
Method Article
Préparation de l’échantillon miniaturisé pour la microscopie électronique à Transmission
* Ces auteurs ont contribué à parts égales
Dans cet article
Résumé
Méthodes pour la préparation du volume des échantillons de taille nanolitres pour microscopie électronique à transmission et un instrument est présenté. Aucune étape de papier-buvard est requise, afin d’éviter les conséquences négatives que cela peut avoir pour les protéines, considérablement réduisant la perte de l’échantillon et permettant l’analyse de lysat pour la protéomique visuel unique cellule.
Résumé
En raison des progrès technologiques récents, cryo microscopie électronique (cryo-EM) devient rapidement une méthode standard pour l’analyse structurale des complexes de protéine à la résolution atomique. Cependant, techniques d’isolation de protéine et les méthodes de préparation d’échantillon pour EM restent un goulot d’étranglement. Un nombre relativement faible (100 000 à quelques millions) des particules de protéines individuelles doivent être photographié pour l’analyse à haute résolution des protéines par la seule particule approche EM, rendant l’échantillon miniaturisé manipulation techniques et principes microfluidiques faisable.
Miniaturisés, papier-buvard-free EM grille PROCEDE de preparation de pré conditionnement d’échantillon, de EM grille d’amorçage et de post-traitement qui ne consomme que nanolitre-volumes d’échantillon est présenté. La méthode utilise un système de distribution avec une précision sub-nanolitres pour absorption liquide contrôle et amorçage grille EM, une plate-forme pour contrôler la température de la grille ainsi déterminer l’humidité relative au-dessus de la grille de l’EM et un pick-et-plongée-mécanisme d’échantillon vitrification. Pour cryo-EM, une grille d’EM est placée sur la scène à température contrôlée et l’échantillon est aspiré dans un tube capillaire. La pointe capillaire est placée à proximité de la surface de la grille, la grille est chargée avec l’échantillon et excès est ré-aspiré dans le microcapillaire. Par la suite, le film de l’échantillon est stabilisé et légèrement éclairci par évaporation de l’eau contrôlée réglementée par le décalage de la température de la plate-forme par rapport au point de rosée. À un moment donné, le mécanisme de choix-et-plongement est déclenché, transférer rapidement la grille EM amorcée en éthane liquide pour la vitrification de l’échantillon. Alternativement, les méthodes de conditionnement d’échantillon sont disponibles pour préparer le volume des échantillons de taille nanolitres pour une coloration négative (NS) EM.
Les méthodologies de considérablement réduisent la consommation de l’échantillon et éviter les approches potentiellement nuisibles aux protéines, telles que le filtre papier buvard utilisées dans les méthodes conventionnelles. En outre, la minuscule quantité d’échantillon nécessaire permet de nouvelles stratégies expérimentales, telles que rapide exemple conditionné, combinaison avec lyse monocellulaires pour « protéomique visuel », ou « sans perte » préparation de l’échantillon total pour l’analyse quantitative des échantillons complexes.
Introduction
Matériel et logiciels pour l’analyse structurale des complexes de protéine par microscopie électronique à transmission (TEM) a massivement avancé au cours de ces dernières années. Les améliorations apportées a ouvert la voie à une « révolution de la résolution »1,2 et fondamentalement changé recherche structurale. La révolution a commencé avec l’avènement de cryo-electron microscopy (cryo-EM)3,4, qui permet la préparation d’échantillons biologiques sous proche des conditions physiologiques tout en diminuant la sensibilité de la radiation et la prévention évaporation d’échantillon dans le vide du microscope électronique à transmission5. Dans les années suivantes, progrès technologique a graduellement augmenté la résolution réalisable. Parmi ces innovations ont été l’application de canons de l’émission de champ6,7et, plus récemment, amélioration des algorithmes d’analyse de données, tels que les méthodes de maximum de vraisemblance8,9. Direct électronique détecteur caméras10,11,12,13, imagerie en mode film et l’accompagnement logiciel développements14,15, 16 , 17, fourni la percée finale requise pour atteindre la résolution atomique pour échantillons biologiques par analyse de particules uniques (pour une revue, voir Cheng, Grigorieff, et al. 18). l’importance de la cryo-EM a récemment été reconnu par l’attribution du prix Nobel de chimie à trois des pionniers.
Pour un échantillon biologique par TEM de l’image, la méthode utilisée pour charger la grille EM avec l’échantillon (par la suite dénommé « préparation de la grille ») doit s’assurer que la couche résultante de l’échantillon (i) est assez mince (< 100 nm) pour éviter les bruits de vastes d’inélastiques ou multiplier dispersés électrons, (ii) résiste au vide poussé du microscope électronique, et (iii) protège les biomolécules des radiolésions. Deux méthodes principales sont utilisées pour remplir ces gratifications : colorant négatif(NS)19,20 procédures (Figure 1A) adsorbent l’échantillon à un film de carbone mince, incorporer les biomolécules dans amorphe métaux lourds, puis permettre à l’Assemblée sécher à l’air. C’est simple et rapide, et les grilles de EM chargés (par la suite dénommés « grilles d’échantillon ») sont faciles à stocker et peuvent être maintenus pendant de longues périodes de temps (en général années). En TEM, les préparations présentent un contraste élevé en raison de la NS et tolèrent des doses plus élevées d’électrons que cryo-préparations, mais la résolution est limitée à environ 20 procédures Å. Cryo-EM (Figure 1B) emploient holey carbone prend en charge. Une couche mince de la solution échantillon est enjambée à travers les trous et la grille de l’EM est plongée dans un cryogène, éthane habituellement liquéfié, pour la refroidir rapidement au-dessous de-150 ° C. Le résultat est un amorphe, vitrifiée, 50 à 100 nm d’épaisseur film de la solution dans les trous de la prise en charge. Ce film mince, amorphe résiste le vide poussé au microscope électronique et, dans le cas idéal, préserve les structures biologiques dans leur état natif. La procédure permet d’échantillons biologiques à être photographiée à haute résolution. Cependant, la grille de l’échantillon doit être conservée à température ci-dessous-150 ° C en permanence pour éviter les devitrification. Il peut être copié à l’aide de doses relativement élevées d’électrons en raison de la basse température, mais le contraste et le rapport signal-bruit est néanmoins faible. Par conséquent, moyenne techniques sont employées pour augmenter le contraste et, pourvu que l’échantillon est photographié sous différents angles, Tèlèchargez la carte en trois dimensions (3D) peut être reconstruite. Le plus couramment utilisé et une méthode très réussie pour la reconstruction 3D à partir de maintenant est l’approche d’une seule particule. Pour une revue récente, voir Cheng al.18.
Colorant négatif TEM (NS-EM) est important pour le dépistage et le contrôle de la qualité, quand le contraste élevé est nécessaire ou lorsqu’il existe seulement une quantité limitée de l’échantillon (adsorption pour le film de carbone concentre généralement l’échantillon). Une seule particule cryo-EM est la méthode de l’étalon-or si les reconstructions 3D haute résolution de la structure de la protéine sont visées pour.

Figure 1: préparation de grille de principes de TEM et la comparaison entre le classique (panneau A, B) et une approche de microfluidique (panneau C, D). A) préparation de grille classique NS-EM : environ 3 µL de l’échantillon sont reversé à la main sur une grille de EM recouverte d’un film de carbone continue (dénommé par la suite une « grille de NS-EM ») (i). Après incubation pendant env. 10 s, papier filtre est utilisé pour sécher à l’excès de liquide du côté (ii), laissant les molécules adsorbées dans un film mince de l’eau. Par la suite, la protéine est incubée dans une solution de sel de métaux lourds, par exemple, 2 % d’acétate d’uranyle, pendant 20 s (iii) et nouveau le liquide est supprimé en tamponnant sur le côté à l’aide de papier filtre (iv). Enfin, l’EM-grille on laisse pour sécher à l’air. B) préparation de grille classique cryo-EM : environ 3 µL de l’échantillon sont distribués par un coup de main sur un film de carbone trouée. Pour former un film mince échantillon, le liquide excédentaire est enlevé par le papier-buvard visage-sur d’une ou deux faces (ii). Enfin, la grille est rapidement plongée dans éthane liquide pour la vitrification (iii). C) préparation de grille de NS-EM en utilisant la configuration de cryoWriter : volume A 5 nL est aspiré de la crosse de l’échantillon à l’aide d’un microcapillaire (i). Pour exemple de conditionnement, la pointe microcapillaire est immergée dans la solution de climatisation, par exemple, 2 % d’acétate d’ammonium. Ions et petites molécules sont échangés par diffusion (ii). Notez que les dimensions de la microcapillaire veille à ce que l’ensemble du processus est piloté par diffusion. Les protéines sont les constantes de diffusion beaucoup plus faible que les ions de sels et ne disparaissent pas significativement24. Enfin, l’échantillon est distribué sur la grille et laisser sécher (iii). D) principes de préparation de grille de cryo-EM à l’aide de la méthode axée sur les cryoWriter : EM An grille recouverte d’un film de carbone holey est placé sur la surface d’une plateforme à température contrôlée et qui s’est tenue par la pince à épiler. La température de la plate-forme est contrôlée à un décalage par rapport à la température de point de rosée de l’environnement de la grille. La grille est déplacée par rapport à la microcapillaire contenant l’échantillon et le microcapillaire s’abaisse jusqu'à ce qu’il est de quelques micromètres au-dessus de la grille. Par la suite, quelques microseringues de l’échantillon sont dispensés d’elle, tandis que la scène est déplacée dans un motif en spirale ; le liquide en excès est ré-aspiré (i). Après la grille EM d’amorçage, le microcapillaire est retiré et la grille reste sur la plateforme contrôlées de température (par la suite appelée stade de point de rosée (DP)) pendant une courte période afin de permettre une quantité contrôlée de l’échantillon de s’évaporer. Pour plongeon de congélation, la grille est rapidement retirée de la scène à l’aide de la pince à épiler (ii), renversée par 90° en position verticale et plongée dans un bain de cryogène (iii) (par la suite dénommé « pick-et-plongeon » mécanisme). S’il vous plaît cliquez ici pour visionner une version agrandie de cette figure.
