
É necessária uma assinatura da JoVE para visualizar este conteúdo. Faça login ou comece sua avaliação gratuita.
Method Article
Detecção em Tempo Real da Produção de Espécies Reativas de Oxigênio na Resposta Imune em Arroz com Ensaio de Quimioluminescência
Neste Artigo
Resumo
Aqui, descrevemos um método para a detecção em tempo real da produção de espécies reativas de oxigênio (EROs) apoplásicas em tecidos de arroz em resposta imune desencadeada por padrão molecular associado a patógenos. Este método é simples, padronizado e gera resultados altamente reprodutíveis em condições controladas.
Resumo
Espécies reativas de oxigênio (ROS) desempenham papéis vitais em uma variedade de processos biológicos, incluindo a detecção de estresses abióticos e bióticos. Após a infecção do patógeno ou desafio com produtos químicos associados a patógenos (padrões moleculares associados a patógenos [PAMPs]), uma série de respostas imunes, incluindo uma explosão de ROS, são rapidamente induzidas em plantas, o que é chamado de imunidade desencadeada por PAMP (PTI). A explosão de ROS é uma resposta característica do PTI, que é catalisada por um grupo de NADPH oxidases localizadas na membrana plasmática - as proteínas da família RBOH. A grande maioria das ERO compreende peróxido de hidrogênio (H2 O2), que pode ser fácil e constantemente detectado por um método de quimioluminescência à base de luminol. A quimioluminescência é uma reação produtora de fótons na qual o luminol, ou seu derivado (como L-012), sofre uma reação redox com ROS sob a ação de um catalisador. Este trabalho descreve um método otimizado de quimioluminescência baseado em L-012 para detectar a produção de ROS de apoplasto em tempo real após a elicitação por PAMP em tecidos de arroz. O método é fácil, estável, padronizado e altamente reprodutível sob condições firmemente controladas.
Introdução
As espécies reativas de oxigênio (EROs) compreendem uma série de derivados quimicamente ativos do oxigênio, incluindo radicais ânion superóxido (O2-) e seus derivados, radicais hidroxila (OH-), peróxido de hidrogênio e produtos de oxigênio singlete ou reações de oxidação-redução, que são constantemente produzidos em plastídios e cloroplastos, mitocôndrias, peroxissomas e outros locais subcelulares 1 . As ERO desempenham papéis importantes em muitos processos biológicos e são essenciais para todas as plantas 2,3,4. O amplo espectro de funções das ROS varia desde a regulação do crescimento e desenvolvimento até a percepção de estresses abióticos e bióticos 5,6,7,8.
No sistema imune vegetal, os receptores localizados na membrana plasmática das células vegetais - os chamados receptores de reconhecimento padrão (PRRs) - percebem padrões moleculares associados a patógenos derivados de patógenos (PAMPs). Esse reconhecimento desencadeia uma série de respostas imunes rápidas, incluindo influxo de cálcio, explosão de ROS e cascata MAPK; assim, essa camada de imunidade é denominada imunidade desencadeada por PAMP (PTI). O ROS burst é uma resposta marcante do ITP, cuja determinação é amplamente aplicada em estudos relacionados a PTI 9,10. A produção de EROs desencadeada por PAMPs é atribuída às proteínas da família NADPH oxidase residentes na membrana plasmática, ou proteínas da família RBOH (respiratory burst oxidase homolog), que transferem elétrons de NADPH citosólico ou NADH para oxigênio extracelular para produzir superóxido (O 2-) que é espontaneamente convertido em peróxido de hidrogênio (H 2 O 2) pela superóxido dismutase8 . A explosão de ROS desencadeada por PAMP é bastante rápida, aparecendo apenas alguns minutos após o tratamento com PAMP e atingindo um pico de ~10-12 min. A grande maioria das moléculas de ROS compreende peróxido de hidrogênio (H2 O2), que pode ser fácil e constantemente detectado com um ensaio de quimioluminescência.
