
このコンテンツを視聴するには、JoVE 購読が必要です。 サインイン又は無料トライアルを申し込む。
Method Article
化学発光アッセイによるイネの免疫応答における活性酸素種産生のリアルタイム検出
要約
ここでは、病原体関連分子パターン誘発免疫応答におけるイネ組織における遊性活性酸素種(ROS)産生をリアルタイムに検出する方法について述べる。この方法はシンプルで標準化されており、制御された条件下で再現性の高い結果を生成します。
要約
活性酸素種(ROS)は、非生物的および生物的ストレスの感知を含む、さまざまな生物学的プロセスにおいて重要な役割を果たします。病原体の感染または病原体関連化学物質(病原体関連分子パターン[PAMP])への挑戦により、ROSバーストを含む一連の免疫応答が植物に迅速に誘導され、これはPAMPトリガー免疫(PTI)と呼ばれます。ROSバーストは特徴的なPTI応答であり、原形質膜に局在するNADPHオキシダーゼ(RBOHファミリータンパク質)のグループによって触媒されます。ROSの大部分は過酸化水素(H2O2)を含み、ルミノールベースの化学発光法によって容易かつ着実に検出することができる。化学発光は、ルミノールまたはその誘導体(L-012など)が触媒の作用下でROSと酸化還元反応を起こす光子生成反応です。本論文では、イネ組織におけるPAMP誘発時にアポプラストROS産生をリアルタイムで検出するための最適化されたL-012ベースの化学発光法について説明します。この方法は、しっかりと制御された条件下で、簡単で、安定しており、標準化されており、再現性が高いです。
概要
活性酸素種(ROS)は、スーパーオキシドアニオンラジカル(O2-)およびその誘導体、ヒドロキシルラジカル(OH-)、過酸化水素、および一重項酸素または酸化還元反応の生成物を含む一連の化学的に活性な酸素誘導体を含み、色素体および葉緑体、ミトコンドリア、ペルオキシソーム、およびその他の細胞内位置で絶えず生成されます1。.ROSは多くの生物学的プロセスにおいて重要な役割を果たしており、すべての植物にとって不可欠です2,3,4。ROS機能の幅広いスペクトルは、成長と発達の調節から非生物的および生物的ストレスの知覚までさまざまです5,6,7,8。
植物免疫系では、植物細胞原形質膜に局在する受容体、いわゆるパターン認識受容体(PRR)が病原体由来の化学物質-病原体関連分子パターン(PAMP)を知覚します。この認識は、カルシウム流入、ROSバースト、MAPKカスケードなどの一連の高速免疫応答を引き起こします。したがって、この免疫層はPAMPトリガー免疫(PTI)と呼ばれます。ROSバーストは特徴的なPTI応答であり、その決定はPTI関連の研究に広く適用されています9,10。PAMPによって引き起こされるROS産生は、細胞質質NADPHまたはNADHから細胞外酸素に電子を移動してスーパーオキシド(O 2-)を生成し、スーパーオキシドジスムターゼ8によって自発的に過酸化水素(H 2 O 2)に変換される原形質膜常在型NADPHオキシダーゼ、または呼吸バーストオキシダーゼホモログ(RBOH)ファミリータンパク質に起因します。.PAMPトリガーROSバーストは非常に速く、PAMP治療後わずか数分で現れ、~10〜12分でピークに達します。ROS分子の大部分は過酸化水素(H 2 O2)を含み、これは化学発光アッセイで容易かつ着実に検出することができる。
