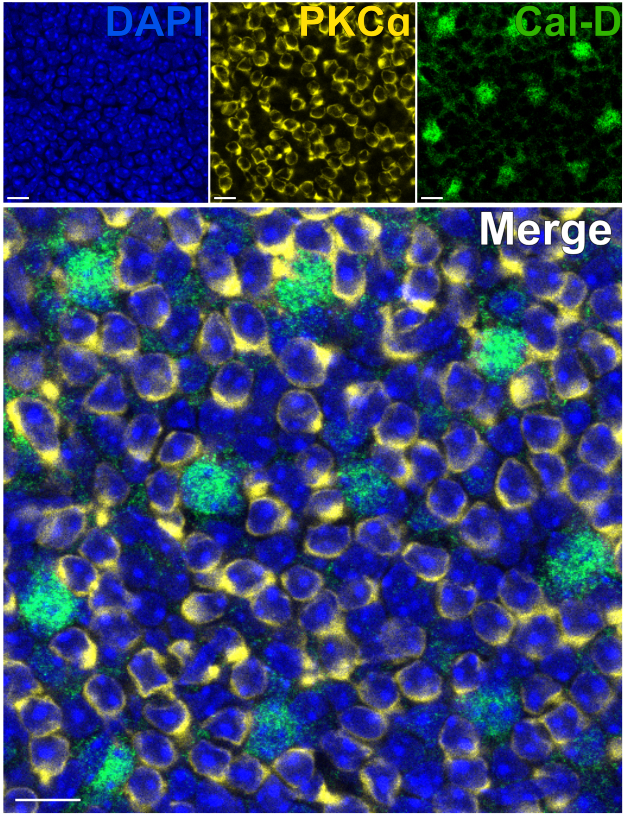
視細胞を除去することで、網膜内ニューロンへの迅速な抗体浸透が可能になります。その結果、網膜内タンパク質ターゲットの標識は、色素標識一次抗体を使用することで、わずか1時間で達成できます。PKCαおよびカルビンジン-Dに対する抗体を使用して、それぞれINLの赤血球およびHCを標識しました(図7)。広視野ニューロンの側方突起を切り捨てる従来の垂直網膜切片とは異なり、スプリット網膜調製物はHCなどの広視野細胞の樹状突起全体を可視化することができます(図6E、 図7)。

図7:分割網膜における網膜内タンパク質の迅速な免疫標識。 フラットマウントの視点から見た裂けた網膜の共焦点免疫蛍光画像。裂けた網膜をPKCα(黄色)およびカルビンジンD(緑)に対する抗体と室温で1時間インキュベートし、それぞれON-BCおよびHCを標識しました。(A)各シングルチャンネル画像は、4つの光学セクションで構成される平均Z投影です:DAPI、平均z10-13;カルビンジン-D、平均z11-14;PKCα、平均z11-14。(B)合成された画像では、同じ投影が重ね合わされています。スケールバー = 10 μm。 この図の拡大版をご覧になるには、ここをクリックしてください。
{kind=link}
補足図1:網膜解離の主要な段階。 すべての画像は、解剖顕微鏡の接眼レンズに取り付けられたスマートフォンのカメラで撮影されました。(A)角膜摘出後のマウスの眼球の上から下へ。(B)レンズを取り外した後のマウスアイカップのトップダウン画像。(C)マウスのアイカップの強膜に小さな切開を行います。矢印は強膜の2つのフラップを示しており、鉗子によって反対方向に引っ張られ、網膜を強膜から分離し始めます。(D)強膜を網膜から部分的に引き離した後、強膜と網膜の間にバンナハサミを挿入し、視神経を切断して網膜を解放します。赤い点線の円は視神経乳頭を示し、はさみは正しい切断軌跡を示しています(強膜と網膜の間にはさみを挿入します)。強膜後の孤立した網膜がこじ開けられます。 このファイルをダウンロードするには、ここをクリックしてください。
補足図2:裂けた網膜におけるOPLのシナプス前部および後部成分の特性評価。 分割網膜内のOPLからの共焦点免疫蛍光画像。分割網膜をGPR179およびPSD95に対する抗体とともに室温で1時間インキュベートし、ON-BCの樹状突起先端および桿体光受容体の末端にそれぞれ標識しました。左と中央の画像は、いくつかの光学セクションの最大投影です。右端の画像にも同じ投影が重ね合わされています。ON-BC樹状突起先端のGPR179点は、桿体視細胞末端と密接に関連していることが見られ、OPL内で無傷のシナプス接触を示しています。スケールバー = 10 μm。 このファイルをダウンロードするには、ここをクリックしてください。
補足図3:トラブルシューティング:裂けた網膜の質の評価。 細胞核を可視化するためにDAPIで染色した分割網膜の蛍光顕微鏡写真。細胞は、核の直径と組織の深さに基づいて識別できます。(A)視細胞核は小さく、明るく、表面的ですが、(B)BC核は大きく、暗く、深くなっています。(C)視細胞が不完全に除去された領域の低倍率画像。焦点が合って見える核はBCからのもので、焦点が合っていないように見える画像の端にある視細胞核よりも深いです。(A)および(B)のスケールバー=20μm。(C) = 50 μm のスケールバー。 このファイルをダウンロードするには、ここをクリックしてください。
ディスカッション
光受容体が光子吸収を神経伝達物質放出に変換した後、BCおよびHCは、視覚信号を処理する最初の網膜ニューロンである23。これらのニューロンの重要性はよく認識されていますが、その機能の多くは完全に理解されていないか、まったく調査されていません。多くのBCおよびHC生理学研究は、横方向の接続性を維持しながらINLニューロンへのアクセスを改善するフラットマウント網膜製剤の恩恵を受ける可能性があります。スプリット網膜法の開発は、フラットマウント配向のBCおよびHCから高品質の電気生理学的記録および顕微鏡データを取得するための簡単なプロトコルを提供するための努力を表しています。ここで説明する分割網膜調製は、特殊な機器を使用せずに、網膜単離後、マウス1匹あたり約20分(網膜あたり10分)で実行できます。この方法は、既存の視細胞除去手順からインスピレーションを得ていますが、シンプルさ、速度、および汎用性が大幅に向上しています4,10,11,12,13。網膜層を分離するための以前の方法とは異なり、網膜の分割は、凍結、凍結乾燥、または網膜への接着剤の繰り返し塗布を必要としません。練習すれば、ほぼすべての光受容体をニトロセルロース膜で1回の裂け目で除去することができます。このアプローチの速さと容易さにより、網膜がカルボゲン化されたエイムから抜け出す時間を最小限に抑えることができ、長期間にわたって高い細胞生存率を得ることができます。分割網膜は、カルボゲン処理されたエイム培地中で、分割後数時間維持することができます。この調製物におけるINLニューロンの健全性の証として、生細胞染色/死細胞染色(図4)およびパッチクランプ電気生理学(図6)により、分裂後の赤血球およびHCの生存率が確認されました。