Na quimioluminescência, o reagente quimioluminescente reage com oxigênio ativo, sob a ação de um catalisador, para produzir os intermediários de estado excitado. Em seguida, os elétrons no produto retornam ao estado fundamental através de transição não radiativa e emitem fótons. Os quimioluminescentes comuns incluem luminol e L-012, com luminol dominando a aplicação11,12,13. No entanto, mais pesquisadores estão escolhendo L-012 para detectar a produção de ROS, uma vez que L-012 tem uma eficiência de emissão de luz muito maior sob condições de pH neutro ou quase neutro em comparação com o luminol.
Este trabalho descreve um método otimizado de quimioluminescência, baseado em L-012, para a detecção em tempo real da produção de ERO após a eliciação de PAMPs em tecidos-discos foliares e bainha de arroz (Oryza sativa). O método aqui apresentado é simples, estável e padronizado, e é altamente adaptável para atender a diferentes necessidades experimentais. Os dados obtidos com este método são altamente reprodutíveis sob condições firmemente controladas.
Protocolo
NOTA: O protocolo é aplicável a diferentes tecidos vegetais. Bainha de arroz e discos foliares foram utilizados neste protocolo para detecção de ROS após elicitação de PAMP. Uma vez que as diferenças surgem principalmente devido ao método de amostragem, apenas os procedimentos comuns são descritos a seguir, sendo mencionadas etapas específicas sempre que necessário.
1. Cultura vegetal
- Esterilizar as sementes de arroz descascadas com etanol 70% por 1 min e, em seguida, com hipoclorito de sódio (NaClO) a 40% por 1 h. Em seguida, enxágue as sementes 5x com água estéril para remover o cloro residual.
- Planear as sementes assepticamente em meio 1/2 MS (2,37 g/L de meio Murashige e Skoog (MS), 30 g/L de sacarose, 2,1 g/L de fitanel, pH 5,7, autoclavado).
- No método da bainha de arroz, plaquear diretamente as sementes no recipiente de vidro estéril com meio MS.