化学発光においては、化学発光試薬は活性酸素と反応し、触媒の作用により、励起状態中間体を生成する。その後、生成物中の電子は非放射遷移によって基底状態に戻り、光子を放出する。一般的な化学発光試薬には、ルミノールおよびL-012が含まれ、ルミノールがアプリケーションを支配しています11、12、13。しかし、L-012はルミノールと比較して中性または中性に近いpH条件下ではるかに高い発光効率を有するため、より多くの研究者がROS産生を検出するためにL-012を選択しています。
本稿では,イネ(Oryza sativa)組織-葉板および鞘中のPAMPs誘発後のROS産生をリアルタイムに検出するためのL-012に基づく最適化された化学発光法について述べる。ここで提供される方法は、シンプルで安定しており、標準化されており、さまざまな実験ニーズを満たすために高度に適応可能です。この方法で得られたデータは、しっかりと制御された条件下で再現性が高いです。
プロトコル
注:このプロトコルは、さまざまな植物組織に適用できます。このプロトコルでは、イネの鞘と葉のディスクを、PAMP誘発時のROS検出に使用しました。主にサンプリングの方法によって違いが生じるため、一般的な手順のみを以下に説明し、必要に応じて具体的な手順について説明します。
1.植物培養
- 脱皮したイネの種子を70%エタノールで1分間殺菌し、次に40%次亜塩素酸ナトリウム(NaClO)で1時間殺菌します。次に、種子を滅菌水で5回すすぎ、残留塩素を除去します。
- 種子を1/2 MS培地(2.37 g / Lムラシゲおよびスクーグ(MS)培地、30 g / Lショ糖、2.1 g / Lフィタゲル、pH 5.7、オートクレーブ滅菌)に無菌的にプレートします。
- 稲鞘法では、MS培地で滅菌ガラス容器内の種子を直接播種する。
- リーフディスク法では、種子をMSプレートに5〜7日間プレートし、成長マトリックスまたは土壌に移植します(図1A)。
- 12時間の明/ 12時間の暗日長の成長室で苗を育てます。
2.組織の準備と前処理
- 米鞘
- ROSアッセイの1日前に、10日齢のイネ苗の鞘を鋭利なカミソリの刃または手術用刃で3 mmのセグメントにカットし、前処理を行います(図1B)。
- 100 μLのddH 2 Oを含む96ウェルマイクロタイタープレートの個々のウェルに5つのシースセグメントを25°Cの暗所で10〜12時間配置すると、創傷損傷関連のイオン漏れと防御応答が軽減されます(図2)。
注意: 引き出し溶液にさらされる一貫した切断表面積を確保するために、カットを垂直に保つように注意することは、再現性の高い結果を得るための重要なステップです。セグメントをそっと動かします。データのばらつきの原因となる可能性のあるセグメントに余分な切り傷や傷を付けないでください。原則として、ROS値の変動が大きいため、各検定には少なくとも5回の反復が含まれている必要があります。反復セットが多いほど、データの信頼性は高くなります。
- リーフディスク
- プランジャー付きの生検パンチを使用して、4〜6週齢のイネからリーフディスク(直径4 mm)を切り取ります。データのばらつきを減らすために、常にメイン耕うん機の2番目の葉の中央3分の1(上から番号が付けられた)からリーフディスクを切り取ります(図1C)。
