
Method Article
Ретровирусное трансдукция Т-хелперы как генетический подход к изучению механизмов, контролирующих их дифференцировку и функции
В этой статье
Резюме
Многие экспериментальные системы были использованы, чтобы понять механизмы, регулирующие развитие и функцию Т-клеток в иммунном ответе. Здесь генетический подход с использованием ретровирусной трансдукции описывается, что экономические, время эффективно, а самое главное, весьма информативным в выявлении регуляторных путей.
Аннотация
Helper T cell development and function must be tightly regulated to induce an appropriate immune response that eliminates specific pathogens yet prevents autoimmunity. Many approaches involving different model organisms have been utilized to understand the mechanisms controlling helper T cell development and function. However, studies using mouse models have proven to be highly informative due to the availability of genetic, cellular, and biochemical systems. One genetic approach in mice used by many labs involves retroviral transduction of primary helper T cells. This is a powerful approach due to its relative ease, making it accessible to almost any laboratory with basic skills in molecular biology and immunology. Therefore, multiple genes in wild type or mutant forms can readily be tested for function in helper T cells to understand their importance and mechanisms of action. We have optimized this approach and describe here the protocols for production of high titer retroviruses, isolation of primary murine helper T cells, and their transduction by retroviruses and differentiation toward the different helper subsets. Finally, the use of this approach is described in uncovering mechanisms utilized by microRNAs (miRNAs) to regulate pathways controlling helper T cell development and function.
Введение
The immune response must be highly regulated to eliminate infections but prevent attacks on self-tissue that lead to autoimmunity. Helper T cells play an essential role in regulating the immune response, and a great deal of effort has been undertaken to understand their development and function (illustrated in several recent reviews 1-3). However, many questions remain, and many approaches have been utilized to study the mechanisms controlling helper T cell development and function. These have ranged from the use of in vitro cell culture systems to whole animals. Cell culture systems, especially those using cell lines, offer the benefit of ease of use and the ability to generate large amount of material to do sophisticated biochemical analyses. However, they suffer from their limited ability to reproduce the actual conditions occurring in an immune response. In contrast, whole animal experiments offer the benefit of relevance, but they can suffer from difficulties in manipulation and the ability to perform precise controls in addition to their large costs and ethical implications. Nevertheless, the vast majority of helper T cells studies today still require the use of whole animal experiments involving primary T cells because of the inability of cell lines to duplicate the exact steps occurring in the whole animal. Therefore, it is essential to utilize cost effective approaches that are highly informative.
Genetics is one powerful tool to study helper T cell development and function, yet traditional methods involving gene knockouts or transgenes are time consuming and expensive so they are often out of reach of small labs. However, retroviral transduction offers a powerful, rapid and, cost effective genetic approach to study the mechanisms of specific gene products. Therefore, it is commonly used in papers studying helper T cell development and function.
We have optimized a procedure for retroviral transduction of helper T cells. It utilizes the pMIG (Murine stem cell virus-Internal ribosomal entry site-Green fluorescent protein) retroviral expression vector, in which the gene of interest can be cloned and thereby expressed from the retrovirus long terminal repeat (LTR) 4. In addition, downstream of the inserted gene of interest is an internal ribosome entry sequence (IRES) followed by the green fluorescent protein (GFP) gene so transduced cells can easily be followed by their expression of GFP. The vector was originally derived from the Murine Stem Cell Virus (MSCV) vectors, which contain mutations in repressor binding sites in the LTRs making them resistant to silencing and thus, giving high expression in many cell types including helper T cells 5,6. Production of high titer retrovirus requires a simple transient transfection protocol of human embryonic kidney (HEK) 293T cells with the MIG vector and a helper virus vector that expresses the retroviral GAG, Pol, and Env genes. For this the pCL-Eco helper virus vector 7 works well in producing high titer replication incompetent retroviruses.
Here these protocols for retroviral production and transduction of primary murine T cells are described in addition to some of our results using this approach to study miRNA regulation of gene expression controlling helper T cell differentiation. miRNAs are small RNAs of approximately 22 nucleotides in length that post-transcriptionally regulate gene expression by targeting homologous sequences in protein encoding messenger RNAs and suppressing translation and inducing message instability 8,9. miRNAs play critical roles in developmental gene regulation. They are essential in the earliest stages of development, as embryos that cannot produce miRNAs die at a very early stage 10. In addition miRNAs are important later on in the development of many tissues. They are thought to function by fine-tuning the expression of genes required for developmental programs 1. In helper T cells miRNAs play multiple roles and are required for regulatory T cell (Treg) development 11-14. We used retroviral transduction as a means to dissect the mechanisms of miRNA regulation of Treg differentiation 15. Through such studies important individual miRNAs were determined by retroviral-mediated overexpression. Subsequently, relevant genes regulated by these miRNAs were identified in order to understand the molecular pathways regulated by miRNAs in helper T cell differentiation.
протокол
Все работы мыши выполнены в этих протоколах была проведена в соответствии с процедурами Животные научных Закона, Великобритании в соответствии с лицензией 70/6965 проекта животного.
1. Производство Ретровирусное
Перед продолжением получения всех необходимых согласований для получения генетически модифицированных организмов и использование ретровирусов в клетках млекопитающих.
- Рост клеток НЕК 293Т
- Grow 293Т HEK на 10 см чашки для культивирования тканей в НЕК 293T среде (среда Игла, модифицированная орла Средний (DMEM) с добавлением 10% фетальной бычьей сыворотки (FBS), 100 ед / мл пенициллина 0,1 мг / мл стрептомицина и 2 мМ L-глутамина) , Для общего прохождения клетки, клетки расщепленные 1:10, когда они достигают слияния путем удаления среды и пипеткой клетки от пластины со свежей средой НЕК 293T.
Примечание: Клетки должны достичь слияния в течение 2-3 дней. 293T НЕК клетки придают неплотно к пластине так трипсинизации не требуется. - 24 ч до трансфекции, разделить conflueнт пластинка клеток от 1:10 до 10 см пластины (одна пластина на трансфекцией), так что на следующий день клетки ~ 50% сплошности.
- Grow 293Т HEK на 10 см чашки для культивирования тканей в НЕК 293T среде (среда Игла, модифицированная орла Средний (DMEM) с добавлением 10% фетальной бычьей сыворотки (FBS), 100 ед / мл пенициллина 0,1 мг / мл стрептомицина и 2 мМ L-глутамина) , Для общего прохождения клетки, клетки расщепленные 1:10, когда они достигают слияния путем удаления среды и пипеткой клетки от пластины со свежей средой НЕК 293T.