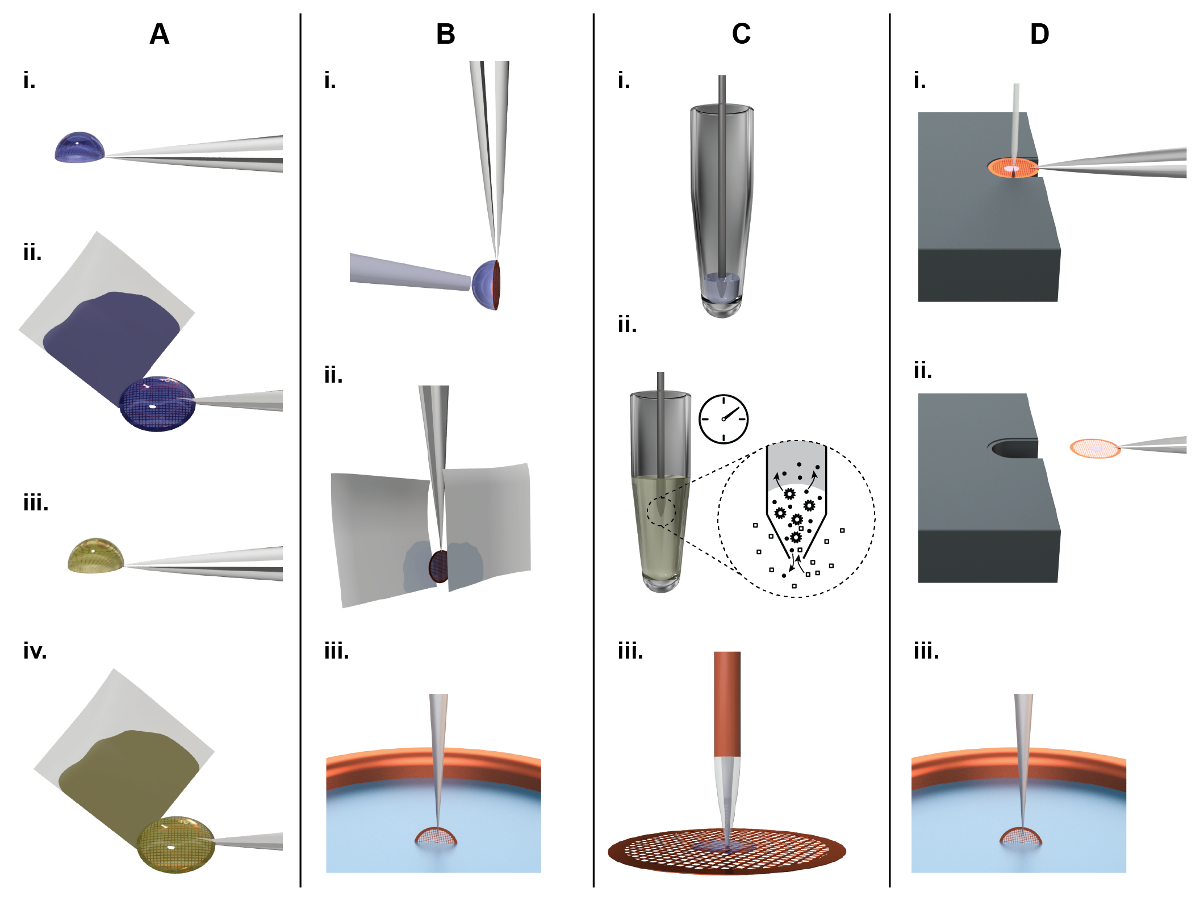{kind=link}
Malheureusement, les méthodes de préparation de grille utilisées pour NS et cryo-EM n’ont pas significativement amélioré depuis qu’ils ont été inventés. Les inconvénients actuels sont la consommation élevée d’échantillon (env. 3 µL de protéine de 1 mg/mL) et la grande quantité (> 99 %) de l’échantillon (Figure 1A, B) a perdu. En outre, la méthode classique utilisée pour préparer les grilles pour cryo-EM est une procédure dure pour les protéines : tout d’abord, il s’agit d’une vaste visage-sur papier-buvard étape (Figure 1B, ii), et, deuxièmement, la protéine est exposée à l’interface air-eau pour une quantité importante de temps21. Ici, une méthode alternative pour échantillon pré conditionnement, préparation des échantillons grille et post-traitement (grille de séchage ou vitrification) pour NS-EM (Figure 1C) ou cryo-EM (Figure 1D) est présenté. Le programme d’installation intégré interne, appelé « cryoWriter », utilise miniaturisé échantillon manutention principes technologiques et microfluidique pour aspirer, condition et de distribuer des échantillons, en évitant de papier buvard complètement et en fournissant des méthodes alternatives aux échantillons minces pour Cryo-EM. Il a considérablement réduit la consommation d’échantillon et améliore le contrôle utilisateur au cours de la préparation de l’échantillon dans son ensemble. En outre, la méthode permet de nouvelles applications expérimentales ; comme la préparation des composants biologiques isolés des cellules individuelles dans une approche appelée « cellule unique visual protéomique »22,23,24,25.
Protocole
Un « cryoWriter » (Figure 2; pour les détails voir précédent travail24,26,27) ou instrumentation équivalente est requise pour les protocoles suivants. Une liste de fournisseurs pour les pièces principales et des consommables est donnée dans la Table des matières.
1. préparation de grille de colorant négatif (NS)
- Mettre l’instrument sous tension et démarrez le logiciel. Initialiser tous les modules nécessaires (régulateur de pompe à seringue, platines motorisées, caméras de surveillance et stade de point de rosée).
- Cool le soutien de l’échantillon et la scène du point de rosée. Si nécessaire, s’assurer que la température de scène de point de rosée est régulée 1-2 ° C au-dessus du point de rosée.
Remarque : La scène est refroidie par un dispositif Peltier commerciaux avec un régulateur PID. - Préparer NS en remplissant un 100 ou 200 µL PCR tube avec 100-150 µL de NS (p. ex., tungstate de méthylamine 2 % disponibles dans le commerce). Placer le tube sur le support d’échantillon refroidi de l’instrument.
- Position de l’échantillon.
- Mettre l’échantillon (0,5 - 1 µM) dans un tube PCR 100 ou 200 µL. Si moins de 50 µL de l’échantillon est disponible, couper le fond d’un tube PCR avec une lame de rasoir et l’utiliser comme un échantillon bien. Cela garantira que le microcapillaire peut atteindre facilement l’échantillon.
Remarque : Il est plus facile à aspirer des échantillons de 100 ou 200 tubes PCR µL, parce que le microcapillaire utilisée ultérieurement pour aspirer l’échantillon est légèrement incliné et la direction de z-AXIS voyage-hauteur est limitée. - Placez le tube PCR/conteneur sur le support d’échantillon refroidi dans l’instrument pour éviter l’évaporation. Alternativement, les échantillons peuvent être aspirés de plats dans l’incubateur haut stade de microscope à température ambiante ; de refroidissement n’est pas implémentée pour les plats.
- Mettre l’échantillon (0,5 - 1 µM) dans un tube PCR 100 ou 200 µL. Si moins de 50 µL de l’échantillon est disponible, couper le fond d’un tube PCR avec une lame de rasoir et l’utiliser comme un échantillon bien. Cela garantira que le microcapillaire peut atteindre facilement l’échantillon.
- Définir des positions. Utilisez le joystick de cryoWriter qui gère les xy-platine motorisée et les boutons de contrôle de logiciel pour le linéaire x, y et étapes d’axe z pour positionner le microcapillaire. La caméra permet de vérifier la position du capillaire.
- Déplacer le microcapillaire vers le réservoir de l’échantillon. Plonger l’extrémité dans le liquide de l’échantillon et enregistrer cette position comme « échantillon ».
- Déplacer le microcapillaire au tube PCR NS, plongez la pointe dans la solution de NS et enregistrer cette position comme « tache ».
- Placez le microcapillaire environ 100µm au-dessus du centre de la fente où la grille EM sera positionné et sauver cette position comme « grid_save ».
- Aspirer l’échantillon et la condition pour NS-EM.
- Si pas déjà installé, installez une seringue de 10 µL (0,46 mm de diamètre intérieur) sur une pompe seringue de précision.
- Coller une extrémité d’un microcapillaire de silice long 30 cm (diamètre extérieur 360 µm, diamètre interne 150 µm) à la sortie de la seringue.
- Connectez l’autre microcapillaire fin le microcapillaire un trop court (5 cm) long conique via connecteur press fit. L’extrémité effilée de la courte microcapillaire constitue l’embout applicateur.
- Remplir la seringue avec de l’eau bidistillée dégazé (ddH2O ; liquide) et éviter la formation de bulles d’air.
- Dispense de quelques dizaines de microseringues de liquide du système et supprimer les gouttes de la microcapillaire avec un chiffon non pelucheux.
- Cliquez deux fois sur la position de l’échantillon enregistrée. Cela positionne bien la microcapillaire dans l’échantillon. Alors que le capillaire est en mouvement, répartir 3 x 0,5 nL de liquide juste avant la pointe microcapillaire est immergée dans l’échantillon pour empêcher la bulle d’air soit emprisonné là (voir note 2 ci-dessous).
- Aspirer 5 ml d’échantillon.
- Cliquez deux fois sur la position enregistrée de NS. Le microcapillaire est retiré du contenant d’échantillon et automatiquement déplacée vers le réservoir de NS. Alors que le capillaire est en mouvement, manuellement distribuer 3 x 0,5 nL de liquide d’essai juste avant la pointe est immergée dans le réservoir pour s’assurer qu’il n’y a pas d’air des bulles (voir note 2 ci-dessous).
Remarque : IMPORTANT : (1) lors du passage d’aspiration au mode de distribution ou vice versa, il y a une petite perte dans la course du piston due au jeu dans les pignons de la pompe à seringue. Selon le fabricant, le contrecoup d’une nouvelle unité se situe entre 7 et 12 µm. Pour notre seringue avec un diamètre de baril de 0,46 mm, cela se traduit par 1-2 nL. Par conséquent, nL 1-2 peut être « servi », avant d’échantillon est effectivement distribué. Habituellement, une minuscule goutte commence à sortir la pointe microcapillaire après le troisième 0.5 nL dispense étape. (2) une bulle d’air piégée au-dessus/au-dessous de l’échantillon serait faire une distribution moins précis et empêcher le conditionnement d’échantillon par diffusion. - Laissez le microcapillaire immergé pour 3-12 min, en fonction de la solution tampon et la géométrie de la buse.
