
需要订阅 JoVE 才能查看此. 登录或开始免费试用。
Method Article
用化学发光法实时检测水稻免疫反应中活性氧的产生
摘要
在这里,我们描述了一种实时检测病原体相关分子模式触发的免疫反应中水稻组织中质外活性氧(ROS)产生的方法。该方法简单、标准化,可在受控条件下产生高度可重复的结果。
摘要
活性氧(ROS)在各种生物过程中起着至关重要的作用,包括非生物和生物胁迫的传感。在病原体感染或病原体相关化学物质(病原体相关分子模式[PAMP])攻击时,一系列免疫反应(包括ROS爆发)在植物中迅速诱导,这称为PAMP触发免疫(PTI)。ROS爆发是一种标志性的PTI反应,由一组质膜定位的NADPH氧化酶 - RBOH家族蛋白催化。绝大多数ROS由过氧化氢(H2O2)组成,可以通过基于鲁米诺的化学发光方法轻松稳定地检测。化学发光是一种产生光子的反应,其中鲁米诺或其衍生物(如L-012)在催化剂的作用下与ROS进行氧化还原反应。本文介绍了一种优化的基于L-012的化学发光方法,用于在水稻组织中引发PAMP时实时检测质外体ROS的产生。该方法简单、稳定、标准化,在严格控制的条件下具有高度可重复性。
引言
活性氧 (ROS) 包括一系列化学活性氧衍生物,包括超氧阴离子自由基 (O2-) 及其衍生物、羟基自由基 (OH-)、过氧化氢以及单线态氧或氧化还原反应的产物,它们在质体和叶绿体、线粒体、过氧化物酶体和其他亚细胞位置中不断产生1.ROS在许多生物过程中起着重要作用,对所有植物都是必不可少的2,3,4。ROS功能的广泛范围从生长发育的调节到非生物和生物胁迫的感知5,6,7,8。
在植物免疫系统中,植物细胞质膜定位的受体 - 所谓的模式识别受体(PRR) - 感知病原体衍生的化学物质 - 病原体相关分子模式(PAMP)。这种识别触发了一系列快速免疫反应,包括钙内流、ROS 爆发和 MAPK 级联反应;因此,这一层免疫被称为PAMP触发免疫(PTI)。ROS突发是PTI响应的标志性特征,其测定被广泛用于PTI相关研究9,10。由 PAMP 触发的 ROS 产生归因于质膜驻留的 NADPH 氧化酶或呼吸爆发氧化酶同系物 (RBOH) 家族蛋白,其将电子从胞质 NADPH 或 NADH 转移到细胞外氧以产生超氧化物 (O 2-),超氧化物通过超氧化物歧化酶自发转化为过氧化氢 (H 2 O 2)8.PAMP触发的ROS突发非常迅速,在PAMP治疗后几分钟出现,并在~10-12分钟达到峰值。绝大多数ROS分子由过氧化氢(H2O2)组成,可以通过化学发光测定轻松稳定地检测。