- Трансфекция по фосфатом кальция Осадки
- Приблизительно 1 ч до трансфекции удалить клетками среду и осторожно добавляют 9 мл свежей среды НЕК 293T, чтобы предотвратить клетки отвязывания от пластины.
- Готовят 2x Hepes - солевом буферном растворе (2x HBS) и свежий 2,5 М CaCl раствора 2 , как описано в таблице 1. Растаяйте запасы ДНК. Для получения наиболее эффективных трансфекциях используют плазмиду качества Prep ДНК.
- В 6 мл с круглым дном или 15 мл коническую трубку добавляют 5 мкг ДНК ретровирусов (pMIG) 4, 5 мкг ДНК вируса Helper (ПКЛ-Eco) 7 и H 2 O в 420 мкл. Добавляют 80 мкл 2,5 М CaCl 2 , доведя конечный объем до 500 мкл.
Примечание: Это наряду со следующим шагом (добавление 2x HBS) может быть сделано на регулярной скамейке до тех пор, как общая STERIle метод используется. - Медленно добавляют 500 мкл 2x HBS по каплям к вышеуказанной смеси ДНК, осторожно встряхивая таким образом , чтобы раствор непрерывно перемешивают и каподастра 4 осаждается в даже размера кристаллов.
- Перенесите в капот культуры ткани и медленно добавляют смесь ДНК CaPO 4 (1 мл) по каплям 9 мл среды , покрывающих клетки в то время как постоянно и осторожно закрученной тарелку. Еще раз быть осторожным, чтобы не отделить клетки от пластины. Поместите клетки обратно в инкубатор.
Примечание: В этот момент осадок CaPO 4 будет легко видны под микроскопом в виде мелких кристаллов размером с бактериями. Они хорошо видны через 30-60 мин, когда кристаллы имели возможность осесть на дно тарелки.
- Сбор вируса в культуре супернатантов.
- На следующий день (где-то от 12-24 ч после трансфекции) удаления среды и питания клеток с 10 млсвежего НЕК 293T среды еще раз осторожно, чтобы не сместить клетки от пластины.
ПРИМЕЧАНИЕ: Это разбавляет из лимфоцитов ингибирующие вещества, что НЕК 293T клетки секретируют в среду во время трансфекции. - Вечером второго дня (~ 24 часов после трансфекции) кормить клеток с 3,5 мл свежей среды НЕК 293T. Будьте осторожны, чтобы пластина помещается ровно уровень в инкубаторе, так что одна сторона не осталась сухой.
- Сбор среды ~ 12 час позже (хранить при температуре 4 ° С) и кормить с 3,5 мл свежей среды НЕК 293T. Повторите сбор 2 раза больше примерно в 12 интервалов ч так, что собрано около 10 мл среды. Это вирус запас.
- Спин любые остаточные НЕК 293Т и клеточного дебриса центрифугированием при 600 х г в течение 5 мин. В качестве альтернативы, фильтр с использованием 0,45 мкм, низкое связывание с белками, шприцевой фильтр. Вирус Хранить при температуре 4 ° С, а для достижения наилучших титрами, использовать в течение дня или двух, так как титр будет медленно уменьшаться при 4 ° С. Не FrEeze, так как это существенно снижает титр.
- На следующий день (где-то от 12-24 ч после трансфекции) удаления среды и питания клеток с 10 млсвежего НЕК 293T среды еще раз осторожно, чтобы не сместить клетки от пластины.
2. Выделение первичной Наивные CD4 + Т - клеток
- Выделение лейкоцитарной клеток
- Эвтаназии мышей путем смещения шейных позвонков, CO 2 удушья или других гуманного протокола соответствующего правилам обращения с животным учреждения.
- Рассеките свеже эвтаназии мышей и удаления селезенки и лимфатических узлов желательно 15. Поместите органы в лунки 6-луночные тканевого культурального планшета, содержащей среде R10 (RPMI среду с 10% инактивированной нагреванием FBS, 100 ед / мл пенициллина, 0,1 мг / мл стрептомицина, 2 мМ L-глутамина и 0,1% бета-меркаптоэтанол) , Используйте для культивирования клеток вытяжку для рассечения мышей и все последующие манипуляции, чтобы свести к минимуму вероятность загрязнения в культурах.
Примечание: Держите среду R10 холодной и выполнять все ниже по течению стадии очистки при 4 ° С. - Поместите 70 мкм сетчатый фильтр для культивирования клеток в 50 мл коническую пробирку, содержащую приблизительно 5 мл R10 среды. Используйте один сетчатый фильтр для селезенки и лимфатических узлов до трех мышей.
- Добавить селезенки и лимфатических узлов на клеточный фильтр и вымачивать, используя конец поршня шприца 5 мл. Полоскание с 1-2 мл среды R10.
- Передача клеток в 15 мл коническую трубку и довести объем до 14 мл с помощью R10. Центрифуга при 600 х г в течение 5 мин при температуре 4 ° С. Удалите супернатант путем заливки его с одним чистым движением, чтобы предотвратить нарушения клеточного осадка.
- Добавляют 2 мл холодной (4 ° C) красных клеток буфера для лизиса (таблица 1) в каждую пробирку. Аккуратно перемешать в течение 3,5 мин, держа трубку на льду.
Примечание: Время проведения лизиса эритроцитов имеет решающее значение для предотвращения гибели лимфоцитов. Таким образом, точные сроки каждой партии буфера для лизиса красных кровяных телец необходимо будет оптимизировать. - Добавить ≈12-13 мл среды R10. Сразу центрифуге и отбросить супернатант, как на стадии 2.1.5. Выполните стадии промывки 2x дополнительной остроумияч R10 среда для удаления остатков красного буфера для лизиса клеток из клеток и избежать гибели лимфоцитов.
- Граф клеток в гемоцитометра разбавление 1:20 в трипановым синим, чтобы определить выход жизнеспособных клеток. Ожидать приблизительно 8-10 × 10 7 клеток на мышь.
- Выделение CD4 + Т - клеток с использованием магнитных шариков Kit для удаления CD8 +, CD11b +, CD16 / 32 +, CD45R +, MHC класса II + и Тер-119 + клеток.
- Аликвоты 8-10 х 10 7 клеток с этапа 2.1.8 в 15 мл коническую пробирку и центрифугируют при 600 х г в течение 5 мин при температуре 4 ° С. Слейте надосадочную с одним чистым движением, чтобы предотвратить нарушения клеточного осадка. Ресуспендируют клеток в каждой пробирке в 100 мкл инактивированной нагреванием FBS, затем добавляют 100 мкл смеси антител из набора. Инкубировать в течение 30 мин при температуре 4 ° С при осторожном перемешивании на валике.
