
需要订阅 JoVE 才能查看此. 登录或开始免费试用。
Method Article
软件辅助跟踪动态细胞突变中蛋白质浓度的图形用户界面
摘要
我们提供了一个软件解决方案,用于半自动跟踪动态细胞突起长度的相对蛋白质浓度。
摘要
Filopodia是与迁移和细胞 - 细胞通信相关联的动态的,指状的细胞突起。为了更好地了解潜伏期起始,延长和随后的稳定或收缩的复杂信号传导机制,确定这些动态结构中的时空蛋白质活性至关重要。为了分析丝状伪足中的蛋白质功能,我们最近开发了一种半自动跟踪算法,适应线形形状变化,从而允许平行分析突起动力学和沿整个长度的整个相对蛋白质浓度。在这里,我们提供了一个详细的逐步协议,用于优化单元处理,图像采集和软件分析。我们进一步提供在图像分析和数据表示期间使用可选功能的说明,以及所有关键步骤的故障排除指南。最后,我们还包括d的比较带有图像分析软件的其他程序可用于丝状伪迹定量。在一起,所提出的方案提供了使用图像分析软件准确分析丝状体突起中的蛋白质动力学的框架。
引言
肌动蛋白调节蛋白的时空控制与纤维素酶动力学1,2相关。随着时间的推移,跟踪沿着整个网络长度的空间分辨蛋白质浓度对于促进我们对这些动态结构的起始,伸长,稳定或崩溃的机理的理解至关重要3,4 。不同于细胞质中的蛋白质分析,其中许多细胞形状变化发生在更大的尺度,丝状伪足是持续弯曲5并弯曲的动态微结构,因此使用简单的方法(例如线扫描)排除分析。
可用于跟踪丝状体形状的不同软件解决方案6,7,8,9。 Likewise,细胞体内蛋白质动力学的比例跟踪软件已经开发10,11 。为了结合自动跟踪网格形状和时空蛋白质分析,我们最近开发了基于凸包算法的图像分析软件12 。这种通过图形用户界面(GUI)操作的新型分析方法首次将沿着丝状体长度和生长速度的相对蛋白质浓度相结合,从而允许精确测量时空蛋白质分布,而与这些蛋白质的运动无关动态结构12 。
软件背后的想法(源代码可以免费获得,见下文),凸包的顶点之一将与圆顶的尖端一致( 图1A )。通过查看后续框架for最近的凸包顶点,可以在整个影片中追踪移动的尖端。一旦在每个框架中检测到尖端,它的位置用于通过将尖端与光栅底部的参考点连接起来绘制轴( 图1B )。最后,使用等距节点,其位置由具有与轴正交的线上的最大强度的中值像素确定,用于确定跟随斑点形状的骨架。利用这种自适应骨架,产生一个色谱图 ,以沿着丝状体长度追踪多达三个通道的丝状体生长和蛋白质浓度( 图1C )。

图1:图像分析软件的工作原理。 ( A )后面的算法该软件。在步骤1中,用户指定参考(基数)和顶点的顶点(tip)。在步骤2-1中,使用具有最大强度值的中值像素获得光栅的骨架。在步骤3中,骨架用于空间蛋白质强度分布。在步骤2-2中,软件会自动跟踪后续帧中的提示。整个过程迭代。 ( B )算法的快照,其中包含真正的光纤引入重要元素,例如用于跟踪的凸包。 ( C )使用算法可以测量的参数概述。这个数字已经从参考文献12中修改过。 请点击此处查看此图的较大版本。
{kind=link}
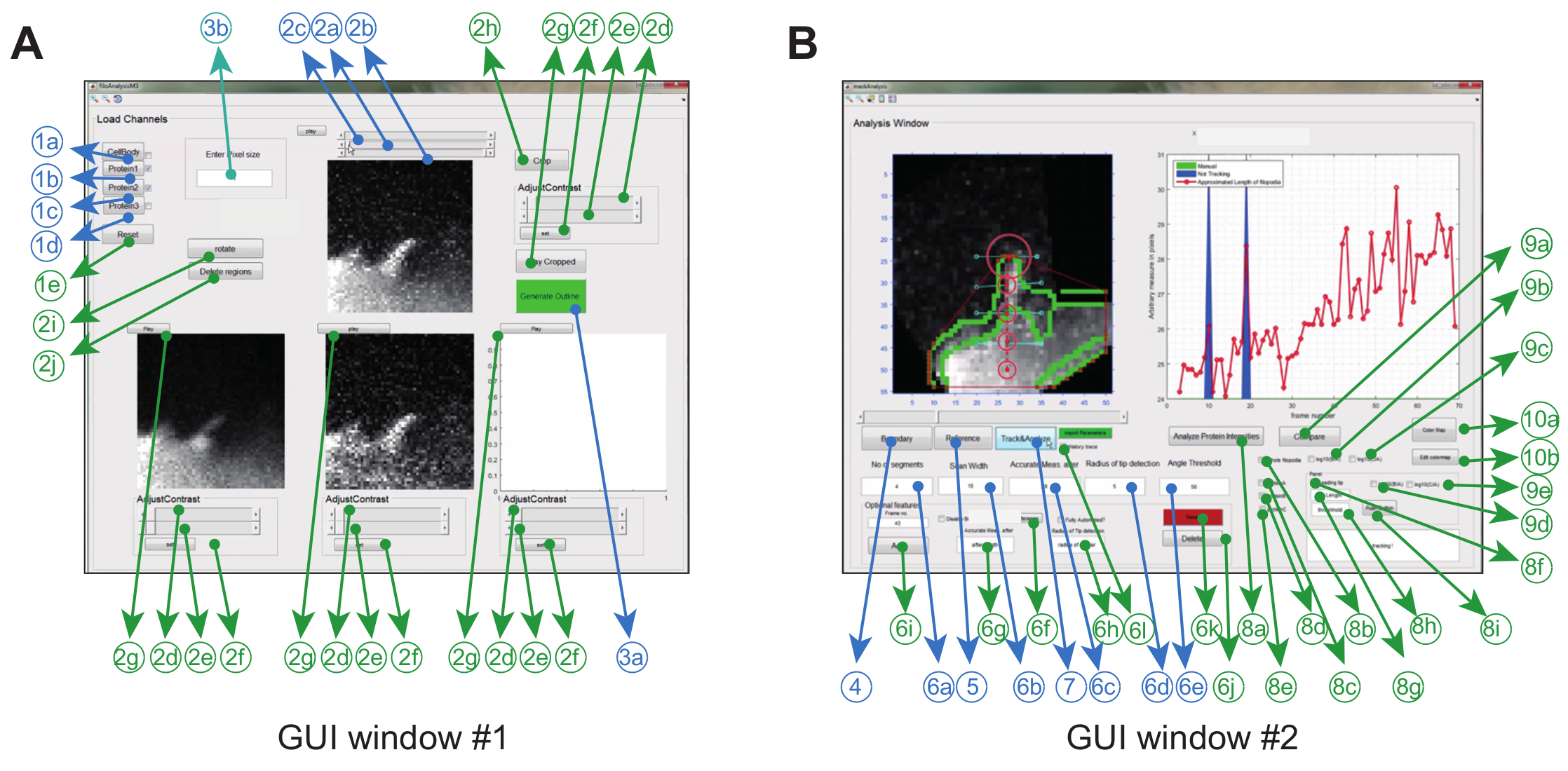
