
Method Article
Методы культивирования для изучения апикально-специфических взаимодействий с использованием кишечных органоидных моделей
В этой статье
Резюме
Здесь мы представляем два протокола, которые позволяют моделировать кишечные апикальные взаимодействия. Органоидные кишечные монослои и культуры воздушно-жидкого интерфейса (ALI) способствуют генерации хорошо дифференцированного эпителия, доступного как с просветной, так и с базолатеральной сторон, тогда как перевернутые по полярности кишечные органоиды обнажают свою апикальную сторону и поддаются высокопроизводимым анализам.
Аннотация
Слизистая оболочка эпителия кишечника состоит из простого слоя специализированных эпителиальных клеток, которые обнажают свою апикальную сторону просвета и реагируют на внешние сигналы. Недавняя оптимизация условий культивирования in vitro позволяет воссоздать нишу стволовых клеток кишечника и развить передовые 3-мерные (3D) системы культивирования, которые рекапитулируют клеточный состав и организацию эпителия. Кишечные органоиды, встроенные во внеклеточный матрикс (ECM), могут поддерживаться для долгосрочной и самоорганизации для создания четко определенного, поляризованного эпителия, который охватывает внутренний просвет и внешнюю открытую базальную сторону. Этот ограничительный характер кишечных органоидов создает проблемы в доступе к апикальной поверхности эпителия in vitro и ограничивает исследование биологических механизмов, таких как поглощение питательных веществ и взаимодействие хозяин-микробиота /хозяин-патоген. Здесь мы описываем два метода, которые облегчают доступ к апикальной стороне органоидного эпителия и поддерживают дифференцировку конкретных типов клеток кишечника. Во-первых, мы покажем, как удаление ECM индуцирует инверсию полярности эпителиальных клеток и позволяет генерацию апикальных 3D-органоидов. Во-вторых, мы описываем, как генерировать 2-мерные (2D) монослои из одноклеточных суспензий, полученных из кишечных органоидов, состоящих из зрелых и дифференцированных типов клеток. Эти методы предоставляют новые инструменты для изучения апикально-специфических взаимодействий эпителия с внешними сигналами in vitro и способствуют использованию органоидов в качестве платформы для облегчения точной медицины.
Введение
Эпителий кишечника является вторым по величине эпителием в организме человека и состоит из поляризованного клеточного слоя, который облегчает поглощение питательных веществ и действует как барьер против экологических оскорблений1. Это различие между апикальной и базолатеральной сторонами позволяет клеткам эпителия выполнять свои разнообразные функции. Апикальный компартмент подвергается воздействию просвета и опосредует эпителиальные взаимодействия со стимулами окружающей среды и микроорганизмами, а также облегчает поглощение питательных веществ. Базолатеральная поверхность содержит межклеточные соединения и клеточно-матричные спайки, при этом сопрягаясь с клетками иммунной системы и другими тканями2. Эти соединения генерируют непроницаемый монослой, прикрепленный к базальной мембране, который действует как барьер и доставляет поглощенные питательные вещества в окружающие ткани организма.
Создание систем культивирования, способных рекапитулировать эти кишечные функции in vitro, было сложной задачей3. Обычные модели in vitro используют трансформированные линии клеток колоректального рака человека, такие как Caco-2, для создания 2D-монослойных культур. Несмотря на способность моделировать множественные функции абсорбционного компартмента, эти модели не могут полностью резюмировать состав и функцию кишечного эпителия, которые ограничивают ключевые функциональные характеристики и приложения4,5.
Появление органоидов как передовой системы 3D-культур, генерируемой из стволовых клеток, которые могут самоорганизовываться и дифференцироваться в орган-специфические типы клеток, стало прорывом в исследовании эпителия кишечника in vitro6. Кишечные органоиды встроены во внеклеточный матрикс (ECM), который напоминает базальную пластинку, и образуют клеточно-матричные соединения, которые позволяют этим культурам сохранять апикобазальную полярность эпителия. Органоиды демонстрируют замкнутую архитектуру, в которой апикальная сторона подвергается воздействию просветного отсека, тем самым имитируя структуру кишечника. Хотя эта закрытая организация предлагает возможность изучать функции, специфичные для ориентации, она ограничивает исследования, требующие доступа к апикальной стороне эпителия. Для преодоления этих ограничений как в 2D, так и в 3D были приняты различные подходы, включая фрагментацию органоидов, микроинъекцию органоидов и генерацию монослойных культур7. Фрагментация органоидов вызывает потерю структурной организации и разрушение клеточных соединений, что позволяет облевать апикальную поверхность эпителия в среду. Этот метод использует регенеративную способность фрагментов для реформирования органоидов при посеве во внеклеточный матрикс и был использован для моделирования инфекционных заболеваний и взаимодействий хозяин-патоген8,9. Однако одновременный доступ как к апикальной, так и к базальной поверхности может также вызывать неспецифические реакции на инфекцию.
Альтернативный подход, обеспечивающий доступ к апикальной поверхности и сохраняющего как структурную архитектуру, так и клеточные соединения, представлен микроинъекцией факторов в просвет органоидов. Этот метод широко использовался для изучения взаимодействий хозяина и патогена и моделирования эффектов Cryptosporidium10, H. pylori11и C. difficile12 на желудочно-кишечный эпителий in vitro. С помощью аналогичных методик был определен мутагенный потенциал pks+ штамма E. coli на кишечном эпителии13. Несмотря на эффективность, органоидная микроинъекция является трудоемкой и неэффективной задачей, учитывая большое количество органоидов, которые необходимо вводить для получения измеримых эффектов, и, следовательно, ограничивает ее применение для высокопроизводительных анализов.
Последние достижения с кишечными органоидами также предоставили методы для создания 2D монослойных органоидных культур, тем самым обнажая их апикальную поверхность14,15,16,17. Эти органоидные монослои повторяют ключевые свойства эпителия кишечника in vivo. Они демонстрируют физиологически значимый клеточный состав, содержащий как дифференцированные, так и стволовые популяции клеток и моделируют разнообразие по оси крипта-ворсо. Поскольку апикобазальная полярность сохраняется, присущие монослойные свойства обеспечивают легкий доступ как к апикальной, так и к базолатеральной сторонам, а обмен сред может имитировать кишечный поток и удаление отходов, что позволяет проводить долгосрочную культуру. Эти особенности делают органоидные монослои поддаются исследованиям, ориентированным на просветные взаимодействия, и обеспечивают превосходную модель целостности и проницаемости эпителиального барьера18,19.
Исследования показали, что полярность эпителиальных клеток жестко регулируется белками ECM в сфероидах MDCK20, 21 и в последнее время в органоидах кишечника человека22. Удаление компонентов ECM или ингибирование рецептора интегрина, опосредует клеточно-матричные соединения, приводит к изменению полярности кишечных органоидов и воздействию апикальной стороны эпителия на среду22. Этот подход привлек интерес исследователей, работающих над инфекционными заболеваниями, поскольку он обеспечивает легкий доступ к апикальной стороне в 3D и делает ее поддаются высокопроизводительным анализам. Здесь мы описываем модифицированный протокол, основанный на недавней работе лаборатории Amieva22,который облегчает генерацию 3D-органоидов кишечника, которые легко обнажают их апикальную сторону. Мы также описываем протокол, который может эффективно и воспроизводимо генерировать кишечные 2D-монослои, полученные из кишечных органоидов.
протокол
Выведение органоидных культур кишечника человека было выполнено, как описано в другом месте23. Органоиды поддерживали в культуре, как описано в Информационном листе продукта (PIS) для среды расширения органоидов кишечника (см. Таблицу материалов).
1. Инверсия полярности органоидов кишечника
ПРИМЕЧАНИЕ: В этом разделе будет описан протокол инвертирования полярности 3D-органоидов кишечника. Этот протокол обеспечивает подробную процедуру установления поляризованных органоидов с открытой апикальной поверхностью во суспензии в культуральной пластине. Этот протокол предназначен для анализа конечной точки, хотя органоиды могут поддерживаться в этой конформации более 2 недель и поддерживать небольшую популяцию стволовых клеток, которая позволяет им восстановить апикальные органоиды при прохождении.