- Хотя вышеуказанные клетки инкубирования, готовят шарики,
- Ресуспендируют бусины в ампуле встряхиванием в течение> 30 сек , то передача по 1 мл шариков на 8-10 х 10 7 клеток , приготовленных в 15 мл коническую пробирку. Добавляют 2 мл среды R10 на мл гранул и аккуратно перемешать.
- Поместите пробирку в магните в течение 1 мин, а затем отбросить супернатант. Удалить трубку от магнита и ресуспендируют бусин в 4 мл среды R10. Это количество шариков достаточно для двух раундов инкубации.
- Промывают клетки в конце инкубации антитела (этап 2.2.1) добавлением 10 мл среде R10 на пробирку. Аккуратно хорошо перемешать с закрученной трубки и центрифужные клеток при 600 мкг в течение 5 мин при 4 ° C. Слейте надосадочную с одним чистым движением, чтобы предотвратить нарушения клеточного осадка. Ресуспендируют клеток в 1 мл среды. R10
- Добавляют 2 мл отмытых бусинок с шага 2.2.2.2 (½ суммы, подготовленной для каждой пробирки) в каждую пробирку антитела клеток, обработанных и инкубировать в течение 30 мин при температуре 4 ° С при осторожном перемешивании на роМюллером.
- Ресуспендируют клеток шарик смесь осторожно пипеткой 5 раз с пипеткой, содержащим узким горлышком. Избегать образования пены. Поместите пробирку в магните бусами в течение 2 мин, а затем передать супернатант, содержащий отрицательно отобранных клеток в новую пробирку. Хранить эти клетки, так как они являются популяция CD4 +.
- Повторите негативный отбор еще один раз. Спин клетки с этапа 2.2.5 при 600 х г в течение 5 мин при температуре 4 ° С. Слейте надосадочную как на этапе 2.2.3 и ресуспендируют в 1 мл среды R10. Последующие шаги 2.2.4 и 2.2.5 снова. После окончательного выбора магнитного шарика, подсчет клеток для определения восстановления (типичные выходы 15-20 х 10 6 клеток на 10 8 лейкоцитов).
- Выделение Наивные CD4 + Т - клеток (CD25 - и CD62L высокого) Использование разделительной ячейки столбцов
- Центрифуга CD4 + Т - клетки при 600 мкг в течение 5 мин при 4 ° С и ресуспендируют в 150-200 мклR10 среда на 10 8 клеток. Добавьте 2 мкл биотинилированного CD25 акций моноклональных антител (клон 7D4). Инкубировать в течение 30 мин при температуре 4 ° С при осторожном перемешивании на валике.
- Промыть клеток путем добавления 10 мл буфера дл разделени клеток на колонке (таблица 1). Центрифуга при 600 х г в течение 5 мин при температуре 4 ° С. Удалить столько супернатант, насколько это возможно, и оценить объем буфера / ячеек остальных. Ресуспендируют до конечного объема примерно 90 мкл на 10 7 клеток.
- Добавьте 20 мкл стрептавидина шариков на 10 7 клеток и смешать с мягкими щелчками. Инкубировать в течение 15 мин при температуре 4 ° С.
- В то время как клетки инкубирования, готовят среда для разделения клеток (MS) столбцы для ручного подхода. Каждый столбец MS сохраняет 10 7 клеток, а так как CD25 + клетки , как правило , составляют около 10% от общего числа CD4 + клеток, используют 1 колонка на 10 8 CD4 + Т - клеток. Добавьте 500 мкл буфера на колонке дл разделени клеток в колонкеи пусть текут через. Повторите еще 2 раза.
- В конце клетки / гранулы инкубации, мыть клетки, добавляя 10 мл колоночного буфера дл разделени клеток и центрифугирование при 600 х г в течение 5 мин при температуре 4 ° С. Ресуспендируют клеток в 500 мкл буфера дл разделени клеток колонки на 10 8 клеток, но до сих пор используют 500 мкл , если меньше клеток.
- Применить 500 мкл клеток / суспензионной в колонну и собирать поток через. Передайте это через колонку 1 больше времени и собрать поток через еще раз. Полоскание в столбце 3 раза 500 мкл буфера для колонки дл разделени клеток, добавляя каждый поток через из этих промывок в собранных клеток. Они являются CD25 - клетки. Граф клеток, чтобы определить выход.
- Если Tregs (CD25 + клетки) желательно изолировать следующим образом .
- Удалить столбец из сепаратора и держать его над открытым универсальной трубки Стараясь для поддержания стерильности. Быстро пипетки 500 мкл буфера для разделения клеток на седловинеитп и прочно вымывать положительную фракцию, используя поршень, поставляемый вместе с колонной.
- Повторите процедуру Смывании еще 2 раза. Пропускают положительную фракцию над свежей MS разделительной колонки клеток и отбросить эту проточные. Твердо вымывать позитивных клеток в три раза, как и раньше, и центрифуг клеток при 600 мкг в течение 5 мин при 4 ° С Ресуспендируют клеток в 1 мл среды R10 и считать для определения выхода.
- Для получения CD25 - CD62L высокие Т - клетки, Центрифуга CD25 - Т - клетки , выделенные выше в п.2.3.6 при 600 х г в течение 5 мин при 4 ° С и ресуспендируют в 150-200 мкл среде R10 на 10 7 клеток. Добавьте 5 мкл акций биотинилированный CD62L моноклонального антитела (клон MEL-14) и инкубировали в течение 30 мин при температуре 4 ° С при осторожном перемешивании на ролик, как и раньше.
- Выполните мытье, Стрептавидином шарик связывания, и Подготовку колонки разделения клеток, как описано для протокола изоляции Treg (2.3.7). Это использование временибольшое разделение клеток (LS) колонка, которая имеет емкость 10 8 клеток.
- Поскольку CD62L высокой Т - клетки , как правило , составляет около 60-70% CD25 - клеток, используют 1 LS колонки разделения клеток на 10 8 клеток. Добавить Смесь клетки / гранулы в LS колонке дл разделени клеток, как описано в п.2.3.6 - на этот раз отбрасывая окончательный поток через и столбец ополаскивателей. Сбор высокие Т - клетки , CD62L , как описано в шаге 2.3.7 для сбора клеток CD25 +.
- Центрифуга клетки при 600 мкг в течение 5 мин при 4 ° С Ресуспендируют клеток в 1 мл R10 и рассчитывать, чтобы определить выход. Разбавить до 0.75-1 Х10 6 клеток на мл в среде R10.
