
Method Article
辅助性T细胞作为一种遗传方法来研究逆转录病毒转导机制控制其分化和功能
摘要
许多实验系统已被利用来理解调节T细胞的发育和功能中的免疫应答的机制。这里使用逆转录病毒转导的基因的方法的描述,这是经济,时间效率,最重要的是,高度信息识别调控通路。
摘要
Helper T cell development and function must be tightly regulated to induce an appropriate immune response that eliminates specific pathogens yet prevents autoimmunity. Many approaches involving different model organisms have been utilized to understand the mechanisms controlling helper T cell development and function. However, studies using mouse models have proven to be highly informative due to the availability of genetic, cellular, and biochemical systems. One genetic approach in mice used by many labs involves retroviral transduction of primary helper T cells. This is a powerful approach due to its relative ease, making it accessible to almost any laboratory with basic skills in molecular biology and immunology. Therefore, multiple genes in wild type or mutant forms can readily be tested for function in helper T cells to understand their importance and mechanisms of action. We have optimized this approach and describe here the protocols for production of high titer retroviruses, isolation of primary murine helper T cells, and their transduction by retroviruses and differentiation toward the different helper subsets. Finally, the use of this approach is described in uncovering mechanisms utilized by microRNAs (miRNAs) to regulate pathways controlling helper T cell development and function.
引言
The immune response must be highly regulated to eliminate infections but prevent attacks on self-tissue that lead to autoimmunity. Helper T cells play an essential role in regulating the immune response, and a great deal of effort has been undertaken to understand their development and function (illustrated in several recent reviews 1-3). However, many questions remain, and many approaches have been utilized to study the mechanisms controlling helper T cell development and function. These have ranged from the use of in vitro cell culture systems to whole animals. Cell culture systems, especially those using cell lines, offer the benefit of ease of use and the ability to generate large amount of material to do sophisticated biochemical analyses. However, they suffer from their limited ability to reproduce the actual conditions occurring in an immune response. In contrast, whole animal experiments offer the benefit of relevance, but they can suffer from difficulties in manipulation and the ability to perform precise controls in addition to their large costs and ethical implications. Nevertheless, the vast majority of helper T cells studies today still require the use of whole animal experiments involving primary T cells because of the inability of cell lines to duplicate the exact steps occurring in the whole animal. Therefore, it is essential to utilize cost effective approaches that are highly informative.
Genetics is one powerful tool to study helper T cell development and function, yet traditional methods involving gene knockouts or transgenes are time consuming and expensive so they are often out of reach of small labs. However, retroviral transduction offers a powerful, rapid and, cost effective genetic approach to study the mechanisms of specific gene products. Therefore, it is commonly used in papers studying helper T cell development and function.
We have optimized a procedure for retroviral transduction of helper T cells. It utilizes the pMIG (Murine stem cell virus-Internal ribosomal entry site-Green fluorescent protein) retroviral expression vector, in which the gene of interest can be cloned and thereby expressed from the retrovirus long terminal repeat (LTR) 4. In addition, downstream of the inserted gene of interest is an internal ribosome entry sequence (IRES) followed by the green fluorescent protein (GFP) gene so transduced cells can easily be followed by their expression of GFP. The vector was originally derived from the Murine Stem Cell Virus (MSCV) vectors, which contain mutations in repressor binding sites in the LTRs making them resistant to silencing and thus, giving high expression in many cell types including helper T cells 5,6. Production of high titer retrovirus requires a simple transient transfection protocol of human embryonic kidney (HEK) 293T cells with the MIG vector and a helper virus vector that expresses the retroviral GAG, Pol, and Env genes. For this the pCL-Eco helper virus vector 7 works well in producing high titer replication incompetent retroviruses.
Here these protocols for retroviral production and transduction of primary murine T cells are described in addition to some of our results using this approach to study miRNA regulation of gene expression controlling helper T cell differentiation. miRNAs are small RNAs of approximately 22 nucleotides in length that post-transcriptionally regulate gene expression by targeting homologous sequences in protein encoding messenger RNAs and suppressing translation and inducing message instability 8,9. miRNAs play critical roles in developmental gene regulation. They are essential in the earliest stages of development, as embryos that cannot produce miRNAs die at a very early stage 10. In addition miRNAs are important later on in the development of many tissues. They are thought to function by fine-tuning the expression of genes required for developmental programs 1. In helper T cells miRNAs play multiple roles and are required for regulatory T cell (Treg) development 11-14. We used retroviral transduction as a means to dissect the mechanisms of miRNA regulation of Treg differentiation 15. Through such studies important individual miRNAs were determined by retroviral-mediated overexpression. Subsequently, relevant genes regulated by these miRNAs were identified in order to understand the molecular pathways regulated by miRNAs in helper T cell differentiation.