在化学发光中,化学发光试剂在催化剂的作用下与活性氧反应,产生激发态中间体。然后,产物中的电子通过非辐射跃迁返回基态并发射光子。常见的化学发光试剂包括鲁米诺和L-012,其中鲁米诺在应用中占主导地位11,12,13。然而,越来越多的研究人员选择L-012来检测ROS的产生,因为与鲁米诺相比,L-012在中性或接近中性的pH条件下具有更高的发光效率。
本文介绍了一种基于L-012的优化化学发光方法,用于实时检测水稻(水稻)组织-叶盘和鞘中PAMPs的产生。本文提供的方法简单、稳定、标准化,适应性强,可满足不同的实验需求。使用这种方法获得的数据在严格控制的条件下具有高度可重复性。
研究方案
注意:该方案适用于不同的植物组织。在该协议中,水稻鞘和叶盘用于PANP诱发时的ROS检测。由于差异主要由取样方法引起,因此下面仅介绍通用程序,并在必要时提及具体步骤。
1. 植物培养
- 用70%乙醇对去壳的水稻种子灭菌1分钟,然后用40%次氯酸钠(NaClO)灭菌1小时。然后,用无菌水冲洗种子5倍以去除余氯。
- 将种子无菌地铺在 1/2 MS 培养基上(2.37 g/L Murashige 和 Skoog (MS) 培养基、30 g/L 蔗糖、2.1 g/L 植物胶、pH 5.7,高压灭菌)。
- 在水稻鞘法中,用MS培养基直接将种子板在无菌玻璃容器中。
- 在叶盘法中,将种子铺在MS平板上5-7天,然后将它们移植到生长基质或土壤中(图1A)。
- 在具有12小时光/ 12小时暗光周期的生长室中生长幼苗。
2. 组织制备和预处理
- 水稻鞘
- 在ROS测定前1天,用锋利的剃须刀片或手术刀片将10天龄水稻幼苗的鞘切成3毫米的段进行预处理(图1B)。
- 将五个鞘段置于含有100μLddH 2 O的96孔微量滴定板的单个孔中,在25°C的黑暗中10-12小时,这使得伤口损伤相关的离子泄漏和防御反应减弱(图2)。
注意:注意保持切割垂直以确保暴露在启发溶液中的一致切割表面积是获得高度可重复结果的重要步骤。轻轻移动线段。不要在段上做额外的切割或伤口,这可能是数据变化的来源。原则上,每个测试必须包含至少五个重复,因为ROS值的变化很大。重复集越多,数据越可靠。
- 叶盘
- 使用带柱塞的活检打孔器从 4-6 周龄的水稻植株上切下叶盘(直径 4 毫米)。始终从主分蘖的第二片叶子的中间三分之一(从顶部编号)切下叶盘,以减少数据变化(图1C)。
- 将一个叶盘放入含有 100 μL ddH 2 O 的 96 孔微量滴定板的单个孔中 10-12 小时进行预处理,这使得与损伤相关的反应减弱,因为这些反应可能会干扰 PAMP 诱导 ROS(图 2)。