- Анализ чистоты клеток путем флуоресцентного активированного сортировки клеток (FACS).
- Стратегия окрашивания FACS
- Для клеток до и после всех этапов изоляции, пятно с CD4-FITC, CD8a-PerCP-Cy5.5 (хелперов и цитотоксических Т-клеток), и MHCII-PE (МНС класса II, выражающие клетки).
- Для клеток до и после выделения CD25, пятно с CD4-FITC и CD25-PE (Tregs по сравнению с другими Т-хелперы).
- Для клеток до и после CD62L изоляции пятно с CD4-FITC, CD62L-PE и CD44-APC (наивным по сравнению с памятью и эффекторных Т-хелперы).
- Для каждого анализа места 10 5 клеток в лунку 96 - луночного U-образным дном. Центрифуга при 600 х г в течение 5 мин при температуре 4 ° С. Слейте среды и промыть клетки один раз 200 мкл ДЗФР. Центрифуга, как описано выше, и слейте DPBS.
- Добавляют 50 мкл ДЗФР на лунку и 0,5 мкл каждого из антител (как указано в 2.4.1) и инкубируют при комнатной температуре в течение 30 мин в темноте, чтобы избежать обесцвечивания флуоресцентной метки.
- Вымойте клетки 2x с ДЗФР, как в 2.4.2 и сразу же анализируют на проточном цитометре с использованием протоколов и руководящих принципов, характерных для инструмента под рукой.
Примечание: После последней стадии выделения хорошей подготовки, 90-95% клеток будетбыть CD4 + CD25 - CD62L высокой Т - клеток.
- Стратегия окрашивания FACS
3. Ретровирусное Трансдукция активированных CD4 + Т - клеток и их дифференцировку в специфические Т - хелперов подмножествах
- Ретровирусное Трансдукция
Примечание: Ретровирусное интеграции и экспрессии генов требует деление клеток. Таким образом, Т-клетки должны быть активированы O / N.- В то время как изолируя наивных Т-клеток, пальто 24 хорошо культуре ткани пластины с анти-CD3 и анти-CD28 антитела разбавленных в ДЗФР. Добавить 250 мкл на лунку. Используйте анти анти-CD3 на 1 мкг / мл. Использование анти-CD28 при 2 мкг / мл, если клетки будут в конечном счете, будет дифференцироваться под Th0, Th1, Th2 и iTregs условиях. Использование анти-CD28 в концентрации 10 мкг / мл для Th9 и Th17 условий. Выдержите ~ 2 ч при 37 ° С в инкубаторе тканевых культур.
- Непосредственно перед культивированием клеток, удалить раствор антител и промойте пластину с ~ 250 мкл на лунку ДЗФР для удаления unbouго антитела. Будьте осторожны, чтобы не позволить пластины высохнуть так обработать максимум 6 лунок одновременно. Удалить DPBS помыть и добавить 1 мл наивных CD4 + Т - клеток в среде R10 в 0,75-1 × 10 6 клеток на мл. Активируйте клетки O / N (14-16 ч).
- На следующий день, центрифужные клетки в пластине при 900 х г в течение 5 мин при 30 ° С, чтобы присоединить клетки к дну лунок.
- Собрать и сохранить среду, стараясь не сместить клетки, и заменить 1 мл вируса супернатанта культуры, полученного из протокола производства вируса. Не позволяйте клетки высыхают так обработать максимум 4 лунок одновременно.
- В каждую лунку добавляют 1 мкл 8 мг / мл полибрена и 10 мкл 1 М HEPES , рН 7,5 , чтобы помочь поглощение вирусов и предотвратить значительное подщелачивание в окружающем СО 2 в течение последующего центрифугирования. Центрифуга клетки в пластине при 900 мкг в течение 90 мин при 30 ° С
- Дифференциация клеток в SpeciFIC Поднаборы
- Тщательно удалите вирус супернатант культуры и заменяют 1 мл среды, собранных выше на этапе 3.1.4, так как он содержит ИЛ-2 и другие факторы роста Т-клеток. Добавьте реагенты, указанные в таблице 2 и культуры клеток в течение 3-4 дней, чтобы дифференцировать клетки в определенных подмножеств.
- Анализ клеток
- Для внутриклеточного окрашивания цитокинов, лечения клеток с 1 мкг / мл каждого из форболовыми 12-миристат-13-ацетата (РМА) и Ionomycin в течение 4 часов. В течение последних 2 ч стимуляции, добавляют 1 мкг / мл брефелдин A.
- Ресуспендируют клетки из нижней части каждой лунки с помощью пипетки вверх и вниз несколько раз. Передача около 10 5 клеток в 96 - луночный U нижней пластины (обычно 100 мкл из 1 мл культуры клеток Th) для фиксации и окрашивания.
- Для окрашивания GFP, зафиксировать клетки с 100 мкл 2% параформальдегида при комнатной температуре в течение ровно 5 мин, в течение длительного времени инкубации будет отбеливать GFP Aго мешают Foxp3 окрашивания. Вымойте клеток путем добавления 100 мкл ДЗФР в каждую лунку и центрифуге при 600 мкг при 22 ° С в течение 5 мин. Слейте надосадочную.
Внимание: параформальдегид является токсичным. Обращайтесь с ним в вытяжном шкафу с кожей и средства защиты глаз. - Закрепить клетки снова с помощью 100 мкл буфера 1x Fixation, сделанный из комплекта для окрашивания Foxp3 (1 объем фиксирующих буфера 3 объема разбавителя). Инкубируйте клетки в течение 45 минут при комнатной температуре, или в качестве альтернативы инкубировать при температуре 4 ° С в течение 16 часов.
- После фиксации проницаемыми клетки путем добавления 100 мкл буфера пермеабилизации из того же комплекта. Инкубируют при комнатной температуре в течение 30-45 мин, затем центрифугировать при 600 х г при 22 ° С в течение 5 мин.
- Для окрашивания антител, удалить фиксации / буфер пермеабилизирующего и добавить 50 мкл свежего буфера пермеабилизации. Добавить требуемое антитело, разведенное в пермеабилизации буфере при 0,5 мкл на 50 мкл буфера на образец. Выдержите в течение 30-45 мин при комнатной температуре в темноте, чтобы избежать отбеливания гриппаorescent этикетки.
- Добавьте 150 мкл буфера и пермеабилизирующего центрифуге при 600 х г при 22 ° С в течение 5 мин. Откажитесь буфер пермеабилизирующего и добавьте 200 мкл DPBS. Анализ на проточном цитометре с использованием протоколов и руководящих принципов, характерных для инструмента под рукой.