图像分析软件通过图形使用在Matlab(简称编程软件)中运行r接口。为了使特定实验设置的灵活性和鲁棒性达到最大化,用户可以调整一系列跟踪参数( 例如允许的弯曲角度和帧间移动),并对电影进行一些修正( 例如裁剪,旋转,删除不需要的对象) ( 图2A和表1) 。
| GUI | 没有。 | 强制性 | 描述 | 名称(在GUI中) | ||||
| #1 | 1A | ÿ | 加载堆叠的.tiff文件,表示单元格主体(框已检入)或从通道创建叠加的单元格体 | CellBody | ||||
| #1 | 1B | ÿ | 加载与蛋白质1相对应的堆叠图像文件 | 蛋白质1 | ||||
| #1 | 1C | ÿ | 加载对应于蛋白质2的堆叠图像文件 | 蛋白质2 | ||||
| #1 | 1D | ÿ | 加载对应于蛋白质3的堆叠图像文件 | 蛋白质3 | ||||
| #1 | 1E | ñ | 将所有内容重置为预先加载的堆叠图像文件 | 重启 | ||||
| #1 | 2A | ÿ | 滚动条确定GUI窗口#2中分析的初始帧 | NA | ||||
| #1 | 2B | ÿ | 滚动条,以确定在GUI窗口#2中进行分析的最终框架 | NA | ||||
| #1 | 2C | ÿ | 代表卷曲的滚动条nt框架 | NA | ||||
| #1 | 2D | ñ | 所有像素将被设置为零以下的像素的灰度值 | NA | ||||
| #1 | 2E | ñ | 所有像素将被设置为最大值的像素的灰度值 | NA | ||||
| #1 | 2F | ñ | 设置由<2e>&<2f>指定的像素的强度值 | 组 | ||||
| #1 | 2克 | ñ | 播放强度调整的电影 | 玩 | ||||
| #1 | 2H | ñ | 作物图像 | 作物 | ||||
| #1 | 2I | ñ | 旋转图像 | 旋转 | ||||
| #1 | 2J | ñ | 删除整个堆栈中的区域 | 删除区域 | ||||
| #1 | 3A | ÿ | 点击打开"分析窗口"(GUI窗口#2) | 跟踪窗口 | ||||
| #1 | 3B | ÿ | 输入像素的大小(以微米为单位) | 输入像素大小 | ||||
| #2 | 4 | ÿ | 单击以生成叠加单元体的边界/边缘图像 | 边界 | ||||
| #2 | 五 | ÿ | 点击以选择网点的基数和提示 | Referernce | ||||
| #2 | 6A | ÿ | 输入段或节点的数量 | 段数 | ||||
| #2 | 6B | ÿ | 输入扫描长度(垂直于轴) | 扫描宽度 | ||||
| #2 | 6C | ÿ | 输入超过该长度的丝网眼开始弯曲的长度 | 准确测量 | ||||
| #2 | 6D | ÿ | 输入尖端检测圆的半径(即顶点可以在下一帧中进行本地化的区域) | 尖端检测半径 | ||||
| #2 | 6E | ÿ | 输入光栅可以从垂直轴弯曲的最大角度 | 角度阈值 | ||||
| #2 | 1207米 | ñ | 为该特定框架的基准和提示添加参考点 | 选择参考 | ||||
| #2 | 6克 | ñ | 输入长度超过该长度的纤维纹身开始弯曲该特定框架 | 准确测量 | ||||