裂けた網膜の視細胞層の除去は、抗体のINLへの拡散時間を劇的に短縮することにより、免疫標識中に大きな利点をもたらします。一次抗体および二次抗体の標識は2時間以内に完了でき、ターゲット5、6、7、8、20、22に応じて72時間以上かかる従来のフラットマウント染色を大幅に改善します。その結果、組織調製と同日に顕微鏡データを取得でき、免疫蛍光実験のペースを大幅に加速できます。mRNAプローブのアニーリングを容易にするために、FISH実験では、通常、免疫標識よりもはるかに長い固定時間(~24時間)が推奨されています18。しかし、ここで紹介する実験では、2時間固定しても例外的なFISH標識が得られることが実証されています(図5)。固定時間を30分から2時間に延長したにもかかわらず、優れた免疫標識を得るために抗原賦活化ステップを行う必要はありませんでしたが、これは抗体または抗原によって異なる場合があります。FISHプロトコルにおけるプロテアーゼ処理は、おそらく標的エピトープの破壊により、抗体標識を妨げる可能性があります。この問題は、複数のエピトープを標的とするポリクローナル抗体を使用することで回避され、エピトープの破壊が免疫標識を妨げる可能性を低減しました。さらに、適度なプロテアーゼ処理(ACDプロテアーゼIII)を使用して、十分な組織浸透性を維持しながら、過剰なエピトープの変化を防止しました。
時折、網膜は代わりに外核層(ONL)を裂き、INL細胞が見えない視細胞体細胞の層を残します。これを防ぐには、網膜がガラス上で完全に平らになり、網膜の周りから残留液体が取り除かれていることを確認する必要があります。絵筆でニトロセルロースをより強く押すと、ONLの裂け目を防ぐのにも役立ちます。膜が濡れすぎたり、網膜が折りたたまれたりすると、裂け目が成功する可能性は大幅に減少します。DAPIを使用して細胞核を染色することは、分裂の質を評価し、残りの光受容体のカバー率を決定するのに役立ちます。視細胞核はより小さく、より明るく、より表面的であるのに対し(補足図3A)、BC核はより大きく、より暗く、より深い(補足図3B)。場合によっては、裂傷の平面が網膜の断片を横切ってわずかに変化し、視細胞体が完全に除去されていない斑点が生じます(補足図3C)。顕微鏡検査や電気生理学のアプリケーションでは、光受容体が適切に除去された領域から高品質のデータを収集する能力が妨げられることはありません。露出した網膜内部の大きな視野は、パッチピペットでイメージングまたは記録すると簡単に見つけることができます。より完全な光受容体除去が望まれる場合は、100%の光受容体除去が保証されるわけではありませんが、ニトロセルロースメンブレンの追加片で2回目の裂傷を行うことができます。したがって、残留光受容体材料が結果に影響を与える可能性のある遺伝子発現またはプロテオミクス研究で分割網膜を使用する場合は注意が必要です。シングルセルアプリケーションの場合、光受容体からのデータを分析から除外できるため、この懸念は杞憂です。
スプリット網膜調製の利点は、おそらく広視野介在ニューロンの電気生理学的記録において最も顕著である。従来の縦型スライスでは広視野細胞の広範なプロセスが切断されますが、スプリット網膜調製ではOPLとIPLがそのまま残るため、縦型スライスでは見落とされてしまうHC24、A17s25、TH AC26、NOS-1AC27 などの広視野細胞からの入力を取り込むことができます。したがって、結果の解釈と網膜スライスから収集された以前のデータとの比較には、慎重な検討が必要です。それにもかかわらず、光刺激の薬理学的模倣を用いた実験では、これらの結果は網膜切片から記録されたデータに類似している19。細胞特異的プロモーター下でChR2を発現させることにより、INLのBCから記録しながら目的の細胞集団を刺激し、目的の細胞が垂直情報経路に及ぼす影響を調べることができます。アマクリン細胞などのより深いINLニューロンから直接記録することも、網膜分裂で実現可能です。この場合、パッチ電極は最初により表面的なINLニューロンを通過する必要がありますが、従来のホールマウント調製と比較すると、その経路を塞ぐ組織が大幅に少なくなります。
この方法は、他のニューロンに対する広視野細胞の影響を測定することに加えて、樹状突起がOPL28で広範なギャップ結合結合ネットワークを形成するHCからの直接単一細胞パッチクランプを可能にします。水平細胞は、網膜を通る垂直方向の情報の伝達を形作る光受容体に重要なフィードバックを送ります。しかし、HCの樹状突起野は垂直スライスで切り捨てられるため、単一細胞の記録データが不足しています。この研究は、解剖学的および生理学的に無傷のHCを提示し、そこからChR2誘発電流が記録されるトリプルトランスジェニックマウス系統(図6 C-E)で紹介しています。ChR2刺激の外側では、分割網膜を使用して、内因性HC電流およびギャップ結合結合を研究することができます28。分裂した網膜は、化学的適用やChR2刺激によって誘導されるシナプス結合やニューロンの活動を研究するための便利なモデルを提供するが、光受容体がないため、自然光応答や光順応メカニズムを直接調べることはできない。
網膜のin situイメージングは、近年、目覚ましい進歩を遂げています。しかし、画像検査の大部分は、網膜全マウント製剤中の神経節細胞層に限定されている29。著者らは、裂けた網膜に光受容体が存在しないことから、OPLとINLにおける生きたカルシウムイメージングの理想的なモデルになると考えている。このモデルは、カルシウムイメージング以外にも、iGluSnFR30,31、iGABASnFR32、pHluorin 33などの遺伝子コードバイオセンサーでの使用に大きな可能性を秘めています。これらの強力なツールをスプリット網膜調製と組み合わせることで、網膜での光処理に寄与するBCおよびHCのシナプス相互作用と生物物理学的特性を探索するための効率的なアプローチを提供する可能性があります。
開示事項
著者らは、競合する金銭的利益がないことを宣言します。
謝辞
この研究は、以下のNIH助成金によって支援されました:NIH助成金R01EY031596(C.M.へ);NIH助成金R01EY029985(C.M.へ);NIH助成金P30EY010572(C.M.へ);NIH助成金R01EY032564(BSへ)。網膜切片の作製における技術サポートをしてくれたTammie Haleyと、この研究で使用したmRNA FISHプローブを惜しみなく提供してくれたCharles Allen博士に感謝します。
