
Method Article
Méthodes de culture pour étudier les interactions spécifiques aux apicales à l’aide de modèles organoïdes intestinaux
Dans cet article
Résumé
Nous présentons ici deux protocoles qui permettent la modélisation des interactions apicales-spécifiques intestinales. Les monocouches intestinales organoïdes-dérivées et les cultures d’interface air-liquide (ALI) facilitent la génération d’épithélium bien différencié accessible des côtés luminal et basolatéral, tandis que les organoïdes intestinaux inversés à polarité exposent leur côté apical et se présentent à des analyses à haut débit.
Résumé
La muqueuse de l’épithélium intestinal est constituée d’une simple couche de cellules épithéliales spécialisées qui exposent leur côté apical à la lumière et répondent aux indices externes. L’optimisation récente des conditions de culture in vitro permet la recréation de la niche des cellules souches intestinales et le développement de systèmes de culture avancés en 3 dimensions (3D) qui récapitulent la composition cellulaire et l’organisation de l’épithélium. Les organoïdes intestinaux incorporés dans une matrice extracellulaire (ECM) peuvent être maintenus à long terme et s’auto-organiser pour générer un épithélium bien défini et polarisé qui englobe une lumière interne et un côté basal exposé externe. Cette nature restrictive des organoïdes intestinaux présente des défis dans l’accès à la surface apicale de l’épithélium in vitro et limite l’étude des mécanismes biologiques tels que l’absorption des nutriments et les interactions hôte-microbiote/hôte-pathogène. Ici, nous décrivons deux méthodes qui facilitent l’accès au côté apical de l’épithélium organoïde et soutenons la différenciation des types intestinaux spécifiques de cellules. Tout d’abord, nous montrons comment l’élimination de l’ECM induit une inversion de la polarité des cellules épithéliales et permet la génération d’organoïdes 3D apicaux. Deuxièmement, nous décrivons comment générer des monocouches à 2 dimensions (2D) à partir de suspensions unicellulaires dérivées d’organoïdes intestinaux, composées de types cellulaires matures et différenciés. Ces techniques fournissent de nouveaux outils pour étudier les interactions apicales spécifiques de l’épithélium avec des indices externes in vitro et promouvoir l’utilisation d’organoïdes comme plate-forme pour faciliter la médecine de précision.
Introduction
L’épithélium intestinal est le deuxième plus grand épithélium du corps humain et se compose d’une couche cellulaire polarisée qui facilite l’absorption des nutriments et agit comme une barrière contre les insultesenvironnementales 1. Cette distinction entre les côtés apical et basolatéral permet aux cellules de l’épithélium de remplir leurs diverses fonctions. Le compartiment apical est exposé à la lumière et négocie les interactions épithéliales avec les stimuli environnementaux et les micro-organismes, tout en facilitant l’absorption des nutriments. La surface basolatérale abrite des jonctions intercellulaires et des adhérences cellule-matrice, tout en s’interfançant avec des cellules du système immunitaire et d’autres tissus2. Ces jonctions génèrent une monocouche imperméable attachée à la membrane basale, qui agit comme une barrière et fournit les nutriments absorbés au tissu corporel environnant.
La mise en place de systèmes de culture capables de récapituler ces fonctions intestinales in vitro a été difficile3. Les modèles in vitro conventionnels utilisent des lignées cellulaires humaines transformées de cancer colorectal, telles que Caco-2, pour générer des cultures monocouches 2D. Bien qu’ils soient capables de modéliser de multiples fonctions du compartiment absorbant, ces modèles ne peuvent pas récapituler complètement la composition et la fonction intestinales de l’épithélium, ce qui limite les caractéristiques fonctionnelles clés et les applications4,5.
L’émergence d’organoïdes en tant que système de culture 3D avancé généré à partir de cellules souches qui peuvent s’auto-organiser et se différencier de types de cellules spécifiques à un organe, a été une percée dans l’étude in vitro de l’épithélium intestinal6. Les organoïdes intestinaux sont incorporés dans une matrice extracellulaire (ECM) qui ressemble à la lame basale et forment des jonctions cellule-matrice qui permettent à ces cultures de conserver la polarité apicobasale de l’épithélium. Les organoïdes présentent une architecture fermée dans laquelle la face apicale est exposée au compartiment luminal, imitant ainsi la structure de l’intestin. Bien que cette organisation fermée offre la possibilité d’étudier des fonctions spécifiques à l’orientation, elle limite les investigations qui nécessitent l’accès à la face apicale de l’épithélium. Différentes approches ont été adoptées pour surmonter ces limitations en 2D et 3D, y compris la fragmentation organoïde, la microinjection organoïde et la génération de cultures monocouches7. La fragmentation organoïde provoque la perte de l’organisation structurelle et la destruction des jonctions cellulaires, ce qui permet l’exposition de la surface apicale de l’épithélium au milieu. Cette technique tire parti de la capacité de régénération des fragments pour reformer les organoïdes lorsqu’ils sont ensemencés dans une matrice extracellulaire et a été utilisée pour modéliser les interactions entre les maladies infectieuses et l’hôte et l’agent pathogène8,9. Cependant, l’accès simultané à la surface apicale et basique peut également obtenir des réponses non spécifiques à l’infection.
Une approche alternative qui permet l’accès à la surface apicale et préserve à la fois l’architecture structurelle et les jonctions cellulaires est représentée par la microinjection de facteurs dans la lumière des organoïdes. Cette méthode a été largement utilisée pour étudier les interactions hôte-pathogène et modéliser les effets de Cryptosporidium10, H. pylori11et C. difficile12 sur l’épithélium gastro-intestinal in vitro. En utilisant des techniques similaires, le potentiel mutagène de la souche pks+ de E. coli sur l’épithélium intestinal a été déterminé13. Bien qu’efficace, la microinjection organoïde est une tâche laborieuse et inefficace compte tenu du nombre élevé d’organoïdes qui doivent être injectés pour obtenir des effets mesurables et limite donc son application pour les essais à haut débit.
Les progrès récents avec les organoïdes intestinaux ont également fourni des méthodes pour l’établissement de cultures organoïdes monocouches 2D, exposant ainsi leur surface apicale14,15,16,17. Ces monocouches dérivées d’organoïdes récapitulent les principales propriétés in vivo de l’épithélium intestinal. Ils présentent une composition cellulaire physiologiquement pertinente, contenant à la fois des populations de cellules différenciées et des populations de cellules souches et modélisent la diversité à travers l’axe crypte-villosité. Au fur et à mesure que la polarité apicobasale est conservée, les propriétés monocouches inhérentes permettent un accès facile aux côtés apical et basolatéral et les échanges de médias peuvent imiter l’écoulement intestinal et l’élimination des déchets, ce qui permet une culture à long terme. Ces caractéristiques rendent les monocouches dérivées d’organoïdes propices aux études axées sur les interactions luminales et fournissent un modèle supérieur pour l’intégrité et la perméabilité de la barrière épithéliale18,19.
Des études ont montré que la polarité des cellules épithéliales est étroitement régulée par les protéines ECM dans les sphéroïdes MDCK20,21 et récemment dans les organoïdes intestinaux humains22. L’élimination des composants ECM ou l’inhibition du récepteur de l’intégrine qui médie les jonctions cellule-matrice entraîne une inversion de polarité des organoïdes intestinaux et l’exposition de la face apicale de l’épithélium au milieu22. Cette approche a suscité l’intérêt des chercheurs travaillant sur les maladies infectieuses car elle permet un accès facile à la face apicale en 3D et la rend facile à des tests à haut débit. Ici, nous décrivons un protocole modifié basé sur les travaux récents du laboratoire Amieva22, qui facilite la génération d’organoïdes intestinaux 3D qui exposent facilement leur côté apical. Nous décrivons également un protocole qui peut générer efficacement et reproductiblement des monocouches 2D intestinales dérivées d’organoïdes intestinaux.
Protocole
Le dérivation des cultures organoïdes intestinales humaines a été réalisée comme décrit ailleurs23. Les organoïdes ont été maintenus en culture comme décrit dans la fiche d’information sur le produit (PIS) pour le milieu d’expansion organoïde intestinal (voir la table des matériaux).