| #2 | 6H | ñ | 输入该特定帧的检测圆的半径 | 尖端检测半径 | ||||
| #2 | 6I | ñ | 输入特定框架的所有参数后,单击以将值存储到内存和文件以供进一步参考 | 加 | ||||
| #2 | 6J | ñ | 单击以删除该框架的设置手动参数 | 删除 | ||||
| #2 | 6K | ñ | 单击以删除手动存储的所有参数,使用所有帧的"可选功能面板" | 重启 | ||||
| #2 | 6升 | ñ | 在跟踪之前检入以将所有跟踪结果存储在内存中,以备将来参考 | 历史追踪 | ||||
| #2 | 7 | ÿ | 点击开始跟踪 | 跟踪及分析 | ||||
| #2 | 8A | ñ | 点击开始跟踪蛋白质通道强度 | 分析蛋白质浓度 | ||||
| #2 | 8B | ñ | 检查沿轨道长度跟踪蛋白质通道强度 | 全丝酵母 | ||||
| #2 | 8C | ñ | 检查参考蛋白或蛋白A的跟踪 | A蛋白 | ||||
| #2 | 8D | ñ | 检查跟踪蛋白B | ProteinB | ||||
| #2 | 8E | ñ | 检查跟踪蛋白质C | P53复合体蛋白 | ||||
| #2 | 8F | ñ | 检查以跟踪平均蛋白质强度提示 | 领先提示 | ||||
| #2 | 8克 | ñ | 输入提示的长度 | 提示长度 | ||||
| #2 | 8小时 | ñ | 输入从底部开始形成尖端的最小距离 | 阈 | ||||
| #2 | 8I | ñ | 点击将顶尖分析结果保存到文件 | 按钮 | ||||
| #2 | 9A | ñ | 点击启动比率蛋白质分析 | 比较 | ||||
| #2 | 9B | ñ | 入住比较蛋白B相对于A | 日志10(B / A) | ||||
| #2 | 9C | ñ | 入住以比较蛋白C相对于A | 日志10(C / A) | ||||
| #2 | 9D | ñ | 入住以比较蛋白B相对于A的尖端 | 日志10(B / A) | ||||
| #2 | 9E | ñ | 入住以比较蛋白C相对于A的尖端 | 日志10(C / A) | ||||
| #2 | 10A | ñ | 选择其他颜色地图(默认值:Jetplot) | 彩色地图 | ||||
| #2 | 10B | ñ | 编辑色彩图 | 编辑色彩图 | ||||
表1:GUI Windows#1和#2中存在的所有功能概述。
一旦完成,该程序将创建一个凸包,并自动跟踪整个电影中的提示。从电影中提取的参数,比如比例计算器,生长速度和光线长度a重新显示并存储在工作文件夹中作为图像和数据文件。然后可以从存储的数据文件中提取和进一步分析其他参数,如光合生存期,生长速率和收缩率( 图2B )。

图2:使用图像分析软件的图形用户界面。 ( A )GUI窗口#1用于加载和处理图像。该程序可以加载多达3个蛋白质通道,从而成对比较2个通道。窗口具有强制性(蓝色)和可选功能(绿色),用于在跟踪之前预处理图像( B )GUI窗口2用于跟踪光点以及时空和比率 - 度量蛋白质分析。再次,可选功能标记为绿色。这个数字已经被修改了参考文献12 。 请点击此处查看此图的较大版本。