資料
| Name | Company | Catalog Number | Comments |
| #1.5 glass coverslips | Fisherbrand | 12544E | |
| 2 pairs of Dumont #5 forceps | Ted Pella | 38125 | |
| 25 gauge needle | Becton Dickenson | 305122 | |
| 470 nm LED | THORLABS | M470L2 | |
| 5-306 curved scissors | Miltex | 5-306 | |
| 9" disposable pasteur pipetes | Fisherbrand | 13-678-20D | for constructing custom transfer pipette |
| Ai80d mouse | Jackson Laboratories | 25109 | RRID: IMSR_JAX:025109 |
| Ames Medium w/L-Glutamate | US Biological | A1372-25 | |
| amplifier control software | Molecular Devices | Clampex 10.3 software | |
| anti-calbindin D28K antibody | Invitrogen | PA-5 85669 | RRID: AB_2792808, host species = rabbit; 1:100 dilution |
| anti-CtBP2 antibody | BD Biosciences | 612044 | RRID: AB_399431, host species = mouse; 1:5000 dilution |
| anti-GPR179 antibody | NA | NA | gift from Kirill Martemyanov; Scripps Research Institute, Jupiter, FL; host species = sheep; 1:1000 dilution |
| anti-PKC alpha antibody | Sigma-Aldrich | P4334 | RRID: AB_477345, host species = rabbit; 1:5000 dilution |
| anti-PKC alpha antibody | Santa Cruz Biotechnology | sc8393 AF594 | RRID: AB_628142, host species = mouse; 1:1000 dilution |
| anti-PSD95 antibody | BD Transduction Laboratories | 610495 | RRID: AB_397862, host species = mouse; 1:1000 dilution |
| anti-RGS11 antibody | NA | NA | gift from Ted Wensel; Baylor College of Medicine, Houston, TX; host species = rabbit; between 1:1000 and 1:5000 dilution |
| anti-Synaptophysin P38 antibody | Sigma | S-S5768 | RRID: AB_477523, host species = mouse; 1:1000 dilution |
| Aquamount mounting media | Epredia | 13800 | slide mounting media |
| C57BL/6J mouse | Jackson Laboratories | 000664 | RRID: IMSR_JAX:000664 |
| carbogen tank | Matheson | NA | 95% O2 and 5% CO2 |
| custom transfer pipette | custom build | NA | Instructions: use scissors to cut off the tip of a plasitc transfer pipette at the point it begins to taper. Use pliers to safely break off the last 2-3 inches of a glass pasteur pipette. Fit the narrow end of the glass pasteur pipette into the wide tip of the plastic transfer pipete. Wrap parafilm around the joint of the two pieces to enhance the seal. |
| Digitical optical power meter | THORLABS | PM100D | |
| dissection microscope | Zeiss | Stemi 2000 | |
| electrophysiology amplifier | Molecular Devices | Axopatch 200B | |
| electrophysiology microscope | Olympus | OLYMPUS, BX50WI | Dodt gradient contrast microscopy |