1. Inversion de la polarité organoïde intestinale
REMARQUE: Cette section décrira le protocole pour inverser la polarité des organoïdes intestinaux 3D. Ce protocole fournit une procédure détaillée pour l’établissement d’organoïdes polarisés avec une surface apicale exposée en suspension dans une plaque de culture. Ce protocole est conçu comme un test final, bien que les organoïdes puissent être maintenus dans cette conformation pendant plus de 2 semaines et maintenir une petite population de cellules souches qui leur permet de rétablir les organoïdes apicaux lors du passage.
- Revêtement d’articles de culture et de tubes avec une solution anti-adhérente
REMARQUE: Pour maximiser le nombre d’organoïdes intestinaux apicaux et prévenir la fixation indésirable à l’article de culture, un pré-enrobage des articles de culture est généralement nécessaire. La section décrite ci-dessous décrit le revêtement de culture et de plastique avec une solution anti-adhérente.- Plaques de culture de revêtement à utiliser dans la partie suspension du protocole comme suit.
- Ajouter 0,5 mL de solution anti-adhérente à chaque puits d’une plaque de culture tissulaire de 24 puits.
- Faire pivoter la plaque pour répartir la solution uniformément sur la surface et les parois des puits.
- Centrifuger la plaque à 1 300 x g pendant 10 min.
- Retirer la solution anti-adhérente de chaque puits à l’aide d’un aspirateur ou d’une pipette de 1 mL.
- Laver les puits avec 1 mL de DMEM/F-12 avec 15mM HEPES (DMEM/F12).
- Remplissez bien chaque puits lavé avec 0,5 mL de DMEM/F-12 et conservez la plaque à 37 °C et 5 % deCO2 jusqu’à utilisation.
REMARQUE: Si elles ne sont pas utilisées immédiatement, les plaques revêtues peuvent être conservées dans l’incubateur pendant au moins 1 semaine.
- Enrobez les tubes coniques de 15 mL utilisés dans ce protocole comme suit.
- Ajouter 4 mL de solution anti-adhérente au tube conique de 15 mL.
- Faire pivoter le tube pour répartir la solution uniformément sur la surface des parois.
- Centrifuger le tube à 1 300 x g pendant 10 min.
- Retirez la solution anti-adhérente du tube.
- Laver le tube 1x avec 5 mL de DMEM/F-12 et aspirer le DMEM/F-12. Les tubes sont maintenant prêts à l’emploi.
REMARQUE: Si vous n’utilisez pas immédiatement, ajouter 5 ml de DMEM/F12 et conserver à 4 °C. Les tubes enduits peuvent être maintenus à 4 °C pendant au moins une semaine.
- Plaques de culture de revêtement à utiliser dans la partie suspension du protocole comme suit.
- Inversion de polarité des organoïdes intestinaux par culture en suspension
REMARQUE: Les étapes ci-dessous décrivent l’inversion d’organoïdes intestinaux précédemment cultivés dans des conditions de dôme ECM standard. Les procédures décrites ici sont pour un puits d’une plaque de 24 puits. Si vous utilisez un autre matériel de culture, ajustez les volumes en conséquence.- Retirez et jetez soigneusement le milieu de chaque puits contenant des organoïdes sans perturber le dôme de la matrice extracellulaire de la membrane sous-sol. Assurez-vous que la taille des organoïdes est de 150 à 250 μm de diamètre (généralement au jour 3-5) avant de commencer le protocole d’inversion.
- Ajouter 1 mL de solution de dissociation glacée dans chaque puits.
- Incuber à température ambiante (15-25 °C) pendant 1 min.
- Ajouter au moins 2 mL de solution anti-adhérente à un tube de 15 mL à utiliser pour le revêtement des pointes en plastique.
- Ajouter au moins 2 mL de DMEM/F-12 à un tube de 15 mL à utiliser pour laver les pointes en plastique rincées avec une solution anti-adhérente.
- Enduire une pointe de pipette de 1 mL avec une solution anti-adhérente, piper 1 mL de solution trois fois dans le tube avec une solution anti-adhérente de l’étape 1.2.4.
- Laver la pointe dans du DMEM/F-12 froid, en piper 1 mL de solution trois fois dans le tube avec du DMEM/F-12 de l’étape 1.2.5.
- À l’aide de la pointe enduite, délogez soigneusement les dômes en pipetant lentement. Veillez à ne pas perturber ou fragmenter les organoïdes.
- Transigeon de la suspension organoïde sur une plaque traitée avec une solution anti-adhérente (à partir de l’étape 1.1.1).
- Placer la plaque sur un agitateur à 4 °C pendant 30 min. Un gyro shaker peut être utilisé à 70 tr / min.
REMARQUE: Évitez les secousses sévères, telles que le vortex de l’échantillon, car cela peut entraîner une fragmentation organoïde. La formation de bulles et de mousse peut indiquer des secousses sévères. - Après 30 min, retirez la plaque. À l’aide d’une pointe de pipette de 1 mL recouverte d’une solution anti-adhérente, pipetter doucement la solution de haut en bas.
- Placer la plaque sur un agitateur à 4 °C pendant 15 min. Un gyro shaker peut être utilisé à 70 tr / min.
- Retirer la plaque et laisser les organoïdes se déposer par gravité (1-2 min à température ambiante). Observez la plaque au microscope pour vérifier la sédimentation organoïde.
REMARQUE: Évitez une agitation importante de la plaque car elle peut provoquer la remise en suspension des organoïdes réglés. - Une fois les organoïdes installés, retirer autant que possible la solution de dissociation et laver en ajoutant 1,5 mL de DMEM/F-12.
- Laisser les organoïdes sédimenter et éliminer autant de surnageant que possible. Répétez l’étape de lavage une fois de plus.
NOTA : Observer la plaque au microscope pour vérifier la sédimentation organoïde avant d’effectuer toute étape de lavage. - Retirez autant que possible le DMEM/F-12 et ajoutez 0,5 mL de milieu d’expansion organoïde intestinal. Incuber à 37 °C et 5 % deCO2.
- Le lendemain, effectuez un changement partiel de milieu en inclinant la plaque sur un angle de 25 à 30 degrés et en enlevant le milieu le long de la paroi du puits. Enlever 0,4 mL de milieu en prenant soin de ne pas enlever les organoïdes en suspension.
- Ajouter 0,4 mL de milieu d’expansion organoïde intestinale. Incuber à 37 °C et 5 % deCO2 pendant 3 jours.
REMARQUE: Après 3 jours, la plupart des organoïdes devraient avoir une polarité apicale, mais de grands agrégats peuvent s’être formés. - Si des granulats se sont formés, utiliser une pipette de 1 mL avec une pointe recouverte d’une solution anti-adhérente (comme décrit aux étapes 1.2.6 et 1.2.7) pour ciselé les granulats en pipent 20 fois de haut en bas tout en appuyant sur l’extrémité de la pointe dans le fond de la plaque.
- Placer la plaque à 37 °C et 5 % deCO2 et effectuer un changement complet du milieu le lendemain avec le milieu d’expansion organoïde intestinale (tel que décrit à la section 1.2.17).
- Après 2 jours (jour 5 en suspension), les organoïdes intestinaux apicaux peuvent être utilisés dans les essais en aval.
2. Établissement de cultures monocouches 2D de cellules intestinales et d’interface air-liquide (ALI) dérivées d’organoïdes intestinaux 3D
REMARQUE: Cette section décrira le protocole pour générer des cultures monocouches 2D à partir d’organoïdes intestinaux. Cette technique offre l’avantage d’établir une culture monocouche confluente et polarisée avec une surface apicale exposée dans une plaque de culture tissulaire contenant un insert de membrane de culture cellulaire. Bien que la monocouche commence à se différencier dans le format monocouche submergé, une différenciation supplémentaire de la monocouche peut être obtenue en passant à une culture ALI après qu’elle ait atteint la confluence. Les protocoles monocouche et ALI ultérieurs sont conçus comme des tests finaux et bien que la culture monocouche maintienne une petite population de cellules souches, ni l’un ni l’autre ne peut être efficacement divisé et passé après son établissement.
- Préparation de milieux et de plaques pour la culture monocouche intestinale
- Préparer le milieu de différenciation organoïde intestinal tel que décrit par le fabricant pour la culture monocouche (voir la liste des matériaux). Ajouter le Y-27632 uniquement à un volume de milieu qui sera utilisé dans un délai de 1 semaine à une concentration finale de 10 μM. S’il n’est pas utilisé immédiatement, conserver à 4 °C.
- Trypsine préchauffée EDTA (0,05 %) à 37 °C.