{kind=link}
在这里,我们提供了一个详细的样品制备和软件处理协议。我们从培养细胞和获取优化图像分析的电影的详细说明开始。本部分数据采集之后是操作图像分析软件的详细说明。在整个协议中,我们介绍了在收集和处理数据时应该考虑的关键步骤和可选功能。最后,我们利用图像分析软件分析来自不同模型系统的虚拟障碍,然后将所描述的图像分析软件与可用于虚拟量化的其他程序进行比较,并讨论限制和未来方向。
研究方案
细胞培养
- 含有4.5g / L D-葡萄糖,L-丙氨酸-L-谷氨酰胺二肽,丙酮酸,10%胎牛血清和10单位/ ml青霉素/链霉素的Dulbecco改良Eagle培养基(DMEM)中的培养HeLa或COS细胞。补充有0.5mM L-丙氨酸-L-谷氨酰胺二肽,无血清神经元补充剂和10单位/ ml青霉素/链霉素的培养基中不含L-谷氨酰胺,谷氨酸或天冬氨酸的培养神经元。
- 一旦达到40%汇合,按照制造商的说明书,使用转染试剂转染选择的构建体。将转染细胞保持在37℃和5%CO 2的培养箱中15-18小时。
- 为了减少图像采集期间的pH和渗透压(由于蒸发)的变化,用含有20mM 4-(2-羟乙基)-1的37℃预热培养基将细胞培养室(成像前)填充至90% - 哌嗪乙磺酸(HEPES)。密封盖子上盖上一层薄薄的真空润滑脂,轻轻按压在含有转染细胞的培养室上。
图像采集
注意:丝状伪足的长度从2-10μm13变化。 Filopodia以0.05-0.1μm/ s的平均速度生长13,14 。
- 使用60X或100X物镜的显微镜拍摄图像,无需像素分类(这里是旋转盘共聚焦显微镜)。使用大于1赫兹(Hz)的采集率来跟踪斑点动力学。为了最小化离焦伪像,靠近基底膜( 即衬底表面)的图像伪迹。
- 为了保证平滑跟踪,调整摄像机的曝光时间和激光强度,使信噪比(SNR)大于4.避免个别通道饱和( 即像素值255位为8位图像,65,535为16位图像),因为这将排除随后的图像分析。
- 为了避免渗色,只能使用与激光线和显微镜的过滤器兼容的荧光标记(详见参考文献15 )。
图像预处理
注意:使用ImageJ或其他可用的软件预处理图像16,17 。
- 如果样品正在移动,则在分析之前使用可用的软件( 例如 https://github.com/NMSchneider/fixTranslation-Macro_for_ImageJ)纠正横向漂移。排除具有轴向偏移的电影(即沿z方向移动)。
- 正确的背景( 即细胞外区域的灰度值),漂白( 即如果荧光蛋白损伤引起的荧光强度持续损失)和可能的出血槽( 即来自一种感染的信号)两个通道中的荧光探针)使用可用的软件( 例如 http://imagej.net/Category:Plugins和16,17 )。
注意:单个通道的荧光强度不会被软件改变。 - 为了确保后续的图像分析,将工作文件夹中的".tiff"堆叠文件格式的灰度级特定蛋白质通道对应的电影保存。
注意:所有通道的堆栈的尺寸( 即大小,长度)必须相同。
4.图像分析 - 步骤1:加载图像
注意:这里描述的软件是用Matlab(简称编程软件)编写的,只能运行这个程序。