- 前処理のために100 μLのddH 2 Oを含む96ウェルマイクロタイタープレートの個々のウェルに1枚のリーフディスクを10〜12時間置くと、PAMPによるROSの誘導を妨げる可能性があるため、創傷関連の応答が軽減されます(図2)。
注意: リーフディスクを穏やかに操作します。実験では、ディスクに余分な切り傷や傷を付けないでください。ROSの誘導は、イネ組織(葉または鞘)の表面が疎水性層で覆われているため、主に切断端の細胞から発生します。切断エッジのセルのみが誘発溶液と接触しています(ディスカッションセクションを参照)。 - 葉の側面に関連する変動を避けるために、水の前処理のために、すべてのリーフディスクを不軸面を上に向けて浮かせたままにします。

図1:アッセイに用いた鞘サンプリング用の水稲苗の生育条件と段階、および鞘と水稲葉の一部。 (A)1/2MS培地で無菌条件下で10日間生育させたイネ苗をROSアッセイ用にサンプリングすることができる。滅菌したイネ種子を1/2 MS培地で培養し、直径8.5 cm、高さ15 cmの透明なガラスバイアルで12時間明/12時間暗日長で成長させました。(B)葉鞘のサンプリング部分の模式図。葉鞘は10日齢のイネ苗から切り取った。葉鞘の位置は根の上と最初の葉の下でした。(C)リーフディスクのサンプリング位置の模式図。リーフディスクは、任意の成長段階で健康なイネの主分げつの2番目の葉の中央3分の1(上から数えて)から切り取ることができます。略語:ROS =活性酸素種;MS = 村重とスクーグ。 この図の拡大版を表示するには、ここをクリックしてください。
{kind=link}

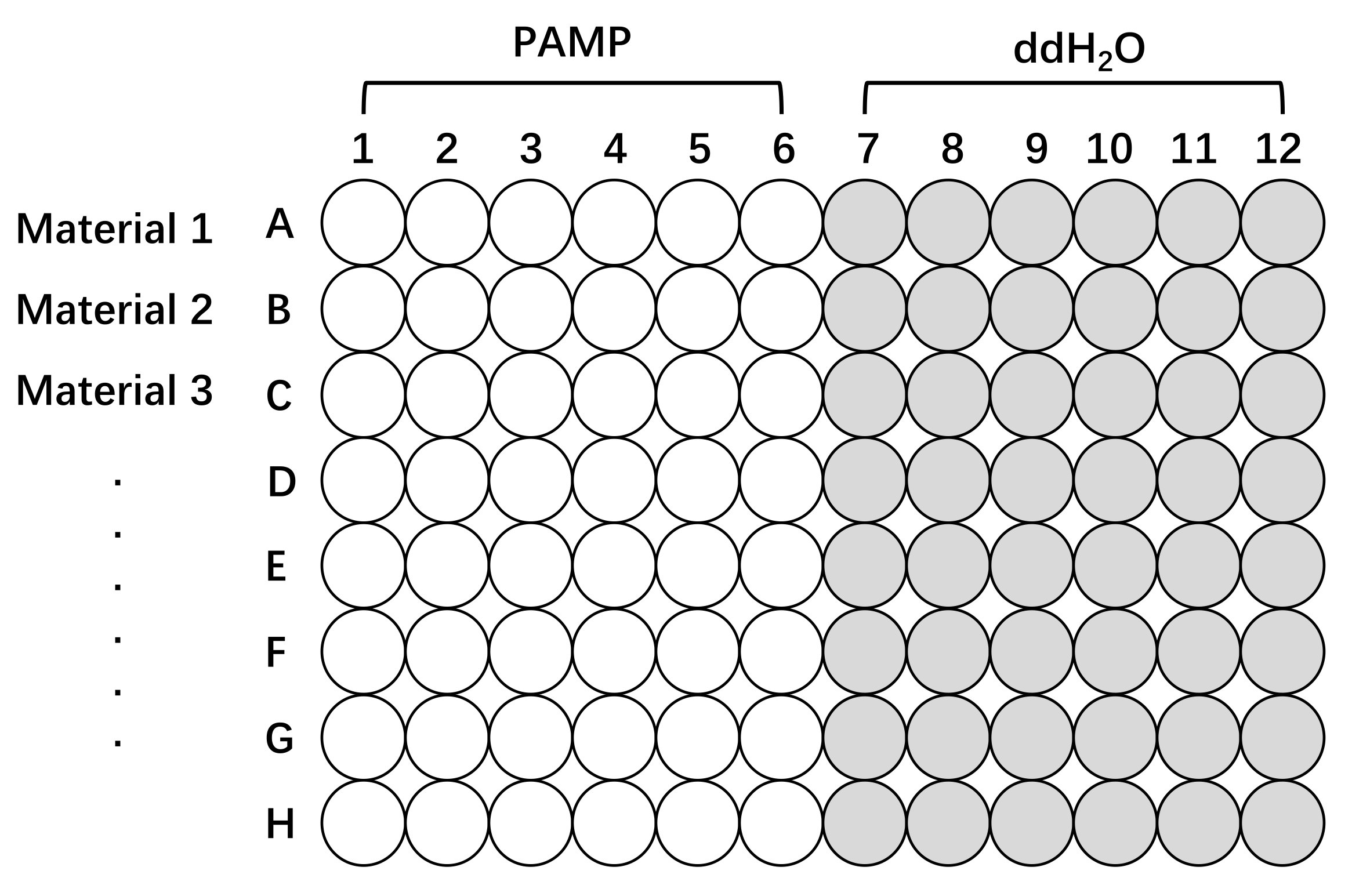
図2: Oryza sativaの異なるラインでROS産生を測定するためのプレートセットアップの概略図。 96ウェルプレートを用いたイネ組織の前処理および試験。ライン1、ライン2、およびライン3(1つのプレート上に最大8ライン)は、関心のある任意の材料、異なる栽培品種、変異体、またはトランスジェニックラインであり得る。組織を、ROS応答を測定するために、PAMP(PAMP、白色)またはPAMP(ddH2O、灰色)を含む誘発溶液で刺激した。テストするサンプルが多いほど、読み取り値の時間間隔が長くなることに注意してください。略語:ROS =活性酸素種;PAMP = 病原体関連分子パターン;ddH2O =二重蒸留水。 この図の拡大版を表示するには、ここをクリックしてください。
{kind=link}
3.誘発溶液の準備
- L-012粉末をddH2Oを含む20 mM(6.23 mg / mL)水溶液に溶解してストック溶液を作ります。その後、ストック溶液を50 mM Tris HClバッファー(pH 7.5)で希釈し、最終濃度500 μM L-012の作業溶液を作製します。使用前にストック溶液を凍結し、作業溶液に希釈してください。
- PAMP、L-012、西洋ワサビペルオキシダーゼ(HRP;ddH2O溶液10 mg/mL)を含む誘発溶液を調製します。10 mLの誘発溶液に対して、9.4 mLの50 mMトリス塩酸塩(pH 7.5)溶液、400 μLのL-012溶液、100 μLのHRP、および100 μLのflg22(PAMP;10 mMのddH2O溶液)を混合します。ネガティブコントロールの場合は、PAMPの代わりに100μLのddH2Oを追加します。