- No método do disco foliar, plaquear as sementes em placas MS por 5-7 dias e transplantá-las para matriz de crescimento ou solo (Figura 1A).
- Cultivar as mudas em sala de crescimento com fotoperíodo de 12 h claro/12 h escuro.
2. Preparo e pré-tratamento dos tecidos
- Bainha de arroz
- Cortar a bainha de mudas de arroz com 10 dias de idade em segmentos de 3 mm com lâmina de barbear afiada ou lâmina cirúrgica para pré-tratamento 1 dia antes do ensaio de ROS (Figura 1B).
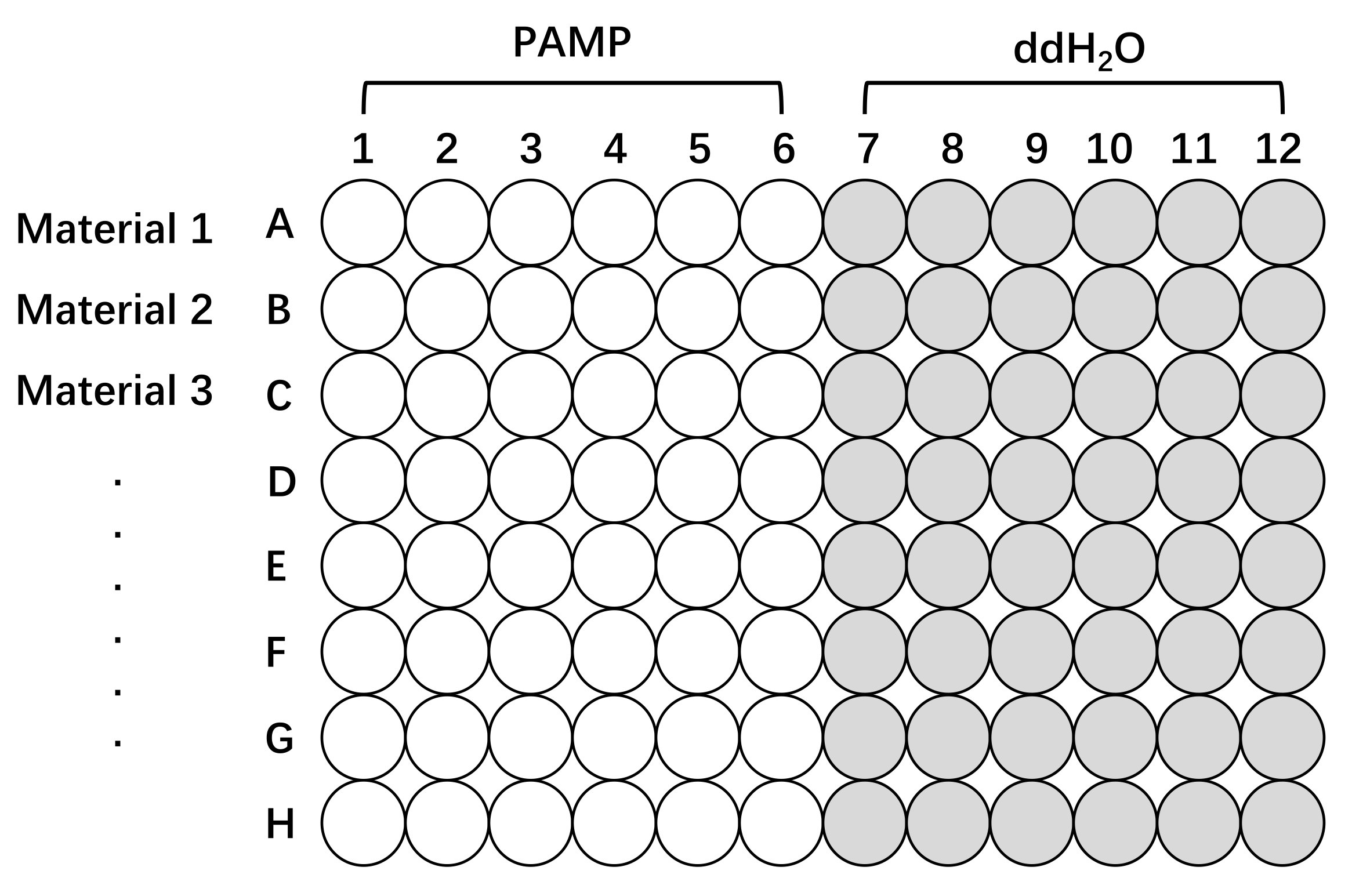
- Colocar cinco segmentos de bainha em um poço individual de uma placa de microtitulação de 96 poços contendo 100 μL deddH 2 O por 10-12 h, no escuro a 25 °C, o que permite que o extravasamento de íons relacionados à lesão da ferida e as respostas de defesa diminuam (Figura 2).
NOTA: Tomar o cuidado de manter os cortes verticais para garantir uma área de superfície de corte consistente exposta à solução de elicitação é um passo importante para obter resultados altamente reprodutíveis. Mova os segmentos suavemente. Não faça cortes ou feridas extras nos segmentos, o que pode ser uma fonte de variação de dados. Como princípio, cada teste deve conter pelo menos cinco repetições, uma vez que a variação do valor de ROS é grande. Quanto mais replicações definidas, mais confiáveis são os dados.
- Disco foliar
- Cortar os discos foliares (4 mm de diâmetro) de plantas de arroz com 4-6 semanas de idade usando um punch de biópsia com um êmbolo. Sempre corte os discos foliares do terço médio da segunda folha (numerados a partir do topo) do perfilhos principal para reduzir a variação dos dados (Figura 1C).
- Colocar um disco foliar em um poço individual de uma placa de microtitulação de 96 poços contendo 100 μL deddH 2 O por 10-12 h para pré-tratamento, o que permite que as respostas relacionadas à ferida diminuam, pois estas podem interferir na indução de ROS por PAMPs (Figura 2).