Remarque : Plus le sel et/ou de phosphate concentration dans la mémoire tampon, plus le temps d’immersion requis. NS (assez rapidement) diffuse dans la prise d’échantillon tandis que les sels de tampon (relativement rapide) et les protéines diffuse out (plus lent). Il permet d’abaisser la concentration de sels de la mémoire tampon dans l’échantillon qui les empêche de se cristallisant quand la grille chargée sèche. En outre, phosphate tend à former un précipité en combinaison avec NS.
- Préparer une grille et déposer une tache de l’échantillon conditionné.
- Alors que l’échantillon est étant conditionnée, prenez un morceau de ruban adhésif et un bloc PDMS et nettoyer le dessus de la PDMS en appliquant et en enlevant le ruban adhésif pour s’assurer qu’il n’y a pas de poussière. Mettre le bloc PDMS dans une boîte de Pétri.
NOTE : Nouveaux blocs de PDMS sont tirées de la salle blanche. - Ramasser soigneusement une grille (p. ex., Cu, maille de 200 ou 400 recouvert de film Parlodion/C). S’assurer de toucher uniquement le bord de la grille avec la pince à épiler. Placez-le sur le bloc PDMS propre avec le film de carbone vers le haut.
- Place le bloc PDMS avec la grille dans une unité de décharge luminescente d’air et la lueur décharger pendant 20 s avec une puissance de 100 W à 0,4 mbar. Stocke la grille dans un plat de Pétri fermée. Lueur décharge plus longs se traduisent généralement par une plus grande diffusion du volume de l’échantillon déposé sur la grille. Par conséquent, la couche de la tache s’amincit (faibles taches).
- 1 min avant que le temps d’immersion est écoulé, saisir la grille lueur-déchargé avec la pince cryoWriter. Assurez-vous que l’électro-aimant est activé ; dans le cas contraire, allumez-le dans le logiciel. Monter la pince sur l’électro-aimant et utilisez la vis manuelle micromanipulateur pour aligner la grille à plat sur la scène du point de rosée, côté film carbone haut. Assurez-vous que la température de scène de point de rosée est régulée 1-2 ° C au-dessus du point de rosée afin de réduire le taux d’évaporation après échantillon est chargé.
- Lorsque le temps d’immersion est terminé, double-cliquez sur « grid_save ». La pointe microcapillaire sera placée en toute sécurité au-dessus de la surface de la grille. Manuellement, amener la pointe microcapillaire en contact avec la grille. Soulevez la buse de 10 µm et positionnez-le au-dessus du centre.
Mise en garde : Certains échantillons contenant des détergents ont tendance à remonter le long de la surface extérieure de la microcapillaire lorsque le liquide s’écoule en raison de la tension superficielle plus faible. Il est important d’être très proche de la surface de la grille afin d’éviter de telles pertes. - Verser 5 ml d’échantillon sur la grille. Un jour sec, quand l’évaporation rapide a lieu à la pointe de microcapillaire, on peut distribuer aussi lentement jusqu'à ce que l’échantillon soit à l’extrémité et puis rapidement distribuer 5 nL.
- Retirer le microcapillaire et sécher l’échantillon conditionné lentement sur la scène du point de rosée (DP-stade).
- Une fois que le spot de l’échantillon est sec, retirez la grille et conserver à température ambiante dans une boîte de grille ou de la boîte de Pétri.
- Distribuer 500 nL de liquide du système de la microcapillaire et enlever avec un chiffon non pelucheux. Rincer le tube capillaire 5 fois avec l’éthanol, détergent ou 1 NaOH M. Cela nettoie la microcapillaire lui permettant d’être utilisé avec un échantillon différent.
- Alors que l’échantillon est étant conditionnée, prenez un morceau de ruban adhésif et un bloc PDMS et nettoyer le dessus de la PDMS en appliquant et en enlevant le ruban adhésif pour s’assurer qu’il n’y a pas de poussière. Mettre le bloc PDMS dans une boîte de Pétri.
2. préparation de la grille de Cryo
- Pour préparer l’instrument et l’échantillon, les étapes 1.1 à 1.5 décrit ci-dessus. Si NS n’est pas nécessaire, omettre les étapes 1.3 et 1.5.2. Pour échanger la solution tampon ou conditionner l’échantillon pour cryo-EM par dialyse, par exemple, pour réduire la concentration du tampon de sels ou pour introduire des additifs (par exemple, tréhalose, détergents) utilisent le tampon souhaité au lieu de NS en étapes 1.3 et 1.5.2.
- Préparer l’éthane liquide dans un récipient standard cryo.
- Assembler la coupe de l’éthane, titulaire de zone de cryo et araignée et remplir le réservoir de cryogène à ras bord avec de l’azote liquide ; Il faut habituellement environ 200 mL. Attendez quelques minutes jusqu'à ce que la coupe de l’éthane est refroidi et est libre de l’azote liquide.
- Ouvrir la bouteille de gaz éthane et laissez lentement le flux de gaz dans la tasse de l’éthane. Laissez-le remplir d’éthane liquide jusqu'à ce que le niveau est de 2 à 3 mm au-dessous du sommet ; Cela prend quelques minutes et nécessite environ 5 mL d’éthane liquide.
- Prenez une boîte de cryo et placez-le dans un emplacement libre dans le conteneur de cryogène.
- Supprimer l’araignée, poser le couvercle de polystyrol dessus et placer le récipient de cryogène sur le montage dans le cryoWriter.
- Lampe à décharge luminescente une grille d’EM.
- Prenez un morceau de ruban adhésif et un bloc de polydiméthylsiloxane (PDMS) et nettoyer le dessus PDMS en appliquant et en enlevant le ruban adhésif (supprime les poussières). Mettre le bloc PDMS dans une boîte de Pétri.
- Prélever soigneusement une grille de la maille. S’assurer de toucher uniquement le bord de la grille avec la pince à épiler. Placez-le sur un bloc PDMS propre avec le film de carbone trouée vers le haut.
- Place le PDMS bloquer avec la grille dans un plasma cleaner et plasma-nettoyer la surface de la grille (p. ex. utilisation 75 % Ar/25% H2, puissance 50 W, pression 25 mTorr). Déposer le bloc PDMS avec la grille de lueur-déchargé dans une boîte de Pétri.
- Placer la grille dans l’instrument
- Saisir la grille lueur-déchargé avec la pince cryoWriter. Assurez-vous que l’électro-aimant est activé ; dans le cas contraire, allumez-le dans le logiciel. Monter la pince sur l’électro-aimant et utiliser la vis de micromanipulateur manuel d’aligner la grille plate sur la scène, côté film carbone vers le haut.
- Double-cliquez sur « grid_save ». Ajustez la position de microcapillaire afin que la pointe soit environ 10 µm, au-dessus de la surface de la grille. Assurez-vous que le microcapillaire peut se déplacer librement sur la grille sans le toucher n’importe où, si nécessaire retire le microcapillaire quelques micromètres.
- Revenez vers le centre de la grille et enregistrer la nouvelle position de « grille ».
Mise en garde : La dénomination correcte est obligatoire pour le script de macro travailler.
- Écrire la grille échantillon et plongeon-gel.
- Rincer le microcapillaire avec quelques dizaines de microseringues de liquide du système et supprimer les gouttes de la microcapillaire avec un chiffon non pelucheux.
- Démarrez le script de macro. La macro vous effectuez les opérations suivantes :
- Pipeter 5 nL (pour éliminer les bulles d’air à l’extrémité) et aller à la position de l’échantillon.
- Aspirat 65 ml d’échantillon. Infuser 5 nL dans le tube d’échantillon. Ce qui représente pour le système de jeu et permettre écriture synchronisée, c’est à dire., pour s’assurer que cette distribution et début du mouvement de la scène en même temps.
- Mettez en position « grille ».
- Lancer « écrit modèle », qui provoquera la microcapillaire pour se déplacer dans la grille, en même temps une distribution 45 ml d’échantillon.
- Ensuite, replacer le microcapillaire dans le centre de la grille, abaisser un autre 10 µm et retirer le liquide d’essai excès.
- Retirer le microcapillaire et hors tension de l’électro-aimant. Ceci libère le plongeon de pincettes et initiés de congélation.
- Tenez la pince à épiler et libérer soigneusement la carte magnétique de plongeur. Transvaser la grille de la coupe de l’éthane dans le récipient de cryogène contenant la boîte de cryo et placer la grille dans un slot libre rapidement.
3. cellule unique préparation Lysate
- Préparer le microcapillaries pour l’électroporation.
Remarque : Les microcapillaries conique en silice fondue (laser-tiré et inspecté) ont un revêtement protecteur de polyimide à l’extérieur, à l’exception de la pointe, où le revêtement est brûlé au cours du processus effilé.- Pour enrober le microcapillaries avec une couche conductrice, les monter à un angle de 45° sur un rail métallique, avec les pointes tournées vers le haut. Utilisez une feuille d’aluminium pour protéger l’extrémité inférieure (2 cm) des conseils, car non revêtus de polyimide constitue le meilleur joint avec les connecteurs press-fit employées plus tard.
- Par pulvérisation cathodique déposer une couche collante de 20 nm Ti/W et puis une couche de 200 nm d’épaisseur du Pt.
- Faire la pointe microcapillaire hydrophobe.
- Préparer une solution de 1 M de 1-dodécanethiol en EtOH.
- Lampe à décharge luminescente la microcapillaire dans l’air pendant 1 min avec une puissance de 100 W à 0,4 mbar.
- Plonger l’extrémité de la microcapillaire dans la solution 1 M de 1-dodécanethiol pendant quelques heures (idéalement une nuit).
Remarque : Il est préférable de préparer les conseils fraîchement un jour avant l’utilisation.
- Installer un nouveau microcapillaire.
- Retirer le microcapillaire de la solution de 1-dodécanethiol, rincer l’extérieur avec de l’éthanol et vider l’intérieur avec de l’éthanol à l’aide d’une seringue.