- Au moins 2 h avant l’ensemencement des cultures monocouches, préparer une solution de revêtement ECM à 2% comme suit.
- Décongeler l’ECM sur la glace et ajouter 1:50 au PBS froid et stérile pour préparer une solution à 2 %. Préparer des quantités suffisantes pour ajouter 100 μL au puits supérieur de chaque insert de membrane de culture cellulaire de 6,5 mm(0,33 cm 2)à utiliser. Ajustez le volume de manière appropriée pour les cultures plus grandes ou plus petites.
- Ajouter 100 μL à chaque puits et incuber chaque plaque à 37 °C et 5 % deCO2 jusqu’à ce qu’elle soit nécessaire (au moins 2 h avant l’ensemencement).
- Organoïdes intestinaux dissociants pour la génération et la culture de monocouches
- Retirer un nombre approprié de puits de culture organoïde de l’incubateur. Veuillez consulter le tableau 1 pour obtenir le nombre recommandé de puits à récolter pour divers articles de culture.
- Aspirer tout le milieu des cultures organoïdes sans perturber le dôme ECM.
- Ajouter 1 mL de solution de dissociation à chaque puits.
- Incuber à température ambiante (15-25 °C) pendant au moins 1 min.
- À l’aide d’une pipette de 1 mL, pipetter vigoureusement de haut en bas pour perturber le dôme et libérer les organoïdes.
- Regrouper les organoïdes suspendus de chaque puits dans un tube conique de 15 mL. Incuber à température ambiante pendant 10 min avec une agitation douce ou un balancement. Les organoïdes pour l’ensemencement de plusieurs puits peuvent être regroupés à cette étape, jusqu’à 10 mL de volume dans chaque tube.
- Centrifuger à 200 x g pendant 5 min à 4 °C.
- Retirez et jetez le surnageant. Ajouter 5 mL de DMEM/F-12 glacé aux organoïdes de remise en suspension.
- Mélanger et centrifuger à nouveau à 200 x g pendant 5 min à 4 °C.
- Aspirez le surnageant, en enlevant autant que possible, en faisant attention à ne pas déranger le granulé.
NOTA : Une partie de l’ECM résiduel peut rester dans la pastille; cependant, cela ne devrait pas affecter de manière significative la dissociation des organoïdes. - Ajouter 1 mL de trypsine-EDTA préchauffé (37 °C) (0,05 %) aux organoïdes de remise en suspension. Bien mélanger pour assurer une suspension uniforme. Ajoutez jusqu’à 1 mL supplémentaire de Trypsine-EDTA pour un grand nombre de cellules, ou s’il reste une quantité importante d’ECM.
- Incuber à 37 °C pendant 5-10 min.
- Bien mélanger avec une pipette de 1 mL pour perturber autant que possible les organoïdes. Les organoïdes doivent être complètement dissociés en cellules simples ou en petits fragments. S’il reste des fragments plus gros ou des organoïdes entiers après un pipetage approfondi, poursuivre l’incubation avec de la trypsine-EDTA à 37 °C pendant encore 3 à 5 minutes.
REMARQUE: Ne pas incuber avec la trypsine-EDTA pendant plus de 20 minutes, car cela peut entraîner une perte accrue de viabilité cellulaire. - Une fois que les organoïdes sont suffisamment dissociés, ajouter un volume égal de DMEM/F-12 (p. ex. 1 mL de DMEM/F-12 par mL de trypsine-EDTA) et pipeter de haut en bas pour bien mélanger. Désactivez trypsine-EDTA en ajoutant 10% fbs au DMEM/F-12 dans cette étape.
- Fragments de centrifugeuse à 200 x g pendant 5 min à 2-8 °C.
- Si les organoïdes dissociés ne parviennent pas à granuler, ceci est commun et peut être dû à une accumulation de mucus libéré par les cellules dissociées. Dans ce cas, mélangez soigneusement les cellules en les pipetant de haut en bas et centrifugez à nouveau à 200 x g pendant 5 min à 2-8 °C.
- Retirez soigneusement autant de surnageant que possible, en ne laissant que la pastille cellulaire.
- Ressusciter les cellules dans 100 μL de milieu de différenciation organoïde intestinale (avec 10 μM Y-27632) pour chaque puits à ensemencer. Ajustez le volume de manière appropriée pour les puits plus grands ou plus petits.
- Retirez les plaques revêtues de l’incubateur (préparées à l’étape 3.1.4) et retirez l’excès d’ECM de chaque puits.
- Ajouter 100 μL de la suspension cellulaire au puits supérieur de chaque insert de culture cellulaire.
- Ajouter 500 μL de milieu de différenciation organoïde intestinale au puits inférieur de chaque insert de culture cellulaire.
- Incuber à 37 °C et 5 % de CO2.
- Remplacez le milieu dans les puits supérieurs et inférieurs tous les 2 à 3 jours. Les cultures monocouches doivent atteindre la confluence dans les 7 jours et atteignent souvent la confluence dans les 2-3 jours.
- Établissement d’une culture d’interface air-liquide (ALI)
REMARQUE: Si vous le souhaitez, une différenciation supplémentaire d’une culture monocouche intestinale submergée peut être accomplie en faisant passer la culture monocouche submergée à une culture ALI. Cette méthode de culture permettra une augmentation du nombre de types cellulaires différenciés, en particulier les cellules de la lignée sécrétoire, telles que les cellules gobelines et entéréno-endocrines.- Établir une culture monocouche dans un insert de culture cellulaire comme décrit ci-dessus aux étapes 2.2.1-2.2.23 et maintenir cette culture à 100% de confluence pendant au moins 4 jours.
- Pour établir une culture ALI, retirez le milieu des puits supérieurs et inférieurs. Ajouter un milieu frais de différenciation organoïde intestinale (avec Y-27632) au puits inférieur, en laissant le puits supérieur vide.
- Incuber à 37 °C et 5 % deCO2.
- Remplacez le milieu dans le puits inférieur tous les 2-3 jours et laissez la monocouche se différencier pendant au moins 1 semaine.
- Rincez bien la partie supérieure avec du PBS stérile pour éliminer l’accumulation excessive de mucus, si nécessaire.
Dans ces conditions, la culture ALI peut être maintenue pendant au moins 2-3 semaines.
Résultats
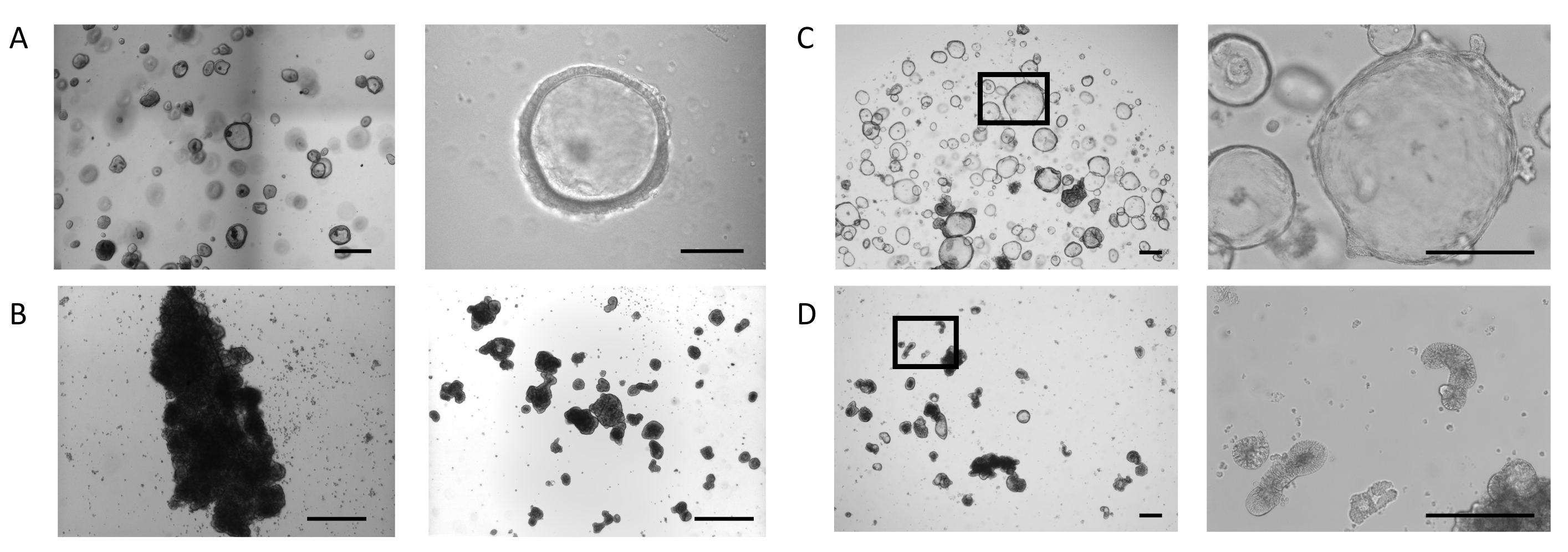
Les organoïdes ont été générés à partir d’échantillons de biopsie suivant le protocole décrit précédemment23 et dans le PIS pour intestinal Organoid Expansion Medium (voir le tableau des matériaux). La figure 1A,panneau gauche,montre le phénotype des organoïdes intestinaux cultivés dans un dôme avec un milieu d’expansion organoïde intestinal. Dans ces conditions de culture, les organoïdes présentent une morphologie kystique définie par un épithélium mince (10-25 μm) qui entoure une lumière(figure 1A,panneau droit). À ce stade, le côté apical de l’épithélium intestinal fait face à la lumière, tandis que le côté basolatéral entre en contact avec la matrice extracellulaire environnante. Lorsque la majorité des organoïdes ont atteint la taille désirée, la matrice extracellulaire a été enlevée, et les organoïdes ont ensuite été cultivés en suspension. La perte de liaison cellulaire à la matrice extracellulaire déclenche un processus d’inversion dans les organoïdes, entraînant une inversion de la polarité de l’épithélium organoïde, exposant la face apicale de l’épithélium au milieu de croissance et internalisant la face basolatérale.