注意:轻轻操作叶盘。请勿在实验中对椎间盘进行额外的切割或伤口,这可能会导致数据变化。ROS的诱导主要发生在切割边缘的细胞中,因为水稻组织(叶子或鞘)的表面覆盖着疏水层。只有切割边缘的细胞与引出溶液接触(请参阅讨论部分)。 - 将所有叶盘漂浮在微量滴定板的孔中,远轴表面朝上,以进行水预处理,以避免叶侧相关的变化。

图1:用于鞘取样的水稻幼苗的生长条件和阶段以及测定中使用的部分水稻鞘和水稻叶。 (A)在无菌条件下在1/2MS培养基上生长10天的稻苗可以采样用于ROS测定。将灭菌的水稻种子在1/2MS培养基上培养,并在透明玻璃小瓶中以12小时光照/ 12小时黑暗光周期生长,直径为8.5cm,高度为15cm。(二)叶鞘取样部分示意图。叶鞘是从10日龄的水稻幼苗上切下来的。叶鞘的位置在根部上方和第一片叶子下方。(三)叶盘取样位置示意图。在任何生长阶段,叶盘都可以从健康水稻植物主分蘖的第二片叶子的中间三分之一(从顶部开始计数)切割。缩写:ROS = 活性氧;MS = Murashige 和 Skoog。 请点击此处查看此图的大图。
{kind=link}

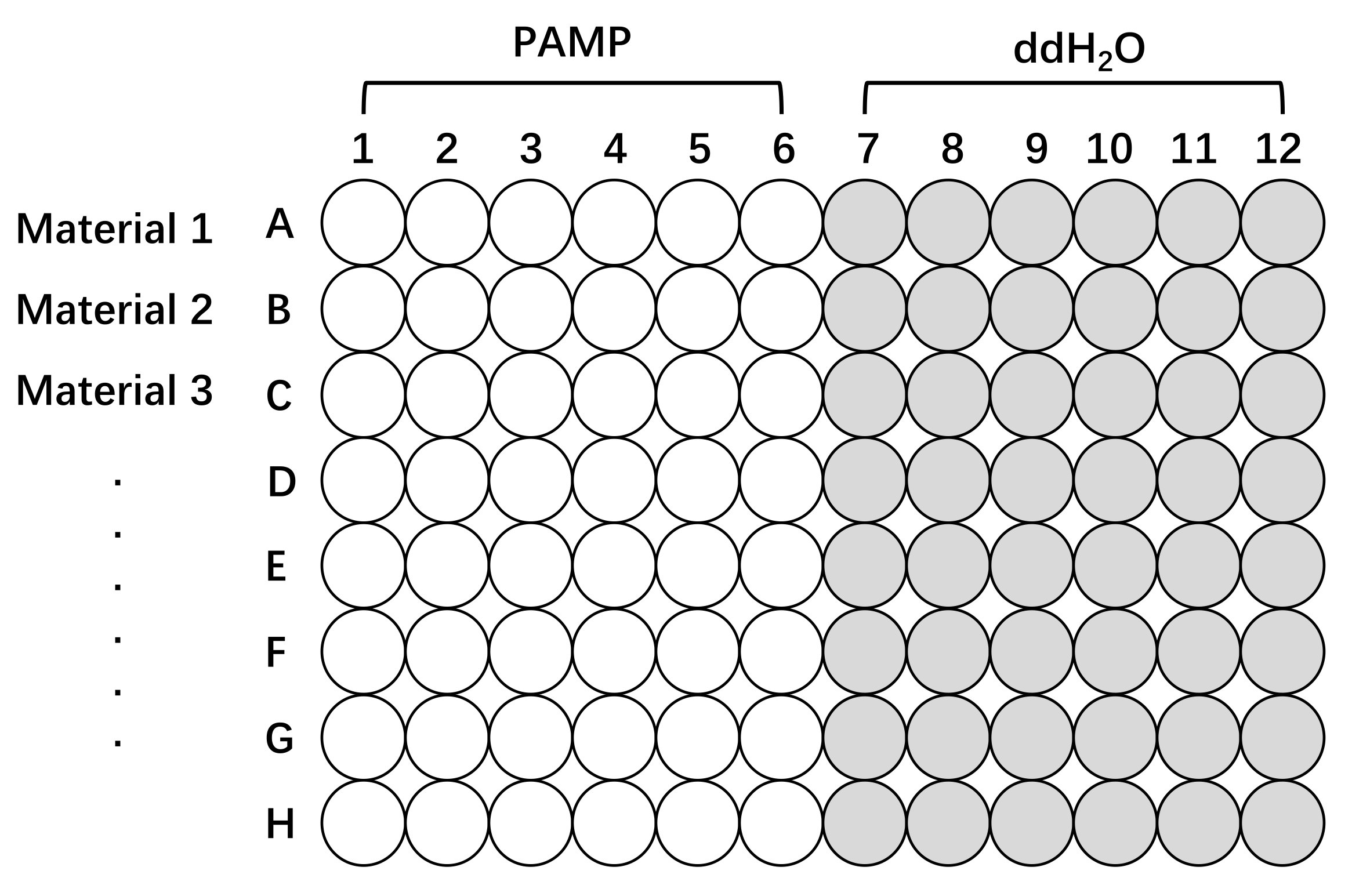
图 2:使用不同生产线的 Oryza sativa 测量 ROS 生产的板设置示意图。 使用96孔板对大米组织进行预处理和测试。第 1 行、第 2 行和第 3 行(一个板上最多 8 行)可以是任何感兴趣的材料、不同的品种、突变体或转基因系。用含有PAMP(PAMP,白色)或不具有PAMP(ddH2O,灰色)的诱发溶液刺激组织以测量ROS反应。应该注意的是,要测试的样品越多,读数之间的时间间隔就越长。缩写:ROS = 活性氧;PAMP = 病原体相关分子模式;ddH2O = 双蒸水。 请点击此处查看此图的大图。
{kind=link}
3. 准备启发溶液
- 将 L-012 粉末溶解到 20 mM (6.23 mg/mL) 含 ddH2O 的水溶液中,制成储备溶液。然后,用50mM Tris HCl缓冲液(pH 7.5)稀释储备溶液,使工作溶液的最终浓度为500μM L-012。使用前保持储备溶液冷冻并稀释至工作溶液。
- 制备含有 PAMP、L-012 和辣根过氧化物酶 (HRP;10 mg/mL 中的 ddH2O)的诱发溶液。对于 10 mL 启发溶液,混合 9.4 mL 50 mM Tris HCl (pH 7.5) 溶液、400 μL L-012 溶液、100 μL HRP 和 100 μL flg22(PAMP; 10 mM 在 ddH2O 中)。对于阴性对照,加入 100 μL ddH2O 代替 PAMP。
