
Method Article
Exogene Verabreichung von Alpha-Synuclein Microsomen verbundenen Aggregate zur primären Neuronen als eine mächtige Zellmodell der Fibrillen Bildung
In diesem Artikel
Zusammenfassung
Das Ziel dieses Protokolls ist, ein Zell-basierte System, das die Bildung von Alpha-Synuclein repliziert in Vivoaggregiert zu bieten. Intrazelluläre Alpha-Synuclein Einschlüsse sind in primären Neuronen durch die Internalisierung ausgesät und Vermehrung von exogenen verabreicht native microsomen-assoziierten Alpha-Synuclein-Aggregaten von erkrankten Alpha-Synuclein transgenen Mäusen isoliert.
Zusammenfassung
Die Unfähigkeit der Bildung von unlöslichen Alpha-Synuclein (αS) Einschlüsse in Zellkulturen zu replizieren ist seit Jahren eine große Einschränkung in der Studie der αS Aggregation bei Parkinson-Krankheit (PD). Vor kurzem hat die Entwicklung von neuen Tiermodellen durch die exogene Impfung von Gehirn Auszüge aus erkrankten αS Transgene Mäuse oder PD-Patienten neue Hoffnungen auf die Möglichkeit der Schaffung eine angemessenere zellmodelle αS Aggregation gegeben. Leider geht es um Zellen in Kulturen, Verwaltung von rohen Gehirn Extrakte nicht so erfolgreich wie in Mäusen nachgewiesen und die Quelle der Wahl von exogenen Aggregaten ist immer noch in-vitro- vorgeformt αS Fibrillen.
Wir entwickelten eine Methode, um die Bildung von intrazellulären αS Einschlüsse im primären Neuronen durch die exogene Gabe von native microsomen-assoziierten αS Aggregate, eine hochgiftige αS Spezies isoliert von erkrankten Bereiche von transgenen Mäusen zu induzieren. Diese Fraktion αS Aggregate, die mit den microsomen Vesikel ist, effizient verinnerlicht und induziert die Bildung von positiven für aggregierte und phosphorylierten αS intrazelluläre Einschlüsse. Im Vergleich zur in-Vitro-vorgeformte Fibrillen aus rekombinanter αS, unsere Methode ist schneller und Garantien, die die pathogenen Aussaat mit authentischen αS Aggregate erfolgt aus erkrankten Tiermodellen der PD, imitiert mehr eng die Art der Einschlüsse erhalten in Vivo. Infolgedessen ist die Verfügbarkeit von Geweben, die reich an αS Einschlüsse obligatorisch.
Wir glauben, dass diese Methode ein vielseitiges zellbasierte Modell um den mikroskopischen Aspekten der αS Aggregation und der damit verbundenen zellulären Pathophysiologie in Vivo zu untersuchen und ein für die Erstellung präziser und anspruchsvolle Zelle Ausgangspunkt Paradigma der PD.
Einleitung
Ansammlung von Alpha-Synuclein (αS) proteinhaltige Einschlüsse ist ein bedeutender und wichtiger Punkt der Parkinson Erkrankung (PD) und Alpha-Synulceinopathies1. Leider während Tiermodelle um eine ausreichende zelluläre und biochemische Umgebung um die Schritte der Aggregation notwendig für die Bildung von Protein Fibrillen2induzieren können, Bildung von komplexen Lewy Body (LB) replizieren-Aggregate wie in Zellkulturen ist schwierig und herausfordernd.
Hier beschreiben wir eine Methode, um die Bildung von αS Einschlüsse, ähnlich wie Protein-Aggregate erhalten Sie in Tiermodellen und PD-Patienten, in kultivierten Zellen mit primären Nervenzellen im Gehirn isoliert Maus zu induzieren. Unser Protokoll basiert auf die exogene Verabreichung von microsomen-assoziierten αS Aggregate von αS symptomatisch transgene (Tg) Mäuse Maus hippocampal oder kortikale primäre Neuronen isoliert. Diese Methode nutzt die Ausbreitung und Vermehrung Fähigkeit αS giftige Arten, die, sobald das Kulturmedium hinzugefügt sind, werden verinnerlicht, und induzieren die Bildung von Reifen αS-positiven Aggregate3,4, 5,6,7,8.
Ursprünglich basierten Standardmethoden zur Bildung von αS Fibrillen in Zellkulturen zu erhalten auf die Überexpression von der entsprechenden αS cDNA durch regelmäßige Transfektion Protokolle oder virale-vermittelte Infektion9. Während im ersten Fall Beschaffung LB-ähnliche αS Aggregate zufällig, waren, zeigten geringen Effizienz, und hing von der Zelltyp, führte das zweite Protokoll zur Bildung von unlöslichen Fibrillen, einschließlich hohes Molekulargewicht (HMW) Arten in 24-48 h vor Infektionen 10. in diesen Methoden, die Bildung von Aggregaten war wohl aufgrund eines übermäßigen und unsymmetrisch in αS Protein-Menge, die unlösliche anstatt einer pathologischen Umwandlung von αS Konformation, die Aggregation bestimmt wird. Stattdessen die Technik, die wir hier präsentieren ändert nicht die αS Expression aber weit verbreitete Proteinaggregation durch die Internalisierung der exogenen Fibrillen induziert. Darüber hinaus die Bildung von αS Aggregate durch die Gabe von exogenen Fibrillen ist ein langwieriger Prozess, der Tage oder Wochen zu erschöpfend ermöglicht es uns, die frühen und mittlere Stadien der αS Einschlüsse Bildung in einem Zeitraffer Mode studieren erfordert und um es mit der zellulären, biochemischen Veränderungen korrelieren. Somit ist unsere Methode eine wertvolle Anwendung zum Erstellen von zellmodellen αS Aggregation, die hilfreich sind, αS Fibrille Bildung mikroskopisch in Bezug auf zelluläre Pathophysiologie zu studieren.
Darüber hinaus obwohl Verwaltung der rohen Gehirn aus erkrankten αS (Tg) Transgenic Mäuse11,Auszüge kann12 oder menschlichen PD Gehirne6,13 induzieren αS Ablagerung in Tg oder Wildtyp (WT) Tiere, Anwendung der gleichen Prozedur, die Zellkulturen erweist nicht sich als erfolgreich, möglicherweise durch die geringe Menge an Aggregate in den Beispielen verwendet und das Fehlen eines Standardverfahrens, native αS toxischer Spezies14zu isolieren. Aus diesem Grund in Vitro vorgeformt Fibrillen (PFFs) von αS wurden die Aggregate-Quelle der Wahl bis jetzt für die Induktion von αS Einschlüsse in den Zellen und Tiermodelle3,4,6,7 ,15,16. Mit unserem Protokoll zeigen wir jedoch, dass microsomen-assoziierten αS aggregierten Arten von αS Tg Mäusen isoliert effizient Anhäufung von intrazellulären LB-ähnliche αS Einschlüsse im primären Neuronen veranlassen können.