- Покрытие культуры и туб антиасцепильным раствором
ПРИМЕЧАНИЕ: Чтобы максимизировать количество апикальных органоидов кишечника и предотвратить нежелательное прикрепление к культуральной посуде, обычно требуется предварительное пометание культуральной посуды. В разделе, описанном ниже, описано покрытие культуры и пластичной посуды антиприлипанным раствором.- Покрытие культурального покрытия пластин, которые должны использоваться в суспензионной части протокола следующим образом.
- Добавьте 0,5 мл антиагезивного раствора в каждую скважину 24-скважинной тканевой культуры.
- Закрутите пластину, чтобы равномерно распределить раствор по поверхности и стенкам колодцев.
- Центрифугировать пластину при 1 300 х г в течение 10 мин.
- Удалите антиадгезивный раствор из каждой скважины с помощью аспиратора или пипетки 1 мл.
- Промывайте скважины 1 мл DMEM/F-12 с 15mM HEPES (DMEM/F12).
- Наполните каждую промытую скважину 0,5 мл DMEM/F-12 и храните пластину при 37 °C и 5% CO2 до использования.
ПРИМЕЧАНИЕ: Если не использовать сразу, пластины с покрытием можно хранить в инкубаторе не менее 1 недели.
- Покрыть 15 мл коническими трубками, используемыми в этом протоколе, следующим образом.
- Добавьте 4 мл антиагезивного раствора в коническую трубку 15 мл.
- Закрутите трубку, чтобы равномерно распределить раствор по поверхности стенок.
- Центрифугировать трубку при 1 300 х г в течение 10 мин.
- Извлеките антиприцепной раствор из пробирки.
- Промыть трубку 1x с 5 мл DMEM/F-12 и аспирировать DMEM/F-12. Теперь трубки готовы к использованию.
ПРИМЕЧАНИЕ: Если не используется немедленно, добавьте 5 мл DMEM/F12 и храните при 4 °C. Трубки с покрытием можно хранить при 4 °C не менее недели.
- Покрытие культурального покрытия пластин, которые должны использоваться в суспензионной части протокола следующим образом.
- Инверсия полярности органоидов кишечника культурой суспензии
ПРИМЕЧАНИЕ: На следующих шагах описывается инверсия кишечных органоидов, ранее культивируемых в стандартных купольных условиях ECM. Процедуры, описанные здесь, предназначены для одной скважины из 24-скважинной пластины. При использовании других культур соответствующим образом отрегулируйте объемы.- Осторожно удалите и выбросьте среду из каждой скважины, содержащей органоиды, не нарушая базальную мембрану купола внеклеточного матрикса. Убедитесь, что размер органоидов составляет 150-250 мкм диаметр (обычно на 3-5 день) до начала протокола инверсии.
- Добавьте в каждую скважину по 1 мл ледяного диссоциационного раствора.
- Инкубировать при комнатной температуре (15-25 °C) в течение 1 мин.
- Добавьте не менее 2 мл антиприлипательного раствора в трубку объемом 15 мл, которая будет использоваться для покрытия пластиковых наконечников.
- Добавьте не менее 2 мл DMEM/F-12 в трубку объемом 15 мл, которая будет использоваться для мытья пластиковых наконечников, промытых антиадгезивным раствором.
- Покрыть наконечник пипетки 1 мл антиприцепным раствором, трижды пипетировку 1 мл раствора в пробирку антиприцепным раствором на этапе 1.2.4.
- Промыть наконечник в холодном DMEM/F-12, трижды пипетировку 1 мл раствора в трубку с DMEM/F-12 с шага 1.2.5.
- Используя покрытый наконечник, осторожно вытесняйте купола, медленно пипетируя. Позаботьтесь о том, чтобы не нарушить и не фрагментировать органоиды.
- Переложить органоидную суспензию на пластину, обработанную анти адгезивным раствором (из этапа 1.1.1).
- Поместите тарелку на шейкер при 4 °C в течение 30 мин. Гиротр шейкер можно использовать при 70 об/мин.
ПРИМЕЧАНИЕ: Избегайте резкого встряхивания, такого как вихрь образца, так как это может привести к фрагментации органоидов. Образование пузырьков и пены может свидетельствовать о резком встряхивании. - Через 30 мин снимите пластину. Используя наконечник пипетки 1 мл, покрытый антиприлипаемым раствором, аккуратно пипеткой повышали раствор вверх и вниз.
- Поместите тарелку на шейкер при 4 °C в течение 15 мин. Гиротр шейкер можно использовать при 70 об/мин.
- Снимите пластину и дайте органоидам отстоять под действием силы тяжести (1-2 мин при комнатной температуре). Понаблюдайте за пластиной под микроскопом, чтобы проверить органоидное осаждение.
ПРИМЕЧАНИЕ: Избегайте обширного перемешивания пластины, так как это может привести к повторному суспендированию оседлых органоидов. - После того, как органоиды осядет, удалите как можно больше диссоциационного раствора и промыть, добавив 1,5 мл DMEM / F-12.
- Позвольте органоидам отстойнику и удалите как можно больше супернатанта. Повторите шаг стирки еще раз.
ПРИМЕЧАНИЕ: Понаблюдайте за пластиной под микроскопом, чтобы проверить органоидное осаждение перед выполнением любого этапа промывки. - Удалите как можно больше DMEM/F-12 и добавьте 0,5 мл кишечной органоидной расширительной среды. Инкубировать при 37 °C и 5% CO2.
- На следующий день выполните частичную смену среды, наклонив плиту под углом от 25 до 30 градусов и удалив среду вдоль стенки скважины. Удалите 0,4 мл среды, заботясь о том, чтобы не удалить взвешенные органоиды.
- Добавьте 0,4 мл кишечной органоидной расширительной среды. Инкубировать при 37 °C и 5% CO2 в течение 3 дней.
ПРИМЕЧАНИЕ: Через 3 дня большинство органоидов должны иметь апикальную полярность, но могут образоваться большие агрегаты. - Если заполнители образовались, используйте пипетку 1 мл с наконечником, покрытым антиприцепляющим раствором (как описано в шагах 1.2.6 и 1.2.7), чтобы скорбеть агрегаты путем пипетки вверх и вниз 20 раз, при нажатии конца наконечника на нижнюю часть пластины.
- Поместите пластину при 37 °C и 5% CO2 и выполните полную смену среды на следующий день с помощью среды расширения органоидов кишечника (как описано в разделе 1.2.17).
- Через 2 дня (5-й день в суспензии) апикальные кишечные органоиды могут быть использованы в последующих анализах.
2. Создание культур 2D-монослоя клеток кишечника и воздушно-жидкостного интерфейса (ALI), полученных из 3D-органоидов кишечника
ПРИМЕЧАНИЕ: В этом разделе будет описан протокол генерации 2D-монослойных культур из кишечных органоидов. Этот метод обеспечивает преимущество создания сливающейся, поляризованной однослойной культуры с открытой апикальной поверхностью в тканевой культуральной пластине, содержащей вставку мембраны клеточной культуры. Хотя монослой начнет дифференцироваться в погруженном, монослойном формате, дополнительная дифференциация монослоя может быть достигнута путем переключения на культуру ALI после того, как она достигнет слияния. Как монослойный, так и последующие протоколы ALI предназначены для анализа конечной точки, и хотя монослойная культура поддерживает небольшую популяцию стволовых клеток, ни один из них не может быть эффективно разделен и пройден после ее создания.
- Подготовка сред и пластин для кишечных однослойных культур
- Подготовьте кишечную органоидную дифференцировочную среду, как указано производителем, для однослойной культуры (см. Список материалов). Добавляйте Y-27632 только к объему среды, которая будет использоваться в течение 1 недели при конечной концентрации 10 мкМ. Если не используется немедленно, хранить при 4 °C.
- Предварительно подогрет трипсин ЭДТА (0,05%) до 37 °C.