| Fluoromount-G | SouthernBiotech | 0100-01 | |
| HC PL APO CS2 40x/1.3 | Leica | 506358 | |
| HC PL APO CS2 63x/1.40 | Leica | 15506350 | |
| Hybridization oven | Robbins Scientific | Model 1000 | for RNAscope protocol only |
| Immedge hydrophobic barrier pen | Vector Laboratories | H-4000 | |
| isoflurane | Piramal Critical Care | 66794-017-25 | |
| Kimwipe (delicate task wipe) | Kimtech Science | 34155 | |
| Leica HC PL APO CS2 40x/1.3 oil immersion objective | Leica | 506358 | |
| Leica HC PL APO CS2 63x/1.40 oil immersion objective | Leica | 15506350 | |
| Leica TCS SP8 X confocal microscope | Leica | discontinued | |
| medium 15 mm petri dish | Corning | 25060-60 | eyes are kept here during retina dissection |
| Merit 97-275 steel scissors | Merit | 97-275 | |
| Micropipette Puller | Sutter Instrument | p-97 | |
| Mm-Gabrd-C2 mRNA probe | ACD | 459481-C2 | |
| mouse euthanasia chamber | NA | NA | custom build; glass petri dish covering a small glass jar. |
| nitrocellulose membrane filters | GE Healthcare Life Sciences; Whatman | 7184-005 | 0.45 µm pore size |
| Picospritzer | General Valve Corporation | Picospritzer II | referred to in the text as microcellular injection unit |
| plastic transfer pipets | Fisherbrand | 13-711-7M | for constructing custom transfer pipette |
| Plastic tubing | Tygon | R-603 | for connection to carbogen tank |
| platinum harp | custom build | NA | for anchoring split retinas within the electrophysiology recording chamber. |
| size 0 paint brush | generic | NA | for flattening retina during splitting. |
| SlowFade Gold antifade reagent | Molecular Probes | S36937 | referred to in the text as anti-fade mounting media |
| small 10 mm petri dish | Falcon | 353001 | eyes are placed here following enucleation |
| small glass pane (7.5 cm x 5 cm) | generic | NA | isolatd retina pieces are placed onto this for the splitting procedure |
| Superfrost plus microscope slides | Fisherbrand | 12-550-15 | electrostatically-charged glass microscope slides |
| Thick-walled borosilicate glass pipettes with filament | Sutter Instrument | BF150-86-10HP | |
| Vannas Scissors; straight | Titan Medical | TMS121 | not brand specific; any comparable scissors will work |
| vGATFLPo mouse | Jackson Laboratories | 29591 | RRID: IMSR_JAX:029591 |
| vGlut2Cre mouse | Jackson Laboratories | 28863, 016963 | RRID: IMSR_JAX:028863, RRID: IMSR_JAX:016963 |
| Zombie NIR Fixable Viability Kit | BioLegend | 423105 | referred to in the text as MI-NIR |
参考文献