In unserem Labor sind microsomen-assoziierten αS aggregierten Arten isoliert aus dem Rückenmark (SpC) Gewebe der erkrankten Tg Mäuse mit dem Ausdruck menschlichen A53T αS Gens unter der Kontrolle der Maus Prion-Protein (PrP) Veranstalter [Prp menschlichen A53T αS Tg Mäuse, Linie G2-3-17]. Diese Mäuse zeigen eine altersabhängige Neurodegenerative Phänotyp, der robuste motor Dysfunktion und Bildung von Einschlüssen in das zentrale Nervensystem enthält aus phosphoryliert, Ubiquitinated und unlöslichen αS, ab 9 Monate alt. Sobald motorische Störungen angezeigt wird, entwickelt sich der Phänotyp schnell zu Lähmungen, ausgehend vom hinteren Gliedmaßen, die bis zum Tod in ca. 2-3 Wochen führt. Ansammlung von αS Aggregate parallelen Manifestation der Krankheit. Mäuse, die zu Beginn der motorischen Dysfunktion geopfert zeigen eine robuste αS Aggregation im SpC, Hirnstamm und Kleinhirn. Es gibt keine Notwendigkeit zu warten, bis die Lähmung setzt auf die Maus verzichten. Präsymptomatische Mäuse werden auf 9 Monate alten Tiere getroffen, die motorische Störungen nicht angezeigt werden.
Protokoll
Die Verwendung von WT und Tg Tieren gebilligt und in voller Höhe von den nationalen und internationalen Gesetze zum Tierschutz Labor und experimentieren (EWG des Rates Richtlinie 86/609, 12. Dezember 1987 und Richtlinie 2010/63/EU, 22. September 2010) erfüllt. Alle Protokolle, die in diesem Dokument beschriebenen Anweisungen Tierbetreuung unserer Institution.
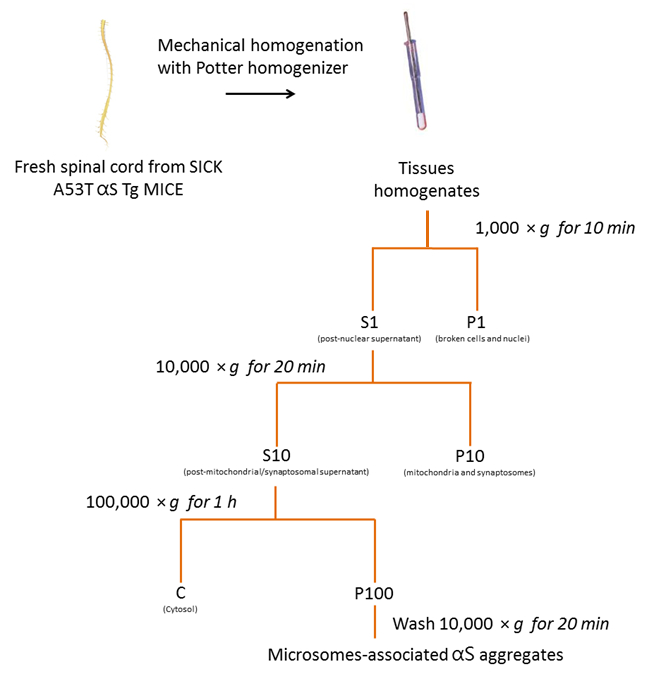
1. Isolierung Microsomen-assoziierten αS Aggregate von erkrankten A53T αS Tg Mäuse
- Vorbereiten des Homogenisierung Puffers, bestehend aus Saccharose 250 mM, 20 mM HEPES, 10 mM KCl, 1,5 mM MgCl2, 2 mM EDTA und 1 X Phosphatase/Protease-Hemmer ist. Halten Sie den Puffer auf Eis.
- Homogenisieren der frisches oder gefrorenen Gewebe in einem 01:10 (w/V) Volumen des eiskalten Homogenisierung Puffer mit einem Teflon zerstoßen Homogenisator, mit 10-15 Schläge.
- Übertragen Sie anfängliche Homogenat (1-2 mL) zu einem Microcentrifuge Schlauch und Zentrifuge bei 1000 × g für 10 min bei 4 ° C mit einer gekühlten Zentrifuge um Kerne und ungebrochene Zellen in der resultierenden Pellet (P1) zu entfernen. P1 zu verwerfen.
- Übertragen Sie den überstand (S1) auf einem sauberen Microcentrifuge Schlauch und Zentrifugieren Sie S1 bei 10.000 × g für 20 min bei 4 ° C mit einer gekühlten Zentrifuge Erlangung der zweiten überstand (S10) und das Pellet (P10).
Hinweis: P10 ist ein Roh Membran-Pellet, der Mitochondrien und Synaptosomes enthält. P10 zu verwerfen. - Den überstand (S10) auf einer Polycarbonat-Flasche übertragen (> 1 mL) und Zentrifugieren S10 bei 100.000 × g für 1 h bei 4 ° C mit einer Ultrazentrifuge und einem festen Winkel-Rotor (90 Ti).
Hinweis: Der Überstand ist der reine Zytosol Anteil während der Pellet, P100, microsomen-assoziierten αS Zuschlagstoffe enthält. - Die P100-Pellet mit 500 µL des Puffers Homogenisierung aufzuwirbeln. Übertragen Sie P100 auf eine saubere Microcentrifuge Schlauch und Zentrifuge bei 10.000 × g für 20 min bei 4 ° C in einer gekühlten Zentrifuge.
- Verwerfen Sie den überstand und Aufschwemmen Sie P100 mit 100 µL der Homogenisierung Puffer.
Hinweis: Diese Fraktion ist die microsomen-assoziierten αS Aggregate. - Beschallen von Proben für 2 s auf Eis [Set Ausgangsleistung 1 Watt (RMS)]. Shop-Proben bei-80 ° C.
- Am nächsten Tag, bestimmen die Protein-Menge mit BCA Analyse.
(2) Western-Blot
Hinweis: Biochemische Charakterisierung der microsomen-assoziierten αS Aggregate wird durch Western-Blot ausgewertet.
- Werfen Sie eine Farbverlauf 4-20 % Tris-Glycin Polyacrylamid-Gel auf einer vertikalen Elektrophorese-Apparatur.
- Laden Sie 1 µg microsomen-assoziierten αS Fraktionen, in der denaturierenden Probenpuffer aufgelöst.
- Laden Sie in einem anderen Brunnen 5 µL Protein standard Marker.
- Führen Sie das Gel bei 100 V in Tris/Glycin/SDB Puffer ausgeführt, bis Protein Marker das Ende des Gels erreicht.
- Transfer von Proteinen auf eine Nitrocellulose-Membran mit einem grundlegenden Karbonat-Puffer (10 mM NaCO3, 3 mM Nahco33, 20 % Methanol) O/N bei 4 ° C, bei 200 mA, konstant.
- Blockieren der Membran mit PBS-nichtionische Tenside 0,05 % (PBS-T) mit 5 % fettfreie Trockenmilch auf einem Orbitalschüttler für 30 min bei RT
- Kurz, spülen Sie die Membran mit PBS-T.