- По меньшей мере за 2 ч до посева однослойных культур готовят 2% раствор для покрытия ECM следующим образом.
- Разморозьте ECM на льду и добавьте 1:50 в холодный, стерильный PBS для приготовления 2% раствора. Подготовьте достаточное количество, чтобы добавить 100 мкл в верхнюю скважину каждой 6,5 мм (0,33см2)клеточной культуры мембранной вставки для использования. Отрегулируйте громкость соответствующим образом для больших или меньших культур.
- Добавьте 100 мкл в каждую скважину и инкубирует каждую пластину при 37 °C и 5% CO2 до тех пор, пока это не понадобится (по крайней мере, за 2 ч до посева).
- Диссоциация кишечных органоидов для генерации и культивирования монослоев
- Извлеките из инкубатора соответствующее количество колодцев органоидной культуры. Рекомендуемое количество скважин для сбора урожая различной посуды приведено в таблице 1.
- Аспирировать всю среду из органоидных культур, не нарушая купол ECM.
- Добавьте 1 мл диссоциационного раствора в каждую скважину.
- Инкубировать при комнатной температуре (15-25 °С) не менее 1 мин.
- Используя пипетку 1 мл, энергично пипетку вверх и вниз, чтобы разрушить купол и освободить органоиды.
- Смешанные органоиды из каждой скважины в коническую трубку 15 мл. Инкубировать при комнатной температуре в течение 10 мин с мягким перемешиванием или раскачивать. Органоиды для посева нескольких скважин могут быть объединены на этом этапе, объемом до 10 мл в каждой трубе.
- Центрифуга при 200 х г в течение 5 мин при 4 °C.
- Удалите и выбросьте супернатант. Добавьте 5 мл холодного льда DMEM/F-12 к ресуспендированию органоидов.
- Снова перемешать и центрифугировать при 200 х г в течение 5 мин при 4 °C.
- Аспирировать супернатант, удаляя как можно больше, стараясь не потревожить гранулу.
ПРИМЕЧАНИЕ: Некоторое остаточное ECM может оставаться в гранулах; однако это не должно существенно влиять на диссоциацию органоидов. - Добавьте 1 мл предварительно подогретого (37 °C) трипсина-ЭДТА (0,05%) для повторного суспендировании органоидов. Тщательно перемешайте, чтобы обеспечить равномерное суспензию. Добавьте до дополнительного 1 мл трипсина-ЭДТА для большого количества клеток, или если остается значительное количество ECM.
- Инкубировать при 37 °C в течение 5-10 мин.
- Тщательно перемешайте с пипеткой 1 мл, чтобы максимально разрушить органоиды. Органоиды должны быть полностью диссоциированы на отдельные клетки или небольшие фрагменты. Если после тщательной пипетки остаются более крупные фрагменты или целые органоиды, продолжают инкубацию с трипсином-ЭДТА при 37 °C еще 3-5 мин.
ПРИМЕЧАНИЕ: Не инкубировать с трипсином-ЭДТА более 20 мин, так как это может привести к повышенной потере жизнеспособности клеток. - Как только органоиды будут достаточно диссоциированы, добавьте равный объем DMEM / F-12 (например, 1 мл DMEM / F-12 на мл трипсина-ЭДТА) и пипетку вверх и вниз, чтобы тщательно перемешать. Инактивировать трипсин-ЭДТА, добавив 10% FBS к DMEM/F-12 на этом этапе.
- Фрагменты центрифуги по 200 х г в течение 5 мин при 2-8 °С.
- Если диссоциированные органоиды не могут гранулироваться, это распространено и может быть связано с накоплением слизи, высвобождаемой диссоциированными клетками. В этом случае тщательно перемешайте ячейки путем пипетирования вверх и вниз и центрифугируют их снова при 200 х г в течение 5 мин при 2-8 °C.
- Осторожно удалите как можно больше супернатанта, оставив только ячейку гранулы.
- Повторное суспендировать клетки в 100 мкл кишечной органоидной дифференцировки (с 10 мкМ Y-27632) для каждой скважины, которая будет засеяна. Отрегулируйте объем соответствующим образом для больших или меньших размеров скважины.
- Извлеките покрытые пластины из инкубатора (подготовленные на этапе 3.1.4) и удалите избыток ECM из каждой скважины.
- Добавьте 100 мкл клеточной суспензии в верхний колодец каждой вставки клеточной культуры.
- Добавьте 500 мкл кишечной органоидной дифференцировочной среды к нижнему колодецу каждой клеточной культуры.
- Инкубировать при 37 °C и 5% CO2.
- Заменяйте среду как в верхней, так и в нижней скважинах каждые 2-3 дня. Однослойные культуры должны достичь слияния в течение 7 дней и часто достигают слияния в течение 2-3 дней.
- Создание культуры взаимодействия воздух-жидкость (ALI)
ПРИМЕЧАНИЕ: При желании дальнейшая дифференциация погружной кишечной монослойной культуры может быть выполнена путем перехода погружной монослойной культуры в культуру ALI. Этот метод культивирования позволит увеличить количество дифференцированных типов клеток, особенно клеток секреторной линии, таких как бокало и энтероэндокринные клетки.- Устанавливают однослойную культуру в клеточной культуре, как описано выше на этапах 2.2.1-2.2.23, и поддерживают эту культуру при 100% слиянии в течение не менее 4 дней.
- Чтобы создать культуру АЛИ, удалите среду из верхней и нижней скважин. Добавьте свежую кишечную органоидную дифференцировку среды (с Y-27632) в нижний колодец, оставив верхний колодец пустым.
- Инкубировать при 37 °C и 5% CO2.
- Заменяйте среду в нижней колодце каждые 2-3 дня и дайте монослою дифференцироваться не менее 1 недели.
- Хорошо промойте верхнюю часть стерильным PBS, чтобы удалить избыточное накопление слизи, если это необходимо.
В этих условиях культура АЛИ может сохраняться не менее 2-3 недель.
Результаты
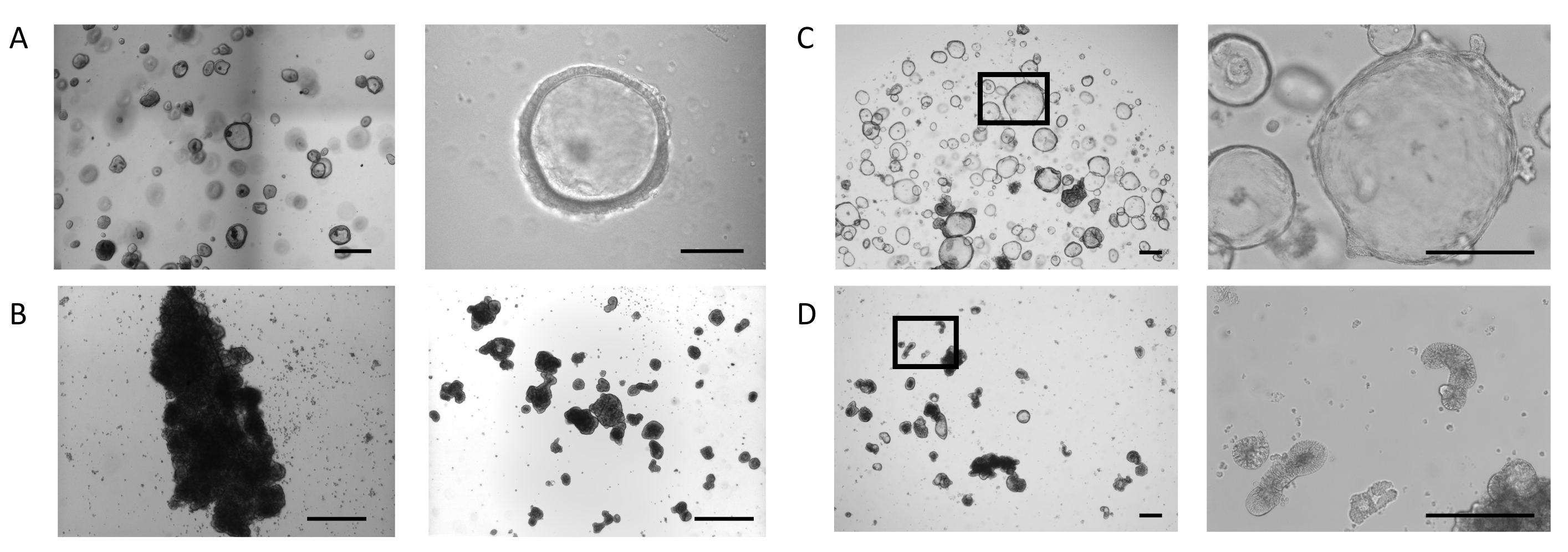
Органоиды были получены из образцов биопсии в соответствии с протоколом, описаннымранее 23, и в PIS для кишечной органоидной расширительной среды (см. Таблицу материалов). На рисунке 1А,левая панель,показан фенотип кишечных органоидов, культивированных в куполе со средой расширения кишечных органоидов. В этих условиях культивирования органоиды демонстрируют кистозную морфологию, определяемую тонким (10-25 мкм) эпителием, который заключает в себе просвет(рисунок 1А,правая панель). На этой стадии апикальная сторона кишечного эпителия обращена к просвету, в то время как базолатеральная сторона контактирует с окружающим внеклеточным матриксом. Когда большинство органоидов достигали желаемого размера, внеклеточный матрикс удаляли, а органоиды затем культивировали во суспензии. Потеря клеточного связывания с внеклеточным матриксом запускает процесс инверсии в органоидах, что приводит к изменению полярности органоидного эпителия, обнажая апикальную сторону эпителия для среды роста и интернализуя базолатеральную сторону.