Remarque : Si aucun microcapillaries fonctionnalisés ne sont disponibles, rapidement lueur décharge la pointe d’un microcapillaire et tremper dans une goutte de traitement de fenêtre de voiture commerciale, qui est un mélange de PDMS et d’acide sulfurique. La fonctionnalisation hydrophobe qui en résulte n’est pas aussi bon qu’avec 1-dodécanethiol, mais généralement suffisante. - S’attacher son extrémité non couchée avec un cutter capillaire.
Remarque : Une coupe nette est importante pour une bonne étanchéité avec le connecteur press-fit. - Utiliser un cure-dent pour appliquer la pâte d’argent à la forte boarder formé entre le verre et le revêtement de polyimide lorsque le revêtement de polyimide était brûlé par le laser pendant capillaire en tirant.
Remarque : Ce renforcement est nécessaire car le revêtement Pt est très faible dans cette région, et la conduction électrique peut facilement se casser vers le bas. - Laver à la fin de polyimide de la microcapillaire avec de l’acétone. Laisser une petite goutte d’acétone à la fin et l’insérer dans l’orifice du connecteur press-fit. Appliquez une légère pression pour former une bonne étanchéité.
- Retirer le microcapillaire de la solution de 1-dodécanethiol, rincer l’extérieur avec de l’éthanol et vider l’intérieur avec de l’éthanol à l’aide d’une seringue.
- Calibrer la position microcapillaire.
- Mettre l’instrument sous tension et démarrez le logiciel comme indiqué au point 1.1. En outre, initialiser le générateur de fonction et de la caméra.
- Placez une lame de microscope de verre dans le support de diapositive sur le microscope.
- Abaisser le microcapillaire à proximité de la lame de verre et la centrer sur l’objectif de microscope jusqu'à ce qu’il apparaît dans la vue de caméra microscope. Utilisez les étapes linéaires x - et y-axe pour positionner la pointe de la microcapillaire au centre de l’image. Puis abaissez lentement le bout jusqu'à ce qu’il touche légèrement la lame de verre.
Mise en garde : Choisir de très petits pas (5 µm) vers l’approche finale. Dans le cas contraire, la pointe peut être endommagée. - Appuyez sur le bouton de « Calibrer la buse ». Cela rétracte le microcapillaire 40 mm et ensembles cette position tant que la maison position.
- Préparation d’affranchie des pièces PDMS et ITO glisse
- Mix PDMS et RETICULATION ratio 10:1, verser dans un grand plat de Pétri jusqu'à une profondeur d’environ 2-3 mm. cuire au four à 60° C pendant quelques heures dans un incubateur de l’hybridation ou un dispositif similaire.
- Avec un marteau et un timbre (12 mm de diamètre), couper des trous dans la couche PDMS. Par la suite, utiliser un scalpel pour découper 2 cm long carrés ou des rectangles dont les trous pour obtenir des pièces embouties qui tiennent sur une lame de microscope. Habituellement, deux pièces embouties sont montées sur lame d’un microscope.
- Lavez les morceaux PDMS et les diapositives de ITO, tout d’abord avec du détergent, puis avec l’éthanol à 70 %. Placez-les, mouillé, dans une boîte de Pétri et laissez l’éthanol s’évaporent complètement dans une étuve à 60° C.
- Les diapositives ITO pendant 1 min avec une puissance de 100 W à 0,4 mbar à décharge luminescente et puis appliquer 1 mL de solution d’enrobage (p. ex., poly-L-lysine (PLL)). Incuber pendant 5 min, retirer la solution avec une pipette et d’appliquer 1 mL de ddH2O. légèrement agiter pendant 1 min et puis retirez la ddH2O à l’aide d’une pipette. Sécher les lames à 60° C.
- Préparer la culture cellulaire. Suivez les cellules adhérentes culture protocole standard (p. ex., Arnold, et al. 24 , 25). graines de cellules de ITO lors de trajets de fractionnement/passage normales.
- Prendre une lame fraîchement PLL revêtement ITO et ajouter un morceau PDMS. Appliquer une pression pour former un joint étanche à l’eau. Ce produit petits puits avec le toboggan comme leur base.
- Ajouter environ 300 µL de cellules en suspension dans un milieu frais dans les puits PDMS. La densité de semis doit être environ 75 000 cellules par puits.
- Incuber les lames ITO pendant 1 à 2 jours dans des conditions normales.
- Préparer à l’expérience de la lyse cellulaire :
- Réchauffez quelques millilitres de la mémoire tampon d’électroporation (p. ex., en solution saline tamponnée au phosphate (PBS)).
- Préparer la mise en place pour la préparation de grille NS-EM
NOTE : Détails pour préparer la mise en place sont indiquées dans les étapes 1,1 à 1,5 (pour NS) ou étape 2.1 à 2.4 (pour cryo). Nous décrivons ici les étapes les plus importantes pour NS-préparation.- Préparer NS en remplissant un 100 ou 200 µL PCR tube avec 100-150 µL de NS (p. ex., tungstate de méthylamine 2 % disponibles dans le commerce). Placer le tube sur le support d’échantillon refroidi de l’instrument.
- Déplacer le microcapillaire au tube PCR NS, plongez la pointe dans la solution de NS et enregistrer cette position comme « tache ».
- Charger les paramètres standard de lysis, par exemple, la tension de la lyse.
- Prendre la diapositive ITO de l’incubateur et installez-le sur l’insert de microscope. Utiliser deux vis pour fixer la lame sur l’insert en aluminium et d’assurer un contact électrique entre l’ITO revêtements de la diapositive et l’armature en aluminium électriquement reliée à la terre.
- Enlever le milieu de culture cellulaire et laver deux fois avec 300 µL de tampon d’électroporation. Garder les cellules dans le tampon d’électroporation.
- Placer l’insert d’aluminium tenant la diapositive ITO dans la scène de l’incubateur de cellules vivantes sur le programme d’installation.
- Localiser la culture de cellules dans la vue au microscope et choisissez un endroit avec aucune cellule. Approcher la pointe à la surface de ITO et doucement toucher, puis retirer la pointe 100 µm et conserver la position de « cellules ».
- Rapidement quitter la culture de cellules et rincer la pointe microcapillaire avec quelques dizaines de microseringues de liquide puis mettez-la à nouveau dans la culture de cellules. Répartir quelques microseringues durant l’immersion dans le puits PDMS à veiller à ce qu’aucun air bulles sont emprisonnées à la pointe.
- Approcher doucement la surface de l’ITO (au départ, vous devez être postées sur « cellules », i.e., 100 µm au-dessus de la surface). Au contact, rétracter pointe 10 µm.
- Sélectionnez une cellule voisine pour la lyse. Placer l’embout de la microcapillaire au-dessus de la cellule cible.
- Lancez le script de macro pour la lyse de la cellule unique.
- Nom de la position du microcapillaire doit être déplacé à après qu’une cellule a été lysée avec succès. Ce sera le réservoir de NS (NS-EM), un tampon de dessalement (cryo-EM) ou la grille d’EM (cryo-EM). Éviter les fautes d’orthographe, puis appuyez sur « OK ».
- La macro produit sans intervention de l’utilisateur :
- i) le microscope stade et cell culture 100 µm sont déplacés vers la gauche, une capture instantanée de la cellule cible est pris et 50 nL de ddH2O liquide sont dispensés de la microcapillaire. Cela déplace et dilue le tampon salin haute et applique une pression osmotique à la cellule.
- II) la scène est déplacée à nouveau pour positionner la pointe au-dessus de la cellule cible à nouveau. L’éclatement de tension prédéfinie est appliquée, et après 500 ms, le système de la pompe commence à aspirer 3 ml d’échantillon à un débit de 2 µL/min.
- III) la scène est déplacée vers la gauche encore une fois, permettant à la cellule pour être inspectés. Une fenêtre apparaît, vous demande d’entrée utilisateur.
- Dire si l’étape de lyse a réussi ou non.
- Si la réponse est non, reprendre, rincer le microcapillaire et cibler une nouvelle cellule.
- Si la réponse est oui, un instantané de la cellule (lysé) supprimé est pris, et ensuite le microcapillaire est déplacé vers l’emplacement spécifié dans 3.8.1. Si c’est NS ou un réservoir tampon, utilisez l’interface utilisateur standard pour distribuer 3 x 0,5 nL de liquide tout en la microcapillaire se déplace pour s’assurer qu’il n’y a pas de bulles d’air. L’extrémité est plongée dans le liquide du réservoir par la macro.
Remarque : Vous pouvez continuer à conditionner l’échantillon et de préparer les grilles comme expliqué dans les sections 1.6.9 - 1.7.9 NS-EM ou une section 2,2-2,6 pour cryo-EM. Ci-dessous, les mesures nécessaires pour la préparation de la NS-grille sont expliqués.
- Laisser le microcapillaire immergé pendant 8-12 min, en fonction de la géométrie de la buse
Remarque : La concentration de phosphate élevées dans la mémoire tampon nécessite un temps d’immersion plus longs pour climatisation. - Pipeter 5 nL de la cellule conditionnée lysate sur la grille.
- Retirer le microcapillaire et laisser l’échantillon conditionné sèche lentement sur la scène du point de rosée (DP-stade) régulée 1-2° C au-dessus de la température de point de rosée.
- Retirez la grille et conserver à température ambiante dans une boîte de grille ou de la boîte de Pétri.