Dans certaines cultures, les organoïdes en suspension s’agrègent et fusionnent, un effet qui est plus profond pendant les 3 premiers jours(figure 1B,panneau gauche). L’application d’une technique de cisaillement permet le détachement des organoïdes et la poursuite des cultures pendant des jours avec une réagrégation minimale(figure 1B,panneau droit).
Les organoïdes intestinaux cultivés dans des dômes ECM continuent de se dilater(figure 1C,panneau gauche)et présentent une formation spontanée de structures bourgeonnantes secondaires, ressemblant à de petites cryptes, sur le côté basolatéral de l’épithélium entourant la lumière(figure 1C,panneau droit). Au même moment, les organoïdes maintenus pendant 5 jours en l’absence de matrice extracellulaire continuent de se développer en suspension(figure 1D,panneau gauche). L’inversion de la polarité est caractérisée par l’épaississement (30-40 μm) de l’épithélium qui entoure le noyau des organoïdes et l’apparition d’une variété de morphologies : allongée(figure 1D,panneau droit et figure supplémentaire 1A),kystique(figuresupplémentaire 1B)et irrégulière(figuresupplémentaire 1C). Ceci est souvent combiné avec un rétrécissement de l’espace luminal dans l’organoïde, impactant leur taille globale.
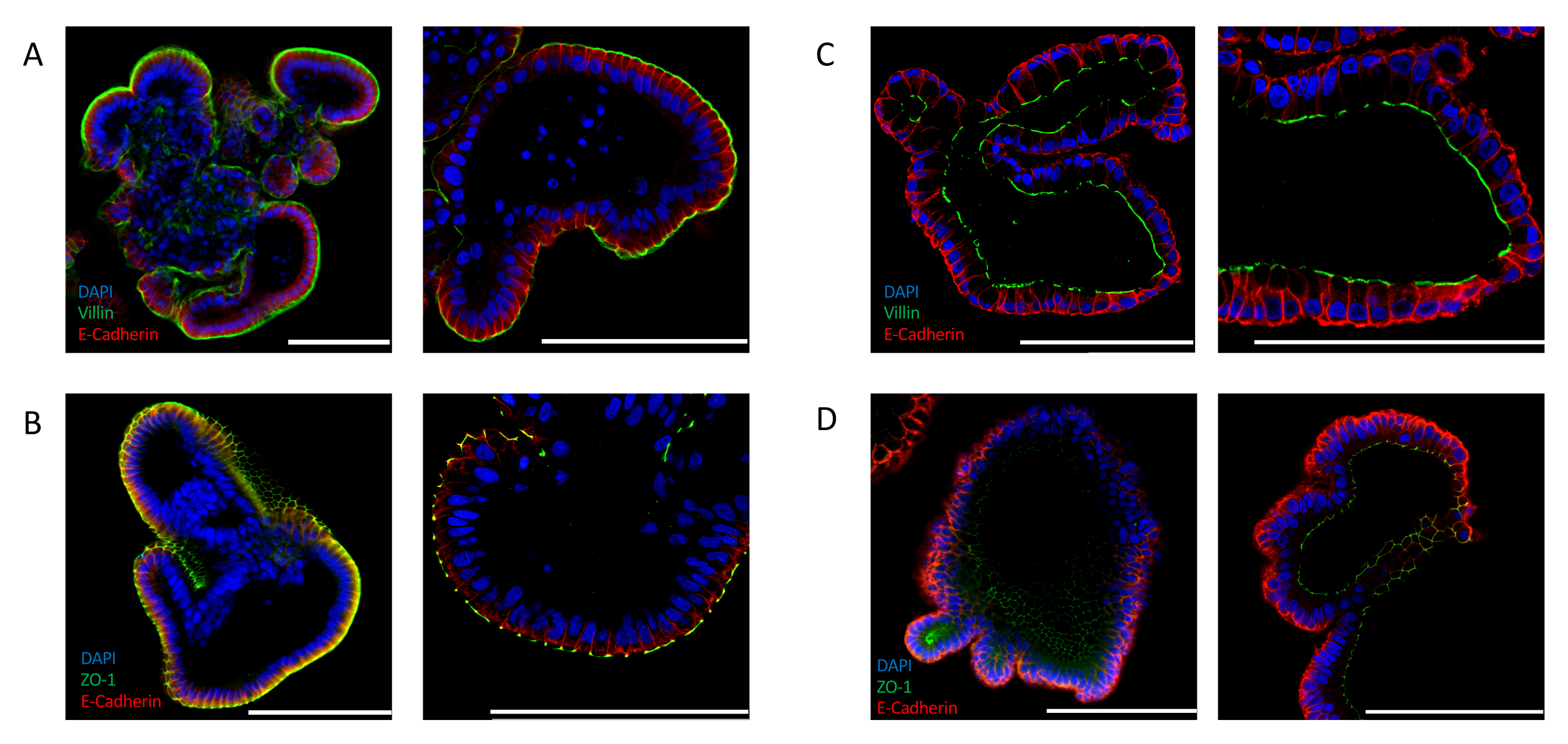
L’inversion efficace peut également être confirmée en analysant l’expression des marqueurs de polarité spécifiques à l’intestin. Les organoïdes intestinaux apicaux montrent une localisation distincte des noyaux vers la lumière de l’organoïde, comme indiqué par le signal 4′,6-diamidino-2-phényléine (DAPI). L’expression de marqueurs apicaux, tels que VILLIN(figure 2A)et ZO-1(figure 2B),est détectée sur la face externe de l’épithélium exposé au milieu. Cette localisation contraste fortement avec celle observée chez les organoïdes intestinaux cultivés dans l’ECM. Les organoïdes incorporés de matrice extracellulaire colorés pour les noyaux (DAPI), VILLIN(figure 2C)et ZO-1(figure 2D)démontrent une polarité apicobasale où la face apicale fait face à la lumière de l’organoïde.
L’élimination complète de l’ECM est nécessaire pour obtenir une inversion efficace de polarité des organoïdes intestinaux. Parfois, une partie des organoïdes trouvés dans des cultures en suspension entourées de résidus d’ECM, montre une morphologie kystique qui suggère une défaillance dans l’inversion de polarité de l’épithélium(figure supplémentaire 2A). L’analyse de la coloration immunofluorescente, effectuée sur ces organoïdes, fournit des preuves de la position basolatérale des noyaux (DAPI) le long de l’épithélium et de l’expression de ZO-1 sur le côté apical qui fait face à la lumière de l’organoïde(figure supplémentaire 2B)confirmant que l’élimination incomplète de l’ECM provoque la rétention de la polarité apicobasale d’une manière similaire aux organoïdes intégrés à l’ECM.