В некоторых культурах органоиды в суспензии агрегируются и сливаются, эффект которого более глубокий в течение первых 3 дней(рисунок 1B,левая панель). Применение техники стрижения позволяет отслоить органоиды и продолжить посевы в течение суток с минимальной реагрегацией(рисунок 1В,правая панель).
Кишечные органоиды, культивируемые в куполах ECM, продолжают расширяться(рисунок 1C,левая панель)и проявляют спонтанное образование вторичных почковых структур, напоминающих небольшие крипты, на базолатеральной стороне эпителия, окружающего просвет(рисунок 1C,правая панель). В то же время органоиды, удерживаемые в течение 5 дней при отсутствии внеклеточного матрикса, продолжают развиваться в суспензии(рисунок 1D,левая панель). Инверсия полярности характеризуется утолщением (30-40 мкм) эпителия, окружающего ядро органоидов, и появлением различных морфологий: вытянутых(рисунок 1D,правая панель и дополнительный рисунок 1A),кистозных(дополнительный рисунок 1B)и нерегулярных(дополнительный рисунок 1C). Это часто сочетается с сокращением просветного пространства внутри органоида, что влияет на их общий размер.
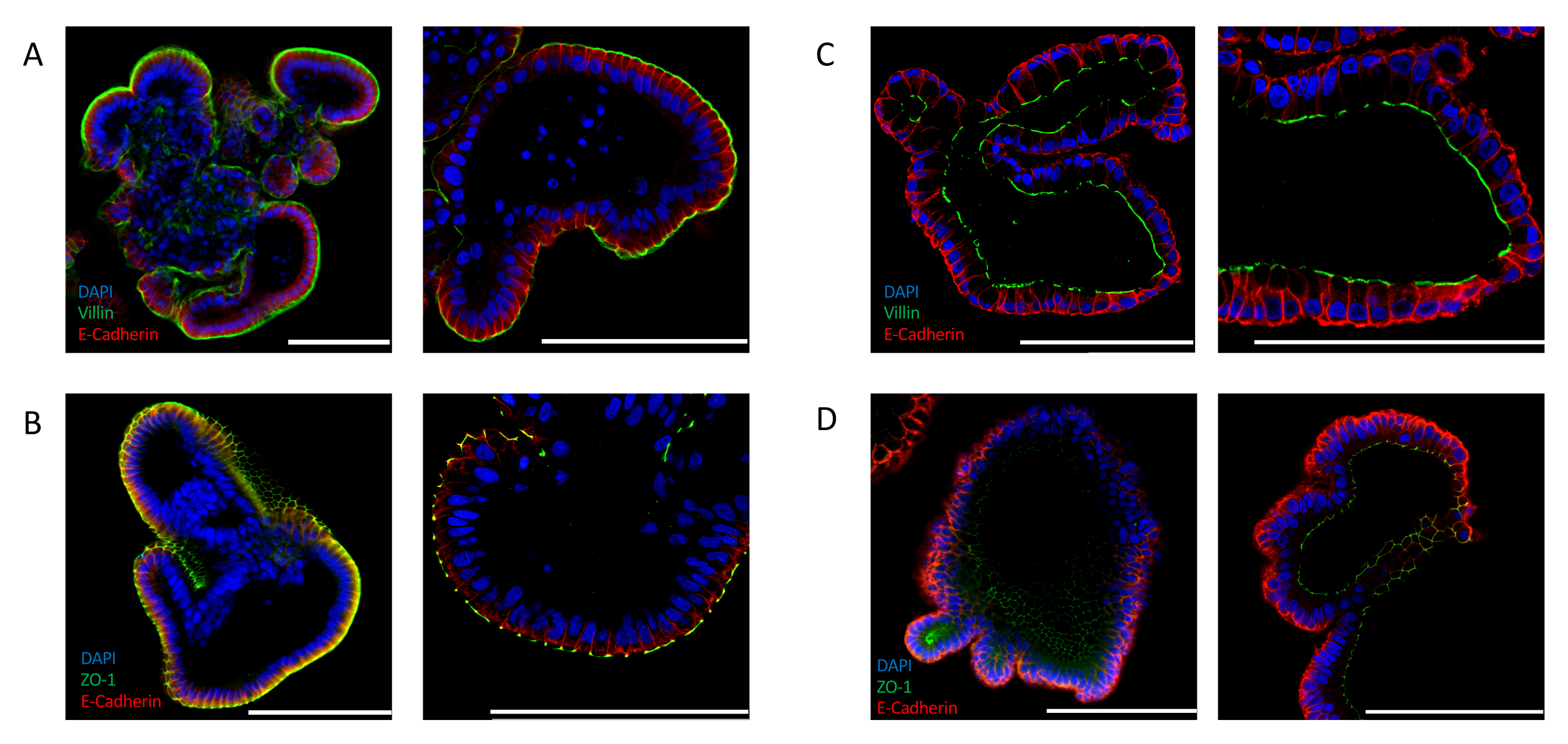
Эффективная инверсия также может быть подтверждена анализом экспрессии маркеров полярности, специфичных для кишечника. Апикальные органоиды кишечника показывают отчетливую локализацию ядер по направлению к просвету органоида, о чем свидетельствует сигнал 4',6-диамидино-2-фенилиндол (DAPI). Экспрессия апикального маркера, такого как VILLIN(Рисунок 2A)и ZO-1(Рисунок 2B),обнаруживается на внешней стороне эпителия, который подвергается воздействию среды. Эта локализация резко контрастирует с той, которая наблюдается в кишечных органоидах, культивируемых в ECM. Внеклеточные матриксные встроенные органоиды, окрашенные для ядер (DAPI), VILLIN(Рисунок 2C)и ZO-1(Рисунок 2D),демонстрируют апикобазальную полярность, где апикальная сторона обращена к просвету органоида.
Полное удаление ECM требуется для получения эффективной инверсии полярности органоидов кишечника. Иногда часть органоидов, обнаруженных в культурах суспензии, окруженных остатками ECM, показывает кистозную морфологию, которая предполагает сбой в инверсии полярности эпителия(дополнительный рисунок 2A). Анализ иммунофлуоресцентного окрашивания, выполненный на этих органоидах, дает доказательства базолатерального положения ядер (DAPI) вдоль эпителия и экспрессии ZO-1 на апикальной стороне, которая обращена к просвету органоида(дополнительный рисунок 2B),подтверждая, что неполное удаление ECM вызывает сохранение апикобазальной полярности способом, аналогичным ECM-встроенным органоидам.