注:米の組織への冷ストレスを避けるために、調製した誘発溶液を室温に保管してください。キチン(最終濃度で20 ng / mL)など、必要に応じて他のPAMPも治療に使用できます。L-012は感光性ですので、L-012溶液を含むすべてのチューブをアルミホイルで覆います。
4.ソフトウェアを起動し、参照されているマイクロプレートリーダーでプロトコルを設定します(材料の表を参照)
注意: マイクロプレートリーダーソフトウェアのパラメータの設定には時間がかかります。誘発ソリューションを追加する前に、マシンとプロトコルを準備することをお勧めします(ワンクリックで続行)。
- ソフトウェアを起動します。[ 実験 ] ボタンをクリックして、新しいプロトコルを作成するか、既存のプロトコルを使用します。
- ポップアップの [プロシージャ ] をクリックして、プレートを設定します。監視するプレートからウェルを選択します。
- [キネティックの開始]をクリックして、合計実行時間と読み取り間隔を設定します。実験の要件に応じて、実行時間を 35 分以上に設定します。できるだけ頻繁に読み取り値を取得するには、[最小間隔]を選択します。積分時間については、信号強度に応じて1秒以上を選択します。
注:読み取り間隔は、サンプル数と信号積分時間によって異なります。 - [ 検証] |OK をクリックして設定を確認します。
- ポップアップで[新しいプレートを検出]をクリックし、ソフトウェアが[荷重 プレート] ダイアログボックスを表示するのを待ちます。テストするプレートをキャリアに置きます。
- ここで停止して、誘発システムが確立されるのを待ちます(次のセクション)。誘発システムの準備ができたらすぐに、[ 実行 ]をクリックして読み取りを開始します。
5. 誘発システムの確立とリアルタイムROS生成の測定
- 前処理された組織を含むウェルからddH2Oを慎重に取り除き、組織の損傷や乾燥を避けます。
- マルチチャンネルピペットを使用して、組織を含むウェルに200 μLの誘発溶液を追加します。
- 軽く振って混ぜます。[ 実行 ] をクリックして検出を開始します。
注:PAMP処理では、植物組織は非常に迅速に反応し、ROSを生成します。したがって、複数の治療がある場合に、操作時間を短縮するためにPAMPなしの陰性対照を最初に治療することが示唆される。治療間の誘発遅延を減らすために、できるだけ早く操作してください。誘発溶液の添加から検出開始までの時間が短いほど、重要な実験データのキャプチャが向上します。
結果
ここでは、flg22処理で生成されるROSを決定するために、イネ材料を例に挙げます。誘発後のROSの生成は一過性である。イネでは、ROS産生の増加は1-2分で最初に検出され、10-12分でピークに達し、~30-35分でベースラインに戻りました(図3)。誘発溶液中にPAMPが存在せず、明らかなROS誘導を示さなかった対照試験と比較して、特定のROSバーストは、flg22またはキチンなどの他の...
ディスカッション
本研究の目的は、イネ組織におけるPAMPに応答した初期ROS産生を非常に効率的に定量する方法を確立することである。この方法は、処理されたイネ組織から産生されたアポプラストROSをリアルタイムで測定するための標準化された手順を提供します。この方法は、操作が簡単で、低コストで、組成が明確で、市販のキットとは無関係です。この方法を使用すると、研究者は、植物が生物的また...