- Morgans, C. W. Neurotransmitter release at ribbon synapses in the retina. Immunology & Cell Biology. 78 (4), 442-446 (2000).
- Euler, T., Haverkamp, S., Schubert, T., Baden, T. Retinal bipolar cells: elementary building blocks of vision. Nature Reviews Neuroscience. 15 (8), 507-519 (2014).
- Barnes, S., Grove, J. C. R., McHugh, C. F., Hirano, A. A., Brecha, N. C. Horizontal Cell Feedback to Cone Photoreceptors in Mammalian Retina: Novel Insights From the GABA-pH Hybrid Model. Frontiers in Cellular Neuroscience. 14, (2020).
- Walston, S. T., Chang, Y. C., Weiland, J. D., Chow, R. H. Method to remove photoreceptors from whole mount retina in vitro. Journal of Neurophysiology. 118 (5), 2763-2769 (2017).
- Stefanov, A., Novelli, E., Strettoi, E. Inner retinal preservation in the photoinducible I307N rhodopsin mutant mouse, a model of autosomal dominant retinitis pigmentosa. Journal of Comparative Neurology. 528 (9), 1502-1522 (2020).
- Matsuoka, R. L., Nguyen-Ba-Charvet, K. T., Parray, A., Badea, T. C., Chédotal, A., Kolodkin, A. L. Transmembrane semaphorin signaling controls laminar stratification in the mammalian retina. Nature. 470 (7333), 259-263 (2011).
- Matsuoka, R. L., et al. Guidance-Cue Control of Horizontal Cell Morphology, Lamination, and Synapse Formation in the Mammalian Outer Retina. Journal of Neuroscience. 32 (20), 6859-6868 (2012).
- Wässle, H., Puller, C., Müller, F., Haverkamp, S. Cone Contacts, Mosaics, and Territories of Bipolar Cells in the Mouse Retina. Journal of Neuroscience. 29 (1), 106-117 (2009).
- Thoreson, W. B., Dacey, D. M. Diverse Cell Types, Circuits, and Mechanisms for Color Vision in the Vertebrate Retina. Physiological Reviews. 99 (3), 1527-1573 (2019).
- Guido, M. E., et al. A simple method to obtain retinal cell preparations highly enriched in specific cell types. Suitability for lipid metabolism studies. Brain Research Protocols. 4 (2), 147-155 (1999).
- Rose, K., Walston, S. T., Chen, J. Separation of photoreceptor cell compartments in mouse retina for protein analysis. Molecular Neurodegeneration. 12 (1), 28(2017).
- Todorova, V., et al. Retinal Layer Separation (ReLayS) method enables the molecular analysis of photoreceptor segments and cell bodies, as well as the inner retina. Scientific Reports. 12 (1), 20195(2022).
- Shiosaka, S., Kiyama, H., Tohyama, M. A simple method for the separation of retinal sublayers from the entire retina with special reference to application for cell culture. Journal of Neuroscience Methods. 10 (3), 229-235 (1984).