- 从以下站点下载包含用于图像分析的所有必需文件的压缩文件夹:https://campus.uni-muenster.de/en/einrichtungen/impb0/nanoscale-forces-in-cells/softw是/。将文件解压缩并复制到工作文件夹中。
- 安装完成后,打开编程软件并运行'filopodiaAnalysisM3.fig'。 GUI窗口#1将打开, 如图2A所示。
- 使用蛋白质A的<1b>,蛋白质B的<1c>,蛋白C的<1d>加载对应于GUI窗口#1中的特定蛋白质的保存的堆积的'.tiff'文件。参见图2和表1详细信息。
注意:GUI窗口#1中显示的蛋白A作为最终比例分析的参考通道。 - 通过点击<1a>从蛋白质通道创建单元格的叠加图像。
- 单击<2a>分配用于分析的最后一帧的第一个帧和<2b>。
- 可选地,使用按钮<2h>裁剪包含感兴趣的感兴趣区域的感兴趣区域(ROI),旋转图像u唱按钮<2i>,或使用自由绘图工具<2j>删除不需要的区域。
注意:为了简化图像分析,建议使用ImageJ将ROI( 即裁剪,旋转,删除)与其他预处理步骤(背景减除,漂白校正等)隔离开来。 - 移动每个框架的滑块<2c>进行质量控制,并检查整个影片中是否保持清晰可见。
5.图像分析 - 步骤2:生成跟踪
- 点击GUI窗口#1中的按钮<3a>打开GUI窗口#2( 见图2B )。
- 点击GUI窗口#2中的按钮<4>,生成叠加单元格的蒙版(点击<1a>后在GUI窗口#1中生成)。该程序还生成掩模的边界,其中实施了凸包以获得顶点。
- CLIck按钮<5>;将出现一个光标。使用光标选择基底(从其中测量细丝尖端的距离),然后在其首次出现的框架中的丝状伪足的尖端。为此,在窗口#2中移动滑块。
注意:为了最大限度地减少数据输出中的错误,请将基准点沿着轴线垂直放置在光栅尖端的下方。将基点定位在其他位置( 例如横向偏移)可能在细胞体内的荧光值中引入方向偏置。 - 使用<6c>选择阈值长度(高于该长度的丝状伪足会弯曲)。
注意:该长度定义为基点(使用<5>选择)与细胞体的边界( 即 ,丝状伪足开始生长的区域)之间的距离。测量的距离是以像素为单位。 - 在<6a>框中指定用于近似丝状伪迹形状的段数。
注意:最少数量的片段取决于光合丝达到的最大长度,而且还如何弯曲。不要选择大于基座和尖端之间像素数的段数( 即阈值长度)。选择比步骤5.4中定义的阈值更多的段。 ( 即基数和提示之间的像素数)将导致对光栅长度的过高估计。 - 指定扫描宽度,作为放置节点的水平扫描仪,位于<6b>中。
注意:程序使用这些节点来将连接基座和尖端的线与弧形体的主体( 即创建骨架)配合使用。作为起点,将像素值等于最大长度乘以因子 。
。 - 在<6d>框中指定扫描半径(以像素为单位)。使用比ob大约50%的值提供了跨框架尖端位移。
注意:使用非常大的扫描半径可能会在不存在真实尖端尖端( 例如 ,平面外移动或低SNR)的帧中抓取不需要的凸包点。 - 在<6e>框中指定弯曲角度。