- Inkubieren Sie die Membran mit Syn-1 (1:5 000) oder pSer129-αS Antikörper (1:5 000) in 2,5 % fettfreie Trockenmilch im PBS-T, O/N bei 4 ° C auf einem Orbitalschüttler.
- Waschen Sie die Membran für 10 min bei RT mit PBS-T auf einem Orbitalschüttler.
- Inkubieren Sie die Membran mit Anti-Maus oder Anti-Kaninchen HRP-konjugierten Sekundärantikörper (1:3 000) in 2,5 % fettfreie Trockenmilch im PBS-T für 1 h bei RT
- Waschen Sie die Membran für 10 min bei RT mit PBS-T auf einem Orbitalschüttler. Wiederholen Sie 3 X.
- Erhalten Sie das Signal durch die regelmäßigen Chemilumineszenz-Kits.
(3) neuronalen Primärkulturen
Hinweis: Neuronale Primärkulturen wurden aus dem WT Neugeborenen (P0) Maus Hippocampus oder Kortex (Linie C57BL/6) vorbereitet. Das gesamte Verfahren, abzüglich der Zentrifugation Schritte wird unter einer Zelle Kultur Haube, unter sterilen Bedingungen durchgeführt.
- Behandlung von Deckgläsern (18 mm Ø) mit 65 % Salpetersäure-Lösung für mindestens 12 h bei RT
- Entfernen Sie die Salpetersäure. Deckgläsern zweimal mit PBS 10 x und mehrere Male mit destilliertem Wasser abspülen.
- Legen Sie Deckgläsern in 24 Wohlen Teller und bestreichen Sie sie mit Poly-D-Lysin (0,1 mg/mL in sterilem destilliertem Wasser oder PBS 1 X) für 1 h bei 37 ° C.
- Poly-D-Lysin zu entfernen und die Deckgläsern dreimal mit destilliertem Wasser zu waschen. Speichern sie bei 4 ° C bis benötigt.
- Die Maus-Welpen durch Enthauptung einschläfern und die Köpfe aus dem Körper zu trennen. Legen Sie die Köpfe auf Gerichte und sezieren Sie die Haut sanft zu.
- Mit feinen Schere Öffnen des Schädels durch einen Schnitt an den Basen der Gehirne zu machen. Trennen Sie die beiden Hälften der Schädel und entfernen Sie vorsichtig.
- Durch die Verwendung einer Pinzette, aus den Gehirnen von den Basen Kneifen und der Kortex und Hippocampus zu isolieren. In zwei separate Küche mit Hank es ausgewogen Salz Lösung (HBSS) 1 % Penicillin/Streptomycin-haltigem Medium überführen.
- Wiederholen Sie die Schritte 3.6 und 3.7 für jedes Tier. Beachten Sie, dass der gesamte Prozess nicht mehr als 30-45 min dauern sollte.
- Zerkleinere die gesammelten Gewebe und übertragen Sie sie in zwei 50 mL konische Röhrchen (eine für Hippocampus) und eine für Kortex mit 10 mL HBSS Medium mit Trypsin 0,1 %. Im Wasserbad für 7 min bei 37 ° c inkubieren
Hinweis: Keine Unruhe ist erforderlich. - Fügen Sie 1 mL DMEM mit 10 % fetalen bovine Serum (FBS) und 10 µg/mL DNase, das Homogenat, Trypsin-Aktivität zu blockieren.
- Das resultierende dissoziierte Gewebe 200 × g für 5 min bei RT auf einer Zentrifuge zentrifugieren und den überstand zu entfernen.
Hinweis: Das Pellet repräsentiert die Zellen aus dem Gewebe seziert. - Aufschwemmen der Zellen in der Beschichtung Medium, mit 2 % B27, 2 mM Glutamin, 6 mg/mL Glukose 10 % FBS, 12,5 µM Glutamat und 10 µg/mL Gentamicin.
- Platte dissoziiert Neuronen auf Poly-D-Lysin Deckgläsern in 24-well-Platten entsprechend dem Verhältnis: 1 Rinde/12 Brunnen und 1 Hippocampus/6 Brunnen, was in etwa 150.000/200.000 Zellen pro Bohrloch. Halten Sie die Zellen im Inkubator bei 37 ° C.
- Am Folgetag (Tag in Vitro, DIV 1), ersetzen Sie die Beschichtung Medium mit 2 % B27, 10 µg/mL Gentamicin und 2 mM Glutamin-haltigem Medium.
- Entfernen Sie bei DIV 2 1/3 des Mediums zu und fügen Sie 1/3 des frischen Mediums mit 2,5 µM Cytosin Arabinoside für 48 h, die Glia Kontamination zu reduzieren.
- Pflegen Sie Neuronen bei 37 ° C und ersetzen Sie die Hälfte des Mediums alle drei Tage.
(4) Neuronen Behandlung
Hinweis: Die Behandlung wurde bei DIV 7 durchgeführt. Alle Schritte werden unter einer Zelle Kultur Haube unter sterilen Bedingungen betrieben. Ein Beispiel für kortikale neuronale Kultur Dichte bei DIV 7 ist in Abbildung 1dargestellt.
- Pool microsomen-assoziierten αS Aggregate aus dem Rückenmark von drei verschiedenen krankhaften Tg Mäusen erhalten, um eine 1 µg/µL-Lösung, mit der ursprünglichen Homogenisierung-Puffer für die Verdünnung haben.
- Entfernen Sie 1/3 des Mediums zu und ersetzen Sie ihn sanft mit dem frischen Medium, 2 % B27, 1 x Gentamicin und 2 mM Glutamin enthalten.
- Die mittlere Zelle 1 µg der gepoolten microsomen-assoziierten αS Aggregate hinzufügen. Rückkehr von Neuronen im Inkubator bei 37 ° C.
- Alle 3 Tage für 1 Woche, fügen Sie 1/3 der frisches Medium. Ersetzen Sie das Medium nicht. Fügen Sie es einfach.
- Entfernen Sie nach 1 Woche Behandlung 1/3 des Mediums zu und ersetzen Sie ihn sanft mit dem frischen Medium, 2 % B27, 10 µg/mL Gentamicin und 2 mM Glutamin enthalten. Wiederholen Sie alle 3 Tage.
- Neuronen nach 2 Wochen der Behandlung (DIV 21) zu beheben.
5. Immunfluoreszenz
- Beheben von Neuronen mit 2 % Paraformaldehyd in PBS 1 X und 5 % Saccharose-Lösung für 15 min bei RT ohne schütteln, unter einer Chemikalie fume Haube.
- Entfernen Sie die Befestigungsschraube Lösung. Kurz, waschen Sie mit PBS 1 x 3-Mal.
Hinweis: Alle Schritte waschen sanft weil primäre Neuronen nicht fest an Poly-Lysin Deckgläsern halten. - Permeabilize Neuronen mit 0,3 % nichtionische Tenside mit PBS-Puffer 1 X für 5 min bei RT
- Kurz, waschen Sie mit PBS 1 x 3-Mal.
- Inkubieren Sie Neuronen mit 3 % FBS mit PBS-Puffer 1 X für 30 min bei RT, unspezifischen Bindungsstellen auf einem Orbitalschüttler blockieren.