NOTA: Opere os discos de folha suavemente. Não faça cortes ou feridas extras nos discos no experimento, o que poderia resultar em variação de dados. A indução de ERO ocorre principalmente a partir das células da borda de corte, uma vez que as superfícies dos tecidos do arroz (folhas ou bainhas), são cobertas por camadas hidrofóbicas. Apenas as células das arestas de corte estão em contato com a solução de elicitação (consulte a seção de discussão). - Manter todos os discos foliares flutuando, com a superfície abaxial voltada para cima, nos poços de uma placa de microtitulação para pré-tratamento com água para evitar variação associada ao lado da folha.

Figura 1: Condições de crescimento e estádios das plântulas de arroz para amostragem da bainha e partes da bainha e folhas de arroz utilizadas no ensaio. (A) Mudas de arroz cultivadas em meio 1/2 MS em condições estéreis por 10 dias podem ser amostradas para o ensaio de ERO. Sementes de arroz esterilizadas foram cultivadas em meio MS 1/2 e cultivadas em fotoperíodo claro de 12 h/12 h escuro em frasco de vidro transparente, com 8,5 cm de diâmetro e 15 cm de altura. (B) Diagrama esquemático das partes de amostragem das bainhas foliares. Bainhas foliares foram cortadas de mudas de arroz com 10 dias de idade. As posições das bainhas foliares estavam acima das raízes e abaixo da primeira folha. (C) Diagrama esquemático da posição de amostragem dos discos foliares. Os discos foliares podem ser cortados a partir do terço médio da segunda folha (contagem a partir do topo) do perfilhos principal de plantas saudáveis de arroz em qualquer fase de crescimento. Abreviações: ROS = espécies reativas de oxigênio; MS = Murashige e Skoog. Clique aqui para ver uma versão maior desta figura.
{kind=link}

Figura 2: Diagrama esquemático do setup da placa para medição da produção de ROS com diferentes linhas de Oryza sativa. Pré-tratamento e teste de tecidos de arroz usando uma placa de 96 poços. A Linha 1, Linha 2 e Linha 3 (até oito linhas em uma placa) podem ser qualquer material de interesse, diferentes cultivares, mutantes ou linhas transgênicas. Os tecidos foram estimulados com soluções de elicitação com PAMP (PAMP, branco) ou sem PAMP (ddH2O, cinza) para medir a resposta das ROS. Ressalta-se que quanto maior o número de amostras a serem testadas, maior o intervalo de tempo entre as leituras. Abreviações: ROS = espécies reativas de oxigênio; PAMP = padrão molecular associado a patógenos; ddH2O = água bidestilada. Clique aqui para ver uma versão maior desta figura.
{kind=link}
3. Preparando a solução de elicitação
- Dissolver L-012 pó em solução aquosa de 20 mM (6,23 mg/mL) com ddH2O para fazer a solução-mãe. Em seguida, diluir a solução-mãe com tampão Tris HCl 50 mM (pH 7,5) para fazer a solução de trabalho na concentração final de 500 μM L-012. Manter a solução-mãe congelada e diluir para a solução de trabalho antes de utilizar.
- Preparar a solução de elicitação contendo PAMP, L-012 e peroxidase de raiz forte (HRP; 10 mg/mL em ddH2O). Para uma solução de elicitação de 10 mL, misturar 9,4 mL de solução de Tris HCl (pH 7,5) 50 mM, 400 μL da solução L-012, 100 μL de HRP e 100 μL de flg22 (PAMP; 10 mM em ddH2O). Para o controle negativo, adicionar 100 μL de ddH2O em vez de PAMP.
NOTA: Manter as soluções de elicitação preparadas à temperatura ambiente para evitar o stress frio dos tecidos do arroz. Outros PAMPs também podem ser usados para tratamento conforme necessário, como quitina (20 ng/mL na concentração final). Como o L-012 é sensível à luz, cubra todos os tubos contendo a solução L-012 com folha de alumínio.