研究方案
在这些协议进行的所有小鼠的工作被按照动物项目许可六千九百六十五分之七十零下动物科学程序法,英国进行的。
1.逆转录病毒生产
在此之前继续获得所有必要的批准生产转基因生物和在哺乳动物细胞中使用逆转录病毒。
- HEK 293T细胞生长
- 上生长的HEK 293T介质10厘米组织培养皿的HEK 293T细胞(含10%胎牛血清的Dulbecco氏改良的Eagle氏培养基(DMEM)(FBS),100单位/ ml青霉素0.1mg / ml的链霉素和2mM L-谷氨酰胺) 。对于一般细胞传代,拆分单元格1:10,当他们通过去除中,吹打关闭单元的新鲜HEK 293T介质板达到汇合。
注意:细胞应在2-3天达到汇合。 293T HEK细胞松散附着到这样的胰蛋白酶消化不需要板。 - 24小时前转,拆分conflue细胞1:10至10cm平板(每个转染一个板),使得在第二天,细胞是〜50%汇合的核苷酸板。
- 上生长的HEK 293T介质10厘米组织培养皿的HEK 293T细胞(含10%胎牛血清的Dulbecco氏改良的Eagle氏培养基(DMEM)(FBS),100单位/ ml青霉素0.1mg / ml的链霉素和2mM L-谷氨酰胺) 。对于一般细胞传代,拆分单元格1:10,当他们通过去除中,吹打关闭单元的新鲜HEK 293T介质板达到汇合。
- 通过转染磷酸钙沉淀
- 转染前约1小时,从细胞中取出,小心可加9毫升新鲜HEK 293T介质,以防止细胞的分离板。
- 制备2倍的Hepes缓冲盐水(2×HBS)和一个新鲜的2.5M的CaCl 2溶液,如表1所述。解冻的DNA股。为了最有效地使用转染质粒制备高质量的DNA。
- 在6毫升圆底或15毫升锥形管中添加5微克的逆转录病毒的DNA(pMIG)4,辅助病毒DNA(PCL-ECO)7 5微克,和H 2 O至420微升。添加80微升的2.5M的CaCl 2使终体积为500微升。
注:此下一步骤(另外2个HBS)的沿可在常规替补,只要做一般韦伯斯特使用ILE技术。 - 慢慢逐滴加入500μl2×HBS下降到上述DNA混合物中,同时轻轻地涡旋以使溶液连续地混合,该课4在偶数大小的晶体沉淀。
- 转移到组织培养罩和缓慢的CAPO 4 DNA混合物(1毫升)逐滴添加到9毫升培养基覆盖细胞,同时不断地和轻轻摇动板。再次要注意不要从板块分离细胞。放置细胞放回培养箱中。
注意:此时该课4沉淀物将是一个显微镜对细菌的尺寸小晶体下容易看见。当该晶体有机会沉降到板的底部,这些很容易被后30-60分钟看到。
- 培养上清病毒的集合。
- 翌日(从转染后12-24小时的任何地方)除去培养基并用10毫升喂细胞新鲜HEK 293T中再次小心不要从板块打跑细胞。
注意:这冲淡了该HEK 293T细胞分泌到转染在媒体淋巴细胞抑制物质。 - 在第二天(〜24小时后转染)晚上喂3.5毫升新鲜HEK 293T中的细胞。请注意,该板置于水平究竟在孵化器,使一方不剩干燥。
- 收集中等〜12小时后(在4℃存储)和饲料3.5毫升新鲜HEK 293T介质。重复收集2次以上在大致使约10ml培养基的收集12小时的时间间隔。这是在病毒原液。
- 通过在600 xg离心离心5分钟旋出任何残余的HEK 293T细胞和细胞碎片。可替代地,过滤用0.45微米,低蛋白质结合,注射器过滤器。商店病毒在4℃,最佳滴度,用一两天之内,因为滴度会慢慢在4℃减弱。不要酒店frEEZE,因为这显著降低滴度。
- 翌日(从转染后12-24小时的任何地方)除去培养基并用10毫升喂细胞新鲜HEK 293T中再次小心不要从板块打跑细胞。
2.主朴素的 CD4 + T细胞的分离
- 白细胞细胞的分离
- 安乐死颈椎脱位,CO 2窒息,或适合于机构的动物处理规则等人性化协议老鼠。
- 解剖新鲜安乐死的小鼠并取出脾脏和所需的淋巴结15。放置器官中含有R10培养基(含10%热灭活的FBS,100单位/ ml青霉素,0.1mg / ml的链霉素,2mM的L-谷氨酰胺和0.1%β巯基乙醇的RPMI培养基)6-孔组织培养板的孔中。使用细胞培养罩老鼠和所有后续操作的解剖,以尽量减少在培养污染的机会。
注:保持R10培养基冷并在4℃下执行所有的下游纯化步骤。 - 将70微米的细胞培养过滤成50毫升含有约5mL R10培养基的锥形管中。使用最多三个小鼠的脾和淋巴结节点的一个过滤器。
- 添加脾和淋巴结的细胞过滤并用5ml的针筒柱塞的端部浸渍。用1-2毫升R10介质的冲洗。
- 细胞转移到15毫升锥形管,并把音量高达14毫升R10。离心机在600×g离心在4℃下5分钟。通过一个干净的运动浇,以防止干扰细胞沉淀弃上清。
- 2毫升冷(4℃)红细胞裂解缓冲液( 表1)添加到每个管中。轻轻混合3.5分钟,保持管冰。
注:红细胞裂解的时间是关键的,以防止淋巴细胞死亡。因此,红细胞裂解缓冲液的每批的精确定时将需要进行优化。 - 加入≈12-13毫升R10介质。立即离心弃上清如步骤2.1.5。执行洗涤步骤2倍多才子ħR10培养基从细胞中除去任何残余的红细胞裂解缓冲液,并避免淋巴细胞死亡。