- Inkubieren Sie Neuronen mit entsprechenden Primärantikörper aufgelöst in 3 % FBS mit PBS-Puffer 1 X, O/N bei 4 ° C auf einem Orbitalschüttler.
Hinweis: αS (1: 200), pser129-αS (1:1 000) und Tau (01:10, 000) syn303 (1:1, 000), Maus Antikörper verwendet wurden. - Entfernen Sie die Antikörperlösung und waschen, kurz, mit PBS 1 x 3-Mal.
- Inkubieren Sie Neuronen mit entsprechenden fluoreszierende Sekundärantikörper aufgelöst in 3 % FBS mit PBS-Puffer für 1 h bei RT im Dunkeln auf einem Orbitalschüttler.
- Entfernen Sie die Antikörperlösung und waschen, kurz, mit PBS 1 x 3-Mal.
- Färben von Neuronen mit DAPI-Lösung (0,1 µg/mL mit PBS-Puffer 1 X) für 15 min bei RT in der Dunkelheit auf einem Orbitalschüttler.
- Entfernen Sie die Antikörperlösung und waschen, kurz, mit PBS 1 x 3-Mal.
- Montieren Sie auf einer Folie mit antifade Eindeckmittel Deckgläsern.
Ergebnisse
Nach dem Protokoll oben beschrieben und in Abbildung 2zusammengefasst, wir gereinigt microsomen-assoziierten αS Aggregate von drei erkrankten A53T αS Tg Mäusen (Abbildung 3). Microsomen sind Roh Membran-Pellet-Fraktionen, die das endoplasmatische Retikulum, Golgi und kleine synaptischen Vesikel enthalten. Der Reinheitsgrad des mikrosomale Pellets im Vergleich zu anderen Fraktionen war zuvor mit bestimmten Organell Marker18ausgewertet.
Einmal isoliert, wird die biochemische Charakterisierung der αS Aggregate durch denaturierenden SDS-Page, gefolgt von Inkubation mit Syn-1 oder phosphorylierten αS an Serin 129 (pSer129-αS) Antikörper beurteilt. Im Vergleich zu präsymptomatische (PreS) und Alter abgestimmt nicht-Tg (nTg) Mäuse, microsomen isoliert von Kranke Mäuse Co Niederschlag mit αS Aggregate. Microsomen-assoziierten αS Arten zeigen die typischen Merkmale der αS Aggregate, wie z.B. Ansammlung von HMW Waschmittel-resistente Spezies, Phosphorylierung an Serin 12919 und C- und N-terminale abgeschnitten Fragmente (für eine vollständige Charakterisierung der microsomen-assoziierten αS Aggregaten siehe Referenz18,20,21). Dies sind grundlegende Anforderungen, da Monomere αS nicht effizient verinnerlicht bekommen und nicht dazu, αS Ablagerung7,15,22 veranlassen. Es ist wichtig, keine Reinigungsmittel (Ionische oder nichtionischen) nicht um P100 Pellets aufzuwirbeln, da es schädlich für Zellen sein kann. Außerdem werden zur Vermeidung von Probe Variabilität microsomen-assoziierten αS Aggregate aus drei verschiedenen erkrankten Mäusen für neuronale Behandlung werden gebündelt.
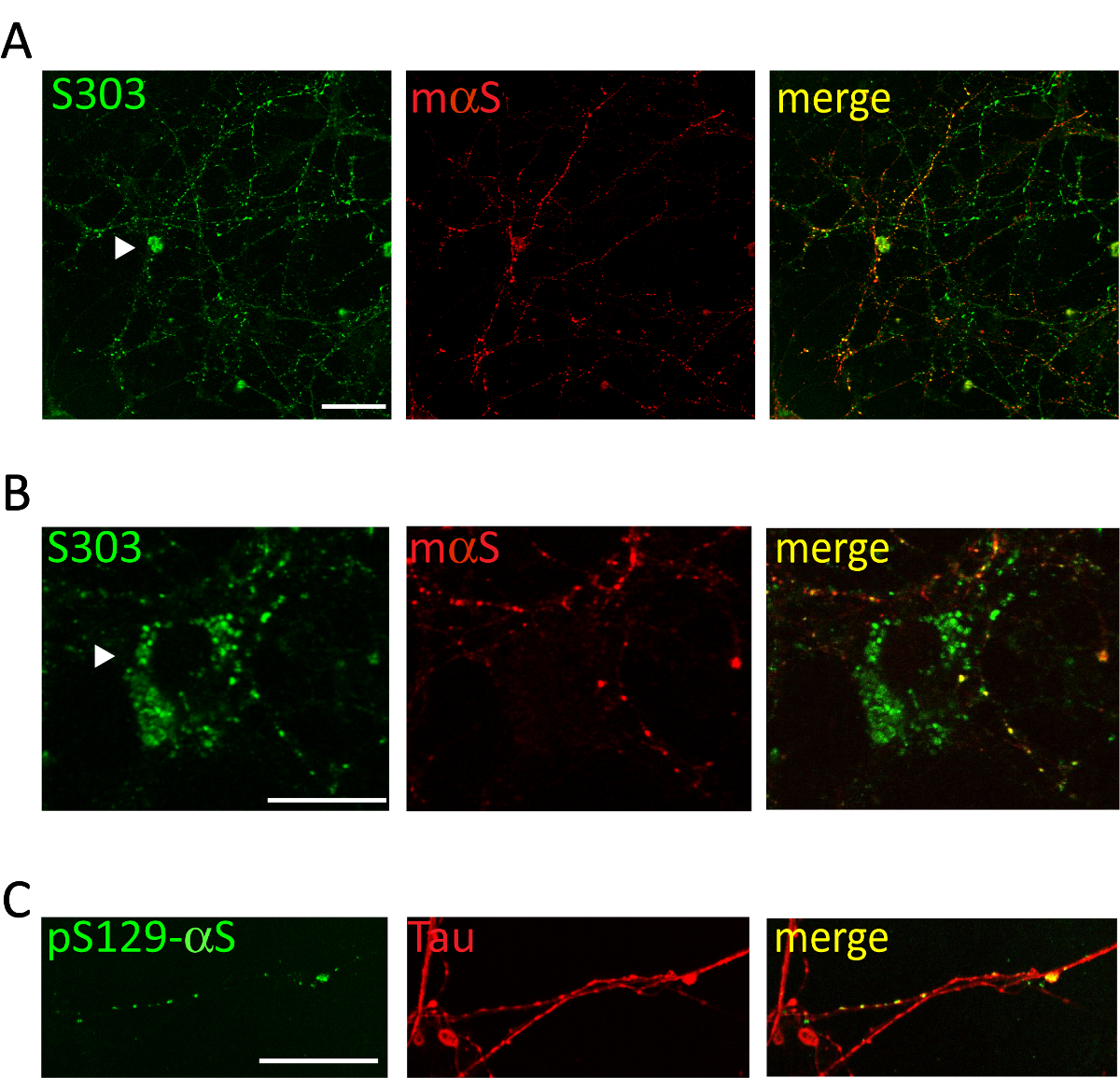
Verwaltung von 1 µg der gepoolten microsomen-assoziierten αS Aggregate von erkrankten A53T Mäusen auf dem Kulturmedium von kortikalen oder hippocampal Neuronen induziert eine zeitabhängige Formation αS Einschlüsse, αS Aggregate-spezifische Antikörper wie Syn303 (Abbildung 4, 5) oder pser129-αS (Abbildung 5). Nach zwei Tagen der Behandlung diese Aggregate erscheinen als kleine, verstreute Puncta, die häufiger zu späteren Zeitpunkten werden (2d). Nach zwei Wochen αS Einschlüsse ähnlich lange und Reifen Perlen-ähnliche Strukturen, stark verbreitete sich in den neuronalen Kulturen, eine Neurit Muster folgen und teilweise gemeinsam mit präsynaptischen Lokalisierung und Neuriten Marker (Abbildung 5). Gelegentlich sehen neu gebildeten αS Einschlüsse Co lokalisieren und Färben der Zelle Soma oder decken den gesamten Prozess, ähnlich nekrotische Neuriten.