Le protocole pour l’établissement des cultures monocouches intestinales se traduit par une culture monocouche confluente dans les 7 jours suivant l’ensemencement et la culture atteindra souvent la confluence en aussi peu que 2-3 jours. L’un des principaux déterminants du succès est le nombre et la qualité des cellules utilisées pour ensemencer la monocouche. La figure 3A,panneau gauche, fournit un exemple d’une densité d’ensemencement idéale d’environ 150 000 cellules dans un insert de membrane de culture cellulaire de 6,5 mm. Ce nombre n’est pas fixe et peut être très variable en fonction du donneur et de la qualité de la culture organoïde source; par conséquent, le numéro de cellule doit être optimisé en fonction de ces variables. Si la densité d’ensemencement est trop faible ou de mauvaise qualité(figure 3A,panneau droit), il se peut qu’il n’y ait pas suffisamment d’accessoires pour former une culture monocouche confluente.
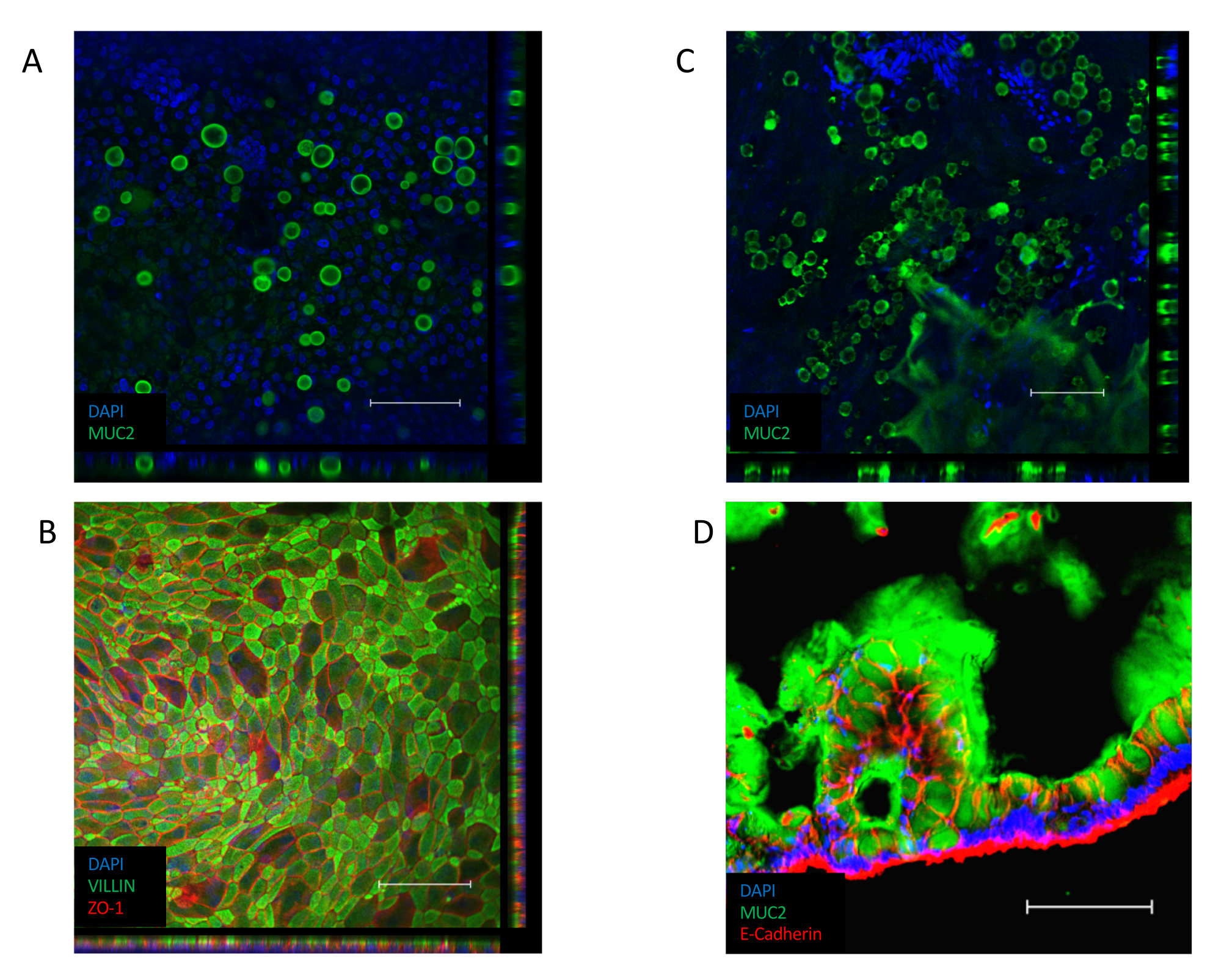
Une fois la monocouche établie(Figure 3B),les cellules forment des jonctions serrées, créant un aspect pavé(Figure 3B,Panneau gauche). S’ils ne parviennent pas à former une monocouche confluente(Figure 3B,Panneau de droite),l’apparence de la monocouche sera souvent « inégale », avec des régions de bonne qualité de fixation cellulaire, mais avec des espaces plus importants entre ces régions. Ces cultures ne fournissent pas de barrière fonctionnelle entre les compartiments basal et apical et ne conviennent pas aux essais décrits. Une monocouche confluente oriente sa bordure de brosse contenant VILLIN vers la face apicale de l’épithélium, avec son noyau positionné vers le pôle basolatéral de la cellule(figure 4B). Entre les cellules, des jonctions intercellulaires constituées de complexes multiprotéiques, dont ZO-1, se forment(figure 4B). Leur présence est essentielle pour assurer la fonction barrière de la culture épithéliale.
Une fois confluente, la transition vers une culture ALI induit une différenciation supplémentaire de la culture(figure 3C). De petites cellules rondes apparaissent et la monocouche elle-même prend un aspect plus plié. Bien que les cellules de gobelet soient présentes dans l’épithélium de la culture submergée(figure 4A),elles sont plus proéminentes après la différenciation d’ALI. Les cellules de gobelet présentes dans l’épithélium sécrètent du mucus, conduisant à un aspect brumeux sur le dessus de l’épithélium. Les cellules de gobelet et le mucus sécrété peuvent être visualisés par coloration pour la protéine de mucine sécrétée, MUC2(figure 4A,C et D)et l’augmentation de la population de cellules de gobelet peut être mesurée par une augmentation de l’expression de MUC2 (figure supplémentaire 3A). Il n’est pas nécessaire d’enlever cette couche de mucus semblable à un gel et elle collera à la surface de l’épithélium et restera après des lavages répétés. Si l’élimination est nécessaire, le lavage de la culture avec un composé mucolytique, tel que la N-acétyl cystéine 10 mM ou la TNT 50 μg/mL élimine l’excès de mucus. En plus de l’augmentation de la population de cellules de gobelet, l’interface ALI augmente également la présence de cellules entéroendocrines (comme indiqué par l’expression CHGA) (Figure supplémentaire 3B)et d’entérocytes matures (comme indiqué par l’expression KRT20) (Figure supplémentaire 3C).

Figure 1: Stades de la génération d’organoïdes intestinaux apicaux. (A) Images représentatives d’un dôme avec des organoïdes de la taille désirée au jour 4 (Panneau de gauche, Barre d’échelle = 500 μm). Les organoïdes sont à paroi mince, avec un compartiment luminal ouvert (panneau de droite, barre d’échelle = 100 μm). (B) Image représentative d’un puits avec une agrégation étendue après 3 jours en suspension (Panneau gauche, Barre d’échelle = 200 μm). Image de fragments de touffe directement après le cisaillement (panneau de droite, barre d’échelle = 200 μm). (C) Image représentative des organoïdes intestinaux en dôme au jour 7. Les organoïdes présentent une lumière élargie avec la formation de petits bourgeons sur le côté basolatéral de l’épithélium (grossissement gauche 20x, grossissement droit 100x de la région marquée, barre d’échelle = 200 μm). (D) Image représentative des organoïdes intestinaux après l’élimination de l’ECM et la culture en suspension ultérieure pendant 5 jours. Les organoïdes obtiennent une morphologie dense avec un épithélium épaissi et exposent leur face apicale au milieu. (Grossissement gauche 20x, grossissement 100x droit de la région marquée, barre d’échelle = 200 μm). Veuillez cliquer ici pour voir une version plus grande de cette figure.