- 计数血球在锥虫蓝稀释1:20,确定活细胞的细胞产量。期望每只小鼠大约8-10×10 7个细胞。
- 利用磁珠套件CD4 + T细胞的分离删除CD8 +细胞CD11b +,CD16 / 32 +,+ CD45R,MHC II +类和泰尔-119 +细胞。
- 等分试样8-10×10 7细胞从步骤2.1.8每的15毫升锥形管中并在600×g离心5分钟离心在4℃下。倒出与一种清洁运动上清液,以防止干扰细胞沉淀。悬浮细胞在100微升的热灭活的FBS,然后从该试剂盒中添加100μl的抗体混合物的每管中。带在辊温和混合孵育30分钟,在4℃。
- 虽然上述细胞培养,准备珠。
- 在小瓶中通过涡旋> 30秒重悬珠子然后用1毫升每8-10×10 7制备的细胞珠转移到15毫升锥形管中。加入2毫升R10中每毫升的珠子,轻轻混匀。
- 放置管在磁体1分钟,然后弃上清。从磁体和重悬珠子在4毫升R10培养基中除去管。珠这一数额足以两轮孵化。
- 通过加入10毫升,每管R10培养基洗细胞在抗体孵育(步骤2.2.1)的端部。轻轻地用管和离心细胞回旋在600 xg离心在4℃下充分混合5分钟。倒出与一种清洁运动上清液,以防止干扰细胞沉淀。悬浮细胞在1ml R10培养基中。
- 从步骤2.2.2.2(½每个管中制备的量),以抗体处理的细胞的每个管中加入2毫升的洗涤珠子,并在4℃下与一ロ温和混合孵育30分钟米勒。
- 通过用含有一个窄尖端开口吸管轻轻吹打5倍重悬细胞 - 珠混合物。避免起泡。放置管中的珠磁铁为2分钟,然后载负选择的细胞的上清转移到新管中。保持这些细胞,因为它们是CD4 +群体。
- 重复阴性选择一次。旋转细胞从步骤2.2.5在600 xg离心在4℃下5分钟。倒掉上清如步骤2.2.3重悬在1毫升R10介质。按照步骤2.2.4和2.2.5一次。最终磁珠选择之后,计数细胞,以确定恢复(典型产率是每10 8白细胞15-20×10 6个细胞)。
- 使用细胞分离柱-朴素的 CD4 + T细胞(和CD62L 高 CD25)的分离
- 离心机CD4 + T细胞在600×g离心5分钟,在4℃,重悬在150-200微升每10 8个细胞R10网上平台。加入2微升股票生物素的CD25单克隆抗体(克隆7D4)的。带在辊温和混合孵育30分钟,在4℃。
- 通过加入10ml的细胞分离柱缓冲液( 表1)洗涤细胞。离心机在600×g离心在4℃下5分钟。除去尽可能多的上清液尽可能并估计剩余缓冲器/细胞的体积。悬浮到每10 7个细胞大约90微升的最终体积。
- 加入20微升每10 7个细胞链亲和素珠和温柔的笔触混合。孵育在4℃下15分钟。
- 当细胞培养,制备培养基细胞分离(MS)列手动方法。每个MS柱保留10 7个细胞,并自CD25 +细胞通常代表总CD4 +细胞的约10%,每10 8 CD4 + T细胞使用1列。加入500微升细胞分离柱缓冲液向柱让流过。重复2次以上。
- 在细胞/珠温育结束时,通过加入10ml的细胞分离的柱缓冲液中,并在4℃以600 xg离心离心5分钟洗涤细胞。重悬在500微升每10 8个细胞的细胞分离柱缓冲液的细胞,但仍使用500微升,如果有较少的细胞。
- 应用500微升细胞/珠悬液到柱,并收集通过的流动。通过这个在第1列更多的时间,并通过再次收集流程。冲洗柱用500μl细胞分离柱缓冲液3次,将每个从这些漂洗流经收集到的细胞中。这些是CD25 -细胞。计数细胞,以确定产量。
- 如果调节性T细胞(CD25 +细胞)被需要的话,隔离如下。
- 从分离器中取出柱,守住一个开放的通用管照顾它保持无菌。快速吸取500微升细胞分离缓冲到山坳柱,并牢固地冲洗出阳性部分,使用与列提供的柱塞。
- 重复冲洗步骤2次以上。越过一个新的MS细胞分离柱的阳性率和丢弃此流过。牢牢冲洗出阳性细胞三次,如前离心细胞以600×g离心5分钟,在4°C重悬细胞于1ml R10培养基中并计数,以确定产率。
- 为了获得CD25 - CD62L 高的T细胞,离心后CD25 -上面的步骤2.3.6在600 XG在150-200微升每10 7细胞R10介质中分离出5分钟,在4°重悬T细胞。添加5微升的库存的生物素化的CD62L单克隆抗体(克隆MEL-14)和如前孵育在4℃下温和混合在辊30分钟。
- 执行洗涤,链霉珠结合,和细胞分离柱制备作为用于调节性T细胞分离方案(2.3.7)中所述。这次使用大细胞分离(LS)柱,其具有10 8个细胞的容量。
- 由于CD62L 高的T细胞通常表示CD25的约60-70% -细胞,利用每10 8个细胞1 LS细胞分离柱。细胞/珠的混合物添加到LS细胞分离柱如步骤2.3.6描述 - 这时候放弃最后流过和列冲洗。收集CD62L 高 T细胞在步骤2.3.7为CD25 +细胞采集描述。
- 离心细胞以在1毫升R10 600×g离心5分钟,在4°C悬浮细胞并计数,以确定产率。稀释至0.75-1 X10每毫升6细胞R10网上平台。
- 通过荧光激活细胞分选(FACS)分析细胞纯度。
- FACS染色策略