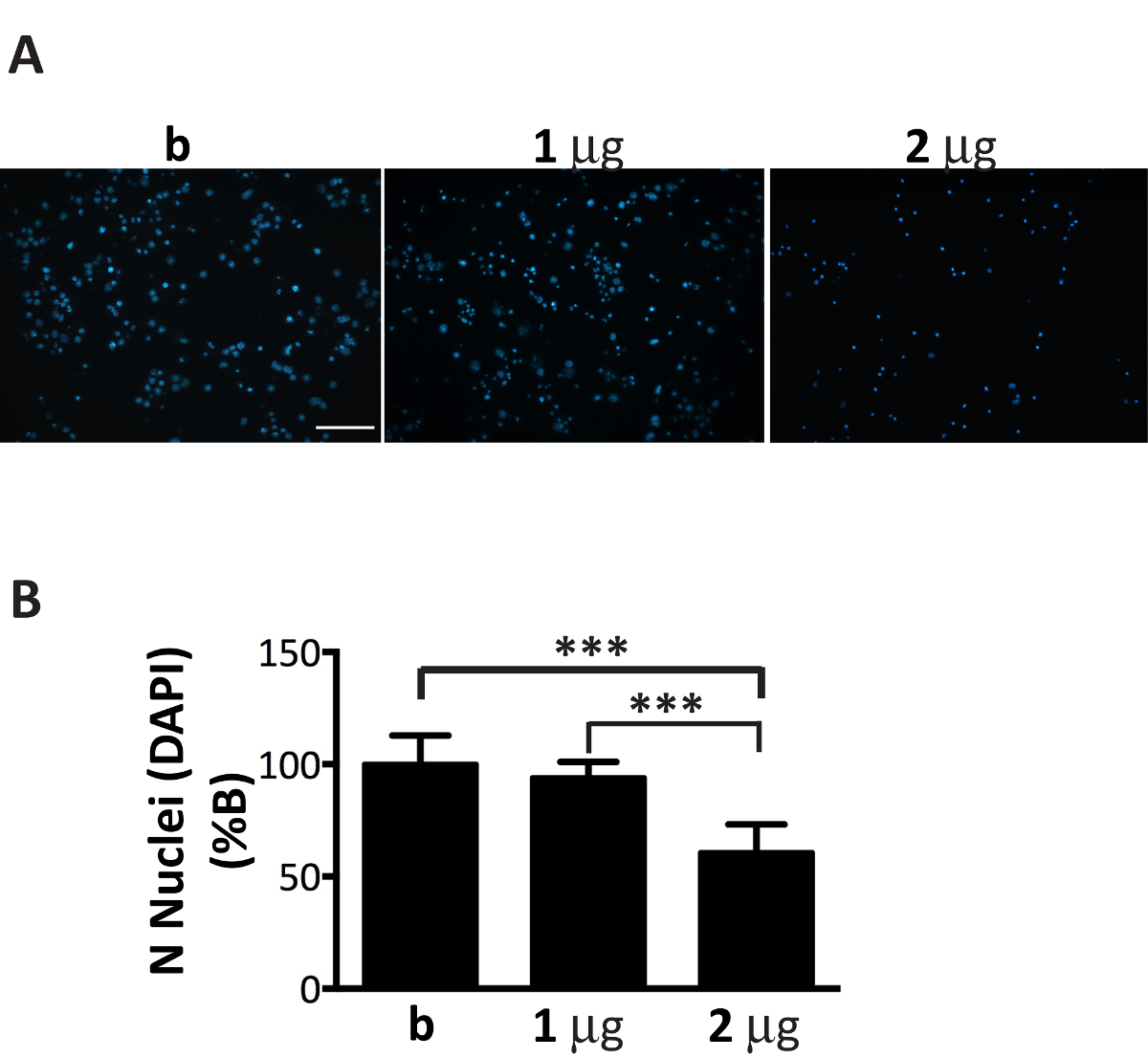
Obwohl microsomen-assoziierten αS Aggregate Brüche effizient in neuronalen Kulturen ausbreiten können, muss deren Höhe fein abgestimmt werden, um die Anzahl der Neuronen beschichtet (Abbildung 1). In der Tat mehr als das empfohlene Verhältnis µg microsomen verbundenen Aggregate: Anzahl der Neuronen, vorzeitigen Zelltod innerhalb von ein paar Tagen (Abbildung 6), zu induzieren, während eine unzureichende Menge an microsomen-assoziierten αS Aggregate zu führen wird ein knapp und reduzierte Anzahl der Einschlüsse nach zwei Wochen der Behandlung, ähnlich dem, was war zu früheren Zeitpunkten (Abb. 4A) erhalten.

Abbildung 1 . Kortikale neuronale Kulturen. Repräsentatives Bild zeigt Dichte bei DIV 7 kortikale neuronale Kulturen. Bilder wurden mit einem inversen Licht Mikroskop, 10 X-Objektiv aufgenommen. Maßstabsleiste = 100 µm. Bitte klicken Sie hier für eine größere Version dieser Figur.
{kind=link}

Abbildung 2 . Isolierung von microsomen-assoziierten αS Aggregate aus Maus SpC. Flussdiagramm des Protokolls Reinigung microsomen-assoziierten αS Aggregate von SpC von erkrankten Mäusen. Bitte klicken Sie hier für eine größere Version dieser Figur.