注意:角度阈值由整个分析期间钢筋弯曲的最大角度决定。指定角度阈值可帮助软件排除跟踪丝状伪足向细胞体弯曲时从丝状伪足一侧生长的不需要的结构。该程序可靠地工作,具有与垂直轴小于45度的倾斜角度的丝眼伪装。 - 要开始跟踪,请单击GUI窗口#2中的<跟踪和分析>按钮。单击GUI窗口#2中的"历史记录跟踪"框以保存整个跟踪协议,以备将来参考。
注意:跟踪过程完成后,每个框架中的圆顶的长度存储在"dynamics.xlsx"的"length_vel"表格供将来参考。同样,所有其他跟踪参数都存储在"dynamics.xlsx"的名为"parameters"的工作表中。 - 或者,如果在所有帧中没有自动检测到虚拟网元,请使用以下步骤手动更正它。
- 使用窗口#2中的滑块访问相应的框架,并手动选择顶点。
注意:在GUI窗口#2的跟踪窗口中的蓝色区域将表示没有检测到凸包点的框架。在启动跟踪程序以访问该帧中的节点坐标之前,必须先检查"历史记录跟踪"按钮。 - 在GUI窗口#2中使用<6f>选择参考点(基础后跟提示)。指定其他参数,如"扫描长度","精确测量后"和"最大弯曲角度";对于该帧,如步骤5.4-5.8所述。
- 点击<6i>按钮保存新的参数(特定于该帧)。对由蓝色区域指示的所有帧重复这些步骤。
- 完成后,使用<7>重新初始化跟踪程序。
注意:如果程序在一个电影中突然从一个文字转换到另一个文字,也可以使用此修正。另一种方法是考虑修剪包含多个信封的电影。
- 使用窗口#2中的滑块访问相应的框架,并手动选择顶点。
图像分析 - 步骤3:时空蛋白质分析
- 对于时空分析,选择由<8b>代表的框,然后选择感兴趣的蛋白质通道(<8c>和/或<8d>和/或<8e>)。
注意:由于使用蛋白A作为参考,因此在比例分析(<9a>,<9b>和/或<9c>)之前必须选择蛋白A。 - 点击<8a>使用步骤5.9中生成的痕迹沿着丝状体长度启动蛋白质跟踪。
7.图像分析 - 步骤4:比率 - 度量蛋白质分析
- 复选框<9b>或<9c>,然后单击按钮<9a>获得时空比例图。
注意:为了避免相对蛋白质浓度的错误表示,比例图像不绘制为X / Y,而是作为log(X / Y)。为了将来使用,比例图将以".png"和".fig"格式文件导出,原始绘图数据存储在"dynamics.xlsx"文件中。
8.图像分析 - 步骤5:虚拟提示分析
- 复选框<8f>,并使用<8g>和<8h>从底部指定刀尖长度和阈值长度。点击<按钮>将数据保存到文件'dynamics.xlsx'。点击<分析蛋白质强度>来生成痕量蛋白质强度的斑点尖端和保存比例分析。
注意:提示长度决定了用于分析的尖端的像素数。该软件将返回每个帧顶端像素的平均强度值。阈值长度决定了从底部到尖端的最小距离,超过该最小距离开始生长(斑点尖端)(当程序开始跟踪尖端的强度值时)。所以尖端长度必须小于阈值长度。 - 点击所需的比例,使用<9d>或<9e>进行分析。
- 点击比较按钮<9a>生成比例数据。
结果
使用用丝状肌动蛋白(f-tractin 18 ,红色)标记和细胞溶质参考(绿色)转染的COS细胞,我们发现肌动蛋白丰富的丝状体突起( 图3A ,上图 )。时间序列表明,丝状伪足迅速伸缩( 图3A ,中间图 )。使用图像分析软件,我们跟踪单个的虚拟眼镜。通过手测量的光?...