- 对于细胞前后的孤立的各个阶段,污渍与CD4-FITC,CD8A-PerCP-经Cy5.5(辅助性和细胞毒性T细胞),并MHCIⅠ-PE(MHC II类表达细胞)。
- 对于细胞前后的CD25隔离,染色与CD4-FITC和CD25-PE(调节性T细胞与其他辅助性T细胞)。
- 对于细胞前,后CD62L隔离带污迹的CD4-FITC,CD62L-PE和CD44-APC(天真与内存和效应辅助性T细胞)。
- 对于每个分析地点10 5个细胞在96-孔U型底的板的孔中。离心机在600×g离心在4℃下5分钟。倒出培养基,并用200μlDPBS洗一次细胞。如上离心倒出DPBS。
- 添加每孔的DPBS 50和0.5μl各抗体(如2.4.1所示)的孵育在室温下进行30分钟,在黑暗中以避免荧光标记的漂白。
- 洗涤细胞2倍用DPBS如在2.4.2和立即分析关于流式细胞仪使用特定于手头的仪器的协议和准则的流程。
注:做好准备的最后分离步骤后,细胞的90-95%将是CD4 + CD25 - CD62L 高的T细胞。
- FACS染色策略
3.逆转录病毒激活CD4 + T细胞转导并分化为特定的T辅助亚群
- 逆转录病毒转导
注:逆转录病毒整合和基因的表达需要细胞分裂。因此,T细胞必须被激活O / N。- 而分离幼稚T细胞,外套24孔组织培养板用抗CD3和在DPBS稀释的抗CD28抗体。加入每孔250微升。在1微克/毫升使用防抗CD3。使用抗CD28在2微克/毫升,如果细胞将最终的Th0,Th1细胞,Th2和iTregs的条件下加以区分。使用抗CD28以10微克/毫升为TH9和Th17的条件。孵育在37℃〜2小时在组织培养箱。
- 培养所述细胞之前,为了,除去抗体溶液并用〜250微升每孔的DPBS除去unbou的洗涤板ND抗体。小心不要让板干出这么一次最多6口井的处理。除去DPBS洗,并在每毫升0.75-1×10 6细胞加入1毫升幼稚的 CD4 + T细胞在R10介质。活化细胞O / N(14-16小时)。
- 第二天,离心细胞,在900×g离心5分钟,该板在30℃下附着的细胞至孔的底部。
- 收集并保存小心不要置换细胞的培养基,并使用从病毒生产方案制备1毫升病毒培养上清液更换。不要让细胞干出这么一次最多4口井处理。
- 向每个孔中加入8毫克/毫升聚凝胺的1微升和10μl1M HEPES pH为7.5的以有助于病毒的摄取,并防止下面的离心分离期间在环境CO 2显著碱化。离心细胞,在900×g离心90分钟板在30℃。
- 细胞向规格分化FIC子集
- 小心地取出病毒培养物上清液并用1ml的培养基中步骤3.1.4上述收集替换,因为它包含的IL-2和其它的T细胞生长因子。加入表2和培养细胞表示3-4天分化细胞分化成特定子集试剂。
- 细胞分析
- 对于细胞因子的细胞内染色,处理细胞用1μg/ ml的每种的佛波醇12-肉豆蔻酸酯13-乙酸酯(PMA)和离子霉素为4小时。在刺激后2小时,加入1μg/ ml的布雷菲德菌素A的
- 从每个孔的底部悬浮细胞通过上下吹打数次。输送约10 5个细胞在96孔U底板(通常为100微升从1ml的Th细胞的培养物),用于固定和染色。
- 对于GFP染色,固定用100微升2%多聚甲醛的细胞在室温整整5分钟,因为一个较长的培育时间将漂一GFPND与Foxp3的染色干扰。通过在22℃在600 xg离心加入100微升DPBS的每个孔中,离心5分钟洗涤细胞。倒出上清液。
注意:多聚甲醛是有毒的。处理它与皮肤和眼睛的保护在通风橱。 - 固定再次使用从所述的Foxp3染色试剂盒(1体积固定缓冲液至3体积的稀释剂)由100微升1×固定缓冲液的细胞。孵育45分钟的细胞在室温或4℃备选地孵育16小时。
- 固定后,通过从相同的试剂盒,加入100微升透化缓冲液的通透细胞。孵育在室温30-45分钟,然后在600 xg离心22℃离心5分钟。
- 对于抗体染色,除去固定/通透缓冲液,加入50微升清新通透缓冲。加入通透缓冲液稀释所需的抗体在每个样品每50微升0.5微升缓冲。在黑暗中孵育30-45分钟,在室温,以避免感漂白orescent标签。
- 在600 XG添加150微升透化缓冲液和离心机在22℃5分钟。丢弃通透缓冲区,并添加200微升DPBS。流式细胞仪上使用特定手头的仪器协议和指导方针流量分析。
注意:一定要包括细胞因子染色适当控制,例如同型对照抗体染色,分析非激活或激活的Th0细胞等
结果
本实验系统的成功需要T细胞和高滴度逆转录病毒制剂的高纯度的人群。代表性的结果在这里显示为成功的实验的实例。 图1示出了在幼稚辅助性T细胞的分离协议的各个阶段前后选定人群的典型纯度。 图2和3说明反转录病毒生产的分析,通过GFP表达在转染的HEK 293T细胞( 图2)和转导的T细胞( 图3)。在HEK 293T细胞的转染效率可以用不同的逆转录病毒构建显著变化,但这常常不与用GFP + T细胞的数量观察逆转录病毒生产水平相关。此外,GFP + T细胞的数量可以根据偏振条件而变化。此外,m个的GFP EAN表达水平和插入的基因可以根据集成病毒拷贝的数目而变化,上转录的整合位点的影响,以及影响病毒转录物的转录后调节机制。