ПРИМЕЧАНИЕ: Не забудьте включить соответствующие элементы управления для окрашивания цитокина , таких как окрашивание с контрольными изотипа антител, анализируя неактивированной или активированными Th0 клетки и т.д.
Результаты
Успех этой экспериментальной системы требует высокой степенью чистоты популяции Т-клеток и препаратов ретровируса с высоким титром. Типичные результаты приведены здесь в качестве примеров успешных экспериментов. На рисунке 1 показана типичная чистота пред- и пост отобранных групп населения на каждом этапе наивным протокола изоляции клеток помощник T. Рисунок 2 и 3 иллюстрируют анализ производства ретровируса с помощью выражения GFP в трансфицированных НЕК 293T клеток (рис 2) и трансдуцированных Т - клетки (рисунок 3). Эффективность трансфекции клеток НЕК 293Т может существенно изменяться в зависимости от различных ретровирусов конструкциями, но это часто не коррелирует с уровнем производства ретровируса наблюдаемого с числом GFP + Т - клеток. Кроме того , количество GFP + Т - клеток может изменяться в зависимости от условий поляризации. Кроме того, мЕАН уровень экспрессии GFP и введенного гена может изменяться в зависимости от количества вирусных копий интегрированных, эффект сайта интеграции по отношению к транскрипции, и пост-транскрипционные регуляторные механизмы, влияющие на вирусный транскрипт.
И, наконец, на рисунке 4 показаны некоторые типичные результаты , которые мы наблюдали с дифференциацией Т - хелперного , когда микроРНК микроРНК-15b / 16 гиперэкспрессии. Эти результаты показывают некоторые из изменчивости, которая может произойти в пределах отдельного эксперимента так истинные эффекты должны быть обоснованы с помощью статистического анализа многократных повторных экспериментах с использованием различных препаратов Т-хелперы. В этих опытах Th2 реакции может быть трудно наблюдать в / 6 линии C57BL используется здесь, потому что они склонны к Th1 ответа. Аналогично, IL-9 окрашивание может быть трудно обнаружить выше фона. Поэтому крайне важно, чтобы сделать контроль изотипа и установить надлежащую компенсацию для обеспечения правильного гатин экспрессии цитокина. В наших результатах , мы обнаружили , что микроРНК-15b / 16 усиливает iTreg индукцию путем ингибирования MTOR сигнальный путь через подавление экспрессии Rictor компонентов и МРМ 15. микроРНК-15b / 16 могут иногда влиять на Th0, Th1 и Th17 дифференциацию в отдельных экспериментах, но нет никакого существенного влияния при рассмотрении в нескольких повторных экспериментах. В отличие от микроРНК-15b / 16 избыточная экспрессия не приводит к существенному подавления Th9 дифференциации (см ссылку 18).

Рисунок 1. Типичная чистота Т - хелперов на каждой стадии выделения. Представитель проточной цитометрии результаты указанных антигенов показаны от ворот живых клеток , обозначенных в Forward Scatter (FSC) и бокового рассеяния (SSC) участков. (А) до и после CD4 негативный отбор. Профили экспрессии CD4,CD8a и MHCII показаны. Они иллюстрируют обогащение Т-хелперов и потерю цитотоксических Т-клеток и МНС класса II, выражающие клетки. Хорошая очистка должна привести к ~ 90% CD4 + T - клеток на данном этапе. (Б) выбор CD25. Слева являются профили экспрессии CD4, CD8a и MHCII, а справа являются CD4 и экспрессия CD25 