Résultats
L’installation de « cryoWriter » a été développée (illustrée à la Figure 2) afin de tester les procédures de préparation de grille EM miniaturisés proposés en Figure 1C, D. Figure 2 A montre une vue d’ensemble des différents composants monté sur un microscope à fluorescence inverse. Un module de culture de cellules est installé sur le côté gauche du microscope ; un module pour la préparation de grille EM se trouve sur la droite. Le module de culture de cellules (Figure 2B) permet la croissance des cellules eucaryotes adhérentes et imagerie de cellules vivantes de la culture cellulaire par microscopie. Les cellules sont lysées par l’action combinée d’un choc osmotique, électroporation et l’aspiration du contenu cellulaire dans un microcapillaire (Figure 2B, Figure 6A)24,25. L’échantillon de lysat aspiré permet ensuite de préparer les grilles pour les NS - ou cryo-EM. Une solution stock de protéines dans un tube PCR peut également être la source de l’échantillon. Le microcapillaire (Figure 2B) employée est connecté à un système de pompe de haute précision permettant des volumes d’échantillon être aspiré et dosé avec précision sub-nL. Tel que décrit dans les protocoles, tous les traitement des échantillons est effectué au sein de cette microcapillaire ou sur la grille de EM elle-même sans transport d’échantillon significatif. Par exemple, le même microcapillaire sert à lyser les cellules eucaryotes individuels, aspirer le lysat, condition il et enfin passer outre aliquotes sur grilles de EM. Le module de préparation de grille se compose d’une DP-scène mobile qui permet à la température de la grille de EM placée dessus pour être précisément contrôlé (Figure 2C). Pour NS-TEM, la grille de l’échantillon préparé simplement soient retirée de la scène froide et laisser sécher à l’air à température ambiante. Cependant, les effets dits de café-anneau qui peuvent alors entraîner doivent être évitées pour TEM quantitative où la protéine « particules » sont comptés. Pour ce faire, grilles sont séchées lentement sur la scène DP à l’aide d’un gradient de température augmente graduellement à ralentir à évaporation liquide. Pour cryo-EM, la température de la grille est maintenue près du point de rosée ; un offset positif d’environ 8 ° C est choisi, ce qui permet l’évaporation contrôlée de liquide d’essai pour la stabilisation de la couche mince et amincissement, qui peut être surveillée par un capteur si nécessaire26. Après l’heure fixée éclaircie, un mécanisme de choix-et-plongement est activé et l’échantillon est vitrifiée (Figure 2C). Notez que ce mécanisme plongeant n’est pas nécessaire pour les grilles de NS-EM, qui sont stockés à température ambiante.

Figure 2: vue d’ensemble de l’installation de cryoWriter. A) vue d’ensemble de l’installation de cryoWriter monté sur un microscope à lumière inverse (1). B) médaillon de zone indiquée sur le côté gauche au panneau de compartiment de culture cellulaire A. (2), avec un microcapillaire (3) pour la lyse placée au-dessus d’une plaque de culture de cellules de base PDMS miniaturisés (4) de la manipulation et de cellule échantillon. C) médaillon de zone indiquée sur le côté droit au mécanisme de panneau a. « Pick-et-grand saut ». Une grille d’EM film carbone holey (5) est montée entre les extrémités des pinces (6) et positionnée horizontalement en contact direct avec la phase de contrôle de la température (7), appelé le stade de point de rosée (DP-stade) dans le texte principal. La température de l’étape est étroitement contrôlée par un régulateur PID et un élément Peltier refroidi à l’eau, garder à ou proche de la température de point de rosée, selon le milieu ambiant. La DP-scène (7) est montée sur un axe xy motorisée pour déplacer la grille par rapport à la microcapillaire. Le microcapillaire lui-même est monté sur un z-stade et peut être abaissée jusqu'à ce qu’il est très proche de la surface de la grille de l’EM et utilisé pour dispenser nanolitres taille des volumes sur le support de l’échantillon en le couvrant (une couche de carbone mince continu pour NS-EM ou d’un film de carbone trouée pour cr Yo-EM). Notez qu’absorption de liquide et de distribution s’effectue à l’aide d’un système de pompage de haute précision (8). Le liquide dosé peut être distribué en déplaçant la grille par rapport à la microcapillaire en forme de spirale. Pour la préparation de la cryo-EM, le mécanisme de gel de pick-et-plongeon (9) transfère rapidement la grille échantillon-chargé en éthane liquide (10) pour un refroidissement rapide et la vitrification de l’échantillon. S’il vous plaît cliquez ici pour visionner une version agrandie de cette figure.
{kind=link}
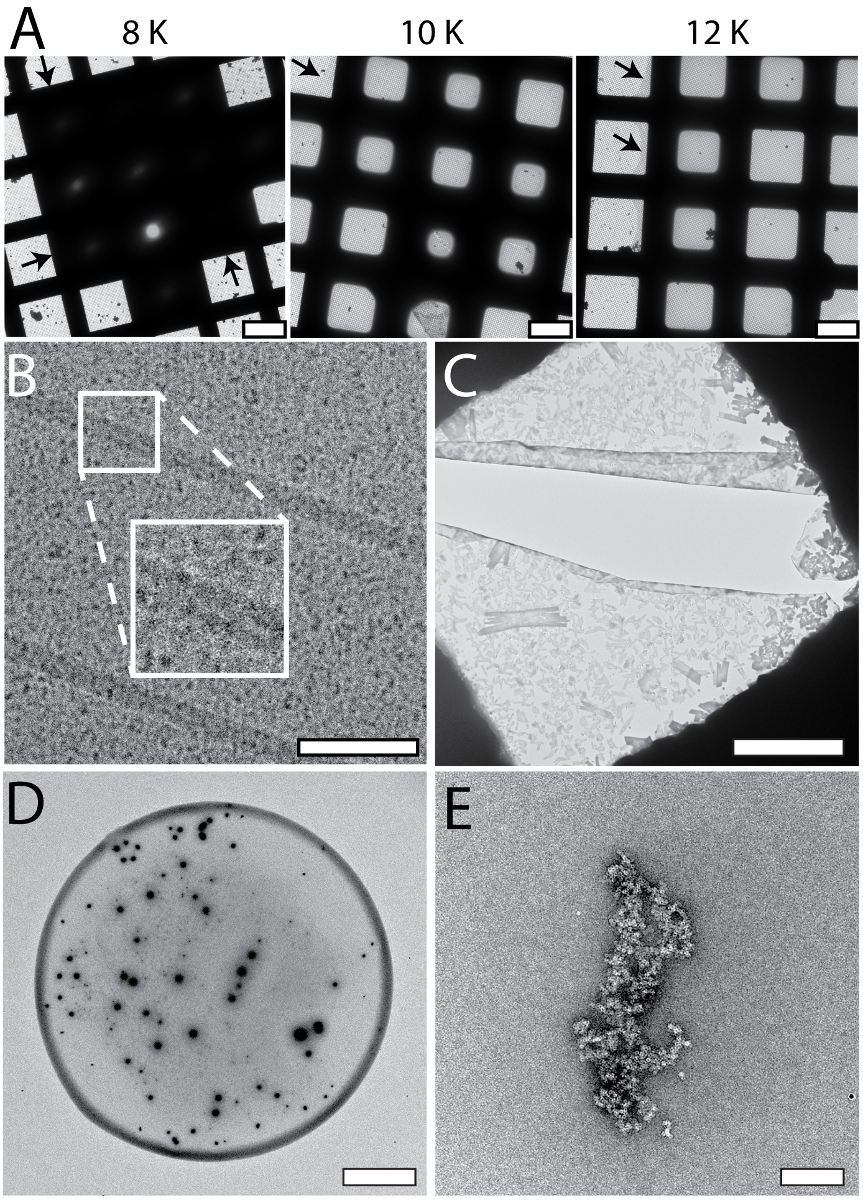
La figure 3 illustre représentant résultats obtenus pour les grilles de NS-EM préparés à l’aide de la configuration de cryoWriter. La pointe de la microcapillaire a été chargée avec 5 nL d’échantillon d’une solution mère et plongé dans un réservoir de solution NS (tungstate de méthylamine 2 %) pendant plusieurs minutes pour permettre l’échange diffusif de NS et des ions de sels (pour une discussion théorique voir Arnold, et coll. 24). par la suite, l’échantillon conditionné a été distribué sur le film de carbone mince d’une grille de NS-EM et séché. Figure 3 A montre l’utilisation d’une grille de fente de la même façon de visualiser la goutte complete, tel que requis pour TEM quantitative. Afin d’éviter l’effet d’anneau de café, la grille fraîchement déchargées lueur était au départ qui s’est tenue à la température de point de rosée (aucune évaporation de l’eau) et ensuite lentement réchauffée sur la DP-scène. Notez que pour la plupart des applications (par exemple, contrôle de la qualité de l’échantillon ou l’analyse structurelle) ce processus de séchage lent n’est pas nécessaire. NS de haute qualité-préparations sont obtenues sans lui, comme le montre la Figure 3B, C. Fois de conditionnement pour les tampons sels faibles sans phosphate sont environ 3 min, par exemple, avec une faible teneur en sel Tris-tampon (20 mM Tris-HCl pH 7,4 avec 50 mM NaCl), comme illustré à la Figure 3B à l’aide de virus de la mosaïque du tabac (TMV) comme exemple. Figure 3 C présente un scénario du pire que le TMV était dans le tampon PBS (2,7 mM KCl, 1,5 mM KH2PO4, 136,9 mM NaCl, 8,9 mM Na2HPO4·7H2O, pH 7,4). Ions phosphates forment des cristaux transitoires avec les ions de métaux lourds de la Nouvelle-Écosse (voir Figure 5C), allonger le temps de conditionnement requis (7 min). Autres sels de métaux lourds peuvent également être utilisés avec le module de préparation de grid, par exemple, 2 % méthylamine vanadate ou ammonium molybdate (voir aussi Arnold, et al. 24). Cependant, l’acétate d’uranyle n’est pas approprié ; l’effet de la réticulation de cette tache conduit aux agrégats si l’échantillon de protéine est conditionnée dans une solution, avant de l’adsorption d’un carbone film (voir Figure 5E)23.

Figure 3: résultats typiques pour les grilles de NS préparés à l’aide de la configuration de cryoWriter comme il est indiqué dans la Figure 1C. A) image d’aperçu d’une goutte de nL 3 distribué sur une grille de fente après conditionnement avec tungstate de méthylamine 2 %. B) VMT dans un tampon TRIS 20 mM. L’encart montre un agrandissement de 3 x de la région indiquée. Adapté de Arnold, Al24 (autres autorisations liées au matériau extrait doivent être adressées à l’ACS). C) virus de la mosaïque du tabac (TMV) dans le tampon PBS. L’encart montre un agrandissement de 3 x de la région indiquée. Adapté de Arnold, Al24 (autres autorisations liées au matériau extrait doivent être adressées à l’ACS). Barreaux de l’échelle : A, 100 µm ; B, 50 nm ; C, 80 nm. S’il vous plaît cliquez ici pour visionner une version agrandie de cette figure.