最后, 图4示出了我们已经与辅助性T细胞的分化时观察到的miRNA的的miR-15b中/ 16过表达的一些典型结果。这些结果表明,一些可以个别实验所以真正的效果,必须通过使用辅助T细胞的不同制剂的多个重复试验的统计分析得到证实内发生的变异性。在这些实验中Th2应答可能难以在这里使用,因为它们很容易Th1应答的C57BL / 6行观察。同样地,IL-9染色可能难以检测高于背景。因此,当务之急是要做到同型对照并设置适当的补偿,以确保正确嘎细胞因子表达婷。在我们的结果,我们发现miR-15B / 16通过抑制mTOR的通过抑制部件Rictor的和mTOR的15的表达信号通路增强iTreg诱导。的miR-15b中/ 16有时可以影响的Th0,Th1细胞,和Th17分化在个别实验,但在多个重复试验检查时,没有显著效果。与此相反的miR-15B / 16表达不显著抑制TH9分化(见参考文献18)。

图在隔离的每个阶段辅助性T细胞的1.典型纯度。代表性流式细胞仪所指示的抗原的结果是从在前向散射(FSC)和侧向角散射(SSC)图指定活细胞的栅极所示。 ( 一 )前和后的CD4阴性选择。 CD4的表达谱,CD8A,和MHCII被示出。这些说明辅助性T细胞的富集和细胞毒性T细胞的损失和II类MHC表达细胞。一个好的净化应导致〜在此阶段90% 的 CD4 + T细胞。 (B)CD25选择。在左边是CD4,CD8A,和MHCII的表达谱,和在右边是CD4和CD25的表达概况前后选择。在CD25负选定单元格的这点"95%应该是CD4 + CD25 - 。 (C)CD62L选择。 CD4,CD8A,和MHCII表达谱显示在左侧。在右边显示与选择后的细胞CD4和CD62L表达谱沿着前,后CD62L选择细胞CD62L和CD44的表达谱。 CD62L选择之后几乎所有的存储单元(CD44 +)的除去,留下包含10-15%的效应细胞(CD62L 低 )幼稚辅助性T细胞的高度富集的群体。对全部FACS型材,相当于设置和尺度特定参数整个维持。数字代表细胞的门控群体中的百分比。在初始选择后的细胞大小略有减少,大概是由于该协议中的机械应力。 请点击此处查看该图的放大版本。
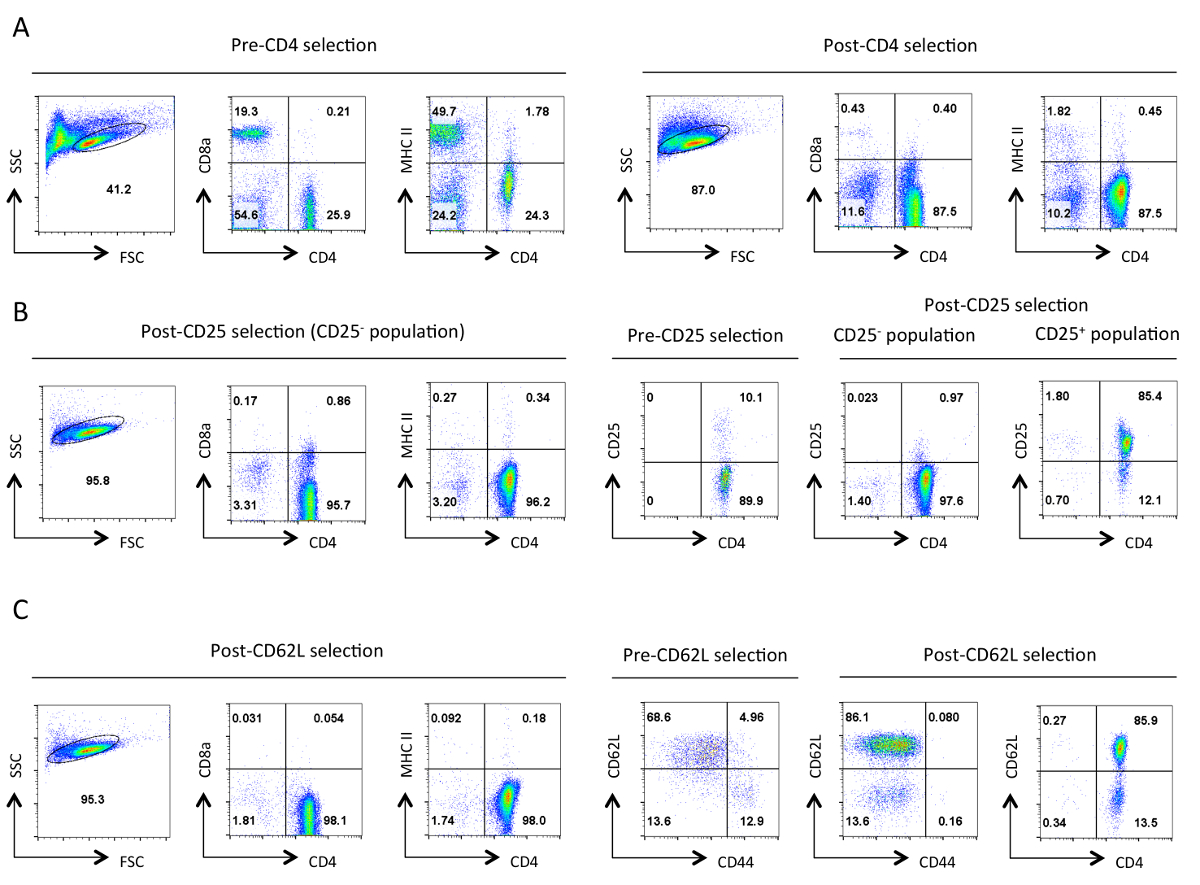{kind=link}

图2.逆转录病毒转染的HEK 293T细胞的分析。GFP表达显示在该要么未转染或转染和病毒培养物上清液的采集后分析的HEK 293T细胞。的GFP分析从FSC的栅极和SSC情节在所述第一面板上的活细胞进行。数字代表的GFP +细胞的选通区域内的百分比。典型Ťransfection效率介于30-90%之间。 请点击此处查看该图的放大版本。
{kind=link}

图3.逆转录病毒转导辅助性T细胞的分析。GFP表达示于逆转录病毒转导的辅助性T细胞中的Th0,Th1细胞,Th2细胞,TH9,Th17的分化后,和三天的Treg偏振条件。分析选通在FSC / SSC面板显示实况和活化细胞。转导效率可以根据构建体和偏振条件10-75%之间变化。同样,GFP表达的平均荧光强度会有所不同。 请点击此处查看该图的放大版本。
{kind=link}

图4. /效果的miR-15b的表达16在不同的极化条件下辅助性T细胞的分化。代表性的细胞因子谱显示在GFP +从图3的细胞群。 请点击此处查看该图的放大版本。
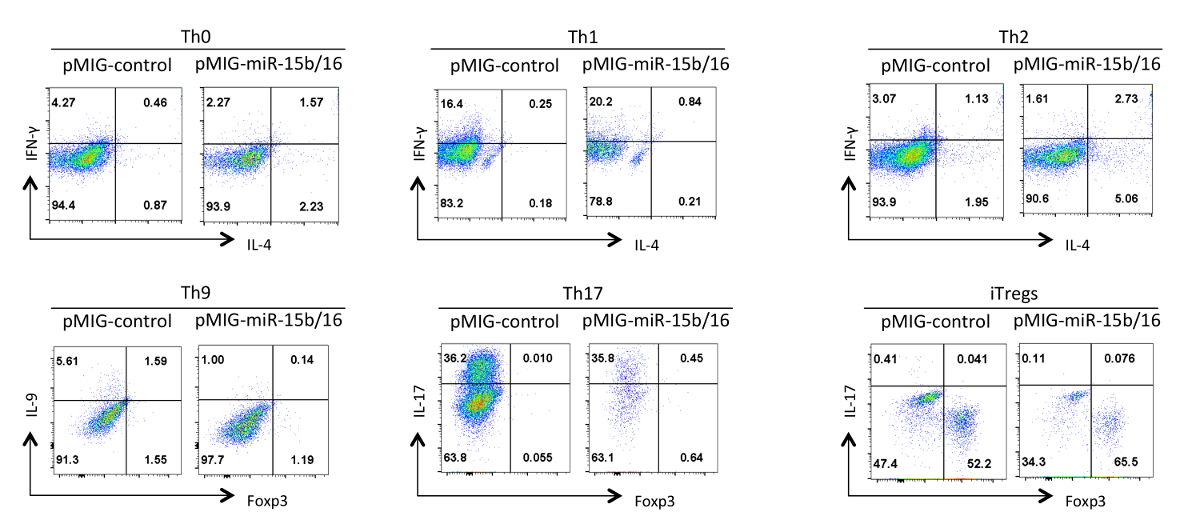{kind=link}

表 1: 在这些协议中使用的缓冲液 ,请点击此处下载此表为Excel电子表格。
| 帮手T细胞极化条件 | ||
| TH0 | 抗IL-4 | 5微克/毫升 |
| 抗IFN-γ的 | 5微克/毫升 | |
| TH1 | 重组IL-12 | 20纳克/毫升 |
| 抗IL-4 | 5微克/毫升 | |
| Th2细胞 | 重组IL-4 | 40纳克/毫升 |
| 抗IFN-γ的 | 5微克/毫升 | |
| TH9 | 重组型TGF-β | 2.5纳克/毫升 |
| 重组IL-4 | 40纳克/毫升 | |
| 抗IFN-γ的 | 10微克/毫升 | |
| TH17 | 重组型TGF-β | 2.5纳克/毫升 |
| 重组IL-6 | 50纳克/毫升 | |
| 抗IFN-γ的 | 5微克/毫升 | |
| 抗IL-4 | 5微克/毫升 | |
| 抗IL-2 | 5微克/毫升 | |
| 调节性T细胞 | 重组型TGF-β | 2.5纳克/毫升 |
| 重组IL-2 | 5毫微克/毫升 | |
表2: 辅助性T细胞亚群的极化条件。
讨论