профилей до и после выбора. На данный момент> 95% CD25 отрицательно отобранных клеток должны быть CD4 + CD25 -. (С) CD62L выбор. CD4, CD8a и MHCII профили экспрессии показаны на левой стороне. Справа профили экспрессии для CD62L и CD44 показаны для пред- и пост-CD62L выбранных клеток наряду с CD4 и CD62L профиля экспрессии почтовых выбранных ячеек. После выбора CD62L практически все ячейки памяти (CD44 +), удаляются , оставляя высокообогащенный популяцию наивных Т - хелперов , который содержит 10-15% эффекторных клеток (CD62L низкий). Для всехFACS профили, эквивалентные параметры и шкалы для определенного параметра были сохранены во всем. Числа представляют процент клеток в пределах огороженной населения. Небольшое уменьшение размеров клеток после первоначального отбора, предположительно , из - за механического напряжения во время протокола. Пожалуйста , нажмите здесь , чтобы посмотреть увеличенную версию этой фигуры.
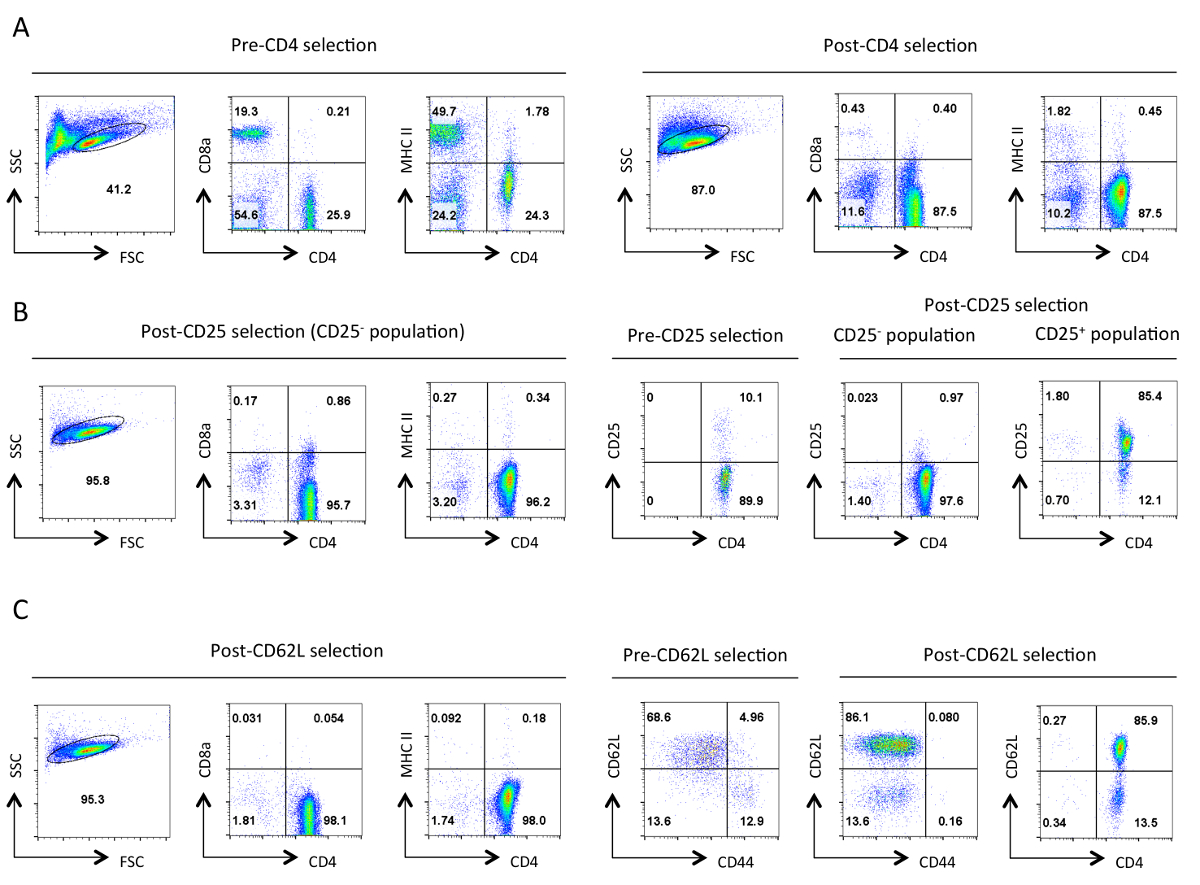{kind=link}

Рисунок 2. Анализ ретровирусоподобных трансфецировали клетки 293Т НЕК. Экспрессии GFP показан на НЕК 293Т , которые были либо нетрансфицированные или трансфицируют и анализировали после сбора вирусных супернатантов культуры. Анализ GFP был сделан на живые клетки от ворот на FSC и SSC участок в первой панели. Числа представляют процент GFP + клеток в зоне интереса . Типичный тЭффективность ransfection в диапазоне от 30-90%. Пожалуйста , нажмите здесь , чтобы посмотреть увеличенную версию этой фигуры.
{kind=link}

Выражение GFP Рисунок 3. Анализ ретровируса трансдуцированных Т - хелперов. Показана на ретровирусных-трансдуцированных Т - хелперы после дифференцировки в Th0, Th1, Th2, Th9, Th17, и условия поляризации Treg в течение трех дней. Анализ был закрытого типа на живых и активированных клеток, указанных в панели FSC / SSC. Эффективность трансдукции может колебаться в пределах 10-75%, в зависимости от конструкции и условий поляризации. Кроме того, средняя интенсивность флуоресценции GFP экспрессии может изменяться. Пожалуйста , нажмите здесь , чтобы посмотреть увеличенную версию этой фигуры.
{kind=link}

Рисунок 4. Влияние MIR-15b / 16 суперэкспрессия на хелперов дифференциации Т - клеток в различных условиях поляризации. Типичные профили цитокина показаны на GFP + популяции клеток из рисунка 3. Пожалуйста , нажмите здесь , чтобы посмотреть увеличенную версию этой фигуры.
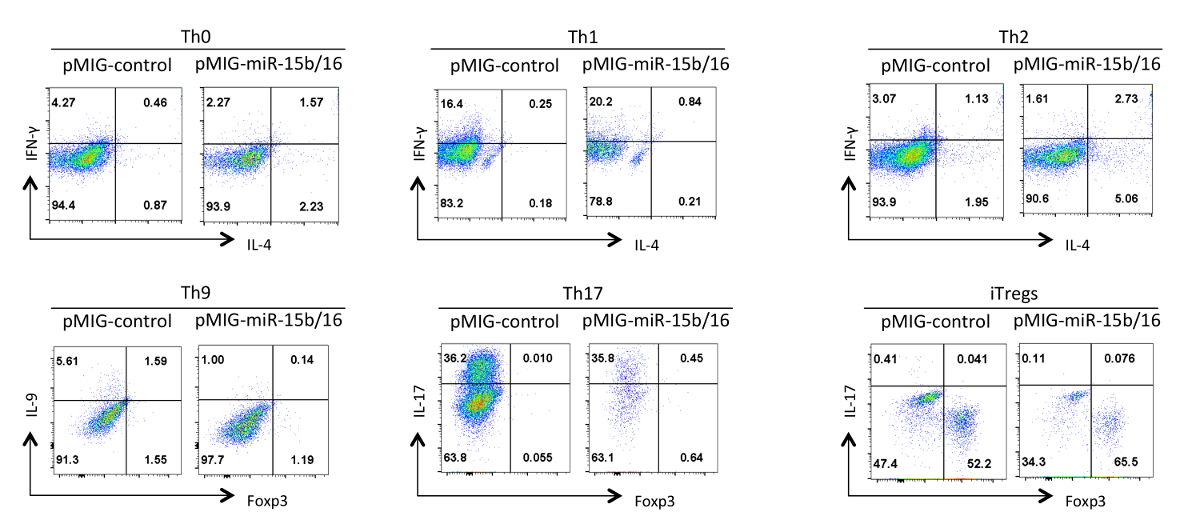{kind=link}

Таблица 1:. Буферы , используемые в этих протоколах Пожалуйста , нажмите здесь , чтобы загрузить эту таблицу в виде таблицы Excel.
| помощникТ условия клеточной поляризации | ||
| Th0 | анти-IL-4 | 5 мкг / мл |
| анти-IFN-γ | 5 мкг / мл | |
| Th1 | Рекомбинантный IL-12 | 20 нг / мл |
| анти-IL-4 | 5 мкг / мл | |
| Th2 | рекомбинантный IL-4 | 40 нг / мл |
| анти-IFN-γ | 5 мкг / мл | |
| Th9 | рекомбинантный TGF-β | 2,5 нг / мл |
| рекомбинантный IL-4 | 40 нг / мл | |
| анти-IFN-γ | 10 мкг / мл | |
| Th17 | рекомбинантный TGF-β | 2,5 нг / мл |
| рекомбинантный IL-6 | 50 нг / мл | |
| анти-IFN-γ | 5 мкг / мл | |
| анти-IL-4 | 5 мкг / мл | |
| анти-IL-2, | 5 мкг / мл | |
| Tregs | рекомбинантный TGF-β | 2,5 нг / мл |
| рекомбинантный IL-2 | 5 нг / мл | |
Таблица 2: Т - хелперного подмножество условия поляризации.