- Ivanova, E., Hwang, G. S., Pan, Z. H. Characterization of transgenic mouse lines expressing Cre-recombinase in the retina. Neuroscience. 165 (1), 233-243 (2010).
- Sarria, I., Orlandi, C., McCall, M. A., Gregg, R. G., Martemyanov, K. A. Intermolecular Interaction between Anchoring Subunits Specify Subcellular Targeting and Function of RGS Proteins in Retina ON-Bipolar Neurons. The Journal of Neuroscience. 36 (10), 2915-2925 (2016).
- Orlandi, C., Cao, Y., Martemyanov, K. A. Orphan Receptor GPR179 Forms Macromolecular Complexes With Components of Metabotropic Signaling Cascade in Retina ON-Bipolar Neurons. Investigative Ophthalmology & Visual Science. 54 (10), 7153-7161 (2013).
- Dikshit, A., Zong, H., Anderson, C., Zhang, B., Ma, X. -J. Simultaneous Visualization of RNA and Protein Expression in Tissue Using a Combined RNAscopeTM In Situ Hybridization and Immunofluorescence Protocol. Methods in Molecular Biology. 2148, Clifton, N.J. 301-312 (2020).
- Wang, F., et al. RNAscope. The Journal of Molecular Diagnostics. 14 (1), 22-29 (2012).
- Morgans, C. W., et al. TRPM1 is required for the depolarizing light response in retinal ON-bipolar cells. Proceedings of the National Academy of Sciences of the United States of America. 106 (45), 19174-19178 (2009).
- Alessio, E., Zhang, D. Q. Immunostaining of whole-mount retinas with the CLARITY tissue clearing method. Investigative Ophthalmology & Visual Science. 61 (7), 5054(2020).
- Ferguson, L. R., Dominguez, J. M., Balaiya, S., Grover, S., Chalam, K. V. Retinal Thickness Normative Data in Wild-Type Mice Using Customized Miniature SD-OCT. PLoS ONE. 8 (6), e67265(2013).
- Ivanova, E., Toychiev, A. H., Yee, C. W., Sagdullaev, B. T. Optimized Protocol for Retinal Wholemount Preparation for Imaging and Immunohistochemistry. Journal of Visualized Experiments JoVE. (82), e51018(2013).
- Kolb, H. Neurotransmitters in the Retina. Webvision: The Organization of the Retina and Visual System. , University of Utah Health Sciences Center. (1995).
- Chaya, T., et al. Versatile functional roles of horizontal cells in the retinal circuit. Scientific Reports. 7 (1), 5540(2017).
- Egger, V., Diamond, J. S. A17 Amacrine Cells and Olfactory Granule Cells: Parallel Processors of Early Sensory Information. Frontiers in Cellular Neuroscience. 14, 600537(2020).
- Dacey, D. M. The dopaminergic amacrine cell. The Journal of Comparative Neurology. 301 (3), 461-489 (1990).
- Park, S. J., et al. Connectomic analysis reveals an interneuron with an integral role in the retinal circuit for night vision. eLife. 9, 56077(2020).
- Janssen-Bienhold, U., et al. Connexin57 is expressed in dendro-dendritic and axo-axonal gap junctions of mouse horizontal cells and its distribution is modulated by light. The Journal of Comparative Neurology. 513 (4), 363-374 (2009).
- Jain, V., et al. The functional organization of excitation and inhibition in the dendrites of mouse direction-selective ganglion cells. eLife. 9, 52949(2020).
- Marvin, J. S., et al. Stability, affinity, and chromatic variants of the glutamate sensor iGluSnFR. Nature Methods. 15 (11), 936-939 (2018).
- Strauss, S., et al. Center-surround interactions underlie bipolar cell motion sensitivity in the mouse retina. Nature Communications. 13 (1), 5574(2022).
- Marvin, J. S., et al. A genetically encoded fluorescent sensor for in vivo imaging of GABA. Nature Methods. 16 (8), 763-770 (2019).
- Beckwith-Cohen, B., Holzhausen, L. C., Wang, T. M., Rajappa, R., Kramer, R. H. Localizing Proton-Mediated Inhibitory Feedback at the Retinal Horizontal Cell-Cone Synapse with Genetically-Encoded pH Probes. The Journal of Neuroscience: The Official Journal of the Society for Neuroscience. 39 (4), 651-662 (2019).
転載および許可
このJoVE論文のテキスト又は図を再利用するための許可を申請します
許可を申請さらに記事を探す
This article has been published
Video Coming Soon
Copyright © 2023 MyJoVE Corporation. All rights reserved