{kind=link}

Abbildung 3 . Isolierung von microsomen Fraktionen aus erkrankten, präsymptomatische A53T αS Tg und nTg Mäuse. Western-Blot Analyse zeigt gereinigt microsomen-assoziierten αS Aggregate isoliert von drei erkrankten (krank), präsymptomatische (PreS) und im Alter abgestimmt nTg Mäuse. Kranke Mäuse sind A53T αS Tg Mäuse, die zeigen, die motorische und neurologische Dysfunktion, einschließlich die Anhäufung von αS Einschlüsse, während präsymptomatische Tiere gesunde A53T αS Tgs von 9 Monaten des Alters, die nicht zeigen noch αS Pathologie bezogene Phänotyp. nTg-Mäuse sind, wurfgeschwistern von erkrankten Mäusen die tragen nicht des αS Transgens und entwickeln daher keine αS Pathologie induziert. 1 µg der einzelnen gereinigten Fraktionen waren laufen auf einer denaturierenden SDS-Page, übertragen auf eine Nitrocellulose-Membran und getupft mit Syn-1 oder pSer129-αS Antikörper. Nur microsomen Fraktionen isoliert von Kranke Mäuse enthaltenen HMW Reinigungsmittel resistent αS Aggregate, die bei 129 Serin phosphoryliert und zeigte C- und N-terminale abschneiden. Diese Zahl wurde angepasst von Colla Et al. 18. Bitte klicken Sie hier für eine größere Version dieser Figur.
{kind=link}

Abbildung 4 . Zeitabhängige Induktion von αS Absetzung nach der Verabreichung von microsomen-assoziierten αS Aggregate. (A) Immunfluoreszenz von primären hippocampal Neuronen behandelt mit 1 µg microsomen-assoziierten αS aggregiert Bruchteile von drei verschiedenen erkrankten Mäusen gebündelt. Neuronen wurden an 2 Tagen behoben (2d), 1 Woche (1w) oder 2 Wochen (2w) Behandlungund Immunostained mit syn303 (S303, 1: 1000), ein Antikörper, der spezifisch für oxidiert und aggregiert αS. Zellen wurden mit DAPI counterstained. Konfokale Bilder wurden mit einem Laser-scanning-confocal Mikroskop, 63 X Objektiv. Maßstabsleiste = 50 µm. (B) Quantitative Analyse des gesamten Fluoreszenzpräparate nach Hintergrundabzug, erfolgte mit dem Partikel-Graf-Plugin Bild J-Software. Werte wurden für die Anzahl der Kerne pro Feld (DAPI Graf) normalisiert und ausgedrückt als Prozentsatz des S303 Fluoreszenz-Signals am 2D. Werte sind als Mittelwert ± SD gegeben (n = 5). ** p < 0,001 *** p < 0.00001, One-Way ANOVA, gefolgt von Fisher LSD Post-hoc-Test. Bitte klicken Sie hier für eine größere Version dieser Figur.
{kind=link}

Abbildung 5 . Intrazelluläre αS Einschlüsse colocalize mit kortikalen Neuriten Netzwerk. Repräsentative konfokale Bilder von kortikalen Neuronen mit microsomen-assoziierten αS Aggregate von erkrankten Tg Mäuse erhalten behandelt. Nach 2 Wochen der Behandlung Neuronen feste und Doppel waren befleckt mit Aggregaten-spezifische Antikörper [S303 (A, B) oder pser129-αS, 1:1,000 (C)] und Neurit Marker [Maus αS, 1: 200 (A, B) oder Tau, 1: 10.000 (C)]. Co Kennzeichnung der fluoreszierenden Signale zeigte teilweise Co Lokalisierung der neugeformten αS Wulst-artige Strukturen mit dem Neuriten Netzwerk. Sammeln gelegentlich αS Einschlüsse innerhalb der neuronalen Soma (A, B, Pfeilspitzen). Gestapelten Bilder wurden mit einem Laser-scanning-confocal Mikroskop, 63 X Objektiv erworben. Maßstabsleiste = 50 µm. Diese Zahl wurde von Colla Et Al. modifiziert 20. Bitte klicken Sie hier für eine größere Version dieser Figur.
{kind=link}

Abbildung 6 . Suboptimale Menge microsomen-assoziierten αS Aggregate ist giftig für Neuronen. DAPI Färbung von Neuronen behandelt mit zunehmender Konzentration der microsomen-assoziierten αS Aggregate zum Zelltod führt. (A) kortikale Neuronen wurden behandelt mit 1, 2 µg microsomen-assoziierten αS Aggregate aus erkrankten Mäusen oder extrahiert nur Puffer, die nicht enthält (B) aggregiert und mit DAPI gefärbt. Fluoreszierende Bilder wurden mit einem Epi-Fluoreszenz-Mikroskop mit einem 20 X Objektiv erworben. Maßstabsleiste = 100 µm.()B) DAPI-positiven Zellen gezählt wurden, mit Hilfe der Software Bild J. Das Diagramm zeigt einen Rückgang der Zahl der Kerne mit zunehmender Konzentration von microsomen-assoziierten αS Aggregate hinzugefügt, um die neuronalen Medien. Werte werden als % der b ausgedrückt und sind als Mittelwert ± SD gegeben (n = 5), *** p < 0,0001, One-Way ANOVA, gefolgt von Fisher LSD Post-hoc-Test. Bitte klicken Sie hier für eine größere Version dieser Figur.
{kind=link}
Diskussion
Wir beschrieben, eine Methode zur Bildung von αS Einschlüsse im Gehirn abgeleitet neuronalen Primärkulturen von WT-Mäusen, durch Zusatz von gereinigtem microsomen-assoziierten αS Aggregate von αS Tg Tiermodellen isoliert zu erhalten.
Wichtige Schritte dieses Protokolls sind die folgenden: das Verhältnis von µg microsomen-assoziierten αS Aggregate/Neuronen und die Quelle der αS Aggregate. Wie in der Ergebnissitzung gezeigt, es ist wichtig, das Verhältnis von µg microsomen-assoziierten αS Aggregate/Anzahl von Neuronen zu optimieren, da unter suboptimalen Bedingungen arbeiten zu den vorzeitigen Zelltod oder zu knapp intrazellulären Aggregation führen kann (siehe die Abschnitt "repräsentative Ergebnisse"). Aus diesem Grund ist es sehr wichtig, die Dichte der neuronalen Kultur an DIV 7 (siehe Abbildung 1) vor Beginn der Behandlung zu bewerten. Darüber hinaus haben αS Aggregate von erkrankten αS Tg Mäusen, d.h.gereinigt werden. aus Tiermodellen, die sich ansammeln LB-ähnliche Einschlüsse von phosphorylierten und Waschmittel unlösliche αS HMW Fibrillen geprägt.
Eventuelle Änderungen dieses Protokolls betrachten das Gewebe, gefroren oder frisch, aus welcher microsomen isoliert werden können und der Ausgangsbetrag. Während wir SpC Tg Mäuse aufgrund des hohen Gehaltes an αS unlösliche Aggregate verwendet, eignet sich das Protokoll microsomen verbundenen Aggregate aus jedem Gewebe isoliert unter der Voraussetzung, dass das Gebiet verfügt über einen hohen Gehalt an αS Aggregaten. Gefrorene Proben können auch verwendet werden, da Einfrieren Reinigungsschritte microsomen verbundenen Aggregate oder die Aggregate per senicht beeinflusst. Zwar die empfohlenen Einwaage für Gewebe ca. 100-150 mg, dieses Protokoll eignet sich für den Erhalt der microsomen von so niedrig wie 50 mg des Rohstoffs (keine maximale Gewichtsgrenze). Bei dem Betrag von weniger als 100 mg werden das Verhältnis der entsprechenden Homogenisierung 01:20 (w/V) jedoch um mindestens 1 mL der überstehenden S10 auf der Polycarbonat-Flasche für Ultrazentrifugen Niederschlag geladen haben. In der Tat führen laden kleiner als 1 mL Volumen der Zusammenbruch der Rohr- und Probe Verlust. Erhöhung des Volumens der Homogenisierung führt zu mehr verdünnte überstand jedoch die Konzentration des Pellet-mikrosomale bleibt davon aber unberührt.