{kind=link}
Résultats typiques obtenus pour les grilles de cryo-EM préparés à l’aide de la configuration de cryoWriter sont représentés dans la Figure 4. Panneau 4 a montre un atlas de la grille de la zone couverte par exemple vitrifié. Groupe 4 b montre l’homogénéité de la glace vitreuse dans une fente de grille sélectionnée. Dans les deux cas, l’échantillon était de 25 millimètres HEPES-KOH pH 7.5, un tampon 50 mM NaCl contenant 0,05 % de Fos 14 détergent. De nombreux échantillons et tampons ont été testés et une glace vitrifiée de qualité comparables ont été obtenus, mais les conditions requises sont dépendants de tampon (voir aussi la discussion de la Figure 5). Panneau 4C montre apoferritine particules et un bactériophage dans un tampon Tris-HCl (20 mM Tris-HCl 50 mM NaCl ; pH 7,4) photographié à défocalisation haute pour augmenter le contraste. Panneau D 4 montre une protéine de membrane de 200 kDa stabilisée par amphipoles.

Figure 4: résultats typiques pour les grilles de cryo-EM préparés à l’aide de la configuration de cryoWriter comme il est indiqué dans la Figure 1D. Les échantillons et les tampons varient dans les exemples montrés. Tous les échantillons ont été chargés sur des films carbone trouée. A) Collage d’images d’aperçu (« atlas de la grille ») d’un échantillon contenant une protéine de la membrane 150 kDa, la périphérie de la glace vitrifiée est indiqué par les flèches blanches. B) fente grille agrandie d’une grille préparée avec le même tampon, la projection du film de carbone holey avec glace vitrifiée. Certains trous ne sont pas remplis de tampon échantillon comme indiqué par les flèches blanches. C) trou carbone vitrifié échantillon contenant des complexes de protéine apoferritine et les bactériophages. En médaillon : double l’élargissement montrant la queue d’un bactériophage. L’astérisque blanche indique le film carbone. Notez que l’image a été enregistrée avec la défocalisation haute pour augmenter le contraste. D) A 200 kDa protéine membranaire reconstitué dans amphipols. En médaillon : un 2 x l’élargissement de la région indiquée avec un contraste accru. Barreaux de l’échelle : A, 100 µm ; B, 10 µm ; C et D, 80 nm. S’il vous plaît cliquez ici pour visionner une version agrandie de cette figure.
{kind=link}
La configuration cryoWriter permet systématique de dépistage des conditions optimales de préparation des EM-grille ; un exemple est illustré à la Figure 5A (apoferritine dans 25 millimètres HEPES-KOH pH 7.5, 50 mM NaCl, 0.05 % 14 Fos). Dans cette expérience, la glace vitreuse « éclaircie » température variait, mais le temps éclaircie (c'est-à-dire, l’écart de temps entre l’exemple d’application et plongeante gel) est resté constant (1 s). À basse température de décalage (par exemple, 8 K), la couche de l’échantillon était trop épaisse. Aux températures élevées de décalage, la glace vitreuse dans les trous était plus mince (10 K, 12 K), jusqu'à ce qu’à un certain stade (au-dessus de 18 K), la grille est devenu complètement sec (non illustré). Dans les résultats présentés ici, un décalage de 12 K conduit à une grande surface homogène de glace vitreuse comme indiqué par les flèches noires. De telles expériences d’optimisation peuvent être effectuées avec le tampon de l’échantillon cible à l’aide de protéines « test » (comme l’apoferritine). Les meilleures conditions sont ensuite appliquées à l’échantillon cible. En outre, les grilles avec des paramètres loin de l’optimum peuvent souvent être reconnus au cours de la procédure de préparation et n’ont pas besoin d’être projeté au microscope électronique, gain de temps considérable. La figure 5 illustre également une galerie de typique cryo-EM (groupe B) et les artefacts de NS-EM (panneaux C à E) spécifiques à la configuration de cryoWriter. La grille de cryo-EM présentée au panneau 5 b avec TMV dans du PBS contenant 0,1 % décyl-β-D-maltopyranoside (2,7 mM KCl, 1,5 mM KH2PO4, 136,9 mM NaCl, 8,9 mM Na2HPO4·7H2O, pH 7,4, 0.1%DM) était excessivement amincie. L’arrière-plan de l’image est granuleuse, parce que la concentration en sel est devenu trop élevée. En général, l’aspect visuel d’un échantillon ne semble pas être une fonction linéaire de la concentration en sel ; grains de soudainement devient importants lorsqu’un seuil de concentration est atteint au cours du processus d’amincissement. Notez que les substances indésirables peuvent être retirés par une étape de conditionnement avant la préparation de la grille, comme pour le NS-EM dans protocole point 1.6. NS peut causer des autres artefacts. Dans l’exemple illustré dans panneau 5C, tampon PBS (2,7 mM KCl, 1,5 mM KH2PO4, 136,9 mM NaCl, 8,9 mM Na2HPO4·7H2O, pH 7,4) sans échantillon était conditionnée en tungstate de méthylamine 2 % à 3 min. précipités et cristaux sont évident et exercent des forces sur la surface de carbone conduisant à des fissures. La seule forme de précipités dans une certaine concentration de PBS et NS varient et peuvent être évités en conditionnant l’échantillon pour le plus long (cf. Figure 3C). Panneau 5D montre la périphérie d’une goutte d’échantillon NS dosée présentant un « anneau de café ». Cela perturberait l’analyse quantitative, total et peut être évité en ralentissant le processus de séchage, c'est-à-direen gardant la grille de l’EM à la température de point de rosée au cours de l’exemple d’application et puis en augmentant graduellement la température pour sécher) Voir la Figure 3A). 5E groupe (apoferritine dans 20 mM HEPES, pH 7.0, conditionné 3 min) montre l’activité de réticulation de teinté d’acétate d’uranyle 2 %, qui ne permet pas d’échantillons de protéines condition avant ils sont adsorbés aux supports de film carbone.

Figure 5: changements systématiques et artefacts observés lorsque la mise en place de cryoWriter a été utilisé pour préparer les grilles pour les NS - et cryo-EM. A) variation épaisseur de glace vitreuse systématique ; optimisation de la préparation de la grille de cryo-EM. La température de la DP-étape a été variée (de 8 à 12 K) gardant la constante de temps éclaircie (1 s). Les flèches indiquent la périphérie de la couche de l’échantillon. B) sel effets ; trop fortement concentré, c'est-à-direl’étape éclaircie était trop long. L’encart représente un élargissement de 2 x de la région indiquée. C) sel précipité formée par tampon PBS en présence de sels de métaux lourds. Tampon PBS contenant les échantillons doivent être conditionnés plus d’échantillons dans les autres tampons. Ici, tampon PBS sans échantillon a été conditionnée en tungstate de méthylamine 2 % pendant 3 min, un temps typique pour les autres tampons de l’échantillon. Notez la fissure dans le film de carbone très probablement en raison des forces puissantes de précipités agissant sur le support mince lors du séchage. D) effet « Cercle café ». E) apoferritine conditionné dans l’acétate d’uranyle de 2 % à 3 min. des ions uranyle pièce activité significative de réticulation et l’apoferritine grappes forment des agrégats grand. Barreaux de l’échelle : A, 80 µm ; B, 80 nm C, 12 µm ; D, 80 µm ; E, 200 nm. S’il vous plaît cliquez ici pour visionner une version agrandie de cette figure.
{kind=link}
La petite quantité et le volume requis pour la préparation de grille EM en utilisant la configuration de cryoWriter permet de nouveaux types d’expériences. Par exemple, le contenu total d’une seule cellule soient collecté et préparé pour NS - et cryo-EM. La procédure est indiquée dans la Figure 6A. Une cellule eucaryote adhérente (HEK 293) est lysée par électroporation simultanée et échantillon aspiration (panneau 6 a)25. Un volume total de 3 nL est aspiré, qui contient le lysat cellulaire et est conservée dans le microcapillaire pour un traitement ultérieur. Pour le NS-EM est indiqué dans le tableau 6 b, le milieu de culture de cellules a été échangé avec le tampon PBS (2,7 mM KCl, 1,5 mM KH2PO4, 136,9 mM NaCl, 8,9 mM Na2HPO4·7H2O, pH 7,4) avant la lyse cellulaire. Le contenu de la cellule ont été aspiré dans 3 nL du tampon et conditionné dans un réservoir de NS comme indiqué dans la Figure 1C pendant environ 10 minutes plus tard, un volume de nL 5 a été distribué sur le film de carbone continue d’une grille de NS-EM. Protéines individuelles, par exemple, filaments d’actine et patches de membrane avec protéines ci-joint peuvent être reconnus dans l’image. Pour la cryo-EM, illustré à la Figure6 C, un volume de 3 nL a été distribué sur une grille de holey carbone EM sans ré-aspiration pour enlever le liquide. Le film relativement épais d’échantillon formé a été largement dilué avant vitrification. Pour ce faire, la température de la DP-stade a augmenté progressivement, à partir de la température de point de rosée. Le processus d’amincissement était surveillé par un système de capteurs en temps réel jusqu'à un seuil prédéterminé déclenchant le mécanisme de « pick-et-plongeon » et la vitrification d’échantillon (pour détails, voir Arnold, et al. 26). structures membranaires et les protéines peuvent être reconnus dans l’image.

Figure 6: Single cell protéomique visuelle à l’aide de la mise en place de cryoWriter. A) la lyse d’une seule cellule eucaryote adhérente. La cellule est cultivée (1, vert) sur une fonctionnalisés, ITO enduit (rouge)-lame de verre (2) dans un miniaturisés de Petri (3)25. La couche d’ITO est relié à la terre. La cellule est approchée par le microcapillaire (4), qui est recouvert de platine. Un choc osmotique initial (ne pas indiqué) est donné pour faciliter la lyse, qui s’effectue par une série d’impulsions électriques et par les forces de cisaillement exercées lors de l’aspiration du lysat cellulaire. Le processus peut être surveillé par microscopie photonique ; l’objectif du microscope est indiquée (5). Pour plus d’informations, consultez notre précédent travail24,25. B) image de NS-EM du lysat d’une cellule individuelle de HEK 293. Filamenteuses taches actine et de la membrane avec des protéines ci-joint sont visibles. Panneau adapté de Arnold, Al24 (autres autorisations liées au matériau extrait doivent être adressées à l’ACS). C) image de Cryo-EM du lysat d’une cellule individuelle de HEK 293. L’encart montre un agrandissement de 2 x de la région indiquée où les structures de la membrane typique avec protéines associées sont visibles. Panneau C adapté de Arnold, Al 26 barres d’échelle : B, 50 nm ; C, 80 nm. S’il vous plaît cliquez ici pour visionner une version agrandie de cette figure.