Протокол создания кишечных однослойных культур приводит к слиянию однослойной культуры в течение 7 дней после посева, и культура часто достигает слияния всего за 2-3 дня. Одним из основных факторов, определяющих успех, является количество и качество клеток, используемых для засева монослоя. На рисунке 3A,Левая панель приведен пример идеальной плотности посева приблизительно 150 000 клеток во вставке мембраны клеточной культуры толщиной 6,5 мм. Это число не является фиксированным и может сильно вариабельно в зависимости от донора и качества исходной органоидной культуры; поэтому номер ячейки должен быть оптимизирован на основе этих переменных. Если плотность посева слишком низкая или низкого качества(рисунок 3А,правая панель), может отсечения недостаточно для образования сливающейся однослойной культуры.
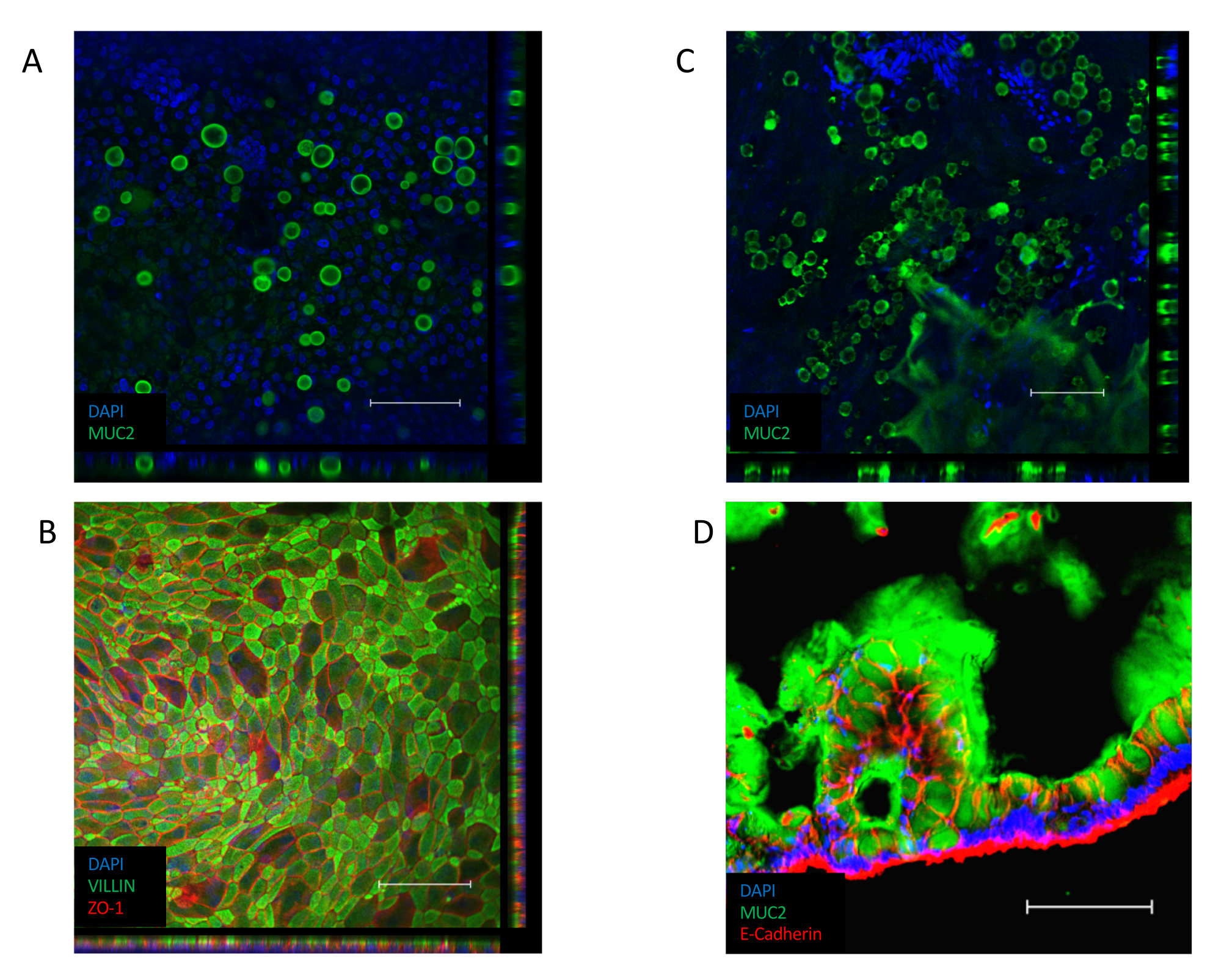
После того, как монослой установлен(рисунок 3B),ячейки образуют плотные соединения, создавая внешний вид булыжника(рисунок 3B,левая панель). Если им не удается сформировать слеженный монослой(рисунок 3B,правая панель),внешний вид монослоя часто будет «пятнистым», с областями хорошего качества клеточного прикрепления, но с большими промежутками между этими областями. Эти культуры не обеспечивают функционального барьера между базальным и апикальным отсеками и не подходят для описанных анализов. Слеивая монослой ориентирует свою VILLIN-содержащую границу кисти к апикальной стороне эпителия, а его ядро расположено к базолатеральному полюсу клетки(рисунок 4B). Между клетками образуются межклеточные соединения, состоящие из мультибелковых комплексов, включая ZO-1(рисунок 4B). Их присутствие является ключом к обеспечению барьерной функции эпителиальной культуры.
После слияния переход к культуре ALI вызывает дальнейшую дифференциацию культуры(рисунок 3C). Появляются небольшие круглые клетки-кувы, а сам монослой приобретает более складчатый вид. Хотя камеровидные клетки присутствуют в эпителии погруженной культуры(рисунок 4A),они более заметны после дифференцировки ALI. Клетки-голки, присутствующие в эпителии, выделяют слизь, что приводит к появлению тумана над верхней частью эпителия. Клетки чаба и секретируемая слизь могут быть визуализированы окрашиванием для секретируемого белка муцина MUC2(Рисунок 4A,C и D),а увеличение популяции клеток чаба может быть измерено увеличением экспрессии MUC2 (Дополнительный рисунок 3A). Нет необходимости удалять этот гелеобразный слой слизи и он прилипнет к поверхности эпителия и останется после повторных промываний. При необходимости удаления промывание культуры слизистым соединением, таким как 10 мМ N-ацетилцистеина или 50 мкг/мл DTT, удаляет избыток слизи. В дополнение к увеличению популяции бокаловидных клеток, интерфейс ALI также увеличивает присутствие энтероэндокринных клеток (как указано экспрессией CHGA) (дополнительный рисунок 3B)и зрелых энтероцитов (как указано экспрессией KRT20) (дополнительный рисунок 3C).