基因的逆转录病毒介导的过表达是分析在辅助性T细胞功能的有力方式,因为它们的发育和功能通常是由关键调节的表达水平来确定。然而,结果的谨慎的解释是必要的,因为表达水平显著高于内源基因的引入可许多文物。因此,这种技术应该与其他组合以验证的函数的相关性。例如,表达应使用的siRNA或基因敲除(如果可用)降低表达的补充。随着miRNA的,我们通过使用过表达人工miRNA目标是充当竞争性抑制剂的miRNA的15个站点的病毒结合,与阻断过度的实验。逆转录病毒转导的细胞,也可在涉及RNA和蛋白质分析生物化学分析使用。然而,这些实验的一个主要限制是转导资源的效率ulting在转导和未转导的细胞的混合群。因此,这些试验将最有可能需要将GFP +群体的排序。最后, 在体外分化测定法应在体内实验相结合,这是可以实现的一种方式是由导的T细胞过继转移入小鼠和以下它们的分化和它们对免疫反应的影响。
一的该系统的主要限制是,可以打包成逆转录病毒衣壳的RNA基因组的大小。根据我们的经验,为MIG逆转录病毒系统的最大插入尺寸提供了良好的病毒生产3-3.5 KB。因此,更大的基因不能与本系统进行分析,因为它们给差病毒滴度。然而,所以该系统是用于各种基因研究的有用大多数基因比该尺寸更小。
与逆转录病毒转导,这些protoco内的几个备选方案LS已经使用。许多研究者已经利用稳定表达逆转录病毒的基因(例如参照图16)的包装细胞系。然而,我们已获得使用标准的HEK 293T细胞与PCL-生态辅助病毒载体的共转染的最高滴度。幼稚辅助性T细胞的分离,也可以通过细胞分选,而不是磁珠与细胞分离柱协议来实现,但这需要访问一个细胞分选仪,和用于排序时间的费用通常比珠试剂高。最后,还有上用于区分的辅助T细胞分化成不同子集的激活条件的变化。例如,细胞太久的TCR刺激暴露在调节性T细胞诱导条件才能抑制其诱导16。因为逆转录病毒表达需要通过细胞的刺激诱导的细胞分裂这可能是一个问题。然而,我们发现使用此协议与O / N ACTIV高效的调节性T细胞的诱导通货膨胀之前的逆转录病毒转导。
内的这些协议,成功的应用需要几个因素。高滴度逆转录病毒制剂需要的HEK 293T细胞所以高质量DNA并精确地制备2×HBS的有效转染是重要的。此外,HEK 293T细胞的细胞密度需要是在转染的点大约50%,因为转染DNA的良好表达需要将细胞活跃生长的,这将被抑制,如果所述细胞是过于稀疏或密集。在转染过程中最佳密度细胞应在在病毒收集步骤某一点达到汇合,但他们将继续一路过关斩将到最后一集生产高滴度的病毒股票。辅助性T细胞的有效分化需要良好的小区质量,以便确保分离的细胞是在图1所示的纯度。同样地,小区的质量取决于小鼠FROM他们被隔离。对于这些研究,我们使用6-8周龄C57BL / 6小鼠。老年小鼠可具有少幼稚细胞,以及其他菌株可以在其分化不同。例如,如上所述,C57BL / 6的T细胞可能难以诱导Th2应答BALB / c小鼠更容易比C57BL / 6小鼠17这样的Th2应答。此外,任何的分化条件可从实验室稍有变化,以实验室,和基因表达的效果可能仅成为在亚最佳条件表观所以在各偏振条件的细胞因子的浓度可能需要被滴定。最后,对细胞增殖的过表达的基因的作用或偏振条件可以影响转导效率的目的基因的,以便测量效果可能需要优化偏振试剂的定时和浓度。优化所有这些因素应导致该系统信息的结果。
披露声明
The authors have no conflicting interest in the publication of this work.
致谢
This work was supported by a Biotechnology and Biological Sciences Research Council (BBSRC) grant (BB/H018573/1) and a BD Biosciences grant.
材料
| Name | Company | Catalog Number | Comments |
| RPMI | Sigma | R8758 | |
| DMEM | Sigma | D5671 | |
| Penicillin Streptomycin solution | Sigma | P4333 | |
| L-Glutamine | Sigma | G7513 | |
| β-mercaptoethanol | Sigma | M3148 | |
| DPBS | Sigma | D8537 | |
| MIG vector | Addgene | Plasmid 9094 | |
| pCL-Eco vector | Addgene | Plasmid 12371 | |
| Cell strainer | BD Falcon | 352350 | |
| Magnetic beads mouse CD4 cell kit | Invitrogen (Dynabeads) | 11415D | |
| Streptavidin Beads | Miltenyi Biotech | 130-048-102 | |
| MS cell separation columns | Miltenyi Biotech | 130-042-201 | |
| LS cell separation columns | Miltenyi Biotech | 130-042-401 | |