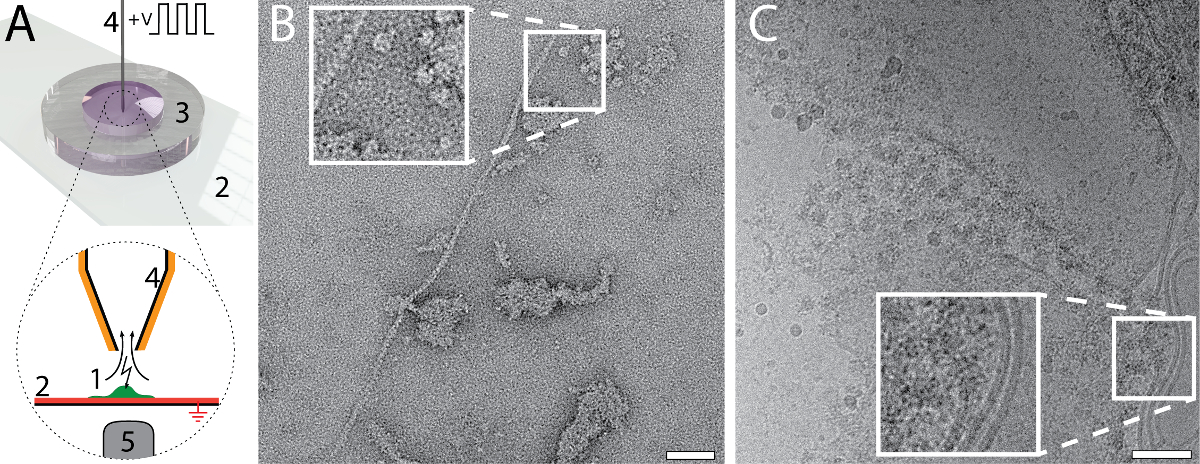{kind=link}
Discussion
L’instrument de « cryoWriter » et les protocoles requis pour préparer les grilles échantillon pour NS - et cryo-EM des volumes d’échantillon total nL taille et complètement éviter l’étape de papier-buvard classique sont présentés. Principes de microfluidique et un système de micromécanique sont combinés dans le cryoWriter pour rendre cela possible.
Notre expérience montre que lorsque les méthodes miniaturisés présentées dans ce manuscrit sont utilisés, l’espace de paramètre pour la préparation de l’EM-grille est plus grand que des méthodes classiques et sous un contrôle plus strict utilisateur. Ce qui est important, la reproductibilité accrue atteinte permet de pré-visualiser avec système de tampon échantillon, complété par une protéine test facilement disponibles, pour déterminer les paramètres optimaux avant l’expérience réelle ne soit effectuée. Ceci maintient la consommation de l’échantillon d’intérêt à l’absolu minimum et est fortement recommandée. Les étapes cruciales pour les deux préparation de grille NS - et cryo-EM sont : (i) d’amorçage du système pompe ; pour sup-nanolitres toute distribution de volume, le liquide (eau) doit être dégazée et bubble gratuit. (ii) contrôle précis de la lampe à décharge luminescente ou plasma nettoyage étape ; les caractéristiques de surface de la grille de l’EM sont essentiels pour obtenir des résultats reproductibles. (iii) conditionnement d’échantillon ; le temps requis pour le conditionnement, par exemple, avec NS, dépend le type de tampon, la teneur en sel et la concentration (Figure 3), ainsi que sur la géométrie de la buse de la microcapillaire24. (iv) taux d’évaporation des préparatifs de la NS-EM EM quantitative ; l’effet du café-anneau peut interdire l’analyse quantitative des préparations de NS-EM et doit être supprimée par des taux d’évaporation lente contrôlées par la DP-scène.
Différents aspects des méthodes de préparation de la grille présentée librement combinables permettant l’élaboration de protocoles polyvalents pour des exemples spécifiques. Des exemples typiques seraient l’élimination d’une substance qui empêche les EM à haute résolution, par exemple, glycérol, par une étape de conditionnement avant la préparation de la grille pour cryo-EM ; l’introduction de molécules de médiateur, comme ligands, par conditionnement avant que les grilles sont préparées ; ou l’examen de la seule cellule de lysat de NS - ou cryo-EM (Figure 6).
L’utilisation de la microfluidique et des quantités d’échantillon minimale dans les méthodes présentées supprime complètement le besoin d’étapes papier-buvard. Il s’agit d’un grand avantage, car le papier buvard est un traitement sévère pour les protéines, potentiellement contaminer l’échantillon avec les ions indésirables et intrinsèquement conduisant à la perte massive d’échantillon. En revanche, les effets potentiels de l’interface air-eau du film mince échantillon formé quand les échantillons de cryo-EM sont préparés à la manière classique sont évités pas lorsque le cryoWriter est utilisé. Grilles pour cryo-EM peuvent être préparés avec un temps d’attente inférieur à 0,2 s entre l’exemple d’application et de la vitrification (données non présentées). Cependant, comme protéines voyagent quelques dixièmes de nanomètre en quelques nanosecondes par diffusion, il est encore assez de temps pour eux d’entrer en collision avec l’interface air-eau d’un film d’épaisseur d’échantillon nm 100 plusieurs fois. Cependant, la quantité de protéines s’en tenir à l’interface air-eau peut être significativement réduite par ces lacunes de peu de temps et pourrait empêcher la dénaturation de la protéine ou restreinte d’orientation de la particule. Une autre approche prometteuse qui pourrait protéger les protéines sensibles de l’interface air-eau est de couvrir le film échantillon de tensio-actifs de faible poids moléculaire. Ces composés pourraient être rapidement présentés par une étape de conditionnement dans le cryoWriter avant la préparation de la grille. Le rapport surface-volume élevé des systèmes microfluidiques est une autre limite de le cryoWriter, comme échantillon peut potentiellement être perdu par adsorption non spécifique à la surface microcapillaire et perturber l’analyse quantitative par comptage des particules. Le problème est abordé de deux façons : tout d’abord, l’échantillon ne déplace pas dans le microcapillaire de longues distances. En effet, le volume d’échantillon nanolitres reste à la pointe capillaire tout au long du traitement. En second lieu, la surface à rapport volumétrique est plus réduite en utilisant microcapillaries avec relativement grands diamètres intérieurs, par exemple, 180 µm. Troisièmement, les surfaces de la microcapillaries peuvent être facilement passivés, si nécessaire, par exemple, en les traitant avec glycols d’éthanol polylysine commercialement disponibles (PLL-PEG).
L’analyse à haute résolution des protéines par l’approche d’une seule particule utilisée dans EM exige seulement 100 000 pour quelques images de millions de particules de protéines individuelles. Cela signifie que les techniques microfluidiques peuvent fournir assez complexes de protéine pour l’enquête structurelle. Une méthode immuno-précipitation miniaturisés pour l’isolement rapide de complexes protéiques (environ 1 heure) des montants minimes cellulaire (env. 40 000 cellules) a été développée plus tôt28. Cette méthode est maintenant directement liée à la phase de préparation d’échantillon miniaturisée de la cryoWriter. L’objectif final est de développer un pipeline intégré microfluidiques pour protéine ultra-rapide d’isolement et cryo-EM grille préparation qui nécessite moins de deux heures en tout. En outre, comme le montre la Figure 6, la quantité infime et le volume du matériel nécessaire pour la préparation des échantillons et la climatisation presque sans perte et la procédure de préparation de grille réalisée à l’aide de la cryoWriter, permettent d’étudier la protéine complexes de cellules individuelles. Ensemble, la méthode d’immuno-précipitation miniaturisés et le cryoWriter jeter les fondements d’une nouvelle méthode de protéomique, appelée « cellule unique visual protéomique », comme nous l’a récemment démontré pour le choc thermique expériences24. Les algorithmes d’analyse de données axées sur l’analyse d’images « protéomique visuelle » sont actuellement à l’essai.
Déclarations de divulgation
Les auteurs Stefan A. Arnold, Henning Stahlberg et Thomas Braun déclarent l’intérêt financier concurrent suivant : le concept de cryoWriter fait partie de la demande de brevet PCT / EP2015/065398 et EP16194230.
Remerciements
Les auteurs aimeraient remercier l’atelier de la Biozentrum de l’Université de Bâle pour leur soutien, S. A. Müller de discussions critiques et de lire attentivement le manuscrit, A. Fecteau-LeFebvre pour l’assistance technique avec EM, Ricardo Adaixo, Frank Lehmann pour protéine membranaire tester des échantillons (tous des C-CINA, Biozentrum, Université de Bâle) et A. Engel, émérite Université Bâle pour ses conversations inspirantes. Les échantillons ont été gracieusement fournis par P. Ringler, m.-a Mahi et T. Schwede (Biozentrum, Université de Bâle), P. Leiman (laboratoire de biologie structurale et biophysique, EPFL) et R. Diaz-Avalos (Structural Biology Center de New York, États-Unis). Le projet a été soutenu par l’Institut suisse de nanosciences (SNI, projet P1401, projet ARGOVIA MiPIS) et le Swiss National Science Foundation (SNF, projet 200021_162521).