注意:将准备好的引出溶液保持在室温下,以避免对稻米组织的冷应激。其他PAMP也可根据需要用于治疗,例如甲壳素(终浓度为20 ng/mL)。由于L-012对光敏感,因此请用铝箔覆盖所有含有L-012溶液的试管。
4.启动软件并使用参考的酶标仪设置实验方案(参见材料表)
注意:设置酶标仪软件的参数需要一些时间。建议在添加启发解决方案之前准备好机器和协议(一键继续)。
- 启动软件。单击 "实验 "按钮以创建新协议或使用现有协议。
- 单击弹出窗口中的 程序 以设置板。从要监测的板中选择要监测的孔。
- 单击启动动力学以设置总运行时间和读取间隔。将运行时间设置为 35 分钟或更长,具体取决于实验要求。要尽可能频繁地获取读数,请选择最小间隔。对于积分时间,请选择 1 秒或更长,具体取决于信号强度。
注意:读取间隔取决于样本数量和信号积分持续时间。 - 单击 "验证 |确定 以确认设置。
- 在弹出窗口中单击检测新板,然后等待软件提示加载 板 对话框。将要测试的板放在托架上。
- 停在这里等待启发系统的建立(在下一节中)。启发系统准备就绪后,单击" 运行 "开始读取。
5. 建立启发系统并测量实时ROS生产
- 小心地从含有预处理组织的孔中取出ddH2O,避免任何组织损伤或干燥。
- 使用多通道移液器将 200 μL 的启发溶液添加到含有组织的孔中。
- 轻轻摇晃混合。单击" 运行 "开始检测。
注意:通过PAMP处理,植物组织反应并产生ROS非常快。因此,建议在有多种治疗时,首先处理没有PAMP的阴性对照以减少手术时间。尽快操作以减少治疗之间的引出延迟。从添加启发溶液到开始检测之间的时间越短,重要实验数据的捕获就越好。
结果
在这里,我们以大米材料为例来确定flg22处理产生的ROS。引出后ROS的产生是短暂的。在水稻中,ROS产量的增加首先在1-2分钟内检测到,在10-12分钟达到峰值,并在~30-35分钟内恢复到基线(图3)。对照试验中,诱发溶液中没有PAMP,导致没有明显的ROS诱导,与对照测试相比,只有当含有flg22或其他PAMP(如几丁质)的诱发溶液时,才会诱导特定的ROS爆发。同时,ROS的总量可以从曲?...
讨论
本研究的目的是建立一种高效的方法来量化水稻组织中PAMP响应的早期ROS产生。该方法为实时测定由处理过的大米组织产生的质外体ROS提供了一种标准化程序。该方法操作简单,成本低,成分清晰,独立于商业试剂盒。使用这种方法,研究人员可以研究植物受到生物或非生物胁迫时质外体ROS的实时产生。
在该协议中,L-012被选为化学发光试剂,因为它是一种无毒化学物质。鲁?...