| CD25 Biotenylated MAb | BD Biosciences | 85059 | clone 7D4 |
| CD62L Biotenylated MAb | BD Biosciences | 553149 | clone MEL-14 |
| Polybrene (Hexadimethrine Bromide) | Sigma | 107689 | |
| Anti-CD3 | eBiosciences | 16-0031-85 | clone 145-2C11 |
| Anti-CD28 | eBiosciences | 16-0281-85 | clone 37.51 |
| Anti-IL-4 | BD Biosciences | 559062 | clone 11B11 |
| Anti-IFN-gamma | BD Biosciences | 559065 | clone XMG1.2 |
| Anti-IL-2 | BD Biosciences | 554425 | cloneJES6-5H4 |
| Recombinant IL-12 p70 | eBiosciences | 14-8121 | |
| Recombinant IL-4 | BD Biosciences | 550067 | |
| Recombinant TGF-beta | eBiosciences | 14-8342-62 | |
| Recombinant IL-6 | eBiosciences | 14-8061 | |
| Recombinant IL-2 | eBiosciences | 14-8021 | |
| PMA | Sigma | P8139 | |
| Ionomycin | Sigma | I0634 | |
| Brefeldin A | eBiosciences | 00-4506 | |
| Paraformaldehyde | Sigma | 16005 | Paraformaldehyde is toxic so use appropriate caution when handling |
| Foxp3 staining buffer set | eBiosciences | 00-5523 | |
| Anti-CD4 FITC | eBiosciences | 11-0041 | clone GK1.5 |
| Anti-CD8a perCP-cy5.5 | eBiosciences | 45-0081-80 | clone 53-6.7 |
| Anti-MHCII PE | eBiosciences | 12-0920 | clone HIS19 |
| Anti-CD25 PE | eBiosciences | 12-0251-82 | clone PC61.5 |
| Anti-CD62L PE | eBiosciences | 12-0621-82 | clone MEL-14 |
| Anti-CD44 APC | eBiosciences | 17-0441 | clone IM7 |
| Anti-IFN-gamma FITC | eBiosciences | 11-7311-81 | clone XMG1.2 |
| Anti-IL-4 PE | BD Biosciences | 554435 | clone 11B11 |
| Anti-IL-9 PE or APC | eBiosciences/Biolegend | 50-8091-82/514104 | clone RM9A4 |
| Anti-IL-17a PE | BD Biosciences | 559502 | clone TC11-18H10 |
| Anti-Foxp3 APC or PE | eBiosciences | 17-5773-82/12-5773-80 | clone FJK-16s |
| NaCl | Sigma | S7653 | |
| KCl | Sigma | P9333 | |
| Na2HPO4-2H2O | Sigma | 71643 | |
| Dextrose/Glucose | Sigma | G7021 | |
| HEPES, free acid | Sigma | H3375 | |
| NH4Cl | Sigma | A9434 | |
| Disodium EDTA | Sigma | D2900000 | |
| KHCO3 | Sigma | 237205 | |
| CaCl2 | Sigma | C5670 |
参考文献
- Baumjohann, D., Ansel, K. M. MicroRNA-mediated regulation of T helper cell differentiation and plasticity. Nat Rev Immunol. 13 (9), 666-678 (2013).