開示事項
著者は開示する利益相反を持っていません。
謝辞
本研究は、上海 自然科学基金会(研究課題番号:21ZR1429300/BS1500016)、上海交通大学(Agri-Xプログラム、研究課題番号:AF1500088/002)、上海農業種子共同イノベーションセンター(研究課題番号:ZXWH2150201/001)から江波ファン氏への助成金、上海交通大学医工連携プロジェクト(課題番号:21X010301734)のカン・リー氏の助成を受けて行われました。
資料
| Name | Company | Catalog Number | Comments |
| 96-well microtiter plate | WHB | WHB-96-01 | |
| Ethanol absolute | Innochem | A43543 | |
| flg22 | Sangon Biotech | p20973 | PAMP |
| Gen5 | BioTek | software | |
| L-012 | FUJIFILM | 120-04891 | 8-amino-5-chloro-7-phenyl-2,3-dihydropyrido [3,4-d] pyridazine-1,4-dione, CAS #:143556-24-5 |
| Microplate reader | BioTek | Synergy 2 | |
| MS Medium | Solarbio | M8521 | |
| NaCLO | Aladdin | S101636 | |
| Peroxidase from horseradish (HRP) | Sigma | P8375 | |
| Phytagel | Sigma | P8169 | |
| Sampler | Miltex | 15110-40 | |
| Sucrose | Sangon Biotech | A502792 | |
| Tris | Sangon Biotech | A610195 |
参考文献
- Gechev, T. S., Van Breusegem, F., Stone, J. M., Denev, I., Laloi, C. Reactive oxygen species as signals that modulate plant stress responses and programmed cell death. Bioessays. 28 (11), 1091-1101 (2006).
- Mittler, R. ROS are good. Trends in Plant Science. 22 (1), 11-19 (2017).
- Gilroy, S., et al. ROS, calcium, and electric signals: key mediators of rapid systemic signaling in plants. Plant Physiology. 171 (3), 1606-1615 (2016).
- Mittler, R., Vanderauwera, S., Gollery, M., Van Breusegem, F. Reactive oxygen gene network of plants. Trends in Plant Science. 9 (10), 490-498 (2004).
- Marino, D., Dunand, C., Puppo, A., Pauly, N. A burst of plant NADPH oxidases. Trends in Plant Science. 17 (1), 9-15 (2012).
- Mittler, R., Zandalinas, S. I., Fichman, Y., Van Breusegem, F. Reactive oxygen species signalling in plant stress responses. Nature Reviews Molecular Cell Biology. 23 (10), 663-679 (2022).
- Suzuki, N., Koussevitzky, S., Mittler, R., Miller, G. ROS and redox signalling in the response of plants to abiotic stress. Plant, Cell & Environment. 35 (2), 259-270 (2012).
- Suzuki, N., et al. Respiratory burst oxidases: the engines of ROS signaling. Current Opinion in Plant Biology. 14 (6), 691-699 (2011).
- Kadota, Y., Shirasu, K., Zipfel, C. Regulation of the NADPH oxidase RBOHD during plant immunity. Plant and Cell Physiology. 56 (8), 1472-1480 (2015).
- Segonzac, C., Zipfel, C. Activation of plant pattern-recognition receptors by bacteria. Current Opinion in Microbiology. 14 (1), 54-61 (2011).
- Roda, A., et al. Progress in chemical luminescence-based biosensors: A critical review. Biosensors and Bioelectronics. 76, 164-179 (2016).
- Hong, D., Joung, H. -. A., Lee, D. Y., Kim, S., Kim, M. -. G. Attomolar detection of cytokines using a chemiluminescence immunoassay based on an antibody-arrayed CMOS image sensor. Sensors and Actuators B: Chemical. 221, 1248-1255 (2015).
- Nishinaka, Y., et al. et al. new sensitive chemiluminescence probe, L-012, for measuring the production of superoxide anion by cells. Biochemical and Biophysical Research Communications. 193 (2), 554-559 (1993).
- Grundy, J., Stoker, C., Carre, I. A. Circadian regulation of abiotic stress tolerance in plants. Frontiers in Plant Science. 6, 648 (2015).
転載および許可
このJoVE論文のテキスト又は図を再利用するための許可を申請します
許可を申請さらに記事を探す
This article has been published
Video Coming Soon
Copyright © 2023 MyJoVE Corporation. All rights reserved