Eine Einschränkung dieses Protokolls betrifft das ineffiziente Kreuz-seeding bei der Bildung von αS Einschlüsse, die kürzlich in der Rechtssache αS PFFs menschlichen Ursprungs murinen neuronalen Kulturen im Gegensatz zur Maus αS PFFs24gemeldet wurden. Da Erhöhung der exogenen αS Fibrillen, murine Kulturen gegeben dieses Problem umgehen kann, empfehlen wir, die Menge an microsomen verbundenen Aggregate bei der Verabreichung von Fibrillen aus anderen αS Varianten oder aus verschiedenen fein abzustimmen Spezies Maus neuronalen Kulturen als das, was wir beschrieben.
Exogene αS Aggregate hinzugefügt, um die Kultur-Medien können aus verschiedenen Quellen sein. In-vitro- αS PFFs wurden zuvor als Ausgangswert Vorlage der intrazellulären αS Aggregate in Zellkulturen, primären Neuronen und Tiermodellen3,7,8,15verwendet. Im Vergleich zu unserer Methode wo microsomen-assoziierten αS Aggregate in paar Stunden isoliert werden können, ist die Bildung von PFFs, langwierig und mühsam, erfordert mehrere Schritte von Reinigungen, gefolgt von zusätzlichen Tests zu überprüfen, dass αS Aggregate Confomations25 . Darüber hinaus PFFs aus bakteriellen ausgedrückt Mensch oder Maus αS, typisch für Eukaryoten, d.h. fehlende posttranslationale Modifikationen können präsentieren verschiedene Konformationen mit selektiven seeding und pathogenen Eigenschaften nach die Keimbildung Protokolle gefolgt (d.h. Bänder Vs Fibrillen)5,8 führt zu unterschiedlichen Ergebnissen und Schlussfolgerungen. Stattdessen garantiert einheitliche Verwaltung von in Vivo gereinigt αS Aggregate die Übertragung von authentischer pathogenen Vorlagen, imitiert eng den Prozess der Bildung von αS Einschlüssen in Tiermodellen und Parkinson-Patienten.
Als zukünftige Anwendung dieser Technik glauben wir, dass dieses Protokoll erfolgreich αS pathogenen Samen aus dem Gehirn von Parkinson-Patienten oder anderen αS Tg Tiermodellen, isolieren eingesetzt werden kann, vorausgesetzt, dass die erkrankten Bereich aus dem microsomen isoliert sind, sind reich an αS Einschlüsse.
Nach unserer Kenntnis ist dies die erste Methode, die die Reinigung des einheimischen giftigen Arten von αS von in Vivo PD Modelle als Vorlage für den Erhalt der Bildung von αS Einschlüsse im primären Neuronen Aussaat verwendet werden kann.
Wir glauben, dass diese Methode äußerst vielseitig ist und eine außergewöhnliche zellbasierte Modell bieten um die verschiedenen Aspekte der αS Aggregation und ihren Einfluss auf die Zelle Pathophysiologie zu studieren. Da die Bildung von αS Einschlüsse bilden einen komplexen Prozess, der schwierig, in kultivierten Zellen repliziert wurde, sind wir zuversichtlich, dass dieses Modell tolle Einblicke in akuten pathogenen Mechanismen, schwer bietet zu erkennen, in chronischen und aufwendigere Systeme sind z. B. Tiermodelle.
Offenlegungen
Die Autoren haben nichts preisgeben.
Danksagungen
Diese Arbeit wurde vom italienischen Ministerium für Universität und Forschung (MIUR) durch das Beihilfeprogramm Karriere Wiedereingliederung (RLM-Programm für junge Forscher) unterstützt und von Scuola Normale Superiore. Wir danken Prof. Dr. Michael Lee von der University of Minnesota, USA, für die Bereitstellung der Prp menschlichen A53T αS Tg Mäuse, aus denen Aggregate isoliert sind.
Materialien
| Name | Company | Catalog Number | Comments |
| Sucrose | Sigma-Aldrich | 84097-1KG | |
| Hepes | Sigma-Aldrich | H0887-100ML | 1M pH=7-7.6 |
| EDTA | Sigma-Aldrich | 0390-100ml | pH=8 0.5M |
| MgCl2 | Sigma-Aldrich | M8266-100G | |
| NaCO3 | Sigma-Aldrich | S7795-500G | |
| NAHCO3 | Sigma-Aldrich | S5761-500G | |
| Methanol | Sigma-Aldrich | 322415-6X1L | |
| KCl | Sigma-Aldrich | P9541-500G | |
| cOmplete Mini | Roche | 11836170001 | protease inhibitor |
| PhosStop | Roche | 4906837001 | phosphatase inhibitor |
| BCA Protein Assay Kit | Euroclone | EMPO14500 | |
| Criterion TGX 4-20% Stain Free, 18 wells | Biorad | 5678094 | |
| Supported Nitrocellulose membrane | Biorad | 1620097 | 0.2 μm |
| Blotting-Grade Blocker | Biorad | 1706404 | Non-fat dry milk |
| SuperSignal West Pico Chemiluminescent Substrate | Termo Fisher Scientific | 34077 | |
| Nitric acid | Sigma-Aldrich | 1004411000 | 65% |
| Glass Coverslips | Termo Fisher Scientific | 1014355118NR1 | 18 mm x |
| Poly-D-Lysine | Sigma-Aldrich | P7280 | |
| Hank's Balanced Salt Solution | Termo Fisher Scientific | 14170-500 mL | |
| Penicillin/Streptomycin | Termo Fisher Scientific | 15140122 | 10,000 U/mL, 100 mL |
| Dulbecco’s Modified Eagle’s Medium | Termo Fisher Scientific | D5796-500 mL | |
| Trypsin-EDTA | Termo Fisher Scientific | 15400054 | 0.50% |
| B27 Supplement | Termo Fisher Scientific | 17504044 | 50X |
| Glutamax | Termo Fisher Scientific | 35050-038 | 100x |
| DNAse | Sigma-Aldrich | D5025 | |
| Fetal bovine serum | Euroclone | EC50182L | |
| Glutamate | Sigma-Aldrich | 1446600-1G | |
| Gentamicin | Termo Fisher Scientific | 15710 | 10 mg/ml |
| Neurobasal Medium | Termo Fisher Scientific | 10888-022 | |
| Cytosine arabinoside (AraC) | Sigma-Aldrich | C3350000 | |
| VECTASHIELD antifade mounting medium | Vector Laboratories | H-1000 | |
| DAPI | Termo Fisher Scientific | 62247 | |
| 90 Ti rotor | Beckman | N/A | Ultracentrifuge rotor |
| Optima L-90K Ultracentrifuge | Beckman | N/A | |
| Syn-1 antibody, clone 42 | BD Biosciences | 610786 | anti-mouse WB: 1:5000 |
| Syn303 antibody | BioLegend | 824301 | anti-mouse IF: 1:1000 |
| Tau antibody | Synaptic Systems | 314 002 | anti-rabbit IF: 1:10,000 |
| pser129-αS antibody | A gift from Fujiwara et al, reference 19 | anti-rabbit WB: 1:5000 | |
| pser129-αS antibody | Abcam | ab51253 | anti-rabbit IF: 1:1000 |
| Mouse αS (D37A6) XP | Cell Signaling | 4179 | anti-rabbit IF 1:200 |
| Alexa fluor 555-conjugated anti-rabbit antibody | Termo Fisher Scientific | A27039 | |
| Alexa fluor 488-conjugated anti-mouse antibody | Termo Fisher Scientific | A-11029 | |
| Microson XL-2000 | Misonix | Sonicator | |
| Ultra Bottles (Oakridge Bottles), PCB, 16x76mm, Assembly, Noryl Cap, Beckman-type | Science Service EU | S4484 | Ultracentrifuge tubes |
| AXIO Observer Inverted Light Microscope | Zeiss | N/A | |
| TCS SP2 laser scanning confocal microscope | Leica | N/A | |
| Inverted epi-fluorescence microscope | Nikon | N/A | |
| Triton x-100 | Sigma-Aldrich | X100-500ML | Nonionic surfactant |
Referenzen
- Goedert, M., Spillantini, M. G., Del Tredici, K., Braak, H. 100 years of Lewy pathology. Nat. Rev. Neurol. 9, 13-24 (2012).