4. Iniciar o software e configurar o protocolo com o leitor de microplacas referenciado (ver Tabela de Materiais)
NOTA: Leva algum tempo para configurar os parâmetros do software leitor de microplacas. Recomenda-se preparar a máquina e o protocolo (um clique para prosseguir) antes de adicionar a solução de elicitação.
- Inicie o software. Clique no botão Experimentos para criar um novo protocolo ou usar um protocolo existente.
- Clique em Procedimento no pop-up para configurar a placa. Selecione os poços da placa a ser monitorada.
- Clique em Iniciar cinética para configurar o tempo total de execução e o intervalo de leitura. Defina o tempo de execução para 35 min ou mais, dependendo dos requisitos experimentais. Para obter leituras com a maior frequência possível, selecione Intervalo mínimo. Para tempo de integração, escolha 1 s ou mais, dependendo da intensidade do sinal.
NOTA: O intervalo de leitura depende do número de amostras e da duração da integração do sinal. - Clique em Validar | OK para confirmar as configurações.
- Clique em Detectar a nova placa no pop-up e aguarde até que o software solicite a caixa de diálogo da placa de carga. Coloque a placa a ser testada no suporte.
- Pare aqui para aguardar o estabelecimento do sistema de elicitação (na próxima seção). Assim que o sistema de elicitação estiver pronto, clique em Executar para iniciar a leitura.
5. Estabelecer o sistema de elicitação e medir a produção de ROS em tempo real
- Remover cuidadosamente o ddH2O dos orifícios que contêm os tecidos pré-tratados, evitando qualquer dano tecidual ou dessecação.
- Use uma pipeta multicanal para adicionar 200 μL da solução de elicitação aos poços contendo os tecidos.
- Agite suavemente para misturar. Clique em Executar para iniciar a detecção.
NOTA: Com o tratamento com PAMP, os tecidos vegetais respondem e produzem ROS muito rapidamente. Portanto, sugere-se que o controle negativo sem PAMP seja tratado primeiramente para reduzir o tempo de operação, quando há múltiplos tratamentos. Operar o mais rápido possível para reduzir o atraso de elicitação entre os tratamentos. Quanto menor o tempo entre a adição da solução de elicitação e o início da detecção, melhor será a captura de dados experimentais importantes.
Resultados
Aqui, tomamos o material de arroz como exemplo para determinar as ROS produzidas com o tratamento flg22. A geração de ROS após a elicitação é transitória. No arroz, o aumento na produção de ROS foi detectado pela primeira vez em 1-2 min, atingiu o pico em 10-12 min e retornou à linha de base em ~30-35 min (Figura 3). Em comparação com o teste controle, no qual PAMP estava ausente na solução de elicitação, resultando em indução de ROS sem óbvia, um burst específico de ROS ...
Discussão
O objetivo deste estudo foi estabelecer um método altamente eficiente para quantificar a produção precoce de ERO em resposta ao PAMP em tecidos de arroz. Este método fornece um procedimento padronizado para a determinação em tempo real de ROS de apoplasto produzido a partir de tecidos de arroz tratados. Este método é simples na operação, baixo custo, claro na composição e independente de kits comerciais. Usando este método, os pesquisadores podem estudar a produção em tempo real de ROS apoplasto quando as ...
Divulgações
Os autores não têm conflitos de interesse a declarar.
Agradecimentos
Este trabalho foi apoiado por subsídios da Shanghai Natural Science Foundation (Número de Bolsa: 21ZR1429300/BS1500016), Shanghai Jiao Tong University (Programa Agri-X, Número de Bolsa:AF1500088/002), Shanghai Collaborative Innovation Center of Agri-Seeds (Número de Bolsa: ZXWH2150201/001) para Jiangbo Fan, e pelo Projeto de Colaboração de Engenharia Médica da Shanghai Jiao Tong Univesity (Número de Bolsa: 21X010301734) para Can Li.