Рисунок 1:Стадии апикально-выхаченной генерации органоидов кишечника. (A) Репрезентативные изображения купола с органоидами нужного размера на 4-й день (Левая панель, шкала = 500 мкм). Органоиды тонкостенные, с открытым просветным отсеком (правая панель, шкала = 100 мкм). (B)Репрезентативное изображение скважины с обширной агрегацией после 3 дней в суспензии (левая панель, шкала = 200 мкм). Изображение фрагментов сгустков непосредственно после стрижения (правая панель, шкала = 200 мкм). (C) Репрезентативное изображение кишечных органоидов в куполе на 7-й день. Органоиды отображают расширенный просвет с образованием мелких почек на базолатеральной стороне эпителия (левое 20-кратное увеличение, правое 100-кратное увеличение отмеченной области, шкала бар = 200 мкм). (D) Репрезентативное изображение органоидов кишечника после удаления ECM и последующей культуры суспензии в течение 5 дней. Органоиды получают плотную морфологию с утолщенным эпителием и подвергают свою апикальную сторону среде. (Левое 20-кратное увеличение, Правое 100-кратное увеличение отмеченной области, Шкала = 200 мкм). Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}

Рисунок 2:Иммунофлуоресцентное окрашивание для маркеров полярности клеток в органоидах кишечника. Апикально-аутовые(A,B)и апикально-в(C,D), ориентированные на кишечник органоиды, окрашивали апикальными маркерами ZO-1 и VILLIN, а также эпителиальным маркером E-CADHERIN (красный). DAPI (синий) использовался для визуализации ядер. Левые панели отображают изображения, сделанные с 25-кратным увеличением, а правые панели отображают изображения различных органоидов с 63-кратным увеличением (только панель C отображает 25-кратное и 63-кратное увеличение одного и того же органоида). (A)Апикально-выявленные кишечные органоиды, окрашенные VILLIN (зеленый) и E-CADHERIN (красный), указывают на воздействие апикальной стороны на среду. (B)Апикальные кишечные органоиды, окрашенные ZO-1 (зеленый) и E-CADHERIN (красный), показывают наличие плотных соединений и возврат апикобазальной полярности. (C)Матригель-внедренный кишечный органоид, окрашенный VILLIN (зеленый) и E-CADHERIN (красный), показывающий апикальную сторону, обращенную к органоидному просвету. (D)Матригель-внедренные кишечные органоиды, окрашенные ZO-1 (зеленый) и E-CADHERIN (красный), указывающие на наличие апикальных плотных соединений, обращенных к просвету органоида. (Шкала = 100 мкм). Органоиды окрашивали иммунофлуоресценцией и визуализировали с использованием ранее опубликованных протоколов24,25. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}

Рисунок 3:Создание монослойных культур кишечника. (А) Репрезентативное изображение 3D органоидов после лечения 0,05% трипсина-ЭДТА. Органоиды диссоциируют на одиночные клетки или мелкие клеточные сгустки при подготовке к посеву однослойных культур. Левая панель: пример оптимальной плотности посева для однослойной культуры, приблизительно 150 000 клеток на 100 мкл на 6,5 мм клеточной культуральной мембранной вставке. Правая панель: пример субоптимальной плотности посева при <50 000 клеток на 100 мкл на мембране клеточной культуры толщиной 6,5 мм. (B) Репрезентативный образ погружной однослойной культуры. Левая панель: 100% слизый слой с характерным булыжником. Правая панель: приблизительно 50% сливного монослоя. Пробелы, наблюдаемые в монослое (обозначенные пунктирной линией), со временем закрываются из-за продолжающейся пролиферации кишечных стволовых клеток. (C) Репрезентативный яркий образ дифференцированной культуры ALI через 7 дней. (Шкала = 200 мкм). Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}

Рисунок 4:Иммунофлуоресцентное окрашивание для дифференцированных клеточных маркеров в монослойных культурах. (A)Z-стек изображение иммунофлуоресцентного окрашивания погружной монослойной культуры для белка муцина MUC2, указывающее на наличие камерочных клеток внутри монослойной культуры (зеленый = MUC2, синий = DAPI). (B) Z-образное изображение иммунофлуоресцентного окрашивания погружного монослоя. Окрашивание VILLIN (зеленый) вдоль апикального конца эпителия указывает на наличие границы кисти, а окрашивание ZO-1 (красный) указывает на наличие плотных соединений между клетками (синий = DAPI). (C)Z-стековое изображение иммунофлуоресцентного окрашивания ALI-дифференцированной монослойной культуры для белка муцина MUC2, указывающее на наличие значительно большего числа камерочных клеток в монослойной культуре ALI (зеленый = MUC2, синий = DAPI). (D)Криосекция ALI-дифференцированной монослойной культуры, окрашенной на наличие MUC2 (зеленый) и E-CADHERIN (красный), что указывает на наличие бокалочных клеток в эпителии и секрецию слизи вдоль апикальной стороны монослойной культуры. (Шкала = 200 мкм). Однослойные культуры окрашивали иммунофлуоресценцией и визуализировали с использованием ранее опубликованных протоколов26,27. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}
Дополнительный рисунок 1: Спектр фенотипов апикально-выпученного кишечного органоида в культуральной суспензии. (А,В,С) Дополнительные репрезентативные изображения морфологии органоидов кишечника сохраняются в суспензии через 5 дней после удаления ЭХМ. Органоидная полярность перевернулась. Органоиды стали более плотными с утолщенным эпителием, а верхушечная сторона органоидов обращена наружу. Органоиды могут проявлять различные морфологии: вытянутые(А),кистозные(В)и нерегулярные(С). (Шкала = 100 мкм). Пожалуйста, нажмите здесь, чтобы загрузить этот файл.
Дополнительный рисунок 2: Органоиды кишечника не могут инвертировать полярность в присутствии остаточной матриксной среды базальной мембраны в культурах суспензии. (A) Репрезентативное изображение неполного удаления ECM и неспособности инвертировать полярность органоидов. Остатки матригеля присутствуют вокруг органоидов и способствуют поддержанию полярности эпителия, ориентированной на апикальный в. Органоиды показывают кистозную морфологию с тонким эпителием, окружающим просвет (шкала бар = 200 мкм). (B) Репрезентативное изображение неинвертированного органоида, обнаруженного в условиях суспензии культуры. Ядра (синий = DAPI) и E-CADHERIN (красный) расположены на базолатеральной стороне, ZO-1 (зеленый) выражен на апикальной стороне, которая обращена к просвету органоида. (Шкала = 100 мкм). Пожалуйста, нажмите здесь, чтобы загрузить этот файл.
Дополнительный рисунок 3: Экспрессия генов дифференцированных клеточных маркеров в монослойных культурах. (А,В,С) Экспрессия MUC2, CHGAи KRT20 в погруженной и ALI-дифференцированной монослойной культуре, полученной в кишечной органоидной дифференцированной среде по сравнению с 3D-органоидной культурой, выращенной с помощью кишечной органоидной расширительной среды, установленной с помощью qPCR. Создание погружной однослойной культуры повышает экспрессию каждого дифференцированного клеточного маркера; однако дифференциация как культура ALI экспоненциально увеличивает экспрессию каждого маркера. Панели ошибок = +/- SEM. Пожалуйста, нажмите здесь, чтобы загрузить этот файл.
| МОНОСЛОЙНАЯ КУЛЬТУРНАЯ ПОСУДА | КОЛИЧЕСТВО КОЛОДЦЕВ КИШЕЧНЫХ ОРГАНОИДОВ ДЛЯ СБОРА (от 50 мкл купола/ на одну скважину, которую необходимо засеять) |
| 6,5 мм Трансвелл вставка | 1 - 2 скважины |
| 12 мм Трансвелл вставка | 3 - 4 скважины |
| 6-скважинная плита | 6 - 8 скважин |
| Плита из 24 скважин | 3 - 4 скважины |
| 96-скважинная плита | 1 - 2 скважины |
Таблица 1: Количество колодцев кишечных органоидов для сбора различной культуры
Обсуждение
Эпителиальные органоидные модели стали мощными платформами, которые могут быть использованы для моделирования организации тканей, прогрессирования заболевания и идентификациитерапевтических средств 23,28,29. Органоидная микроинъекция добавила ценность способности органоидов моделировать инфекционные заболевания, поскольку она позволяет патогенное взаимодействие с апикальной стороной эпителия хозяина. Последние достижения в области методов микроинъекции оптимизировали скорость инъекций в органоидах и достигли скорости до 90 вводимых органоидов в час. Барьерная функция в вводимых органоидах сохранялась, а низкая концентрация кислорода внутри просвета позволяла прижиться обязательно-анаэробным вводимымбактериям 30. Тем не менее, исследования отметили наличие гетерогенности в органоидных популяциях в пределах одной скважины. Эти различия наблюдались в размерах и форме31,уровнях экспрессии ключевых генов32,а также скорости пролиферации33. Дифференциальные реакции в пределах одной и той же органоидной популяции на такие соединения, как форсколин и PGE2, или на холериновый токсин, также были описаны28,33. Эти результаты подчеркивают необходимость высоких количеств органоидов в исследованиях и ограничивают использование люминальной инъекции.