披露声明
作者没有利益冲突需要披露。
致谢
这项工作得到了 上海市自然科学基金(批准号:21ZR1429300/BS1500016)、上海交通大学(Agri-X计划,批准号:AF1500088/002)、上海市农业种子协同创新中心(批准号:ZXWH2150201/001)对范江波的资助,以及上海交通大学医学工程合作项目(批准号:21X010301734)对李灿的资助。
材料
| Name | Company | Catalog Number | Comments |
| 96-well microtiter plate | WHB | WHB-96-01 | |
| Ethanol absolute | Innochem | A43543 | |
| flg22 | Sangon Biotech | p20973 | PAMP |
| Gen5 | BioTek | software | |
| L-012 | FUJIFILM | 120-04891 | 8-amino-5-chloro-7-phenyl-2,3-dihydropyrido [3,4-d] pyridazine-1,4-dione, CAS #:143556-24-5 |
| Microplate reader | BioTek | Synergy 2 | |
| MS Medium | Solarbio | M8521 | |
| NaCLO | Aladdin | S101636 | |
| Peroxidase from horseradish (HRP) | Sigma | P8375 | |
| Phytagel | Sigma | P8169 | |
| Sampler | Miltex | 15110-40 | |
| Sucrose | Sangon Biotech | A502792 | |
| Tris | Sangon Biotech | A610195 |
参考文献
- Gechev, T. S., Van Breusegem, F., Stone, J. M., Denev, I., Laloi, C. Reactive oxygen species as signals that modulate plant stress responses and programmed cell death. Bioessays. 28 (11), 1091-1101 (2006).
- Mittler, R. ROS are good. Trends in Plant Science. 22 (1), 11-19 (2017).
- Gilroy, S., et al. ROS, calcium, and electric signals: key mediators of rapid systemic signaling in plants. Plant Physiology. 171 (3), 1606-1615 (2016).
- Mittler, R., Vanderauwera, S., Gollery, M., Van Breusegem, F. Reactive oxygen gene network of plants. Trends in Plant Science. 9 (10), 490-498 (2004).
- Marino, D., Dunand, C., Puppo, A., Pauly, N. A burst of plant NADPH oxidases. Trends in Plant Science. 17 (1), 9-15 (2012).
- Mittler, R., Zandalinas, S. I., Fichman, Y., Van Breusegem, F. Reactive oxygen species signalling in plant stress responses. Nature Reviews Molecular Cell Biology. 23 (10), 663-679 (2022).
- Suzuki, N., Koussevitzky, S., Mittler, R., Miller, G. ROS and redox signalling in the response of plants to abiotic stress. Plant, Cell & Environment. 35 (2), 259-270 (2012).
- Suzuki, N., et al. Respiratory burst oxidases: the engines of ROS signaling. Current Opinion in Plant Biology. 14 (6), 691-699 (2011).
- Kadota, Y., Shirasu, K., Zipfel, C. Regulation of the NADPH oxidase RBOHD during plant immunity. Plant and Cell Physiology. 56 (8), 1472-1480 (2015).
- Segonzac, C., Zipfel, C. Activation of plant pattern-recognition receptors by bacteria. Current Opinion in Microbiology. 14 (1), 54-61 (2011).
- Roda, A., et al. Progress in chemical luminescence-based biosensors: A critical review. Biosensors and Bioelectronics. 76, 164-179 (2016).
- Hong, D., Joung, H. -. A., Lee, D. Y., Kim, S., Kim, M. -. G. Attomolar detection of cytokines using a chemiluminescence immunoassay based on an antibody-arrayed CMOS image sensor. Sensors and Actuators B: Chemical. 221, 1248-1255 (2015).
- Nishinaka, Y., et al. et al. new sensitive chemiluminescence probe, L-012, for measuring the production of superoxide anion by cells. Biochemical and Biophysical Research Communications. 193 (2), 554-559 (1993).
- Grundy, J., Stoker, C., Carre, I. A. Circadian regulation of abiotic stress tolerance in plants. Frontiers in Plant Science. 6, 648 (2015).
转载和许可
请求许可使用此 JoVE 文章的文本或图形
请求许可探索更多文章
This article has been published
Video Coming Soon
版权所属 © 2025 MyJoVE 公司版权所有,本公司不涉及任何医疗业务和医疗服务。