- Visanji, N. P., et al. α-Synuclein-Based Animal Models of Parkinson's Disease: Challenges and Opportunities in a New Era. Trends Neurosci. 39, 750-762 (2016).
- Luk, K. C., et al. Pathological α-Synuclein Transmission Initiates Parkinson-like Neurodegeneration in Nontransgenic Mice. Science. 338, 949-953 (2012).
- Rey, N. L., Petit, G. H., Bousset, L., Melki, R., Brundin, P. Transfer of human α-synuclein from the olfactory bulb to interconnected brain regions in mice. Acta Neuropathol. (Berl). 126, 555-573 (2013).
- Guo, J. L., et al. Distinct α-Synuclein Strains Differentially Promote Tau Inclusions in Neurons. Cell. 154, 103-117 (2013).
- Masuda-Suzukake, M., et al. Prion-like spreading of pathological α-synuclein in brain. Brain. 136, 1128-1138 (2013).
- Volpicelli-Daley, L. A., et al. Exogenous α-Synuclein Fibrils Induce Lewy Body Pathology Leading to Synaptic Dysfunction and Neuron Death. Neuron. 72, 57-71 (2011).
- Peelaerts, W., et al. α-Synuclein strains cause distinct synucleinopathies after local and systemic administration. Nature. 522, 340-344 (2015).
- Lázaro, D. F., Pavlou, M. A. S., Outeiro, T. F. Cellular models as tools for the study of the role of alpha-synuclein in Parkinson's disease. Exp. Neurol. 298, 162-171 (2017).
- Lee, H. -. J., Patel, S., Lee, S. -. J. Intravesicular localization and exocytosis of alpha-synuclein and its aggregates. J. Neurosci. Off. J. Soc. Neurosci. 25, 6016-6024 (2005).
- Luk, K. C., et al. Intracerebral inoculation of pathological α-synuclein initiates a rapidly progressive neurodegenerative α-synucleinopathy in mice. J. Exp. Med. 209, 975-986 (2012).
- Mougenot, A. -. L., et al. Prion-like acceleration of a synucleinopathy in a transgenic mouse model. Neurobiol. Aging. 33, 2225-2228 (2012).
- Recasens, A., et al. Lewy body extracts from Parkinson disease brains trigger α-synuclein pathology and neurodegeneration in mice and monkeys: LB-Induced Pathology. Ann. Neurol. 75, 351-362 (2014).
- Woerman, A. L., et al. Propagation of prions causing synucleinopathies in cultured cells. Proc. Natl. Acad. Sci. 112, 4949-4958 (2015).
- Luk, K. C., et al. Exogenous alpha-synuclein fibrils seed the formation of Lewy body-like intracellular inclusions in cultured cells. Proc. Natl. Acad. Sci. U. S. A. 106, 20051-20056 (2009).
- Sacino, A. N., et al. Intramuscular injection of α-synuclein induces CNS α-synuclein pathology and a rapid-onset motor phenotype in transgenic mice. Proc. Natl. Acad. Sci. U. S. A. 111, 10732-10737 (2014).
- Lee, M. K., et al. Human alpha-synuclein-harboring familial Parkinson's disease-linked Ala-53 --> Thr mutation causes neurodegenerative disease with alpha-synuclein aggregation in transgenic mice. Proc. Natl. Acad. Sci. U. S. A. 99, 8968-8973 (2002).
- Colla, E., et al. Endoplasmic Reticulum Stress Is Important for the Manifestations of -Synucleinopathy In Vivo. J. Neurosci. 32, 3306-3320 (2012).
- Fujiwara, H., et al. alpha-Synuclein is phosphorylated in synucleinopathy lesions. Nat. Cell Biol. 4, 160-164 (2002).
- Colla, E., et al. Toxic properties of microsome-associated alpha-synuclein species in mouse primary neurons. Neurobiol. Dis. 111, 36-47 (2018).
- Colla, E., et al. Accumulation of Toxic -Synuclein Oligomer within Endoplasmic Reticulum Occurs in -Synucleinopathy In Vivo. J. Neurosci. 32, 3301-3305 (2012).
- Volpicelli-Daley, L. A., Luk, K. C., Lee, V. M. -. Y. Addition of exogenous α-synuclein preformed fibrils to primary neuronal cultures to seed recruitment of endogenous α-synuclein to Lewy body and Lewy neurite-like aggregates. Nat. Protoc. 9, 2135-2146 (2014).
- Li, W., et al. Aggregation promoting C-terminal truncation of alpha-synuclein is a normal cellular process and is enhanced by the familial Parkinson's disease-linked mutations. Proc. Natl. Acad. Sci. U. S. A. 102, 2162-2167 (2005).
- Luk, K. C., et al. Molecular and Biological Compatibility with Host Alpha-Synuclein Influences Fibril Pathogenicity. Cell Rep. 16, 3373-3387 (2016).
- Volpicelli-Daley, L. A., Kirik, D., Stoyka, L. E., Standaert, D. G., Harms, A. S. How can rAAV-α-synuclein and the fibril α-synuclein models advance our understanding of Parkinson's disease. J. Neurochem. 139, 131-155 (2016).
Nachdrucke und Genehmigungen
Genehmigung beantragen, um den Text oder die Abbildungen dieses JoVE-Artikels zu verwenden
Genehmigung beantragenThis article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. Alle Rechte vorbehalten