Materiais
| Name | Company | Catalog Number | Comments |
| 96-well microtiter plate | WHB | WHB-96-01 | |
| Ethanol absolute | Innochem | A43543 | |
| flg22 | Sangon Biotech | p20973 | PAMP |
| Gen5 | BioTek | software | |
| L-012 | FUJIFILM | 120-04891 | 8-amino-5-chloro-7-phenyl-2,3-dihydropyrido [3,4-d] pyridazine-1,4-dione, CAS #:143556-24-5 |
| Microplate reader | BioTek | Synergy 2 | |
| MS Medium | Solarbio | M8521 | |
| NaCLO | Aladdin | S101636 | |
| Peroxidase from horseradish (HRP) | Sigma | P8375 | |
| Phytagel | Sigma | P8169 | |
| Sampler | Miltex | 15110-40 | |
| Sucrose | Sangon Biotech | A502792 | |
| Tris | Sangon Biotech | A610195 |
Referências
- Gechev, T. S., Van Breusegem, F., Stone, J. M., Denev, I., Laloi, C. Reactive oxygen species as signals that modulate plant stress responses and programmed cell death. Bioessays. 28 (11), 1091-1101 (2006).
- Mittler, R. ROS are good. Trends in Plant Science. 22 (1), 11-19 (2017).
- Gilroy, S., et al. ROS, calcium, and electric signals: key mediators of rapid systemic signaling in plants. Plant Physiology. 171 (3), 1606-1615 (2016).
- Mittler, R., Vanderauwera, S., Gollery, M., Van Breusegem, F. Reactive oxygen gene network of plants. Trends in Plant Science. 9 (10), 490-498 (2004).
- Marino, D., Dunand, C., Puppo, A., Pauly, N. A burst of plant NADPH oxidases. Trends in Plant Science. 17 (1), 9-15 (2012).
- Mittler, R., Zandalinas, S. I., Fichman, Y., Van Breusegem, F. Reactive oxygen species signalling in plant stress responses. Nature Reviews Molecular Cell Biology. 23 (10), 663-679 (2022).
- Suzuki, N., Koussevitzky, S., Mittler, R., Miller, G. ROS and redox signalling in the response of plants to abiotic stress. Plant, Cell & Environment. 35 (2), 259-270 (2012).
- Suzuki, N., et al. Respiratory burst oxidases: the engines of ROS signaling. Current Opinion in Plant Biology. 14 (6), 691-699 (2011).
- Kadota, Y., Shirasu, K., Zipfel, C. Regulation of the NADPH oxidase RBOHD during plant immunity. Plant and Cell Physiology. 56 (8), 1472-1480 (2015).
- Segonzac, C., Zipfel, C. Activation of plant pattern-recognition receptors by bacteria. Current Opinion in Microbiology. 14 (1), 54-61 (2011).
- Roda, A., et al. Progress in chemical luminescence-based biosensors: A critical review. Biosensors and Bioelectronics. 76, 164-179 (2016).
- Hong, D., Joung, H. -. A., Lee, D. Y., Kim, S., Kim, M. -. G. Attomolar detection of cytokines using a chemiluminescence immunoassay based on an antibody-arrayed CMOS image sensor. Sensors and Actuators B: Chemical. 221, 1248-1255 (2015).
- Nishinaka, Y., et al. et al. new sensitive chemiluminescence probe, L-012, for measuring the production of superoxide anion by cells. Biochemical and Biophysical Research Communications. 193 (2), 554-559 (1993).
- Grundy, J., Stoker, C., Carre, I. A. Circadian regulation of abiotic stress tolerance in plants. Frontiers in Plant Science. 6, 648 (2015).
Reimpressões e Permissões
Solicitar permissão para reutilizar o texto ou figuras deste artigo JoVE
Solicitar PermissãoThis article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. Todos os direitos reservados