{kind=link}

Figure 2: Coloration immunofluorescente pour les marqueurs de polarité cellulaire dans les organoïdes intestinaux. Apical-out(A, B),et apical-in (C, D) ont orienté des organoids intestinaux ont été souillés avec les marqueurs apicaux ZO-1 et VILLIN, et avec le marqueur épithélial E-CADHERIN (rouge). DAPI (bleu) a été utilisé pour visualiser les noyaux. Les panneaux de gauche affichent des images prises à un grossissement de 25x et les panneaux de droite affichent des images de différents organoïdes à un grossissement de 63x (seul le panneau C affiche un grossissement de 25x et 63x du même organoïde). (A) Les organoïdes intestinaux apicaux colorés avec VILLIN (vert) et E-CADHÉRINE (rouge) indiquent l’exposition de la face apicale au milieu. (B) Les organoïdes intestinaux apicaux colorés avec ZO-1 (vert) et E-CADHERIN (rouge) montrent la présence de jonctions serrées et la réversion de la polarité apicobasale. (C) Organoïde intestinal matrigel-incorporé coloré avec VILLIN (vert) et E-CADHERIN (rouge) montrant la face apicale face à la lumière organoïde. (D) Organoïdes intestinaux matrigel-incorporés colorés avec ZO-1 (vert) et E-CADHERIN (rouge) indiquant la présence de jonctions apicales serrées face à la lumière de l’organoïde. (Barre d’échelle = 100 μm). Les organoïdes ont été colorés par immunofluorescence et essionés à l’aide des protocoles précédemment publiés24,25. Veuillez cliquer ici pour voir une version plus grande de cette figure.
{kind=link}

Figure 3: Établissement de cultures monocouches intestinales. (A)Image représentative d’organoïdes 3D après traitement à 0,05% de Trypsine-EDTA. Les organoïdes sont dissociés en cellules individuelles ou en petites touffes cellulaires en vue de l’ensemencement de cultures monocouches. Panneau de gauche : exemple d’une densité d’ensemencement optimale pour une culture monocouche, environ 150 000 cellules par 100 μL sur un insert de membrane de culture cellulaire de 6,5 mm. Panneau de droite : exemple de densité d’ensemencement sous-optimale à <50 000 cellules par 100 μL sur un insert de membrane de culture cellulaire de 6,5 mm. (B) Image représentative d’une culture monocouche submergée. Panneau de gauche: couche 100% confluente avec l’aspect pavé caractéristique. Panneau de droite: environ 50% de monocouches confluentes. Les lacunes observées dans la monocouche (indiquées par une ligne pointillée) se ferment au fil du temps en raison de la prolifération continue des cellules souches intestinales. (C) Image représentative brightfield d’une culture ALI différenciée à 7 jours. (Barre d’échelle = 200 μm). Veuillez cliquer ici pour voir une version plus grande de cette figure.
{kind=link}

Figure 4: Coloration immunofluorescente pour marqueurs cellulaires différenciés dans les cultures monocouches. (A) Image de pile Z de la coloration immunofluorescente d’une culture monocouche immergée pour la protéine de mucine MUC2, indiquant la présence de cellules de gobelet dans la culture monocouche (vert = MUC2, bleu = DAPI). (B) image de Z-pile de la souillure immunofluorescente d’une monocouche immergée. La coloration VILLIN (vert) le long de l’extrémité apicale de l’épithélium indique la présence d’une bordure de brosse et la coloration ZO-1 (rouge) indique la présence de jonctions serrées entre les cellules (bleu = DAPI). (C) image de Z-pile de la souillure immunofluorescente d’une culture monocouche ALI-différenciée pour la protéine de mucine MUC2, indiquant la présence d’un nombre significativement plus grand de cellules de gobelet dans la culture monocouche d’ALI (vert = MUC2, bleu = DAPI). (D) Cryosection de la culture monocouche ali-différenciée, colorée pour la présence de MUC2 (vert) et e-CADHERIN (rouge) indiquant la présence de cellules gobelines dans l’épithélium et la sécrétion de mucus le long de la face apicale de la culture monocouche. (Barre d’échelle = 200 μm). Des cultures monocouches ont été colorées par immunofluorescence et essées à l’aide des protocoles précédemment publiés26,27. Veuillez cliquer ici pour voir une version plus grande de cette figure.
{kind=link}
Figure supplémentaire 1 : Spectre des phénotypes d’organoïdes intestinaux apicaux dans la suspension de culture. (A,B,C) Images représentatives additionnelles des morphologies intestinales d’organoïde maintenues en suspension pendant 5 jours après déplacement d’ECM. La polarité organoïde s’est inversée. Les organoïdes sont devenus plus denses avec un épithélium épaissi et la face apicale des organoïdes est tournée vers l’extérieur. Les organoïdes peuvent montrer une variété de morphologies: allongées(A),kystiques(B)et irrégulières(C). (Barre d’échelle = 100 μm). Veuillez cliquer ici pour télécharger ce fichier.
Figure supplémentaire 2 : Les organoïdes intestinaux ne parviennent pas à inverser la polarité en présence d’un milieu matriciel membranaire sous-sol résiduel dans les cultures en suspension. (A) Image représentative de l’élimination incomplète de l’ECM et de l’incapacité d’inverser la polarité des organoïdes. Des restes de Matrigel sont présents autour des organoïdes et contribuent au maintien de la polarité de l’épithélium orientée apicale-in. Les organoïdes montrent une morphologie kystique avec un épithélium mince entourant la lumière (barre d’échelle = 200 μm). (B) Image représentative d’un organoïde non inversé trouvé dans des conditions de culture en suspension. Les noyaux (bleu = DAPI) et E-CADHERIN (rouge) sont positionnés sur le côté basolatéral, ZO-1 (vert) est exprimé sur le côté apical qui fait face à la lumière de l’organoïde. (Barre d’échelle = 100 μm). Veuillez cliquer ici pour télécharger ce fichier.
Figure supplémentaire 3 : Expression génique de marqueurs cellulaires différenciés dans des cultures monocouches. (A,B,C) Expression de MUC2, CHGAet KRT20 dans une culture monocouche submergée et différenciée d’ALI générée dans un milieu de différenciation organoïde intestinal par rapport à une culture organoïde 3D cultivée avec un milieu d’expansion organoïde intestinal établi via qPCR. L’établissement d’une culture monocouche submergée augmente l’expression de chaque marqueur cellulaire différencié; cependant, la différenciation en tant que culture ALI augmente l’expression de chaque marqueur de manière exponentielle. Barres d’erreur = +/- SEM. Veuillez cliquer ici pour télécharger ce fichier.
| CULTUREWARE MONOCOUCHE | NOMBRE DE PUITS D’ORGANOÏDES INTESTINAUX À RÉCOLTER (à partir de 50 μL de dôme/par puits à ensemencer) |
| Insert transwell de 6,5 mm | 1 - 2 puits |
| Insert transwell de 12 mm | 3 - 4 puits |
| Plaque de 6 puits | 6 - 8 puits |
| Plaque de 24 puits | 3 - 4 puits |
| Plaque de 96 puits | 1 - 2 puits |
Tableau 1 : Nombre de puits d’organoïdes intestinaux à récolter pour divers articles de culture
Discussion
Les modèles organoïdes épithéliaux sont devenus de puissantes plateformes qui peuvent être utilisées pour modéliser l’organisation des tissus, la progression de la maladie et identifier les traitements23,28,29. La microinjection organoïde a ajouté de la valeur à la capacité des organoïdes de modéliser les maladies infectieuses car elle permet l’interaction pathogène avec la face apicale de l’épithélium hôte. Les progrès récents des techniques de microinjection ont optimisé la vitesse d’injection dans les organoïdes et ont atteint un taux allant jusqu’à 90 organoïdes injectés par heure. La fonction barrière dans les organoïdes injectés a été préservée, et la faible concentration en oxygène à l’intérieur de la lumière a permis la survie des bactéries injectées obligatoirement-anaérobies30. Cependant, des études ont noté la présence d’hétérogénéité dans les populations organoïdes au sein d’un même puits. Ces différences ont été observées dans la taille et la forme31,les niveaux d’expression des gènes clés32,ainsi que les taux de prolifération33. Des réponses différentielles au sein d’une même population organoïde à des composés tels que la forskoline et la PGE2, ou à la toxine du choléra, ont également été décrites28,33. Ces résultats soulignent la nécessité d’un nombre élevé d’organoïdes dans les études et limitent l’utilisation de l’injection luminale.