- Wilson, C. B., Rowell, E., Sekimata, M. Epigenetic control of T-helper-cell differentiation. Nat Rev Immunol. 9 (2), 91-105 (2009).
- Pearce, E. L., Poffenberger, M. C., Chang, C. H., Jones, R. G. Fueling immunity: insights into metabolism and lymphocyte function. Science. 342 (6155), 1242454 (2013).
- Cherry, S. R., Biniszkiewicz, D., Van Parijs, L., Baltimore, D., Jaenisch, R. Retroviral Expression in Embryonic Stem Cells and Hematopoietic Stem Cells. Mol and Cell Biol. 20 (20), 7419-7426 (2000).
- Miller, A. D., Rosman, G. J. Improved retroviral vectors for gene transfer and expression. BioTechniques. 7 (9), 980-990 (1989).
- Grez, M., Akgün, E., Hilberg, F., Ostertag, W. Embryonic stem cell virus, a recombinant murine retrovirus with expression in embryonic stem cells. Proc Nat Acad Sci USA. 87 (23), 9202-9206 (1990).
- Naviaux, R. K., Costanzi, E., Haas, M., Verma, I. M. The pCL vector system: rapid production of helper-free, high-titer, recombinant retroviruses. J Virol. 70 (8), 5701-5705 (1996).
- Filipowicz, W., Bhattacharyya, S. N., Sonenberg, N. Mechanisms of post-transcriptional regulation by microRNAs: are the answers in sight?. Nat Rev Genetics. 9 (2), 102-114 (2008).
- He, L., Hannon, G. J. MicroRNAs: small RNAs with a big role in gene regulation. Nat Rev Genetics. 5 (7), 522-531 (2004).
- Bernstein, E., Kim, S. Y., et al. Dicer is essential for mouse development. Nat Genetics. 35 (3), 215-217 (2003).
- Cobb, B. S., Hertweck, A., et al. A role for Dicer in immune regulation. J Exp Med. 203 (11), 2519-2527 (2006).
- Chong, M. M. W., Rasmussen, J. P., Rudensky, A. Y., Rundensky, A. Y., Littman, D. R. The RNAseIII enzyme Drosha is critical in T cells for preventing lethal inflammatory disease. J Exp Med. 205 (9), 2005-2017 (2008).
- Liston, A., Lu, L. F., O'Carroll, D., Tarakhovsky, A., Rudensky, A. Y. Dicer-dependent microRNA pathway safeguards regulatory T cell function. J Exp Med. 205 (9), 1993-2004 (2008).
- Zhou, X., Jeker, L. T., et al. Selective miRNA disruption in T reg cells leads to uncontrolled autoimmunity. J Exp Med. 205 (9), 1983-1991 (2008).
- Singh, Y., Garden, O. A., Lang, F., Cobb, B. S. MicroRNA-15b/16 Enhances the Induction of Regulatory T Cells by Regulating the Expression of Rictor and mTOR. J Immunol. 195 (12), 5667-5677 (2015).
- Sauer, S., Bruno, L., et al. T cell receptor signaling controls Foxp3 expression via PI3K, Akt, and mTOR. Proc Nat Acad Sci USA. 105 (22), 7797-7802 (2008).
- Yagi, J., Arimura, Y., Takatori, H., Nakajima, H., Iwamoto, I., Uchiyama, T. Genetic background influences Th cell differentiation by controlling the capacity for IL-2-induced IL-4 production by naive CD4+ T cells. Int Immunol. 18 (12), 1681-1690 (2006).
- Singh, Y., Garden, O. A., Lang, F., Cobb, B. S. MicroRNAs regulate T-cell production of interleukin-9 and identify hypoxia-inducible factor-2a as an important regulator of T helper 9 and regulatory T-cell differentiation. Immunology. 149, 74-86 (2016).
转载和许可
请求许可使用此 JoVE 文章的文本或图形
请求许可探索更多文章
This article has been published
Video Coming Soon
版权所属 © 2025 MyJoVE 公司版权所有,本公司不涉及任何医疗业务和医疗服务。