Обсуждение
Ретровирусное опосредованной избыточная экспрессия генов является мощным средством для анализа функции в Т-хелперов, так как их развитие и функция часто определяется уровнем экспрессии ключевых регуляторов. Тем не менее, осторожность интерпретации результатов не требуется, поскольку уровни экспрессии значительно выше тех эндогенного гена можно ввести множество артефактов. Таким образом, этот метод следует сочетать с другими, чтобы проверить актуальность функции. Например, избыточная экспрессия должна быть дополнена снижением экспрессии с помощью миРНК или нокаут гена если таковые имеются. С микроРНК, мы дополнены ГИПЕРЭКСПРЕССИЯ эксперименты с теми блокировки с помощью вирусов, суперэкспрессированный искусственного микроРНК таргетирования сайтов , которые действовали в качестве конкурентных ингибиторов для микроРНК 15. Ретровирусные трансдуцированных клеток также могут быть использованы в биохимических анализов, включающих в себя РНК и анализа белков. Однако, главное ограничение этих экспериментов эффективность трансдукции Резulting в смешанной популяции трансдуцированных и untransduced клеток. Таким образом, эти анализы, скорее всего , потребуется сортировка GFP + населения. И, наконец, в пробирке дифференциации анализа должны быть объединены с экспериментами в естественных условиях, и один из способов это может быть достигнуто , является адаптивно передачи трансдуцированных Т - клеток в организм мышей и после их дифференцировки и их влияние на иммунный ответ.
Одним из ключевых ограничений для этой системы является размер генома РНК, которые могут быть упакованы в ретровируса капсида. По нашему опыту, максимальный размер вставки для системы ретровирусов MIG, которая дает хорошую продукцию вируса составляет 3-3,5 кб. Таким образом, более крупные гены не могут быть проанализированы с помощью этой системы, так как они дают плохие вирусные титры. Тем не менее, большинство генов меньше, чем этот размер так что эта система полезна для широкого круга исследований генов.
С помощью ретровирусов трансдукции, несколько альтернатив в рамках этих Protocoиспользовались Ls. Многие исследователи использовали упаковочные линии клеток , которые стабильно экспрессируют ретровирусные гены (например , ссылка 16). Тем не менее, мы получили самые высокие титры с использованием стандартных НЕК 293Т с котрансфекции вируса помощника вектора-Eco PCl. Выделение наивным Т-хелперов, также может быть достигнуто с помощью сортировки клеток, а не магнитного протокола колонке шарика и разделения клеток, но это требует доступа к мобильному сортировщика, а затраты на время сортировки, как правило, выше, чем бусинки реагентов. Наконец, существуют вариации на условиях активации, используемых для дифференциации Т-хелперы в различные подмножества. Например, TCR стимуляция клеток слишком долго перед воздействием Treg условий , побуждающих может ингибировать их индукции 16. Это может быть проблемой, так как ретровирусный выражение требует деление клеток, индуцированную стимуляции клеток. Тем не менее, мы нашли эффективную индукцию Treg, используя этот протокол с O / N эквания до ретровирусной трансдукции.
В рамках этих протоколов, успешное применение требует нескольких факторов. Высокий титр препараты ретровирус должны эффективно трансфекции клеток 293Т НЕК настолько высокого качества ДНК и точно подготовленную 2x HBS имеют важное значение. Кроме того, плотность клеток из клеток НЕК 293Т должен быть примерно 50% в точке трансфекции, так как хорошее выражение трансфицированных ДНК требует, чтобы клетки активно растут, и это будет запрещено, если клетки слишком разреженным или плотным. Клетки при оптимальной плотности во время трансфекции должна достигать впадения в какой-то момент в течение этапов сбора вируса, но они будут продолжать производить вирусные акции с высоким титром весь путь вплоть до последней коллекции. Эффективное дифференцировка Т - хелперов требует хорошего качества клеток таким образом гарантировать , что отдельные клетки в чистоте , показанного на фиг.1. Подобным же образом, качество клеток зависит от мышей фрО.М. которой они были выделены. Для этих исследований мы использовали 6-8 неделю 6 мышей C57BL / старый. Более старые мыши могут иметь менее наивные клетки, и другие штаммы могут отличаться в их дифференциации. Например, BALB / C мышей , более склонны к Тh2 - ответов , чем мыши C57BL / 6 17 так , как было указано выше, C57BL / 6 Т - клетки могут быть трудно вызвать Th2 ответ. Кроме того, любое из условий дифференциации могут незначительно отличаться от лаборатории к лаборатории, и эффект гена сверхэкспрессии может только стать очевидными в неоптимальных условиях так концентрации цитокинов в различных условиях поляризации может потребоваться титрование. И, наконец, эффекты суперэкспрессированный гена или условия поляризации на пролиферацию клеток может влиять на эффективность трансдукции так измерения эффекты представляющего интерес гена может потребовать оптимизации времени и концентрации поляризационными реагентов. Оптимизация все эти факторы должны привести к информативных результатов с этой системой.
Раскрытие информации
The authors have no conflicting interest in the publication of this work.
Благодарности
This work was supported by a Biotechnology and Biological Sciences Research Council (BBSRC) grant (BB/H018573/1) and a BD Biosciences grant.