La culture organoïde conventionnelle est basée sur l’encapsulation et la propagation d’organoïdes dans un hydrogel. Cependant, les hydrogels peuvent poser des limites à la diffusion et introduire des gradients de concentration, ce qui peut augmenter l’hétérogénéité34. De plus, une grande variabilité a été documentée, non seulement entre les cultures et les donneurs, mais aussi dans des conditions expérimentales individuelles. La source donneuse, les propriétés biochimiques de l’hydrogel et l’hétérogénéité intrinsèque de l’organoïde en tant que système de culture sont des facteurs importants qui peuvent augmenter la variabilité expérimentale et limiter la reproductibilité des résultats obtenus dans les applications en aval. Les deux méthodes décrites ici fournissent un moyen simple d’exposer la face apicale de l’épithélium, permettant la modélisation de composés et d’agents pathogènes d’intérêt en les ajoutant directement au milieu de culture. La réduction de l’utilisation des hydrogels peut limiter la variabilité expérimentale provenant de sources techniques d’erreur.
Les organoïdes intestinaux apicaux conservent les caractéristiques clés du système de modèle organoïde et leur évolutivité les rend plus sensibles aux tests à haut débit, par rapport à la monocouche 2D. Cependant, comme les organoïdes conservent leur structure 3D, l’accessibilité du côté basal est limitée et peut entraver les études nécessitant un accès simultané aux deux côtés.
Nous avons démontré que l’inversion de polarité des organoïdes intestinaux repose sur l’élimination efficace et absolue de l’ECM, tout en préservant la structure intacte des organoïdes. L’utilisation de la solution de dissociation pour éliminer l’ECM et de la solution anti-adhérente pour empêcher l’adhésion des organoïdes à la vaisselle a contribué à améliorer l’efficacité globale du protocole publié par Co, J. Y. et ses collègues22,en particulier en ce qui concerne le nombre d’organoïdes apicaux produits pour des applications en aval.
En outre, nous avons observé que notre protocole prend en charge une inversion plus efficace des organoïdes inférieurs à 250 μm et que l’utilisation d’organoïdes plus gros peut entraîner une réduction de la production d’organoïdes, en raison de la fragmentation causée par le pipetage. Les pointes à alésage large, telles que celles indiquées dans la table des matériaux,peuvent permettre l’utilisation d’organoïdes plus gros. Cependant, les pointes à alésage large sont moins efficaces dans la dissociation ECM par rapport aux pointes standard en raison de la force mécanique appliquée plus faible. Par conséquent, il peut être nécessaire de répéter les étapes 1.2.9 à 1.2.11 pour perturber suffisamment et éliminer complètement tous les restes du MEC lorsque l’on travaille avec des organoïdes plus gros.
Les organoïdes en suspension peuvent survivre pendant au moins 2 semaines. Après cette période de temps, nous avons observé des changements de morphologie et un plus grand nombre de mort cellulaire. La présence de cellules proliférantes dans les organoïdesapicaux 22 permet le rétablissement de cultures organoïdes intestinales apicales. Ceci peut être réalisé en dissociant les organoïdes apicaux en cellules individuelles et en les intégrant dans l’ECM avec le milieu d’expansion organoïde intestinal.
Une limitation fréquemment rencontrée dans les protocoles décrivant l’établissement d’organoïdes intestinaux dans les cultures en suspension est la génération de grands agrégats. Cela affecte plusieurs variables telles que l’efficacité, la reproductibilité des caractéristiques morphologiques, la perméabilité aux composés et la signalisation paracrine. Semblable au protocole publié par Co, J. Y. et collègues, ici nous confirmons avoir obtenu l’inversion de polarité d’au moins 97% de tous les organoïdes suspendus sans cisaillement après 3 jours en suspension. Cependant, contrairement à la publication, nous avons introduit une étape de dissociation mécanique afin de réduire la formation de grands agrégats et d’augmenter le rendement. Puisque ce procédé peut endommager l’épithélium des organoïdes, nous avons prolongé la période d’incubation d’organoïdes pendant 2 jours supplémentaires pour permettre la récupération complète d’épithélium et assurer des cultures de haute qualité pour des applications en aval. L’introduction d’une agitation constante avec l’utilisation d’un agitateur d’incubateur ou d’une fiole de fileuse pourrait potentiellement réduire les événements de fusion, minimiser la fragmentation et augmenter l’oxygénation. Ces approches alternatives peuvent maintenir les cultures pendant de plus longues durées, réduire la mort cellulaire et permettre une différenciation plus poussée des organoïdes intestinaux apicaux.
L’établissement d’une monocouche 2D dérivée d’organoïdes offre plusieurs avantages et inconvénients par rapport aux organoïdes de polarité inversée. Le protocole décrit ici permet l’établissement rapide d’une culture monocouche confluente, typiquement, en moins de 7 jours et l’option de maintien à long terme des cultures pendant une période prolongée (jusqu’à 10 semaines). Le protocole et les milieux utilisés ici permettent également la différenciation efficace d’un nombre significatif de cellules, que l’on ne retrouve pas toujours dans d’autres cultures monocouches dérivées d’organoïdes16. L’établissement d’une monocouche sur une membrane d’insertion de culture cellulaire permet un accès simultané aux côtés apical et basolatéral de l’épithélium, ce qui les rend idéaux pour les études sur l’intégrité de la barrière et le transport épithélial. Cet accès simplifié les rend également plus susceptibles de faire l’objet d’études sur les infections et le traitement de la toxicomanie. En outre, ces cultures maintiennent bon nombre des caractéristiques propres au donneur, maintenant leur pertinence pour les études spécifiques au patient. La méthode de culture ALI facilite également la différenciation d’un épithélium plus fonctionnel composé de types de cellules sécrétoires et absorbives, ce qui le rend plus représentatif de l’épithélium intestinal humain. La stabilité relative de ces cultures permet également de les maintenir pendant une période prolongée, offrant la possibilité d’études à long terme. Cependant, les limites de cette approche sont le nombre élevé de cellules requises pour établir une monocouche confluente et la nécessité de maintenir une confluence complète pour avoir une séparation fonctionnelle entre les chambres apicales et basolatérales. L’architecture caractéristique de la crypte, qui peut être modélisée dans les cultures organoïdes 3D, est également perdue lors de l’établissement d’une culture monocouche. Néanmoins, le format expérimentalement convivial de la culture et la facilité d’accès aux côtés apicaux et basolatéraux de l’épithélium en font un outil puissant pour l’étude de la physiologie intestinale.
Déclarations de divulgation
G. S., W.C. et S. S. sont des employés de STEMCELL Technologies Ltd., Cambridge (Royaume-Uni). M. S., F. E., S. L., A. E. et R. K.C. sont des employés de STEMCELL Technologies Inc., Vancouver (Canada).
Remerciements
Cette recherche a été financée par la subvention Horizon 2020 OrganoVIR 812673 sur le projet Organoids for Virus Research - An innovative training-ITN programme.
matériels
| Name | Company | Catalog Number | Comments |
| Anti-Adherence Rinsing Solution | STEMCELL Technologies Inc. | 7010 | For coating cultureware. Referred as anti-adherent solution into the main text. |
| Conical tubes, 15 mL | STEMCELL Technologies Inc. | 38009 | |
| Corning Matrigel Matrix, Growth Factor Reduced (GFR), Phenol Red-Free | Corning | 356231 | Extracellular matrix (ECM) for maintenance and establishment of organoid lines. |
| Costar 6.5 mm or 12 mm Transwell inserts | STEMCELL Technologies Inc. | 38023/38024 | For 2D Monolayer culture. |
| Costar 24 Well Flat-Bottom, Tissue culture-treated plate | STEMCELL Technologies Inc. | 38017 | For maintenance and establishment of organoid lines. |
| D-PBS (Without Ca++ and Mg++) | STEMCELL Technologies Inc. | 37350 | For washing |
| Dimethyl sulfoxide (DMSO) | Millipore Sigma | D2650 | Reconstitution of small molecules |
| DMEM/F-12 with 15 mM HEPES | STEMCELL Technologies Inc. | 36254 | For washing |
| Gentle Cell Dissociation Reagent (GCDR) | STEMCELL Technologies Inc. | 7174 | For Matrigel removal. Referred as dissociation reagent into the main text. |
| IntestiCult Organoid Growth Medium (Human) | STEMCELL Technologies Inc. | 6010 | For expansion of organoid lines prior to differentiation. Referred as Intestinal Organoid Expansion Medium into the main text. |
| IntestiCult Organoid Differentiation Medium (Human) | STEMCELL Technologies Inc. | 100-0214 | For establishment of monolayers and 3D differentiation. Referred as Intestinal Organoid Differentiation Medium into the main text. |
| Trypsin-EDTA (0.05%) | STEMCELL Technologies Inc. | 7910 | For 2D Monolayer establishment. |
| Y-27632 | STEMCELL Technologies Inc. | 72302 | RHO/ROCK pathway inhibitor, Inhibits ROCK1 and ROCK2. Used for 2D monolayer establishment. |
| Wide bore tips | Corning | #TF-1005-WB-R-S | Organoids handling |
Références
- Gehart, H., Clevers, H. Tales from the crypt: new insights into intestinal stem cells. Nature Reviews Gastroenterology and Hepatology. 16 (1), 19-34 (2019).