讨论
在这里,我们提出了一个详细的协议,用于通过凸壳算法跟踪丝状体生长动力学和这些动态结构中相对蛋白质浓度的分析。使用该软件,可以在单次运行中将多达3个通道成对比较,从而在整个延伸/缩回循环中确定两个通道( 即蛋白质)的相对浓度,并将其作为图像和数据文件存储在不同的文件夹中。除了常规操作之外,软件还提供了许多可以进行修改的参数,以优化相应实验的分析。这?...
披露声明
作者没有什么可以披露的。
致谢
作者承认了DFG(EXC-1003至MG)的资助。
材料
| Name | Company | Catalog Number | Comments |
| DMEM | Life Technologies | 31966-021 | |
| 10% Fetal bovine serum | Biochrom AG | L11-044 | |
| Lipofectamine 2000 | Life Technologies | 11668-027 | |
| 1% penicillin/streptomycin | Biochrom AG | 12212 | |
| Neurobasal Medium | Life Technologies | 21103-049 | |
| B27 | Life Technologies | 17504-044 | |
| HEPES (1M stock solution) | Life Technologies | 15630 | |
| Citrine-N1 | Addgene | 54593 | |
| Labtech | Thermo | 155411 | |
| Glutamax-I | Thermo | 35050-061 | |
| Hela | Leibniz Institute DSMZ | ACC-57 | |
| COS 7 | Leibniz Institute DSMZ | ACC-60 | |
| 3T3 cells | Leibniz Institute DSMZ | ACC-59 | |
| Microscope | Nicon Eclipse | ||
| Camera | Andor | DU888 Ultra | |
| Confocal Unit | Yokagawa | CSU-X1 | |
| Pyruvate | Gibco | 31966-021 |
参考文献
- Dunaevsky, A., Tashiro, A., Majewska, A., Mason, C., Yuste, R. Developmental regulation of spine motility in the mammalian central nervous system. Proc Natl Acad Sci U S A. 96 (23), 13438-13443 (1999).
- Matus, A., Brinkhaus, H., Wagner, U. Actin dynamics in dendritic spines: a form of regulated plasticity at excitatory synapses. Hippocampus. 10 (5), 555-560 (2000).
- Galic, M., et al. Dynamic recruitment of the curvature-sensitive protein ArhGAP44 to nanoscale membrane deformations limits exploratory filopodia initiation in neurons. Elife. 3, e03116 (2014).
- Hotulainen, P., et al. Defining mechanisms of actin polymerization and depolymerization during dendritic spine morphogenesis. J Cell Biol. 185 (2), 323-339 (2009).
- Leijnse, N., Oddershede, L. B., Bendix, P. M. Helical buckling of actin inside filopodia generates traction. Proc Natl Acad Sci U S A. 112 (1), 136-141 (2015).
- Tsygankov, D., et al. CellGeo: a computational platform for the analysis of shape changes in cells with complex geometries. J Cell Biol. 204 (3), 443-460 (2014).