Материалы
| Name | Company | Catalog Number | Comments |
| RPMI | Sigma | R8758 | |
| DMEM | Sigma | D5671 | |
| Penicillin Streptomycin solution | Sigma | P4333 | |
| L-Glutamine | Sigma | G7513 | |
| β-mercaptoethanol | Sigma | M3148 | |
| DPBS | Sigma | D8537 | |
| MIG vector | Addgene | Plasmid 9094 | |
| pCL-Eco vector | Addgene | Plasmid 12371 | |
| Cell strainer | BD Falcon | 352350 | |
| Magnetic beads mouse CD4 cell kit | Invitrogen (Dynabeads) | 11415D | |
| Streptavidin Beads | Miltenyi Biotech | 130-048-102 | |
| MS cell separation columns | Miltenyi Biotech | 130-042-201 | |
| LS cell separation columns | Miltenyi Biotech | 130-042-401 | |
| CD25 Biotenylated MAb | BD Biosciences | 85059 | clone 7D4 |
| CD62L Biotenylated MAb | BD Biosciences | 553149 | clone MEL-14 |
| Polybrene (Hexadimethrine Bromide) | Sigma | 107689 | |
| Anti-CD3 | eBiosciences | 16-0031-85 | clone 145-2C11 |
| Anti-CD28 | eBiosciences | 16-0281-85 | clone 37.51 |
| Anti-IL-4 | BD Biosciences | 559062 | clone 11B11 |
| Anti-IFN-gamma | BD Biosciences | 559065 | clone XMG1.2 |
| Anti-IL-2 | BD Biosciences | 554425 | cloneJES6-5H4 |
| Recombinant IL-12 p70 | eBiosciences | 14-8121 | |
| Recombinant IL-4 | BD Biosciences | 550067 | |
| Recombinant TGF-beta | eBiosciences | 14-8342-62 | |
| Recombinant IL-6 | eBiosciences | 14-8061 | |
| Recombinant IL-2 | eBiosciences | 14-8021 | |
| PMA | Sigma | P8139 | |
| Ionomycin | Sigma | I0634 | |
| Brefeldin A | eBiosciences | 00-4506 | |
| Paraformaldehyde | Sigma | 16005 | Paraformaldehyde is toxic so use appropriate caution when handling |
| Foxp3 staining buffer set | eBiosciences | 00-5523 | |
| Anti-CD4 FITC | eBiosciences | 11-0041 | clone GK1.5 |
| Anti-CD8a perCP-cy5.5 | eBiosciences | 45-0081-80 | clone 53-6.7 |
| Anti-MHCII PE | eBiosciences | 12-0920 | clone HIS19 |
| Anti-CD25 PE | eBiosciences | 12-0251-82 | clone PC61.5 |
| Anti-CD62L PE | eBiosciences | 12-0621-82 | clone MEL-14 |
| Anti-CD44 APC | eBiosciences | 17-0441 | clone IM7 |
| Anti-IFN-gamma FITC | eBiosciences | 11-7311-81 | clone XMG1.2 |
| Anti-IL-4 PE | BD Biosciences | 554435 | clone 11B11 |
| Anti-IL-9 PE or APC | eBiosciences/Biolegend | 50-8091-82/514104 | clone RM9A4 |
| Anti-IL-17a PE | BD Biosciences | 559502 | clone TC11-18H10 |
| Anti-Foxp3 APC or PE | eBiosciences | 17-5773-82/12-5773-80 | clone FJK-16s |
| NaCl | Sigma | S7653 | |
| KCl | Sigma | P9333 | |
| Na2HPO4-2H2O | Sigma | 71643 | |
| Dextrose/Glucose | Sigma | G7021 | |
| HEPES, free acid | Sigma | H3375 | |
| NH4Cl | Sigma | A9434 | |
| Disodium EDTA | Sigma | D2900000 | |
| KHCO3 | Sigma | 237205 | |
| CaCl2 | Sigma | C5670 |
Ссылки
- Baumjohann, D., Ansel, K. M. MicroRNA-mediated regulation of T helper cell differentiation and plasticity. Nat Rev Immunol. 13 (9), 666-678 (2013).
- Wilson, C. B., Rowell, E., Sekimata, M. Epigenetic control of T-helper-cell differentiation. Nat Rev Immunol. 9 (2), 91-105 (2009).
- Pearce, E. L., Poffenberger, M. C., Chang, C. H., Jones, R. G. Fueling immunity: insights into metabolism and lymphocyte function. Science. 342 (6155), 1242454 (2013).
- Cherry, S. R., Biniszkiewicz, D., Van Parijs, L., Baltimore, D., Jaenisch, R. Retroviral Expression in Embryonic Stem Cells and Hematopoietic Stem Cells. Mol and Cell Biol. 20 (20), 7419-7426 (2000).
- Miller, A. D., Rosman, G. J. Improved retroviral vectors for gene transfer and expression. BioTechniques. 7 (9), 980-990 (1989).
- Grez, M., Akgün, E., Hilberg, F., Ostertag, W. Embryonic stem cell virus, a recombinant murine retrovirus with expression in embryonic stem cells. Proc Nat Acad Sci USA. 87 (23), 9202-9206 (1990).
- Naviaux, R. K., Costanzi, E., Haas, M., Verma, I. M. The pCL vector system: rapid production of helper-free, high-titer, recombinant retroviruses. J Virol. 70 (8), 5701-5705 (1996).
- Filipowicz, W., Bhattacharyya, S. N., Sonenberg, N. Mechanisms of post-transcriptional regulation by microRNAs: are the answers in sight?. Nat Rev Genetics. 9 (2), 102-114 (2008).
- He, L., Hannon, G. J. MicroRNAs: small RNAs with a big role in gene regulation. Nat Rev Genetics. 5 (7), 522-531 (2004).
- Bernstein, E., Kim, S. Y., et al. Dicer is essential for mouse development. Nat Genetics. 35 (3), 215-217 (2003).
- Cobb, B. S., Hertweck, A., et al. A role for Dicer in immune regulation. J Exp Med. 203 (11), 2519-2527 (2006).
- Chong, M. M. W., Rasmussen, J. P., Rudensky, A. Y., Rundensky, A. Y., Littman, D. R. The RNAseIII enzyme Drosha is critical in T cells for preventing lethal inflammatory disease. J Exp Med. 205 (9), 2005-2017 (2008).
- Liston, A., Lu, L. F., O'Carroll, D., Tarakhovsky, A., Rudensky, A. Y. Dicer-dependent microRNA pathway safeguards regulatory T cell function. J Exp Med. 205 (9), 1993-2004 (2008).
- Zhou, X., Jeker, L. T., et al. Selective miRNA disruption in T reg cells leads to uncontrolled autoimmunity. J Exp Med. 205 (9), 1983-1991 (2008).
- Singh, Y., Garden, O. A., Lang, F., Cobb, B. S. MicroRNA-15b/16 Enhances the Induction of Regulatory T Cells by Regulating the Expression of Rictor and mTOR. J Immunol. 195 (12), 5667-5677 (2015).
- Sauer, S., Bruno, L., et al. T cell receptor signaling controls Foxp3 expression via PI3K, Akt, and mTOR. Proc Nat Acad Sci USA. 105 (22), 7797-7802 (2008).
- Yagi, J., Arimura, Y., Takatori, H., Nakajima, H., Iwamoto, I., Uchiyama, T. Genetic background influences Th cell differentiation by controlling the capacity for IL-2-induced IL-4 production by naive CD4+ T cells. Int Immunol. 18 (12), 1681-1690 (2006).
- Singh, Y., Garden, O. A., Lang, F., Cobb, B. S. MicroRNAs regulate T-cell production of interleukin-9 and identify hypoxia-inducible factor-2a as an important regulator of T helper 9 and regulatory T-cell differentiation. Immunology. 149, 74-86 (2016).
Перепечатки и разрешения
Запросить разрешение на использование текста или рисунков этого JoVE статьи
Запросить разрешениеСмотреть дополнительные статьи
This article has been published
Video Coming Soon
Авторские права © 2025 MyJoVE Corporation. Все права защищены