Традиционная органоидная культура основана на инкапсуляции и размножении органоидов в гидрогеле. Однако гидрогели могут создавать ограничения на диффузию и вводить градиенты концентрации, которые могут увеличивать гетерогенность34. Кроме того, была задокументирована высокая изменчивость не только между культурами и донорами, но и в отдельных экспериментальных условиях. Донорский источник, биохимические свойства гидрогеля и внутренняя гетерогенность органоида как системы культуры являются важными факторами, которые могут увеличить экспериментальную изменчивость и ограничить воспроизводимость результатов, полученных в последующих приложениях. Оба способа, описанные здесь, обеспечивают простое средство обнажения апикальной стороны эпителия, позволяя моделировать интересующие соединения и патогены путем непосредственного добавления их в культуральную среду. Сокращение использования гидрогеля может ограничить экспериментальную изменчивость от технических источников ошибок.
Апикальные кишечные органоиды сохраняют ключевые характеристики системой модельных органоидов, а их масштабируемость делает их более поддаются высокопроизводительным анализам по сравнению с 2D-монослоем. Однако, поскольку органоиды сохраняют свою 3D-структуру, доступность базальной стороны ограничена и может препятствовать исследованиям, требующим доступа к обеим сторонам одновременно.
Мы продемонстрировали, что инверсия полярности кишечных органоидов зависит от эффективного и абсолютного удаления ECM, сохраняя при этом неповрежденную структуру органоидов. Как использование диссоциационного раствора для удаления ECM, так и антиагезивного раствора для предотвращения прилипания органоидов к пластиковому посовму способствовало повышению общей эффективности протокола, опубликованного Co, J. Y. и коллегами22,особенно в отношении количества апикальных органоидов, произведенных для последующих применений.
Кроме того, мы заметили, что наш протокол поддерживает более эффективную инверсию органоидов размером менее 250 мкм, и использование более крупных органоидов может привести к снижению выхода органоидов из-за фрагментации, вызванной пипеткой. Широкоствольный наконечник, такой как те, которые указаны в Таблице материалов,может позволить использовать более крупные органоиды. Тем не менее, широкостворные наконечники менее эффективны при диссоциации ECM по сравнению со стандартными наконечниками из-за более низкой приложенной механической силы. Поэтому повторение шагов 1.2.9-1.2.11 может потребоваться для достаточного разрушения и полного удаления всех остатков ECM при работе с более крупными органоидами.
Органоиды в суспензии могут выживать не менее 2 недель. По истечении этого периода времени мы наблюдали изменения морфологии и увеличение числа гибели клеток. Наличие пролиферающих клеток в апикально-ауторганоидах22 позволяет восстановить апикальные органоидные культуры кишечника. Это может быть достигнуто путем диссоциации апикальных органоидов на отдельные клетки и встраивания их в ECM со средой расширения органоидов кишечника.
Ограничением, часто встречающимся в протоколах, описывающих установление кишечных органоидов в культурах суспензии, является генерация больших агрегатов. Это влияет на несколько переменных, таких как эффективность, воспроизводимость морфологических признаков, проницаемость для соединений и паракриновая сигнализация. Подобно протоколу, опубликованному Co, J. Y. и коллегами, здесь мы подтверждаем, что получили инверсию полярности по меньшей мере 97% всех взвешенных органоидов без сдвига после 3 дней в суспензии. Однако, в отличие от публикации, мы ввели этап механической диссоциации с целью уменьшения образования крупных агрегатов и повышения урожайности. Поскольку эта процедура может повредить эпителий органоидов, мы продлили инкубационный период органоидов еще на 2 дня, чтобы обеспечить полное восстановление эпителия и обеспечить высокое качество культур для последующего применения. Введение постоянного перемешивания с использованием шейкера инкубатора или колбы спиннера может потенциально уменьшить события синтеза, свести к минимуму фрагментацию и увеличить оксигенацию. Эти альтернативные подходы могут поддерживать культуры в течение более длительных периодов времени, уменьшать гибель клеток и допускать дальнейшую дифференцировку апикальных органоидов кишечника.
Создание органоидного 2D-монослоя обеспечивает ряд преимуществ и недостатков по сравнению с органоидами с перевернутой полярностью. Описанный здесь протокол допускает быстрое создание сливаемой однослойной культуры, как правило, менее чем за 7 дней и возможность длительного поддержания культур в течение длительного периода времени (до 10 недель). Протокол и среда, используемые здесь, также позволяют эффективно дифференцировать значительное количество клеток, не всегда встречающихся в других органоидных монослойных культурах16. Установка монослоя на мембране вставки клеточной культуры обеспечивает одновременный доступ как к апикальной, так и к базолатеральной сторонам эпителия, что делает их идеальными для исследований барьерной целостности и переноса эпителия. Этот упрощенный доступ также делает их более поддаются исследованиям инфекций и лечения наркомании. Кроме того, эти культуры сохраняют многие характеристики, уникальные для донора, сохраняя их актуальность для исследований, ориентированных на конкретного пациента. Метод культивирования ALI также облегчает дифференцировку более функционального эпителия, состоящего как из секреторных, так и из абсорбирующих типов клеток, что делает его более репрезентативным для кишечного эпителия человека. Относительная стабильность этих культур также позволяет поддерживать их в течение длительного периода времени, обеспечивая возможность для долгосрочных исследований. Однако ограничениями этого подхода являются большое количество ячеек, необходимых для создания слижащегося монослоя, и необходимость поддержания полного слияния для функционального разделения между апикальной и базолатеральной камерами. Характерная архитектура крипты, которая может быть смоделирована в 3D органоидных культурах, также теряется при создании однослойной культуры. Тем не менее, экспериментально дружественный формат культуры и легкость, с которой можно получить доступ к апикальной и базолатеральной сторонам эпителия, делают ее мощным инструментом для изучения физиологии кишечника.
Раскрытие информации
G. S., W.C. и S. S. являются сотрудниками STEMCELL Technologies Ltd., Кембридж (Великобритания). M.S., F.E., S.L., A.E., и R.K.C. являются сотрудниками STEMCELL Technologies Inc., Ванкувер (Канада).
Благодарности
Это исследование финансировалось грантом Horizon 2020 OrganoVIR 812673 в рамках проекта Organoids for Virus Research - Инновационная программа обучения - ITN.
Материалы
| Name | Company | Catalog Number | Comments |
| Anti-Adherence Rinsing Solution | STEMCELL Technologies Inc. | 7010 | For coating cultureware. Referred as anti-adherent solution into the main text. |
| Conical tubes, 15 mL | STEMCELL Technologies Inc. | 38009 | |
| Corning Matrigel Matrix, Growth Factor Reduced (GFR), Phenol Red-Free | Corning | 356231 | Extracellular matrix (ECM) for maintenance and establishment of organoid lines. |
| Costar 6.5 mm or 12 mm Transwell inserts | STEMCELL Technologies Inc. | 38023/38024 | For 2D Monolayer culture. |
| Costar 24 Well Flat-Bottom, Tissue culture-treated plate | STEMCELL Technologies Inc. | 38017 | For maintenance and establishment of organoid lines. |
| D-PBS (Without Ca++ and Mg++) | STEMCELL Technologies Inc. | 37350 | For washing |
| Dimethyl sulfoxide (DMSO) | Millipore Sigma | D2650 | Reconstitution of small molecules |
| DMEM/F-12 with 15 mM HEPES | STEMCELL Technologies Inc. | 36254 | For washing |
| Gentle Cell Dissociation Reagent (GCDR) | STEMCELL Technologies Inc. | 7174 | For Matrigel removal. Referred as dissociation reagent into the main text. |
| IntestiCult Organoid Growth Medium (Human) | STEMCELL Technologies Inc. | 6010 | For expansion of organoid lines prior to differentiation. Referred as Intestinal Organoid Expansion Medium into the main text. |
| IntestiCult Organoid Differentiation Medium (Human) | STEMCELL Technologies Inc. | 100-0214 | For establishment of monolayers and 3D differentiation. Referred as Intestinal Organoid Differentiation Medium into the main text. |
| Trypsin-EDTA (0.05%) | STEMCELL Technologies Inc. | 7910 | For 2D Monolayer establishment. |
| Y-27632 | STEMCELL Technologies Inc. | 72302 | RHO/ROCK pathway inhibitor, Inhibits ROCK1 and ROCK2. Used for 2D monolayer establishment. |
| Wide bore tips | Corning | #TF-1005-WB-R-S | Organoids handling |
Ссылки
- Gehart, H., Clevers, H. Tales from the crypt: new insights into intestinal stem cells. Nature Reviews Gastroenterology and Hepatology. 16 (1), 19-34 (2019).