- Schneeberger, K., Roth, S., Nieuwenhuis, E. E. S., Middendorp, S. Intestinal epithelial cell polarity defects in disease: lessons from microvillus inclusion disease. Disease Models & Mechanisms. 11 (2), (2018).
- Klunder, L. J., Faber, K. N., Dijkstra, G., Van Ijzendoorn, S. C. D. Mechanisms of cell polarity - Controlled epithelial homeostasis and immunity in the intestine. Cold Spring Harbor Perspectives in Biology. 9 (7), 0227888 (2017).
- DiMarco, R. L., Hunt, D. R., Dewi, R. E., Heilshorn, S. C. Improvement of paracellular transport in the Caco-2 drug screening model using protein-engineered substrates. Biomaterials. 129, 152-162 (2017).
- Sun, H., Chow, E. C., Liu, S., Du, Y., Pang, K. S. The Caco-2 cell monolayer: usefulness and limitations. Expert Opinion on Drug Metabolism & Toxicology. 4 (4), 395-411 (2008).
- Clevers, H. Modeling development and disease with organoids. Cell. 165 (7), 1586-1597 (2016).
- Dutta, D., Heo, I., Clevers, H. Disease modeling in stem cell-derived 3D organoid systems. Trends in Molecular Medicine. 23 (5), 393-410 (2017).
- Zhang, Y. -. G., Wu, S., Xia, Y., Sun, J. Salmonella-infected crypt-derived intestinal organoid culture system for host-bacterial interactions. Physiological Reports. 2 (9), 12147 (2014).
- Nigro, G., Rossi, R., Commere, P. -. H., Jay, P., Sansonetti, P. J. The cytosolic bacterial peptidoglycan sensor Nod2 affords stem cell protection and links microbes to gut epithelial regeneration. Cell Host & Microbe. 15 (6), 792-798 (2014).
- Heo, I., et al. Modelling Cryptosporidium infection in human small intestinal and lung organoids. Nature Microbiology. 3 (7), 814-823 (2018).
- Bartfeld, S., et al. In vitro expansion of human gastric epithelial stem cells and their responses to bacterial infection. Gastroenterology. 148 (1), 126-136 (2015).
- Leslie, J. L., et al. Persistence and toxin production by Clostridium difficile within human intestinal organoids result in disruption of epithelial paracellular barrier function. Infection and Immunity. 83 (1), 138-145 (2015).
- Pleguezuelos-Manzano, C., et al. Mutational signature in colorectal cancer caused by genotoxic pks(+) E. coli. Nature. 580 (7802), 269-273 (2020).
- Fernando, E. H., et al. A simple, cost-effective method for generating murine colonic 3D enteroids and 2D monolayers for studies of primary epithelial cell function. American journal of physiology. Gastrointestinal and Liver Physiology. 313 (5), 467-475 (2017).
- Moon, C., VanDussen, K. L., Miyoshi, H., Stappenbeck, T. S. Development of a primary mouse intestinal epithelial cell monolayer culture system to evaluate factors that modulate IgA transcytosis. Mucosal Immunology. 7 (4), 818-828 (2014).
- Wang, Y., et al. Long-term culture captures injury-repair cycles of colonic stem cells. Cell. 179 (5), 1144-1159 (2019).
- Noel, G., et al. A primary human macrophage-enteroid co-culture model to investigate mucosal gut physiology and host-pathogen interactions. Scientific Reports. 7, 45270 (2017).
- VanDussen, K. L., et al. Development of an enhanced human gastrointestinal epithelial culture system to facilitate patient-based assays. Gut. 64 (6), 911-920 (2015).
- Thorne, C. A., et al. Enteroid monolayers reveal an autonomous WNT and BMP circuit controlling intestinal epithelial growth and organization. Developmental Cell. 44 (5), 624-633 (2018).
- Wang, A. Z., Ojakian, G. K., Nelson, W. J. Steps in the morphogenesis of a polarized epithelium: I. Uncoupling the roles of cell-cell and cell-substratum contact in establishing plasma membrane polarity in multicellular epithelial (MDCK) cysts. Journal of Cell Science. 95 (1), 137-151 (1990).
- Wang, A. Z., Ojakian, G. K., Nelson, W. J. Steps in the morphogenesis of a polarized epithelium: II. Disassembly and assembly of plasma membrane domains during reversal of epithelial cell polarity in multicellular epithelial (MDCK) cysts. Journal of Cell Science. 95 (1), 153-165 (1990).
- Co, J. Y., et al. Controlling epithelial polarity: a human enteroid model for host-pathogen interactions. Cell Reports. 26 (9), 2509-2520 (2019).
- Sato, T., et al. Long-term expansion of epithelial organoids from human colon, adenoma, adenocarcinoma, and Barrett's epithelium. Gastroenterology. 141 (5), 1762-1772 (2011).
- Dodt, H. -. U., et al. Ultramicroscopy: three-dimensional visualization of neuronal networks in the whole mouse brain. Nature Methods. 4 (4), 331-336 (2007).
- Fujii, M., et al. Human intestinal organoids maintain self-renewal capacity and cellular diversity in niche-inspired culture condition. Cell Stem Cell. 23 (6), 787-793 (2018).
- Crowley, S. M., et al. Intestinal restriction of Salmonella Typhimurium requires caspase-1 and caspase-11 epithelial intrinsic inflammasomes. PLoS Pathogens. 16 (4), 1008498 (2020).
- Rees, W. D., et al. Enteroids derived from inflammatory bowel disease patients display dysregulated endoplasmic reticulum stress pathways, leading to differential inflammatory responses and dendritic cell maturation. Journal of Crohn's & Colitis. 14 (7), 948-961 (2020).
- Dekkers, J. F., et al. A functional CFTR assay using primary cystic fibrosis intestinal organoids. Nature Medicine. 19 (7), 939-945 (2013).
- Almeqdadi, M., Mana, M. D., Roper, J., Yilmaz, &. #. 2. 1. 4. ;. H. Gut organoids: mini-tissues in culture to study intestinal physiology and disease. American Journal of Physiology. Cell Physiology. 317 (3), 405-419 (2019).
- Williamson, I. A., et al. A high-throughput organoid microinjection platform to study gastrointestinal microbiota and luminal physiology. Cellular and Molecular Gastroenterology and Hepatology. 6 (3), 301-319 (2018).
- Kassis, T., Hernandez-Gordillo, V., Langer, R., Griffith, L. G. OrgaQuant: human intestinal organoid localization and quantification using deep convolutional neural networks. Scientific Reports. 9 (1), 12479 (2019).
- Gracz, A. D., et al. A high-throughput platform for stem cell niche co-cultures and downstream gene expression analysis. Nature Cell Biology. 17 (3), 340-349 (2015).
- Gunasekara, D. B., et al. Development of arrayed colonic organoids for screening of secretagogues associated with enterotoxins. Analytical Chemistry. 90 (3), 1941-1950 (2018).
- Shin, W., et al. Spatiotemporal gradient and instability of Wnt induce heterogeneous growth and differentiation of human intestinal organoids. iScience. 23 (8), 101372 (2020).
Réimpressions et Autorisations
Demande d’autorisation pour utiliser le texte ou les figures de cet article JoVE
Demande d’autorisationThis article has been published
Video Coming Soon
À PROPOS DE JoVE
Copyright © 2025 MyJoVE Corporation. Tous droits réservés.