matériels
| Name | Company | Catalog Number | Comments |
| Liquid handling | |||
| Microcapillary tip | New Objective | FS360-100-30-N-20 | SilicaTips 30 µm tip ID, 100 µm ID , pk. 20 |
| FS360-150-30-N-20 | SilicaTips 30 µm tip ID, 150 µm ID , pk. 20 | ||
| Conductive sleeve | New Objective | CONGAS-1 | Conductive Elastomer for HV contact, 12" long |
| Fused silica tubing | BGB-Analytik | TSP-150375 | TSP Standard FS Tubing, 150 µm ID, 363 µm OD, 5 meter |
| PressFit Connector | BGB-Analytik | 2525LD | Deact. PressFit Connector 0.25 to 0.25 mm ID, pk. 25 |
| Syringe 10 µl | BGB-Analytik | HA-80001 | Hamilton 1701 LT - Luer Tip (needle not included) |
| Syringe pump controller | Cetoni | A3921000093 | neMESYS controller |
| Syringe pump dosing unit | Cetoni | A3921000095 | neMESYS dosing unit |
| Temperature control | |||
| Dew point sensor | Meltec | UFT75-AT | Humidity/Temperature sensor, dew point calculation, USB and embedded DLL |
| Temperature sensor | Sensorshop24.ch | LS7-PT1000-1.0-3L | Surface mountable temperature sensor Pt1000 |
| Peltier controller | Cooltronic | TC2812 | PID temperature controller for activation of Peltier driven systems with constant voltage output |
| Peltier element | Distrelec.ch | PE-127-14-15-S | 40x40 mm |
| Water-cooling block | - | - | Water cooling block for 40x40 mm peltier element, copper base |
| Water pump | Digitec.ch | 294877 | XSPC X20 750 Dual 5.25 Bay Reservoir Pump Black V4 |
| Water heat exchanger block | - | - | Water heat exchanger block with 2 fans to cool down circulating water |
| Small water coolers | PCHC.ch | 223050 | Alphacool MCX ram copper edition 2 pcs |
| Small peltier elements | Distrelec.ch | PE-031-10-15-S | Laird 15x15 mm 3.4 A 8.1 W 3.8 V 74 °C, 2 pcs |
| Mechanics | |||
| Linear stage z-axis | Dyneos AG | M-404.2PD | High-load precision translational stage, 50 mm travel range, DC motor |
| Stage controller | Dyneos AG | C-863 | Mercury Servo Controller |
| Microscope xy-axis stage | Prior Scientific | H117EIL5 | Motorized stage for Zeiss Axiovert 200 inverted microscope |
| Small permanent magnets | Supermagnete | S-05-08-N | Rod magnet Ø 5 mm, height 8.47 mm, neodymium, N45, nickel-plated, pk. 10 |
| Small electromagnet | Schramme | EG1025A02/110 | Electromagnet 24V 220 N 150 N 2,5 W |
| Controller electromagnet | Conrad.ch | MST-1630.001 7 | Electromagnet control PCB board |
| Solenoid hub | Distrelec.ch | HD8286-R-F-24V100% | Kuhnke Solenid Hub 30 mm 16 N 2 N 16 W |
| Mini tweezers | Electron Microscopy Sciences | 0302-M5S-PS | Dumont Type 5 mini, super thin tips |
| Various optomechanical parts | Thorlabs | - | - |
| Various mechanical parts | Workshop Biozentrum, University Basel | - | Custom designed adapter plates, holders, etc. see attached CAD files |
| Optics | |||
| Laser diode | Thorlabs | CPS780S | 780 nm laser diode module 2.5 mW |
| Photo detector | Thorlabs | PDA100A-EC | Si Switchable Gain Detector, 320-1100 nm, 2.4 MHz BW, 100 mm2 |
| EM grids | |||
| DURASIN FILM TEM | emsdiasum.com | DTF-03523 | 30 nm DuraSiN™ silicon nitride membranes for TEM, pack of 5 |
| Quantifoil Cu 200 mesh R 2/1 | Quantifoil Micro Tools GmbH | ||
| Quantifoil Cu 200 mesh R 1.2/1.3 | Quantifoil Micro Tools GmbH | ||
| Buffers / Neg. stain | |||
| PBS | Sigma | D8537 SIGMA | Dulbecco’s Phosphate Buffered Saline |
| NanoVan 2% | nanoprobes.com | Negative stain vanadium based | |
| NanoW 2% | nanoprobes.com | Negative stain tungsten based | |
| Software | |||
| openBEB | The openBEB 2 framework is avaible upon request, version 3 will be downloadable | ||
| CryoWriter control software (openBEB plugin) | Avaible upon request, extra fees for 3rd party driver software might apply. |
Références
- Kuhlbrandt, W. Biochemistry: The resolution revolution. Science. 343 (6178), 1443-1444 (2014).
- Bai, X. -. c., McMullan, G., Scheres, S. H. W. How cryo-EM is revolutionizing structural biology. Trends Biochem Sci. 40 (1), 49-57 (2015).
- Dubochet, J., Adrian, M., Chang, J. J., Homo, J. C., Lepault, J., McDowall, A. W., Schultz, P. Cryo-electron microscopy of vitrified specimens. Q Rev Biophys. 21 (2), 129-228 (1988).
- Lepault, J., Booy, F. P., Dubochet, J. Electron microscopy of frozen biological suspensions. J Microsc. 129, 89-102 (1983).
- Baker, L. A., Rubinstein, J. L. Radiation damage in electron cryomicroscopy. Methods Enzymol. , (2010).
- Crewe, A. V., Eggenberger, D. N., Wall, J., Welter, L. M. Electron gun using a field emission source. Rev Sci Instrum. , (2003).
- Zemlin, F. Expected contribution of the field-emission gun to high-resolution transmission electron microscopy. Micron. 25 (3), 223-226 (1994).
- Scheres, S. H. RELION: implementation of a Bayesian approach to cryo-EM structure determination. J Struct Biol. 180 (3), 519-530 (2012).
- Grigorieff, N. FREALIGN: high-resolution refinement of single particle structures. J Struct Biol. 157 (1), 117-125 (2007).
- Li, X., Mooney, P., Zheng, S., Booth, C. R., Braunfeld, M. B., Gubbens, S., Agard, D. a., Cheng, Y. Electron counting and beam-induced motion correction enable near-atomic-resolution single-particle cryo-EM. Nat methods. 10, 584-590 (2013).
- Milazzo, A. -. C., Cheng, A., Moeller, A., Lyumkis, D., Jacovetty, E., Polukas, J., Ellisman, M. H., Xuong, N. -. H., Carragher, B., Potter, C. S. Initial evaluation of a direct detection device detector for single particle cryo-electron microscopy. J Struct Biol. 176 (3), 404-408 (2011).
- Ruskin, R. S., Yu, Z., Grigorieff, N. Quantitative characterization of electron detectors for transmission electron microscopy. J Struct Biol. 184 (3), 385-393 (2013).
- Veesler, D., Campbell, M. G., Cheng, A., Fu, C. -. y., Murez, Z., Johnson, J. E., Potter, C. S., Carragher, B. Maximizing the potential of electron cryomicroscopy data collected using direct detectors. J Struct Biol. 184 (2), 193-202 (2013).
- Campbell, M. G., Cheng, A., Brilot, A. F., Moeller, A., Lyumkis, D., Veesler, D., Pan, J., Harrison, S. C., Potter, C. S., Carragher, B., Grigorieff, N. Movies of ice-embedded particles enhance resolution in electron Cryo-microscopy. Structure. 20 (11), 1823-1828 (2012).
- Ripstein, Z. A., Rubinstein, J. L. Processing of Cryo-EM movie data. Methods in Enzymology. 579, 103-124 (2016).
- Li, X., Mooney, P., Zheng, S., Booth, C. R., Braunfeld, M. B., Gubbens, S., Agard, D. A., Cheng, Y. Electron counting and beam-induced motion correction enable near-atomic-resolution single-particle cryo-EM. Nature Methods. 10 (6), 584-590 (2013).
- McLeod, R. A., Kowal, J., Ringler, P., Stahlberg, H. Robust image alignment for cryogenic transmission electron microscopy. J Struct Biol. 197 (3), 279-293 (2017).
- Cheng, Y., Grigorieff, N., Penczek, P. A., Walz, T. A primer to single-particle cryo-electron microscopy. Cell. 161 (3), 438-449 (2015).
- Brenner, S., Horne, R. W. A negative staining method for high resolution electron microscopy of viruses. Biochim Biophys Acta. 34, 103-110 (1959).
- De Carlo, S., Harris, J. R. Negative staining and cryo-negative staining of macromolecules and viruses for TEM. Micron. 42 (2), 117-131 (2011).
- Glaeser, R. M. How good can cryo-EM become. Nat Methods. 13, 28-32 (2016).
- Engel, A., Graslund, A., Rigler, R., Widengren, J. in . Single Molecule Spectroscopy in Chemistry, Physics and Biology. 96, 417-431 (2010).
- Kemmerling, S., Ziegler, J., Schweighauser, G., Arnold, S. A., Giss, D., Müller, S. A., Ringler, P., Goldie, K. N., Goedecke, N., Hierlemann, A., Stahlberg, H., Engel, A., Braun, T. Connecting µ-fluidics to electron microscopy. J Struct Biol. 177 (1), 128-134 (2012).
- Arnold, S. A., Albiez, S., Opara, N., Chami, M., Schmidli, C., Bieri, A., Padeste, C., Stahlberg, H., Braun, T. Total sample conditioning and preparation of nanoliter volumes for electron microscopy. ACS nano. 10 (5), 4981-4988 (2016).
- Kemmerling, S., Arnold, S. A., Bircher, B. A., Sauter, N., Escobedo, C., Dernick, G., Hierlemann, A., Stahlberg, H., Braun, T. Single-cell lysis for visual analysis by electron microscopy. J Struct Biol. 183, 467-473 (2013).
- Arnold, S. A., Albiez, S., Bieri, A., Syntychaki, A., Adaixo, R., McLeod, R. A., Goldie, K. N., Stahlberg, H., Braun, T. Blotting-free and lossless cryo-electron microscopy grid preparation from nanoliter-sized protein samples and single-cell extracts. J Struct Biol. 197 (3), 220-226 (2017).
- Ramakrishnan, C., Bieri, A., Sauter, N., Roizard, S., Ringler, P., Müller, S. A., Goldie, K. N., Enimanev, K., Stahlberg, H., Rinn, B., Braun, T. openBEB: open biological experiment browser for correlative measurements. BMC Bioinformatics. 15, 84 (2014).
- Giss, D., Kemmerling, S., Dandey, V., Stahlberg, H., Braun, T. Exploring the interactome: microfluidic isolation of proteins and interacting partners for quantitative analysis by electron microscopy. Anal Chem. 86, 4680-4687 (2014).
Réimpressions et Autorisations
Demande d’autorisation pour utiliser le texte ou les figures de cet article JoVE
Demande d’autorisationThis article has been published
Video Coming Soon
À PROPOS DE JoVE
Copyright © 2025 MyJoVE Corporation. Tous droits réservés.