- Schneeberger, K., Roth, S., Nieuwenhuis, E. E. S., Middendorp, S. Intestinal epithelial cell polarity defects in disease: lessons from microvillus inclusion disease. Disease Models & Mechanisms. 11 (2), (2018).
- Klunder, L. J., Faber, K. N., Dijkstra, G., Van Ijzendoorn, S. C. D. Mechanisms of cell polarity - Controlled epithelial homeostasis and immunity in the intestine. Cold Spring Harbor Perspectives in Biology. 9 (7), 0227888 (2017).
- DiMarco, R. L., Hunt, D. R., Dewi, R. E., Heilshorn, S. C. Improvement of paracellular transport in the Caco-2 drug screening model using protein-engineered substrates. Biomaterials. 129, 152-162 (2017).
- Sun, H., Chow, E. C., Liu, S., Du, Y., Pang, K. S. The Caco-2 cell monolayer: usefulness and limitations. Expert Opinion on Drug Metabolism & Toxicology. 4 (4), 395-411 (2008).
- Clevers, H. Modeling development and disease with organoids. Cell. 165 (7), 1586-1597 (2016).
- Dutta, D., Heo, I., Clevers, H. Disease modeling in stem cell-derived 3D organoid systems. Trends in Molecular Medicine. 23 (5), 393-410 (2017).
- Zhang, Y. -. G., Wu, S., Xia, Y., Sun, J. Salmonella-infected crypt-derived intestinal organoid culture system for host-bacterial interactions. Physiological Reports. 2 (9), 12147 (2014).
- Nigro, G., Rossi, R., Commere, P. -. H., Jay, P., Sansonetti, P. J. The cytosolic bacterial peptidoglycan sensor Nod2 affords stem cell protection and links microbes to gut epithelial regeneration. Cell Host & Microbe. 15 (6), 792-798 (2014).
- Heo, I., et al. Modelling Cryptosporidium infection in human small intestinal and lung organoids. Nature Microbiology. 3 (7), 814-823 (2018).
- Bartfeld, S., et al. In vitro expansion of human gastric epithelial stem cells and their responses to bacterial infection. Gastroenterology. 148 (1), 126-136 (2015).
- Leslie, J. L., et al. Persistence and toxin production by Clostridium difficile within human intestinal organoids result in disruption of epithelial paracellular barrier function. Infection and Immunity. 83 (1), 138-145 (2015).
- Pleguezuelos-Manzano, C., et al. Mutational signature in colorectal cancer caused by genotoxic pks(+) E. coli. Nature. 580 (7802), 269-273 (2020).
- Fernando, E. H., et al. A simple, cost-effective method for generating murine colonic 3D enteroids and 2D monolayers for studies of primary epithelial cell function. American journal of physiology. Gastrointestinal and Liver Physiology. 313 (5), 467-475 (2017).
- Moon, C., VanDussen, K. L., Miyoshi, H., Stappenbeck, T. S. Development of a primary mouse intestinal epithelial cell monolayer culture system to evaluate factors that modulate IgA transcytosis. Mucosal Immunology. 7 (4), 818-828 (2014).
- Wang, Y., et al. Long-term culture captures injury-repair cycles of colonic stem cells. Cell. 179 (5), 1144-1159 (2019).
- Noel, G., et al. A primary human macrophage-enteroid co-culture model to investigate mucosal gut physiology and host-pathogen interactions. Scientific Reports. 7, 45270 (2017).
- VanDussen, K. L., et al. Development of an enhanced human gastrointestinal epithelial culture system to facilitate patient-based assays. Gut. 64 (6), 911-920 (2015).
- Thorne, C. A., et al. Enteroid monolayers reveal an autonomous WNT and BMP circuit controlling intestinal epithelial growth and organization. Developmental Cell. 44 (5), 624-633 (2018).
- Wang, A. Z., Ojakian, G. K., Nelson, W. J. Steps in the morphogenesis of a polarized epithelium: I. Uncoupling the roles of cell-cell and cell-substratum contact in establishing plasma membrane polarity in multicellular epithelial (MDCK) cysts. Journal of Cell Science. 95 (1), 137-151 (1990).
- Wang, A. Z., Ojakian, G. K., Nelson, W. J. Steps in the morphogenesis of a polarized epithelium: II. Disassembly and assembly of plasma membrane domains during reversal of epithelial cell polarity in multicellular epithelial (MDCK) cysts. Journal of Cell Science. 95 (1), 153-165 (1990).
- Co, J. Y., et al. Controlling epithelial polarity: a human enteroid model for host-pathogen interactions. Cell Reports. 26 (9), 2509-2520 (2019).
- Sato, T., et al. Long-term expansion of epithelial organoids from human colon, adenoma, adenocarcinoma, and Barrett's epithelium. Gastroenterology. 141 (5), 1762-1772 (2011).
- Dodt, H. -. U., et al. Ultramicroscopy: three-dimensional visualization of neuronal networks in the whole mouse brain. Nature Methods. 4 (4), 331-336 (2007).
- Fujii, M., et al. Human intestinal organoids maintain self-renewal capacity and cellular diversity in niche-inspired culture condition. Cell Stem Cell. 23 (6), 787-793 (2018).
- Crowley, S. M., et al. Intestinal restriction of Salmonella Typhimurium requires caspase-1 and caspase-11 epithelial intrinsic inflammasomes. PLoS Pathogens. 16 (4), 1008498 (2020).
- Rees, W. D., et al. Enteroids derived from inflammatory bowel disease patients display dysregulated endoplasmic reticulum stress pathways, leading to differential inflammatory responses and dendritic cell maturation. Journal of Crohn's & Colitis. 14 (7), 948-961 (2020).
- Dekkers, J. F., et al. A functional CFTR assay using primary cystic fibrosis intestinal organoids. Nature Medicine. 19 (7), 939-945 (2013).
- Almeqdadi, M., Mana, M. D., Roper, J., Yilmaz, &. #. 2. 1. 4. ;. H. Gut organoids: mini-tissues in culture to study intestinal physiology and disease. American Journal of Physiology. Cell Physiology. 317 (3), 405-419 (2019).
- Williamson, I. A., et al. A high-throughput organoid microinjection platform to study gastrointestinal microbiota and luminal physiology. Cellular and Molecular Gastroenterology and Hepatology. 6 (3), 301-319 (2018).
- Kassis, T., Hernandez-Gordillo, V., Langer, R., Griffith, L. G. OrgaQuant: human intestinal organoid localization and quantification using deep convolutional neural networks. Scientific Reports. 9 (1), 12479 (2019).
- Gracz, A. D., et al. A high-throughput platform for stem cell niche co-cultures and downstream gene expression analysis. Nature Cell Biology. 17 (3), 340-349 (2015).
- Gunasekara, D. B., et al. Development of arrayed colonic organoids for screening of secretagogues associated with enterotoxins. Analytical Chemistry. 90 (3), 1941-1950 (2018).
- Shin, W., et al. Spatiotemporal gradient and instability of Wnt induce heterogeneous growth and differentiation of human intestinal organoids. iScience. 23 (8), 101372 (2020).
Перепечатки и разрешения
Запросить разрешение на использование текста или рисунков этого JoVE статьи
Запросить разрешениеСмотреть дополнительные статьи
This article has been published
Video Coming Soon
Авторские права © 2025 MyJoVE Corporation. Все права защищены