- Xiong, Y., et al. Automated characterization of cell shape changes during amoeboid motility by skeletonization. BMC Syst Biol. 4, 33 (2010).
- Styner, M., Gerig, G., Lieberman, J., Jones, D., Weinberger, D. Statistical shape analysis of neuroanatomical structures based on medial models. Med Image Anal. 7 (3), 207-220 (2003).
- Blum, H., Wathen-Dunn, W. A transformation for extracting new descriptors of shape. Models for the Perception of Speech and Visual Form: Proceedings of a Symposium. , 362-380 (1967).
- Barry, D. J., Durkin, C. H., Abella, J. V., Way, M. Open source software for quantification of cell migration, protrusions, and fluorescence intensities. J Cell Biol. 209 (1), 163-180 (2015).
- Machacek, M., et al. Coordination of Rho GTPase activities during cell protrusion. Nature. 461 (7260), 99-103 (2009).
- Saha, T., et al. Automated analysis of filopodial length and spatially resolved protein concentration via adaptive shape tracking. Mol Biol Cell. 27 (22), 3616-3626 (2016).
- Argiro, V., Bunge, M. B., Johnson, M. I. A quantitative study of growth cone filopodial extension. J Neurosci Res. 13 (1-2), 149-162 (1985).
- Mogilner, A., Rubinstein, B. The physics of filopodial protrusion. Biophys J. 89 (2), 782-795 (2005).
- Shaner, N. C., Steinbach, P. A., Tsien, R. Y. A guide to choosing fluorescent proteins. Nat Methods. 2 (12), 905-909 (2005).
- Abramoff, M. D., Magalhaes, P. J., Ram, S. J. Image Processing with ImageJ. Biophotonics International. 11, 36-42 (2004).
- Courtney, J., Woods, E., Scholz, D., Hall, W. W., Gautier, V. W. MATtrack: A MATLAB-Based Quantitative Image Analysis Platform for Investigating Real-Time Photo-Converted Fluorescent Signals in Live Cells. PLoS One. 10 (10), e0140209 (2015).
- Schell, M. J., Erneux, C., Irvine, R. F. Inositol 1,4,5-trisphosphate 3-kinase A associates with F-actin and dendritic spines via its N terminus. J Biol Chem. 276 (40), 37537-37546 (2001).
- Korobova, F., Svitkina, T. Molecular architecture of synaptic actin cytoskeleton in hippocampal neurons reveals a mechanism of dendritic spine morphogenesis. Mol Biol Cell. 21 (1), 165-176 (2010).
- Cheadle, L., Biederer, T. The novel synaptogenic protein Farp1 links postsynaptic cytoskeletal dynamics and transsynaptic organization. J Cell Biol. 199 (6), 985-1001 (2012).
- Tarnok, K., et al. A new tool for the quantitative analysis of dendritic filopodial motility. Cytometry A. 87 (1), 89-96 (2015).
- Hendricusdottir, R., Bergmann, J. H. F-dynamics: automated quantification of dendrite filopodia dynamics in living neurons. J Neurosci Methods. 236, 148-156 (2014).
- Fanti, Z., Martinez-Perez, M. E., De-Miguel, F. F. NeuronGrowth, a software for automatic quantification of neurite and filopodial dynamics from time-lapse sequences of digital images. Dev Neurobiol. 71 (10), 870-881 (2011).
- Costantino, S., et al. Semi-automated quantification of filopodial dynamics. J Neurosci Methods. 171 (1), 165-173 (2008).
- Nilufar, S., Morrow, A. A., Lee, J. M., Perkins, T. J. FiloDetect: automatic detection of filopodia from fluorescence microscopy images. BMC Syst Biol. 7, 66 (2013).
转载和许可
请求许可使用此 JoVE 文章的文本或图形
请求许可探索更多文章
This article has been published
Video Coming Soon
版权所属 © 2025 MyJoVE 公司版权所有,本公司不涉及任何医疗业务和医疗服务。